
Environmental associations of shortfin squid Illex argentinus (Cephalopoda: Ommastrephidae) in the...
16
Fisheries Research 76 (2005) 401–416 Environmental associations of shortfin squid Illex argentinus (Cephalopoda: Ommastrephidae) in the Northern Patagonian Shelf Gast´ on Bazzino a,∗ , Renato A. Qui ˜ nones b , Walter Norbis a a Direcci´ on Nacional de Recursos Acu ´ aticos (DINARA), Constituyente 1497, Montevideo, Uruguay b Centro de Investigaci´ on Oceanogr´ afica en el Pacif´ ıco Sur Oriental (COPAS) & Departamento de Oceanograf´ ıa, Universidad de Concepci´ on, Casilla 160-C, Concepci ´ on, Chile Received 3 March 2005; received in revised form 19 July 2005; accepted 26 July 2005 Abstract We identified environmental associations (depth, bottom temperature and bottom salinity) of the shortfin squid Illex argentinus in the Northern Patagonian Shelf. Data are from 15 research bottom trawl surveys conducted in the Argentine–Uruguayan Common Fishing Zone (AUCFZ) between 1985 and 1998. Squid distribution in the surveyed area showed significant associations (p < 0.05) mostly with depth and bottom temperature. Bathymetric associations of squid detected in March–April, May–June and July–August cruises revealed a preference of squid for deep depths (80–280 m, intermediate and external shelf) and cool bottom temperatures (4–10 ◦ C, subantarctic water) within the survey area. In contrast, the results observed during November–December cruises showed a preference of squid for shallower depths (70–90 m) and warmer bottom temperatures. The majority of two- way environmental analyses showed significant associations, suggesting the existence of a strong correlation between the environmental factors used in our analyses. The identification of significant associations between squid distribution and bottom temperature seem to be related to the presence of strong thermal gradients within the study area. This could be supporting the existence of an association between I. argentinus with the Brazil–Malvinas confluence front and also with the shelf break front. © 2005 Elsevier B.V. All rights reserved. Keywords: Shortfin squid; Illex argentinus; Environmental associations; Northern Patagonian Shelf ∗ Corresponding author at: Centro de Investigaciones Biol´ ogicas del Noroeste (CIBNOR), Programa de Doctorado – Ecologia Pes- quera, Mar Bermejo 195, Colonia Playa Palo de Santa Rita, C.P. 23090, La Paz, Baja California Sur (BCS), M´ exico. Tel.: +52 612 1198998. E-mail address: [email protected] (G. Bazzino). 1. Introduction The shortfin squid Illex argentinus (Castellanos, 1960) is a neritic-oceanic species distributed along the continental shelf and slope of Southwestern Atlantic Ocean (SWAO) between latitudes 20 ◦ S and 55 ◦ S, occurring more abundantly in the area 35–52 ◦ S (Brunetti, 1988; Haimovici et al., 1998). It is the 0165-7836/$ – see front matter © 2005 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2005.07.005
Transcript of Environmental associations of shortfin squid Illex argentinus (Cephalopoda: Ommastrephidae) in the...

Fisheries Research 76 (2005) 401–416
Environmental associations of shortfin squidIllex argentinus (Cephalopoda: Ommastrephidae)
in the Northern Patagonian Shelf
Gaston Bazzinoa,∗, Renato A. Quinonesb, Walter Norbisa
a Direccion Nacional de Recursos Acuaticos (DINARA), Constituyente 1497, Montevideo, Uruguayb Centro de Investigacion Oceanografica en el Pacifıco Sur Oriental (COPAS) & Departamento de Oceanografıa,
Universidad de Concepcion, Casilla 160-C, Concepcion, Chile
Received 3 March 2005; received in revised form 19 July 2005; accepted 26 July 2005
Abstract
We identified environmental associations (depth, bottom temperature and bottom salinity) of the shortfin squidIllex argentinusin the Northern Patagonian Shelf. Data are from 15 research bottom trawl surveys conducted in the Argentine–UruguayanCommon Fishing Zone (AUCFZ) between 1985 and 1998. Squid distribution in the surveyed area showed significant associations(p < 0.05) mostly with depth and bottom temperature. Bathymetric associations of squid detected in March–April, May–June andJuly–August cruises revealed a preference of squid for deep depths (80–280 m, intermediate and external shelf) and cool bottomtemperatures (4–10◦C, subantarctic water) within the survey area. In contrast, the results observed during November–Decemberc ity of two-w etween thee nd bottomt porting thee ront.©
K
dq2T
,ongtern
2
0
ruises showed a preference of squid for shallower depths (70–90 m) and warmer bottom temperatures. The majoray environmental analyses showed significant associations, suggesting the existence of a strong correlation bnvironmental factors used in our analyses. The identification of significant associations between squid distribution a
emperature seem to be related to the presence of strong thermal gradients within the study area. This could be supxistence of an association betweenI. argentinus with the Brazil–Malvinas confluence front and also with the shelf break f2005 Elsevier B.V. All rights reserved.
eywords: Shortfin squid;Illex argentinus; Environmental associations; Northern Patagonian Shelf
∗ Corresponding author at: Centro de Investigaciones Biologicasel Noroeste (CIBNOR), Programa de Doctorado – Ecologia Pes-uera, Mar Bermejo 195, Colonia Playa Palo de Santa Rita, C.P.3090, La Paz, Baja California Sur (BCS), Mexico.el.: +52 612 1198998.
E-mail address: [email protected] (G. Bazzino).
1. Introduction
The shortfin squidIllex argentinus (Castellanos1960) is a neritic-oceanic species distributed althe continental shelf and slope of SouthwesAtlantic Ocean (SWAO) between latitudes 20◦S and55◦S, occurring more abundantly in the area 35–5◦S(Brunetti, 1988; Haimovici et al., 1998). It is the
165-7836/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.fishres.2005.07.005

402 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
most important cephalopod species in the SWAO,not only because of its wide distribution and largeabundance (annual catches attained as much as500,000–750,000 ton;FAO, 1997), but for its ecolog-ical role in the ecosystem due to trophic interactions(Santos and Haimovici, 2000).
I. argentinus is an opportunistic species (strategyr; Brunetti, 1988) having rapid growth, a short lifecycle (annual approx.;Arkhipkin, 1990), opportunis-tic feeding (Ivanovic and Brunetti, 1994; Bazzinoand Quinones, 2001), and a high degree of intra-population differentiation (Brunetti, 1988; Nigmat-ullin, 1989; Arkhipkin, 1993, 2000). Furthermore, it isa species with seasonal migrations (Arkhipkin, 1993)from cold-water feeding areas (Malvinas/FalklandCurrent) to warmer spawning areas (Brazil Cur-rent). Some of these characteristics are relevantfor the management ofI. argentinus’ fishery inthe SWAO (Caddy, 1983; Bazzino and Quinones,1999).
Neritic-oceanic squid species (ommastrephids; e.g.,I. argentinus) tend to distribute themselves in relation tolarge ocean current systems (O’Dor and Coelho, 1993)and in relation to the fronts that are generated wherewater masses with different hydrographic characteris-tics converge (Caddy, 1983). In fact, the life cyclesof several squid species are carefully synchronizedwith the movements of the water masses they inhabit(Hatanaka et al., 1985; Rodhouse et al., 1992; Bakunand Csirke, 1998; Dawe et al., 2000; Anderson andR aleo nver-g ttie 8;W
O( ug-g bun-da s,dn cef al.,1 -i t ism ent( slo,2
The Northern Patagonian Shelf is a highly com-plex region from an oceanographic point of view.The circulation in the upper layers (0–2000 m) isdominated by the opposing flow of the Brazil Cur-rent and the Malvinas/Falkland Current (Peterson,1992). The Brazil Current (warm) moves southwardalong the edge of South America, whereas the Malv-inas/Falkland Current (cold) flows northward along thecontinental shelf, originating as a branch of the Antarc-tic Circumpolar Current (Peterson and Stramma,1991).
The Brazil and Malvinas/Falkland Currents meetaround the 33–39◦S area, causing an important ther-mohaline front that separates the subtropical watersfrom the subantarctic waters (Brazil–Malvinas Con-vergence Front;Legeckis and Gordon, 1982; Gordon,1989; Olson et al., 1988). Strong thermal and salinegradients can be observed in this convergence area,due to the interaction between both currents, but alsoto the influence of the Patagonian Current (also knownas the western branch of the Malvinas/Falkland Cur-rent) and the discharge of the Rıo de La Plata, whichgenerates high levels of primary production (Hubold,1980a,b; Haimovici et al., 1998). Productivity is fur-ther favored by another front, located over the shelfbreak that indicates the border between shelf waters(Patagonian Current and Rıo de La Plata discharge)and waters of Malvinas/Falkland Current (Bakun andParrish, 1991).
Considering the ecological and commercial impor-t plor-ih ae slo,2 per-s thed ec tionso
enso ianS ingg1 r-t otht ingfl ter( wn-
odhouse, 2001) and associated with the mesoscceanographic processes that are found in these coence zones or fronts (Rodhouse et al., 1996; Bertolot al., 1996; Gonzalez et al., 1997; Brunetti et al., 199aluda et al., 2001a).The variability of main current systems in SWA
Brazil and Malvinas/Falkland Current) has been sested as one of determining factors in the aance and distribution ofI. argentinus (Andersonnd Rodhouse, 2001). According to some authoruring spawning and first life stages,I. argenti-us is associated with Brazil–Malvinas confluenront (Hatanaka, 1988; Leta, 1992; Rodhouse et995; Haimovici et al., 1998), whereas dur
ng maturation, growth, and feeding stages, iostly associated with Malvinas/Falkland Curr
Rodhouse et al., 1995; Laptikhovsky and Reme001).
ance of this squid species in the SWAO, studies exng the environmental associations ofI. argentinusave been intensified (Brunetti et al., 1998; Waludt al., 1999, 2001a,b; Laptikhovsky and Reme001). Nevertheless, a high degree of uncertaintyists as to which environmental variables influenceistribution ofI. argentinus in certain stages of its lifycle, such as during the reproductive concentraver the Northern Patagonian Shelf.
According to the existing records, the specimf I. argentinus captured in the Northern Patagonhelf correspond mostly to the winter shelf spawnroup (Bonaerensis–North Patagonic stock;Brunetti,988; Leta, 1989). This stock is the most impo
ant from a fishing perspective, being caught by bhe Uruguayan and Argentine jigging and trawleets, principally during austral autumn and winApril–September). The presence of the spring spa

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 403
ing group and the summer spawning group in theNorthern Patagonian Shelf is only detected duringaustral spring (October–December) (Brunetti, 1981;Hatanaka, 1986).
In this context, the objective of this study is toidentify the environmental associations (depth, bottomtemperature and bottom salinity) of squidI. argenti-nus in the Northern Patagonian Shelf, specificallywithin the Argentine–Uruguayan Common FishingZone (AUCFZ).
2. Materials and methods
Squid catch and hydrographic data were obtainedfrom 15 research bottom trawl surveys carried outbetween 1985 and 1998 (Table 1) in the Northern Patag-onian Shelf within the Argentine–Uruguayan Com-mon Fishing Zone (AUCFZ, 34◦00′–39◦30′S; Fig. 1).The surveys were based on a stratified random design(Erhardt et al., 1977) with 24 strata defined by latitudeand depths intervals (Fig. 1).
Standard bottom trawls were performed in allcruises with an Engel trawl (100 mm mesh in the wingsand 60 mm mesh in the cod ends, 4 m vertical openingand 15 m horizontal aperture). Each tow lasted 30 min
Table 1R tago-n onF
C
MMM
MMM
JJJ
SSS
NND
of effective bottom trawling time at an average speed of3 knots. All tows were performed during daytime whenshortfin squid is located near ocean bottom. The catchof I. argentinus was recorded after each tow and thefollowing oceanographic parameters were registered:tow depth (m), bottom temperature (◦C) and bottomsalinity.
The depth (m) was estimated by averaging the start-ing and final tow depths. Bottom temperature and salin-ity were obtained from water samples collected withNansen bottles in those surveys performed within theperiod 1985–1989; and directly using a CTD in theperiod 1994–1998.
2.1. Spatial distribution of squid abundance
I. argentinus biomass estimates per strata were cal-culated in each research cruise through swept areamethod (Alverson and Pereyra, 1969). We classifiedstrata with high and low squid abundance in orderto analyze the spatial distribution within the AUCFZ.The strata with high squid abundance were thosewhose biomass was equal or greater than the aver-age biomass obtained with all the strata during eachcruise.
2.2. Identification of habitat associations
The method ofPerry and Smith (1994)was appliedto determine the existence of significant environmentala nS asso-c UE( dy,t idc -t thea -t
em ns:( on-m edw if-f eduref en-t h(
esearch bottom trawl surveys performed in the Northern Paian Shelf, specifically within the Argentine–Uruguayan Commishing Zone (AUCFZ), during the period 1985–1998
ruise Year
arch–April 1987arch 1988arch–April 1998
ay–June 1985ay–June 1986ay–June 1995
une–July 1988une–July 1994uly–August 1989
eptember 1986eptember–October 1988eptember–October 1994
ovember–December 1985ovember 1986ecember 1987
ssociations inI. argentinus over Northern Patagoniahelf. These authors developed a non-parametriciation test for an environmental variable and the CPcatch per unit of effort) of a given species. In our stuhe CPUE ofI. argentinus was represented by squatch per fishing tow (CPUEsquid= catch/30 min botom trawling). The environmental variables used innalyses were depth, bottom temperature (◦C) and bot
om salinity.The method ofPerry and Smith (1994)calculates th
aximum absolute difference between two functioi) the cumulative frequency distribution of the envirental variablef(t), and (ii) the same function weightith the squid CPUEg(t). The maximum absolute d
erence is used as a statistical test in a random procor evaluating the existence of significant environmal associations. For further details seePerry and Smit1994).
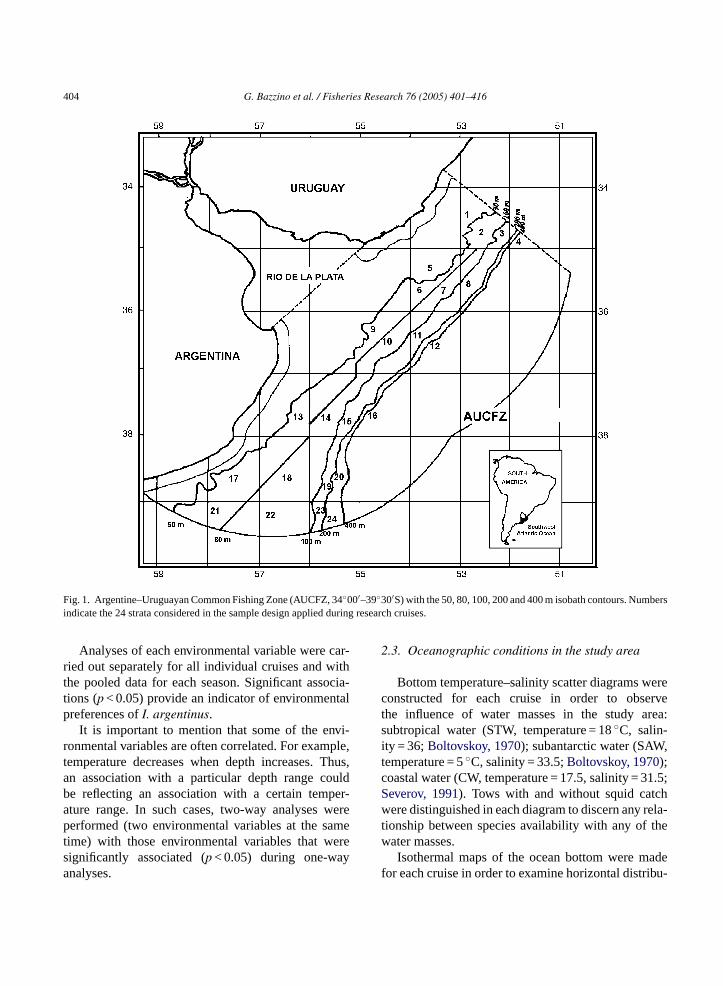
404 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Fig. 1. Argentine–Uruguayan Common Fishing Zone (AUCFZ, 34◦00′–39◦30′S) with the 50, 80, 100, 200 and 400 m isobath contours. Numbersindicate the 24 strata considered in the sample design applied during research cruises.
Analyses of each environmental variable were car-ried out separately for all individual cruises and withthe pooled data for each season. Significant associa-tions (p < 0.05) provide an indicator of environmentalpreferences ofI. argentinus.
It is important to mention that some of the envi-ronmental variables are often correlated. For example,temperature decreases when depth increases. Thus,an association with a particular depth range couldbe reflecting an association with a certain temper-ature range. In such cases, two-way analyses wereperformed (two environmental variables at the sametime) with those environmental variables that weresignificantly associated (p < 0.05) during one-wayanalyses.
2.3. Oceanographic conditions in the study area
Bottom temperature–salinity scatter diagrams wereconstructed for each cruise in order to observethe influence of water masses in the study area:subtropical water (STW, temperature = 18◦C, salin-ity = 36; Boltovskoy, 1970); subantarctic water (SAW,temperature = 5◦C, salinity = 33.5;Boltovskoy, 1970);coastal water (CW, temperature = 17.5, salinity = 31.5;Severov, 1991). Tows with and without squid catchwere distinguished in each diagram to discern any rela-tionship between species availability with any of thewater masses.
Isothermal maps of the ocean bottom were madefor each cruise in order to examine horizontal distribu-

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 405
tion of bottom temperature (presence of strong thermalgradients).
3. Results
3.1. Spatial distribution of squid abundance
The spatial distribution of squid abundance withinthe AUCFZ revealed a considerable seasonal variabil-ity between the research cruises analyzed (Fig. 2).The strata showing high squid abundance duringMarch–April cruises were mainly located in the south-ern sector of the AUCFZ and at greater depths(100–400 m). In May–June cruises the strata withhigh squid abundance were located more northern inthe AUCFZ and at intermediate depths (80–200 m).This pattern became more evident in June–July andSeptember–October cruises. On the other hand, dur-ing November–December cruises those strata with highsquid abundance were found in the southern sector ofthe AUCFZ and at shallower depths (50–100 m).
3.2. Identification of habitat associations
March–April: significant associations (p < 0.05)were found for I. argentinus with depth and bot-tom temperature in all the analyses done with indi-
vidual March–April cruises and also with pooledMarch–April data (Table 2). On the other hand, theonly cruise that reflected a significant association withbottom salinity was March–April 1998.
Squid were strongly associated (p < 0.01) with: (1)deep depths, as 98% of the population occurred wherebottom depths were deeper than 80 m (mostly withinthe range 80–280 m,Fig. 3), which represented thedeepest 50% of the survey area; (2) cooler bottom tem-peratures, as 95% of the population occurred within therange 4.0–8.0◦C, at the lower 40% of available bottomtemperatures (Fig. 4).
The results obtained with the two-way analysesshowed a significant association between squid withthe simultaneous distribution of depth and bottom tem-perature in the study area (p < 0.05;Table 3).
May–June: analyses of May–June pooled datarevealed significant associations (p < 0.05) for I.argentinus with depth and bottom temperature(Table 2). However, these associations were not con-sistent in the individual cruises analyses. Only thecruise of May–June 1986 reflected significant associ-ations with all the environmental variables, includedbottom salinity. No significant environmental associ-ations were observed in the other May–June cruises(Table 2).
In those analyses that reflected significant environ-mental associations (May–June 1986 and May–June
F CFZ. B ndance.W
ig. 2. Spatial distribution ofI. argentinus biomass within the AUhite strata: no squid. For further details see Section2.
lack strata: high squid abundance. Grey strata: low squid abu

406 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Fig. 3. Cumulative frequency distributions of observed depth (solid line) and squid CPUE weighted data (dashed line) from (A) March–April,(B) May–June, (C) June–August, (D) September–October and (E) November–December cruises.

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 407
Fig. 4. Cumulative frequency distributions of observed bottom temperature (solid line) and squid CPUE weighted data (dashed line) from (A)March–April, (B) May–June, (C) June–August, (D) September–October and (E) November–December cruises.

408 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Table 2Test results ofPerry and Smith (1994)one-way analyses
Table entries are probability (p) values for having a test statistic, from the Monte Carlo simulation, equal or greater than the test statistic observed.* Significant environmental associations (p < 0.05).
pooled data), squid showed preferences for: (1) deepdepths, as 74% of the population occurred deeper than80 m (mostly within the ranges 80–200 and 360–400 m,Fig. 3), which represented the deepest 50% of the avail-able depths; (2) cooler bottom temperatures, as 80% ofthe squid occurred within the range 5–10.5◦C, at thecoldest 50% of available bottom temperatures (Fig. 4).
No significant associations were found betweensquid and the simultaneous distribution of depthand bottom temperature in the two-way analyses ofMay–June pooled data (Table 3). For this reason wecompared the results of one and two-way analyses.
During one-way analyses, the maximum absolute dif-ference between the functionsf(t) andg(t) was observedat 140 m in the case of depth and at 9.5◦C in the caseof bottom temperature. In two-way analyses, the max-imum absolute difference between both functions wasobserved at 140 m for depth, which coincided withthe one-way results, whereas the bottom temperaturechanged to 22.0◦C.
June–August: the analyses of June–August pooleddata revealed significant associations (p < 0.05) of I.argentinus with depth, bottom temperature and bottomsalinity (Table 2). However, these significant associ-

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 409
Table 3Test results ofPerry and Smith (1994)two-way analyses
Bottom temperature Bottom salinity
March–AprilDepth <0.01* –Bottom temperature –
May–JuneDepth 0.16 –Bottom temperature –
June–AugustDepth 0.07 <0.01*
Bottom temperature <0.01*
September–OctoberDepth – –Bottom temperature –
November–DecemberDepth <0.01* –Bottom temperature –
The analyses were performed with the pooled data and only withthose environmental variables that reflected significant associationsduring one-way analyses. Table entries are probability (p) values forhaving a test statistic, from the Monte Carlo simulation, equal orgreater than the test statistic observed.
* Significant environmental associations (p < 0.05).
ations were not consistent during the analyses withindividual cruises.
Squid were significantly associated with: (1) deepdepths, as 78% of the population occurred where bot-tom depths were deeper than 80 m (mostly within therange 80–150 m,Fig. 3), which represented the deepest50% of the survey area; (2) cooler bottom temperatures,as 87% of the squid population occurred within therange 5.5–9.5◦C, which represented the lower 45% ofavailable bottom temperatures (Fig. 4); (3) lower bot-tom salinities, as 91% of squid population occurredwithin the range 32.9–33.7 at the lower 50% of avail-able bottom salinities in study area (Fig. 5).
All two-way analyses performed with June–Augustpooled data registered strong associations (Table 3).Squid showed an effective association with the simul-taneous distribution of depth, bottom temperature andbottom salinity.
September–October: individual and pooled dataanalyses for September–October cruises did not regis-tered significant environmental associations (Table 2).
November–December: analyses of November–December pooled data revealed significant associations(p < 0.05) forI. argentinus with depth and bottom tem-
perature (Table 2). However, these significant associa-tions were not consistent with the individual analysesof November–December cruises.
Squid showed a strong association (p < 0.01) with:(1) shallow depths, as 97% of the population occurredwithin the range 70–90 m which represented the shal-lowest portion of the surveyed area (Fig. 3); (2)warmer bottom temperatures, as 80% of the populationoccurred within the range 9.5–12◦C which representedthe warmer 50% of available bottom temperatures inthe surveyed area (Fig. 4).
Two-way analyses performed with the November–December pooled data reflected significant associa-tions (p < 0.05) betweenI. argentinus with the simul-taneous distribution of depth and bottom temperature(Table 3). Both environmental variables were effec-tively associated with November–December squid dis-tribution in the study area.
Annual variability: environmental significant asso-ciations (p < 0.05) were found forI. argentinus withdepth and bottom temperature all across the yearexcept during September–October cruises (Table 2).The associations detected in March–April, May–Juneand July–August cruises revealed a preference of squidfor deep depths and cool bottom temperatures withinthe survey area. In contrast, the results obtained duringNovember–December cruises showed a preference ofsquid for shallower depths and warmer bottom temper-atures.
The majority of two-way analyses reflected signifi-c eousds ntala ses(
3
cteds them era-t easeo picalw andN sr towsw ass( nt).
ant associations between squid with the simultanistribution of environmental factors (Table 3). Thetronger (p < 0.01) and more consistent environmessociations were found during March–April cruiTable 2).
.3. Oceanographic conditions in the study area
Oceanographic conditions in the study area refleeasonal changes which were associated withovements of different water masses. Bottom temp
ure–salinity scatter diagrams reflected an incrf subantarctic water and a decrease of subtroater in the study area between March–Aprilovember–December cruises (Fig. 6). These diagram
evealed a close association between those fishingith positive squid catch and subantarctic water m
Malvinas/Falkland Current and Patagonian Curre

410 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Fig. 5. Cumulative frequency distributions of observed bottom salinity (solid line) and squid CPUE weighted data (dashed line) from (A)March–April, (B) May–June, (C) June–August, (D) September–October and (E) November–December cruises.

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 411
Fig. 6. Bottom temperature–salinity scatter diagrams from (A) March–April, (B) May–June, (C) June–August, (D) September–October and (E)November–December cruises. SAW: subantarctic water; STW: subtropical water; CW: coastal water. Black symbols: fishing tows with squidcatch. Cross symbols: fishing tows without squid catch.

412 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Fig. 7. Spatial distribution of bottom temperature (isotherms;◦C) in the surveyed area during the following cruises: (A) March 1988; (B)May–June 1986; (C) June–July 1988; (D) July–August 1989; (E) September–October 1988; (F) December 1987.

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 413
Bottom isothermal maps of some monthly cruisesshowed strong thermal gradients in the study area(March–April cruises,Fig. 7), reflecting the presenceof Brazil–Malvinas confluence front and shelf breakfront.
4. Discussion
The results obtained using the method ofPerry andSmith (1994)indicated significant environmental asso-ciations of I. argentinus in the Northern PatagonianShelf, principally with depth and bottom temperature.However, the majority of these associations were notconsistent across cruises carried out in the same sea-son but in different years. A possible explanation of thisinconsistency is that shortfin squid may have a broadtolerance to environmental factors within the availablerange in the study area, which correspond to a smallproportion of the overall geographic range of species.The same type of inconsistency was also observed byBrodziak and Hendrickson (1999)in the northwesternAtlantic Ocean for another squid species of the genusIllex (Illex illecebrosus).
Depth and bottom temperature were the environ-mental variables that registered the higher number(n = 11) of significant associations (p < 0.05) with squiddistribution. The influence of these environmental vari-ables was observed all around the year except inSeptember–October cruises. In contrast, the influencee mem por-t stentw losea
pthi sesr atedo m).A rvedw uidb itht ra-t tra-t lledB ;Bn er
(Brunetti, 1988; Leta, 1989). These concentrations aremade up of adult specimens that have initiated their sex-ual maturation or are sexually mature, a prerequisite foremigration toward spawning areas in deeper waters ofthe oceanic region outside the AUCFZ (Brunetti, 1988;Hatanaka, 1986, 1988).
On the other hand, bathymetric associations of squidduring November–December cruises showed prefer-ence for shallower depths (70–90 m) which coincideswith the bathymetric distribution of pre-reproductiveconcentrations of the summer spawning group andreproductive concentrations of the spring spawninggroup (Brunetti, 1981; Hatanaka, 1986). In fact, squidbiomasses during November–December cruises weremostly concentrated in those strata with depths between50 and 100 m.
The significant associations registered with bot-tom temperature in March–April, May–June andJuly–August cruises showed a preference of squidfor cooler temperatures (4–10◦C) included within therange known for the subantarctic water mass (Severov,1991). This suggests an association between squid dis-tribution and the influence of subantarctic water massin the study area. The existence of this link was alsodemonstrated with the results observed in the bottomtemperature–salinity scatter diagrams which revealeda close association between those fishing tows withpositive squid catch and the subantarctic water mass(Malvinas/Falkland Current and Patagonian Current).
The lack of significant environmental associationsd finals ingg iono
vedb ceo esses( -i t thes ents( nta-ta os-s tingt ausen oci-a ationb yses.
xerted by bottom salinity was only detected in soonths, mostly during June–July cruises. The im
ance of depth and bottom temperature is consiith demersal behavior of squid and reflects a cssociation with bottom hydrographic features.
The significant associations detected with den March–April, May–June, and July–August cruieflect a preference of squid for those regions locver the intermediate and external shelf (80–280similar depth preference range was also obse
ith the analyses of the spatial distribution of sqiomasses within the AUCFZ. This coincides w
he bathymetric distribution mentioned in the liteure for pre-reproductive and reproductive concenions of the winter shelf spawning group, also caonaerensis–North Patagonic stock (Brunetti, 1988runetti et al., 1997; Haimovici et al., 1998) observedorth of latitude 43◦S between April and Septemb
uring September–October maybe is reflecting thetages in the life cycle of the winter shelf spawnroup which is completing the reproductive migratutside the AUCFZ toward spawning areas.
Our results coincide partially with those obsery Waluda et al. (2001a), who analyzed the influenf bathymetry and mesoscale oceanographic procthermal gradients) onI. argentinus around the Malvnas/Falkland Islands. These authors found thaquid was directly associated with thermal gradimainly observed over the 200 m isobath, represeive of the shelf break front;Bertolotti et al., 1996),nd indirectly with depth. In our study, it was not pible to determine the environmental variable exerhe greatest influence on squid distribution becearly all two-way analyses showed significant asstions, suggesting the existence of a strong correletween the environmental factors used in our anal

414 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Stronger and more consistent environmental associ-ations were detected during March–April cruises whichcoincide with the presence of intense thermal gradi-ents in the study area. These thermal gradients corre-sponded partially to the Brazil–Malvinas Confluencefront and partially to the shelf break front that indicatesthe limit between the shelf waters (Patagonian Currentand Rıo de La Plata discharge) and those of the Malv-inas/Falkland Current (Bakun and Parrish, 1991).
The association between shortfin squid with thepresence of oceanographic fronts in the study area(Brazil–Malvinas confluence front and shelf breakfront), could be related to higher food availability.According toOlson et al. (1994), frontal regions areareas of high nutrient concentration in which phy-toplankton, zooplankton, and nekton abundances canchange significantly due to greater productivity andprey availability. Feeding and spawning concentrationsof commercially important oceanic species are com-monly seen at frontal boundaries.
A better understanding of the influence of physi-cal oceanographic processes onI. argentinus’ distri-bution would be considered very useful for the man-agement of its fishery in the Southwestern AtlanticOcean (SWAO). Considering the annual nature ofI.argentinus life cycle, the high interannual abundancevariability and the weak stock–recruit relationships, itis very difficult to predict the recruitment strength ofthis species (Csirke, 1987; Basson et al., 1996; Bakunand Csirke, 1998). For an adequate management of thist t ofe ncep a,b;R en-c tionw s asi iona orthP
ncew ex-p ra iron-m oa a-tw eda n the
Malvinas/Falkland Islands and the Argentine ExclusiveZone suffered an early closure in 2004 and 2005.
Acknowledgements
We express our appreciation to the authorities of theDireccion Nacional de Recursos Acuaticos (DINARA;Montevideo, Uruguay) for providing the data used inthis study. Also, we thank the CIPO/IAI project “Con-sorcio para la Investigacion del Cambio Global delPacıfico Oriental” (EPCOR-IAI, Eastern Pacific Con-sortium of the InterAmerican Institute for Research onGlobal change) for partially financing this study.
References
Alverson, D.L., Pereyra, W.T., 1969. Demersal fish explorations inthe northeastern Pacific Ocean – an evaluation of exploratoryfishing methods and analytical approaches to stock size and yieldforecasts. J. Fish. Res. Board Can. 26, 1985–2001.
Anderson, C.I.H., Rodhouse, P.G., 2001. Life cycles, oceanographyand variability: ommastrephid squid in variable oceanographicenvironments. Fish. Res. 54, 133–143.
Arkhipkin, A.I., 1990. Edad y crecimiento del calamar (Illex argenti-nus). Frente Marıtimo 6 (A), 25–35 (in Spanish).
Arkhipkin, A.I., 1993. Age, growth, stock structure and migratoryrate of pre-spawning short-finned squidIllex argentinus based onstatolith ageing investigations. Fish. Res. 16, 313–338.
Arkhipkin, A.I., 2000. Intrapopulation structure of winter-spawnedArgentine shortfin squid,Illex argentinus (Cephalopoda:Ommastrephidae), during its feeding period over the Patagonian
B astal
48,
B tments.),e6.
B Pur-ent ofst
Be-na
BmSpan-
ype of resource, it is very useful to know the effecnvironmental variables on distribution and abundaatterns (Caddy, 1983; Waluda et al., 1999, 2001odhouse, 2001). For example, Japanese fishery agies use oceanographic information in combinaith pre-recruit and recruit abundance estimate
mportant indicators when predicting the distributnd abundance of several squid species in the Nacific Ocean (Caddy, 1983).The aforementioned takes on greater releva
hen considering the high probability of an overloitation inI. argentinus fishery that could be furtheggravated by the presence of unfavorable envental conditions (Haimovici et al., 1998; Bazzinnd Quinones, 1999). Actually, there are some indic
ors that suggest a decline in theI. argentinus’ fisheryithin the SWAO: (1) the total catches in 2004 showstrong deployment, and (2) the fishing seasons i
Shelf. Fish. Bull. 98, 1–13.akun, A., Parrish, R.H., 1991. Comparative studies of co
pelagic fish reproductive habitats: the anchovy (Engraulisanchoita) of the Southwestern Atlantic. ICES J. Mar. Sci.343–361.
akun, A., Csirke, J., 1998. Environmental processes and recruivariability. In: Rodhouse, P.G., Dawe, E.G., O’Dor, R.K. (EdSquid Recruitment Dynamics. The GenusIllex as a Model. ThCommercialIllex Species. Influences on Variability, vol. 37FAO Fish. Tech. Pap., pp. 105–124.
asson, M., Beddington, J.R., Crombie, J.A., Holden, S.J.,chase, L.V., Tingley, G.A., 1996. Assessment and managemannual squid stocks: theIllex argentinus fishery in the SouthweAtlantic as an example. Fish. Res. 28, 3–29.
azzino, G., Quinones, R.A., 1999. Caracterısticas biologicas yecologicas del calamarIllex argentinus relevantes para el ordnamiento de su pesquerıa en el Atlantico Sudoccidental. Gaya63 (2), 87–100 (in Spanish).
azzino, G., Quinones, R.A., 2001. Alimentacion del calamarIllexargentinus (Cephalopoda: Ommastrephidae) en la Zona Counde Pesca Argentino–Uruguaya. Gayana 65 (2), 181–192 (inish).

G. Bazzino et al. / Fisheries Research 76 (2005) 401–416 415
Bertolotti, M.I., Brunetti, N.E., Carreto, J.I., Prenski, L.B., Sanchez,R.P., 1996. Influence of shelf-break fronts on shellfish and fishstocks off Argentina. ICES C.M., S/41.
Boltovskoy, E., 1970. Masas de agua (caracterısticas, distribucion,movimientos) en la superficie del Atlantico sudoeste, segun indi-cadores biologicos-foraminıferos. Publicacion del Servicio deHidrografıa Naval H643, Buenos Aires, Argentina, 99 pp. (inSpanish).
Brodziak, J., Hendrickson, L., 1999. An analysis of environmentaleffects on survey catches of squidsLoligo pealei andIllex ille-cebrosus in the northwest Atlantic. Fish. Bull. 97 (1), 9–24.
Brunetti, N.E., 1981. Distribucion de tallas y biologıa reproductivadel calamarIllex argentinus en el Mar Argentino. In: Angelescu,V. (Ed.), Campanas del B/I “Shinkai Maru” 1978–1979, vol.383. Contr. Inst. Invest. Desarrollo Pesq. INIDEP, Mar del Plata,Argentina, pp. 105–119 (in Spanish).
Brunetti, N.E., 1988. Contribucion al conocimiento biologico-pesquero del calamar argentino (Cephalopoda: Ommastrephidae,Illex argentinus). Tesis de Doctorado. Universidad Nacional deLa Plata (in Spanish).
Brunetti, N.E., Ivanovic, M.L., Pineda, S., Elena, B., Rossi, G.R.,Aubone, A., 1997. Calamar (Illex argentinus). Caracterizacionbiologica y evaluacion del estado de explotacion. InformeTecnico del INIDEP (Mar del Plata, Argentina) 98/96 (in Span-ish).
Brunetti, N.E., Elena, B., Rossi, G.R., Ivanovic, M.L., Aubone, A.,Guerrero, R., Benavides, H., 1998. Summer distribution, abun-dance and population structure ofIllex argentinus on the Argen-tine shelf in relation to environmental features. S. Afr. J. Mar.Sci. 20, 175–186.
Caddy, J.F., 1983. The cephalopods: factors relevant to their pop-ulation dynamics and to the assessment and management ofstocks. In: Caddy, J.F. (Ed.), Advances in Assessment of WorldCephalopod Resources, vol. 231. FAO Fish. Tech. Pap., pp. 416–452.
C entinooda).
C6,
D ntal
E .,osyae
).F dings
G Inter-y of
.
G s. 36,
Haimovici, M., Brunetti, N.E., Rodhouse, P.G., Csirke, J., Leta, R.H.,1998.Illex argentinus. In: Rodhouse, P.G., Dawe, E.G., O’Dor,R.K. (Eds.), Squid Recruitment Dynamics. The GenusIllex as aModel. The CommercialIllex species. Influences on Variability,vol. 376. FAO Fish. Tech. Pap., pp. 27–58.
Hatanaka, H., 1986. Growth and life span of short-finned squid,Illexargentinus, in the waters off Argentina. Bull. Jpn. Soc. Sci. Fish.52, 11–17.
Hatanaka, H., 1988. Feeding migration of short-finned squidIllexargentinus in the waters off Argentina. Nippon Suisan Gakkaishi54, 1343–1349.
Hatanaka, H., Kawahara, S., Uozumi, Y., Kasahara, S., 1985. Com-parison of five ommastrephid squids fished by Japan:Todarodespacificus, Illex illecebrosus, Illex argentinus, Nototodarus sloaniandNototodarus sloani gouldi. NAFO Sci. Coun. Stud. 9, 59–68.
Hubold, G., 1977. Hydrography and plankton off Southern Braziland Rio de La Plata August–November. Atlantica 4 (1), 1–22.
Hubold, G., 1978. Second report on hydrography and plankton offSouthern Brazil and Rıo de La Plata; Autumn Cruise: April–June.Atl antica 4 (1), 23–42.
Ivanovic, M.L., Brunetti, N.E., 1994. Food and feeding ofIllexargentinus. Antartic Sci. 6 (2), 185–193.
Laptikhovsky, V.V., Remeslo, A.V., 2001. The Falkland Current as agovernor of biological patterns in the shortfin squid,Illex argenti-nus on the high seas (45–47◦S). ICES C.M. K:16, 8.
Legeckis, R., Gordon, A., 1976. Satellite observations of the Braziland Falkland Currents 1975 to 1976. Deep-Sea Res. 29, 375–401.
Leta, H.R., 1989. Pesca exploratoria y experimental del calamarrojo (Ommastrephes bartramii) y del calamar comun (Illexargentinus) en el sector Uruguayo de la Zona Comun de PescaArgentino–Uruguaya (Invierno de 1986). Frente Marıtimo 5 (A),29–38 (in Spanish).
Leta, H.R., 1992. Abundance and distribution ofIllex argentinusrhynchoteuthion larvae (Cephalopoda: Ommastrephidae) in theSouthwestern Atlantic. S. Afr. J. Mar. Sci. 12, 927–941.
N s-
O bigcentsity
O po-nts.
O .W.,nts.
P s ofwest
P entine
P the
R ries in
astellanos, Z.A., 1960. Una nueva especie de calamar argOmmastrephes argentinus sp. nov. (Mollusca, CephalopNeotropica 6 (20), 55–58.
sirke, J., 1987. Los recursos pesqueros patagonicos y las pesquerıasde altura en el Atlantico sudoccidental. FAO Doc. Tec. Pesca 2875 (in Spanish).
awe, E.G., Colbourne, E.B., Drinkwater, K.F., 2000. Environmeeffects on recruitment of short-finned squid (Illex illecebrosus).ICES J. Mar. Sci. 57, 1002–1013.
rhardt, G., Arena, G., Abella, A., Varela, Z., Sanchez, E., Rios, CDe Moratorio, N.B., 1977. Evaluacion preliminar de los recursdemersales de la Zona Comun de Pesca Argentino–Urugua1975–1976, vol. 11. Informe Tecnico del Instituto Nacional dPesca (INAPE), Montevideo, Uruguay, 176 pp. (in Spanish
AO, 1997. Yearbook of Fishery Statistics. Catches and Lan1995, vol. 80. Rome, FAO, 714 pp.
onzalez, A.F., Trathan, P.N., Yau, C., Rodhouse, P.G., 1997.action between oceanography, ecology and fishery biologthe ommastrephid squidMartialia hyadesi in the South AtlanticMar. Ecol. Prog. Ser. 152, 205–215.
ordon, A.L., 1984. Brazil–Malvinas confluence. Deep-Sea Re359–384.
igmatullin, Ch.M., 1989. Las especies de calamar mas abundantedel Atlantico sudoccidental y sinopsis sobre la ecologıa del calamar (Illex argentinus). Frente Marıtimo 5 (A), 71–81.
’Dor, R.K., Coelho, M.L., 1993. Big squid, big currents andfisheries. In: Okutani, T., O’Dor, R., Kubodera, T. (Eds.), ReAdvances in Cephalopod Fisheries Biology. Tokai UniverPress, pp. 385–396.
lson, D.B., Podesta, G.P., Evans, R.H., Brown, O.B., 1988. Temral variation in the separation of the Brazil and Malvinas CurreDeep-Sea Res. 35, 1971–1990.
lson, D.B., Mariano, A.J., Ashjian, C.J., Peng, G., Nero, RPodesta, G.P., 1994. Life on the edge: marine life and froOceanography 7, 52–60.
erry, R.I., Smith, S.J., 1994. Identifying habitat associationmarine fishes using survey data: an application to the NorthAtlantic. Can. J. Fish. Aquat. Sci. 51, 589–602.
eterson, R.G., 1992. The boundary currents in the western ArgBasin. Deep-Sea Res. 39, 623–644.
eterson, R.G., Stramma, L., 1991. Upper-level circulation inSouth Atlantic Ocean. Prog. Oceanogr. 26, 1–73.
odhouse, P.G., 2001. Managing and forecasting squid fishevariable environments. Fish. Res. 54, 3–8.

416 G. Bazzino et al. / Fisheries Research 76 (2005) 401–416
Rodhouse, P.G., Symon, C., Hatfield, E.M.C., 1992. Early life cycleof cephalopods in relation to the major oceanographic featuresof the southwest Atlantic Ocean. Mar. Ecol. Prog. Ser. 89, 183–195.
Rodhouse, P.G., Barton, J., Hatfield, E.M.C., Symon, C., 1995.Illexargentinus: life cycle, population structure and fishery. ICES Mar.Sci. Symp. 199, 425–432.
Rodhouse, P.G., Prince, P.A., Trathan, P.N., Hatfield, E.M.C.,Watkins, D.G., Bone, D.G., Murphy, E.J., White, M.G., 1996.Cephalopods and mesoscale oceanography at the Antartic PolarFront: satellite tracked predators locate pelagic trophic interac-tions. Mar. Ecol. Prog. Ser. 136, 37–50.
Santos, R.A., Haimovici, M., 2000. The Argentine short-finned squidIllex argentinus in the food webs of southern Brazil. Sarsia 85,49–60.
Severov, D.N., 1991. Masas de agua en elarea del Atlantico Sudoc-cidental, sus caracterısticas y distribucion. Frente Marıtimo 9,93–102 (in Spanish).
Waluda, C.M., Trathan, P.N., Rodhouse, P.G., 1999. Influence ofoceanographic variability on recruitment in theIllex argentinus(Cephalopoda: Ommastrephidae) fishery in the South Atlantic.Mar. Ecol. Prog. Ser. 183, 159–167.
Waluda, C.M., Rodhouse, P.G., Trathan, P.N., Pierce, G.J., 2001a.Remotely sensed mesoscale oceanography and the distributionof Illex argentinus in the South Atlantic. Fish. Oceanogr. 10 (2),207–216.
Waluda, C.M., Rodhouse, P.G., Podesta, G.P., Trathan, P.N., Pierce,G.J., 2001b. Surface oceanography of the inferred hatchinggrounds ofIllex argentinus (Cephalopoda: Ommastrephidae) andinfluences on recruitment variability. Mar. Biol. 139, 671–679.




















