
Cuvier meets Watson and Crick: the utility of molecules as classical homologies
Upload
independentCategory
view
5download
0
EI.SEVIER J. Exp. Mar. Bid. Ecol. 177 il994) 1X7-20.!
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Feeding behaviour and selection of bivalve prey by ~c~~~~~~ vulgaris Cuvier
C. D. ~cQu~id
Abstract
Ocfo~~~s pz&&y in South Africa preys primarily on mussels. Small mussels are simply pulled open, while large ones are drilled first and then pulled open. Medium mussels fall into an overlap size range and may be opened by either method. Actively pulling a mussel open was estimated to have an energetic cost 4.5 times that of resting metabolism, but active pulling lasts for only seconds and forms a very small proportion of handling time. The energetic cost per minute of handling prey while using force is only 1.29 times greater than when they are drilled. Curves of rate of energetic return (E/H) against mussel size were derived from laboratory measurements. Curves varied for individual octopuses and the technique used to open mussels. They were monotonic with sharp discontjnuities between the two techniques in the overlap size range. Drilling takes much longer than using force alone and produces lower E/H values. Mussels just smaller than the length at which SOY,, are drilled (L,,,JO) are “optimal” in terms of maximum E:H values. L,,i,,SO increases with octopus weight. In the laboratory octopuses selectively took mussels with maximum E/H. The force required to remove mussels from rocks in the laborator>
is less than that needed to open them, but increases with mussel size. Previously published data on prey selection in the field indicate that octopuses of > 500 g take the largest mussel size classes. Below 500 g, mean prey size in the field is related to octopus weight. Small octopuses may take smaller mussels because of they are unable to remove large ones from the rocks.
Ke)* worcis: Feeding behaviour; Octopus n&uric; Pemu pernu; Prey selection
1. Introduction
For motile species feeding entails actively seeking out food, or foraging. During foraging an animal can move randomly or it can concentrate its attentions on a spe- cific part of its habitat. It can also either accept and consume food as it is encountered
~(~~~-~981~94~$7.00 0 1994 Eisevier Science B.V. A11 rights reserved SSDl 0022-098 l(93)EO 175-X

IXX C.D. McQurrrd :I J. E.vp. Mar. Bid. Ewl. 177 /I 994) 147-202
or decide to accept or discard each food item individually. Optimal foraging theory
attempts to model and predict such decision making by animals during foraging (see
Schoener. 1987 for a history of the theory and Hughes, 1980 for its application to marine
organisms). A central assumption of the theory is that natural selection will favour behaviour patterns that maximise fitness (e.g. Burrows & Hughes, 1990). although “fitness” can be difficult to define (Sih, 1984). The theory predicts that maximum fitness is achieved by selection for foraging behaviour that will maximise or minim& some
consequence of behaviour (Abrams, 1982). For example. it might maximise the number of encounters with potential food items or minimise exposure to predation during foraging.
The most obvious and commonly used denominator of “fitness” is the rate of energy intake (Pyke et al., 1977; Pyke, 1984). The use of energy intake as a currency for measuring optimum behaviour assumes that other factors (such as predation) are un- important (Pyke et al., 1977). In this paper the predictions of the theory are applied to prey selection by Octopus. Prey selection and prey value curves were measured in the laboratory and compared with a previous study on prey selection in the field.
Octopus vulg~wis is an abundant predator in the intertidal and shallow subtidal zones of much of the South African coast. Densities of 0.007 m -’ (i.e. an arca of
12 x 12 m per octopus) have been recorded in Natal (Buchan & Smale, 198 1) whcrc the main prey is the brown mussel Prtxaperm (L.). Octopus wlgmis attains much lower densities and biomass than other major predators, such as rock lobsters, but has higher consumption rates and is calculated to be the most important sublittoral predator of mussels (Buchan & Smale, 1981; Griffiths & Hockey, 1987).
Ocrop~s vulgaris takes many prey species, larger individuals taking a wider variety (Smale & Buchan. 19X1), but the use of a number of prey species greatly increases the complexity of predicting prey selection. The application of optimal foraging theorb is most likely to be successful when there is a clear hierarchy of “profitability”. as when different sized individuals of a single prey species are considered. In this study differ- ent size classes of Prrm prrntr were used to provide prey of different cncrgetic value and handling difficulty.
2. Methods
There arc two aspects to handling mussel prey: removing them from the rocks and opening the valves. Both aspects were examined on a mechanical level, followed by an evaluation of feeding behaviour of octopuses when selecting and opening mussels.
The force required to detach mussels from the substratum was measured in the laboratory. Mussels were left for 12 days in running seawater to attach in clumps of three to rocks collected from the shore. Each rock was then removed from the aquarium, the mussel shells wiped dry and metal tags glued to them with epoxy glue. After a further 12 h in the aquarium a fine piano wire was hooked to each tag. The three wires were

C.D. McQuaid 1 J. Exp. Mar. Biol. Ecol. I77 (1994) 187-202 18’)
attached to a fourth wire leading via a pulley to a container. The wires were adjusted
to give even pull along the longitudinal axis of each mussel and the container slowly
filled with sand until the mussel clump detached. The sand was then weighed. This weight was divided by 3 and plotted against the mean size of the three mussels.
The force required to pull open individual mussels was measured similarly by glueing a metal tag to each valve. One was hooked to a heavy weight and the other, via a pulley, to a container slowly filled with sand. The effect of pull is a function of duration as well as magnitude of pull. No attempt was made to simulate the pull exerted by an octopus, and the weight of sand was increased at the same rate in all cases
(Z400 g’ 10 s -I>.
2.2. Octopus behaviour
Eight octopuses were used to examine two aspects of behaviour: prey handling and
prey selection. To predict prey size selection it is necessary to rank the prey by their “profitability”.
Larger prey contain more flesh, but may also take longer to capture, subdue and eat (i.e. they require a longer handling time). Profitability is therefore estimated as the rate
of energetic return, measured as E/H; the energetic value of the prey (E) divided by handling time (H). The dry mass of mussel flesh was used as a measure of energetic value, as the two are directly proportional (unpubl. data).
Handling times were recorded with a stop-watch to the nearest second by continu- ous direct observation of six octopuses in individual aquaria. Octopuses were starved overnight before being offered unattached prey, to which they normally reacted imme-
diately. Timing began once the octopus had settled with a mussel and had begun try- ing to open it. This was an obvious behaviour during which the octopus arranged the
mussel within the web to gain purchase with as many arms as possible. Pulling was clearly marked by tensing of the arms and ended with a sharp release of tension as the
mussel opened, the suckers tore free, or the octopus relaxed. Clearly defined pulling behaviour allowed accurate measurement of the duration of each pull. An octopus that resorted to drilling a mussel rearranged it within the web and then sat motionless for prolonged periods with occasional changes of grip. Drilling was terminated by pulling behaviour. After drilling a single pull was usually sufficient to open a mussel. Once a mussel was opened, feeding involved a pulsing action of the head as flesh was torn away
from the shells and ended when the empty valves were blown out from under the web by a jet of water.
The data collected were used to produce plots of E/H against mussel length. These plots predict the prey size classes which are energetically most profitable, or “optimal”, for the octopuses to eat. These size classes were compared with those selected when offered an overabundance of food. Octopuses in the laboratory ate 6-7 mussels per day. For this experiment six individuals were offered 25 mussels each day, covering the complete range of mussel sizes available in the field. Each morning all mussels were removed, measured and replaced by another 25. In three cases all prey size classes were offered in identical proportions. Each experiment was run for 3 to 6 wk.

C.D. McQuuid / J. E.vp. Mur. Biol. Ewl. 177 ilY94/ 187-202 I90
3. Results
3.1. Mechanics
The diameter of the posterior adductor muscle was significantly correlated with mussel length (y = 0.096 x1.062; n = 53; r = 0.87; p <O.Ol), as was the weight of sand required to pull the valves apart (Fig. 1). The weight of sand required to pull mussels
from the substratum was also closely related to the mean size of the mussels in a clump
and markedly lower than that required to open a mussel of the same size (Fig. 1).
3.2. Feeding hehaviour
During the use of force to open shells, the mussel was held close to the mouth, usually with the apex pointing away from the octopus and the dorsal ligament either up or down rather than to the side. Each pull lasted z 5 s and often ended when the epithelium of the suckers tore away. With a small mussel pulling continued with - 1 pull I min ‘. until
the adductor muscle of the bivalve ruptured or, rarely, the shell broke. With a larger
mussel the octopus changed to drilling with the salivary papilla after 5 or 10 min.
I I 8
4 5 6
SHELL LENGTH (CM)
Fig. I. Relationship between weight of sand required to remove a mussel from the substratum and mean length of mussels in a clump (0: wt = -2.879 + 0.76 length; n = 16; Y= 0.84; p<O.Ol) and between weight of sand required to pull the valves open mechanically and mussel length (0: wt = 456 + 108.35 length: II = 70: r=O.58; /~<0.01).

191
ANTERIOR
SHELL THICKNESS
Fig. 2. Position of drill sites (0) on a number of mussel shells, recorded separately for three octopuses. 0 indicates sites of incomplete holes.
Octopuses invariably attempted to force a mussel open before drilling it, but with a very large mussel this was perfunctory and they resorted to drilling almost immediately, after
perhaps a single preliminary pull. Small octopuses frequently drilled more than one complete hole before attempting to use force. The drilled holes were irregular slits with
sharply tapering sides. The inner aperture was much narrower than the external aper- ture and often failed to penetrate the innermost, transparent layer of the shell.
Usually the shell was drilled in the mid-dorsal region (i.e. above the heart) often preceded by an initial, incomplete hole in the vicinity of the umbo (Fig. 2). The time required to drill through the shell depended on mussel size and the site of the drill hole
II 0 100 200 300 400
OCTOPUS WT (9)
Fig. 3. The relationship between the length at which SOT, = 0.5588 x0 J;
of mussels are drilled and octopus weight (I+,,,,50 n = 7: r= 0.89; pccO.01). Ldi,li50 was estimated ~aphicali~ for individual octopuses from data
on ‘;, mussels drilled in each mussel size class.
IQ? C.D. McQuuid / J. E.rp. Mar. Biol. Ecol. I77 11994) 1X7-202
and ranged from 30 min to over 8 h. There was no clearly defined size at which an
octopus switched from one technique to the other. Small mussels were regularly forced
open and large ones drilled, but there was an overlap in size range within which mussels might be opened by either method. For larger octopuses this overlap occurred at larger mussel sizes and the size of mussel at which 50”‘, of the prey were drilled (L,,i,,50) was
greater (Fig. 3).
3.3. Handling time
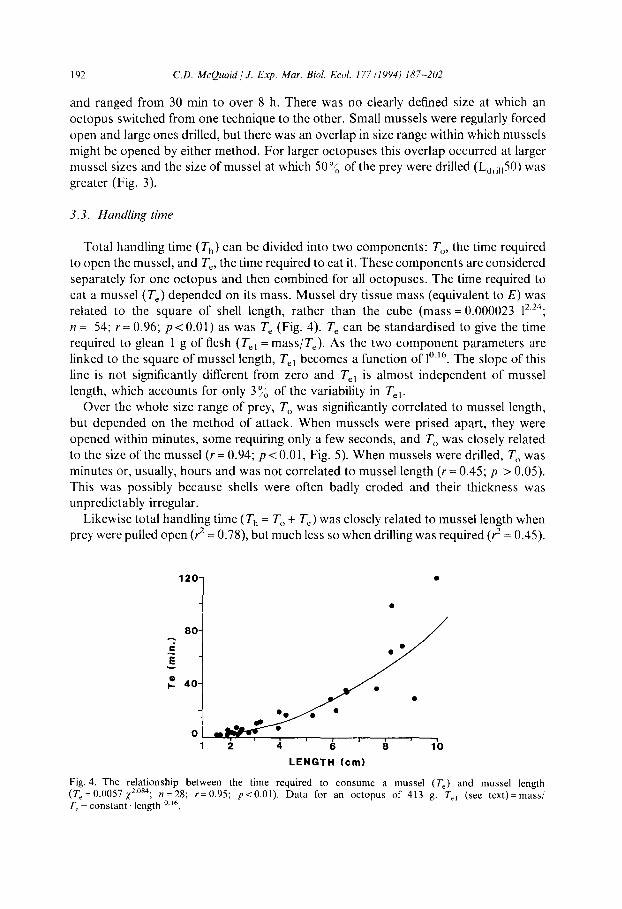
Total handling time (T,,) can be divided into two components: T,,, the time required to open the mussel, and T,, the time required to eat it. These components are considered separately for one octopus and then combined for all octopuses. The time required to
eat a mussel (T,) depended on its mass. Mussel dry tissue mass (equivalent to E) was related to the square of shell length, rather than the cube (mass = 0.000023 12.24; n = 54; Y = 0.96; p<O.Ol) as was T, (Fig. 4). T, can be standardised to give the time required to glean 1 g of flesh (T,, = mass/T,). As the two component parameters are linked to the square of mussel length, T,, becomes a function of l”.16. The slope of this line is not significantly different from zero and T,, is almost independent of mussel length, which accounts for only 3 % of the variability in 7’,,
Over the whole size range of prey, To was significantly correlated to mussel length,
but depended on the method of attack. When mussels were prised apart, they were opened within minutes, some requiring only a few seconds, and T, was closely related to the size of the mussel (r = 0.94; p < 0.0 1, Fig. 5). When mussels were drilled, T,, was minutes or, usually, hours and was not correlated to mussel length (v = 0.45; p > 0.05). This was possibly because shells were often badly eroded and their thickness was unpredictably irregular.
Likewise total handling time ( Th = T,, + T,) was closely related to mussel length when prey were pulled open (? = 0.78), but much less so when drilling was required (2 = 0.45).
120- .
.
80-
.5 L
: 40-
0 - * 1 2 4
I 6 8 10
LENGTH (cm)
Fig. 4. The relationship between the time required to consume a mussel (T,) and mussel length (T, = 0.0057 xzos4; n = 28; r = 0.95; T, = constant length ‘.I’.
pt0.01). Data for an octopus of 413 g. T,, (see text)= mass/

C.D. McQuaid / J. Exp. Mtrr. Biol. Ecol. 177 11994) 187-202 193
6-
0 I 1 1 2 3 4 6
LENGTH (cm)
Fig. 5. The relationship between the time required to open a mussel by force and mussel length (T, = 2.07 + 0.14 x : n = 17: I= 0.94; p<O.Ol). Data for an octopus of 413 g. T, for drilled mussels is not predictable
from length (r = 0.45; p > 0.05).
Nevertheless, T,, was significantly correlated with mussel length over the whole size range, i.e. when data for both methods were combined (Fig. 6, r = 0.87; p<O.Ol).
Th also varied for individual octopuses. When force was used, Th for a given mussel
length depended on octopus size; bigger octopuses generally pulled mussels open faster (Fig. 7). This was not the case if drilling was required; then there was no relationship between octopus size and Th for given mussel sizes (Fig. 8). Thus T,, was predictable
.
.
. I/ l
DRILL
.
1 2 4 6 6 10
LENGTH (cm)
Fig. 6. The relationship between total handling time (&) and mussel length for an octopus of 413 g. 0 are data for mussels opened by force; 0 for drilled mussels. All data were combined in the calculation of the correlation coefficient. n = 38; I = 0.87; p < 0.01.

193 C.D. MrQwid : J. Esp. Mur. Bioi. EmI. I77 (1994) 187-202
i 1
0
5cm
0
0
5
0
0
l 0 ’
l *
0 100 200 300 400
OCTOPUS WT (9)
Fig. 7. The rciationship between octopus weight and iotai handling time (I;,) for mussels of 3 cm (II = h: Y = 0.90: p < 0.05) and 5 cm (II = 6: r = 0.52; pc 0.05). Data are for mussels opened by force only.
400
1
4139
( r*=0.76 n = 38
//I? 3709
( r*=:o.86 n = 40
LENGTH (cm)
Fig. 8. The relationship between total handling time (7h) for individual octopuses (wetghts given in grams) and mussel Icngth. Curves were calculated from data for mussels opened by either drilling or force. p < 0.0 1 ill ail casts.

C.D. McQuuid : I. E.ty. Mm. Bid. Etoi. 177 fIYY4) 187-202 195
from mussel length for individual octopuses, but was only related to octopus size if
mussels could be opened without drilling.
3.4. E/H CUPIY.S
The effects described above necessitate separate plots of E/H for each octopus. Because of substantial differences in handling time for drilled and forced mussels.
separate E/H curves are also required for each technique. ‘There is the additional consideration that H represents energetic expenditure during
handling. With energetically equivalent methods of handling, H is proportional to handling time (Th). In the present case, the use of force is energetically more expen-
sive than drilling. This is apparent from changes in respiration rate (visible as changes in rate of exhalation) when animals change handling method in the laboratory (pers.
obs.). The relative metabolic costs of the two methods can be estimated assuming (a) energy
expenditure during drilling approximates resting metabolic rate and (b) 20”” eficiencj of the muscles involved in pulling the valves apart (Alexander, 1968). The work done in opening a mussel is given by the force used multiplied by the distance over which it is applied. The mass of sand required to pull open an 8 cm mussel {the largest size class regularly opened without drilling) is 9.1 kg (Fig. 1). This is converted to a force by multiplying by the acceleration due to gravity (9.8 m * se’). The adductor muscle ruptures when the shell valves have been pulled apart < 1 cm (pers. obs.). The work
done is then:
(9.1 kg x 9.8 m ‘s ‘) x 0.01 m = 0.89 J
With 20”,, efficiency this gives a metabolic cost of ~4.5 times resting inetabolisln, i.e.
4.5 times the cost of drilling per unit time. Each pull lasts 2 5 s, with z 1 pull. min I.
Expressing the cost of resting metabolism as 1 unit. s , drilling for 1 min costs 60 units.
The use of force for 1 min costs (55 units) + (5 x 4.5 units) = 77.25 units. Pulling mussels open therefore costs 1.29 times the cost of drilling. In energetic terms,
T,,pcn(force) = ‘T;,,,,(drilling) x 1.29. This correction factor is much lower for smaller mussels, which require less force to be opened (e.g. it is 1.16 for a 5 cm mussel). r,, could not be divided into T,, and T, for all mussels and so individual corrections could not bc made. Therefore E/H curves were plotted after multiplying T,, by 1.29 for prey cvhich were pulled open, giving a conservative estimate of E/H for these mussels. Drilling entails an initial period, ranging from a few seconds to 10 min, during which octopuses attempt to prise the prey open without drilling. The energetic cost of pulling for 10 min was incorporated in the caIculation of E/H for drilled mussels. Drilling also
rcquircs pulling the mussel open after it has been drilled. This usually entails a single. brief pull for which no correction was made.
There was an ill-defined maximum size to the mussels which an octopus could open without drilling, but no upper limit to the size of mussels which could be opened by drilling.
Even using an over-estimate of H for mussels that were opened by force, their E/H values were generally much higher than for mussels opened by drilling. This difference

196 C.D. McQurrid / J. Exp. Mar. Bid. Ecol. 177 (I 994) 187-202
was particularly obvious in the size range within which a mussel might be attacked by
either method (Fig. 9). Individual octopuses differed in the ease with which they could
open mussels. When force was used, large octopuses often (but not always) opened
mussels more quickly (Fig. 7), and E/H values for given prey sizes tended to be higher. Because there was no relationship between octopus size and handling time when drilling
18
1 65g
6
18-
I
2
6-
Y-0 2 4 6 8 10
LENGTH (cm)
Fig. 9. The relationship between E/H (g’min -‘) and mussel length for six octopuses (weights given in grams). 0 are values for mussels which were forced open, 0, for drilled mussels. The shaded area is the size range within which mussels may be attacked by tither method.

C.B. Ms(c?uaid /.f_ Exp. &fur. Biol. Ecoi. 177 (1994) 187-202 191
was used (Fig. S), E/H for these mussels varied for individual octopuses without re- gard to their size (Fig. 9).
E/H cannot be predicted accurately from mussel length, but in most cases E/H in- creases with mussel Length. Size classes of mussels which could invariably be forced open (i.e. those below the overlap zone in Fig. 9) gave a smaller absolute energetic return than large mussels, but generally gave a higher E/H value (i.e. rate of energetic return). Within the overlap zone mussels might produce either maximum or minimum E/H
values, depending on whether drilling was eventually necessary. Above the length at which 50:/, of prey are drilled (L,,i,,50; Fig. 3) there is an increasing probability that the prey will have to be drilled and that E/H wili be very low. Optimum size classes may then be defined as those at, or just below L,,illSO and vary for individual octopuses.
40
%
12 a 4 5 6 7 6,910
7 2235
LENGTH(cml
Fig. IO. Observed (shaded bars) and expected fopen bars) size distributions of mussels in six choice ex- periments. The weight of the octopus in each experiment is given. n varied between experiments, being 40-83 for observed and 165-I 177 for expected distributions.

19x C.D. McQttuid )/ J. Eup. Mur. Bd. Ed. 177 flYY4i 187-202
3.5. Size selection in the laborator>
Octopuses exhibited non-random prey choice, selecting particular size classes when
offered an excess of mussels (x2, calculated using availability of size classes as expected values, ~~0.05 in all cases, Fig. 10). Both maximum and mean sizes of mussels se-
lected were greater for larger octopuses (Fig. 11). Actual prey choice was compared with the choice predicted by E/H values for each
octopus using r-tests to compare mean size of prey taken with LC,,i,,50 for that size of octopus. Mean size of mussels taken was less than (65-, 223- and 350-g animals in
Fig. 10; one-tailed t-test, p< 0.05) or not significantly different from (50-, 100-, 413-g animals. f-test, p> 0.01) L,,,,,50 in all cases. Thus, all octopuses took prey of a mean
length equal to, or less than, L,,,,,50. The same effect can be demonstrated using the combined data for all the octopuses
by comparing two regressions: (1) octopus weight against mean mussel length eaten in the laboratory (Llab = “mean” in Fig. 11); (2) octopus weight against L,,.,,,50 (Fig. 3). There was no significant difference in slope between Llab and I,,,,,,50 (t-test; ~7 > 0. I). but the J,-intercept was significantly lower for Llnh (p<O.OOl). This indicates that. in the laboratory, octopuses conform to the predictions of optimal foraging theory by
selecting prey with a mean size just below L,,,,,50.
11
I
maximum
9 0
81 mean
iy F-f-7-f 0 100 200 300 400
OCTOPUS WT (9)
Fig. 1 I. The relationship between the maximum and mean ( F SD) lengths of mussels taken In choice ex- periments and octopus size. Maximum = 1.65 f2’ (?I= 6; r= 0.96; p<0.01). Mean = 0.87 I”~“” (n = 6: r=O.94; p<O.Ol).

C.D. McQuuid )/ .J. EYJI. Mrw. Biol. Ecol. 177 11994) 187-X2 I99
01 I r 1
0 500 1000 1500 2000 2500
OCTOPUS WT (g)
Fig. 12. The relationship between the length of musscl~ taken in the field and octopus siLc. Data are from octopus middens. points represent mean values. lines are upper and lower limits of mussels eaten. Redra\\n from Smale & Buchan. 1981.
3.6. Size selection in the field
Smale & Buchanan (198 1) provide data on the size distribution of Pernu prrna in middens at the mouths of dens of South African 0. vulguris (Fig. 12).
For octopuses of less than 500 g (i.e. within the size range used in this study), the mean size of mussels found in middens was closely related to octopus size (mean length = 1.93 octopus Wt0.6’J; II = 23; Y= 0.88; p < 0.01, calculated from data in Fig. 12).
Regressions of L,,i,,50 and of mean size of mussels in middens against octopus weight have unequal slopes (t-test, p>O.O5) and cross at an octopus weight of ~200 g. Octopuses smaller than 200 g took mussels of less than L,,,,,50, while octopuses of 200-500 g took mussels larger than L,,,,,50, but smaller than the maximum size avail- able in the field.
4. Discussion
The feeding behaviour of 0. vulgaris is very plastic and minimises the time required to open mussels. Like lobsters (Griffiths & Seiderer, 1980; Robles et al., 1990) and crabs (Du Preez, 19X4), octopuses have two methods for opening bivalves. An initial attempt is made to pull mussels open, whatever their size, using the tentacles (the structure and mechanics of octopus suckers are discussed by Kier & Smith, 1990 and
Smith, 1991) and the mussel is only drilled if force fails. Octopus saliva can initiate external digestion of crabs (Nixon, 1984; Boyle, 1990) but the main effect in the case of mussels seems to be to paralyse the adductor muscles which hold the shell valves closed (see Cariello & Zanetti, 1977). This effect allows the shell to be pulled open more easily and even the very largest mussels can be opened eventually by drilling. European 0. vulgaris regularly drill the bivalve Mytilus galloprovincialis through the umbo, close

200 C.D. McQuuid 1 J. Exp. Mar. Bid. Ed. 177 (1994) 187-202
to the myostracum of the anterior adductor muscle (Nixon & Maconnachie, 1988).
Perna lacks an anterior adductor muscle (Siddall, 1980) and in the case of South
African O.vufguris feeding on Perna pernu, initial holes near the umbo are very rarely
complete. Mussels are normally drilled in the mid-dorsal region (Fig. 2) which is not the thinnest part of the shell, but is in the region of the heart.
As mussel length increases, body mass, and thus energetic value, increase. However, so too do both the diameter of the posterior adductor muscles, which hold the valves of the shell closed, and the thickness of the shell, both of which render the mussel more difficult to open using either method of attack. Pulling mussels open is energetically
expensive, but each pull lasts for only a few seconds and the much longer handling time required for drilling over-rides this difference. So, although the absolute energy content of large mussels is high, they produce low E/H values because they require drilling
(Fig. 9). Elner & Hughes (1978) found that, for crabs eating mussels. the time required to glean
1 g of tissue (T,,) was low for large prey due to satiation, and for small prey due to difficulty in manipulation. In the case of 0. vulgaris, T,, is almost constant for all mussel lengths, indicating that satiation and manipulation are not problems for such large,
dextrous predators. Small mussels can be opened and eaten quickly, but contain relatively little meat so that both E and E/H are low. Thus E/H is low for large prey due to high values of H, and for small prey, because E is low. E/H was greatest for the largest size classes which could be forced open (Fig. 9). The energetically optimum diet was predicted to be those size classes just below L,,i,,50. L,,,i,,50 depends on octopus size, consequently different size classes are optimal for individual octopuses.
Choice experiments show that octopuses select particular size classes and larger octopuses generally take larger mussels. Further evidence for active selection of spe- cific sizes is found when minimum prey sizes are considered. Hughes (1980) observed
that large predators may not take small prey either because they are not energetically worthwhile, or because they are too difficult to manipulate. If larger 0. vulguris are considered, both the maximum and minimum sizes of prey selected increase (Figs. 11 and 12). However, 0. vulgaris will take very small prey if starved (pers. obs.), indicating that they can take small mussels, but choose not to.
The prey size classes favoured in the laboratory include those with maximum E/H
values and the mean length of mussels taken by each octopus was equal to, or just below the L,,,,,50 for an octopus of that size. This observation conforms with the predictions of optimal foraging theory. There was, however, considerable variation in the size classes taken and this may be related to the difficulty of predicting which method of attack will be required. The force required to pull a mussel open is significantly cor- related with its length (Fig. l), but has a very low predictability (2 = 0.34).
Mather (1991) has found that there is no relationship between the size of a prey item and the likelihood that 0. vulguvk will consume it in situ during a foraging bout, rather than carrying it back to the den. Thus, Smale & Buchan’s data (Fig. 12) give a good indication of the size classes of mussels eaten under field conditions. These data show that, in the field, octopuses take different size classes of mussels from those eaten in the laboratory, possibly because their behaviour is different in laboratory conditions or because there are constraints on actual availability of prey size classes in the field. For

C.D. McQuaid 1 J. Exp. Mar. Biol. Ecol. I77 (1994) 187-202 20 I
example, Tokeshi (1989) found that small sun-stars, Heliaster, avoid large mussels as
they are more vulnerable to wave action when attacking them and Mather and O’Dor
(1991) suggest that the foraging behaviour of juvenile 0. vulgaris is influenced by the
threat of predation. Smale and Buchan’s field data cannot be compared with the pre- dictions of optimal foraging theory for two reasons:
(a) there is no information on the time required to remove mussels from the rocks in the field and this could be an important component of handling time (i.e. optimal
size classes may be different in the field). (b) there is no information on the availability of different sizes of prey in the field.
However, prey size availability was very uniform within Smale & Buchan’s study area and there is no reason to suspect that it differed for different sizes of octopus (M.J.
Smale, pers. comm.). One can, therefore, tentatively compare prey selection by octo- pusus of different sizes within their study. Octopuses of over 500 g all took approxi- mately the same size classes of mussels in the field (Fig. 12). These are the biggest
mussels generally available. For smaller octopuses, of the size range used in this study, the mean length of mussel taken was closely correlated to octopus weight, but differed from laboratory results. Octopuses of > 200 g took larger mussels than in the labora- tory while octopuses of ~200 g took smaller mussels than in the laboratory. One constraint on size selection in the field could be the force required to remove mussels from the rocks. In the laboratory this is less than the force needed to open them, but it increases rapidly with the size of the mussel (Fig. 1). More importantly, mussels
produce more byssus threads under wave exposed conditions (Hepburn et al., 1979) and so laboratory measures of this force are likely to be extremely conservative. For small octopuses, the major constraint on prey selection in the field may be the ability to remove mussels from the rocks. This ability was not measured, but very small octopuses in the laboratory cannot tear large mussels free. The fact that, under natural conditions, octopuses of <500 g take prey smaller than the maximum size available could reflect their inability to collect large mussels.
5. Acknowledgements
I am grateful to S.G.A. Compton and R.N. Hughes for constructive comments on the manuscript and to M. Balarin and J.B. Lang for help in capturing and caring for octopuses. Permission to reproduce Fig. 12 from Smale & Buchan (1981) was kindly provided by the authors and Springer Verlag. Financial support was provided by the Joint Research Committee of Rhodes University.
6. References
Abrams. P.A., 1982. Functional responses of foragers. Am. ME, Vol. 120, pp. 382-390.
Alexander, R.. t968. Animal mechunics. Sidgwick & Jackson, London, 346 pp. Boyle. P.R., 1990. Prey handling and salivary secretions in octopuses. In, Trophic relationships in the marine

eurironnwzt, edited by M. Barnes & R.N. Gibson, Proceedings of the 24th European Marine Biolog!
Symposium, Aberdeen University Press. Aberdeen, pp. 541-552.
Buchan, P.R. & M.J. Smale, 1981. Estimates of biomass, consumption and production of Ocrop~s ~ulgori.~
Cuvier of%’ the east coast of South Africa. Oceurzugr. Res. Inst. Itwest. Rep.. Vol. 50, pp. I-9.
Burrows, M.T. & R.N. Hughes. 1990. Variation in growth and consumption among individuals and popu-
lations of dogwhelks, &uiucellu ~~~pj~~z~.~: a link between foraging behaviour and fitness. J. nt~ilrt. Ec<zf., Vol.
59% pp. 713-742.
Cariello, L. & L. Zanetti, 1977. Alpha and beta-cephalotoxin: two paraiysing proteins from posterior sali-
vary glands of Octqzrs v~1gf~?s. Ct>ntp. Biode?>z. Phmid.. Vol. 57. pp. 169- 173.
Du Precr. H.H.. 1984. Molluscan predation by O~uli/~,s punc’trtru.~ (De Haan) (Crustacca: Brachyura: Por-
tunidac). J. .E\-l,. MU. Bid. Ed. Vol. 83, pp. 55-71.
Elner, R.W. & R.N. Hughes, 1978. Energy maximisation in the diet of the shore crab, Curcimv ~m~cw~~~. J.
,l,lb?Z. E&.. Vol. 47, pp. 103-t 16.
Griffith. C.L. & P.A.R. Hockey, 1987. A model describing the interactive roles of predation. competition
and tidal elevation in structuring mussel populations. S. Ati. J. Mar. Sci., Vol. 5, pp. 547-556.
Grisths, C.L. & J.L. Seiderer, 1980. Rock-lobsters and muss& - linlitations and prcfcrcnces in a prcq-
predator interaction. J. Exp. Mar. Bid. Ed., Vol. 44, pp. 95-105.
Hepburn. H.R.. G.S. Sinclair, M.R. Davidoff. E.A. Moss, H.D. Chandler. Sr S. Ripley. 1979. Of hyssus
threads and their tenacity. S. .4f/. J. Sri., Vol. 75. pp. 187-189.
Hughes, R.N., 1980. Optimal foraging thc.il L :n !he marine context. Occ~crnqpr. Mm. Bid. Am Rev., Vol.
IX. pp. 423-481.
Kicr. W.M. & A.M. Smith, 1990. The morphology and mechanics of octopus suckers. Biol. Bull.. Vol. 17X.
pp. 126-136.
Mathcr. J.A., 1991. Foraging, feeding and prey remains in middens of juvenile ~~~(~~~~.s ~~~/~~~~j.~ (Moliusca: C~phalopoda). J. .&ml. ttmd., Vol. 224. pp. 27-39.
Mathcr. J.A. & R.K. O’Dor, 1991. Foraging strategies and nlzdation risk shape the natural history of’ju-
venile Ocro~us wlguris. Bull. Mur. Sci., Vol. 49. pp. 256-269.
Nixon, M.. 1984. Is there external digestion by OCII)I)US? J. Zool. Lo&., Vol. 202, pp. 441-447.
Nixon, M. & E. Maconnachic, 1988. Drilling by OC~XI.F vulgururis (Mollusca: Ccphalopoda) in the Mediter-
ranean. J. Zoof. Land., Vol. 216. pp. 687-7 16.
Pykc. G.H., 1984. Optimal foraging theory: a critical review. Anrr. Rev. Erof. S~sr., Vol. 15, pp. 523-575.
Pyke. G.H.. H.R. Pulliam, & EL. Charnov. 1977. Optimal foraging: a selective review of theory and tests.
Q. Res. Bid., Vol. 52, pp. 137-154.
Robles, C.. D. Sweetnam & J. Eminike. 1990. Lobster predation on mussels: shore-level differences in prey
vulner~Ibility and predator preference. Ecofq~, Vol. 71, pp. 1564-1577.
Schoener. T.W., 1987. A brief history of optimal foraging ecology. In. F&&R hehrrviour, edited by A.C.
Kamil, J.R. Krebs & H.R. Pulliam. Plenum Press, New York. pp. 5-67.
Siddall, S.E., 1980. A clarification of the genus Perrw. Buff. Mar. Sci.. Vol. 30. pp. 858-870.
Sih, A.. 19X4. Optimal bchaviour and density-dependent predation. .4,~1. NUT.. Vol. 123. pp. 314-326.
Smale, M.J. & P.R. Buchan. 1981. Biology of Ocropus vtrlguris off the east coast of South Africa. Mm. Bid., Vol. 65. pp. I-12.
Smith, A.M.. 1991. Negative pressure generated by octopus suckers: a study of the ten& strength of water
in nalure. J. Esp. &of., Vol. 157, pp. 257-171.
Tokeshi, M., 1989. Development of a foraging model for a field population of the South American sun-star
Heficrster ~~liu~l~hl~.~. J. Aniru. Ecof.. Vol. 58, pp. 139-206.
Copyright © 2022 FDOKUMEN




















