
“Descripción anatómica e histológica de la concha nácar Pteria sterna, Mollusca: Pteriidae
Upload
independentCategory
view
2download
0
Aquaculture 314 (2011) 215–220
Contents lists available at ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r.com/ locate /aqua-on l ine
Amino acid mobilization and growth of juvenile Octopus maya (Mollusca:Cephalopoda) under inanition and re-feeding
Arturo George-Zamora a, Maria-Teresa Viana b, Sergio Rodríguez c, Gabriela Espinoza c, Carlos Rosas d,⁎a Posgrado en Ciencias del Mar y Limnología (PCMyL), Universidad Nacional Autónoma de México (UNAM), Puerto de Abrigo S/N, Sisal, Yucatán, Mexicob Instituto de Investigaciones Oceanológicas (IIO), Universidad Autónoma de Baja California (UABC), Ensenada, Baja California, Mexicoc Facultad de Química, Unidad de Química-Sisal, Universidad Nacional Autónoma de México (UNAM), Puerto de Abrigo S/N, Sisal, Yucatán, Mexicod Unidad Multidisciplinaria de Docencia e Investigación (UMDI), Facultad de Ciencias, Universidad Nacional Autónoma de México (UNAM), Puerto de Abrigo S/N, Sisal, Yucatán, Mexico
⁎ Corresponding author. Tel.: +52 988 9120147/49; fE-mail address: [email protected] (C. Rosas).
0044-8486/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.aquaculture.2011.02.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 22 September 2010Received in revised form 15 February 2011Accepted 15 February 2011Available online 21 February 2011
Keywords:Amino acidsGrowthCephalopodaOctopus mayaFastingMéxico
Octopus maya is an endemic cephalopod from the Yucatán Peninsula in Mexico, with interest to develop theircommercial culture. Like all cephalopods, protein is an important nutrient both for growing and energeticmetabolism. This condition results in a high demand for protein and specifically for certain amino acids (AAs).In this study it is examined the effects of inanition and re-feeding on growth and AA content in soft tissue todetect the amino acid (AA) mobilization to identify the principal metabolic reserves in juvenile O. maya. After25 days re-feeding of all starved groups, the octopuses were unable to reach the similar weight as the controlgroup. However, SGR of some groups were greater than that of the controls, although the differences were notsignificant due to variability in the data. Therefore, it is assumed that the juveniles of O. maya would need alonger period of time to reach the control group. It is therefore demonstrated that juveniles of O. maya have awide plasticity to tolerate, at least 10 days of food deprivation without any apparently physiological damage.Moreover, during inanition the juveniles of O. maya used preferentially Thr, Phe, Ile, Ala, Glu and Ser,suggesting a strong mobilization of both essential and non essential AA to maintain the homeostasis. Prove ofthat is that survival of the animals during fasting and re-feeding period was not affected by treatments.
ax: +52 988 9120020.
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
The octopus Octopus maya (Voss and Solís-Ramírez, 1966) is anendemic cephalopod from theYucatan Peninsula inMexico (CONAPESCA,2008) being one of the most important resources in the fishery sector.Earlier studies have demonstrated that O. maya has a high potential foraquaculture due to their well adaptability to captivity conditions (VanHeukelem, 1977; Boletzky and Hanlon, 1983; Hanlon and Forsythe1985; DeRusha et al., 1989) and direct development from hatchingwithout a larval transition (Boletzky, 1974). Moreover, O. maya showslarger eggs than those observed in other cephalopods (up to 17 mm).They are able to accept dead preys or formulated diets even during thefirst life cycle stadiums (Aguila et al., 2007), resulting in high growthrates (up to 8% wet body weight, wBW, per day) as a result of highfeeding rates (FR) and food conversion efficiencies (FC) (VanHeukelem,1983; Hanlon and Forsythe, 1985; Domingues et al., 2007; Rosas et al.,2007).
Dependence on natural diets together with a long larval period hasbeen the bottleneck in the cephalopod aquaculture (Domingues,1999; Lee, 1994; Iglesias et al., 2000; Iglesias et al., 2007), due to the
high cost associated to obtain the juveniles for the growth-out phase(Nabhitabhata, 1995). Therefore, several formulated diets have beentested in O. maya, efforts that have resulted in moderate growth rates,below to those achieved with natural diets (Aguila et al., 2007;Domingues et al., 2007; Rosas et al., 2007; Rosas et al., 2008). Similarresults were found in cuttlefish (Castro et al., 1993; Castro and Lee,1994) concluding that protein quality is an important factor to gethigh digestibility coefficients and better growth rates, being necessaryto generate more information on the amino acid requirements topromote higher growth rates. Le Bihan et al., (2006) found thatcuttlefish fed with surimi from the shrimp Crangon crangon soaked infish silage resulted in better growth and conversion rates than thosefed a similar diet without the fish silage. The difference between bothdiets was the peptides and AA present in the fish silage. However, it isnot clear if the peptides and AA promoted a better assimilation or dueto the solubility inherent to the soluble protein resulted in a higheringestion rate due to a better feed attractability and palatability.
Like all cephalopods, O. maya is a carnivorous species and proteinis an important nutrient for tissue accretion and energy source(Segawa and Hanlon, 1988; Rosas et al., 2007), therefore, cephalopodsrequire high amounts of protein and amino acids (AAs) for optimumgrowth (Lee, 1994; Domingues et al., 2005; Solorzano et al., 2009).However, at present the protein and amino acid requirements havenot been determined for O. maya.

216 A. George-Zamora et al. / Aquaculture 314 (2011) 215–220
In rearing conditions octopuses are able to survive long periodswithout food due to several stress factors like system failures (pumpingandpower energyproblems) or density stress (Domingues et al., 2010).After a period of food deprivation organisms are able to catch up theirweight by increasing their growth rates as a compensatory growth.Howeverwhen food deprivation is for longer periods the organisms areunable to catch up their weight compared to well feed siblings, callingthis physiological phenomenon as the point of no return. Thereforewhen culturing organisms that undergo long periods of food depriva-tion it is important to know those limits to be able to catch up their lostweight without fatal consequences. Taking into consideration thatoctopuses have a low lipid reserves it is possible to think that duringfasting condition octopuses will mobilize amino acids as the principalsource of energy in attempt to maintain the homeostasis (Vidal et al.,2002; Villanueva et al., 2004; Iglesias et al., 2006; Vidal et al., 2006;Grigoriou and Richardson, 2009; Solorzano et al., 2009). Recently,Villanueva et al., (2004) determined that Leu, Lys and Arg representalmost half of the essential amino acids (EAAs) required by Octopusvulgaris, Sepia officinalis and Loligo vulgaris during early life stages.According to Akagi and Ohmori (2004) Thr could be the optimumsubstrate for D-lactate formation, suggesting that the principalbiochemical pathway to obtain energy in cephalopods couldbe throughgluconeogenesis, and therefore Thr may be limiting in the diet fromthese species as indicated by D'Mello (2003). Domingues et al. (2007)found similar results for the juvenile O. maya.
In the present study it was evaluated the inanition and re-feedingeffects on the overall performance to detect the amino acid (AA)mobilization and overall performance for the juvenile O. maya.
2. Materials and methods
2.1. Broodstock collection and rearing of juvenile octopuses
Wild females from O. maya were caught on the coastal area fromthe Yucatan Peninsula (21° 9′ 55″ N, 90° 1′ 50″ W) using artisan lineswith blue crabs Callinectes spp. as bait. Females were transported in120 L circular tanks containing seawater to the UMDI-UNAMlaboratory, situated at 300 m inland. In the laboratory, the femaleswere also maintained in 250 L black tanks until the eggs were layed(Moguel et al., 2010). Hatchlings were fed for 15–20 days with livingArtemia adults and crab paste until they passed 0.5 g living weight(Rosas et al., 2008; Avila-Poveda et al., 2009; Moguel et al., 2010).
2.2. Feeding acclimation of post-hatching
Juvenile O. maya of 32 days post-hatching (DPH) were weighed(wet body weight (wBW) mean value of 0.63±0.10 SD g; N=120octopuses). Juveniles, dried with a paper towel, were individuallyweighed in an electronic balance (±0.001 g), randomly individualized(1 L tanks), and distributed in six experimental groups (N=20animals per group). Juveniles were fed a single diet consisting infresh crab meat mixed with 5% natural gelatin, fed at 30% of wBW(Rosas et al., 2008; Quintana et al., 2010) twice a day at 0900 and1700 h. After 12 days all octopuses from the six groups wereindividually weighed again (wBW; mean value of 0.90±0.21 SD g;N=65 octopuses).
2.3. Inanition and re-feeding treatments
To determine the effect of food deprivation on the overallperformance of juvenile O. maya, six groups (N=20 per group)were deprived of food for 2, 4, 6, 8 and 10 days, while the sixth groupwas maintained with food as a control under a similar protocol asduring the acclimation period. Inanition began at the same time for alloctopuses from five experimental groups. At the end of each fastingperiod all octopuses from the six groups were weighed. After the
inanition period, the octopuses from each treatment were re-feedingfor 25 days,with the samediet and under similar protocol as explainedabove. The octopuses were weighed at the end of re-feeding period.
2.4. Growth rate
Growth rate (g day−1) was determined as the difference betweenthe initial and final weight for octopuses during acclimation period(days 0 and 12), at the endof each fastingperiods (2, 4, 6, 8 and 10 days)and latest day of re-feeding period.
Specific growth rate (SGR, %day−1) was determined as:
SGR = lnW2−lnW1ð Þ= t½ � � 100
where W2 and W1 are the final and initial wet weights of the octopus(obtained during acclimation period, fasting periods or re-feedingperiod), ln is the natural logarithm, and t the duration of each timeperiod (days).
2.5. Amino acid analysis
The amino acid content (g AA/100 g protein) of individual samplesfrom juvenile O. maya arms (N=3 octopus per treatment) from theacclimation period (control on day 12), and after the inanition period(on the 4 and 8 days food deprivation)was determined. Defatted tissuesamples (20 mg) were hydrolyzed with 200 μL of 6 N HCl and 0.06%phenol in a closed vial and heated to 110 °C for 24 h. Amino acid profileswere determined following Waters AccQ-Tag™ procedure as follows:(1)Hydrolyzed samplesweredried in a termicmonoblock at 60 °Cwithnitrogen and rehydrated with 1 mL water HPLC grade. (2) Sampleswere then filtered (0.45 μm) and maintained at −20 °C until used.(3) Samples were derivatized using the Waters system AccQ-Tag™.(4) Samples were chromatographed through a reverse phase column(3.9×150 mm) 4 μm Nova Pak™ C-18, using the water-acetonitrilegradient recommended by the Waters AccQ-Tag™ system (Milford,MA, USA), in a Waters™ HPLC and a fluorescence detector (excitationand emissionwavelength; 250 and 395 nm, respectively). (6) Analyseswere conducted at a constant temperature of 39 °C. (7) HPLC signalcalibration and standard curves were obtained by using an amino acidstandard solution at three different concentrations containing from18.75 to 150 pmol of each amino acid. Taking into account thatMet andCys were partially destroyed by acid hydrolysis, the results of bothamino acids were taken with caution.
2.6. Statistical analysis
A one way analysis of variance was used to known the effect offasting period on wet weight and growth rate (SGR, %day−1), andwhen applied, amultiple comparison of means (Tuckey test) was usedto known differences between groups. A significance level of Pb0.05was established. All statistical analyses were performed using theStatistica@ program (Version 6.1).
3. Result
3.1. Acclimation period
No differences were detected among during the acclimationperiod. Mean values of 0.63±0.02 g, 0.90±0.04 g and 2.81±0.4%day−1 were obtained for initial, final and specific growth rate (SGR)respectively, during acclimation period, (Table 1; Figs. 1 and 2).
3.2. Inanition effect on growth
After the food deprivation period, the final weight showeddifferences between treatments with 9% of wet weight lost in animals
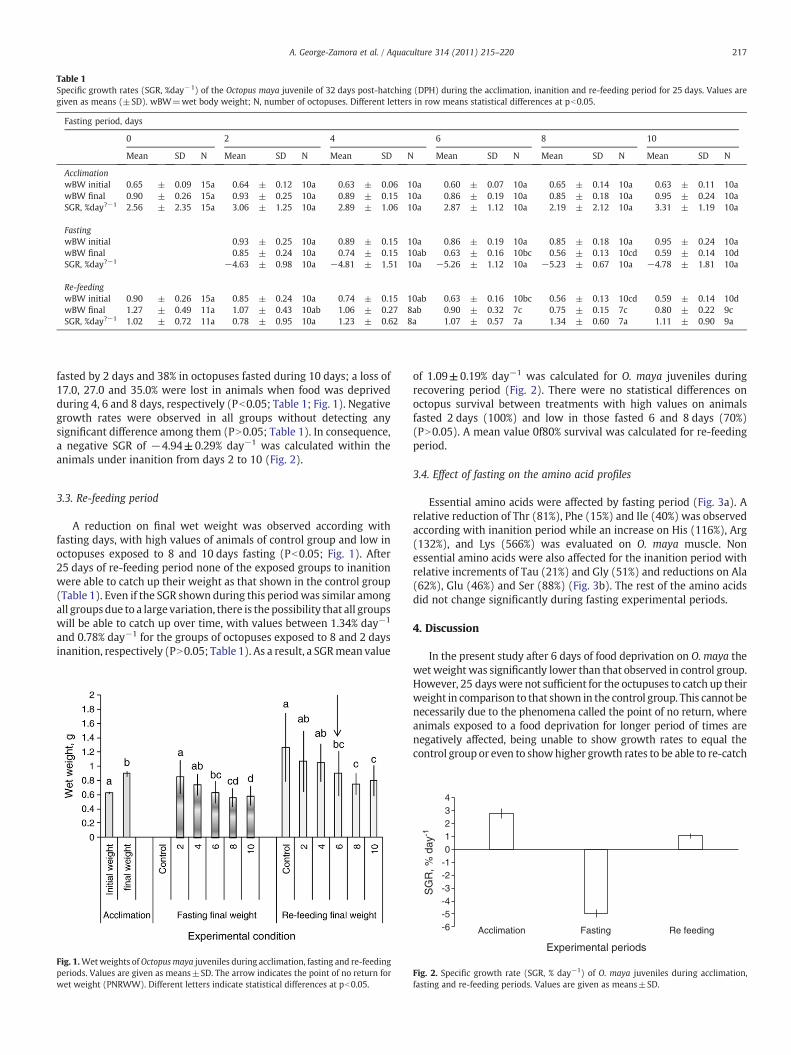
Table 1Specific growth rates (SGR, %day−1) of the Octopus maya juvenile of 32 days post-hatching (DPH) during the acclimation, inanition and re-feeding period for 25 days. Values aregiven as means (±SD). wBW=wet body weight; N, number of octopuses. Different letters in row means statistical differences at pb0.05.
Fasting period, days
0 2 4 6 8 10
Mean SD N Mean SD N Mean SD N Mean SD N Mean SD N Mean SD N
AcclimationwBW initial 0.65 ± 0.09 15a 0.64 ± 0.12 10a 0.63 ± 0.06 10a 0.60 ± 0.07 10a 0.65 ± 0.14 10a 0.63 ± 0.11 10awBW final 0.90 ± 0.26 15a 0.93 ± 0.25 10a 0.89 ± 0.15 10a 0.86 ± 0.19 10a 0.85 ± 0.18 10a 0.95 ± 0.24 10aSGR, %day?−1 2.56 ± 2.35 15a 3.06 ± 1.25 10a 2.89 ± 1.06 10a 2.87 ± 1.12 10a 2.19 ± 2.12 10a 3.31 ± 1.19 10a
FastingwBW initial 0.93 ± 0.25 10a 0.89 ± 0.15 10a 0.86 ± 0.19 10a 0.85 ± 0.18 10a 0.95 ± 0.24 10awBW final 0.85 ± 0.24 10a 0.74 ± 0.15 10ab 0.63 ± 0.16 10bc 0.56 ± 0.13 10cd 0.59 ± 0.14 10dSGR, %day?−1 −4.63 ± 0.98 10a −4.81 ± 1.51 10a −5.26 ± 1.12 10a −5.23 ± 0.67 10a −4.78 ± 1.81 10a
Re-feedingwBW initial 0.90 ± 0.26 15a 0.85 ± 0.24 10a 0.74 ± 0.15 10ab 0.63 ± 0.16 10bc 0.56 ± 0.13 10cd 0.59 ± 0.14 10dwBW final 1.27 ± 0.49 11a 1.07 ± 0.43 10ab 1.06 ± 0.27 8ab 0.90 ± 0.32 7c 0.75 ± 0.15 7c 0.80 ± 0.22 9cSGR, %day?−1 1.02 ± 0.72 11a 0.78 ± 0.95 10a 1.23 ± 0.62 8a 1.07 ± 0.57 7a 1.34 ± 0.60 7a 1.11 ± 0.90 9a
217A. George-Zamora et al. / Aquaculture 314 (2011) 215–220
fasted by 2 days and 38% in octopuses fasted during 10 days; a loss of17.0, 27.0 and 35.0% were lost in animals when food was deprivedduring 4, 6 and 8 days, respectively (Pb0.05; Table 1; Fig. 1). Negativegrowth rates were observed in all groups without detecting anysignificant difference among them (PN0.05; Table 1). In consequence,a negative SGR of −4.94±0.29% day−1 was calculated within theanimals under inanition from days 2 to 10 (Fig. 2).
3.3. Re-feeding period
A reduction on final wet weight was observed according withfasting days, with high values of animals of control group and low inoctopuses exposed to 8 and 10 days fasting (Pb0.05; Fig. 1). After25 days of re-feeding period none of the exposed groups to inanitionwere able to catch up their weight as that shown in the control group(Table 1). Even if the SGR shown during this periodwas similar amongall groupsdue to a large variation, there is the possibility that all groupswill be able to catch up over time, with values between 1.34% day−1
and 0.78% day−1 for the groups of octopuses exposed to 8 and 2 daysinanition, respectively (PN0.05; Table 1). As a result, a SGRmean value
Fig. 1.Wetweights ofOctopusmaya juveniles during acclimation, fasting and re-feedingperiods. Values are given as means±SD. The arrow indicates the point of no return forwet weight (PNRWW). Different letters indicate statistical differences at pb0.05.
of 1.09±0.19% day−1 was calculated for O. maya juveniles duringrecovering period (Fig. 2). There were no statistical differences onoctopus survival between treatments with high values on animalsfasted 2 days (100%) and low in those fasted 6 and 8 days (70%)(PN0.05). A mean value 0f80% survival was calculated for re-feedingperiod.
3.4. Effect of fasting on the amino acid profiles
Essential amino acids were affected by fasting period (Fig. 3a). Arelative reduction of Thr (81%), Phe (15%) and Ile (40%) was observedaccording with inanition period while an increase on His (116%), Arg(132%), and Lys (566%) was evaluated on O. maya muscle. Nonessential amino acids were also affected for the inanition period withrelative increments of Tau (21%) and Gly (51%) and reductions on Ala(62%), Glu (46%) and Ser (88%) (Fig. 3b). The rest of the amino acidsdid not change significantly during fasting experimental periods.
4. Discussion
In the present study after 6 days of food deprivation on O. maya thewet weight was significantly lower than that observed in control group.However, 25 dayswere not sufficient for the octupuses to catch up theirweight in comparison to that shown in the control group. This cannot benecessarily due to the phenomena called the point of no return, whereanimals exposed to a food deprivation for longer period of times arenegatively affected, being unable to show growth rates to equal thecontrol group or even to showhigher growth rates to be able to re-catch
-6-5-4-3-2-101234
Acclimation Fasting Re feeding
SG
R, %
day
-1
Experimental periods
Fig. 2. Specific growth rate (SGR, % day−1) of O. maya juveniles during acclimation,fasting and re-feeding periods. Values are given as means±SD.

Fig. 3. Amino acid content (gAA/100 g protein) of individual samples of arms of theOctopus maya juvenile of 32 days post-hatching (DPH) during the acclimation period(control on day 12), and at the fasting period (on the 4 and 8 days fasting). Each aminoacid was ordered from major to minor content respect to the control. Values are givenas means±SD. NEAA, nonessential amino acid; EAA, essential amino acid.
218 A. George-Zamora et al. / Aquaculture 314 (2011) 215–220
their weight. It is interesting to note that the growth rate during fastingand re-feeding periods was not affected by treatments, demonstratingthatO. maya juveniles have a wide plasticity to tolerate, at least 10 daysof fastingwithout any apparently physiological damage that could affectthe mechanisms involved into the growth rate of animals. Aquaticanimal are generally exposed to long periods of inanition and thereforethey are easily able to support long periods of food deprivation beforesuffering the point of no return by several mechanisms like loweringtheir metabolic rate and being able to efficiently use their reserves.Althoughwe don't know exactly what kind ofmechanisms are involvedon such physiological plasticity, the results of the present study showsthat AA were being used as an energetic support during the inanition.Besides the tissue that was lost during the food deprivation (−4.63 to−5.26% day−1), the O. maya juveniles the AA like Thr, Phe, Ile, Ala, Gluand Ser, were relatively more used, suggesting a strong mobilization ofboth essential and non essential AA to maintain the homeostasis. Proveof that is that survival of animals during fasting and re-feeding periodwas not affected by treatments. In fact a similar survival of control groupwas observed on fasting groups suggesting that natural mortalityprovoked by isolation or manipulation could be present. Leporati et al.,(2007) found 78% survival of O. pallidus hatchlings, while in the presentstudyameansurvival valueof 80%was found forO.maya early juveniles.
No studies have examined the fasting consequences on octopusearly juveniles mainly due that there are not many research teamsthat have a production of octopus juveniles to make such type ofstudies. Although at the date there are some studies on the effects of
fasting condition on cephalopod paralarvae, that results should betaking with precaution because the differences between both lifestages (Villanueva and Norman, 2008). Taking into account that thereare not many research on this topics, the results obtained in thepresent study were compared with other obtained both octopus andsquid paralarvae; that comparison should be taken with cautionbecause amino acid profile obtained on O. maya juveniles were fromarms while in paralarvae studies it was obtained on whole animalsVidal et al., (2006) showed that Loligo opalescens 15 days age,tolerated 3 days without food at 16 °C with no recuperation after3 days fasting. In that study they observed that the mechanismsaffected by starvation involved protein synthesis (RNA/DNA ratio),and the general lost of squid homeostasis. It was also observed thatparalarvae fasted for 3 days or less not only survive but they showed acompensatory growth rate that permitted to animals reach a final wetweight similar to control group. Other different characteristic ofoctopus paralarvae is that after hatch they have available yolkreserves that help to animal during the first days of plancktonic life.According to Villanueva and Norman (2008) octopus paralarvae cantolerate up to 15 days of starving conditions depending on size ofhatchlings while hatchlings of L. opalescens can survive until 6 daysthanks to yolk reserves (Vidal et al., 2002). Several studies have shownthat the ability of a marine organism to resist deprivation of fooddepends on temperature and the developmental stage; as larvae growtheir sensitivity to lack of food diminishes (His and Seaman, 1992;Strüssmann and Takashima, 1992; Vidal et al., 2002; Zheng et al., 2005;Iglesias et al., 2006; Vidal et al., 2006; Viana et al., 2007). Thus, higherfasting resistance should be expected for older organisms; in fact,O. maya as all octopuses should tolerate starving conditions during the50 days of maternal care period after spawn (Solis, 1967), whenanimals should use mainly protein to support its life.
There are few studies on body AA composition in juvenilecephalopods. Among the most recent, Villanueva et al. (2004) foundhigh concentrations of Lys, Leu and Arg in O. vulgaris paralarvae,representing nearly half of total AA. Moreover, in studies made on sub-adult and adult animals of O. maya have revealed that Lys, Leu and Argare also abundant (Aguila et al., 2007). However the results obtained inthe present study demonstrated that the amino acidic composition ofoctopus tissues could be not necessarily the most important forenergetic metabolism. In fasting conditions Thr, Phe, Ile, Ala, Glu andSer were mobilized to maintain animals without physiological damagedemonstrating that the amino acidic requirement for the energeticdemands of cephalopods could be around those amino acids, more thanAAs that are abundant in tissues. Threoninehas been suggested to be thebest substrate for D-lactate formation in octopus tentacle, fructose-1,6-biphosphate and Gly (Akagi and Ohmori, 2004). Also, Fujisawa et al.,(2005) detected increments of D-lactate dehydrogenase, together withlow values of D-lactate in the mantle of fasted Octopus ocellatus,demonstrating that the gluconeogenic pathway could be a moregeneralized process on cephalopods. These authors suggested that Thrand Gly contribution to total protein are high enough to formmethylglyoxal in octopuses. The animals appear to use methylglyoxalmore actively during starvation than under normal conditions. Apossible explanation for the active utilization of methylglyoxal is thatit is formed from amino-acetone, which in turn is formed from Gly andThr. Fasted octopusesmust use Thr or Gly even frombody protein. Thus,it appears that fasted octopuses achieve energy acquisition moreactively through methylglyoxal than from pyruvate. In O. maya, thismetabolic route seems to be similar to that of O. ocellatus. The resultsobtained in thepresent study, suggests active “consumption”of Thr. Arg,His, Lys, Gly, Tau over the fasting days period. Arginine is vigorouslymetabolized in cephalopods (Hochachka and Fields, 1982; Hochachkaet al., 1983) and participates in anaerobicmetabolism. During anaerobicwork, Arg-phosphate is hydrolyzed, leading to increased availability ofArg for condensation with glucose-derived pyruvate to form octopine,the main anaerobic end product that accumulates in adult cephalopods

219A. George-Zamora et al. / Aquaculture 314 (2011) 215–220
during periods of exercise and stress (Hochachka et al., 1976; Storeyet al., 1979; Gäde, 1980). When octopuses must urgently escape from apredator or live anaerobically for a time, Arg-phosphate is drawn(Fujisawa et al., 2005). In the case of O. maya, the increment in Argconcentration and utilization of other AA (mainly NEAA and Thr) duringstarvation may be linked to a strategy aimed at obtaining energeticreserves to survive stressing conditions. Thismay also be the case forHisand Lys, detected at high proportion of total protein during fastingconditions. Lysine was the only ketogenic amino acid which showedsignificant changesduring fasting condition (D'Mello, 2003; all theotherAA are glucogenic).
InO.mayaAlawas themost abundant, followed by Tau, Asp andGlu.References on the specific function of Ala in cephalopods are lacking.Aspartic acid and Glu acid were reported also as the most abundant AAof diverse cephalopods tissues as: mantle of O. vulgaris (Rosa et al.,2002), gonad of O. vulgaris and O. defilippi (Rosa et al., 2004), gonad ofIllex coindetii and Todaropsis eblanae (Rosa et al., 2005), gonad anddigestive gland of Rossia macrosoma (Rosa et al., 2006). Also, on earlystages of S. officinalis, L. vulgaris and O. vulgaris and hatchlings and wildjuveniles of L. vulgaris and O. vulgaris (Villanueva et al., 2004), and O.maya (Aguila et al., 2007; Domingues et al., 2007), were found highproportion of Tau of total protein. Taurine has been shown to play ametabolic role in the regulation of cell osmotic pressure (Norton, 1979;Bishopet al., 1983), and in the anaerobicmetabolismofbivalvemollusks(Wells and Baldwin, 1995).
In general,O.maya appears to use Thr, Ser andAlamainly asmetabolicfuel to face starvation. Simultaneous increases in His, Arg and Lysconcentrations suggest possible accumulation of these AAs. It should bekept in mind that previous work to meet nutritional requirements inother mollusks and fishes species has shown that high concentrations ofcertainAAsarenot the result ofdenovo synthesisbut of proteinhydrolysis(Gadomski and Petersen, 1988; Moran and Manahan, 2004). Our resultsprovide insights on the relative utilization of AAs in octopuses, in order todesign novel diets that could include different relative concentrations oftheseAAs tomeet specific nutritional requirements. Since octopuses havelow fat reserves, the amino acids may be used for energy supply duringfasting period. Accordingly, when glycogen stores are depleted as a resultof fasting, energy must be obtained from increased catabolism of aminoacids and even of body proteins (Storey and Storey, 1983; Fujisawa et al.,2005), just to maintain the homeostasis. That adaptation could bereflecting capability of octopus to tolerate starvation during ecologicalconstrains of food and during maternal care period.
Acknowledgements
This study is part of the Ph.D. thesis of George-Zamora A. The presentstudy was partially supported by DGAPA-UNAM project no. IN202909and CONACYT-Básico 24743. Thanks are given to “Posgrado en Cienciasdel Mar y Limnologia, PCMyL” and to the “Pulpo Program from Sisal” ofUMDI-UNAM, for the possibility to participate in CIAC-09 SymposiumandWorkshops. Also thanks are given to “Posgrado enCienciasdelMaryLimnologia, PCMyL” forfinancial support during the stay at the “Institutode Investigaciones Oceanologicas, at the Universidad Autónoma de BajaCalifornia”. Claudia Caamal-Monsreal, Richard Mena and Luis Yan fromUMDI-Sisal UNAM, the Comite Sistema Producto Pulpo “CSPP”Moluscosdel Mayab, and Victoria Patiño-Suarez from CINVESTAV-Mérida helpedwith technical assistance during the experiment and maintenance ofoctopuses. Omar Hernando Avila-Poveda is also thanked by reviewingand editing the manuscript.
References
Aguila, J., Cuzon, G., Pascual, C., Domingues, P.M., Gaxiola, G., Sánchez, A., Maldonado, T.,Rosas, C., 2007. The effects of fish hydrolysate (CPSP) level on Octopus maya (Vossand Solis) diet: digestive enzyme activity, blood metabolites, and energy balance.Aquaculture 273, 641–655.
Akagi, S., Ohmori, S., 2004. Threonine is the best substrate for D-lactate formation inoctopus tentacle. Amino Acids 26, 169–174.
Avila-Poveda, O.H., Colin-Flores, R.F., Rosas, C., 2009. Gonad development during theearly life of O. maya (Mollusca: Cephalopoda). The Biological Bulletin 21, 94–102.
Bishop, S., Ellis, L., Burcham, J., 1983. Amino acid metabolism in molluscs. In:Hochachka, P.W. (Ed.), The Mollusca. Metabolic Biochemistry and MolecularBiomechanics, 1. Academic Press, New York, pp. 243–327.
Boletzky, S.V., 1974. Elevage de Céphalopodes en aquarium. Vie et Milieu 24,309–340.
Boletzky, S.V., Hanlon, R.T., 1983. A review of the laboratory maintenance, rearing andculture of cephalopod molluscs. Memoirs of Museum Victoria 44, 147–187.
Castro, B.G., Lee, P.G., 1994. The effects of semi-purified diets on growth and conditionof Sepia officinalis L. (Mollusca: Cephalopoda). Comparative Biochemistry andPhysiology. Part A, Physiology 109A, 1007–1016.
Castro, B.G., DiMarco, F.P., DeRusha, R.H., Lee, P.G., 1993. The effects of surimi andpelleted diets on the laboratory survival, growth and feeding rate of the cuttlefishSepia officinalis. Journal of Experimental Marine Biology and Ecology 170, 241–252.
Comisión Nacional de Acuacultura y Pesca (CONAPESCA), 2008. Anuario Estadístico deAcuacultura y Pesca 2008. SAGARPA-CONAPESCA. http://www.conapesca.sagarpa.gob.mx/wb/cona/cona_anuario_estadistico_de_pesca. Available at.
D'Mello, J.P.F., 2003. Amino acids as multifunctional molecules. In: D'Mello, J.P.F. (Ed.),Amino Acids in Animal Nutrition. CABI Publishing. Cambridge, USA, pp. 1–14.
Development of Alternative Diets for the Mass Culture of the European Cuttlefish Sepiaofficinalis. Ph.D. Thesis, University of the Algarve, Portugal.
Domingues, P.M., DiMarco, P.F., Andrade, J.P., Lee, P.G., 2005. Effect of artificial diets ongrowth, survival and condition of adult cuttlefish, Sepia officinalis Linnaeus, 1758.Aquaculture International 13, 423–440.
Domingues, P.M., López, N., Muñoz, J.A., Maldonado, T., Gaxiola, G., Rosas, C., 2007.Effects of a dry pelleted diet on growth and survival of the Yucatan octopus, Octopusmaya. Aquaculture Nutrition 13, 273–280.
Domingues, P., Garcia, S., Garrido, D., 2010. Effects of three culture densities on growthand survival of Octopus vulgaris. Aquaculture International 18, 165–174.
Fujisawa, T., Akagi, S., Kawase, M., Yamamoto, M., Ohmori, S., 2005. D-Lactatemetabolism in starved Octopus ocellatus. Journal of Experimental Zoology. Part A:Comparative Experimental Biology 303A, 489–496.
Gäde, G., 1980. Biological role of octopine formation inmarinemolluscs. Marine BiologyLetters 1, 121–135.
Gadomski, D.M., Petersen, J.H., 1988. Effects of food deprivation in larvae of twoflatfishes. Marine Ecology Progress Series 44, 103–111.
Grigoriou, P., Richardson, C.A., 2009. Effect of body mass, temperature and fooddeprivation on oxygen consumption rate of common cuttlefish Sepia officinalis.Marine Biology 156, 2473–2481.
Hanlon, R.T., Forsythe, J.W., 1985. Advances in the laboratory culture of octopuses forbiomedical research. Laboratory Animal Science 35, 33–40.
His, E., Seaman, M.N.L., 1992. Effects of temporary starvation on the survival, and onsubsequent feeding and growth, of oyster (Crassostrea gigas) larvae. Marine Biology114, 277–279.
Hochachka, P.W., Fields, J.H.A., 1982. Arginine, glutamate and proline as substrates foroxidation and for glycogenesis in cephalopod tissues. Pacific Science 36, 325–335.
Hochachka, P.W., Hartline, P.H., Fields, J.H.A., 1976. Octopine as an end product ofanaerobic glycolysis in the chambered nautilus. Science 195, 72–74.
Hochachka, P.W., Mommsen, T.P., Storey, J., Storey, K.B., Johansen, K., French, C.J., 1983.The relationship between arginine and proline metabolism in cephalopods. MarineBiology Letters 4, 1–21.
Iglesias, J., Sánchez, F.J., Otero, J.J., Moxica, C., 2000. Culture of Octopus (Octopus vulgaris,Cuvier): Present knowledge, problems and perspectives. Cahiers Options Médi-terranéennes 77, 313–321.
Iglesias, J., Fuentes, L., Sánchez, L., Otero, J.J., Moxica, C., Lago, M.J., 2006. First feeding ofOctopus vulgaris Cuvier, 1797 paralarvae using Artemia: Effect of prey size, preydensity and feeding frequency. Aquaculture 261, 817–822.
Iglesias, J., Sánchez, F.J., Bersano, J.G.F., Carrasco, J.F., Dhont, J., Fuentes, L., Linares, F.,Muñoz, J.L., Okumura, S., Roo, J., van der Meeren, T., Vidal, E.A.G., Villanueva, R.,2007. Rearing of Octopus vulgaris paralarvae: present status, bottlenecks andtrends. Aquaculture 266, 1–15.
Le Bihan, E., Perrin, A., Koueta, N., 2006. Influence of diet peptide content on survival,growth and digestive enzymes activities of juvenile cuttlefish Sepia officinalis. Vie etMilieu 56, 139–145.
Lee, P.G., 1994. Nutrition of cephalopods: fueling the system. Marine and FreshwaterBehaviour and Physiology 25, 35–51.
Leporati, E., Pecl, G., Semmens, J.M., 2007. Cephalopod hatchling growth: the effects ofinitial size and seasonal temperatures. Marine Biology 151, 1375–1383.
Moguel, C., Mascaró, M., Avila-Poveda, O.H., Caamal-Monsreal, C., Sanchez, A., Pascual,C., Rosas, C., 2010. Morphological, physiological, and behavioral changes duringpost-hatching development of Octopus maya (Mollusca: Cephalopoda) with specialfocus on the digestive system. Aquatic Biology 9, 35–48.
Moran, A.L., Manahan, D.T., 2004. Physiological recovery from prolonged “starvation” inlarvae of the Pacific oyster Crassostrea gigas. Journal of Experimental MarineBiology and Ecology 306, 17–36.
Nabhitabhata, J., 1995. Mass culture of cephalopods in Thailand. World Aquaculture 26,25–29.
Norton, R.S., 1979. Identification of mollusc metabolites by natural-abundance 13CNMR studies of whole tissue and tissue homogenates. Comparative Biochemistryand Physiology. Part B: Biochemistry & Molecular Biology 63B, 67–72.
Quintana, D., Rosas, C., Moreno-Villegas, E., 2010. Relationship between nutritional andrearing parameters of Octopus maya juveniles fed with different rations of crabpaste. Aquaculture Nutrition. doi:10.1111/j.1365-2095.2010.00772.x.

220 A. George-Zamora et al. / Aquaculture 314 (2011) 215–220
Rosa, R., Nunes, L., Sousa-Reis, C., 2002. Seasonal changes in the biochemicalcomposition of Octopus vulgaris Cuvier, 1797, from three areas of the Portuguesecoast. Bulletin of Marine Science 71, 739–751.
Rosa, R., Costa, P.R., Nunes, M.L., 2004. Effect of sexual maturation on the tissuebiochemical composition of Octopus vulgaris and O. defilippi (Mollusca: Cephalo-poda). Marine Biology 145, 563–574.
Rosa, R., Pereira, J., Nunes, M.L., 2005. Biochemical composition of cephalopods withdifferent life strategies, with special reference to a giant squid, Architeuthis sp.Marine Biology 146, 739–751.
Rosa, R., Pereira, J., Costa, P.R., Nunes, M.L., 2006. Distribution, abundance, biology andbiochemistry of the stout bobtail squid Rossia macrosoma from the Portuguesecoast. Marine Biology Research 2, 14–23.
Rosas, C., Cuzon, G., Pascual, C., Gaxiola, G., Chay, D., López, N., Maldonado, T.,Domingues, P.M., 2007. Energy balance of Octopus maya fed crab and artificial diet.Marine Biology 152, 371–381.
Rosas, C., Tut, J., Baeza, J., Sánchez, A., Sosa, V., Pascual, C., Arena, L., Domingues, P.M.,Cuzon, G., 2008. Effect of type of binder on growth, digestibility, and energeticbalance of Octopus maya. Aquaculture 275, 291–297.
Segawa, S., Hanlon, R.T., 1988. Oxygen consumption and ammonia excretion rates inOctopus maya, Loligo forbesi and Lolliguncula brevis (Mollusca: Cephalopoda).Marine Behaviour and Physiology 13, 389–400.
Solis, M., 1967. Aspectos biológicos del pulpo Octopus maya Voss y Solis. InstitutoNacional de Investigaciones Biológico Pesqueras (México) 18, 1–90.
Solorzano, Y., Viana, M.T., López, L.M., Correa, J.G., True, C.C., Rosas, C., 2009. Response ofnewly hatched Octopus bimaculoides fed enriched Artemia salina: growthperformance, ontogeny of the digestive enzyme and tissue amino acid content.Aquaculture 289, 84–90.
Storey, K.B., Storey, J.M., 1983. Carbohydrate metabolism in cephalopod molluscs. In:Hochachka, P.W. (Ed.), The Mollusca. Metabolic Biochemistry and MolecularBiomechanics, vol. 1. Academic Press, New York, pp. 91–136.
Storey, K.B., Storey, J.M., Johansen, K., Hochachka, P.W., 1979. Octopine metabolism inthe cuttlefish, Sepia officinalis. Effect of hypoxia and metabolite loads on the blood
levels of octopine and related compounds. Canadian Journal of Zoology 57,2331–2336.
Strüssmann, C.A., Takashima, F., 1992. Larval growth and point-of no-return in fish.Oceánis 18, 27–39.
Van Heukelem, W.F., 1977. Laboratory maintenance, breeding, rearing and biomedicalresearch potential of the Yucatan octopus (Octopus maya). Laboratory AnimalScience 27, 852–859.
Van Heukelem, W.F., 1983. Octopus maya. In: Boyle, P.R. (Ed.), Cephalopod Life Cycles,vol. I. Academic Press, London, pp. 311–323.
Viana, M.T., D'Abramo, L.R., Gonzalez, M.A., García-Suárez, J.V., Shimada, A., Vásquez-Peláez, C., 2007. Energy and nutrient utilization of juvenile green abalone (Haliotisfulgens) during starvation. Aquaculture 264, 323–329.
Vidal, E.A.G., DiMarco, F.P., Wormuth, J.H., Lee, P.G., 2002. Influence of temperature andfood availability on survival, growth and yolk utilization in hatchling squid. Bulletinof Marine Science 71, 915–931.
Vidal, E.A.G., DiMarco, F.P., Lee, P.G., 2006. Effects of starvation and recovery on thesurvival, growth and RNA/DNA ratio in loliginid squid paralarvae. Aquaculture 260,94–105.
Villanueva, R., Norman, M.D., 2008. Biology of the planktonic stages of benthicoctopuses Oceanogr. Marine Biology Annual Review 46, 105–202.
Villanueva, R., Riba, J., Ruíz-Capillas, C., González, A.V., Baeta, M., 2004. Amino acidcomposition of early stages of cephalopods and effects of amino acid dietarytreatments on Octopus vulgaris paralarvae. Aquaculture 242, 455–478.
Voss, G.L., Solís-Ramírez, M., 1966. Octopus maya, a new species from the Bay ofCampeche. Bulletin of Marine Science 16, 615–625.
Wells, R.M.G., Baldwin, J., 1995. A comparison of metabolic stress during air exposure intwo species of New Zealand abalone, Haliotis iris and Haliotis australis: implicationsfor the handling and shipping of live animals. Aquaculture 134, 361–370.
Zheng, H., Ke, C., Zhou, S., Li, F., 2005. Effects of starvation on larval growth, survival andmetamorphosis of Ivory shell Babylonia formosae habei Altena et al., 1981(Neogastropoda: Buccinidae). Aquaculture 243, 357–366.
Copyright © 2022 FDOKUMEN




















