
Bridging “Romer’s Gap”: Limb Mechanics of an Extant Belly-Dragging Lizard Inform Debate on...
18
1 23 Evolutionary Biology Evolutionary Biology ISSN 0071-3260 Evol Biol DOI 10.1007/s11692-013-9266-z Bridging “Romer’s Gap”: Limb Mechanics of an Extant Belly-Dragging Lizard Inform Debate on Tetrapod Locomotion During the Early Carboniferous John A. Nyakatura, Emanuel Andrada, Stefan Curth & Martin S. Fischer
Transcript of Bridging “Romer’s Gap”: Limb Mechanics of an Extant Belly-Dragging Lizard Inform Debate on...

1 23
Evolutionary BiologyEvolutionary Biology ISSN 0071-3260 Evol BiolDOI 10.1007/s11692-013-9266-z
Bridging “Romer’s Gap”: Limb Mechanicsof an Extant Belly-Dragging Lizard InformDebate on Tetrapod Locomotion Duringthe Early Carboniferous
John A. Nyakatura, Emanuel Andrada,Stefan Curth & Martin S. Fischer

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media New York. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

FOCAL REVIEWS
Bridging ‘‘Romer’s Gap’’: Limb Mechanics of an Extant Belly-Dragging Lizard Inform Debate on Tetrapod Locomotion Duringthe Early Carboniferous
John A. Nyakatura • Emanuel Andrada •
Stefan Curth • Martin S. Fischer
Received: 9 July 2013 / Accepted: 3 December 2013
� Springer Science+Business Media New York 2013
Abstract Devonian stem tetrapods are thought to have
used ‘crutching’ on land, a belly-dragging form of syn-
chronous forelimb action-powered locomotion. During the
Early Carboniferous, early tetrapods underwent rapid
radiation, and the terrestrial locomotion of crown-group
node tetrapods is believed to have been hindlimb-powered
and ‘raised’, involving symmetrical gaits similar to those
used by modern salamanders. The fossil record over this
period of evolutionary transition is remarkably poor (Ro-
mer’s Gap), but we hypothesize a phase of belly-dragging
sprawling locomotion combined with symmetrical gaits.
Since belly-dragging sprawling locomotion has differing
functional demands from ‘raised’ sprawling locomotion,
we studied the limb mechanics of the extant belly-dragging
blue-tongued skink. We used X-ray reconstruction of
moving morphology to quantify the three-dimensional
kinematic components, and simultaneously recorded single
limb substrate reaction forces (SRF) in order to calculate
SRF moment arms and the external moments acting on the
proximal limb joints. In the hindlimbs, stylopodal long-axis
rotation is more emphasized than in the forelimbs, and
much greater vertical and propulsive forces are exerted.
The SRF moment arm acting on the shoulder is at a local
minimum at the instant of peak force. The hindlimbs dis-
play patterns that more closely resemble ‘raised’ sprawling
species. External moment at the shoulder of the skink is
smaller than in ‘raised’ sprawlers. We propose an evolu-
tionary scenario in which the locomotor mechanics of
belly-dragging early tetrapods were gradually modified
towards hindlimb-powered, raised terrestrial locomotion
with symmetrical gait. In accordance with the view that
limb evolution was an exaptation for terrestrial locomotion,
the kinematic pattern of the limbs for the generation of
propulsion preceded, in our scenario, the evolution of
permanent body weight support.
Keywords Tiliqua scincoides � XROMM � X-ray
motion analysis � Substrate reaction force � Moment arm �External moment � Belly-dragging � Early tetrapods
Romer’s Gap Obscures Late Stem Group Tetrapod
Locomotor Evolution
In 1956, A.S. Romer recognized a hiatus in the continental
fossil record spanning the Early Carboniferous. During this
Tournaisian to mid-Visean time span, later called ‘‘Ro-
mer’s Gap’’ (Coates and Clack 1995), some key features of
the first terrestrial vertebrates were acquired (e.g., Ahlberg
and Milner 1994; Clack 2002a, b; Smithson et al. 2012).
Prior to the gap, late Devonian early tetrapods retained a
predominantly aquatic lifestyle and had very limited ter-
restrial locomotor capabilities (Clack 2002b; Pierce et al.
2012). Known late Devonian stem-group tetrapods such as
the iconic fossils Acanthostega and Ichthyostega were
relatively large animals (measuring between 0.5 and 1.2 m
in length), with the main bony elements of the tetrapod
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11692-013-9266-z) contains supplementarymaterial, which is available to authorized users.
J. A. Nyakatura (&) � S. Curth � M. S. Fischer
Institut fur Spezielle Zoologie und Evolutionsbiologie mit
Phyletischem Museum, Friedrich-Schiller-Universitat,
Erbertstraße 1, 07743 Jena, Germany
e-mail: [email protected]
E. Andrada
Institut fur Sportwissenschaft, Bewegungswissenschaft,
Friedrich-Schiller-Universitat, Seidelstraße 20, 07743 Jena,
Germany
123
Evol Biol
DOI 10.1007/s11692-013-9266-z
Author's personal copy

limbs already present, but with paddle-like feet and large,
heavy heads (Clack 2002b). By the mid-Carboniferous a
wide range of representatives of the lineages now assigned
to the crown-group of tetrapods were present and had
evolved varied morphologies including penta- or tetra-
dactylus limbs, secondary limb reduction or even loss,
different skull shapes, and smaller-bodied forms of about
0.1 m in length (Carroll et al. 1998; Paton et al. 1999; Janis
and Keller 2001; Clack 2002b; Smithson et al. 2012).
Importantly, species that seem to have lived predominantly
terrestrially were clearly present by the mid-Visean
(Smithson et al. 1994; Paton et al. 1999).
Although Romer’s Gap is increasingly being filled by
spectacular fossil findings such as Pederpes (Clack 2002a),
Ossinodus (Warren and Turner 2004), and various tetra-
pods from Tournaisian localities of Scotland (Smithson
et al. 2012; Smithson and Clack 2013), our understanding
of early tetrapod locomotor evolution close to the crown-
group node remains uncertain. For the predominantly
aquatic late Devonian stem tetrapod Ichthyostega, Pierce
et al. (2013a) propose a form of belly-dragging, synchro-
nous forelimb action-driven terrestrial locomotion with
little contribution from the hindlimbs, a locomotor mode
they hypothesize to have been widespread amongst
Devonian stem tetrapods on the occasions they ventured
onto land (Pierce et al. 2013a). Modern mudskippers
(Periophthalmini: Actinopterygii) are cited as functional
analogues to this inferred locomotor mode (Pierce et al.
2012, 2013a; see also Pace and Gibb 2009). In contrast, the
primitive (i.e., crown-group node) tetrapod condition (e.g.,
Edwards 1977; Gans and de Gueldre 1992) is often con-
sidered to be most closely approximated by the locomotion
of modern terrestrial salamanders. Salamander locomotion
is marked by (1) a symmetrical walking gait (either in
lateral sequence or trot; see Hildebrand 1966, Pridmore
1994), (2) hindlimb-driven propulsion (Kawano and Blob
2013), and (3) the ability to lift the belly off the ground.
Kawano and Blob (2013) point out that from the perspec-
tive of locomotor mechanics there is a striking difference
between the locomotor mode recently inferred for Devo-
nian stem tetrapods and that of extant salamanders. This
paper aims to help conceptualize this evolutionary transi-
tion in locomotor mechanics.
Given the low limb length to body mass ratios of early
crown-group tetrapods compared to extant sprawling liz-
ards (approximated from the ratio of limb length to hum-
eral and femoral shaft diameter put forward by Bakker
1971), early crown-group tetrapod locomotion has been
suggested to have been more clumsy, lumbering and less
agile (e.g., Bakker 1971; Reilly et al. 2006). Pridmore
proposes that late Devonian (predominantly aquatic) tet-
rapods were certainly belly-dragging on land because at
least three limbs would have had to be down at all times
just to support and lift the early tetrapod and prevent it
from falling over, and because this would necessitate more
complex brain control, it may not have been achieved
immediately (Pridmore 1994). Some support for the belly-
dragging theory is provided by potential body drag marks
tentatively assigned to tetrapod track makers in the pre-
served trackways of the Genoa River locality in Australia
(trackway II) and the Hoy Sandstone of Orkney in Scotland
(Warren and Wakefield 1972; Clack 1997). Subsequent
evolutionary transformation by terrestrially adapted mid-
Carboniferous crown-group tetrapods towards salamander-
like terrestrial locomotion remains obscured by the per-
sisting scarcity of body fossils and lack of unequivocal
terrestrial fossil tracks (e.g., Clack 1997).
The body mass of an approximately 1 m-long animal
such as Pederpes or Ossinodus (fossils which are closer to
the crown-group node of tetrapods than the stem tetrapods
Acanthostega and Ichthyostega mentioned above) would
have been many times greater than that of a 60 g extant
terrestrial salamander (such as Dicamptodon tenebrosus or
Ambystoma tigrinum, species often used to study salaman-
der locomotor mechanics, see Edwards 1977; Frolich and
Biewener 1992; Ashley-Ross 1994a, b; Ashley-Ross and
Bechtel 2004; Reilly et al. 2006; Ashley-Ross et al. 2013;
Kawano and Blob 2013). Body mass scales with the third
power of length, whereas the cross-sectional area of bone
and muscle (associated with structural strength and force
exertion) scale with the second power of length. It would
therefore need to be shown that specific compensatory
morphological differences in the musculoskeletal system of
relatively large early tetrapods were present before it could
be fully accepted that they were functionally similar to
extant salamanders in terrestrial, weight-bearing locomo-
tion. As an alternative view, we propose that early tetrapods
in the Early Carboniferous were not able to permanently
raise their belly off the ground when on land. While similar
on the whole, belly-dragging sprawling locomotion has
different functional demands than ‘raised’ sprawling loco-
motion without ventral trunk-ground contact (Fig. 1).
Analysis of Limb Mechanics During Belly-Dragging
Sprawling Locomotion in the Blue-Tongued Skink
Sprawling locomotion with belly-dragging potentially
represents an intermediate stage between the locomotor
mode inferred by Pierce et al. (2012, 2013a) for Devonian
stem tetrapods and the salamander-like locomotion pro-
posed to be primitive to crown tetrapods (e.g., Edwards
1977; Gans and de Gueldre 1992). We here use a relatively
large, belly-dragging lizard—the blue-tongued skink (Til-
iqua scincoides intermedia: Scincidae, Mitchell 1955)—to
study limb mechanics during belly-dragging sprawling
Evol Biol
123
Author's personal copy

locomotion. T. scincoides shares various features with late
Devonian stem tetrapods, including fore- and hindlimbs of
similar length, and manus and pes that point forward during
locomotion (Kubo 2010; Curth et al., in press). The skink
also shares low limb length to body length ratios with early
tetrapods (see supplemental information), and while not
quite equaling the size of the known *1 m fossils,
northern blue-tongued skinks reach up to 60 cm in total
length and more than 1.3 kg in body mass (Horner 2000).
Additionally, in contrast to many other skinks, T. scinco-
ides possess girdles and limbs that are not vestigial, and
full pentadactyl autpodia (Moch and Senter 2011).
Detailed three-dimensional (3D) kinematics, especially
long axis rotation (LAR) in the humerus and femur, have
rarely been studied in vivo and have never been quantified
in a belly-dragging lizard (Jenkins and Goslow 1983;
McElroy et al. 2012; Karakasiliotis et al. 2012). Neither
has any previous study examined the substrate reaction
forces (SRFs) exerted by the limbs during locomotion in a
belly-dragging lizard. To fill this gap in our knowledge, we
quantified the 3D stance-phase kinematics of the proximal
limb joints (shoulder, elbow, hip and knee) using X-ray
reconstruction of moving morphology (XROMM; Brainerd
et al. 2010; Gatesy et al. 2010). Simultaneously, we mea-
sured the SRFs exerted by individual limbs, and calculated
the SRF moment arms for each joint. Finally, these data
were combined and the external moments acting on the
proximal limb joints during locomotion were inferred. We
use the data to address the following questions regarding
comparative limb function in belly-dragging and ‘raised’
sprawling locomotion:
A. Do kinematic differences in fore- and hindlimbs to
‘raised’ sprawlers exist? Gray (1968) identified three
components that simultaneously contribute to progres-
sion: girdle rotation, active limb retraction and limb
rotation. Limb rotation propels the body forward by
means of the ‘double crank mechanism’(Barclay 1946),
in which the LAR of the stylopodia caused by the
contraction of the muscles that insert on the crista
ventralis of the humerus and femur, respectively, is
transmitted back to the manus and pes (see Edwards 1977;
Ashley-Ross 1994a for illustrative figures). We quantify
the relative contribution to the progression of the body of
the major kinematic aspects of sprawling tetrapod
locomotion in the fore- and hindlimbs, respectively.
B. Do the SRF profiles of the fore- and hindlimbs differ
in non-belly-dragging sprawlers? Chen et al. (2006),
Willey et al. (2004), and Kawano and Blob (2013)
certainly found differential leg function in trotting
geckos, alligators, and tiger salamanders, respectively,
with net-braking forelimbs (i.e., the limbs exert a
negative impulse during stance) and net-propulsive
hindlimbs (i.e., the limbs exert a positive impulse
during stance) during near-constant locomotion. On
the basis of these findings Kawano and Blob suggest
this to be a general pattern with an ancestry deep in the
use of stepping locomotion (Kawano and Blob 2013).
C. Are the moment arms of the net SRF vector-modulated
during stance? When SRF magnitudes are identical,
decreasing the moment arm will reduce external moments.
To maintain equilibrium, muscles must exert opposing
and equal net moments about the joints to balance
moments incurred by SRFs, gravity and inertia (e.g.
Biewener and Full 1992; Hutchinson and Garcia 2002).
Net musculo-skeletal forces at the joints will decrease if
the SRF moments about the joints are minimized.
Finally, we synthesize our findings with previous work
to infer aspects of the locomotor mechanics of a hypo-
thetical, belly-dragging evolutionary phase in tetrapod
locomotion that occurred during the Early Carboniferous.
Fig. 1 Schematic drawings to illustrate the differences in functional
demands between a belly-dragging (e.g., T. scincoides) and b non-
belly-dragging (e.g., green iguana) sprawling locomotion. During
belly-dragging sprawling locomotion the animal’s weight partially
rests on the trunk via direct contact to the substrate (see supplemen-
tary material). During non-belly-dragging sprawling locomotion
larger moments (blue arrows) need to be produced by the musculo-
skeletal system about the shoulder/hip and elbow/knee joints to bear
the animal’s weight. Thus, elbow/knee extensors and shoulder/hip
adductors need to be stronger. Schematic drawing not to scale (Color
figure online)
Evol Biol
123
Author's personal copy

Our results can be regarded as predictive and remain to be
validated by future fossil finds that add to the ‘population’
of Romer’s Gap.
Materials and Methods
Experimental Subjects
A pair of T. scincoides was purchased from an authorized
dealer (M&S Reptilien, Weigheim, Germany). The animals
were legal captive offspring from 2009 and were kept
separately in spacious terrariums (2.0 9 0.6 9 0.6 m).
Each terrarium contained sand as a substrate and various
places for the animals to hide. The terrariums were lit and
heated (12 h/day) by an ultraviolet lamp and a metal halide
lamp controlled by a clock timer. Temperatures were kept
at 28–35 �C during the day and 22�–25� at night. Water
was available ad libitum and the animals were given
appropriate food once a week. The female (body mass
1,150 g; total length 56 cm) was slightly larger than the
male (965 g; 54 cm). All animal care and experiments
were carried out in strict adherence to the animal welfare
regulations of the federal state of Thuringia, Germany and
were approved by the Thuringian committee for animal
research (license for husbandry: J-SHK-2684-05-04/11-1;
license for experiments: Reg.-Nr. 02-008/11).
Experimental Design
The experimental setup combined biplanar, high-speed
X-ray videography (for the specifics of the equipment see
Nyakatura et al. 2010) with the simultaneous measurement
of single limb SRFs using two synchronized, custom built
radiolucent force plates (for details see Andrada et al.
2013) which were placed within the capture volume of the
biplanar X-ray. The animals were motivated to traverse the
instrumented trackway (1.0 9 0.3 m) by gentle touches
with a stick on the tail that usually resulted in a series of up
to eight consecutive strides. To distinguish limb SRFs from
those resulting from ventral body contact, the forceplates
were arranged side-by-side relative to the direction of
motion to maximize the likelihood of a single limb making
contact. Synchronized X-ray sequences from the ventral
and lateral projection were recorded by the 38 cm diameter
image intensifiers at 500 frames per second at a resolution
of 1,536 9 1,024 pixels.
All trials in which more than a single limb made contact
with a plate were discarded. 43 successful trials were
recorded (often containing useful data from both forelimbs
and hindlimbs), on the basis of which the kinematics and
spatio-temporal characteristics of 20 forelimb strides and
20 hindlimb strides were analyzed. As the overall speed
range was minimal and inter-individual differences in gait
parameters consistently non-significant (Table 1), forelimb
and hindlimb data from the two individuals were pooled.
From this dataset, the ten hindlimb (four from the male and
six from the female) and ten forelimb (seven from the male
and three from the female) contact phases most similar in
speed were selected for further dynamic analysis. In this
step, kinematics and SRF vectors, magnitudes and moment
arm lengths were combined to determine the external
moments acting on the limb joints (see below).
X-ray Reconstruction of Moving Morphology
‘‘X-ray reconstruction of moving morphology’’ (XROMM,
Brainerd et al. 2010) was performed to estimate the kine-
matics of the fore- and hindlimbs. We made use of the
totally non-invasive approach known as ‘‘scientific roto-
scoping’’ (SR, Gatesy et al. 2010) which removes the need
to implant internal markers. In SR, an articulated 3D model
of the specimen’s skeleton (derived from computed
tomography in our case) is manually posed using the 3D
animation package Autodesk Maya� 2012 (Autodesk, San
Rafael, CA, USA) so that it overlays the individual’s X-ray
Table 1 Spatio-temporal parameters of the two experimental subjects
Female (average ± SD;
N = 10)
Male (average ± SD;
N = 10)
t test Average ± SD
(N = 20)
Speed (m/s) 0.095 ± 0.010 0.100 ± 0.011 P = 0.721* 0.097 ± 0.010
Diagonality 0.472 ± 0.025 0.482 ± 0.029 P = 0.415* 0.477 ± 0.027
Forelimb duty factor 0.679 ± 0.034 0.664 ± 0.040 P = 0.381* 0.672 ± 0.037
Forelimb stride length (m) 0.099 ± 0.019 0.097 ± 0.017 P = 0.801* 0.098 ± 0.018
Hindlimb duty factor 0.604 ± 0.045 0.579 ± 0.051 P = 0.264* 0.592 ± 0.048
Hindlimb stride length (m) 0.076 ± 0.016 0.082 ± 0.018 P = 0.440* 0.079 ± 0.017
Unpaired t tests were used to check for significant differences between the individuals (significance level P \ 0.05). Because none were found,
we decided to pool data from the two individuals
* Non-significant
Evol Biol
123
Author's personal copy

shadow in both biplanar views. This process is repeated for
key frames of the X-ray video which can then be interpo-
lated between (cubic–spline interpolation) to produce
smooth movements that closely approximate the recorded
kinematics.
Prior to SR, raw X-ray videos must be corrected for
distortion (Brainerd et al. 2010; Gatesy et al. 2010) and the
orientation of the X-ray image intensifiers in relation to the
subject is determined by recording a calibration object
(0.2 9 0.12 9 0.12 m) with metal beads inserted at regu-
lar distances that is placed within the biplanar field of view.
We used the freely available MATLAB routine developed
at Brown University, Providence, USA (www.xromm.org).
In order to obtain bone models, both individuals were
cooled to ca. 15 �C to reduce movement and scanned using
a scanner belonging to the Friedrich Schiller University
Hospital, Jena, Germany. Sedation was not necessary.
Virtual reconstruction of bones was performed using the
segmentation editor in the Amira software package (VSG,
Burlington, MA, USA).
In SR, bone models are linked to form a hierarchical
chain (Gatesy et al. 2010). Anatomical coordinate systems
are implemented at each joint to measure the movement of
the distally adjacent bone relative to the proximal bone
directly from the Maya animation. In theory, a whole six
degrees of freedom (DOF) can be derived for each joint
using this method, but since translations within limb joints
were too small to be reliably measured we only considered
the rotations. All movements are measured relative to a
reference pose. The reference pose was aligned to a
right-handed global coordinate system placed in the
trackway with positive x pointing in the direction of
movement, positive y pointing to the animals’ right, and
positive z pointing upwards. All bone model coordinate
systems were aligned to the axes of the global coordinate
system (see Nyakatura and Fischer 2010a). In order to
obtain anatomically meaningful data we used non-physio-
logical fully extended reference poses for both the fore-
and the hindlimbs (Sullivan 2007; Nyakatura and Fischer
2010a, b). To avoid the singularity problem, the rotation
order in each joint was set to have the largest expected
movement as the dominant axis (cf. Sullivan 2007).
For the pectoral model reference pose, coordinate sys-
tems were placed in (1) the interclavicle to measure the
movement of the girdle relative to the global coordinate
system (the girdle was oriented to have both shoulder joints
at the same height and the x-axis running through the
cranial tip of the interclavicle and the caudal tip of the
mesosternum), (2) the shoulder joint to measure humeral
movement relative to the pectoral girdle (the long axis of
humerus pointed in the direction of positive global x with
the flexor side of the elbow joint pointing ventral), (3) the
elbow joint to measure ulnar movement relative to the
humerus (the long axis of the ulna pointed in the direction
of positive global x), and (4) the wrist joint to be able to
animate manus placement.
For the pelvic model reference pose, coordinate systems
were placed (1) midway between the ilia to measure pelvic
movement relative the global coordinate system (the girdle
was oriented to have both hip joints and the ishiadic
tuberosity at the same height), (2) in the hip joint to
measure femoral movement relative to the pelvis (the long
axis of the femur was oriented to point in the direction of
positive global x with the flexor side of the knee facing
ventral), (3) in the knee joint to measure tibial movement
relative to the femur (the long axis of the tibia pointed in
the direction of positive global x), and (4) in the joint space
distal to the tibia to be able to animate pes placement.
In SR, the quality of the results is highly dependent on
the fidelity of the X-ray images (visibility of the bony
structures) and—since the mapping of the model onto the
X-ray-images is an approximation—the experience and
skill of the investigator. We were unable to reliably mea-
sure the movements of the bony elements of the pectoral
girdle relative to one another and of the diminutive carpal
and tarsal bones, and nor did we analyze the movement of
the pectoral girdle relative to the vertebral column. Our
analysis is thus restricted to the shoulder, hip, elbow and
knee joints—the constituent kinematic components of the
double crank mechanism. We also quantified the overall
movements of the pectoral girdle (treating it as one object)
and the pelvic girdle relative to a global coordinate system
placed in the midpoint of the runway. All data was
exported into Excel (v. 2010, Microsoft, Redmond, WA,
USA) and each trial was normalized to the same duration
(101 points; Nyakatura et al. 2010) in order to make the
kinematic profiles (i.e., plots of kinematic data vs. percent
of stance) of trials of slightly different duration directly
comparable.
One of the merits of XROMM is the opportunity it
affords to conduct ‘virtual experiments’. This enabled us to
work out the relative contribution of individual kinematic
components of complex movements (e.g., LAR vs.
retraction in the shoulder and hip) as a percentage of total
manus and pes displacement, respectively, relative to the
body as it is moved over the fixed manus and pes during
stance. Because the autopodia are fixed to the ground
during stance, this percentage is a proxy for the contribu-
tion of a kinematic component to the progression of the
body. To work it out, we used the XROMM animation of
each trial and ‘muted’ (inactivated) the movement com-
ponent under consideration at touch-down, while all the
other components were kept active during stance (Nyaka-
tura and Fischer 2010a). The relative trunk displacement
that resulted was then compared to the displacement
observed in the original trial. The relative contribution of
Evol Biol
123
Author's personal copy

the inactivated kinematic component to trunk progression
is defined herein as the difference between the virtual
experimental trial and the original trial.
Substrate Reaction Forces and External Moments
In order to avoid having metal in the runway (and hence the
occlusion of the X-ray beam) the force plates were custom-
built from carbon fiber (Andrada et al. 2013). As transducer
elements, we used 6-DOF force-torque transducers (ATI
nano17; ATI Industrial Automation, Apex, NC, USA). The
force plates were flush with the runway and, to prevent the
lizards from slipping, the runway was covered with emery
paper. The set-up permitted the resolution of the SRF into
antero-posterior (or cranio-caudal; Fx), medio-lateral (Fy)
and vertical (Fz) components. To match the kinematic data
sampling rate, we collected SRFs at 500 Hz using cus-
tomized software for LabView 2009 (NI USB-6229,
National Instruments Germany GmbH, Munich, Germany).
We synchronized force and X-ray data acquisition by
matching the X-ray frame of apparent touch-down with the
initial spike of Fz. After smoothing using a low pass zero-
phase filter, the components of the SRF force trace were
again normalized to the same duration (101 points, see
above) using a custom MATLAB routine provided by B.
Hesse (FSU Jena, Germany). All SRFs were normalized to
body weight units (BW). To be able to pool data for the left
and right body sides, we set positive Fy to always point
medially.
External moment is the product of the moment arm, or
distance vector, and the SRF vector. The orientation of the
SRF vector for every 5 % increment in stance duration was
animated in Maya using a Maya ‘‘locator’’ in accordance
with the magnitudes of Fx, Fy, and Fz relative to a coordi-
nate system placed at the center of pressure (CoP) and in
line with the inherent coordinate system of the force plate
(Fig. 2). The positions of this locator were spline interpo-
lated in Maya for all in-between frames. An irregular tri-
angle was spanned between the CoP, the SRF orientation
locator and the joint. We used the Maya distance mea-
surement tool to measure, over time, the lengths a, b and c
(Fig. 2) of the triangle (it has previously been shown using
the known span of a caliper that measuring lengths in the
virtual experimental environment is relatively accurate; see
Nyakatura and Fischer 2010a). In accordance with the law
of cosines, the known lengths were used to calculate the
angle a, where
a ¼ arccosb2 þ c2 � a2
2bc
� �ð1Þ
The external moment arm (R) in 3D space, then, is part
of a right-angled triangle in which b is the hypotenuse and
R is opposite the angle a. The moment arm was calculated
as R = sin a 9 b (Fig. 2). Finally, the moment acting on a
joint was calculated as R 9 magnitude of SRF vector. R is
the shortest distance between the SRF vector and the joint,
but does not relate to any anatomical frame of reference
and can be regarded as a 3D moment arm of the instanta-
neous SRF. External moment, induced by load and inertia,
is here used as an indicator of the magnitude of net mus-
culoskeletal forces that exert opposing and equal net
moments to maintain equilibrium.
Fig. 2 Estimating external moments acting on limb joints during
stance phase—method shown here for the left shoulder for one frame.
The SRF vector (orange) of a trial was animated within the
corresponding XROMM animation. The origin of the SRF vector
(CoP) was assumed to be underneath the third metacarpal-phalangeal
(third metatarsal-phalangeal in the hindlimb) of digit III. CoP center
of pressure; a distance between joint and tip of SRF vector; b distance
between CoP and joint; c distance between CoP and tip of SRF vector.
For further explanation see text (Color figure online)
Evol Biol
123
Author's personal copy
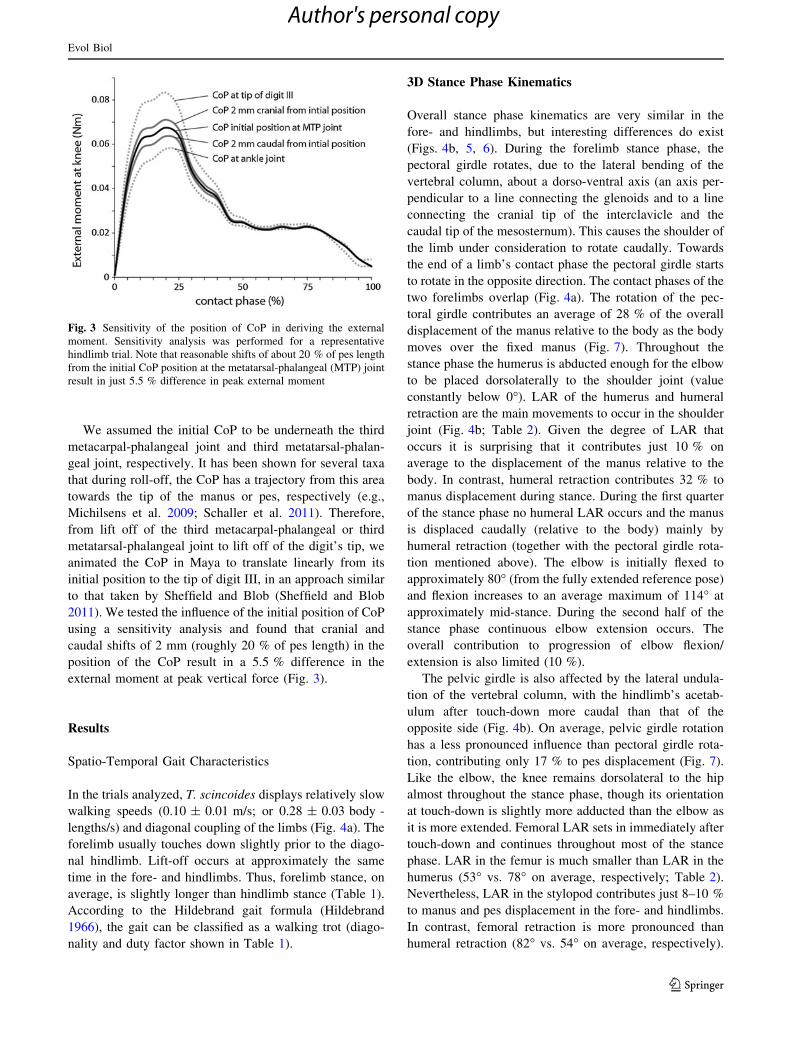
We assumed the initial CoP to be underneath the third
metacarpal-phalangeal joint and third metatarsal-phalan-
geal joint, respectively. It has been shown for several taxa
that during roll-off, the CoP has a trajectory from this area
towards the tip of the manus or pes, respectively (e.g.,
Michilsens et al. 2009; Schaller et al. 2011). Therefore,
from lift off of the third metacarpal-phalangeal or third
metatarsal-phalangeal joint to lift off of the digit’s tip, we
animated the CoP in Maya to translate linearly from its
initial position to the tip of digit III, in an approach similar
to that taken by Sheffield and Blob (Sheffield and Blob
2011). We tested the influence of the initial position of CoP
using a sensitivity analysis and found that cranial and
caudal shifts of 2 mm (roughly 20 % of pes length) in the
position of the CoP result in a 5.5 % difference in the
external moment at peak vertical force (Fig. 3).
Results
Spatio-Temporal Gait Characteristics
In the trials analyzed, T. scincoides displays relatively slow
walking speeds (0.10 ± 0.01 m/s; or 0.28 ± 0.03 body -
lengths/s) and diagonal coupling of the limbs (Fig. 4a). The
forelimb usually touches down slightly prior to the diago-
nal hindlimb. Lift-off occurs at approximately the same
time in the fore- and hindlimbs. Thus, forelimb stance, on
average, is slightly longer than hindlimb stance (Table 1).
According to the Hildebrand gait formula (Hildebrand
1966), the gait can be classified as a walking trot (diago-
nality and duty factor shown in Table 1).
3D Stance Phase Kinematics
Overall stance phase kinematics are very similar in the
fore- and hindlimbs, but interesting differences do exist
(Figs. 4b, 5, 6). During the forelimb stance phase, the
pectoral girdle rotates, due to the lateral bending of the
vertebral column, about a dorso-ventral axis (an axis per-
pendicular to a line connecting the glenoids and to a line
connecting the cranial tip of the interclavicle and the
caudal tip of the mesosternum). This causes the shoulder of
the limb under consideration to rotate caudally. Towards
the end of a limb’s contact phase the pectoral girdle starts
to rotate in the opposite direction. The contact phases of the
two forelimbs overlap (Fig. 4a). The rotation of the pec-
toral girdle contributes an average of 28 % of the overall
displacement of the manus relative to the body as the body
moves over the fixed manus (Fig. 7). Throughout the
stance phase the humerus is abducted enough for the elbow
to be placed dorsolaterally to the shoulder joint (value
constantly below 0�). LAR of the humerus and humeral
retraction are the main movements to occur in the shoulder
joint (Fig. 4b; Table 2). Given the degree of LAR that
occurs it is surprising that it contributes just 10 % on
average to the displacement of the manus relative to the
body. In contrast, humeral retraction contributes 32 % to
manus displacement during stance. During the first quarter
of the stance phase no humeral LAR occurs and the manus
is displaced caudally (relative to the body) mainly by
humeral retraction (together with the pectoral girdle rota-
tion mentioned above). The elbow is initially flexed to
approximately 80� (from the fully extended reference pose)
and flexion increases to an average maximum of 114� at
approximately mid-stance. During the second half of the
stance phase continuous elbow extension occurs. The
overall contribution to progression of elbow flexion/
extension is also limited (10 %).
The pelvic girdle is also affected by the lateral undula-
tion of the vertebral column, with the hindlimb’s acetab-
ulum after touch-down more caudal than that of the
opposite side (Fig. 4b). On average, pelvic girdle rotation
has a less pronounced influence than pectoral girdle rota-
tion, contributing only 17 % to pes displacement (Fig. 7).
Like the elbow, the knee remains dorsolateral to the hip
almost throughout the stance phase, though its orientation
at touch-down is slightly more adducted than the elbow as
it is more extended. Femoral LAR sets in immediately after
touch-down and continues throughout most of the stance
phase. LAR in the femur is much smaller than LAR in the
humerus (53� vs. 78� on average, respectively; Table 2).
Nevertheless, LAR in the stylopod contributes just 8–10 %
to manus and pes displacement in the fore- and hindlimbs.
In contrast, femoral retraction is more pronounced than
humeral retraction (82� vs. 54� on average, respectively).
Fig. 3 Sensitivity of the position of CoP in deriving the external
moment. Sensitivity analysis was performed for a representative
hindlimb trial. Note that reasonable shifts of about 20 % of pes length
from the initial CoP position at the metatarsal-phalangeal (MTP) joint
result in just 5.5 % difference in peak external moment
Evol Biol
123
Author's personal copy
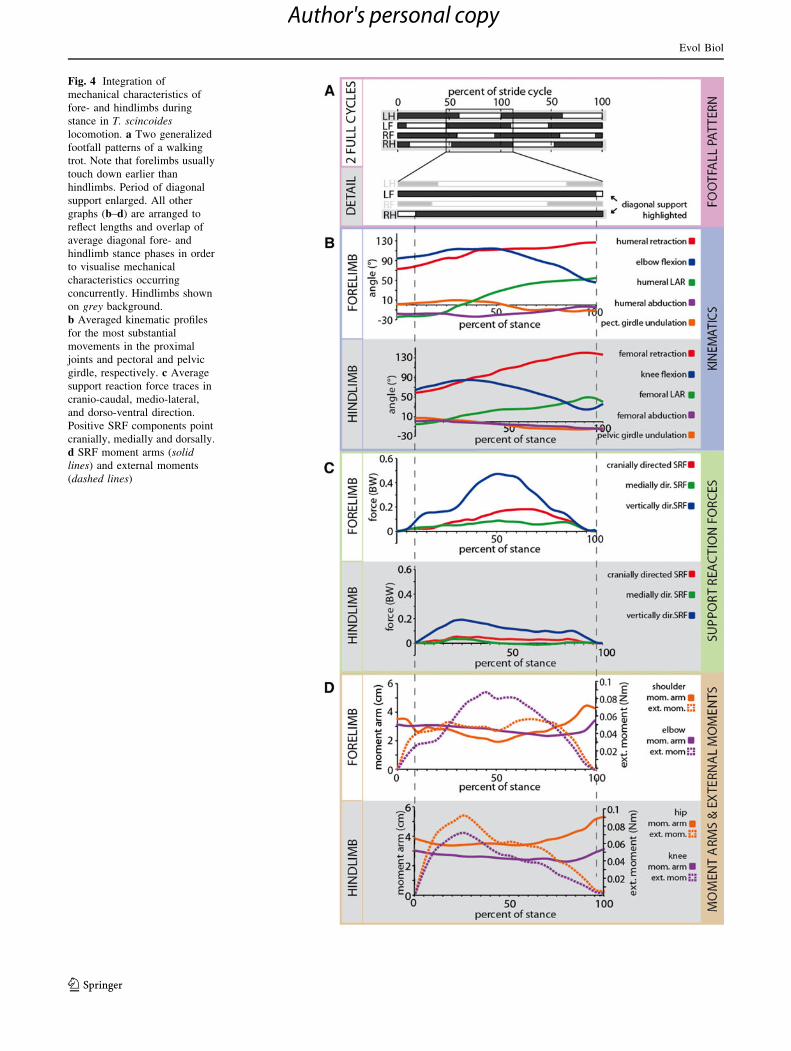
Fig. 4 Integration of
mechanical characteristics of
fore- and hindlimbs during
stance in T. scincoides
locomotion. a Two generalized
footfall patterns of a walking
trot. Note that forelimbs usually
touch down earlier than
hindlimbs. Period of diagonal
support enlarged. All other
graphs (b–d) are arranged to
reflect lengths and overlap of
average diagonal fore- and
hindlimb stance phases in order
to visualise mechanical
characteristics occurring
concurrently. Hindlimbs shown
on grey background.
b Averaged kinematic profiles
for the most substantial
movements in the proximal
joints and pectoral and pelvic
girdle, respectively. c Average
support reaction force traces in
cranio-caudal, medio-lateral,
and dorso-ventral direction.
Positive SRF components point
cranially, medially and dorsally.
d SRF moment arms (solid
lines) and external moments
(dashed lines)
Evol Biol
123
Author's personal copy
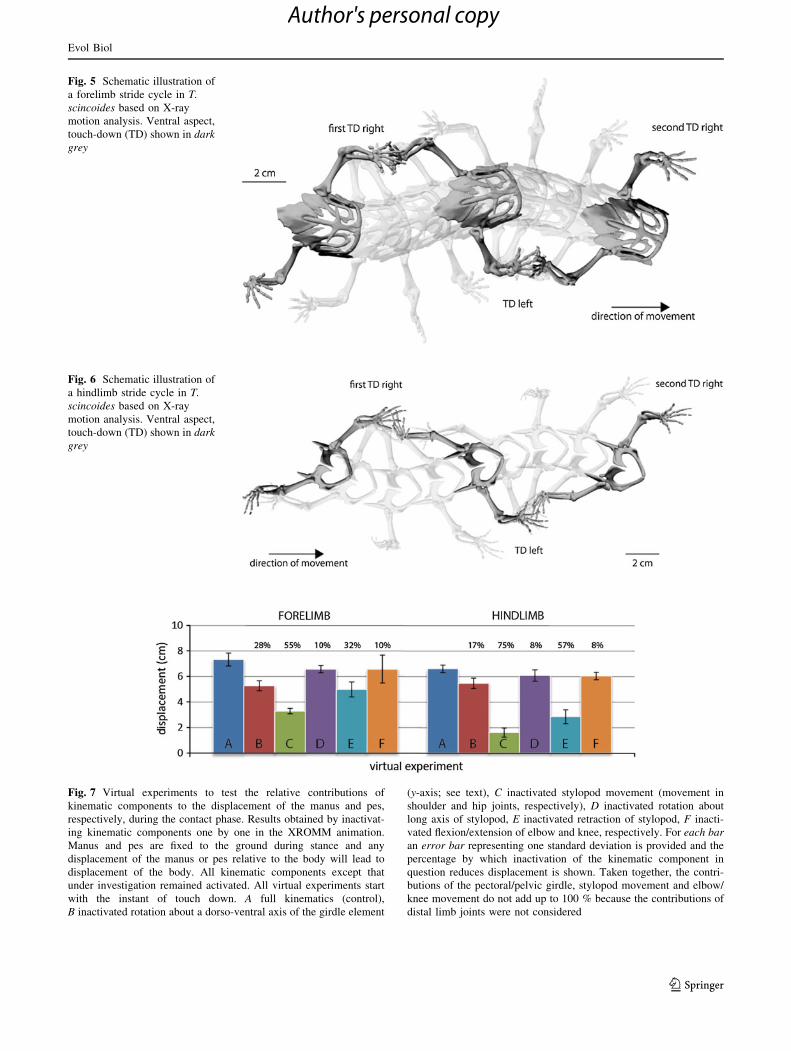
Fig. 5 Schematic illustration of
a forelimb stride cycle in T.
scincoides based on X-ray
motion analysis. Ventral aspect,
touch-down (TD) shown in dark
grey
Fig. 6 Schematic illustration of
a hindlimb stride cycle in T.
scincoides based on X-ray
motion analysis. Ventral aspect,
touch-down (TD) shown in dark
grey
Fig. 7 Virtual experiments to test the relative contributions of
kinematic components to the displacement of the manus and pes,
respectively, during the contact phase. Results obtained by inactivat-
ing kinematic components one by one in the XROMM animation.
Manus and pes are fixed to the ground during stance and any
displacement of the manus or pes relative to the body will lead to
displacement of the body. All kinematic components except that
under investigation remained activated. All virtual experiments start
with the instant of touch down. A full kinematics (control),
B inactivated rotation about a dorso-ventral axis of the girdle element
(y-axis; see text), C inactivated stylopod movement (movement in
shoulder and hip joints, respectively), D inactivated rotation about
long axis of stylopod, E inactivated retraction of stylopod, F inacti-
vated flexion/extension of elbow and knee, respectively. For each bar
an error bar representing one standard deviation is provided and the
percentage by which inactivation of the kinematic component in
question reduces displacement is shown. Taken together, the contri-
butions of the pectoral/pelvic girdle, stylopod movement and elbow/
knee movement do not add up to 100 % because the contributions of
distal limb joints were not considered
Evol Biol
123
Author's personal copy
This difference is reflected in the relative contributions to
pes displacement of femoral retraction (57 %) and humeral
retraction (32 %). Like the elbow, the knee undergoes
initial flexion followed by extension later in stance
(Fig. 4b). However, starting from a more extended initial
pose (angle at touch-down 64� vs. 94� in the elbow;
Table 2), maximal flexion (84�) is already reached by
about 25 % of stance. The contributions of elbow and knee
flexion/extension to pes displacement are relatively small
(Fig. 7).
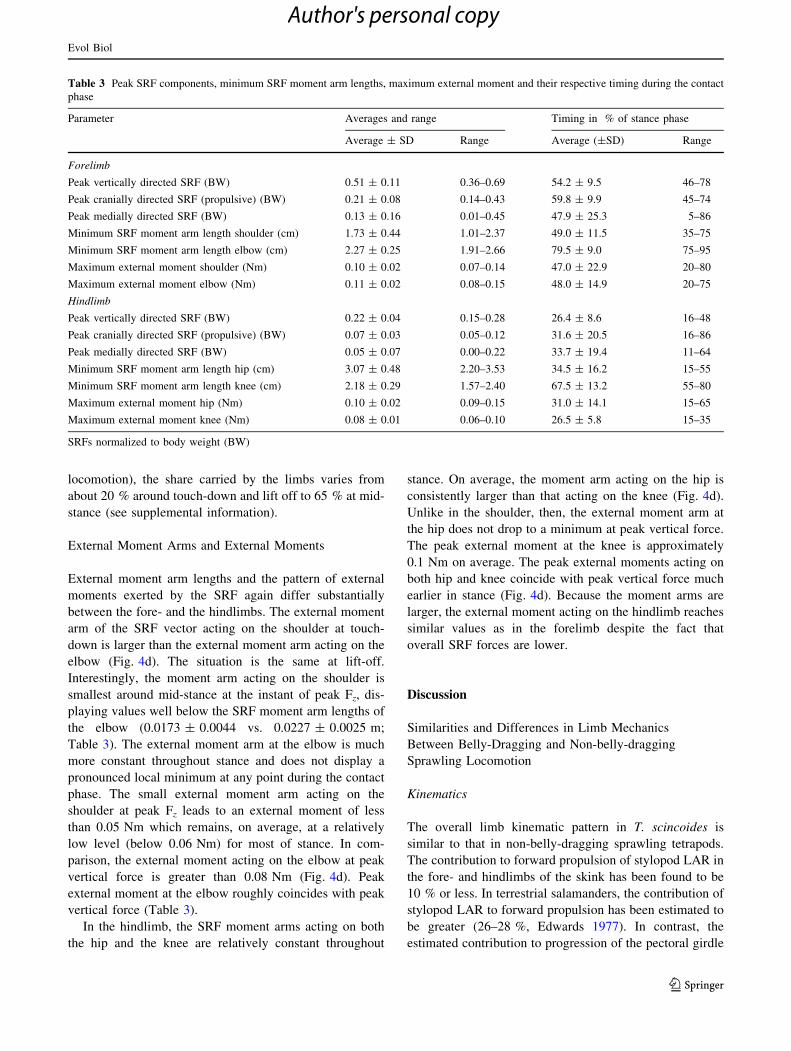
Substrate Reaction Forces
In contrast to the kinematic aspects documented here,
substrate reaction forces differ substantially between the
fore- and the hindlimbs (Fig. 4c). Forces are much greater
in the forelimbs, with the vertical component being the
largest in both the fore- and the hindlimbs. In the forelimb,
Fz increases moderately but not abruptly after touch-down
and reaches its maximum shortly after mid-stance (peak
Fz = 0.51 ± 0.11 BW; Table 3). Peak Fz occurs concur-
rently with maximum elbow flexion, but as humeral
rotation already has positive values, the forearm is some-
what ventro-caudally orientated. On average, Fx is always
positive (peak Fx = 0.21 ± 0.08 BW; Fig. 4c), exerting a
propulsive impulse throughout stance. Fy is comparatively
small (average peak Fy is just 0.13 ± 0.16 BW) and, on
average, medially directed throughout the stance phase.
In the hindlimb, peak Fz is exerted much earlier in the
stance phase (at 26.4 ± 8.6 % of stance). Just as peak
forelimb Fz and maximum elbow flexion coincide, peak
hindlimb Fz coincides with peak flexion of the knee
(Fig. 4). At peak Fz, less than a quarter of the body weight
is brought to bear on the support (Table 3). Values for peak
Fx and peak Fy are smaller than in the forelimb (Table 3),
but like in the forelimb there is a constant propulsive
impulse, even though it is smaller in overall magnitude.
Medio-lateral forces are small and slightly laterally direc-
ted after midstance.
The pattern of forces exerted on the ground during
stance means that the share of the total body weight carried
by the limbs is non-constant. The remaining share is car-
ried by the trunk via ventral contact with the support.
Neglecting the effects of inertia (assuming quasi-static
Table 2 Kinematic parameters at touch down, mid contact and lift off
Touch down angle (�) Mid contact angle (�) Lift off angle (�)
Average (±SD) Range Average (±SD) Range Average (±SD) Range
Pectoral girdle
x-Axis rotation 1.2 ± 2.1 -2.7 to 4.3 2.8 ± 3.6 -3.4 to 7.6 1.0 ± 7.6 -12.6 to 10.9
y-Axis rotation -5.0 ± 3.8 -9.7 to 1.6 -4.9 ± 4.9 -8.9 to 7.8 -9.5 ± 2.7 -12.6 to -4.0
z-Axis rotation 2.0 ± 9.4 -17.8 to 14.8 6.6 ± 1.5 3.7 to 8.5 -12.2 ± 9.1 -23.5 to 4.4
Shoulder
x-Axis rotation -24.3 ± 6.3 -36.2 to -10.2 19.5 ± 12.4 3.5 to 46.8 54.7 ± 12.2 43.7 to 75.7
y-Axis rotation -17.9 ± 9.5 -31.8 to -3.5 -22.3 ± 10.9 -43.8 to -7.1 -4.6 ± 4.1 -11.8 to 8.8
z-Axis rotation 72.9 ± 13.8 54.9 to 89.4 111.3 ± 16.1 92.7 to 129.5 126.0 ± 6.6 117.5 to 137.4
Elbow
x-Axis rotation 9.7 ± 8.3 -0.5 to 23.8 10.2 ± 6.6 -0.1 to 25.0 13.2 ± 8.3 -3.3 to 26.7
y-Axis rotation 94.2 ± 22.4 69.4 to 131.2 114.3 ± 10.1 97.7 to 134.8 45.4 ± 12.9 26.9 to 67.5
z-Axis rotation 45.4 ± 12.9 -26.8 to 7.2 7.9 ± 9.9 -9.4 to 25.1 12.4 ± 13.2 -11.3 to 41.9
Pelvic girdle
x-Axis rotation -0.4 ± 0.9 -1.7 to 1.1 -4.6 ± 2.2 -7.3 to -1.1 6.9 ± 5.6 0.9 to 15.4
y-Axis rotation -15.5 ± 0.5 -16.8 to -14.9 -15.5 ± 0.9 -16.7 to -13.7 -18.3 ± 0.5 -19.1 to -17.4
z-Axis rotation 7.5 ± 2.5 4.2 to 11.2 -5.4 ± 2.2 -8.9 to -1.9 -12.2 ± 4.5 -17.4 to -5.5
Hip
x-Axis rotation -5.2 ± 7.2 -14.7 to 6.0 24.7 ± 6.8 13.7 to 35.1 40.6 ± 6.0 32.1 to 52.8
y-Axis rotation 1.2 ± 3.6 -6.4 to 6.8 -4.5 ± 2.9 -8.9 to 1.8 -15.7 ± 5.4 -22.8 to -5.1
z-Axis rotation 58.9 ± 4.0 52.2 to 65.7 104.7 ± 7.9 99.8 to 126.2 136.2 ± 16.2 105.9 to 161.4
Knee
x-Axis rotation -19.9 ± 9.1 -24.5 to 5.0 -19.7 ± 6.7 -29.5 to -4.3 -15.1 ± 9.8 -32.4 to 1.9
y-Axis rotation 64.6 ± 7.2 53.3 to 77.1 76.5 ± 5.9 69.2 to 89.6 36. 2 ± 21.3 8.0 to 81.8
z-Axis rotation -2.5 ± 2.9 -5.9 to 1.7 -5.8 ± 7.5 -18.9 to 4.2 -3.2 ± 6.1 -14.9 to 8.3
Evol Biol
123
Author's personal copy
locomotion), the share carried by the limbs varies from
about 20 % around touch-down and lift off to 65 % at mid-
stance (see supplemental information).
External Moment Arms and External Moments
External moment arm lengths and the pattern of external
moments exerted by the SRF again differ substantially
between the fore- and the hindlimbs. The external moment
arm of the SRF vector acting on the shoulder at touch-
down is larger than the external moment arm acting on the
elbow (Fig. 4d). The situation is the same at lift-off.
Interestingly, the moment arm acting on the shoulder is
smallest around mid-stance at the instant of peak Fz, dis-
playing values well below the SRF moment arm lengths of
the elbow (0.0173 ± 0.0044 vs. 0.0227 ± 0.0025 m;
Table 3). The external moment arm at the elbow is much
more constant throughout stance and does not display a
pronounced local minimum at any point during the contact
phase. The small external moment arm acting on the
shoulder at peak Fz leads to an external moment of less
than 0.05 Nm which remains, on average, at a relatively
low level (below 0.06 Nm) for most of stance. In com-
parison, the external moment acting on the elbow at peak
vertical force is greater than 0.08 Nm (Fig. 4d). Peak
external moment at the elbow roughly coincides with peak
vertical force (Table 3).
In the hindlimb, the SRF moment arms acting on both
the hip and the knee are relatively constant throughout
stance. On average, the moment arm acting on the hip is
consistently larger than that acting on the knee (Fig. 4d).
Unlike in the shoulder, then, the external moment arm at
the hip does not drop to a minimum at peak vertical force.
The peak external moment at the knee is approximately
0.1 Nm on average. The peak external moments acting on
both hip and knee coincide with peak vertical force much
earlier in stance (Fig. 4d). Because the moment arms are
larger, the external moment acting on the hindlimb reaches
similar values as in the forelimb despite the fact that
overall SRF forces are lower.
Discussion
Similarities and Differences in Limb Mechanics
Between Belly-Dragging and Non-belly-dragging
Sprawling Locomotion
Kinematics
The overall limb kinematic pattern in T. scincoides is
similar to that in non-belly-dragging sprawling tetrapods.
The contribution to forward propulsion of stylopod LAR in
the fore- and hindlimbs of the skink has been found to be
10 % or less. In terrestrial salamanders, the contribution of
stylopod LAR to forward propulsion has been estimated to
be greater (26–28 %, Edwards 1977). In contrast, the
estimated contribution to progression of the pectoral girdle
Table 3 Peak SRF components, minimum SRF moment arm lengths, maximum external moment and their respective timing during the contact
phase
Parameter Averages and range Timing in % of stance phase
Average ± SD Range Average (±SD) Range
Forelimb
Peak vertically directed SRF (BW) 0.51 ± 0.11 0.36–0.69 54.2 ± 9.5 46–78
Peak cranially directed SRF (propulsive) (BW) 0.21 ± 0.08 0.14–0.43 59.8 ± 9.9 45–74
Peak medially directed SRF (BW) 0.13 ± 0.16 0.01–0.45 47.9 ± 25.3 5–86
Minimum SRF moment arm length shoulder (cm) 1.73 ± 0.44 1.01–2.37 49.0 ± 11.5 35–75
Minimum SRF moment arm length elbow (cm) 2.27 ± 0.25 1.91–2.66 79.5 ± 9.0 75–95
Maximum external moment shoulder (Nm) 0.10 ± 0.02 0.07–0.14 47.0 ± 22.9 20–80
Maximum external moment elbow (Nm) 0.11 ± 0.02 0.08–0.15 48.0 ± 14.9 20–75
Hindlimb
Peak vertically directed SRF (BW) 0.22 ± 0.04 0.15–0.28 26.4 ± 8.6 16–48
Peak cranially directed SRF (propulsive) (BW) 0.07 ± 0.03 0.05–0.12 31.6 ± 20.5 16–86
Peak medially directed SRF (BW) 0.05 ± 0.07 0.00–0.22 33.7 ± 19.4 11–64
Minimum SRF moment arm length hip (cm) 3.07 ± 0.48 2.20–3.53 34.5 ± 16.2 15–55
Minimum SRF moment arm length knee (cm) 2.18 ± 0.29 1.57–2.40 67.5 ± 13.2 55–80
Maximum external moment hip (Nm) 0.10 ± 0.02 0.09–0.15 31.0 ± 14.1 15–65
Maximum external moment knee (Nm) 0.08 ± 0.01 0.06–0.10 26.5 ± 5.8 15–35
SRFs normalized to body weight (BW)
Evol Biol
123
Author's personal copy

in T. scincoides (28 %) is slightly higher than the 10–18 %
estimated by Edwards (1977) for salamanders. The esti-
mated contribution to progression of the pelvic girdle in T.
scincoides is 17 %, while that in salamanders is again
10–18 % (Edwards 1977). The retraction of the humerus
and femur contributes the most to the progression of the
body during stance in both blue-tongued skinks and sala-
manders (56–62 % in salamanders, Edwards 1977). How-
ever, while we found a large difference in the contributions
of humeral retraction (32 %) and femoral retraction
(57 %), Edwards (1977) does not mention any differences
between the fore- and the hindlimbs.
3D forelimb kinematic data on sprawling locomotion is
largely absent from the published literature, but Karaka-
siliotis et al. (2012) recently published a detailed X-ray
motion analysis of the terrestrial locomotion of the Iberian
ribbed newt (Pleurodeles waltl) using the same equipment
we used. In comparison to the blue-tongued skink in our
study, the newts displayed greater amplitudes of femoral
LAR. Furthermore, unlike the belly-dragging skink they
also displayed greater amplitudes of humeral retraction
than femoral retraction (Karakasiliotis et al. 2012). Inter-
estingly, the observation that humeral LAR is much greater
than femoral LAR in T. scincoides coincides with the
observation that the forelimbs produce greater overall
forces and the SRF moment arm acting on the shoulder is
smallest at peak SRF (see ‘‘Discussion’’ below).
Data on femoral LAR and femoral retraction during burst
locomotion in the Florida scrub lizard (Sceloporus woodi)
show overall values comparable to T. scincoides (McElroy
et al. 2012). Tiger salamanders (Ambystoma tigrinum) also
displayed constant limb adduction and similar femoral
retraction amplitudes as T. scincoides (Sheffield and Blob
2011). White tegus (Tupinambis merianae), green iguanas
(Iguana iguana) and American alligators (Alligator missis-
sippiensis) all have even larger amplitudes of femoral pro-
and retraction (Blob and Biewener 2001; Sheffield et al.
2011). According to our quantitative data, the LAR of the
stylopodia contributes less to the progression of the body
than the rotation of the pectoral and pelvic girdles (Fig. 7)—
even in a skink characterized by comparatively little exter-
nally visible undulation of the vertebral column (Daan and
Belterman 1968; supplemental information). LAR is an
essential factor in overall limb kinematics and has effects on
the function of the lower arm and leg (Rewcastle 1983;
Landsmeer 1983). In our virtual experimental trials in which
stylopodial long-axis rotation was inactivated, autopodia
swept through the ground at all times.
SRF Patterns and Their Timing in Fore- and Hindlimbs
With regard to weight support distribution between the
fore- and the hindlimbs, geckos (Hemidactylus garnotii)
studied by Chen and coworkers (Chen et al. 2006) expe-
rienced greater vertical forces in their forelimbs than in
their hindlimbs. The running gait used by these geckos
makes the data in question difficult to compare to this
study, but greater forelimb support was also evident in T.
scincoides. Walking salamanders studied by Kawano and
Blob (2013) carried around 0.5 BW on their forelimbs
(Fig. 1 in Kawano and Blob 2013)—a figure similar to that
found in T. scincoides. However, the Fz exerted by a single
tiger salamander hindlimb also peaked at *0.5 BW
(Sheffield and Blob 2011; Kawano and Blob 2013). Thus,
support in salamanders is distributed approximately
equally between the fore- and the hindlimbs. In contrast,
the forelimbs of the skink carried more than double the
load of the isolated hindlimbs (which bore just *0.2 BW
at peak Fz). In the slow trotting or lateral sequence gait of
salamanders, where the duty factors are high, the consid-
erable time spent with all four appendages in simultaneous
ground contact apparently permits a slight lifting of the
trunk off the ground. Other available hindlimb data per-
taining to ‘raised’ sprawlers also exceeds the peak Fz of the
skink’s hindlimbs in every case [gecko: *1 BW (Chen
et al. 2006); green iguana: *1 BW (Blob and Biewener
2001); white tegu: *0.45 BW (Sheffield et al. 2011)].
Although peak Fz occurs much earlier during the contact
phase in the hindlimb than the forelimb in T. scincoides, it
coincides in both cases with maximum flexion in the knee
or elbow (Fig. 4). In geckos, on the other hand, peak Fz
occurs earlier in the forelimb, and well within the first half
of contact (Chen et al. 2006). In tiger salamanders and
white tegus, as in T. scincoides, peak Fz occurs much
earlier during contact in the hindlimbs than it does in the
forelimbs (Sheffield and Blob 2011; Sheffield et al. 2011).
American alligators and green iguanas, in contrast, exert
peak Fz at approximately mid-stance (Blob and Biewener
2001; Reilly et al. 2005). In none of these species do
maximum knee flexion and peak Fz seem to coincide as in
the hindlimbs of T. scincoides. In species that do not have
constant ventral contact to the substrate, no clear pattern
exists, with peak Fz occurring before (tiger salamander,
white tegu; Sheffield and Blob 2011; Sheffield et al. 2011)
or after (green iguana, American alligator; Blob and
Biewener 2001; Reilly et al. 2005) maximum knee flexion.
However, in those species that exert Fz before or at max-
imum knee flexion, the femur is not yet retracted and still
points somewhat cranio-laterally (tiger salamander, white
tegu, blue-tongued skink). Vice versa, in those species that
exert peak Fz after maximum knee flexion (i.e., as the knee
is extending), the femur is already in a retracted position
and pointing somewhat caudo-laterally (American alliga-
tor, green iguana). More belly-dragging species need to be
analyzed to test whether a clearer pattern may be detectable
during this type of sprawling tetrapod locomotion.
Evol Biol
123
Author's personal copy

While geckos (Chen et al. 2006; Autumn et al. 2006)
and salamanders (Kawano and Blob 2013) both exerted a
braking impulse with the forelimbs, T. scincoides did not
brake (on average), but very slightly propelled the body in
the direction of movement with both fore- and hindlimbs
on a constant basis. As the animals were not observed to
accelerate, this propulsive impulse clearly just offsets the
constant braking effected by the belly as a consequence of
ventral contact. The hindlimbs of tiger salamanders and
white tegus were not found to exert braking impulses either
(Sheffield and Blob 2011; Sheffield et al. 2011; Kawano
and Blob 2013). In stark contrast to these non-belly-drag-
ging species, however, T. scincoides produces more pro-
pulsive impulse in the forelimbs than in the hindlimbs,
making it an exception among sprawling tetrapods,
according to the available data.
SRF Moment Arms and External Moments Acting
at Proximal Limb Joints
Reducing the moment arm of the SRF acting at a specific
joint by directing the SRF vector more closely to the joint
is, to some extent, a way of reducing the muscle moments
acting at the same joint (Biewener 1989; Full et al. 1991).
However, in multi-segmented bent limbs, simultaneously
minimizing the moment arms of the SRF vector at all joints
would be impossible and counterproductive. It would also
complicate joint control, because the closer the SRF vector
passes to a joint, the easier it switches sides, thus triggering
the need for net joint moments to switch between flexion
and extension. In T. scincoides the minimum moment arm
length of the SRF vector acting at the shoulder occurred at
the moment of peak Fz, but this did not apply to the SRF
moment arm acting at the hip (Fig. 4). Unlike T. scinco-
ides, geckos and salamanders seem not to direct the SRF
vector to pass close to the shoulder and thus experience
relatively short external moment arms at the elbow (Chen
et al. 2006; Kawano and Blob 2013). Given that a shorter
external moment arm in the elbow is partially offset by
higher peak vertical forces in ‘raised’ sprawlers, the mus-
cular force required at the elbow will not differ dramati-
cally between belly-dragging and ‘raised’ sprawlers
(Fig. 8). In contrast, ‘raised’ sprawlers seem to be able to
generate—thanks to strong pectoral muscles—a much lar-
ger adducting moment in the shoulder, especially given the
large external moment arm and larger peak vertical force
they exert compared to the belly-dragging T. scincoides.
The small external moment arm at the shoulder at peak
vertical force might, then, simply reflect the inability of the
pectoral muscles of T. scincoides to produce more ad-
ducting moment. In the hindlimbs, the SRF vector at peak
force is directed to pass more closely to the knee than the
hip in geckos (Chen et al. 2006), tiger salamanders
(Sheffield and Blob 2011), green iguanas (Blob and
Biewener 2001), white tegus (Sheffield et al. 2011), and the
blue-tongued skink.
Functional Consequences of the Adoption of Belly-
Dragging Locomotion in Early Tetrapod Evolution
The patchy fossil record near the crown-group node and the
apparently rapid radiation and diversification of tetrapods
in the Early Carboniferous (e.g., Smithson and Clack 2013)
mean caution is called for when attempting to reconstruct
the last common ancestor of crown-group tetrapods.
However, the notion of a belly-dragging phase is substan-
tiated by the considerable size of fossils such as Pederpes
(Clack 2002a), Whatcheeria (Lombard and Bolt 1995),
Tulerpeton (Lebedev and Coates 1995) and Ossinodus
(Warren and Turner 2004), and by low limb length to body
mass ratios (see Bakker 1971). The dramatic change in
functional role from pectoral appendages that drag the
body via in-phase crutching, as proposed for Devonian
stem tetrapods, to those that contribute to propulsion via
stepping, as proposed for crown-group node tetrapods, has
been pointed out by Kawano and Blob (2013). This reflects
the notion that the similarity in function between fore- and
hindlimbs was initially absent in Devonian tetrapods and
later evolved convergently (see Diogo et al. 2013). Anal-
yses of the locomotor mechanics in belly-dragging T.
scincoides reveal an intermediate character combination
that potentially affords insights into a transformational
state of early tetrapods close to the crown-group node.
According to Pierce et al. (2012), the forelimbs played a
crucial role in terrestrial locomotion much earlier than the
hindlimbs during the fin-to-limb transition. In this regard it
is interesting to note that T. scincoides generate more
propulsive thrust in their forelimbs than in their hindlimbs.
Kawano and Blob (2013) suggest that while propulsion was
‘‘hindlimb-driven’’, the forelimbs of early tetrapods with
heavy tails probably still made a substantial contribution.
Our data demonstrates that sprawling locomotion is not
always ‘‘hindlimb-driven’’.
The relatively large and heavy Devonian stem tetrapods
usually had their weight supported by water and were
likely to be lacking the strong shoulder and hip adductors
and knee and elbow extensors needed for non-belly-drag-
ging sprawling locomotion (Fig. 1). An explanation for the
ability of ‘raised’ sprawlers to produce large adducting
moments in the shoulder might lie in the more vertical
orientation of the SRF vector documented in ‘raised’
sprawling tetrapod forelimbs compared to belly-dragging
T. scincoides (Chen et al. 2006; Kawano and Blob 2013;
this study). In T. scincoides the vertical component of the
SRF reaches a maximum of only about 50 % of body
weight in the forelimbs and considerably less in the
Evol Biol
123
Author's personal copy

hindlimbs, leading to smaller overall magnitudes of the
SRF vector. With similar mechanics as documented here
for T. scincoides, a belly-dragging early tetrapod would
have been able to move on land with relatively weakly
developed pectoral muscles (Fig. 8). Having body weight
partially supported by direct substrate contact can be con-
sidered an intermediate condition between having it sup-
ported by water and having to raise it off the ground against
gravity. In the light of the considerable size of known early
tetrapods and the relationship between cross-sectional area
and muscle volume, we suggest that belly-dragging meant
that smaller, and thus initially more attainable, anti-gravity
muscles were needed to achieve terrestrial locomotion.
A Scenario of Tetrapod Locomotor Evolution During
the Early Carboniferous
Accepting the possibility of a belly-dragging phase in early
tetrapod evolution during the Early Carboniferous, the
study of the extant belly-dragging T. scincoides or other
belly-dragging tetrapods (e.g., gharials, giant salamanders)
has the potential to help conceptualize an evolutionary
scenario for the transition of early tetrapod locomotor
mechanics. The transition starts with locomotion marked
by ventral contact and synchronous forelimb action-gen-
erated propulsion—the form of locomotion presumably
embodied by Ichthyostega (and likely other tetrapods) of
the Devonian (Pierce et al. 2012, 2013a, b; Kawano and
Blob 2013). Later, the salamander-like ‘raised’ terrestrial
locomotion inferred for diverse Early- and mid-Carbonif-
erous terrestrial tetrapods evolved (e.g., Clack 2002b),
marked by symmetrical gaits, hindlimb propulsion and full
support of body weight by the limbs. The present study
suggests a link between belly-dragging and forelimb
dominance in weight bearing and provision of propulsion.
We thus propose that during the hypothesized belly-drag-
ging phase of tetrapod locomotor evolution, locomotion
was still forelimb-driven with regard to provision of pro-
pulsion, initially with little assistance from the hindlimbs.
Dragging substantial parts of the body along increases the
propulsive role of the limbs as a means of countering the
net braking effect of the body (Willey et al. 2004). After
this intermediate, forelimb-dominated belly-dragging
phase, intermittent and later permanent lifting of the trunk
(effected by the action of the shoulder adductors and elbow
extensors in the forelimb and the hip adductors and knee
extensors in the hindlimb) evolved, while the role of the
hindlimbs in weight-bearing and the provision of propul-
sion gradually increased. Salamander-like terrestrial loco-
motion was finally achieved either by an increase in the
relative mass of muscles with an anti-gravity role and/or an
overall reduction in body size (documented in the fossil
record by the first appearance of very small-sized animals
such as Casineria in the Mid-Visean; Paton et al. 1999, for
example). Hence, in the concept of evolutionary transition
proposed herein, the locomotor mechanics of initially
belly-dragging Devonian tetrapods were gradually modi-
fied, as opposed to the de-novo appearance of salamander-
like terrestrial locomotion in the alternative scenario.
Importantly, our hypothesis predicts that the mode of
propulsion generation, i.e., the overall kinematic pattern,
precedes the evolution of permanent body weight support
by the limbs. That is, we suggest, in accordance with
accepted accounts, that limbs and initial limb kinematics
likely evolved to serve purposes in water (Clack 2002b;
Pierce et al. 2012), with subsequent smaller modifications
of limb mechanics representing an exaptation for terrestrial
locomotion.
This evolutionary scenario based on a biomechanical
analysis of an extant lizard needs to be shown to be in
agreement with biomechanical inferences about the ter-
restrial locomotor capabilities of specific fossil specimens.
Dynamic musculoskeletal modeling of the limbs in belly-
dragging and non-belly-dragging extant sprawlers could be
used to anchor musculoskeletal models of available
Fig. 8 Functional consequences of a non-belly-dragging and b belly-
dragging locomotion. The nearly vertical orientation of the SRF
vector (orange arrows) documented in the forelimbs of non-belly-
dragging sprawling tetrapods (Chen et al. 2006; Kawano and Blob
2013) results in a short moment arm (R) at the elbow, but long R at the
shoulder and a high external moment. The shortened R at peak Fz and
low overall SRFs during belly-dragging locomotion documented here
result in a moderate external moment. Please note that the combi-
nation of a 50 % reduction in SRF magnitude and a 50 % reduction in
R will result in just 25 % of the net joint torque needed to maintain
equilibrium (Color figure online)
Evol Biol
123
Author's personal copy

well-preserved fossil specimens (e.g., Tulerpeton, Peder-
pes or Ossinodus) close to the crown-group tetrapod node.
However, no stem group specimen can be claimed to
represent an immediate ancestor: they are offshoots from
the lineage leading to the last common ancestor of modern
tetrapods. Hopefully, future findings of ichnofossils and
fossil specimens that further populate Romer’s Gap (see
Smithson et al. 2012) will help to narrow down possible
scenarios of locomotor evolution in early tetrapods.
Acknowledgments The authors would like to thank Rommy Pet-
ersohn and Ingrid Weiss for technical help during X-ray motion
analyses. Vivian R. Allen and Brandon M. Kilbourne provided
helpful comments on earlier versions of the manuscript. Professional
language polishing by Lucy Cathrow improved the final version of the
manuscript. CT scans were recorded at the Institut fur Interventionelle
und Diagnostische Radiologie at the University Hospital of the
Friedrich-Schiller-Universitat Jena. This project is financed by the
Volkswagen Foundation (Grant No.: AZ85857 to JAN and MSF).
References
Ahlberg, P. E., & Milner, A. R. (1994). The origin and early
diversification of tetrapods. Nature, 368(6471), 507–514.
Andrada, E., Nyakatura, J. A., Bergmann, F., & Blickhan, R. (2013).
Adjustments of global and local properties of the hindlimb
during the terrestrial locomotion of the common quail (Coturnix
coturnix). Journal of Experimental Biology, 216, 3906–3916.
Ashley-Ross, M. A. (1994a). Hind limb kinematics during terrestrial
locomotion in a salamander (Dicamptodon tenebrosus). Journal
of Experimental Biology, 193, 255–283.
Ashley-Ross, M. A. (1994b). Metamorphic and speed effects on hind
limb kinematics during terrestrial locomotion in the salamander
Dicamptodon tenebrosus. Journal of Experimental Biology, 193,
285–305.
Ashley-Ross, M. A., & Bechtel, B. F. (2004). Kinematics of the
transition between aquatic and terrestrial locomotion in the newt
Taricha torosa. Journal of Experimental Biology, 207(3),
461–474.
Ashley-Ross, M. A., Hsieh, S. T., Gibb, A. C., & Blob, R. W. (2013).
Vertebrate land invasions—Past, present, and future: an intro-
duction to the symposium. Integrative and Comparative Biology.
doi:10.1093/icb/ict048.
Autumn, K., Hsieh, S. T., Dudek, D. M., Chen, J., Chitaphan, C., &
Full, R. J. (2006). Dynamics of geckos running vertically.
Journal of Experimental Biology, 209(2), 260–272.
Bakker, R. T. (1971). Dinosaur physiology and the origin of
mammals. Evolution, 25, 636–658.
Barclay, O. R. (1946). The mechanics of amphibian locomotion.
Journal of Experimental Biology, 23(2), 177–203.
Biewener, A. A. (1989). Scaling body support in mammals: Limb
posture and muscle mechanics. Science, 245(4913), 45–48.
Biewener, A. A., & Full, R. J. (1992). Force platform and kinematic
analysis. In A. A. Biewener (Ed.), Biomechanics (structures and
systems): A practical approach (pp. 45–73). New York: Oxford
University Press.
Blob, R. W., & Biewener, A. A. (2001). Mechanics of limb bone
loading during terrestrial locomotion in the green iguana (Iguana
iguana) and American alligator (Alligator mississippiensis).
Journal of Experimental Biology, 204(6), 1099–1122.
Brainerd, E. L., Baier, D. B., Gatesy, S. M., Hedrick, T. L., Metzger,
K. A., Gilbert, S. L., et al. (2010). X-ray reconstruction of
moving morphology (XROMM): Precision, accuracy and appli-
cations in comparative biomechanics research. Journal of
Experimental Zoology Part A: Ecological Genetics and Physi-
ology, 313(5), 262–279.
Carroll, R. L., Bossy, K. A., Milner, A. C., Andrews, S. M., &
Wellstead, C. F. (1998). Handbuch der Palaoherpetologie (Vol.
1). Pfeil: Lepospondyli. Munich.
Chen, J. J., Peattie, A. M., Autumn, K., & Full, R. J. (2006).
Differential leg function in a sprawled-posture quadrupedal
trotter. Journal of Experimental Biology, 209(2), 249–259.
Clack, J. A. (1997). Devonian tetrapod trackways and trackmakers; a
review of the fossils and footprints. Palaeogeography, Palaeo-
climatology, Palaeoecology, 130(1), 227–250.
Clack, J. A. (2002a). An early tetrapod from ‘Romer’s Gap’. Nature,
418(6893), 72–76.
Clack, J. A. (2002b). Gaining ground: The origin and evolution of
tetrapods. Bloomington: Indiana University Press.
Coates, M. I., & Clack, J. A. (1995). Romer’s gap—Tetrapod origins
and terrestriality. In M. Arsenault, H. Leliere, & P. Janvier
(Eds.), Studies on early vertebrates (Vol. 17, pp. 373–388).
Paris: Bulletin du Museum National d’Hisoire Naturelle.
Curth, S., Fischer, M. S., & Nyakatura, J. A. (in press). Ichnology of
an extant belly-dragging lizard—Analogies to early reptile
locomotion? Ichnos.
Daan, S., & Belterman, T. (1968). Lateral bending in locomotion of
some lower tetrapods. In Proceedings of the Koninklijke
Nederlandse Akademie van Wetenschappen. Series C: Biological
and Medical Sciences (Vol. 71, pp. 245–266).
Diogo, R., Linde-Medina, M., Abdala, V., & Ashley-Ross, M. A.
(2013). New, puzzling insights from comparative myological
studies on the old and unsolved forelimb/hindlimb enigma.
Biological Reviews, 88(1), 196–214.
Edwards, J. L. (1977). The evolution of terrestrial locomotion. In M.
K. Hecht, P. C. Goody, & B. M. Hecht (Eds.), Major patterns in
vertebrate evolution. New York: Plenum Press.
Frolich, L. M., & Biewener, A. A. (1992). Kinematic and electro-
myographic analysis of the functional role of the body axis
during terrestrial and aquatic locomotion in the salamander
Ambystoma tigrinum. Journal of Experimental Biology, 162(1),
107–130.
Full, R. J., Blickhan, R., & Ting, L. H. (1991). Leg design in hexapedal
runners. Journal of Experimental Biology, 158(1), 369–390.
Gans, C., & de Gueldre, G. (1992). Striated muscle: Physiology and
functional morphology. In M. E. Feder & W. W. Burggren
(Eds.), Environmental physiology of the amphibians. Chicago:
University of Chicago Press.
Gatesy, S. M., Baier, D. B., Jenkins, F. A., & Dial, K. P. (2010). Scientific
rotoscoping: A morphology-based method of 3-D motion analysis
and visualization. Journal of Experimental Zoology Part A:
Ecological Genetics and Physiology, 313(5), 244–261.
Gray, S. J. (1968). Animal locomotion. London: Weidenfeld & Nicolson.
Hildebrand, M. (1966). Analysis of the symmetrical gaits of tetrapods.
Folia Biotheoretica, 6, 9–22.
Horner, P. (2000). Der Nordliche Blauzungenskink Tiliqua scincoides
intermedia (Mitchell, 1955). In A. Hauschild, K. Henle, R. Hitz,
G. M. Shea, & H. Werning (Eds.), Blauzungenskinke: Beitrage
zu Tiliqua und Cyclodomorphus (pp. 161–168). Munster: Natur
und Tier-Verlag.
Hutchinson, J. R., & Garcia, M. (2002). Tyrannosaurus was not a fast
runner. Nature, 415(6875), 1018–1021.
Janis, C. M., & Keller, J. C. (2001). Modes of ventilation in early
tetrapods: Costal aspiration as a key feature of amniotes. Acta
Palaeontologica Polonica, 46(2), 137–170.
Jenkins, F. A., & Goslow, G. E. (1983). The functional anatomy of the
shoulder of the savannah monitor lizard (Varanus exanthemat-
icus). Journal of Morphology, 175(2), 195–216.
Evol Biol
123
Author's personal copy

Karakasiliotis, K., Schilling, N., Cabelguen, J. M., & Ijspeert, A. J.
(2012). Where are we in understanding salamander locomotion:
Biological and robotic perspectives on kinematics. Biological
Cybernetics, 107(5), 529–544.
Kawano, S. M., & Blob, R. W. (2013). Propulsive forces of
mudskipper fins and salamander limbs during terrestrial loco-
motion: Implications for the invasion of land. Integrative and
Comparative Biology, 53(2), 283–294.
Kubo, T. (2010). Extant lizard tracks: Variation and implications for
paleoichnology. Ichnos, 17(3), 187–196.
Landsmeer, J. M. (1983). The mechanism of forearm rotation in
Varanus exanthematicus. Journal of Morphology, 175(2),
119–130.
Lebedev, O. A., & Coates, M. I. (1995). The postcranial skeleton of
the Devonian tetrapod Tulerpeton curtum Lebedev. Zoological
Journal of the Linnean Society, 114(3), 307–348.
Lombard, R. E., & Bolt, J. R. (1995). A new primitive tetrapod,
Whatcheeria deltae, from the Lower Carboniferous of Iowa.
Palaeontology, 38(3), 471–494.
McElroy, E. J., Archambeau, K. L., & McBrayer, L. D. (2012). The
correlation between locomotor performance and hindlimb kine-
matics during burst locomotion in the Florida scrub lizard,
Sceloporus woodi. The Journal of Experimental Biology, 215(3),
442–453.
Michilsens, F., Aerts, P., Van Damme, R., & D’Aout, K. (2009).
Scaling of plantar pressures in mammals. Journal of Zoology,
279(3), 236–242.
Moch, J. G., & Senter, P. (2011). Vestigial structures in the
appendicular skeletons of eight African skink species (Squamata,
Scincidae). Journal of Zoology, 285(4), 274–280.
Nyakatura, J. A., & Fischer, M. S. (2010a). Three-dimensional
kinematic analysis of the pectoral girdle during upside-down
locomotion of two-toed sloths (Choloepus didactylus, Linne
1758). Front Zool, 7(1), 21.
Nyakatura, J. A., & Fischer, M. S. (2010b). Functional morphology
and three-dimensional kinematics of the thoraco-lumbar region
of the spine of the two-toed sloth. The Journal of Experimental
Biology, 213(24), 4278–4290.
Nyakatura, J. A., Petrovitch, A., & Fischer, M. S. (2010). Limb
kinematics during locomotion in the two-toed sloth (Choloepus
didactylus, Xenarthra) and its implications for the evolution of
the sloth locomotor apparatus. Zoology, 113(4), 221–234.
Pace, C. M., & Gibb, A. C. (2009). Mudskipper pectoral fin
kinematics in aquatic and terrestrial environments. Journal of
Experimental Biology, 212(14), 2279–2286.
Paton, R. L., Smithson, T. R., & Clack, J. A. (1999). An amniote-like
skeleton from the early Carboniferous of Scotland. Nature,
398(6727), 508–513.
Pierce, S. E., Ahlberg, P. E., Hutchinson, J. R., Molnar, J. L.,
Sanchez, S., Tafforeau, P., et al. (2013a). Vertebral architecture
in the earliest stem tetrapods. Nature, 494(7436), 226–229.
Pierce, S. E., Clack, J. A., & Hutchinson, J. R. (2012). Three-
dimensional limb joint mobility in the early tetrapod Ichthyo-
stega. Nature, 486(7404), 523–526.
Pierce, S. E., Hutchinson, J. R., & Clack, J. A. (2013b). Historical
perspectives on the evolution of tetrapodomorph movement.
Integrative and Comparative Biology. doi:10.1093/icb/ict022.
Pridmore, P. A. (1994). Submerged walking in the Epaulette shark
Hemiscyllium ocellatum (Hemiscyllidae) and its implications for
locomotion in rhipidistian fisches and early tetrapods. Zoology,
98(4), 278–297.
Reilly, S. M., McElroy, E. J., Odum, R. A., & Hornyak, V. A. (2006).
Tuataras and salamanders show that walking and running
mechanics are ancient features of tetrapod locomotion. Proceed-
ings of the Royal Society B: Biological Sciences, 273(1593),
1563–1568.
Reilly, S. M., Willey, J. S., Biknevicius, A. R., & Blob, R. W. (2005).
Hindlimb function in the alligator: Integrating movements,
motor patterns, ground reaction forces and bone strain of
terrestrial locomotion. Journal of Experimental Biology, 208(6),
993–1009.
Rewcastle, S. C. (1983). Fundamental adaptations in the lacertilian
hind limb: A partial analysis of the sprawling limb posture and
gait. Copeia, 1983(2), 476–487.
Schaller, N. U., D’Aout, K., Villa, R., Herkner, B., & Aerts, P. (2011).
Toe function and dynamic pressure distribution in ostrich
locomotion. The Journal of experimental biology, 214(7),
1123–1130.
Sheffield, K. M., & Blob, R. W. (2011). Loading mechanics of the
femur in tiger salamanders (Ambystoma tigrinum) during
terrestrial locomotion. The Journal of Experimental Biology,
214(15), 2603–2615.
Sheffield, K. M., Butcher, M. T., Shugart, S. K., Gander, J. C., &
Blob, R. W. (2011). Locomotor loading mechanics in the
hindlimbs of tegu lizards (Tupinambis merianae): Comparative
and evolutionary implications. The Journal of Experimental
Biology, 214(15), 2616–2630.
Smithson, T. R., Carroll, R. L., Panchen, A. L., & Andrews, S. M.
(1994). Westlothiana lizziae from the Visean of East Kirkton,
West Lothian, Scotland. Transactions of the Royal Society of
Edinburgh, Earth Sciences, 84, 417–431.
Smithson, T. R., & Clack, J. A. (2013). Tetrapod appendicular
skeletal elements from the Early Carboniferous of Scotland.
Comptes Rendus Palevol, 12(7), 405–417.
Smithson, T. R., Wood, S. P., Marshall, J. E., & Clack, J. A. (2012).
Earliest Carboniferous tetrapod and arthropod faunas from
Scotland populate Romer’s Gap. Proceedings of the National
Academy of Sciences, 109(12), 4532–4537.
Sullivan, C. S. (2007). Function and evolution of the hind limb in
Triassic archosaurian reptiles. Dissertation, Harvard University.
ProQuest.
Warren, A., & Turner, S. (2004). The first stem tetrapod from the
Lower Carboniferous of Gondwana. Palaeontology, 47(1),
151–184.
Warren, J. W., & Wakefield, N. A. (1972). Trackways of tetrapod
vertebrates from the Upper Devonian of Victoria, Australia.
Nature, 238(5365), 469–470.
Willey, J. S., Biknevicius, A. R., Reilly, S. M., & Earls, K. D. (2004).
The tale of the tail: Limb function and locomotor mechanics in
Alligator mississippiensis. Journal of Experimental Biology,
207(3), 553–563.
Evol Biol
123
Author's personal copy




















