
Simulation of Pedestrian Behaviour using a Discrete Choice Model Calibrated on Actual Motion Data
Upload
independentCategory
view
1download
0
Carbonates calibrated against oceanographic parameters along alatitudinal transect in the Gulf of California, Mexico
J . HALFAR*, L. GODINEZ-ORTA� , M. MUTTI� , J . E. VALDEZ-HOLGUIN§ andJ. M. BORGES�*Institut fur Geologie und Palaontologie, Universitat Stuttgart, Herdweg 51, 70174 Stuttgart, Germany(E-mail: [email protected])�Centro Interdisciplinario de Ciencias Marinas, Ave. IPN s/n, Apdo. Postal 592, La Paz, B.C.S., 23096,Mexico�Institut fur Geowissenschaften, Universitat Potsdam, Postfach 60 15 53, D-14415 Potsdam, Germany§DICTUS, Rosales y Ninos Heroes s/n, Col. Centro, Hermosillo, Sonora 83000, Mexico
ABSTRACT
Trophic resources are an important control governing carbonate production.
Though this importance has long been recognized, no calibration exists to
quantitatively compare biogenic assemblages within trophic resource fields.
This study presents a field calibration of carbonate producers in a range of
settings against high-resolution in situ measurements of nutrients, temperature
and salinity. With its latitudinal extent from 30� to 23� N, the Gulf of
California, Mexico, spans the warm-temperate realm and encompasses
nutrient regimes from oligo-mesotrophic in the south to eutrophic in the
north. Accordingly, from south to north carbonates are characterized by: (i)
coral-dominated shallow carbonate factories (5–20 m water depth) with
average sea-surface temperatures of 25 �C (min. 18 �C, max. 31 �C), average
salinities of 35Æ06& and average chlorophyll a levels, which are a proxy for
nutrients, of 0Æ25 mg Chl a m)3 (max. 0Æ48, min. 0Æ1). (ii) Red algal-dominated
subtidal to inner-shelf carbonate formation (10–25 m) in the central Gulf of
California exhibiting average temperatures of 23 �C (min. 18 �C, max. 30 �C),
average salinities of 35Æ25&, and average Chl a levels of 0Æ71 Chl a m)3 (max.
5Æ62, min. 0). (iii) Molluskan bryozoan-rich inner to outer shelf factories in the
northern Gulf of California (20–50 m) with average sea surface temperatures of
only 20 �C (min. 13 �C, max 29 �C), average salinities of 35Æ01&, and average
contents of 2Æ2 mg Chl a m)3 (max. 8Æ38, min. 0). By calibrating sedi-
mentological data with in situ measured oceanographic information in
different environments, the response of carbonate producers to environ-
mental parameters was established and extrapolated to carbonates on a global
scale. The results demonstrate the importance of recognizing and quantifying
trophic resources as a dominant control determining the biogenic composition
and facies character of both modern and fossil carbonates.
Keywords Gulf of California, heterozoan, nutrients, photozoan, rhodoliths,trophic resources.
INTRODUCTION
Tropical and non-tropical carbonates differ intheir platform geometry, character of sediments,facies dynamics, stratal architecture, sedimentaccumulation rates, and type of carbonate-produ-
cing biota (Nelson, 1988; James, 1997). Climateand ocean circulation, which determine tempera-ture, salinity and nutrient availability, largelycontrol the composition of the biogenic compo-nents forming the carbonates. The dominant rolesof temperature and salinity influencing the
Sedimentology (2006) 53, 297–320 doi: 10.1111/j.1365-3091.2005.00766.x
� 2006 International Association of Sedimentologists 297

biogenic composition of carbonates have beenlong recognized and have resulted in a number ofclassifications (Lees & Buller, 1972; Lees, 1975;Nelson, 1988; James, 1997). Currently, differentterminologies are used in the literature for de-limiting temperature regimes of carbonate-produ-cing settings (Mutti & Hallock, 2003). In additionto temperature and salinity, nutrient availabilityis recognized as an important control on biogeniccomposition in modern and ancient settings (e.g.Littler & Littler, 1985; Hallock & Schlager, 1986;Birkeland, 1987; Carannante et al., 1988; Hallock,1988, 2001; Mutti & Hallock, 2003; Vecsei, 2003;Halfar et al., 2004a; Wilson & Vecsei, 2005). Evenat temperatures typical for formation of tropicalcarbonates, a calcareous flora and fauna can occurthat is more characteristic of transitional or cool-water settings when nutrient levels are high (e.g.lack of coral reefs) (Hallock, 1988; Wilson &Vecsei, 2005). Along a gradient of decreasingnutrient input, whether on a regional or a localscale, the predominant occupants of primarysubstrata in shallow water change from hetero-trophic suspension feeders to benthic plants tophototrophic animal–plant symbionts (Birkeland,1987). For example, corals primarily build reefsin oligotrophic settings, whereas coralline redand calcareous green algae are more abundantin mesotrophic environments. Heterotrophiccarbonate producers (e.g. bryozoa, barnacles,mollusks) in turn are most common in eutro-phic settings (Littler & Littler, 1985; Hallock &Schlager, 1986; Birkeland, 1987; Hallock, 1988,2001). Input of dissolved organic, inorganic, orparticulate organic nutrients (e.g. nitrate, nitrite,phosphate and silicate) must be in bioavailableform. Nutrients stimulate growth of phytoplank-ton, which reduces water transparency, limitingdepth ranges of zooxanthellate corals and calcar-eous algae, and thereby reducing phototrophiccarbonate production (Hallock & Schlager, 1986).Hence, if nutrient and food resources are plenti-ful, small, fast-growing species like filamentousalgae, barnacles, and bryozoans are superiorcompetitors for space to corals (Birkeland, 1987).
The influence of nutrients has also beencontemplated in the formation of a number offossil carbonate systems (Gischler et al., 1994;Carannante et al., 1995; Carannante & Simone,1996; Dupraz & Strasser, 2002; Samankassou,2002; Corda & Brandano, 2003; Mutti & Bernoulli,2003; Pomar et al., 2004). However, the difficultyof separating the relative roles of temperature andnutrient availability on carbonate-producing biotais a key issue in the interpretation of ancient
deposits (see Discussion in Mutti & Hallock,2003). Temperature and nutrients are oftenintimately linked variables, as in many carbonatesettings nutrients are introduced by upwelling ofcold subsurface waters. By compiling data frommodern carbonates worldwide, Halfar et al.(2004a) were able to establish a first set of criteriato distinguish carbonate systems formed underhigh nutrient levels from those formed under lownutrient levels based on the biogenic assemblagespresent.
Recognizing the effect of nutrient availability inassociation with other oceanographic parameters,such as temperature and salinity, as boundaryconditions on different types of carbonates re-quires a quantitative approach in a range ofsettings. To gain a better understanding of car-bonate formation and their oceanographic con-trols and to improve interpretation of fossilcarbonates, a comprehensive calibration of arange of modern carbonate environments tooceanographic parameters in different parts ofthe world is essential. The first calibration ispresented here from the Gulf of California, Mex-ico, which is located at the transition betweentropical and temperate latitudes and thereforeencompasses a suite of carbonate systems develop-ing under contrasting temperature and nutrientconditions along a latitudinal gradient.
REGIONAL SETTING
The Gulf of California is a mid-latitude marginalocean basin that is delimited in the west by themountainous and arid Baja California Peninsula(Fig. 1). Arid conditions are responsible for theabsence of continuously flowing river systemsentering the western Gulf of California, whereonly sporadic input of clastic material occursonto the shelf during winter storm or late summerhurricane events. Colorado River discharge ofsediment and nutrients into the gulf has becomenegligible in recent years because of upstreamdamming (Kowalewski et al., 2000). However,even before dam construction, Colorado-derivedsediments were distributed only in the northern-most Gulf of California (Carriquiry et al., 2001).Being an active rift basin, the Gulf of California ischaracterized by steep margins and narrow shelfareas, resulting in restricted accommodationspace for shallow-water carbonate formation.
In oceanographic terms, the warm-temperateGulf of California can be defined as an evapora-tive ocean basin with net outflow at the surface
298 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

and net inflow at depth (Bray, 1988). Surfacewater masses are influenced by interplay of thecold California Current and the warm NorthEquatorial Current entering the mouth of the gulf.Surface circulation patterns are complex but canbe generally characterized by anticyclonic circu-lation during winter and cyclonic circulationduring summer, with incoming and outgoingflows intensified along the coast (Beier, 1997).Shallow nutrient-rich subsurface water massesare able to sustain high rates of primary produc-tivity through seasonal upwelling (Alvarez-Borr-ego & Lara-Lara, 1991). Upwelling and seasonallyreversing near-surface currents in turn result fromthe monsoonal climate of the Gulf of Californiaregion (Pride et al., 1999). During late springand summer this climate pattern is responsiblefor north–south oriented wind systems, which
initiate the temporary upwelling events along thewestern gulf. Upwelling in turn is closely tied toshort-term surface water temperature reductions.In addition to upwelling, year-round tidal mixinghas been observed to be an important processdelivering nutrients to the macrotidal northernGulf of California shelf regions (Alvarez-Borrego,1983).
METHODS
Surface sediment grab samples were collectedalong transects in four selected shallow-watercarbonate environments distributed along a lati-tudinal gradient spanning almost the entire west-ern shelf of the Gulf of California (ca 800 km) inwater depths ranging from shallow subtidal to120 m (Fig. 1). Groups of carbonate-producingorganisms were distinguished during pointcounting of 40 sediment samples from each site(400 points/sample, 63 lm to 2 mm fraction;average of 150 points per sample, > 2 mm frac-tion). In order to distinguish biofacies, biotamaking up more than 5% of carbonate producersat an individual site were subjected to a Q-modecluster analysis, which was conducted usingSPSS 9.0.1 statistical software package by apply-ing Ward’s method with Euclidean distances as asimilarity index. In addition, carbonate content ofall samples collected (entire size fraction) wasdetermined using a coulometer following proce-dures outlined in John et al. (2003).
At each of the four sampling sites, an oceano-graphic mooring with a near-surface buoy waspositioned at 25 m depth containing StowawayTidbit temperature loggers at 25 m and 10 m anda self-contained underwater fluorometer (SCUFA,Turner Designs, Sunnyvale, CA, USA) at 10 m forrecording Chl a, a proxy for nutrients. In addition,a salinometer (37SM MicroCAT, Sea-Bird Elec-tronics, Bellevue, WA, USA) was installed at thesouthernmost site at 10 m. All equipment loggeddata at hourly intervals, and with the exceptionof the salinometer, was operating from March2002 to March 2003. The salinometer wasmoored between September 2002 and September2003. Furthermore, salinity–depth profiles andSecchi disc depths, indicative of light penetra-tion, were recorded at 3 month intervals ateach site between March 2002 and March 2003.Water samples at 10 m depth were taken at3 month intervals using Niskin bottles, frozen inliquid nitrogen immediately upon retrieval, andanalysed for nutrients and Chl a. Chl a content of
113°W
28°N
26°N
24°N
22°N
60
Gulf ofCalifornia
Baj
a C
alifo
rnia
>
N
Bahía de Los Angeles
Cabo Pulmo
Isla San Jose
Punta Chivato
Site I
Site II
Site III
Site IV
200 km
Eutroph
Mesotroph
Oligotroph
18 °C
16 °C
14 °C
200 m
0
Fig. 1. Location map of Gulf of California indicatingfour study sites. In addition, lowest monthly winterseawater temperature isotherms at 10 m, averagenutrient levels, and 200 m bathymetry contour areindicated.
Trophic resources and carbonate production 299
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

water samples was used for calibration of SCUFA-derived fluorescence values. As a result of inter-mittent malfunctioning of the SCUFA fluorome-ters, the in situ Chl a data were supplementedwith remote-sensed SeaWiFS information, whichwas obtained for each site at weekly resolutionfrom http://seawifs.gsfc.nasa.gov. The validity ofthis approach was confirmed by the close corre-lation of averaged in situ and SeaWiFS data at allsites (R2 ¼ 0Æ94). Analyses of inorganic nutrientsin water samples were conducted using a nutrientautoanalyzer (Skalar, Breda, the Netherlands) atthe Universidad Autonoma de Baja California,Mexico, with four channels for simultaneousmeasurements of nitrate + nitrite (NO2�
3 + NO2),phosphate (PO4) and silicate (Si). The analyticalprecision achieved was 0Æ01 lmol l)1 forphosphate, and 0Æ05 lmol l)1 for the othernutrients.
RESULTS
Oceanography
The western margin of the Gulf of California ischaracterized by a shelf width of a few kilometresor less. Hence, only a limited number of areas aresuitable for the formation of shallow-water car-bonate factories, which are of small lateral extent(up to 80 km2). Oceanographic variables weremeasured at each of the four selected settings(Fig. 1).
SalinityBased on quarterly measurements to 30 m depth,salinity at each of the four carbonate producingsettings fluctuated little throughout the year andamong sites and, was centred ca 35Æ00–35Æ30&. Inaddition, hourly high-resolution measurements ofsalinity at the southernmost site at 10 m depthindicated few short-term fluctuations and averagevalues of 35Æ00&. Small negative salinity depar-tures could be correlated to precipitation eventsand were of short duration ranging from hours toa few days.
TemperatureHigh-resolution temperature data were recordedat depths of 10 and 25 m at each of the fourmooring localities (Fig. 2), which encompassedthe average depth of carbonate formation in all ofthe settings. In general, near-surface temperaturesat mooring sites increased from north to south,with minimum and average monthly values
increasing by 5 �C from 14 to 19 �C and 20–25 �C, respectively, whereas maximum valuesincreased only by 1 �C (from 30 to 31 �C). Theamplitude of intra-annual temperature variationsin the south was 12 �C and increased towards thenorth to 16 �C. Maximum temperatures occurred
12
14
16
18
20
22
24
26
28
30
Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
02 03
Tem
pera
ture
(°C
)Site II
}Site III
Site IV
Site I
12
14
16
18
20
22
24
26
28
30
12
14
16
18
20
22
24
26
28
30Te
mpe
ratu
re (
°C)
12
14
16
18
20
22
24
26
28
30
Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
02 03
Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
02 03
Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
02 03
10 m
25 m
Tem
pera
ture
(°C
)Te
mpe
ratu
re (
°C)
5m
Fig. 2. Daily temperature variations at all sites recor-ded at 10 and 25 m depth. Data show development andbreakdown of thermoclines at three southern sites, andwell mixed-water column throughout the year at SiteIV. Missing data for June at Site I were supplementedwith temperature data from 5 m depth.
300 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

in September, whereas minimum temperatureswere recorded in April (two northern sites) andduring upwelling events in June (two southernsites). The temporal variability of the temperaturerecorded showed mainly fortnightly, diurnal andsemidiurnal scales, which were more pronouncedduring spring and summer. In addition, theinfluence of short-duration upwelling eventswhich lasted from a few days to 2 weeks andsignificantly lowered temperatures in spring andsummer months, was visible from the data series(Fig. 2). Development of a shallow thermocline inspring and summer was most pronounced in thesouth where temperature differences of up to10 �C between 10 and 25 m were recorded.Thermocline breakdown occurs in the beginningof the fall season starting in the southern gulf andpropagating to the north. A thermocline did notdevelop at Site IV because of strong windsprevailing year-round (Fig. 2). At Site IV, temper-atures measured at quarterly intervals alongdepth profiles to 50 m were similar to tempera-tures at 10 and 25 m, confirming the well-mixedcharacter of the upper water column in thenorthern Gulf of California.
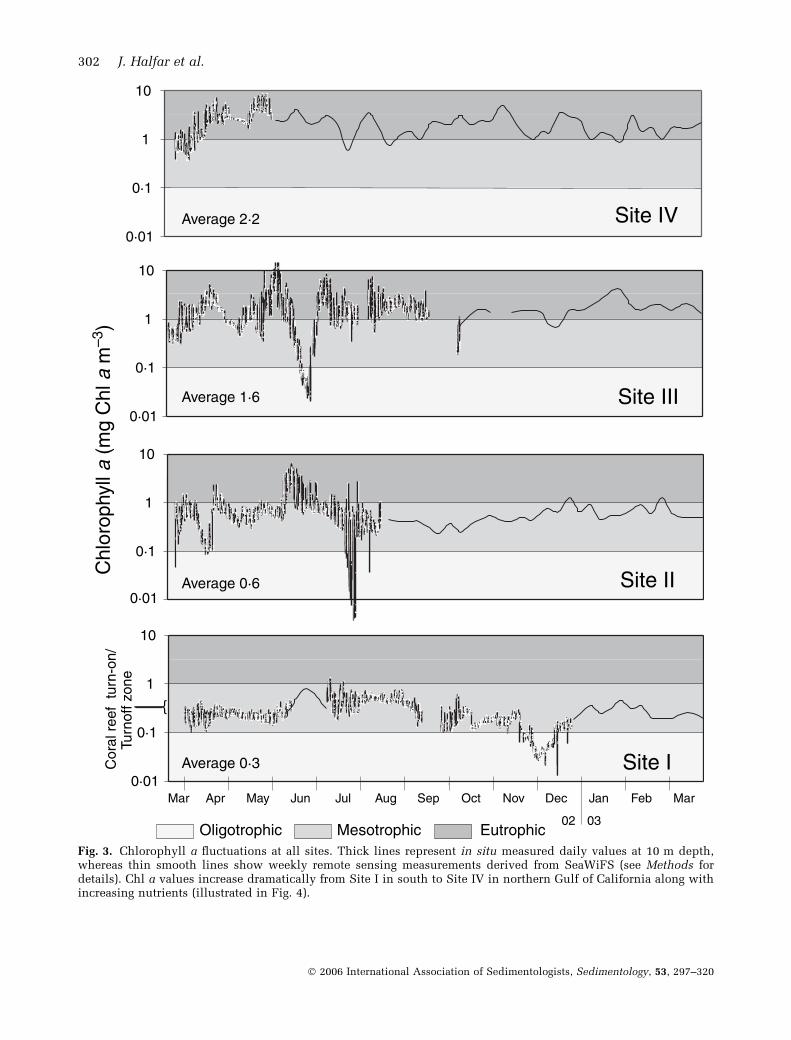
Chl a and nutrientsAt the southern Site I, results of hourly in situmeasurements of Chl a concentrations showedChl a values fluctuating between lower meso-trophic and oligotrophic conditions, with highestvalues of 1Æ1 mg Chl a m)3 being reached duringsummer months when upwelling delivered nutri-ents and cooled surface waters (Figs 1 and 3).Trophic conditions are defined on the basis of Chla values according to the subdivisions of Hallock(2001). Conditions for coral-reef development atSite I are marginal since the average Chl aconcentration of 0Æ3 mg Chl a m)3 is within theso-called coral reef turn-on/turn-off zone (Hal-lock, 2001). Chl a concentrations above that levelwill generally not allow for the development ofcoral-reef communities, whereas at lower values,coral reefs can flourish (Hallock, 2001). To thenorth, Chl a levels increased in the Gulf ofCalifornia, reaching mesotrophic to temporarilyeutrophic conditions at the central Sites II and III(average 0Æ6 and 1Æ6 mg Chl a m)3), with extremevalues of 7 and 10Æ5 mg Chl a m)3 respectively(Fig. 3). Almost entirely eutrophic conditionsprevailed at the northernmost Site IV, withaverage values of 2Æ2 mg Chl a m)3 and short-term extremes of near 9 mg Chl a m)3. Eventhough frequency and amplitude of upwelling-related Chl a spikes were higher in the north than
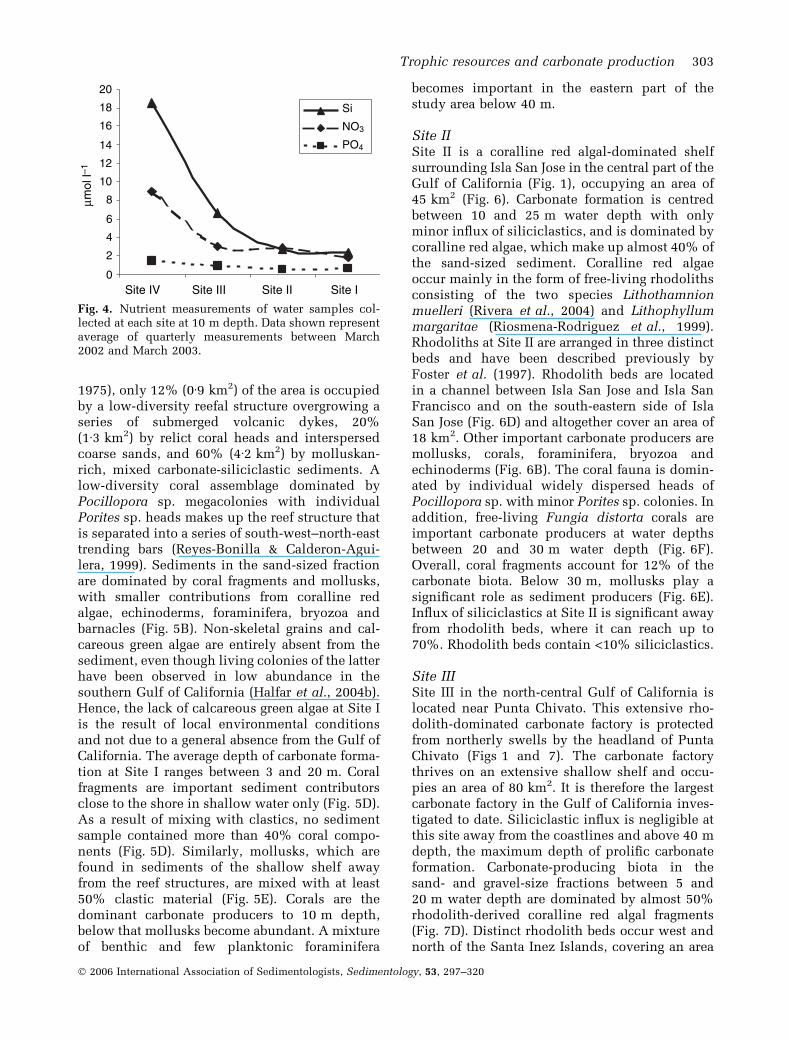
in the south, duration of events was similar at allsites. Inorganic nutrients, determined by analysisof water samples at quarterly intervals at all sites,showed a strongly increasing south-to-north trendsimilar to that observed in Chl a values (Fig. 4).Annually averaged silicate values at all sites were> 2 lmol l)1 reaching up to 18 lmol l)1 at thenorthernmost site. Nitrate values measured atSites II to IV (3–9 lmol l)1) resembled valuestypical for temperate seas, whereas the southern-most Site I had average nitrate values of1Æ8 lmol l)1, which is indicative of tropical con-ditions. Similarly, phosphate concentrations de-creased from north to south from 1Æ5 to0Æ6 lmol l)1. It should be kept in mind thatinorganic nutrients reported here are annuallyaveraged values, which are expected to fluctuatesignificantly throughout the year in accordancewith the observed Chl a cyclicity.
Light penetrationQuarterly Secchi disc measurements for a periodof 1 year revealed shallowest Secchi disc depthsof 8 m at the northernmost site, where lightpenetration is restricted by high phytoplanktonconcentrations in the surface waters. Sites II andIII exhibited Secchi disc depths of 12 and 11 m,respectively, whereas the oligotrophic–meso-trophic Site I had Secchi disc depths of up to17 m (average 13 m). Furthermore, light reachingthe carbonate-producing organisms was restrictedby seasonal macroalgal overgrowth, which wasobserved at the three northern sites only, but was,apart from isolated macroalgal patches, largelyabsent from the southernmost site.
Carbonate environments
Site IThe southernmost Site I is located at 23Æ4� Nlatitude at the bay of Cabo Pulmo in a warm-temperate oligo-mesotrophic environment (Figs1–3). Site I occupies an area of 6Æ5 km2 andexhibits a 2–3 km wide shelf that drops offsteeply to the east (Fig. 5A). The setting isdominated by mixed carbonate-siliciclastics, withcarbonate-rich areas being confined to nearshorelocalities in the northern and southern part of thebay. Carbonates are diluted by coarse granite-derived siliciclastics that are introduced into thebay via a perennial river system. This system isactive only during infrequent late summer hurri-cane events and sporadic winter rains. Eventhough Site I is known as the northernmost coralreef of the eastern Pacific (Brusca & Thomson,
Trophic resources and carbonate production 301
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
0·1
1
10
0·01
0·1
1
10
Site IV
0·01
0·1
1
10
Site III
Site II
0·01
0·1
1
10
Site I
Mar Apr May Jun Jul Aug Sep Oct Nov Dec Jan Feb Mar
02 03Oligotrophic Mesotrophic Eutrophic
Cor
al r
eef
turn
-on/
Turn
off z
one
{
Average 0·3
Average 2·2
Average 1·6
Average 0·60·01
Chl
orop
hyll
a (m
g C
hl a
m–3
)
Fig. 3. Chlorophyll a fluctuations at all sites. Thick lines represent in situ measured daily values at 10 m depth,whereas thin smooth lines show weekly remote sensing measurements derived from SeaWiFS (see Methods fordetails). Chl a values increase dramatically from Site I in south to Site IV in northern Gulf of California along withincreasing nutrients (illustrated in Fig. 4).
302 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
1975), only 12% (0Æ9 km2) of the area is occupiedby a low-diversity reefal structure overgrowing aseries of submerged volcanic dykes, 20%(1Æ3 km2) by relict coral heads and interspersedcoarse sands, and 60% (4Æ2 km2) by molluskan-rich, mixed carbonate-siliciclastic sediments. Alow-diversity coral assemblage dominated byPocillopora sp. megacolonies with individualPorites sp. heads makes up the reef structure thatis separated into a series of south-west–north-easttrending bars (Reyes-Bonilla & Calderon-Agui-lera, 1999). Sediments in the sand-sized fractionare dominated by coral fragments and mollusks,with smaller contributions from coralline redalgae, echinoderms, foraminifera, bryozoa andbarnacles (Fig. 5B). Non-skeletal grains and cal-careous green algae are entirely absent from thesediment, even though living colonies of the latterhave been observed in low abundance in thesouthern Gulf of California (Halfar et al., 2004b).Hence, the lack of calcareous green algae at Site Iis the result of local environmental conditionsand not due to a general absence from the Gulf ofCalifornia. The average depth of carbonate forma-tion at Site I ranges between 3 and 20 m. Coralfragments are important sediment contributorsclose to the shore in shallow water only (Fig. 5D).As a result of mixing with clastics, no sedimentsample contained more than 40% coral compo-nents (Fig. 5D). Similarly, mollusks, which arefound in sediments of the shallow shelf awayfrom the reef structures, are mixed with at least50% clastic material (Fig. 5E). Corals are thedominant carbonate producers to 10 m depth,below that mollusks become abundant. A mixtureof benthic and few planktonic foraminifera
becomes important in the eastern part of thestudy area below 40 m.
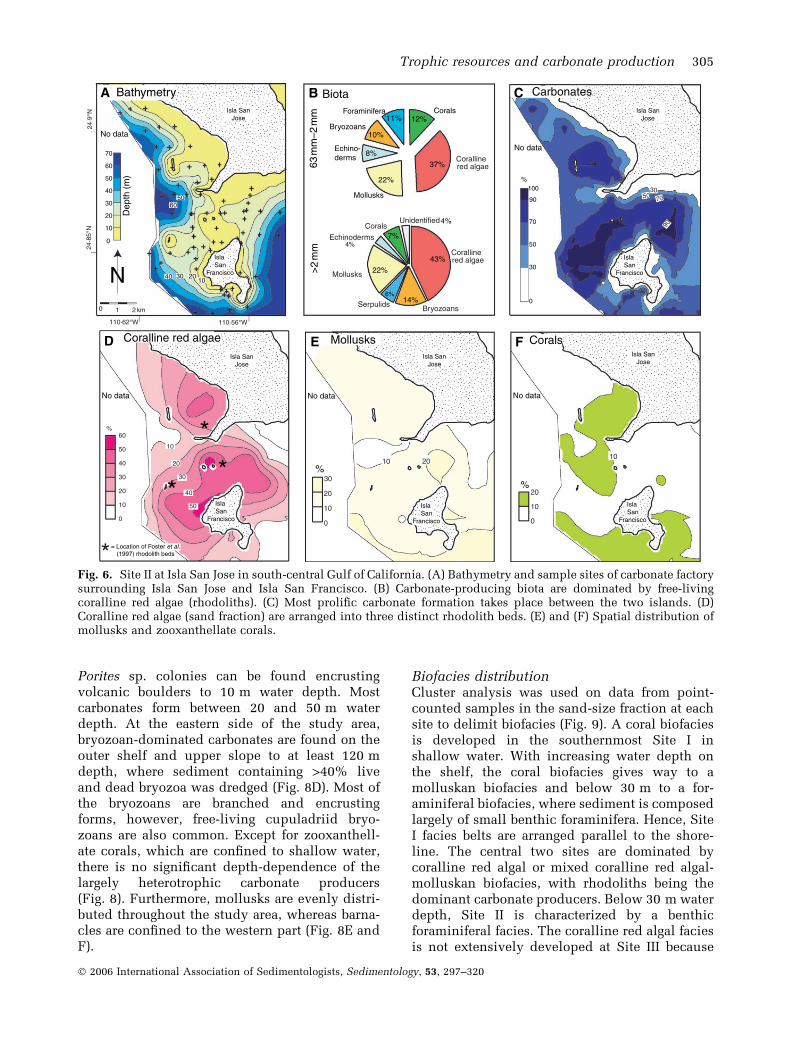
Site IISite II is a coralline red algal-dominated shelfsurrounding Isla San Jose in the central part of theGulf of California (Fig. 1), occupying an area of45 km2 (Fig. 6). Carbonate formation is centredbetween 10 and 25 m water depth with onlyminor influx of siliciclastics, and is dominated bycoralline red algae, which make up almost 40% ofthe sand-sized sediment. Coralline red algaeoccur mainly in the form of free-living rhodolithsconsisting of the two species Lithothamnionmuelleri (Rivera et al., 2004) and Lithophyllummargaritae (Riosmena-Rodriguez et al., 1999).Rhodoliths at Site II are arranged in three distinctbeds and have been described previously byFoster et al. (1997). Rhodolith beds are locatedin a channel between Isla San Jose and Isla SanFrancisco and on the south-eastern side of IslaSan Jose (Fig. 6D) and altogether cover an area of18 km2. Other important carbonate producers aremollusks, corals, foraminifera, bryozoa andechinoderms (Fig. 6B). The coral fauna is domin-ated by individual widely dispersed heads ofPocillopora sp. with minor Porites sp. colonies. Inaddition, free-living Fungia distorta corals areimportant carbonate producers at water depthsbetween 20 and 30 m water depth (Fig. 6F).Overall, coral fragments account for 12% of thecarbonate biota. Below 30 m, mollusks play asignificant role as sediment producers (Fig. 6E).Influx of siliciclastics at Site II is significant awayfrom rhodolith beds, where it can reach up to70%. Rhodolith beds contain <10% siliciclastics.
Site IIISite III in the north-central Gulf of California islocated near Punta Chivato. This extensive rho-dolith-dominated carbonate factory is protectedfrom northerly swells by the headland of PuntaChivato (Figs 1 and 7). The carbonate factorythrives on an extensive shallow shelf and occu-pies an area of 80 km2. It is therefore the largestcarbonate factory in the Gulf of California inves-tigated to date. Siliciclastic influx is negligible atthis site away from the coastlines and above 40 mdepth, the maximum depth of prolific carbonateformation. Carbonate-producing biota in thesand- and gravel-size fractions between 5 and20 m water depth are dominated by almost 50%rhodolith-derived coralline red algal fragments(Fig. 7D). Distinct rhodolith beds occur west andnorth of the Santa Inez Islands, covering an area
0
2
4
6
8
10
12
14
16
18
20
NO3
PO4
Si
µmol
l–1
Site IV Site III Site II Site I
Fig. 4. Nutrient measurements of water samples col-lected at each site at 10 m depth. Data shown representaverage of quarterly measurements between March2002 and March 2003.
Trophic resources and carbonate production 303
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

of > 16 km2. Mollusks, which make up 28% ofthe sand-size fraction and 43% of the > 2 mmfraction, are distributed evenly throughout thesampling area and occur at all water depths(Fig. 7E). Porites sp. corals account for only 5%of the sand fraction, and are therefore signifi-cantly reduced in abundance compared with SiteII (Fig. 7F).
Site IVThe northernmost Site IV is located near Bahıade Los Angeles at 29� N, where the carbonatefactory extends over an area of at least 53 km2
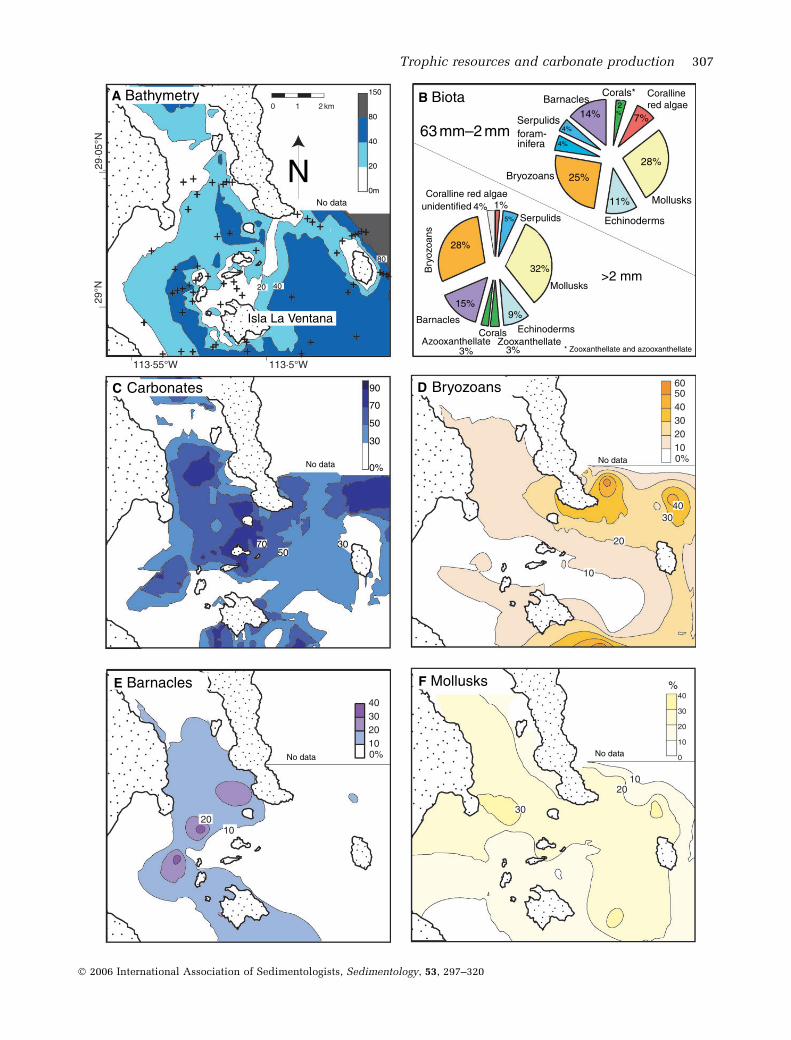
and surrounds a group of islands away from thesiliciclastics dominated coastline (Figs 1 and8A,C). Bryozoans and mollusks dominate car-bonates in the sand- and gravel-size ractions,with echinoderms and barnacles being addi-tional important constituents (Fig. 8B). On aver-age, zooxanthellate coral fragments make up only3% of the rare gravel-size fraction and <1% ofthe more abundant sand-size components.Nevertheless, zooxanthellate coral fragments aresignificant carbonate producers over a limitedarea of 4 km2 surrounding a cluster of smallislands near Isla Ventana. Here monospecific
Corals
33%
Corallinered algae
10%
Mollusks
31%
Echino-derms
6%
Bryozoans4%
Foraminifera5%
Barnacles3%
Others
8%
23·4
°N23
·43°
N0 1 2 km
10
2030
40
5
5060
10
10
50
30
CoralD
B
0
10
20
30
40
50
60
70m
0
30
50
70
10
%
0
10
20
30
40
50
60%
10
20
30
40
50
C
N
A
1020
30
40
0
10
20
30
40
50
%
E F
%
0
5
10
20
5
Mollusks Coralline red algae
CarbonatesBiotaBathymetry
Fig. 5. Southernmost study Site I at bay of Cabo Pulmo. (A) Bathymetry of bay and sample sites are indicated. (B)Major carbonate producers in sand-size fraction are corals and mollusks. (C) Siliciclastics are dominant sedimentcomponents (all size fractions) with carbonates being important in northern and southern near-coastal zones. (D)Coral-derived sand-sized carbonates are abundant in near-coastal shallow waters where low-diversity reefal structureexists. (E) Mollusks become dominant carbonate producers with increasing water depth, whereas (F) coralline algaeare present in low abundance and show patchy distribution throughout study area.
304 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
Porites sp. colonies can be found encrustingvolcanic boulders to 10 m water depth. Mostcarbonates form between 20 and 50 m waterdepth. At the eastern side of the study area,bryozoan-dominated carbonates are found on theouter shelf and upper slope to at least 120 mdepth, where sediment containing >40% liveand dead bryozoa was dredged (Fig. 8D). Most ofthe bryozoans are branched and encrustingforms, however, free-living cupuladriid bryo-zoans are also common. Except for zooxanthell-ate corals, which are confined to shallow water,there is no significant depth-dependence of thelargely heterotrophic carbonate producers(Fig. 8). Furthermore, mollusks are evenly distri-buted throughout the study area, whereas barna-cles are confined to the western part (Fig. 8E andF).
Biofacies distributionCluster analysis was used on data from point-counted samples in the sand-size fraction at eachsite to delimit biofacies (Fig. 9). A coral biofaciesis developed in the southernmost Site I inshallow water. With increasing water depth onthe shelf, the coral biofacies gives way to amolluskan biofacies and below 30 m to a for-aminiferal biofacies, where sediment is composedlargely of small benthic foraminifera. Hence, SiteI facies belts are arranged parallel to the shore-line. The central two sites are dominated bycoralline red algal or mixed coralline red algal-molluskan biofacies, with rhodoliths being thedominant carbonate producers. Below 30 m waterdepth, Site II is characterized by a benthicforaminiferal facies. The coralline red algal faciesis not extensively developed at Site III because
0
10
20
30
40
50
60
70
Dep
th (
m)
No data
110·62°W 110·56°W
24·8
5°N
24·9
°N
1 2 km
Corals12%
37%
Mollusks
22%
Echino-derms 8%
Bryozoans10%
Foraminifera11%
N
Biota
Bryozoans Serpulids
Mollusks
Echinoderms
Corals Unidentified
Corallinered algae
4%
43%
14%
22%
4%
6%
7%
Corallinered algae
>2
mm
63 m
m–2
mm
0
6050
40 203010
Bathymetry
0
30
50
70
100
90
%
3050 70
90
Carbonates
Coralline red algae
10
20
30
40
50
No data
No data
Isla SanJose
Isla San
Francisco
Isla SanJose
Isla San
Francisco
Isla SanJose
Isla San
Francisco
*
**
= Location of Foster et al. (1997) rhodolith beds
No data
Isla SanJose
Isla San
Francisco
Corals
0
10
20%
10
Mollusks
No data
Isla SanJose
Isla San
Francisco0
10
20
%10 20
30
A B C
FED
60%
50
40
30
20
10
0
Fig. 6. Site II at Isla San Jose in south-central Gulf of California. (A) Bathymetry and sample sites of carbonate factorysurrounding Isla San Jose and Isla San Francisco. (B) Carbonate-producing biota are dominated by free-livingcoralline red algae (rhodoliths). (C) Most prolific carbonate formation takes place between the two islands. (D)Coralline red algae (sand fraction) are arranged into three distinct rhodolith beds. (E) and (F) Spatial distribution ofmollusks and zooxanthellate corals.
Trophic resources and carbonate production 305
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

mollusks are abundant. A distinct bryozoan bio-facies consisting of encrusting, branched andmobile bryozoans is developed at Site III, mainlybetween 20 to 40 m water depth. Site IV ischaracterized by extensive bryozoan and mollus-kan biofacies, with bryozoans occupying the
substratum below 40 m water depth and mollusksdominating in shallower zones. In addition, asmall coral biofacies is developed at the nor-thernmost site, even though zooxanthellate coralsare overall insignificant contributors to the car-bonate sediment.
111·96°W 111·9°W
27·0
2°N
27·0
5°N Coral
5%
46%
Mollusks
28%
Echino-derms
11%
Bryozoans
10%N10 20 30 40
50
60
A Bathymetry
C Carbonates
B Biota
Others5%
3%
>2 mm63 mm–2 mm
D Coralline red algae
70
90
1020
304050
Punta Chivato
Islas SantaInez
0 0 1 2 km
10
20
30
40
50
60
70m
CorallineRed algae
BryozoansCorallinered algae
Echino-dermsMollusks
43%
38%
11%
10
20
E Mollusks
0
10
20
%30
10
5
%20
10
5
0
%60
50
40
30
20
10
0
%100
90
70
50
30
0
F Coral
Fig. 7. Site III at Punta Chivato in central Gulf of California. (A) Bathymetry and sample sites near Punta Chivato. (B)Carbonate-producing biota are dominated by coralline red algae and mollusks. (C) Punta Chivato is most extensiveand prolific carbonate factory in Gulf of California. (D) Coralline red algae in sand-size fraction are most abundant tothe west of Islas Santa Inez. (E) and (F) Spatial distribution of mollusks and zooxanthellate corals.
Fig. 8. Northernmost Site IV at Bahıa de Los Angeles. (A) Bathymetry and sample locations surrounding smallislands. (B) Most dominant carbonate producers are heterotrophic organisms. (C) Carbonate factories are locatedbetween islands and on slope at eastern side of study area. (D–F) Distribution of dominant carbonate producers.
306 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
No data
0m
20
40
80
150
No data
0 1 2 km
113·55°W 113·5°W
29°N
29·0
5°N
N
0%
30
50
70
90
Corals* 2%
7%
Mollusks
28%
Echinoderms
11%
Bryozoans 25%
foram-inifera 4%
Serpulids4%
Barnacles14%
* Zooxanthellate and azooxanthellate
20 40
80
A Bathymetry
3050
70
C Carbonates
B Biota
0%102030405060
No data
10
20
3040
unidentified 4%Serpulids5%
Coralline red algae1%
Bry
ozoa
ns
28%
Barnacles
15%
CoralsAzooxanthellate
3%Zooxanthellate
3%
Echinoderms
9%
Mollusks
32%>2 mm
63 mm–2 mm
Coralline red algae
D Bryozoans
E Barnacles
No data 0%10203040
1020
0
10
20
30
40%F Mollusks
No data
1020
30
Isla La Ventana
Trophic resources and carbonate production 307
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

DISCUSSION
Integrating the above oceanographic and sedi-mentologic data allows for a calibration of theGulf of California carbonate environments toselected oceanographic controls. The calibrationin turn is used below as a baseline for comparisonwith temperature and nutrient controls on mod-ern carbonate environments worldwide. Based onthis a model is established for delimiting therespective influence of temperature and nutrientson modern and fossil carbonates.
Environmental parameters
SalinityAlthough salinity is commonly a major factorcontrolling carbonate-producing environments
(Lees, 1975), in situ measurements in the Gulf ofCalifornia study sites show limited fluctuations ofsalinity and exhibit values ca 35&. Salinity in thewestern Gulf of California is therefore comparableto open marine conditions and intra-annualsalinity fluctuations are negligible. Hence, salin-ity is suitable for a wide range of carbonate-producing communities in the Gulf of California.
TemperatureWater temperature is commonly cited as one ofthe most important oceanographic controls gov-erning shallow-water carbonate formation (Lees &Buller, 1972; Carannante et al., 1988; Nelson,1988; James, 1997). Gulf of California sites arecharacterized by intra-annual temperature fluctu-ations of up to 16 �C as well as rapid temperaturereductions during short-term upwelling events.
Coral facies
Molluskan facies
Coralline algal facies
Bryozoan facies
Benthicforaminiferal facies
Coralline algal-molluskan facies
109·435°W 109·405°W
23·4
1°N
23·4
45°N
0 1 2 km111·97°W 111·91°W
27·0
2°N
27·0
6°N
No data
113·55°W 113·45°W
110·64°W 110·56°W
24·8
5°N
24·9
°N
27·0
227
·06
No data
Site I
Site II
Site III
Site IV
N
No data
Fig. 9. Distribution of biofacies in four study sites as determined by cluster analysis of all point-counted samples insand-size fraction. Note that all four sites are shown at same scale.
308 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
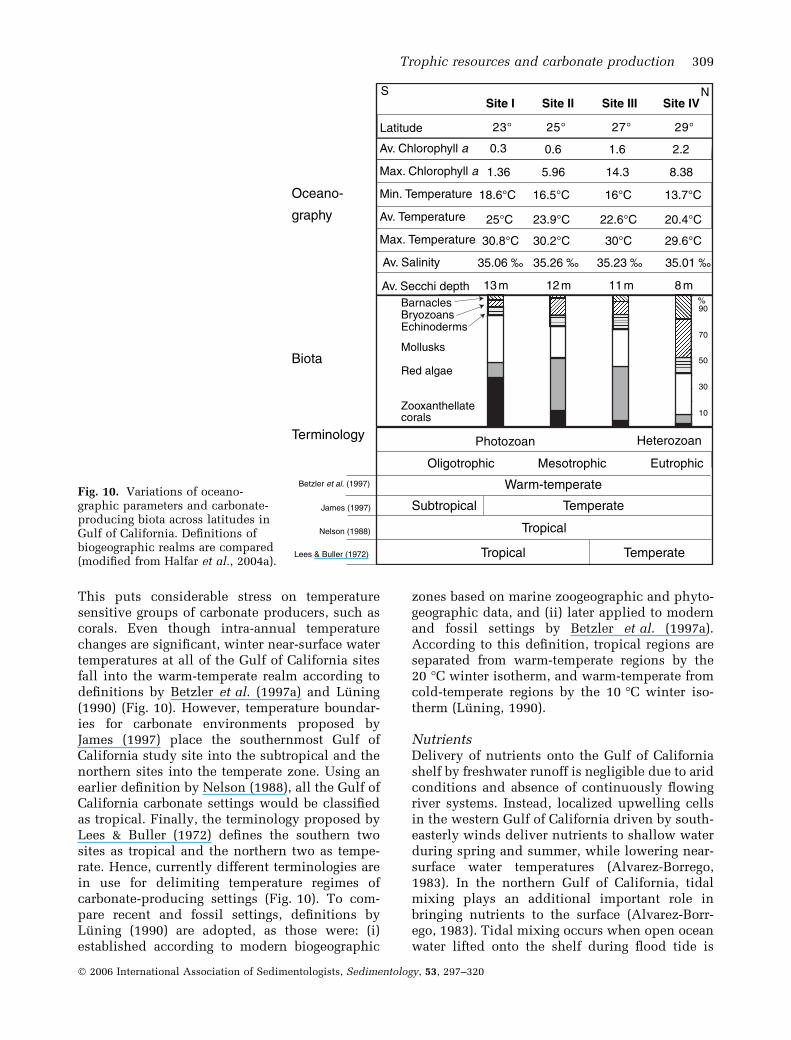
This puts considerable stress on temperaturesensitive groups of carbonate producers, such ascorals. Even though intra-annual temperaturechanges are significant, winter near-surface watertemperatures at all of the Gulf of California sitesfall into the warm-temperate realm according todefinitions by Betzler et al. (1997a) and Luning(1990) (Fig. 10). However, temperature boundar-ies for carbonate environments proposed byJames (1997) place the southernmost Gulf ofCalifornia study site into the subtropical and thenorthern sites into the temperate zone. Using anearlier definition by Nelson (1988), all the Gulf ofCalifornia carbonate settings would be classifiedas tropical. Finally, the terminology proposed byLees & Buller (1972) defines the southern twosites as tropical and the northern two as tempe-rate. Hence, currently different terminologies arein use for delimiting temperature regimes ofcarbonate-producing settings (Fig. 10). To com-pare recent and fossil settings, definitions byLuning (1990) are adopted, as those were: (i)established according to modern biogeographic
zones based on marine zoogeographic and phyto-geographic data, and (ii) later applied to modernand fossil settings by Betzler et al. (1997a).According to this definition, tropical regions areseparated from warm-temperate regions by the20 �C winter isotherm, and warm-temperate fromcold-temperate regions by the 10 �C winter iso-therm (Luning, 1990).
NutrientsDelivery of nutrients onto the Gulf of Californiashelf by freshwater runoff is negligible due to aridconditions and absence of continuously flowingriver systems. Instead, localized upwelling cellsin the western Gulf of California driven by south-easterly winds deliver nutrients to shallow waterduring spring and summer, while lowering near-surface water temperatures (Alvarez-Borrego,1983). In the northern Gulf of California, tidalmixing plays an additional important role inbringing nutrients to the surface (Alvarez-Borr-ego, 1983). Tidal mixing occurs when open oceanwater lifted onto the shelf during flood tide is
0.3 0.6 1.6 2.2Av. Chlorophyll a
30
50
70
90
10
NS
27°25°23° 29°
BarnaclesBryozoansEchinoderms
Mollusks
Red algae
Zooxanthellatecorals
Biota
Min. Temperature 18.6°C 16.5°C 16°C 13.7°C
Terminology Photozoan Heterozoan
Av. Salinity 35.01 ‰35.23 ‰35.26 ‰35.06 ‰
Oligotrophic Mesotrophic Eutrophic
Oceano-
graphy
Av. Secchi depth 13 m 12 m 11 m 8 m
Av. Temperature
Max. Chlorophyll a
Max. Temperature
25°C 23.9°C 22.6°C 20.4°C
30.8°C 30.2°C 30°C 29.6°C
1.36 5.96 14.3 8.38
%
Site IVSite IIISite IISite I
Latitude
Betzler et al. (1997)
James (1997)
Nelson (1988)
Lees & Buller (1972)
Warm-temperate
Tropical Temperate
Subtropical Temperate
Tropical
Fig. 10. Variations of oceano-graphic parameters and carbonate-producing biota across latitudes inGulf of California. Definitions ofbiogeographic realms are compared(modified from Halfar et al., 2004a).
Trophic resources and carbonate production 309
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

mixed with the shelf water, enabling it to transferpart of its nutrient load to the waters on the shelfbefore being removed again during ebb tide(Tomczak & Godfrey, 1994). Particularly favour-able conditions for such mixing are found wherea chain of islands or submarine ridges separatethe shelf from the open sea, such as at Site IV andto some extent at Site III. Unlike upwelling, tidalmixing is a process that takes place year-roundand involves the uppermost water column only,therefore not strongly affecting near-surface watertemperatures. Tidal mixing is most pronouncedin the macrotidal island-bounded northern Gulfof California sites. Hence, extreme nutrient con-ditions during short-duration upwelling eventsare superimposed onto year-round high nutrientlevels in the northern Gulf of California.
The high spatial variability of surface nutrientsled Santamaria-Del-Angel et al. (1994) to dividethe Gulf of California into 14 biogeographicregions based on analysis of remotely sensedcoastal zone colour scanner chlorophyll data.Differences between the four stations studied hereare extreme, with average Chl a as well as silicatevalues being 7 times higher in the north than inthe south (Figs 3 and 4). Similarly, nitrate andphosphate levels differ significantly betweennorth and south. Fixed nitrogen and phosphateions are required by all organisms for synthes-izing proteins and nuclear material for cell main-tenance (Hallock, 2001), whereas phosphorus isknown to be an inhibitor for calcification (Hallock& Schlager, 1986). The nutrient values encoun-tered in this study are typical for temperate zonesinfluenced by upwelling (Crossland, 1987).Furthermore, intra-annual fluctuations of Chl aconcentrations encountered during the high-reso-lution measurements at each site are significantlystronger in the northern than in the southernsites. This is in agreement with Valentine (1973),Birkeland (1987) and Hallock (1987), who indi-cate that nutrient-deficient environments aremore stable, whereas high nutrient levels corre-late with environmental instability, because fac-tors that supply nutrients to sea surface waterstend to be seasonal (e.g. upwelling). In fact,stable high-resource environments are not com-mon in nature, but are usually found only inareas of anthropogenic influence (e.g. sewageoutfalls) (Hallock, 1987). Environmental instabil-ity favours heterotrophic feeders that can adaptmore easily to changing conditions.
The nutrient and temperature gradient alongthe axis of the Gulf of California is paralleled by ashift from phototroph carbonate producers in the
south to a heterotroph community in the north.Corals decrease northwards whereas coralline redalgae increase in the central mesotrophic sitesand heterotrophs such as byrozoans and mollusksdominate at the northern site, where year-roundhigh-nutrient levels are maintained by tidal mix-ing (Fig. 10). Even though coralline algae arefavoured over corals and larger foraminiferaunder higher supplies of nutrients (Brasier,1995), they are not important constituents of thesediment in the northernmost eutrophic site(Fig. 10). Coralline red algae are tolerant of avariety of water temperatures and neither sub-strate type, wave exposure, nor salinity can beinvoked for the paucity of red algae at Site IV.Thus, it is argued that the dominance of corallinered algae decreases under eutrophic conditions asa consequence of reduced depth of light penetra-tion as reflected by Secchi disc depths of only8 m, and overgrowth by filamentous and fleshyalgae.
LightLight penetration, which constrains the extent ofthe euphotic zone, is dependent on trophicresources as well as on suspended fine sedimentin the water column. Double Secchi disc depthcommonly defines the depth of the euphotic zonewhere < 1% of surface irradiance penetrates(Holmes, 1970). Hence the euphotic zone in theGulf of California ranges from 26 m in the southto 16 m in the north (Fig. 10). In the absence of acontinuous supply of fine terrigenous or land-derived organic material from the arid BajaCalifornia peninsula onto the Gulf of Californiashelf, light penetration is expected to fluctuatemainly according to phytoplankton abundanceand, hence, nutrients. Input of fine terrigenousclastics along with a reduction in light penetra-tion play a role only during infrequent seasonalstorms. These are most pronounced in the nor-thern Site IV that is affected by north-east Pacificstorm systems during the winter months. Theeuphotic zone is significantly reduced duringshort-term upwelling events and in regions ofpronounced tidal mixing, limiting available lightfor phototrophic carbonate producers. Light limi-tation is most pronounced at Site IV, wherephototrophs in the form of dwarfed corals areonly present to 10 m depth, and phototrophiccoralline red algae occur in low abundance.Monotypic Porites coral communities are highlyadapted to withstand the low light levels andturbid waters of unstable nutrient-rich environ-ments (Wood, 1993). As a result of the intimate
310 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

link between nutrients and depth of the euphoticzone (Hallock, 1987), it is difficult to distinguishwhich of those two controls is actually limitingphototrophic carbonate producers at the studysites. Furthermore, the observed seasonal macro-algal overgrowth, which increases to the north inthe Gulf of California, can shift an ecosystem fromdominance of corals to benthic algae because oflight limitation (Birkeland, 1987). Light limitationof carbonate production by the influence ofnutrients has been described for the Miocene ofthe Balearic Islands (Pomar et al., 2004), where ashallow chlorozoan factory predominated in theeuphotic zone and a rhodalgal association existedon the oligophotic open shelf.
Influence of siliciclasticsHigh input of fine siliciclastics can result in lightlimitation as well as clogging of suspension-feeding organisms. As a result of the absence ofyear-round river systems in the western Gulf ofCalifornia, terrestrial sediment input into thecarbonate factories is small at the central twosites. There, rhodolith factories, which are lesstolerant to mixing with siliciclastics, are devel-oped. At the southernmost site, abundant sili-ciclastics are mixed with carbonate sediments,whereas prolific carbonate formation takes placein a low-diversity reefal structure, even thoughcorals are known to be most susceptible toterrigenous input. Reasons for siliciclastics atSite I not being detrimental to benthic carbonateproducers are their coarse nature and onlyinfrequent introduction and redistribution inthe system during occasional storms. A similarpattern has been found in the Gulf of Eilat,where deposition of coarse terrigenous clasticsdoes not adversely affect coral reef development(Friedman, 1985). This carbonate/terrigenousclastic relationship may in fact be a characteris-tic of tectonically active areas where a new oceanbasin is forming in a climate suitable for carbon-ate-synthesizing organisms (Friedman, 1985).Furthermore, it has been shown that corals insettings influenced by terrigenous sediments arephysiologically adapted to turbidity associatedstress (Anthony, 2000). At Site IV carbonates aremixed with fine siliciclastics, whereas light-independent organisms adapted to living inmuddy oligophotic settings, such as mollusksand mobile bryozoa, thrive. Even though finesiliciclastics are only infrequently introducedinto the Site IV setting, significant redistributioncan be expected to take place as a result of strongbottom currents (Alvarez et al., 1984), hence
requiring a benthic community adapted to copewith fine clastics.
Water depthSurprisingly, the depth of benthic production ofthe four studied carbonate factories shifts down-wards with decreasing depth of light penetrationcaused by increasing phytoplankton (Chl a) con-tent in the water column (Fig. 10). This down-ward shift is a consequence of increaseddominance of light-independent or low light-tolerant organisms at these sites (e.g. higher lowlight tolerance of coralline red algae in compar-ison with corals). Differences in depth of benthicproduction also depend on: (i) on the shelfprofile, with more shallow-water accommodationspace available at the three southern sites, and (ii)a strong thermocline that develops in the south,with temperatures below 20 m at Site I being upto 10 �C cooler than at 10 m depth during thesummer (Fig. 2). The thermocline therefore re-stricts development of the photozoan assemblageat Site I to depths shallower than 20 m. In thecentral sites, most prolific carbonate formation byrhodoliths occurs in water depths of 20 m. Thiscontrasts with other studies of rhodolith depthdistribution, where most prolific rhodolithgrowth is reported from below 30 m water depth(McMaster & Conover, 1966; Basso, 1998). Hence,interpretations of depth occurrences of rhodalgallithofacies should be made with caution, asrhodoliths can occur at a wide variety of depthsdepending on light availability (Bosence, 1983).At the northernmost site, carbonate formationmainly takes place below 20 m. This reflectsshallow water zones being influenced by a macro-tidal regime and year-round strong winds thatcause significant mixing in the upper watercolumn and thus decrease environmental stabil-ity in the shallow zones.
Carbonate occurrence in the warm-temperate,oligotrophic and mesotrophic Gulf of Californiacarbonate factories decreases significantly withdepth (Sites I to III), whereas the eutrophicfactory (Site IV) exhibits carbonates to the maxi-mum sampled depth (120 m), as a result of theabundance of heterotrophic, light-independentorganisms. This is in agreement with examplesfrom the fossil record, where carbonate formationof some warm-temperate systems has been ob-served to decrease drastically with depth (Brac-hert et al., 1998), whereas in other temperate seasheterozoan-dominated carbonates are found overa broad bathymetric spectrum (James, 1997; Jameset al., 2004).
Trophic resources and carbonate production 311
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

Biogenic assemblages along a latitudinaltransect
Along with a change of oceanographic boundaryconditions, Gulf of California carbonates displaya distinct north–south gradient with respect tothe composition of organisms from heterotroph-dominated to a mixture of phototrophs andheterotrophs (Fig. 10). The transect investigatedshows latitudinal changes both in temperatureand nutrient concentrations. The warm-temperatecarbonates are coral-dominated if oligo-meso-trophic conditions prevail, rhodolith-dominatedif mesotrophic, and bryozoan–molluskan-domin-ated under eutrophic conditions. This is inagreement with Littler & Littler (1985) andHallock (2001) who state that under increasingtrophic resources, a carbonate system changesfrom phototrophs with abundant coral throughalgal and finally to heterotroph-dominatedsystems.
Hence, significant facies and biotic changes areobserved along the 800 km Gulf of Californiatransect, which can be attributed to the fact thatmany organisms in this transitional warm-tem-perate realm are near their growth thresholds, andhence, changes in oceanographic parameters (e.g.nutrients and temperature) bring about rapidlateral biofacies changes (James et al., 1999). Interms of classification, the southernmost sitebelongs to the photozoan association, which asdefined by James (1997) includes zooxanthellatecorals, calcareous green algae and non-skeletalgrains plus or minus contributions from theheterozoan association including coralline redalgae. The northernmost site is a truly heterozoanregime and is made up of non-light-dependentbiota plus or minus coralline red algae (James,1997). Sites II and III, however, contain signifi-cant amounts of both heterozoan and photozoancomponents, and can therefore not be clearlycharacterized as belonging to either association.These settings instead are part of a transitionalphotozoan–heterozoan assemblage, which wascharacterized by Halfar et al. (2004a) as contain-ing 1–20% photozoan components within aheterozoan-dominated association. Transitionalassemblages are not only common in the Recent,but have also been described from a number offossil settings (see discussion below).
The occurrence and abundance of photozoancomponents, such as calcareous green algae andcorals, play an important role in distinguishingbetween heterozoan and photozoan carbonatesystems. Apart from a low-diversity reefal struc-
ture at Site I, zooxanthellate corals are present inthe Gulf of California carbonate environments asnon-framework coral communities (sensu Riegl &Piller, 2000); they decrease in abundance as wellas species diversity to the north. Oligotypiccommunities prevail to the south (11 species),whereas largely monotypic Porites panamensiscommunities occur in the north (Reyes-Bonilla &Lopez-Perez, 1998). Furthermore, corals showsignificantly reduced growth and calcificationsrates from south to north. Similarly, live Halime-da plants were observed in the southernmost sitesonly and were not encountered in the sediment atany site. A similar pattern was found in warm-temperate sites on the south-west Australianshelf, where live Halimeda colonies are encoun-tered underwater, but do not contribute tocarbonate formation in the sediment (James et al.,1999). The authors attributed that to poor calci-fication of the algae.
Depositional profiles and physicalsedimentology of Gulf of California carbonates
The dominant shelf morphology at all sites is thatof a gently sloping, unrimmed shelf, with a steepdrop-off at the shelf edge between 50 and 100 m.Unlike tropical carbonates, the warm-temperateGulf of California carbonates are poorly cemen-ted, facilitating transport of coarse-grained bio-clasts. Poor cementation is characteristic of manynon-tropical carbonates forming under lowertemperatures and in water with a lesser degreeof saturation with respect to CaCO3 than tropicalcarbonate settings (Nelson, 1988). This results insignificant carbonate loss from the shallow car-bonate factory to off-shelf transport. As alreadyestablished in fossil warm-temperate carbonates(Betzler et al., 1997b; Brachert et al., 1998), thestyle of deposition is therefore similar to silicic-lastics. Hence, any transport and redepositionprocesses will function according to conceptsestablished for siliciclastics rather than tropicalcarbonates. A fossil example from the Pliocene ofthe Gulf of California demonstrates this kind ofsediment transport, where uncemented carbonategrains of temperate character were redepositedinto an adjacent basin (Dorsey & Kidwell, 1999).
Currents driven by waves, storms and tidescontrol carbonate-grain dispersal and accumula-tion at all the sites to a much greater extent thanin tropical factories. Where dispersal is limited bybinding and cementation, however, distinct auto-chthonous and parautochthonous biofacies canbe distinguished in the Gulf of California (Fig. 9).
312 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
This observation contrasts with previous sugges-tions that minor biofacies differentiations in theMiocene of southern Spain are characteristic offossil nontropical carbonates because of sedimentdispersal and mixing in the absence of binding(Betzler et al., 1997b). This difference can eitherbe attributed to higher energy conditions havingprevailed in the Miocene southern Spain settingsor to the fact that biofacies variations in the fossilrecord are additionally obscured by differentialdissolution of biogenic particles, which wouldpreferentially remove aragonitic constituents.
Global influence of nutrients and temperatureon modern carbonates
Carbonates containing a mixture of photozoanand heterozoan assemblages (transitional carbon-ates), such as the ones forming in the Gulf ofCalifornia, are common in the fossil record (e.g.Lukasik et al., 2000; Samankassou, 2002; Pomaret al., 2004). The separation of the relative roles oftemperature and nutrients on fossil assemblagesis a key issue in the interpretation of thesesystems, and a number of sedimentological andgeochemical criteria have been proposed to dif-ferentiate these two main controls (Mutti &Hallock, 2003). This study in the Gulf of Califor-nia highlights how certain biogenic assemblagescan be tied to specific nutrient and temperatureranges, thus providing a tool for more preciseinterpretations.
Furthermore, this study serves as a baseline forcomparing carbonate environments worldwide inorder to establish temperature and nutrient bound-aries responsible for production of different typesof carbonates. Data were compiled from moderncarbonate environments and their respectivenutrient and temperature controls in differentparts of the world (Figs 11–13) (Halfar et al.,2004a). Only studies providing at least semiquan-titative information on abundance of carbonateproducers were included in the database. Seventyper cent of published studies lack quantitativeabundance data, therefore in order to facilitatecomparison with present results, published car-bonate components were categorized as rare, com-mon and abundant. Likewise, reliable data onregional oceanography cited within individualstudies are scarce. Oceanographic data were there-fore acquired from global charts (sea-surface tem-perature and salinity; http://www.saa.noaa.gov) orfrom satellite measurements (Chl a). Hence,oceanographic parameters listed in Fig. 13 arederived from surface measurements, even thoughthis might not, in some cases, precisely reflectthe conditions under which benthic carbonatecommunities develop. Annual Chl a valueswere obtained from remote-sensed SeaWiFS data,based on the close correspondence of annuallyaveraged in-situ measurements of Chl a withannual SeaWiFS data in the Gulf of Californiasettings. Even though carbonate environmentsincluded in this compilation are limited to 50 m
0 5 10 15 20 25 30
0·1
1
1
2
3
46
7
8
9 10
11
1213
14
15
17
1819
20
23
30*
24*
31
25
2629
3233
34*
35*36
37
38
39
40
41*
42
43
44
45
Ave
rage
chl
orop
hyll
a (m
g C
hl a
m–3
)
Minimum monthly sea surface temperature (°C)
* Av. salinity >38‰
<1% non-sk. grains + zooxanthell. corals +
green algae + large benthic foraminifera1–20% non-sk. grains + zooxanthell. corals + green
algae + large benthic foraminifera >20% non-sk. grains + zooxanthell. corals +
green algae + large benthic foraminifera
3
Cool-temperate Warm-temperate Tropical
Olig
otro
phic
Eut
roph
icM
esot
roph
ic
Pol
ar
546
16 27
2821
0·05
log-
scal
e
Site IV
Site III
Site II
Site I
22
Heterozoan
Photozoan
Heterozoan-photozoantransitionFig. 11. Compilation of types of
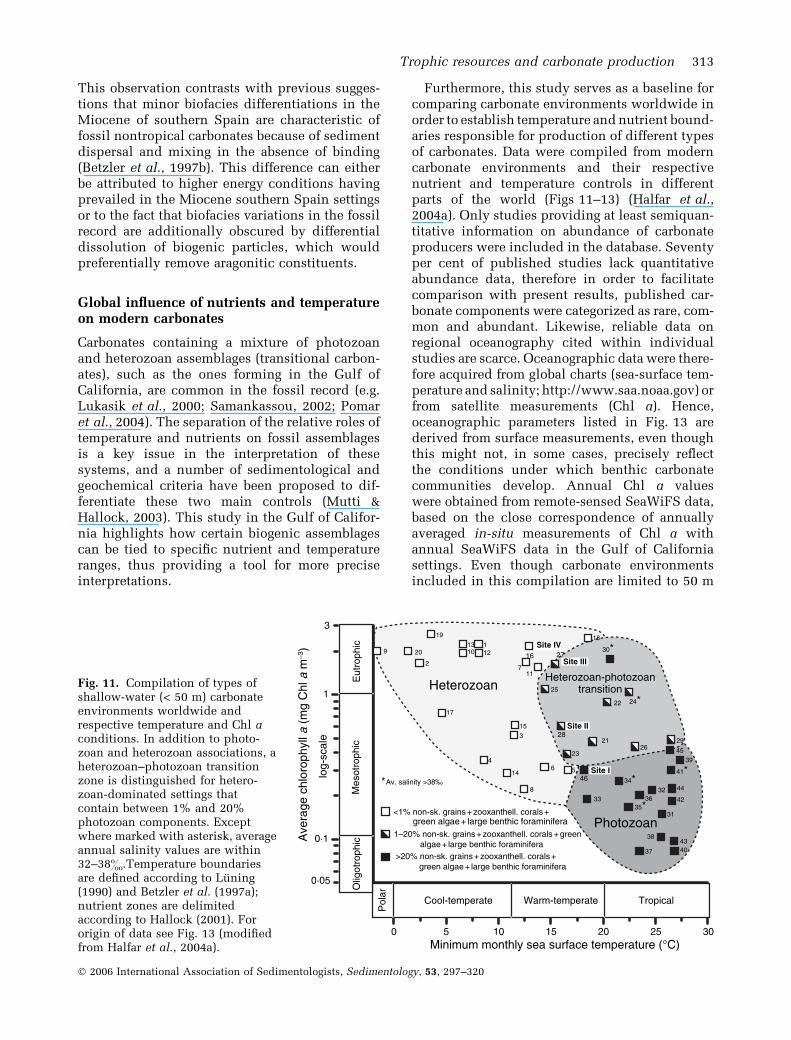
shallow-water (< 50 m) carbonateenvironments worldwide andrespective temperature and Chl aconditions. In addition to photo-zoan and heterozoan associations, aheterozoan–photozoan transitionzone is distinguished for hetero-zoan-dominated settings thatcontain between 1% and 20%photozoan components. Exceptwhere marked with asterisk, averageannual salinity values are within32–38&.Temperature boundariesare defined according to Luning(1990) and Betzler et al. (1997a);nutrient zones are delimitedaccording to Hallock (2001). Fororigin of data see Fig. 13 (modifiedfrom Halfar et al., 2004a).
Trophic resources and carbonate production 313
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

water depth, individual settings usually span awide depth range.
The compilation shows that: (i) as expected,carbonate environments occur under a wide rangeof minimum monthly temperatures and nutrientlevels, (ii) heterozoan and photozoan assemblagesare clearly controlled by both nutrients andtemperatures, and (iii) under high temperaturesand high nutrient levels, a distinct transitionalassemblage can be distinguished (Fig. 11). Thetransitional assemblage is defined as containingbetween 1% and 20% of a combination of non-skeletal grains, zooxanthellate corals, calcareousgreen algae and large benthic foraminifera. Set-tings with these biotic characteristics are onlyfound at minimum monthly temperatures above15 �C and above minimum Chl a contents of0Æ4 mg Chl a m)3. The Chl a level coincides withthe previously described coral reef turn-on, turn-off zone defined by Hallock (2001), which indi-cates that corals, but not coral reefs can occur atnutrient values above that level.
Spatially, these so-called transitional hetero-zoan–photozoan assemblages are found largely inthe warm-temperate realm (e.g. this study; south-ern Brazil – Carannante et al., 1988; Mediterra-nean Sea – Burollet, 1981; Western Australia –
Collins et al., 1997; James et al., 1999) or undertropical temperatures where they are additionallyinfluenced by unusually high or low salinities(Persian Gulf – Hughes Clarke & Keij, 1973;Seibold et al., 1973; East Java Sea – Roberts et al.,1988) (Fig. 11). Transitional heterozoan–photo-zoan carbonate settings, where all three environ-mental controls – temperature, salinity andnutrients – are high, are usually characterizedby a dominance of calcareous green algae. Othercarbonate environments that feature high salini-ties (> 38&), but exhibit oligotrophic or onlyslightly mesotrophic nutrient levels and formunder tropical temperatures, fall within the photo-zoan field. In summary, photozoan carbonatesalways develop under tropical oligotrophic toslightly mesotrophic conditions, whereas hetero-zoan carbonates form from polar to warm-tem-perate and mesotrophic to eutrophic settings. Theheterozoan-photozan transitional assemblage re-quires upper mesotrophic or eutrophic conditionsand warm-temperate to tropical temperatures.
Even though the database indicates a cleardistinction into three different carbonate assem-blages based on amount and type of biota present,it must be noted that the controls studied are notthe only ones influencing carbonate environ-
20°
20°
40°
40° 60° 80° 100° 120° 140° 160°20°0°20°40°80°100°120°140°160°
0°
1627
2846
19
43
3836
37
1520
4031
26
11
22
18
21
39 44
454134
3525
81412
13
110
33
24
2
9
17
29
7
65
30
3
4
23
32
40
42
60°
Fig. 12. Spatial distribution of selected carbonate environments shown in Figs 11 and 13, and their characterizationas photozoan, heterozoan, or transitional photozoan-heterozoan (symbols are same as in Fig. 11).
314 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

ments. Further complexity is added by factorssuch as CO2, aragonite saturation, Mg/Ca ratio,alkalinity, shelf profile, water energy, watertransparency, salinity, oxygen, Ca2+ availability,alkalinity, substrate requirements, and competit-ive displacement as well as biological and evolu-tionary trends (Kleypas et al., 1999; Pomar et al.,2004). In addition, biogeographic differencesneed to be taken into account when comparingcarbonate-producing settings. The southern Gulfof California for example is characterized by alow-diversity assemblage of larger benthic for-aminifera (five species) (Halfar & Ingle, 2003),whereas 11 species of larger benthic foraminiferathrive along the south-western Australian shelf in
an oceanographically comparable setting (Jameset al., 1999).
Implications for fossil carbonates
The calibration of biogenic assemblages to nutri-ent and temperature fields in the Gulf of Califor-nia, complemented with the literature-basedworldwide occurrence of types of carbonatedeposits, provides a new tool to interpret ancientcarbonate deposits. Nutrient influences on car-bonate formation have been inferred from anumber of fossil transitional carbonate environ-ments that exhibit unusual biogenic assemblageswhen compared to their palaeogeographic or
1 Scotland 8 14 35·5 2·2 0 0 0 1 3 2 2 2 1 3 3 Scoffin, 19882 Norway 3 16 34·5 1·6 0 0 0 3 3 2 1 1 0 0 1 Freiwald and Henrich, 19943 New Zealand, Three Kings 12 35·5 0·5 0 0 0 2 3 2 3 2 2 1 3 Nelson et al., 19884 New Zealand, Snares 9 34·5 0·4 0 0 0 1 3 1 3 2 1 2 2 Nelson et al., 19885 Lacepede Shelf, Australia 17 18 35·9 0·3 0 0 0 0 2 0 3 2 0 0 3 James et al., 19926 Southwest Australia, Perth 15 20 35·7 0·3 0 0 0 3 3 1 3 2 1 0 1 James et al., 19997
Southwest Australia, Shark Bay 21 25 35·1 0·9 0 0 0 1 3 1 3 2 1 0 2 James et al., 1999
8 Balearic Platform, Spain 13 26 36·5 0·2 0 0 0 3 3 1 2 2 0 0 2 Fornos Ahr, 19979 Spitsbergen -2 4 34·5 2·0 0 0 0 0 3 1 1 1 0 3 3 Henrich et al., 1997
10 West Shetland Shelf 7 13 35·1 2·0 0 0 0 0 3 1 2 1 2 2 3 Light and Wilson, 199811
Southern Brazil 19 25 36·5 0·5 0 0 1 1 3 2 2 3 0 2 3 Carannante et al., 1988
12 Western English Channel 8 17 34·6 2·0 0 0 0 1 3 3 3 0 0 1 3 Boillot, 196513 Southern Irish Sea 7 16 34·5 2·2 0 0 0 1 3 2 3 1 0 0 3 Dobson et al., 197114 Strait of Otranto (Italy) 11 25 38·3 0·3 0 0 0 1 3 1 1 2 0 0 3 Hesse et al., 197115 US Atlantic shelf 12 26 34·0 0·6 0 0 0 1 3 1 2 3 0 0 3 Pilkey, 196416 Site IV 14 29 35·0 2·2 0 0 0 2 3 2 3 1 1 2 3 this study17 Yellow Sea, China 5 26 31·5 0·8 0 0 0 0 3 2 1 3 0 0 3 Niino and Emery, 196118 West Africa 19 28 34·8 2·4 0 0 0 0 3 0 2 1 1 1 3 McMaster et al., 197119 Alexander Archipelago, Alaska 4 12 32·0 2·6 0 0 0 0 2 2 1 1 0 3 3 Hoskin and Nelson, 196920 Maine, USA 2 17 31·8 2·0 0 0 0 1 3 2 1 1 0 3 3 Raymond and Stetson, 193221
South Africa 15 23 35·5 1·3 0 0 0 1 3 3 2 2 0 0 3 Siesser, 1970
22
Northwest African Shelf 13 26 36·0 1·4 0 0 0 2 3 3 2 3 0 0 2 Summerhayes et al., 1976
23 Houtman Abrolhos, Australia 17 26 35·5 0·4 2 0 0 3 2 1 1 2 0 0 3 Collins et al., 199724 Eastern Persian Gulf 23 33 39·5 1·0 1 0 0 0 3 1 1 2 0 1 3 Seibold et al., 197325 Tunesian Shelf 15 27 38·0 1·1 0 0 3 1 1 0 1 3 0 0 2 Burollet, 198126 Central Brazil 23 27 36·5 0·4 0 0 1 3 2 0 3 2 2 0 2 Carannante et al., 198827 Site III 16 30 35·2 1·6 1 0 0 3 3 2 2 1 1 1 1 this study28 Site II 17 30 35·3 0·6 1 0 0 3 3 2 2 2 1 1 2 this study29 East Java Sea 27 29 31·0 0·5 0 0 3 0 1 0 0 2 0 0 1 Roberts et al., 198830
Lord Howe Island, SW-Pacific 19 25 35·5 0·2 1 0 1 3 1 0 1 1 0 0 1 Kennedy et al., 2002
31 Brasil, Atol de Rocas 26 28 36·5 0·1 1 0 0 3 1 0 0 2 2 0 1 Gherardi and Bosence, 200132 Serranilla Bank, Nicaragua Rise 26 29 36·0 0·2 0 0 3 1 3 0 0 2 1 0 1 Triffleman et al., 199233
Southern Persian Gulf 21 33 40·0 1·7 3 0 0 2 3 2 0 3 0 0 - Hughes Clarke and Keij, 1973
34 Bay of Safaga, Red Sea 22 28 42·5 0·3 3 0 1 2 2 1 1 2 1 0 2 Piller, 199435 Red Sea 23 42·3 0·2 2 1 1 1 2 1 1 2 1 0 3 Gabrie and Montaggioni, 198236 Bahama Bank 24 29 37·0 0·7 2 2 2 1 2 0 0 2 0 0 1 Purdy, 196337 Bahamas, Hogsty Reef 24 28 36·4 0·1 3 1 1 2 2 0 0 1 0 0 1 Milliman, 196738 Grand Cayman 26 29 36·5 0·1 3 0 1 1 2 1 0 2 0 0 1 Kalbfleisch and Jones, 199839 Comoro 28 30 34·5 0·3 3 0 3 2 1 1 0 2 0 0 3 Masse et al., 198940 Northern Brazil 27 25 36·5 0·1 1 0 3 3 2 0 2 2 0 0 2 Carannante et al., 198841 Jeddah, Red Sea 27 32 39·5 0·3 3 0 1 3 2 1 1 1 1 0 1 Montaggioni et al., 198642 Belize, Elmer Reef 27 29 35·5 0·2 3 0 2 1 1 1 0 0 1 1 1 Mazzullo et al., 199243 Jamaica 27 29 35·5 0·1 3 0 3 2 1 1 0 2 0 0 1 Boss Liddell, 198744 Seychelles 27 31 34·8 0·2 3 0 2 3 1 1 0 1 0 0 2 Lewis, 196945 India, East Coast 27 30 33·1 0·5 1 3 0 0 3 1 2 2 0 0 3 Rao, 196446 Site I 19 31 35·1 0·3 3 0 0 2 3 2 1 1 1 1 3 this study
# on
figu
res
11 a
nd 1
2
Site Max
. mon
thly
tem
p. (°
C)
Aver
age
salin
ity (‰
)
Cor
als
(zoo
xant
hella
te)
Ooi
dsC
alca
reou
s gr
een
alga
e
Cor
allin
e re
d al
gae
Mol
lusk
sE
chin
oder
ms
Bryo
zoan
sB
enth
ic fo
ram
inife
raS
erpu
lids
Bar
nacl
esN
on-c
arbo
nate
sed
imen
t
Ref
eren
ce
Min
. mon
thly
tem
p. (°
C)
Av. c
hl a
(mg
Chl
a m
–1)
2218
30
Heterozoan
TransitionalP
hotozoan
and
and
Fig. 13. Shallow-water carbonate environments, their respective oceanographic settings and dominant biota. Assome studies of shallow-water carbonate environments only present semiquantitative data, biota abundances here aregiven as 1 ¼ rare (1–5%), 2 ¼ common (5–20%), 3 ¼ abundant (> 20%). Shading (as in Fig. 11) represents theclassification of carbonate environments into the three associations. Chl a values for each site were derived fromSeaWiFS (avg. of 6 years), whereas temperature and salinity data were obtained from NOAA.
Trophic resources and carbonate production 315
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

palaeoceanographic setting. Shallow-water car-bonates with a cool-water type (heterozoan) orwarm-water type (transitional heterozoan-photo-zoan) assemblage at low-latitude, can often beattributed to higher nutrient supplies. The newcategory defined in this study, the transitionalheterozoan-photozoan assemblage, captures thesecases of low-latitude carbonates dominated byheterozoan assemblages with photozoan compo-nents. Similar cases in ancient settings have beenattributed to upwelling of nutrient-rich watersand associated oligophotic conditions (Saman-kassou, 2002; Wilson & Vecsei, 2005).
In a study of Oligo-Miocene carbonates from theMurray Basin (southern Australia), Lukasik et al.(2000) recognized four bio-lithological associa-tions (large foraminifera–bryozoa, echinoid–bryo-zoan, mollusk and clay facies) and interpretedthem as representing shallow-water (< 50 m)deposition under progressively higher trophicresources, from low mesotrophy to eutrophy. Atemporal shift from inferred high to low meso-trophic conditions was interpreted as reflecting achange in climate from wet to seasonally dryconditions and decreased land-derived nutrientflux. Lower trophic resources during periods ofdry climate resulted in deepening of the euphoticzone, and widespread proliferation of a largerforaminifera photozoan-type fauna. Periods ofwet climate led to increased trophic resources,shallowing of the euphotic zone and widespreaddevelopment of a heterozoan fauna. This case fitsinto the heterozoan–photozoan transitional fieldof Fig. 11, having formed under warm-temperateor tropical meso-eutrophic conditions.
Oligo-Miocene ramp carbonates in the centralMediterranean are characterized by heterozoanfaunas (Mutti et al., 1997; Vecsei & Sanders,1999) with minor contributions from photozoanassemblages (Brandano & Corda, 2002). Faunaldata in association with geochemical criteria(Mutti et al., 1997; Spezzaferri et al., 2001) sug-gest that nutrient supply was a major control overfacies development and variability. Recently,Mutti & Bernoulli (2003) have provided furtherevidence for trophic resource role on the basis of astudy of sea-floor lithification in non-tropicalramp carbonates from the Maiella platform (Italy).Based on the assemblages described, photozoancomponents contribute to the bryozoan-rich car-bonates, placing this Central Apennine settinginto the heterozoan–photozoan transitional fieldof Fig. 11.
Similarly, Pomar et al. (2004) interpret theinfluence of mesotrophic conditions during for-
mation of tropical Miocene rhodalgal sedimentsin the Balearic Islands, based on the fact thatred algae tolerate low light. Photozoan compo-nents, specifically larger benthic foraminifera,occur in the rhodalgal facies of the BalearicIslands, positioning this setting in the hetero-zoan–photozoan transitional field. In contrast,in the middle Miocene Pannonian basin, anassemblage of heterozoans with minor contribu-tions of Porites coral fragments was interpretedby Randazzo et al. (1999) as having formedunder cool-water conditions. Based on theassemblage, however, this system would like-wise plot into the heterozoan–photozoan trans-itional field and should therefore bereinterpreted as a tropical or warm-temperatecarbonate system influenced by mesotrophic oreutrophic conditions. Hence, it is suggested thatquantitative evaluation of the biogenic assem-blages of ancient carbonate successions mayallow for the relative influences of temperatureand nutrients on carbonate production to bedifferentiated.
CONCLUSIONS
The warm-temperate Gulf of California exhibitseutrophic to oligotrophic conditions along anorth–south transect. Fluctuations of nutrientsand temperature as a result of upwelling and tidalmixing have a significant impact on carbonate-producing communities in the Gulf of California.Further controls on carbonate biofacies composi-tion are water depth, and nutrient-dependentlight penetration. Carbonate assemblages rangefrom phototroph-dominated in the southern re-gions to heterotroph-dominated in the northernregions of the Gulf.
A comparison of nutrient and temperatureconditions with modern carbonate compositionson a global scale reveals a progressive reductionof phototrophic carbonate producers withincreasing nutrients and decreasing temperatures.A heterozoan–photozoan transitional assemblageis defined which forms under warm-temperate ortropical mesotrophic or eutrophic conditionscharacterized by low percentages of phototrophicorganisms. Even though in many carbonate-form-ing regions nutrient levels and temperature areintimately linked parameters, application of theseobservations to ancient carbonates should allowfor differentiation of carbonate depositional set-tings characterized by low and high nutrientconditions.
316 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

ACKNOWLEDGEMENTS
We acknowledge the help of Markus Eisele,Steffen Hetzinger and Marcel Strasser in provi-ding additional information on the individualstudy sites. We thank Jose Angel Hinosa foranalysis of water samples, Guillermo Gutierrezfor designing and Gabriel Plata Damian forbuilding oceanographic moorings, and Elvia Plat-a, Tobias Schwennicke, Mario Castro, AntonioResendiz and the CICIMAR staff for field support.This work was supported by the VolkswagenStif-tung, Federal Republic of Germany (grant I/77446). Permission to collect sediment sampleswas granted by SEMARNAT (permit 300702-613-03). Pamela Hallock, Isabel Montanez, and Peir K.Pufahl provided very helpful reviews.
REFERENCES
Alvarez, L.G., Badan-Dangon, A. and Robles, J.M. (1984) La-
grangian observations of near-surface currents in Canal de
Ballenas. CalCOFI Rep., XXV, 35–42.
Alvarez-Borrego, S. (1983) Gulf of California. In: Estuaries andEnclosed Seas (Ed. B.H. Ketchum), Ecosystems of the
World, 26. pp. 427–449. Elsevier, Amsterdam.
Alvarez-Borrego, S. and Lara-Lara, J.R. (1991) The physical
environment and primary productivity of the Gulf of Cali-
fornia. In: The Gulf and Peninsular Province of the Cali-
fornias (Eds B.R.T. Simoneit and J.P. Dauphin), AAPG
Mem., 47, 555–567.
Anthony, K.R.N. (2000) Enhanced particle-feeding capacity in
nearshore reef corals (Great Barrier Reef, Australia). Coral
Reefs, 19, 59–67.
Basso, D. (1998) Deep rhodolith distribution in the Pontian
Islands, Italy: a model for the paleoecology of a temperate
sea. Palaeogeogr. Palaeoclimatol. Palaeoecol., 137, 173–
187.
Beier, E. (1997) A numerical investigation of the annual vari-
ability in the Gulf of California. J. Phys. Oceanogr., 27, 615–
632.
Betzler, C., Brachert, T.C. and Nebelsick, J. (1997a) The warm
temperate carbonate province: a review of the facies, zona-
tions, and delimitations. Courier Forschungs-Institut Senc-
kenberg, 201, 83–99.
Betzler, C., Brachert, T.C., Braga, J.-C. and Martin, J.M.(1997b) Nearshore, temperate, carbonate depositional sys-
tems (lower Tortonian, Agua Amarga Basin, southern
Spain): implications for carbonate sequence stratigraphy.
Sed. Geol., 113, 27–53.
Birkeland, C. (1987) Nutrient availability as a major deter-
minant of differences among coastal hard-substratum com-
munities in different regions of the tropics. In: Differencesbetween Atlantic and Pacific Tropical Marine Coastal Eco-
systems: Community Structure, Ecological Processes, and
Productivity (Ed. C. Birkeland), pp. 45–90. UNESCO Reports
in Marine Science, Paris.
Boillot, G. (1965) Organogenic gradients in the study of neritic
deposits of biological origin: the example of the Western
English Channel. Mar. Geol., 3, 359–367.
Bosence, D.W.J. (1983) The occurrence and ecology of recent
rhodoliths – a review. In: Coated Grains (Ed. T.M. Peryt),
pp. 225–241. Springer-Verlag, Berlin.
Boss, S.K. and Liddell, W.D. (1987) Patterns of sediment
composition of Jamaican fringing reef facies. Sedimentolo-
gy, 34, 77–87.
Brachert, T., Betzler, C., Braga, J.-C. and Martin, J.M. (1998)
Microtaphofazies of a warm-temperate carbonate ramp
(Uppermost Tortonian/Lowermost Messinian, southern
Spain). Palaios, 13, 459–475.
Brandano, M. and Corda, L. (2002) Nutrients, sea level
and tectonics: constrains for the facies architecture of a
Miocene carbonate ramp in central Italy. Terra Nova, 14,257–262.
Brasier, M. (1995) Fossil indicators of nutrient levels, 1:
Eutrophication and climate change. In: Marine Paleoenvi-
ronmental Analysis from Fossils (Eds D.W.J. Bosence and
P.A. Allison), Geol. Soc. Spec. Publ., 83, 133–150.
Bray, N.A. (1988) Water mass formation in the Gulf of Cali-
fornia. J. Geophys. Res., 93, 9223–9240.
Brusca, R.C. and Thomson, D.A. (1975) Pulmo Reef: the only
‘‘coral reef’’ in the Gulf of California. Ciencias Marinas, 2,37–53.
Burollet, P.F. (1981) The Pelagian Sea east of Tunisia: bio-
clastic deposition under temperate climate. Mar. Geol., 44,157–170.
Carannante, G. and Simone, L. (1996) Rhodolith facies in the
central-southern Apennines Mountains, Italy. In: Models for
Carbonate Stratigraphy from Miocene Reef Complexes of
Mediterranean Regions (Eds E.K. Franseen, M. Esteban,
W.C. Ward and J.-M. Rouchy), SEPM Concepts Sedimentol.
Paleontol., 5, 262–275.
Carannante, G., Esteban, M., Milliman, J.D. and Simone, L.(1988) Carbonate lithofacies as paleolatitude indicators:
problems and limitations. Sed. Geol., 60, 333–346.
Carannante, G., Cherchi, A. and Simone, L. (1995) Chlorozoan
versus foramol lithofacies in Upper Cretaceous rudist
limestones. Palaeogeogr. Palaeoclimatol. Palaeoecol., 119,137–154.
Carriquiry, J.D., Sanchez, A. and Camacho Ibar, V.F. (2001)
Sedimentation in the northern Gulf of California after ces-
sation of the Colorado River discharge. Sed. Geol., 144, 37–
62.
Collins, L.B., France, R.E., Zhu, Z.R. and Wyrwoll, K.-H.(1997) Warm-water platform and cool-water shelf carbon-
ates on the Abrolhos Shelf, Southwest Australia. In: Cool-
Water Carbonates (Eds N.P. James and J.A.D. Clarke), SEPM
Spec. Publ., 56, 23–36.
Corda, L. and Brandano, M. (2003) Aphotic zone carbonate
production on a Miocene ramp, Central Apennines, Italy.
Sed. Geol., 161, 55–70.
Crossland, C.J. (1987) Dissolved nutrients in coral reef waters.
In: Perspectives on Coral Reefs (Ed. D.J. Barnes), pp. 56–68.
The Australian Institute of Marine Science, Townsville.
Dobson, M.R., Evans, W.E. and James, K.H. (1971) The sedi-
ment of the floor of the southern Irish Sea. Mar. Geol., 11,27–69.
Dorsey, R.J. and Kidwell, S.M. (1999) Mixed carbonate-silici-
clastic sedimentation on a tectonically active margin:
example from the Pliocene of Baja California Sur, Mexico.
Geology, 27, 935–938.
Dupraz, C. and Strasser, A. (2002) Nutritional modes in coral-
microbialite reefs (Jurassic, Oxfordian, Switzerland): evo-
lution of trophic structure as a response to environmental
change. Palaios, 17, 449–471.
Trophic resources and carbonate production 317
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

Fornos, J.J. and Ahr, W.M. (1997) Temperate carbonates on a
modern, low-energy, isolated ramp: the Balearic Platform,
Spain. J. Sed. Res., 67, 364–373.
Foster, M.S., Riosmena-Rodriguez, R., Steller, D.L. and
Woelkerling, W.M.J. (1997) Living rhodolith beds in the
Gulf of California and their implications for paleoenviron-
mental interpretation. In: Pliocene Carbonates and Related
Facies Flanking the Gulf of California, Baja California,Mexico (Eds M.E. Johnson and J. Ledesma-Vazquez), Geol.
Soc. Am. Spec. Pap., 318, 127–140.
Freiwald, A. and Henrich, R. (1994) Reefal coralline agal
build-ups within the Arctic Circle: morphology and sedi-
mentary dynamics under extreme environmental seasonal-
ity. Sedimentology, 41, 963–984.
Friedman, G.M. (1985) Gulf of Elat (Aquaba). Geological and
sedimentological framework. In: Hypersaline Ecosystems –
The Gavish Sabkha (Eds G.M. Friedman and W.E. Krumb-
ein), Ecological Studies, 53. pp. 39–71. Springer-Verlag,Berlin.
Gabrie, L. and Montaggioni, L. (1982) Sedimentary facies from
the modern coral reefs, Jordan Gulf of Aqaba, Red Sea. Coral
Reefs, 1, 115–124.
Gherardi, D.F. and Bosence, D.W.J. (2001) Composition and
community structure of the coralline algal reefs from Atol
das Rocas, South Atlantic, Brazil. Coral Reefs, 19, 205–
219.
Gischler, E., Grafe, K.-U. and Wiedmann, J. (1994) The Upper
Cretaceous Lacazina limestone in the Basco-Cantabrian and
Iberian Basins of northern Spain: cold-water grain associa-
tions in warm-water environments. Facies, 30, 209–246.
Halfar, J. and Ingle, J.C. (2003) Modern warm-temperate and
subtropical shallow water benthic foraminifera of the
southern Gulf of California, Mexico. J. Foramin. Res., 33,309–329.
Halfar, J., Godinez-Orta, L., Mutti, M., Valdez-Holguin, J.E.and Borges, J.M. (2004a) Nutrient and temperature controls
on modern carbonate production: an example from the Gulf
of California, Mexico. Geology, 32, 213–216.
Halfar, J., Ingle, J.C. and Godinez-Orta, L. (2004b) Modern
heterozoan mixed carbonate-siliciclastic sediments and
environments of the southwestern Gulf of California,
Mexico. Sed. Geol., 165, 93–115.
Hallock, P. (1987) Fluctuations in the trophic resource con-
tinuum: a factor in global diversity cycles? Paleoceano-
graphy, 2, 457–471.
Hallock, P. (1988) The role of nutrient availability in bioero-
sion: Consequences to carbonate buildups. Palaeogeogr.
Palaeoclimatol. Palaeoecol., 63, 275–291.
Hallock, P. (2001) Coral reefs, carbonate sediments, nutrients,
and global change. In: The History and Sedimentology of
Ancient Reef Systems (Ed. G.D. Stanley), pp. 387–427.
Kluwer Academic/Plenum Publishers, New York.
Hallock, P. and Schlager, W. (1986) Nutrient excess and the
demise of coral reefs and carbonate platforms. Palaios, 1,389–398.
Henrich, R., Freiwald, A., Bickert, T. and Schafer, P. (1997)
Evolution of an Arctic open-shelf carbonate platform,
Spitsbergen Bank (Barents Sea). In: Cool-water Carbonates
(Eds N.P. James and J.A.D. Clarke), SEPM Spec. Publ., 56,163–184.
Hesse, R., von Rad, U. and Fabricius, F.H. (1971) Holocene
sedimentation in the Strait of Otranto between the Adriatic
and Ionian Seas (Mediterranean). Mar. Geol., 10, 293–355.
Holmes, R.W. (1970) The secchi disk in turbid coastal waters.
Limnol. Oceanogr., 15, 688–694.
Hoskin, C.M. and Nelson, R.V. (1969) Modern marine car-
bonate sediment, Alexander Archipelago, Alaska. J. Sed.
Pet., 39, 581–590.
Hughes Clarke, M.W. and Keij, A.J. (1973) Organisms as pro-
ducers of carbonate sediment and indicators of environment
in the southern Persian Gulf. In: The Persian Gulf (Ed. B.H.
Purser), pp. 33–56. Springer-Verlag, Berlin.
James, N.P. (1997) The cool-water carbonate depositional
realm. In: Cool-water Carbonates (Eds N.P. James and J.A.D.
Clarke), SEPM Spec. Publ., 56, 1–22.
James, N.P., Bone, Y., von der Borch, C. and Gostin, V.A.(1992) Modern carbonate and terrigenous clastic sediments
on a cool water, high energy, mid-latitude shelf: Lacepede,
southern Australia. Sedimentology, 39, 877–903.
James, N.P., Collins, L.B., Bone, Y. and Hallock, P. (1999)
Subtropical carbonates in a temperate realm: modern sedi-
ments on the southwest Australian shelf. J. Sed. Res., 69,1297–1321.
James, N.P., Feary, D.A., Betzler, C., Bone, Y., Holbourn, A.E.,Quianyu, L., Machiyama, H., Simo, J.A.T. and Surlyk, F.(2004) Origin of late Pleistocene bryozoan reef mounds:
Great Australian Bight. J. Sed. Res., 74, 20–48.
John, C.M., Mutti, M. and Adatte, T. (2003) Mixed carbonate-
siliciclastic record on the North African margin (Malta)-
coupling of weathering processes and mid Miocene climate.
Geol. Soc. Am. Bull., 115, 217–229.
Kalbfleisch, W.B.C. and Jones, B. (1998) Sedimentology of
shallow, hurricane-affected lagoons: Grand Cayman, British
West Indies. J. Coast. Res., 14, 140–160.
Kennedy, D.M., Woodroffe, C.D., Jones, B.D., Dickson, M.E.and Phipps, C.V.G. (2002) Carbonate sedimentation on
subtropical shelves around Lord Howe Island and Balls
Pyramid, southwest Pacific. Mar. Geol., 188, 333–349.
Kleypas, J.A., Buddemeier, R.W., Archer, D., Gattuso, J.-P.,Langdon, C. and Opdyke, B.N. (1999) Geochemical conse-
quences of increased atmospheric carbon dioxide on coral
reefs. Science, 284, 118–120.
Kowalewski, M., Avila-Serrano, G.E., Flessa, K.W. and
Goodfriend, G.A. (2000) Dead delta’s former productivity:
Two trillion shells at the mouth of the Colorado River.
Geology, 28, 1059–1062.
Lees, A. (1975) Possible influence of salinity and temperature
on modern shelf carbonate sedimentation. Mar. Geol., 19,159–198.
Lees, A. and Buller, A.T. (1972) Modern temperate-water and
warm-water shelf carbonates contrasted. Mar. Geol., 13,M67–M73.
Lewis, M.S. (1969) Sedimentary environments and uncon-
solidated carbonate sediments of the fringing coral reefs of
Mahe, Seychelles. Mar. Geol., 7, 95–127.
Light, J.M. and Wilson, J.B. (1998) Cool-water deposition on
the West Shetland Shelf: a modern distally steepened ramp.
In: Carbonate Ramps (Eds V.P. Wright and T.P. Burchette),
Geol. Soc. London, 149, 73–105.
Littler, D.S. and Littler, M.M. (1985) Factors controlling
relative dominance of primary producers on biotic reefs.
Proc. Fifth Int. Coral Reef Congr., Tahiti, vol. 4, pp. 35–
39.
Lukasik, J.J., James, N.P., McGrowan, B. and Bone, Y. (2000)
An epeiric ramp: low-energy, cool-water carbonate facies in
a Tertiary inland sea, Murray Basin, South Australia. Sedi-
mentology, 47, 851–881.
Luning, K. (1990) Seaweeds. Their Environment, Biogeo-graphy, and Ecophysiology. New York, John Wiley & Sons,
589 pp.
318 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

Masse, J.-P., Thomassin, B.A. and Acquaviva, M. (1989) Bio-
clastic sedimentary environments of coral reefs and lagoon
around Mayotte Island (Comoro Archipelago, Mozambique
Channel, SW Indian Ocean). J. Coast. Res., 5, 419–432.
Mazzullo, S.J., Anderson-Underwood, K.E., Burke, C.D. and
Bischoff, W.D. (1992) Holocene coral patch reef ecology and
sedimentary architecture, northern Belize, Central America.
Palaios, 7, 591–601.
McMaster, R.L. and Conover, J.T. (1966) Recent algal stro-
matolites from the Canary Islands. J. Geol., 74, 647–652.
McMaster, R.L., Milliman, J.D. and Ashraf, A. (1971) Con-
tinental shelf and upper slope sediments off Portugese
Guinea, Guinea, and Sierra Leone, West Africa. J. Sed. Pet-
rol., 41, 150–158.
Milliman, J.D. (1967) Carbonate sedimentation on Hogsty Reef,
a Bahamian Atoll. J. Sed. Petrol., 37, 658–676.
Montaggioni, L.F., Behairy, K.A., El-Sayed, M.K. and Yusuf,N. (198) The modern reef complex, Jeddah area, Red Sea: a
facies model for carbonate sedimentation on embryonic
passive margins. Coral Reefs, 5, 127–150.
Mutti, M. and Bernoulli, D. (2003) Early marine lithification
and hardground development on a Miocene ramp (Maiella,
Italy): key surfaces to track changes in trophic resources in
nontropical carbonate settings. J. Sed. Res., 73, 296–308.
Mutti, M. and Hallock, P. (2003) Carbonate systems
along nutrient and temperature gradients: some sedimen-
tological and geochemical constraints. Int. J. Earth Sci., 92,465–475.
Mutti, M., Bernoulli, D. and Stille, P. (1997) Temperate car-
bonate platform drowning linked to Miocene oceanographic
events: Maiella platform margin, Italy. Terra Nova, 9, 122–
125.
Nelson, C.S. (1988) An introductory perspective on non-trop-
ical shelf carbonates. Sed. Geol., 60, 3–12.
Nelson, C.S., Keane, S.L. and Head, P.S. (1988) Non-tropical
carbonate deposits on the modern New Zealand shelf. Sed.
Geol., 60, 71–94.
Niino, H. and Emery, K.O. (1961) Sediments of shallow por-
tions of East China Sea and South China Sea. Geol. Soc. Am.
Bull., 72, 731–762.
Pilkey, O.H. (1964) The size distribution and mineralogy of the
carbonate fraction of United States South Atlantic shelf and
upper slope sediments. Mar. Geol., 2, 121–136.
Piller, W.E. (1994) The northern Bay of Safaga (Red Sea,
Egypt): An actuopalaeontological approach. 4. Thin section
analysis. Beitr. Palaontol. Osterr., 18, 1–73.
Pomar, L., Brandano, M. and Westphal, H. (2004) Environ-
mental factors influencing skeletal grain sediment associa-
tions: a critical review of Miocene examples from the
western Mediterranean. Sedimentology, 51, 627–651.
Pride, C., Thunell, R., Sigman, D., Keigwin, L., Altabet, M. and
Tappa, E. (1999) Nitrogen isotopic variations in the Gulf of
California since the last deglaciation: response to global
climate change. Paleoceanography, 14, 397–409.
Purdy, E.G. (1963) Recent calcium carbonate facies of the
Great Bahama Bank. 2. Sedimentary Facies. J. Geol., 71,472–497.
Randazzo, A.F., Muller, P., Lelkes, G., Juhasz, E. and Hamor,T. (1999) Cool- water limestones of the Pannonian Basinal
System, Middle Miocene, Hungary. J. Sed. Res., 69, 283–
293.
Rao, M.S. (1964) Some aspects of continental shelf sediments
off the east coast of India. Mar. Geol., 1, 44–58.
Raymond, P.E. and Stetson, H.C. (1932) A calcareous beach on
the coast of Maine. J. Sed. Petrol., 2, 51–62.
Reyes-Bonilla, H. and Calderon-Aguilera, L.E. (1999) Popu-
lation density, distribution and consumption rates of three
corallivores at Cabo Pulmo Reef, Gulf of California, Mexico.
Mar. Ecol., 20, 347–357.
Reyes-Bonilla, H. and Lopez-Perez, A. (1998) Biogeographıa
de los Petreos (Scleractinia) del Pacıfico de Mexico. Cien-
cias Marinas, 24, 211–224.
Riegl, B. and Piller, W.E. (2000) Reefs and coral carpets in the
northern Red Sea as models for organism-environment
feedback in coral communities and its reflection growth
fabrics. In: Carbonate Platform Systems: Components andInteractions (Eds E. Insalaco, P.W. Skelton and T.J. Palmer),
Geol. Soc. London Spec. Publ., 178, 71–88.
Riosmena-Rodriguez, R., Woelkerling, W.J. and Foster, M.S.(1999) Taxonomic reassessment of rhodolith-forming spe-
cies of Lithophyllum (Corallinales, Rhodophyta) in the Gulf
of California, Mexico. Phycologia, 38, 401–417.
Rivera, M.G., Riosmena-Rodriguez, R. and Foster, M. (2004)
Age and growth of Lithothamnion muelleri (Corallinales,
Rhodophyta) in the southwestern Gulf of California, Mex-
ico. Ciencias Marinas, 30, 235–249.
Roberts, H.H., Aharon, P. and Phipps, C.V. (1988) Morphology
and sedimentology of Halimeda bioherms from the eastern
Java Sea (Indonesia). Coral Reefs, 6, 161–172.
Samankassou, E. (2002) Cool-water carbonates in a paleo-
equatorial shallow-water environment: the paradox of the
Auernig cyclic sediments (Upper Pennsylvanian, Carnic
Alps, Austria-Italy) and its implications. Geology, 30, 655–
658.
Santamaria-Del-Angel, E., Alvarez-Borrego, S. and Mueller-Karger, F.E. (1994) Gulf of California biogeographic regions
based on coastal zone color scanner imagery. J. Geoph. Res.,
99 (C4), 7411–7421.
Scoffin, T.P. (1988) The environments of production and
deposition of calcareous sediment in the shelf west of
Scotland. Sed. Geol., 60, 107–124.
Seibold, E., Diester, L., Futterer, D., Lange, H., Muller, P.and Werner, F. (1973) Holocene sediments and sedimen-
tary processes in the Iranian part of the Persian Gulf. In:
The Persian Gulf (Ed. B.H. Purser), pp. 57–80. Springer,
Berlin.
Siesser, W.G. (1970) Carbonate components and mineralogy of
the South African coastal limestones and limestones of the
Agulhas bank. Trans. Geol. Soc. South Africa, 73, 49–63.
Spezzaferri, S., Mutti, M. and Spiegler, D. (2001) Integrated
planktonic foraminifera, Bolboforma and carbon isotope
stratigraphy in a mid/early late Miocene carbonate ramp
setting from the Acreide area (Sicily). Mar. Micropaleontol.,43, 223–238.
Summerhayes, C.P., Milliman, J.D., Briggs, S.R., Bee, A.G.and Hogan, C. (1976) Northwest African shelf sediments:
influence of climate and sedimentary processes. J. Geol., 84,277–300.
Tomczak, M. and Godfrey, J.S. (1994) Regional Oceanogra-
phy: An Introduction. Pergamon, Oxford, 442 pp.
Triffleman, N.J.H., Hallock, P. and Hine, A.C. (1992) Mor-
phology, sediments, and depositional environments of a
small carbonate platform; Serranilla Bank, Nicaraguan Rise,
Southwest Caribbean Sea. J. Sed. Petrol., 62, 591–606.
Valentine, J.W. (1973) Evolutionary Paleoecology of the Mar-
ine Biosphere. Prentice-Hall, Princeton, NJ, 511 pp.
Vecsei, A. (2003) Nutrient control of the global occurrence of
isolated carbonate banks. Int. J. Earth Sci., 92, 467–481.
Vecsei, A. and Sanders, D.G.H. (1999) Facies analysis and
sequence stratigraphy of a Miocene warm-temperate car-
Trophic resources and carbonate production 319
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320

bonate ramp, Monagna della Maiella, Italy. Sed. Geol. 123,103–127.
Wilson, M.E.J. and Vecsei, A. (2005) The apparent paradox of
abundant foramol facies in low latitudes: their environ-
mental significance and effect on platform development.
Earth-Sci. Rev, 69, 133–168.
Wood, R. (1993) Nutrients, predation and the history of reef-
building. Palaios, 8, 526–543.
Manuscript received 3 September 2004; revisionaccepted 12 September 2005
320 J. Halfar et al.
� 2006 International Association of Sedimentologists, Sedimentology, 53, 297–320
Copyright © 2022 FDOKUMEN




















