
Calcium induces pro-anabolic effects on human primary osteoblasts associated with acquisition of...
8
Calcium induces pro-anabolic effects on human primary osteoblasts associated with acquisition of mature osteocyte markers Katie J. Welldon a , David M. Findlay a , Andreas Evdokiou b , Renee T. Ormsby a , Gerald J. Atkins a,⇑ a Bone Cell Biology Group, Centre for Orthopaedic & Trauma Research, The University of Adelaide, Adelaide, SA 5005, Australia b Breast Cancer Research Unit, Basil Hetzel Institute, Discipline of Surgery, The University of Adelaide, Woodville, SA 5011, Australia article info Article history: Received 20 March 2013 Received in revised form 16 May 2013 Accepted 11 June 2013 Available online 18 June 2013 Keywords: Human osteoblast Calcium Mineralisation Gene expression OPG/RANKL ASARM abstract Calcium, in combination with vitamin D, is an effective treatment for osteoporosis. Since bone mineralisation occurs concurrently with osteoblast to osteocyte transition, we hypothesised that calcium would stimulate this process. The effect of calcium (1.8–11.8 mM) was tested on human primary osteoblast (NHBC) differen- tiation in vitro. Cultures were assayed for cell-associated mineral and gene expression associated with osteo- blast differentiation and mineralisation. Treatment with calcium resulted in a striking dose- and time- dependent increase in cell-associated mineralisation. Calcium appeared to promote osteoblast to osteocyte differentiation, as indicated by increased expression of osteocalcin (OCN), E11, dentin matrix protein 1 (DMP1) and SOST mRNA. The expression of the osteoclast inhibitor, osteoprotegerin, was dramatically enhanced by calcium. Calcium also increased the ratio of PHEX mRNA expression relative to that of MEPE, sug- gesting a mechanism for the pro-anabolic effect. Consistent with this, calcium-dependent mineralisation was reversed in the presence of MEPE–ASARM peptides. This study suggests that calcium promotes osteoblast to osteocyte transition and concurrent matrix mineralisation, at least in part through the PHEX–MEPE axis. Ó 2013 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Calcium, in combination with vitamin D, has been shown to be effective in the treatment of osteoporosis, a condition typically associated with a low BMD, increased fracture risk and a reduced osteocyte number (Tang et al., 2007; Qiu et al., 2003; Chang et al., 2000). Although these treatments are widely accepted, the exact mechanisms, by which these agents act, are yet to be fully elucidated (Chapuy et al., 1994; Horwood et al., 1998). Blood cal- cium (Ca 2+ ) levels are strictly regulated, with total calcium levels in healthy individuals usually maintained within the range of 2.2–2.6 mM and between 1.1 and 1.3 mM for ionised calcium (Dvorak and Riccardi, 2004). However, soluble calcium levels in bone interstitial fluid may be higher. For example, calcium concen- trations can range from 8 to 40 mM in the immediate vicinity of the resorbing osteoclast (Silver et al., 1988; Berger et al., 2001; Maeno et al., 2005). Osteocytes are also capable of releasing signif- icant amounts of calcium, demonstrated during lactation (Qing et al., 2012), suggesting that they are exposed to elevated concen- trations of extracellular calcium, although actual levels have not been reported. Osteoid osteocytes appear to mediate calcium phos- phate crystal formation in mineralising bone (Atkins and Findlay, 2012; Atkins et al., 2011; Barragan-Adjemian et al., 2006; Prideaux et al., 2012) and these differentiating osteocytes may also be dependent on the surrounding mineral for the full expression of their phenotype. However, understanding the vital process of min- eralisation is challenged by the difficulty of replicating it in vitro. We hypothesised that calcium itself would be a regulator of the mineralisation process, and to that extent would act as an anabolic agent in bone. Using a human primary osteoblast model, we found that calcium potently increased calcium phosphate mineral depo- sition. This response was associated with osteoblast differentiation to an osteocyte-like phenotype and a dramatic increase in the expression of OPG. Since the effect of calcium on mineralisation was blocked in the presence of MEPE–ASARM, our results suggest the involvement of the PHEX–MEPE axis in calcium-mediated min- eralisation. We conclude that human osteoblast to osteocyte tran- sition is sensitive to extracellular calcium. 2. Materials and methods 2.1. Cell culture Normal human bone-derived cells (NHBC), obtained from inter- trochanteric trabecular bone samples from 10 female donors (Do- nors 1–10) undergoing primary hip replacement surgery for osteoarthritis, with informed consent and approval by the Human Ethics Committee of the Royal Adelaide Hospital, were processed for culture and the cells propagated as described previously (Atkins et al., 2009a). Briefly, cells were cultured in a-MEM, which con- 0303-7207/$ - see front matter Ó 2013 Elsevier Ireland Ltd. All rights reserved. http://dx.doi.org/10.1016/j.mce.2013.06.013 ⇑ Corresponding author. Address: Centre for Orthopaedic & Trauma Research, The University of Adelaide, North Terrace, Adelaide, SA 5005, Australia. Tel.: +61 882223107. E-mail address: [email protected] (G.J. Atkins). Molecular and Cellular Endocrinology 376 (2013) 85–92 Contents lists available at SciVerse ScienceDirect Molecular and Cellular Endocrinology journal homepage: www.elsevier.com/locate/mce
Transcript of Calcium induces pro-anabolic effects on human primary osteoblasts associated with acquisition of...

Molecular and Cellular Endocrinology 376 (2013) 85–92
Contents lists available at SciVerse ScienceDirect
Molecular and Cellular Endocrinology
journal homepage: www.elsevier .com/locate /mce
Calcium induces pro-anabolic effects on human primary osteoblastsassociated with acquisition of mature osteocyte markers
0303-7207/$ - see front matter � 2013 Elsevier Ireland Ltd. All rights reserved.http://dx.doi.org/10.1016/j.mce.2013.06.013
⇑ Corresponding author. Address: Centre for Orthopaedic & Trauma Research, TheUniversity of Adelaide, North Terrace, Adelaide, SA 5005, Australia. Tel.: +61882223107.
E-mail address: [email protected] (G.J. Atkins).
Katie J. Welldon a, David M. Findlay a, Andreas Evdokiou b, Renee T. Ormsby a, Gerald J. Atkins a,⇑a Bone Cell Biology Group, Centre for Orthopaedic & Trauma Research, The University of Adelaide, Adelaide, SA 5005, Australiab Breast Cancer Research Unit, Basil Hetzel Institute, Discipline of Surgery, The University of Adelaide, Woodville, SA 5011, Australia
a r t i c l e i n f o a b s t r a c t
Article history:Received 20 March 2013Received in revised form 16 May 2013Accepted 11 June 2013Available online 18 June 2013
Keywords:Human osteoblastCalciumMineralisationGene expressionOPG/RANKLASARM
Calcium, in combination with vitamin D, is an effective treatment for osteoporosis. Since bone mineralisationoccurs concurrently with osteoblast to osteocyte transition, we hypothesised that calcium would stimulatethis process. The effect of calcium (1.8–11.8 mM) was tested on human primary osteoblast (NHBC) differen-tiation in vitro. Cultures were assayed for cell-associated mineral and gene expression associated with osteo-blast differentiation and mineralisation. Treatment with calcium resulted in a striking dose- and time-dependent increase in cell-associated mineralisation. Calcium appeared to promote osteoblast to osteocytedifferentiation, as indicated by increased expression of osteocalcin (OCN), E11, dentin matrix protein 1(DMP1) and SOST mRNA. The expression of the osteoclast inhibitor, osteoprotegerin, was dramaticallyenhanced by calcium. Calcium also increased the ratio of PHEX mRNA expression relative to that of MEPE, sug-gesting a mechanism for the pro-anabolic effect. Consistent with this, calcium-dependent mineralisation wasreversed in the presence of MEPE–ASARM peptides. This study suggests that calcium promotes osteoblast toosteocyte transition and concurrent matrix mineralisation, at least in part through the PHEX–MEPE axis.
� 2013 Elsevier Ireland Ltd. All rights reserved.
1. Introduction et al., 2012) and these differentiating osteocytes may also be
Calcium, in combination with vitamin D, has been shown to beeffective in the treatment of osteoporosis, a condition typicallyassociated with a low BMD, increased fracture risk and a reducedosteocyte number (Tang et al., 2007; Qiu et al., 2003; Changet al., 2000). Although these treatments are widely accepted, theexact mechanisms, by which these agents act, are yet to be fullyelucidated (Chapuy et al., 1994; Horwood et al., 1998). Blood cal-cium (Ca2+) levels are strictly regulated, with total calcium levelsin healthy individuals usually maintained within the range of2.2–2.6 mM and between 1.1 and 1.3 mM for ionised calcium(Dvorak and Riccardi, 2004). However, soluble calcium levels inbone interstitial fluid may be higher. For example, calcium concen-trations can range from 8 to 40 mM in the immediate vicinity ofthe resorbing osteoclast (Silver et al., 1988; Berger et al., 2001;Maeno et al., 2005). Osteocytes are also capable of releasing signif-icant amounts of calcium, demonstrated during lactation (Qinget al., 2012), suggesting that they are exposed to elevated concen-trations of extracellular calcium, although actual levels have notbeen reported. Osteoid osteocytes appear to mediate calcium phos-phate crystal formation in mineralising bone (Atkins and Findlay,2012; Atkins et al., 2011; Barragan-Adjemian et al., 2006; Prideaux
dependent on the surrounding mineral for the full expression oftheir phenotype. However, understanding the vital process of min-eralisation is challenged by the difficulty of replicating it in vitro.We hypothesised that calcium itself would be a regulator of themineralisation process, and to that extent would act as an anabolicagent in bone. Using a human primary osteoblast model, we foundthat calcium potently increased calcium phosphate mineral depo-sition. This response was associated with osteoblast differentiationto an osteocyte-like phenotype and a dramatic increase in theexpression of OPG. Since the effect of calcium on mineralisationwas blocked in the presence of MEPE–ASARM, our results suggestthe involvement of the PHEX–MEPE axis in calcium-mediated min-eralisation. We conclude that human osteoblast to osteocyte tran-sition is sensitive to extracellular calcium.
2. Materials and methods
2.1. Cell culture
Normal human bone-derived cells (NHBC), obtained from inter-trochanteric trabecular bone samples from 10 female donors (Do-nors 1–10) undergoing primary hip replacement surgery forosteoarthritis, with informed consent and approval by the HumanEthics Committee of the Royal Adelaide Hospital, were processedfor culture and the cells propagated as described previously (Atkinset al., 2009a). Briefly, cells were cultured in a-MEM, which con-

86 K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92
tains 1.8 mM calcium, with 10% fetal calf serum FCS, L-glutamine(2 mM), HEPES (1 mM) and ascorbate 2-phosphate (100 lM) at37 �C/5% CO2, in a humidified incubator. Cells grown to confluencewere enzymatically removed from dishes using collagenase, dis-pase, and/or trypsin, and plated into plastic tissue culture multi-well dishes or flasks. Experiments were performed on cells pas-saged up to three times. For experimental purposes, cell culturemedium contained graded concentrations of additional CaCl2 (Sig-ma–Aldrich, St. Louis, MO, USA) from 0.1 mM to 10 mM, with orwithout the CaSR inhibitor NPS2390 (Sigma–Aldrich, St. Louis,MO, USA) (Kwak et al., 2005). Treatments were initiated 24 h aftercell seeding and continued for the duration of the experiment. Insome experiments, the effect of a tri-phosphorylated peptide rep-resentative of the MEPE–ASARM motif (ASARM-PO4) (Atkinset al., 2011; Rowe et al., 2005) was investigated. In these experi-ments, NHBC were cultured under standard mineralising condi-tions for 21 days, whereupon medium was replaced with mediacontaining the indicated concentrations of calcium and ASARM-PO4 (NeoMPS Inc., San Diego, CA), and cultured for a further 7 days.
2.2. Cell viability assays
NHBC viability was determined after 7 days of culture using anumber of methods, including trypan blue dye exclusion and cas-pase 3 activity. Additionally, apoptotic nuclei morphology wasdetermined by 40,6-diamidine-20-phenylindole dihydrochloridestaining (DAPI, Roche Diagnostics, Castle Hill, NSW, Australia), asdescribed previously (Atkins et al., 2002). The cell surface expres-sion of annexin V, an indicator of apoptosis was used in conjunc-tion with staining, using the fluorescent viability stain, 7AAD toinvestigate the viability of NHBC in increasing concentrations ofcalcium, as described previously (Findlay et al., 2002).
2.3. Mineralisation of cells
The effect of calcium on NHBC ability to form a mineralised ma-trix was determined using a method described previously (Gron-thos et al., 1994). Briefly, NHBC were incubated in triplicate inwells of a 96-well plate (seeded at 8 � 103 cells/well) in a-MEMcontaining 10% FCS, dexamethasone (10�8 M), KH2PO4 1.8 mM),and HEPES (10 mM), with varying concentrations of calcium. Med-ium was replaced every 3–4 days, and incubation continued for upto 6 weeks. Mineralisation was determined using a variation of theVon Kossa technique, as described previously (Atkins et al., 2009a).For the purpose of quantification, representative images were ob-tained, using a Nikon D1H digital camera attached to an invertedmicroscope (Nikon TE300) with a x20 objective. These were thenconverted to grayscale, and Image Tool version 2.2 (MolecularDynamics�, Microsoft) was then used to quantify the amount ofmineralisation per image (Atkins et al., 2009a).
To determine the composition of the mineralised layer, separateexperiments were performed on NHBC grown in 8-chamber slides.NHBC were grown under mineralising conditions for 21 days in thepresence of calcein (Sigma–Aldrich) (Atkins et al., 2009b) and variousconcentrations of calcium. The samples were then fixed with 4% para-formaldehyde and fluorescent labeling visualised using a Nikon con-focal microscope. Matched samples were prepared for analysis bycritical point drying and then electron dispersive X-ray spectroscopyanalysis was performed on a XL-20 scanning electron microscope todetermine relative elemental composition of the mineral.
2.4. Proliferation rate of cells
Cell proliferation was measured using 5,6-carboxyfluoresceindiacetate, succinimidyl ester (CFSE) (Molecular probes, Eugene,OR, USA), a nuclear stain that is partitioned evenly between daughter
cells as they divide (Atkins et al., 2009a; Lyons et al., 2001). Afterbeing irreversibly labeled with CFSE, NHBC were seeded into tissueculture plastic and cultured as indicated for 7 days (Atkins et al.,2009a). The level of CFSE staining was then determined by fluores-cence-activated cell scanning analysis (FACS), using a FACStarPLUS
flow cytometer (Becton Dickinson, Sunnyvale, CA, USA). The numberof cell doublings was determined from listmode data using ModFitLT™ software (Verity Software House, Topsham, NJ, USA), as de-scribed previously (Atkins et al., 2009a).
2.5. Expression of osteogenic and osteoclastogenic genes
Total RNA was extracted from NHBC using Trizol™ (Life Technol-ogies, Gaithersburg, MD) and complementary DNA (cDNA; Super-script III, Promega Corp., Madison, WI) was prepared as describedpreviously (Atkins et al., 2007). Gene expression was analysed byreal-time RT-PCR, using the SYBR Green incorporation technique,as we have described previously (Atkins et al., 2007). Relative geneexpression between samples was calculated using the comparativecycle threshold (CT) method (DCT), calculated by the formulaDCT = 2�(CT(x)�CT(GAPDH)), where CT(x) is the CT of the gene of interest,and CT(GAPDH) is the CT for glyceraldehyde-3-phosphate dehydro-genase, a housekeeping gene we have used previously for examininggene expression in long-term human osteoblast cultures (Atkinset al., 2011, 2009a,b,c). Similar results were obtained when 18S ribo-somal RNA (rRNA) was used as a housekeeping gene (data notshown). Messenger RNA-specific oligonucleotide primers were de-signed in-house to flank intron–exon boundaries, and were pur-chased from Geneworks (Thebarton, SA, Australia). Real-timeoligonucleotide primers for the amplification of receptor activatorof NF-jB ligand (RANKL), OPG, SOST/sclerostin, E11/podoplanin,DMP1, OCN, matrix extracellular phosphoglycoprotein (MEPE), tis-sue non-specific alkaline phosphatase (TNAP), 18S rRNA and GAPDHwere described previously (Atkins et al., 2011, 2009a,b,c, 2007; Vin-cent et al., 2009).
2.6. Expression of secreted OPG
Supernatants were collected at intervals for up to 42 days fromcultures under mineralising conditions, with calcium concentra-tions ranging from 1.8–11.8 mM. Secreted OPG levels in culturesupernatants were measured by an in-house enzyme-linkedimmunoassay (ELISA) using matched antibodies purchased fromR&D Systems Inc (Minneapolis, MN, USA) and quantified using astandard curve generated using recombinant human OPG (Pepro-tech, Rocky Hill, NJ) (Atkins et al., 2009a).
2.7. Statistical analysis
One way analysis of variance (ANOVA), followed by Tukey’spost hoc analysis, was used to examine differences in gene expres-sion studies from individual donors’ cells. Paired Student’s T-testswere used to compare effects in data pooled from multiple donors’cells. A p value < 0.05 was considered to be significant.
3. Results
3.1. In vitro mineralisation in response to calcium
NHBC were cultured under mineralising conditions for up to4 weeks in the presence of extracellular calcium concentrationsranging from the standard media content of 1.8 mM up to11.8 mM. Mineralisation of the cell layer was visualised usingVon Kossa staining of phosphate. Although there were donordependent differences in the level of mineralisation, calcium con-
B
H
*
MIN
ERAL
ISED
AR
EA (%
)
0 20406080
100
120
1.8 1.9 2.3 2.8 3.8 6.8 11.8
* *
*
*
[Ca2+] (mM)
2.26 0.40CaCl2 11.8mM
1.84 0.06CaCl2 2.6mM
1.66 0.05CaCl2 1.8mM
0.77 0.02MEM
Ca/PTreatmentC
50µm
Ca 1.8mM
F G 50µm
10µm
D E Ca 2.6mM
10µm
D14 D28 D42
1.8
2.8
3.8
6.8
11.8
[Ca] (mM)A
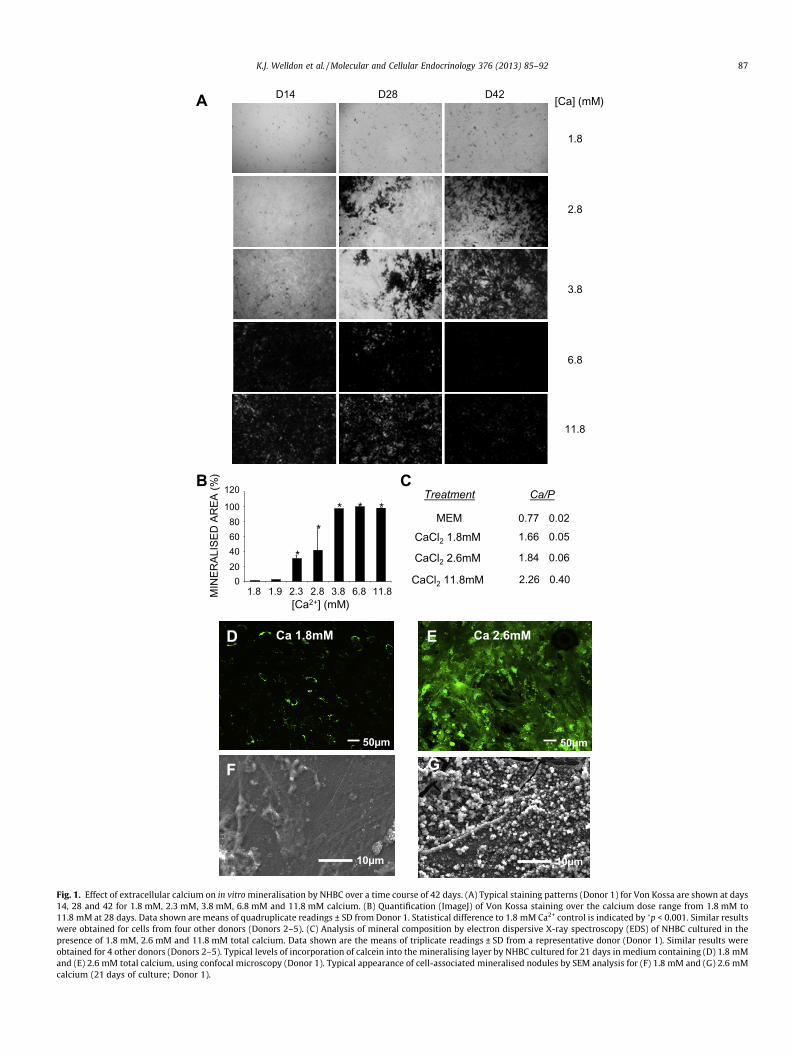
Fig. 1. Effect of extracellular calcium on in vitro mineralisation by NHBC over a time course of 42 days. (A) Typical staining patterns (Donor 1) for Von Kossa are shown at days14, 28 and 42 for 1.8 mM, 2.3 mM, 3.8 mM, 6.8 mM and 11.8 mM calcium. (B) Quantification (ImageJ) of Von Kossa staining over the calcium dose range from 1.8 mM to11.8 mM at 28 days. Data shown are means of quadruplicate readings ± SD from Donor 1. Statistical difference to 1.8 mM Ca2+ control is indicated by �p < 0.001. Similar resultswere obtained for cells from four other donors (Donors 2–5). (C) Analysis of mineral composition by electron dispersive X-ray spectroscopy (EDS) of NHBC cultured in thepresence of 1.8 mM, 2.6 mM and 11.8 mM total calcium. Data shown are the means of triplicate readings ± SD from a representative donor (Donor 1). Similar results wereobtained for 4 other donors (Donors 2–5). Typical levels of incorporation of calcein into the mineralising layer by NHBC cultured for 21 days in medium containing (D) 1.8 mMand (E) 2.6 mM total calcium, using confocal microscopy (Donor 1). Typical appearance of cell-associated mineralised nodules by SEM analysis for (F) 1.8 mM and (G) 2.6 mMcalcium (21 days of culture; Donor 1).
K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92 87

0
1
2
3
4
5
6
7
8
9
1.8 2.8 6.8 11.8Ca2+ CONCENTRATION (mM)
CEL
L D
EATH
(%)
A
B 90
100 *** *** ***
88 K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92
sistently increased mineralisation in all donor cells in a dose andtime-dependent manner (Fig. 1A and B). Mineralisation was max-imal for all donors’ cells tested at 6.8 mM calcium (Fig. 1B). Addi-tionally, analysis of the mineralised layer in cells treated with1.8 mM and 2.6 mM of calcium by EDS analysis demonstratedCa:P ratios similar to those found in hydroxyapatite (Fig. 1C). Atthe highest concentration of calcium tested, there was increasedcalcium relative to phosphorus, indicating an altered crystal struc-ture, due possibly to limiting concentrations of inorganic phos-phate, which was kept constant in these cultures. In separateexperiments, the calcium mimetic, calcein, was used to visualisethe relative capacity of cells to incorporate calcium into their ma-trix at different calcium concentrations. As shown in Fig. 1D and E,a higher extracellular concentration of calcium promoted in-creased calcein uptake. SEM imaging of these samples showedcalcospherulite-like structures associated with cell processes, con-sistent with normal mineralisation and differentiation to osteo-cyte-like cells (Barragan-Adjemian et al., 2006) at 2.6 mMcalcium (Fig. 1F and G).
0
10
20
30
40
50
60
70
80
1.8 2.8 6.8 11.8
DIV
IDED
CEL
LS (%
)
Ca2+ CONCENTRATION (mM)
Fig. 2. Effect of varying calcium concentrations on cell viability and proliferation.(A) NHBC were cultured in 96 well plates with a range of concentrations of calciumfor 7 days and cell viability determined using trypan blue dye exclusion. (B) NHBC
3.2. Effect of calcium on osteoblast viability and proliferation
The effect of extracellular calcium on cell proliferation and sur-vival of NHBC was investigated over a culture period of 7 days.Treatment with calcium at 1.8 mM, 2.8 mM, 6.8 mM and11.8 mM showed no effect on the percentage of viable cells, as as-sessed by trypan blue dye exclusion, which indicated close to 100%viability (Fig. 2A). Other measures of cell death, including caspase 3activity, the expression of annexin V on the cell surface and alterednuclear morphology visualised with DAPI staining, all indicatedthat calcium did not induce apoptosis at any of the concentrationstested (data not shown). The effect of calcium on cell proliferation,examined using the CFSE staining technique, revealed that the per-centage of divided cells increased from that at 1.8 mM but wasmaximal at 2.8 mM calcium (Fig. 2B).
were labelled with CFSE, cultured for 7 days and analysed by flow cytometry. Thepercentage of divided cells was calculated using Modfit software, as described inMaterials and Methods. Data shown are the means ± SEM of quadruplicate samples.Statistical difference to 1.8 mM Ca is indicated by ���(p < 0.001). Data shown wereobtained using cells from Donor 1. Similar results were obtained for cells from thetwo other donors tested (Donors 2 & 3).
3.3. Gene expression effects of calcium
The expression of several osteoblastic genes was examined inNHBC cultured for up to 42 days in media containing a range of cal-cium concentrations, under conditions otherwise permissive formineralisation. Consistent with a maturational effect, calciumdose-dependently increased OCN mRNA at early time points(Fig. 3A) and decreased mRNA encoding TNAP (Fig. 3B). Interest-ingly, the expression of the mineralisation inhibitor, MEPE, wassuppressed in response to calcium for all donors’ cells tested(Fig. 3C). The expression of the endopeptidase PHEX, a pro-miner-alisation gene, was notably increased at the earliest time point as-sayed but was suppressed at the higher concentrations of calciumon days 7 and 14 (Fig. 3D). Increased exposure to calcium appearedto alter the kinetics of expression of other osteoblast differentia-tion-associated genes. Messenger RNA levels of E11, a pre-osteo-cyte-associated gene, was elevated in early cultures at thehighest calcium concentration used and was again elevated at latertime points at the 2.8 and 6.8 mM calcium concentrations (Fig. 3E).A marked increase in expression of the mature osteocyte-associ-ated genes, DMP1 (Fig. 3F) and SOST (Fig. 3G), was also observedin all donors’ cells tested, consistent with osteoblast differentiationinto osteocytes. Since the SOST gene product, sclerostin, is a potentinhibitor of mineralisation by late osteoblasts/pre-osteocytes (At-kins et al., 2011), its elevation suggests a negative feedback re-sponse by the cells to high doses of extracellular calcium andsubsequent mineralisation (see Fig. 1). Overall, these results sug-gest that increased extracellular calcium levels promote differenti-ation of NHBC to an osteocyte-like phenotype.
The response of NHBC to calcium was also investigated interms of a potential effect on bone resorption by analysing genesinvolved in osteoclastogenesis. As depicted in Fig. 4A, RANKLmRNA expression was increased late in cultures (Fig. 4A)concomitant with the late phases of mineralisation (see Fig. 1).Calcium triggered a robust, dose-dependent induction of OPGmRNA expression (Fig. 4B), equating to an increased OPG:RANKLmRNA ratio (Fig. 4C). Calcium also elicited a strong OPG secre-tory response (Fig. 4D) for all donors’ cells tested. Overall, thisgene expression profile is consistent with calcium eliciting anet anabolic effect.
3.4. Effect of MEPE–ASARM peptides
The PHEX–MEPE axis has been shown previously to be centralto the mineralisation process by osteoblasts, with PHEX being animportant osteocyte-expressed inhibitor of MEPE and MEPE–ASARM peptides (Quarles, 2003; Rowe et al., 2004). We haveshown altered expression of both MEPE and PHEX in sclerostin-mediated reduction of mineralisation and bone formation (Atkinset al., 2011). Accordingly, we showed that MEPE–ASARM peptidesare powerful inhibitors of mineralisation by NHBC (Atkins et al.,

A 1.8 mM2.8 mM6.8 mM11.8 mM
[Ca]
0
5
10
15
20
25
30
0 10 20 30 40 50
OC
N:G
APD
H m
RN
A (x
10-3
)
a,ba
a,b
a,b,c
b
TIME (DAY)
B
0
0.5
1.0
1.5
2.0
2.5
0 10 20 30 40 50
TIME (DAY)
TNAP
:GAP
DH
mR
NA
(x10
-5)
a,ba,b
a,b
a,b,c
C
0
1.0
2.0
3.0
4.0
0 10 20 30 40 50
TIME (DAY)
MEP
E:G
APD
H m
RN
A (x
10-5
)
c a a,b
E
0 5
1015202530354045
0 10 20 30 40 50
TIME (DAY)
E11:
GAP
DH
mR
NA
(x10
-3)
a
b,c
b
a
F
TIME (DAY)
0
0.05
0.10
0.15
0.20
0.25
0 10 20 30 40 50
DM
P1:G
APD
H m
RN
A (x
10-3
)
a
a
a
a,b
a,b
b,c
0
0.4
0.8
1.2
1.6
0 10 20 30 40 50
SOST
:GAP
DH
mR
NA
(x10
-2)
TIME (DAY)
a
a,b
a,c
a
G
D
0
1.02.0
3.04.0
5.06.0
7.08.0
9.0
0 10 20 30 40 50TIME (DAY)
PHEX
:GAP
DH
mR
NA
(x10
-5) a,b,c
a,b,c
a b
Fig. 3. Effect of varying the calcium concentration on NHBC mRNA expression of osteoblast and osteocyte markers. NHBC were grown under mineralising conditions for42 days, with Ca at 1.8, 2.8, 6.8 and 11.8 mM. RT-PCR was performed in triplicate on total RNA prepared over the time course for (A) OCN, (B) TNAP, (C) MEPE, (D) PHEX, (E)E11, (F) DMP1 and (G) SOST. Data shown are means ± SEM. a, b, and c indicate significant differences (p < 0.001) for 11.8, 6.8 and 2.8, respectively, using cells from Donor 1.Similar results were obtained from 4 donors’ cells tested (Donors 1–4).
K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92 89

B
a,b
0
2
4
6
8
10
12
14
0 10 20 30 40 50
TIME (DAY)
OPG
:GAP
DH
mR
NA
(x10
-3)
a,b,ca,b,c
a,b,c
a
a
a,c
D
0
100
200
300
400
500
600
0 10 20 30 40 50
TIME (DAY)
SEC
RET
ED O
PG (n
g/m
l)
a,b
a,b,ca,b,c
a,b,c
a,b,ca,b,c
a,b,c
A
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8R
ANKL
:GAP
DH
mR
NA
(x10
-3) a,b,c
a,b a,ca a,b,c a,c
0 10 20 30 40 50
TIME (DAY)
1.82.86.811.8
[Ca] (mM)
0
20
40
60
80
100
120
0 10 20 30 40 50
TIME (DAY)
OPG
:RAN
KL m
RN
A
C
a,b,c
a,b,c
a,b,c
a a,c
Fig. 4. Effect of Ca on bone resorption-associated gene expression. NHBC were grown under mineralising conditions for 42 days in differentiation media containing Ca2+ at1.8, 2.8, 6.8 and 11.8 mM. RT-PCR was performed in triplicate on total RNA prepared over the time course, for (A) RANKL and (B) OPG mRNA, normalised to GAPDH mRNAexpression levels. (C) The OPG:RANKL mRNA ratio for the data shown in A & B. D) Supernatants sampled 3 days after feeding in each case were assayed for OPG secretion byan in-house enzyme-linked immunosorbent assay (ELISA) system, as described in the Methods. In all cases data shown are means ± SD. a, b, and c indicate significantdifferences from control (1.8 mM Ca) (p < 0.01) for 11.8, 6.8 and 2.8 mM, respectively, using cells from Donor 1. Similar results were obtained with cells from 3 other donors’cells tested (Donors 2, 3 and 4).
90 K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92
2011). Since we had observed that calcium dose-dependentlyinhibited the expression of MEPE mRNA (see Fig. 3C) and promotedPHEX mRNA expression at an early time point, we examined theratio of PHEX:MEPE mRNA. This revealed apparent co-ordinateregulation of these genes as a function of calcium concentration.Calcium at 2.8, 6.8 and 11.8 mM all enhanced the PHEX:MEPEmRNA ratio over the first 7 days of NHBC differentiation, withthe highest dose of calcium producing a sustained increase overa 28 day period (Fig. 5A). It was possible that the relative decreasein MEPE expression in response to calcium directly promoted theincorporation of calcium into the matrix. To test this, we generatedmineralised cultures of NHBC and after onset of mineralisation re-placed media with medium containing various concentrations ofcalcium in the presence or absence of ASARM-PO4 peptide, at con-centrations we have previously shown to inhibit human osteoblastmineralisation (Atkins et al., 2011), and cultured for a further7 days. As shown in Fig. 5B, mineralisation was enhanced by in-creased calcium at 2.6 and 5 mM over this period. No effect ofASARM-PO4 was observed in cultures under standard mineralisingconditions (1.8 mM calcium), likely since mineralisation did not in-crease sufficiently over the 7 day period. However, ASARM-PO4 po-tently abrogated the increase seen at the two higherconcentrations of calcium tested. These data imply that calcium-mediated suppression of MEPE and induction of PHEX expressionmay be at least partly responsible for the ability of the cells to in-crease calcium apposition.
4. Discussion
Consistent with previous reports (Maeno et al., 2005; Gabusiet al., 2011; Takagishi et al., 2006), this study demonstrates thatthe process of osteoblast-mediated mineralisation is sensitive tothe extracellular concentration of calcium, up to around 4 mM. Inthe presence of 2–4 mM extracellular calcium, the Ca:P ratio ofaround 1.8 for the mineralised cell layers was similar to that re-ported in bone (Cassella et al., 1995). In addition, the presence ofcalcospherulite-like structures is consistent with bio-mineralisa-tion as it has been described for bone (Barragan-Adjemian et al.,2006; Midura et al., 2009). Our findings suggest that adult humanlong-bone derived osteoblasts are able to tolerate even very highconcentrations of calcium, which is generally consistent with pre-vious findings, although concentrations of calcium between 10 and15 mM exhibited a mild negative effect on cell survival in mouseneonatal calvarial cells (Maeno et al., 2005).
Matrix mineralisation occurs concomitantly with the osteoblasttransition to osteocytes, as elegantly demonstrated by Barragan-Adjemian and colleagues, using the mouse late osteoblast cell line,MLO-A5 (Barragan-Adjemian et al., 2006). Indeed, it appears thatbone matrix mineralisation is required for osteocyte mineralisation(Irie et al., 2008). Consistent with the genetic re-programming ofosteoblasts through this transition, expression of a number ofgenes has been identified to be up or down-regulated during thisprocess. Thus, we found that calcium induced an increase in OCN

UTMEPE- ASARM 1 µM
MEPE- ASARM 10 µM MEPE- ASARM 20 µM
*
0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
1.8 2.6 5
Aliz
arin
Red
(405
nm
)
****
****
*
Ca2+ concentration (mM)
A
B
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
9.0
0 10 20 30 40 50
TIME (DAYS)
PHEX
:MEP
E m
RN
A
b,c
a,b,c
a,b,cb b,c
1.8 mM2.8 mM6.8 mM11.8 mM
[Ca]
Fig. 5. (A) The effect of calcium on the PHEX:MEPE mRNA ratio. NHBC were grownunder mineralising conditions for 42 days, with Ca at 1.8, 2.8, 6.8 and 11.8 mM. RT-PCR was performed for MEPE and PHEX mRNA expression (as depicted in Fig. 3) andPHEX expression was calculated relative to that of MEPE. Data shown aremeans ± SEM. a, b, and c indicate significant differences (p < 0.01) for 11.8, 6.8and 2.8, respectively, using cells from Donor 1. (B) Effect of ASARM-PO4 peptides oncalcium induced mineralisation. NHBC were grown under mineralising conditionsfor 21 days. Additional calcium to the indicated concentrations was then added for7 days ± ASARM-PO4. Data shown are means ± SEM. �p < 0.05, ��p < 0.01. Similarresults were obtained using cells from 2 donors (Donors 9 and 10).
K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92 91
mRNA expression and decreased TNAP expression, similar to find-ings in rat primary osteoblasts and mouse 2T3 cells (Dvorak et al.,2004). Decreased alkaline phosphatase activity has also been re-ported to be associated with the accumulation of calcium in matrixvesicles (Dvorak et al., 2004). The biphasic expression of E11 in ourcultures, a gene preferentially expressed by pre-osteocytes andthought to be involved in cell process formation (Zhang et al.,2006), was consistent with the promotion of cell differentiationby calcium. Sclerostin/SOST, an important inhibitor of bone forma-tion, is a marker of mature osteocytes (Poole et al., 2005; Winkleret al., 2003). In the current study, high concentration of calcium in-duced a large and rapid increase in SOST mRNA expression. Sincewe have shown that sclerostin retards osteoblast differentiationbeyond the pre-osteocyte stage (Atkins et al., 2011), it is possiblethat calcium up-regulation of SOST serves to extend the pre-oste-ocyte phase to accommodate further calcium incorporation. Cal-
cium also increased the mRNA levels of DMP1, another marker ofmature osteocytes and, potentially of relevance here, a nucleatorand promoter of mineralisation (Narayanan et al., 2003; Fenget al., 2006). Calcium also had effects on the expression of PHEX,a gene associated with promotion of mineralisation and inhibitionof MEPE activity (Quarles, 2003; Rowe, 2012), which were mostevident when the ratio of PHEX:MEPE mRNA was calculated. Thecalcium-induced increase in PHEX:MEPE mRNA expression, sug-gested a mechanism, by which NHBC could increase mineral incor-poration. We and others have demonstrated that the post-translational products of the MEPE gene, phosphorylated MEPE–ASARM peptides, are potent inhibitors of mineralisation (Roweet al., 2005, 2004), including that by human primary osteoblasts(Atkins et al., 2011). In the current study, addition of MEPEASARM-PO4 peptides to calcium treated cultures completelyinhibited the calcium-enhanced incorporation of calcium over a7-day period, suggesting that the calcium-induced increase in thePHEX:MEPE mRNA expression ratio may be important in the ensu-ing pro-anabolic response. Calcium also increased RANKL mRNAexpression over time, perhaps consistent with RANKL now beingrecognised as a functional marker of osteocytes in the adult skele-ton (Nakashima et al., 2011; Xiong et al., 2011). Despite this in-crease in RANKL mRNA expression, the overwhelming responseto calcium was a dramatic increase in the expression of the RANKLantagonist, OPG, consistent with a net pro-anabolic effect.
In summary, our findings are consistent with calcium having ananabolic effect on human osteoblasts, increasing the transition ofosteoblasts into an osteocyte-like phenotype. The precise mecha-nisms governing the complex process of bone mineralisation re-main to be fully elucidated but our results support a cellmediated modulation of mineralisation, which is sensitive to extra-cellular calcium levels. Mineralisation appears to be orchestratedby newly formed osteocytes so that the differentiation of osteo-blasts into osteocytes is not separable from physiological minerali-sation (Atkins and Findlay, 2012). Evidence also supports theability of mature osteocytes to remove and replace mineral frombone, in order to contribute to calcium homeostasis (Tazawaet al., 2004; Yajima et al., 2010). Since this ‘osteocytic osteolysis’is most easily observed in situations of high calcium demand, suchas lactation (Qing et al., 2012), it is possible that this process is alsosensitive to extracellular calcium, although this remains to beinvestigated.
Disclosures
The authors have nothing to disclose.
Acknowledgements
The authors thank Professor Peter Rowe (University of KansasMedical Center, KS, USA) for the provision of ASARM-PO4 peptide.We also gratefully acknowledge the orthopaedic surgeons andnursing staff of The Department of Orthopaedics and Trauma, Roy-al Adelaide Hospital for support and cooperation in the collectionof femoral specimens. KW was supported by an Australian Post-graduate Award (APA). This work was supported by the NationalHealth and Medical Research Council of Australia (NHMRC).
References
Atkins, G., Findlay, D., 2012. Osteocyte regulation of bone mineral: a little give andtake. Osteoporos Int., 2067–2079.
Atkins, G.J., Bouralexis, S., Evdokiou, A., Hay, S., Labrinidis, A., Zannettino, A.C.,Haynes, D.R., Findlay, D.M., 2002. Human osteoblasts are resistant to Apo2L/TRAIL-mediated apoptosis. Bone 31, 448–456.
Atkins, G.J., Anderson, P.H., Findlay, D.M., Welldon, K.J., Vincent, C., Zannettino, A.C.,O’Loughlin, P.D., Morris, H.A., 2007. Metabolism of vitamin D(3) in human

92 K.J. Welldon et al. / Molecular and Cellular Endocrinology 376 (2013) 85–92
osteoblasts: Evidence for autocrine and paracrine activities of 1alpha,25-dihydroxyvitamin D(3). Bone 40, 1517–1528.
Atkins, G.J., Welldon, K.J., Halbout, P., Findlay, D.M., 2009a. Strontium ranelatetreatment of human primary osteoblasts promotes an osteocyte-like phenotypewhile eliciting an osteoprotegerin response. Osteoporos. Int. 20, 653–664.
Atkins, G.J., Welldon, K.J., Wijenayaka, A.R., Bonewald, L.F., Findlay, D.M., 2009b.Vitamin K promotes mineralization, osteoblast-to-osteocyte transition, and ananticatabolic phenotype by {gamma}-carboxylation-dependent and -independent mechanisms. Am. J. Physiol. Cell Physiol. 297, C1358–C1367.
Atkins, G.J., Welldon, K.J., Holding, C.A., Haynes, D.R., Howie, D.W., Findlay, D.M.,2009c. The induction of a catabolic phenotype in human primary osteoblastsand osteocytes by polyethylene particles. Biomaterials 30, 3672–3681.
Atkins, G.J., Rowe, P.S., Lim, H.P., Welldon, K.J., Ormsby, R., Wijenayaka, A.R.,Zelenchuk, L., Evdokiou, A., Findlay, D.M., 2011. Sclerostin is a locally actingregulator of late-osteoblast/preosteocyte differentiation and regulatesmineralization through a MEPE–ASARM-dependent mechanism. J. BoneMiner. Res. 26, 1425–1436.
Barragan-Adjemian, C., Nicolella, D., Dusevich, V., Dallas, M.R., Eick, J.D., Bonewald,L.F., 2006. Mechanism by which MLO-A5 late osteoblasts/early osteocytesmineralize in culture: similarities with mineralization of lamellar bone. Calcif.Tissue Int. 79, 340–353.
Berger, C.E., Rathod, H., Gillespie, J.I., Horrocks, B.R., Datta, H.K., 2001. Scanningelectrochemical microscopy at the surface of bone-resorbing osteoclasts:evidence for steady-state disposal and intracellular functionalcompartmentalization of calcium. J. Bone Miner. Res. 16, 2092–2102.
Cassella, J.P., Garrington, N., Stamp, T.C., Ali, S.Y., 1995. An electron probe X-raymicroanalytical study of bone mineral in osteogenesis imperfecta. Calcif. TissueInt. 56, 118–122.
Chang, Y.L., Stanford, C.M., Keller, J.C., 2000. Calcium and phosphatesupplementation promotes bone cell mineralization: implications forhydroxyapatite (HA)-enhanced bone formation. J. Biomed. Mater. Res. 52,270–278.
Chapuy, M.C., Arlot, M.E., Delmas, P.D., Meunier, P.J., 1994. Effect of calcium andcholecalciferol treatment for three years on hip fractures in elderly women. Bmj308, 1081–1082.
Dvorak, M.M., Riccardi, D., 2004. Ca2+ as an extracellular signal in bone. Cell Calcium35, 249–255.
Dvorak, M.M., Siddiqua, A., Ward, D.T., Carter, D.H., Dallas, S.L., Nemeth, E.F.,Riccardi, D., 2004. Physiological changes in extracellular calcium concentrationdirectly control osteoblast function in the absence of calciotropic hormones.Proc. Natl. Acad. Sci. USA 101, 5140–5145.
Feng, J.Q., Ward, L.M., Liu, S., Lu, Y., Xie, Y., Yuan, B., Yu, X., Rauch, F., Davis, S.I.,Zhang, S., Rios, H., Drezner, M.K., Quarles, L.D., Bonewald, L.F., White, K.E., 2006.Loss of DMP1 causes rickets and osteomalacia and identifies a role forosteocytes in mineral metabolism. Nat. Genet. 38, 1310–1315.
Findlay, D.M., Raggatt, L.J., Bouralexis, S., Hay, S., Atkins, G.J., Evdokiou, A., 2002.Calcitonin decreases the adherence and survival of HEK-293 cells by a caspase-independent mechanism. J. Endocrinol. 175, 715–725.
Gabusi, E., Manferdini, C., Grassi, F., Piacentini, A., Cattini, L., Filardo, G., Lambertini,E., Piva, R., Zini, N., Facchini, A., Lisignoli, G., 2011. Extracellular calciumchronically-induced human osteoblasts effects: Specific modulation ofosteocalcin and collagen type XV. J. Cell Physiol..
Gronthos, S., Graves, S.E., Ohta, S., Simmons, P.J., 1994. The STRO-1+ fraction of adulthuman bone marrow contains the osteogenic precursors. Blood 84, 4164–4173.
Horwood, N.J., Elliott, J., Martin, T.J., Gillespie, M.T., 1998. Osteotropic agentsregulate the expression of osteoclast differentiation factor and osteoprotegerinin osteoblastic stromal cells. Endocrinology 139, 4743–4746.
Irie, K., Ejiri, S., Sakakura, Y., Shibui, T., Yajima, T., 2008. Matrix mineralization as atrigger for osteocyte maturation. J. Histochem. Cytochem. 56, 561–567.
Kwak, J.O., Kwak, J., Kim, H.W., Oh, K.J., Kim, Y.T., Jung, S.M., Cha, S.H., 2005. Theextracellular calcium sensing receptor is expressed in mouse mesangial cellsand modulates cell proliferation. Exp. Mol. Med. 37, 457–465.
Lyons, A.B., Hasbold, J., Hodgkin, P.D., 2001. Flow Cytometric Analysis of CellDivision History Using Dilution of Carboxyfluorescein Diacetate SuccinimidylEster, a Stably Integrated Fluorescent Probe. Academic Press.
Maeno, S., Niki, Y., Matsumoto, H., Morioka, H., Yatabe, T., Funayama, A., Toyama, Y.,Taguchi, T., Tanaka, J., 2005. The effect of calcium ion concentration onosteoblast viability, proliferation and differentiation in monolayer and 3Dculture. Biomaterials 26, 4847–4855.
Midura, R.J., Vasanji, A., Su, X., Midura, S.B., Gorski, J.P., 2009. Isolation ofcalcospherulites from the mineralization front of bone. Cells Tissues Organs189, 75–79.
Nakashima, T., Hayashi, M., Fukunaga, T., Kurata, K., Oh-Hora, M., Feng, J.Q.,Bonewald, L.F., Kodama, T., Wutz, A., Wagner, E.F., Penninger, J.M., Takayanagi,H., 2011. Evidence for osteocyte regulation of bone homeostasis through RANKLexpression. Nat. Med. 17, 1231–1234.
Narayanan, K., Ramachandran, A., Hao, J., He, G., Park, K.W., Cho, M., George, A.,2003. Dual functional roles of dentin matrix protein 1. Implications inbiomineralization and gene transcription by activation of intracellular Ca2+
store. J. Biol. Chem. 278, 17500–17508.Poole, K.E., van Bezooijen, R.L., Loveridge, N., Hamersma, H., Papapoulos, S.E., Lowik,
C.W., Reeve, J., 2005. Sclerostin is a delayed secreted product of osteocytes thatinhibits bone formation. Faseb J. 19, 1842–1844.
Prideaux, M., Loveridge, N., Pitsillides, A.A., Farquharson, C., 2012. Extracellularmatrix mineralization promotes E11/gp38 glycoprotein expression and drivesosteocytic differentiation. PLoS One 7, e36786.
Qing, H., Ardeshirpour, L., Pajevic, P.D., Dusevich, V., Jahn, K., Kato, S., Wysolmerski,J., Bonewald, L.F., 2012. Demonstration of osteocytic perilacunar/canalicularremodeling in mice during lactation. J. Bone Miner. Res. 27, 1018–1029.
Qiu, S., Rao, D.S., Palnitkar, S., Parfitt, A.M., 2003. Reduced iliac cancellous osteocytedensity in patients with osteoporotic vertebral fracture. J. Bone Miner. Res. 18,1657–1663.
Quarles, L.D., 2003. FGF23, PHEX, and MEPE regulation of phosphate homeostasisand skeletal mineralization. Am. J. Physiol. Endocrinol. Metab. 285, E1–E9.
Rowe, P.S., 2012. The chicken or the egg: PHEX, FGF23 and SIBLINGs unscrambled.Cell Biochem. Funct. 30, 355–375.
Rowe, P.S., Kumagai, Y., Gutierrez, G., Garrett, I.R., Blacher, R., Rosen, D., Cundy, J.,Navvab, S., Chen, D., Drezner, M.K., Quarles, L.D., Mundy, G.R., 2004. MEPE hasthe properties of an osteoblastic phosphatonin and minhibin. Bone 34, 303–319.
Rowe, P.S., Garrett, I.R., Schwarz, P.M., Carnes, D.L., Lafer, E.M., Mundy, G.R.,Gutierrez, G.E., 2005. Surface plasmon resonance (SPR) confirms that MEPEbinds to PHEX via the MEPE–ASARM motif: a model for impaired mineralizationin X-linked rickets (HYP). Bone 36, 33–46.
Silver, I.A., Murrills, R.J., Etherington, D.J., 1988. Microelectrode studies on the acidmicroenvironment beneath adherent macrophages and osteoclasts. Exp. CellRes. 175, 266–276.
Takagishi, Y., Kawakami, T., Hara, Y., Shinkai, M., Takezawa, T., Nagamune, T., 2006.Bone-like tissue formation by three-dimensional culture of MG63 osteosarcomacells in gelatin hydrogels using calcium-enriched medium. Tissue Eng. 12, 927–937.
Tang, B.M., Eslick, G.D., Nowson, C., Smith, C., Bensoussan, A., 2007. Use of calciumor calcium in combination with vitamin D supplementation to prevent fracturesand bone loss in people aged 50 years and older: a meta-analysis. Lancet 370,657–666.
Tazawa, K., Hoshi, K., Kawamoto, S., Tanaka, M., Ejiri, S., Ozawa, H., 2004. Osteocyticosteolysis observed in rats to which parathyroid hormone was continuouslyadministered. J. Bone Miner. Metab. 22, 524–529.
Vincent, C., Findlay, D.M., Welldon, K.J., Wijenayaka, A.R., Zheng, T.S., Haynes, D.R.,Fazzalari, N.L., Evdokiou, A., Atkins, G.J., 2009. Pro-inflammatory cytokines TNF-related weak inducer of apoptosis (TWEAK) and TNFalpha induce the mitogen-activated protein kinase (MAPK)-dependent expression of sclerostin in humanosteoblasts. J. Bone Miner. Res. 24, 1434–1449.
Winkler, D.G., Sutherland, M.K., Geoghegan, J.C., Yu, C., Hayes, T., Skonier, J.E.,Shpektor, D., Jonas, M., Kovacevich, B.R., Staehling-Hampton, K., Appleby, M.,Brunkow, M.E., Latham, J.A., 2003. Osteocyte control of bone formation viasclerostin, a novel BMP antagonist. Embo J. 22, 6267–6276.
Xiong, J., Onal, M., Jilka, R.L., Weinstein, R.S., Manolagas, S.C., O’Brien, C.A., 2011.Matrix-embedded cells control osteoclast formation. Nat. Med. 17, 1235–1241.
Yajima, A., Inaba, M., Tominaga, Y., Nishizawa, Y., Ikeda, K., Ito, A., 2010. Increasedosteocyte death and mineralization inside bone after parathyroidectomy inpatients with secondary hyperparathyroidism. J. Bone Miner. Res. 25, 2374–2381.
Zhang, K., Barragan-Adjemian, C., Ye, L., Kotha, S., Dallas, M., Lu, Y., Zhao, S., Harris,M., Harris, S.E., Feng, J.Q., Bonewald, L.F., 2006. E11/gp38 selective expression inosteocytes: regulation by mechanical strain and role in dendrite elongation.Mol. Cell Biol. 26, 4539–4552.




















