
Changes in protein expression associated with chronic in vitro exposure of hexavalent chromium to...
11
Changes in protein expression associated with chronic in vitro exposure of hexavalent chromium to osteoblasts and monocytes: A proteomic approach Vijay Krishna Raghunathan, 1 M. Helen Grant, 1 Elizabeth M. Ellis 2 1 Bioengineering Unit, Wolfson Centre, 106 Rottenrow, University of Strathclyde, Glasgow, G4 0NW, United Kingdom 2 Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, Glasgow, G4 0NR, United Kingdom Received 29 August 2008; accepted 20 November 2008 Published online 20 February 2009 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jbm.a.32396 Abstract: Cr (VI) is a well-recognized environmental toxin and carcinogen. It is known to be released from orthopedic metal implants in-situ by biocorrosion and is speculated to play a role in periprosthetic osteolysis. It is hence essential to understand its long-term biological effects. We have assessed the in vitro responses of osteoblasts and monocytes to chronic exposure (3 weeks) to Cr (VI), at concentrations that have been measured in patients with metal implants, using two-dimensional gel electrophoresis. Cr (VI) exposure resulted in a differential time-dependent regulation of gly- colytic, stress, and cytoskeletal proteins. The proteins that have been found to be altered in expression play an essential role in normal cellular functioning such as energy metabo- lism, cell signaling, and proliferation. The results highlight the complex molecular changes that occur in both cell types with long-term exposure to Cr and may be useful in estab- lishing a series of clinically useful biomarkers to monitor long-term use of metallic implants. Ó 2009 Wiley Periodi- cals, Inc. J Biomed Mater Res 92A: 615–625, 2010 Key words: chronic in vitro toxicity; hexavalent chromium; monocytes; osteoblasts; metal orthopaedic implants INTRODUCTION Stainless steel and cobalt-chrome alloys are often used in the manufacture of medical implants for joint replacement. Hip replacement is the second most common elective operation in Europe and often involves the use of metal-on-metal (MOM) articula- tions. MOM implants are associated with high circu- lating levels of metal ions especially cobalt and chro- mium 1 ; however, the long-term effects associated with exposure to metal ions have not been investi- gated. It has been reported that 97% of the chro- mium released on corrosion of stainless steel implants is in its hexavalent form. 2 This ion has been shown to cause direct toxicity to the cells of the immune system. It induces oxidative damage and apoptosis in lymphocytes and in macrophages. 3–5 Although the mechanisms involved in osteolysis induced by metal ions/particles are unclear, there are reports suggesting that tissue damage by necro- sis 6 and cell death by apoptosis 7 occur at the site of the implant. In addition to these overtly toxic effects, the release of metal ions from MOM can also induce an adaptive response in cells, a process that may be influenced by or may influence inflammatory mechanisms. Once released from an implant, Cr (VI) can enter the circulation. It can cross cell membranes via non- specific anionic channels and is reduced to Cr (III) by intracellular reductants (enzymatic and nonenzy- matic), in the process generating reactive intermedi- ates and oxygen species (ROS). The reduction of Cr (VI) by reduced glutathione (GSH) and glutathione reductase has been well documented. 8 These reactive intermediates have been postulated to interfere with cellular signaling cascades, cell proliferation, and activation of cytokines and transcription factors. 9 Cr (VI) is known to lead to toxicity in various cell types in vitro after acute exposure. 4,10 Various bio- chemical and molecular parameters of toxicity have been measured previously. Chromate compounds have been shown to cause cell-cycle arrest, inhibit expression of heme oxygenase-1, 11 disrupt the actin cytoskeleton 12 and cause apoptosis via ROS medi- Correspondence to: M. H. Grant; e-mail: m.h.grant@strath. ac.uk Contract grant sponsor: DePuy International Ltd. Ó 2009 Wiley Periodicals, Inc.
Transcript of Changes in protein expression associated with chronic in vitro exposure of hexavalent chromium to...

Changes in protein expression associated with chronicin vitro exposure of hexavalent chromium to osteoblastsand monocytes: A proteomic approach
Vijay Krishna Raghunathan,1 M. Helen Grant,1 Elizabeth M. Ellis21Bioengineering Unit, Wolfson Centre, 106 Rottenrow, University of Strathclyde, Glasgow, G4 0NW, United Kingdom2Strathclyde Institute of Pharmacy and Biomedical Sciences, University of Strathclyde, Glasgow,G4 0NR, United Kingdom
Received 29 August 2008; accepted 20 November 2008Published online 20 February 2009 in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/jbm.a.32396
Abstract: Cr (VI) is a well-recognized environmental toxinand carcinogen. It is known to be released from orthopedicmetal implants in-situ by biocorrosion and is speculated toplay a role in periprosthetic osteolysis. It is hence essentialto understand its long-term biological effects. We haveassessed the in vitro responses of osteoblasts and monocytesto chronic exposure (3 weeks) to Cr (VI), at concentrationsthat have been measured in patients with metal implants,using two-dimensional gel electrophoresis. Cr (VI) exposureresulted in a differential time-dependent regulation of gly-colytic, stress, and cytoskeletal proteins. The proteins that
have been found to be altered in expression play an essentialrole in normal cellular functioning such as energy metabo-lism, cell signaling, and proliferation. The results highlightthe complex molecular changes that occur in both cell typeswith long-term exposure to Cr and may be useful in estab-lishing a series of clinically useful biomarkers to monitorlong-term use of metallic implants. � 2009 Wiley Periodi-cals, Inc. J Biomed Mater Res 92A: 615–625, 2010
Key words: chronic in vitro toxicity; hexavalent chromium;monocytes; osteoblasts; metal orthopaedic implants
INTRODUCTION
Stainless steel and cobalt-chrome alloys are oftenused in the manufacture of medical implants forjoint replacement. Hip replacement is the secondmost common elective operation in Europe and ofteninvolves the use of metal-on-metal (MOM) articula-tions. MOM implants are associated with high circu-lating levels of metal ions especially cobalt and chro-mium1; however, the long-term effects associatedwith exposure to metal ions have not been investi-gated. It has been reported that 97% of the chro-mium released on corrosion of stainless steelimplants is in its hexavalent form.2 This ion hasbeen shown to cause direct toxicity to the cells of theimmune system. It induces oxidative damage andapoptosis in lymphocytes and in macrophages.3–5
Although the mechanisms involved in osteolysisinduced by metal ions/particles are unclear, there
are reports suggesting that tissue damage by necro-sis6 and cell death by apoptosis7 occur at the site ofthe implant. In addition to these overtly toxic effects,the release of metal ions from MOM can also inducean adaptive response in cells, a process that maybe influenced by or may influence inflammatorymechanisms.
Once released from an implant, Cr (VI) can enterthe circulation. It can cross cell membranes via non-specific anionic channels and is reduced to Cr (III)by intracellular reductants (enzymatic and nonenzy-matic), in the process generating reactive intermedi-ates and oxygen species (ROS). The reduction of Cr(VI) by reduced glutathione (GSH) and glutathionereductase has been well documented.8 These reactiveintermediates have been postulated to interfere withcellular signaling cascades, cell proliferation, andactivation of cytokines and transcription factors.9
Cr (VI) is known to lead to toxicity in various celltypes in vitro after acute exposure.4,10 Various bio-chemical and molecular parameters of toxicity havebeen measured previously. Chromate compoundshave been shown to cause cell-cycle arrest, inhibitexpression of heme oxygenase-1,11 disrupt the actincytoskeleton12 and cause apoptosis via ROS medi-
Correspondence to: M. H. Grant; e-mail: [email protected] grant sponsor: DePuy International Ltd.
� 2009 Wiley Periodicals, Inc.

ated activation of p53,13 NF-jB, AP-1,14 JNK,MAPK,15 Src-family kinases, and caspase-3.16 Ratosteoblast FFC cells exposed to concentrations aslow as 1 lM showed a reduction in viability, alkalinephosphatase activity, and collagen synthesis within48 h.17,18 Previously, Ye and Shi19 have used micro-arrays to demonstrate changes in expression of awide range of genes in A549 epithelial cells exposedto 300 lM potassium chromate for 2 h. However,few previous studies have examined the changes inprotein levels caused by chronic exposure to concen-trations that may be found clinically in patients withmetallic implants.
We have previously reported that chronic (up to 4weeks) exposure to low concentrations of Cr (VI)(0.05, 0.1, and 0.5 lM) can alter GSH levels and theexpression and activities of GSH regulating enzymes[g-glutamate cysteine ligase (GCL) and glutathionereductase] in both FFC (osteoblast) and U937 (mono-cyte) cells.20 This indicates that Cr (VI) treatment isable to induce a protective response. However, therehas been no previous study that has investigated thechronic effects of Cr (VI) on global changes in pro-tein levels.
In this study, we have exposed FFC cell line andU937 cell line to 0, 0.05, 0.1, and 0.5 lM Cr (VI) con-tinuously for 3 weeks and have studied the changesin protein expression using two-dimensional gelelectrophoresis. The aim is to understand the toxiclong-term effects of Cr (VI), to develop a series ofclinically useful biomarkers, and to provide an indi-cator to the susceptibility of cells during prolongedexposure to chromium.
MATERIALS AND METHODS
Cell culture
Immortalized osteoblasts (FFC cell line) were derived inthe laboratory from rat neonatal calvaria by transfectionwith T-antigen and grown as described previously.21 Theywere seeded at 6 3 104 cell/cm2 in Dulbecco’s ModifiedEagle’s Medium (DMEM) supplemented with 1% (v/v)nonessential amino acids. U937 [Human leukemic mono-cyte lymphoma cell line; European Collection of Cell Cul-tures] cells were cultured (1.5 3 105 cells/mL) in RPMI1640 medium supplemented with 1% (v/v) L-glutamine.Both media contained 10% (v/v) foetal bovine serum(FBS), 50 Units/mL penicillin, and 50 lg/mL streptomycin.
Cr (VI) solutions were freshly prepared using chromiumoxide (CrO3) (Alfa Aesar, Johnson Matthey company, UK)and diluted to the required concentrations (0.05 lM, 0.1lM, and 0.5 lM) in growth medium under sterile condi-tions. Both cell lines were grown continuously in the aboveconcentrations of Cr (VI) in 75 cm2 flasks, and passagedtwice every week, for 4 weeks, at a split ratio of 1:10. Sam-
ples were taken every 7 days for measurement of proteinand 2D gel electrophoresis.
2D gel electrophoresis method
Protein samples for two-dimensional gel electrophoresis(2DE) were extracted as follows: Cells were lysed in lysisbuffer (8M urea, 2% (w/v) 3[(3-cholamidopropyl)dimethy-lammonio]-propanesulfonic acid (CHAPS), 0.5% (v/v)ampholytes, 20 mM dithiothreitol (DTT), and 100 lMphenylmethylsulfonyl fluoride). The lysate was then centri-fuged at 3000 rpm for 1 min. The supernatant (cell extract)was collected and protein levels determined by Bradford’sassay.22 The cell extract was then prepared in samplebuffer [8M urea, 2% (w/v) CHAPS, 0.5% (v/v) Ampho-lytes, 20 mM DTT, and a few grains of bromophenol blue]to give a loading concentration of 50 lg protein.
The first-dimensional isoelectric focusing (IEF) was per-formed on an IPGphor unit (Amersham Biosciences) usingprecast 18-cm pH 3–10 NL ImmobilineTM gel strips. Gelstrips were rehydrated at room temperature overnight. IEFwas performed in a stepwise voltage gradient procedure at208C to achieve a total of 28 kVh. After IEF separation,the gel strips were incubated with equilibration buffer I[50 mM Tris-Cl, pH 8.8, 6M urea, 30% (v/v) glycerol, 2%(w/v) sodium dodecylsulphate (SDS), bromophenol blue,and 1% (w/v) DTT] for 15 min followed by incubation inthe same buffer with DTT replaced by 2.5% iodoacetamide(w/v) for an additional 15 min. The second dimensionseparation was performed on 1.0-mm-thick 10% polyacryl-amide gels at 17 W per gel for 4 h using Ettan Daltsix Elec-trophoresis Unit (Amersham Biosciences). The gels werestained with colloidal Coomassie Blue. The stained gelswere scanned by a Fujifilm LAS 3000 Image Reader andanalyzed using the evaluation version of ProgenesisSameSpots v2.0.2644.18003, Progenesis PG240 version 2006(Nonlinear dynamics Ltd., UK).
Protein identification
The pattern of proteins was compared between control(untreated) cells and those cells treated with 0.5 lM Cr(VI). Each 2DE gel array was compared with the control2DE array. Spots which were markedly upregulated/downregulated by Cr treatment were picked using a man-ual one touch spot picker into Eppendorf tubes and storedat 2808C until digestion. Trypsin digestion and mass spec-troscopy of digested spots were performed by Dr. RichardBurchmore at the Sir Henry Wellcome Functional GenomicFacility (Glasgow University, UK) using Tandem massspectroscopy (MS/MS).
Secondary validation
Secondary validation of protein spots of interest wascarried out by SDS-PAGE followed by Western blotting.Goat polyclonal antibody for CapZb (1:500 dilution) waspurchased from Abcam Ltd. (UK). Homogenates fromboth Cr-treated and untreated osteoblasts and monocytes
616 RAGHUNATHAN, GRANT, AND ELLIS
Journal of Biomedical Materials Research Part A
were prepared in Laemmli buffer and 10 lg of proteinwere separated on 10% (w/v) SDS-polyacrylamide gels.The optical densities of the protein bands were then quan-tified using NIH Image J 1.37 v.
Detection of cytoskeletal damage in osteoblastsfollowing exposure to Cr (VI)
Detection of cytoskeletal damage was carried out as pre-viously described by Gunaratnam and Grant.23 FFC cells,grown on coverslips, were exposed to 0, 0.05, 0.1, and 0.5lM Cr (VI) for a week. At the end of the week, mediumwas removed and the cells were washed with phosphatebuffered saline (PBS) at 378C. The cells were fixed with 4%formalin in PBS for 20 min at room temperature. Followingfixing, cells were washed thrice with PBS (5 min eachwash) and incubated with FITC-Phalloidin (1:500 dilution)for 1 h in the dark at room temperature in a humid atmos-phere. Cells were then washed as described earlier andprepared for observation under a Leica Confocal Laser
Scanning Microscope (CLSM) at 488 nm excitation wave-length using 403 oil lens.
RESULTS
Chronic toxicity of chromium to cell lines
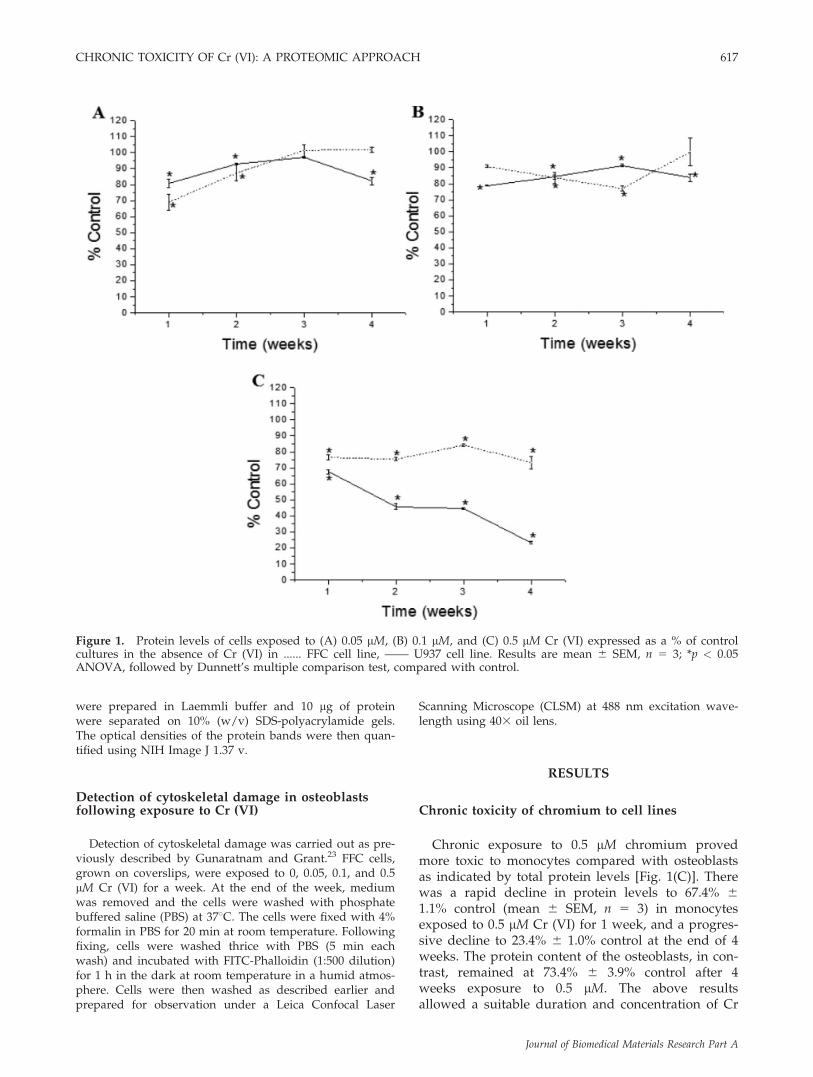
Chronic exposure to 0.5 lM chromium provedmore toxic to monocytes compared with osteoblastsas indicated by total protein levels [Fig. 1(C)]. Therewas a rapid decline in protein levels to 67.4% 61.1% control (mean 6 SEM, n 5 3) in monocytesexposed to 0.5 lM Cr (VI) for 1 week, and a progres-sive decline to 23.4% 6 1.0% control at the end of 4weeks. The protein content of the osteoblasts, in con-trast, remained at 73.4% 6 3.9% control after 4weeks exposure to 0.5 lM. The above resultsallowed a suitable duration and concentration of Cr
Figure 1. Protein levels of cells exposed to (A) 0.05 lM, (B) 0.1 lM, and (C) 0.5 lM Cr (VI) expressed as a % of controlcultures in the absence of Cr (VI) in ...... FFC cell line, —— U937 cell line. Results are mean 6 SEM, n 5 3; *p < 0.05ANOVA, followed by Dunnett’s multiple comparison test, compared with control.
CHRONIC TOXICITY OF Cr (VI): A PROTEOMIC APPROACH 617
Journal of Biomedical Materials Research Part A

(VI) for both cell types to allow changes in theexpression of individual proteins to be investigated.As 0.5 lM Cr (VI) was observed to cause the mostapparent toxic effects in monocytes, we chose to ex-pose both FFC cells and U937 cells continuously to0.5 lM Cr (VI) for 3 weeks.
Protein expression profiles followingchromium exposure
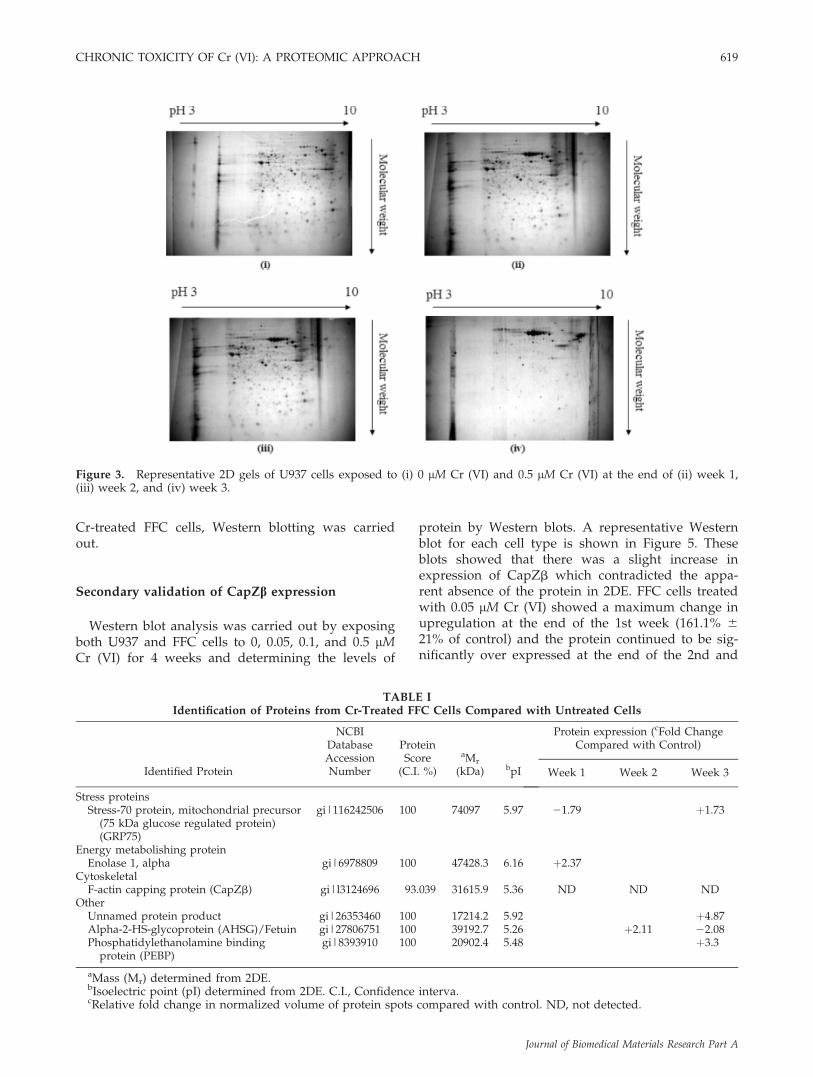
To understand the changes in protein expressionin both cell types (osteoblasts and monocytes) fol-lowing exposure to 0.5 lM Cr (VI), two-dimensionalelectrophoresis was used to perform proteomicprofiling in extracts from Cr (VI)-treated cells andcontrols. Figures 2 and 3 show representative two-dimensional electrophoresis patterns of protein regu-lation exhibited in osteoblasts and monocytes afterexposure to 0.5 lM Cr (VI) over 3 weeks. All gelswere matched using SameSpots software and imageanalysis results were visually confirmed to minimizepotential software errors. Background subtractionwas performed to minimize the likelihood of misin-terpretation of staining streaks as false protein spotsand a 3D tool was further used to confirm the occur-rence of a spot. Differential expression of the proteinspots was determined across chromium-treated anduntreated cells over 3 weeks.
From the protein profiles, it was apparent that, thetotal number of proteins that were expressed at theend of the 3rd week in cells treated with 0.5 lM Cr(VI) was significantly lower than those in controlcells, even taking into account the variation in indi-vidual expression levels. Approximately 1000 spotswere present in untreated cells and in cells treatedwith Cr for 2 weeks. In contrast, approximately 400spots were revealed in Cr-treated cells at the end ofthe 3rd week. Protein spots that were present in thesamples after the 3 week exposure to Cr (VI), andsignificantly up/downregulated, were observed inboth cell lines and were picked and identified bymass spectrometry followed by peptide mass finger-printing. This allowed the identification of six pro-teins in FFC cells and 11 proteins in U937 cells theexpression of which was altered by 3 week exposureto 0.5 lM Cr (VI). The identified spots are listed inTables I and II and include a range of glycolyticenzymes (enolase, glyoxalase), stress proteins, andproteins associated with the cytoskeleton. Changesin protein expression profiles were time, dose, andcell-type dependent as indicated in Tables I and II.From the 2DE array of Cr-treated and untreated FFCcells, it was observed that one protein spot wasapparently expressed only in control cells. FollowingMS/MS this spot was identified as F-actin cappingprotein (CapZb). Figure 4 represents a 3D montageof this protein spot in Cr-treated and untreated FFCcells. To confirm the presence/absence of CapZb in
Figure 2. Representative 2D gels of FFC cell line exposed to (i) 0 lM Cr (VI) and 0.5 lM Cr (VI) at the end of (ii) week 1,(iii) week 2, and (iv) week 3.
618 RAGHUNATHAN, GRANT, AND ELLIS
Journal of Biomedical Materials Research Part A
Cr-treated FFC cells, Western blotting was carriedout.
Secondary validation of CapZb expression
Western blot analysis was carried out by exposingboth U937 and FFC cells to 0, 0.05, 0.1, and 0.5 lMCr (VI) for 4 weeks and determining the levels of
protein by Western blots. A representative Westernblot for each cell type is shown in Figure 5. Theseblots showed that there was a slight increase inexpression of CapZb which contradicted the appa-rent absence of the protein in 2DE. FFC cells treatedwith 0.05 lM Cr (VI) showed a maximum change inupregulation at the end of the 1st week (161.1% 621% of control) and the protein continued to be sig-nificantly over expressed at the end of the 2nd and
Figure 3. Representative 2D gels of U937 cells exposed to (i) 0 lM Cr (VI) and 0.5 lM Cr (VI) at the end of (ii) week 1,(iii) week 2, and (iv) week 3.
TABLE IIdentification of Proteins from Cr-Treated FFC Cells Compared with Untreated Cells
Identified Protein
NCBIDatabaseAccessionNumber
ProteinScore
(C.I. %)
aMr
(kDa) bpI
Protein expression (cFold ChangeCompared with Control)
Week 1 Week 2 Week 3
Stress proteinsStress-70 protein, mitochondrial precursor
(75 kDa glucose regulated protein)(GRP75)
gi|116242506 100 74097 5.97 21.79 þ1.73
Energy metabolishing proteinEnolase 1, alpha gi|6978809 100 47428.3 6.16 þ2.37
CytoskeletalF-actin capping protein (CapZb) gi|l3124696 93.039 31615.9 5.36 ND ND ND
OtherUnnamed protein product gi|26353460 100 17214.2 5.92 þ4.87Alpha-2-HS-glycoprotein (AHSG)/Fetuin gi|27806751 100 39192.7 5.26 þ2.11 22.08Phosphatidylethanolamine binding
protein (PEBP)gi|8393910 100 20902.4 5.48 þ3.3
aMass (Mr) determined from 2DE.bIsoelectric point (pI) determined from 2DE. C.I., Confidence interva.cRelative fold change in normalized volume of protein spots compared with control. ND, not detected.
CHRONIC TOXICITY OF Cr (VI): A PROTEOMIC APPROACH 619
Journal of Biomedical Materials Research Part A
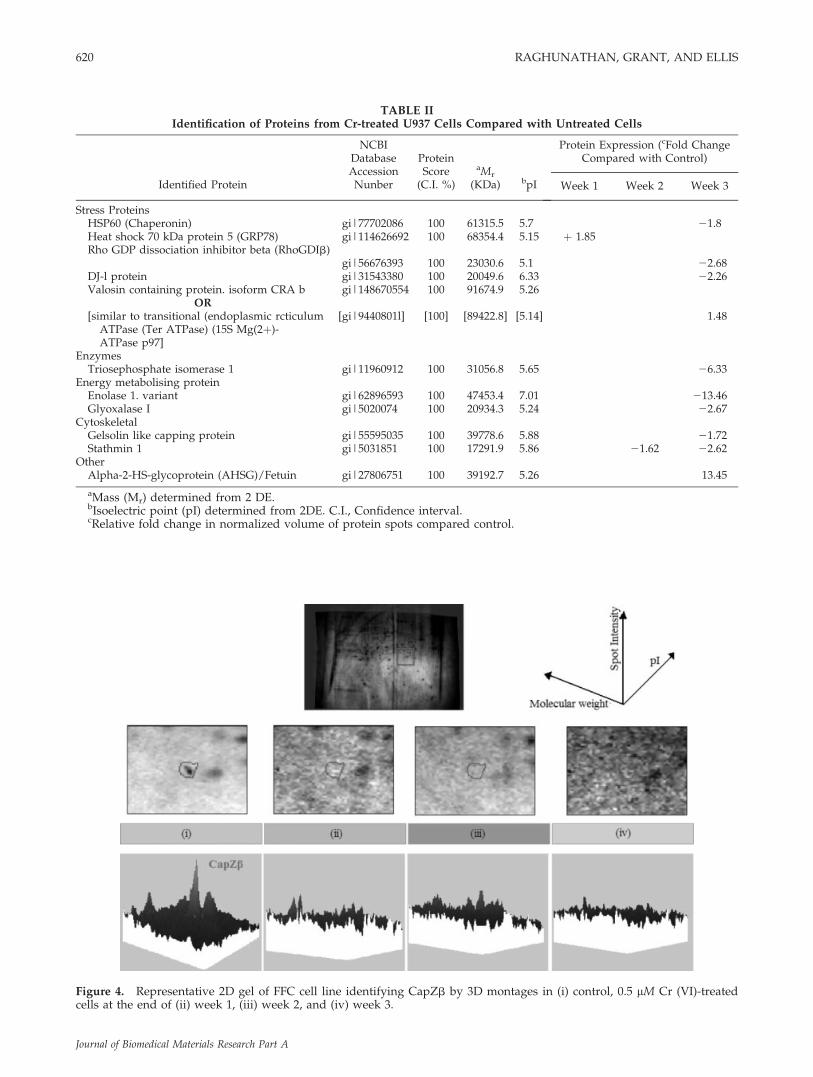
TABLE IIIdentification of Proteins from Cr-treated U937 Cells Compared with Untreated Cells
Identified Protein
NCBIDatabaseAccessionNunber
ProteinScore(C.I. %)
aMr
(KDa) bpI
Protein Expression (cFold ChangeCompared with Control)
Week 1 Week 2 Week 3
Stress ProteinsHSP60 (Chaperonin) gi|77702086 100 61315.5 5.7 21.8Heat shock 70 kDa protein 5 (GRP78) gi|114626692 100 68354.4 5.15 þ 1.85Rho GDP dissociation inhibitor beta (RhoGDIb)
gi|56676393 100 23030.6 5.1 22.68DJ-l protein gi|31543380 100 20049.6 6.33 22.26Valosin containing protein. isoform CRA b gi|148670554 100 91674.9 5.26
OR[similar to transitional (endoplasmic rcticulum
ATPase (Ter ATPase) (15S Mg(2þ)-ATPase p97]
[gi|9440801l] [100] [89422.8] [5.14] 1.48
EnzymesTriosephosphate isomerase 1 gi|11960912 100 31056.8 5.65 26.33
Energy metabolising proteinEnolase 1. variant gi|62896593 100 47453.4 7.01 213.46Glyoxalase I gi|5020074 100 20934.3 5.24 22.67
CytoskeletalGelsolin like capping protein gi|55595035 100 39778.6 5.88 21.72Stathmin 1 gi|5031851 100 17291.9 5.86 21.62 22.62
OtherAlpha-2-HS-glycoprotein (AHSG)/Fetuin gi|27806751 100 39192.7 5.26 13.45
aMass (Mr) determined from 2 DE.bIsoelectric point (pI) determined from 2DE. C.I., Confidence interval.cRelative fold change in normalized volume of protein spots compared control.
Figure 4. Representative 2D gel of FFC cell line identifying CapZb by 3D montages in (i) control, 0.5 lM Cr (VI)-treatedcells at the end of (ii) week 1, (iii) week 2, and (iv) week 3.
620 RAGHUNATHAN, GRANT, AND ELLIS
Journal of Biomedical Materials Research Part A

3rd weeks before the expression was inhibited at theend of the 4th week (83.2% 6 6.1% of control). Theprotein in U937 cells exposed to 0.01 and 0.5 lM Cr(VI) was slightly upregulated (121.6% 6 4.5% and121.9% 6 5.1% of control, respectively) at the end ofweek 1, and its expression declined to 21.1% 6 1.1%of control in monocytes exposed to the highest con-centration of Cr (VI) at the end of the 4th week.
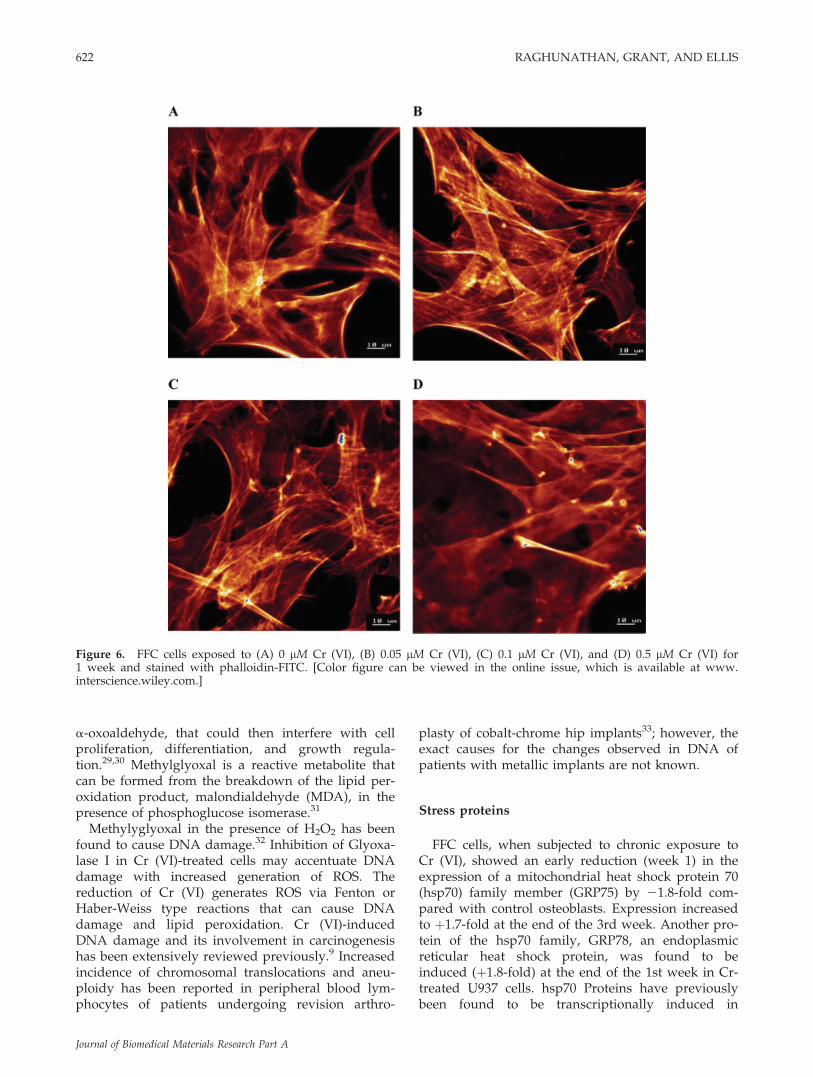
The effect of the differential regulation of CapZbwas also investigated by FITC-Phalloidin staining ofactin fibers (Fig. 6) of Cr-treated and untreatedosteoblasts after exposure for a week. The frame-work of F-actin in the untreated and Cr-treatedosteoblasts was similar. There were no detectablechanges observed in the distribution of F-actin de-
spite the apparent differential expression of the F-actin capping protein in Cr-treated cells.
DISCUSSION
Biological relevance of protein changes
To understand the responses to chronic exposureto Cr (VI) at concentrations that may be found clini-cally in patients with metallic implants, osteoblastsand monocytes were continuously exposed to 0,0.05, 0.1, and 0.5 lM Cr (VI) for 4 weeks. These con-centrations are within the range of in vivo Cr-levelmeasurements in the serum of patients with metalimplants.1 However, in vivo, in patients with metallicimplants, osteoblasts may be exposed to higher con-centrations of metal ions compared with monocytes.Following classification by function it was apparentthat a greater number of proteins involved in stressresponse were altered in their expression in mono-cytes compared with osteoblasts. This indicates thatU937 cells are more susceptible to Cr (VI)-inducedstress compared with FFC cells. U937 cells subjectedto oxidative stress by exposure to 50 lM H2O2 for 24h have previously been shown to alter the expres-sion of many proteins involved in energy metabo-lism, as well as proteins involved in RNA process-ing, protein folding, cell signaling, redox regulation,channel components, and cell structure.24 However,the effect of chronic of exposure to H2O2 is notknown.
Glycolytic proteins
It is apparent that enolase 1, a glycolytic enzyme,was over expressed (þ2-fold) in osteoblasts at theend of the first week whereas its expression was sig-nificantly lower (213-fold) in monocytes after 3weeks. a-Enolase, identified as a heat shock pro-tein,25 was found to be inactivated by glutathionyla-tion in T-lymphocytes exposed to 1 mM H2O2 for 5min.26 Phosphorylation of enolase and tyrosine ki-nases were previously observed in lymphocytestreated with 1 lM Cr (VI).16
Glyoxalase-I, a glycolytic enzyme that detoxifiesmethylglyoxal using GSH as a cofactor, is anotherenzyme associated with glycolysis that was found tobe downregulated in Cr-treated U937 cells at the endof the 3 weeks (22.3-fold). Mitsumoto et al.27 identi-fied glyoxalase I as a nitric oxide responsive proteinin immortalized human endothelial cells and itsphosphorylation was identified as a key step in tu-mor necrosis factor (TNF)-induced cell death.28
Reduction in expression of this enzyme could resultin the intracellular accumulation of methyglyoxal, an
Figure 5. CapZb expression of (A) FFC cells, (B) U936cells exposed to 0.05, 0.1, and 0.5 lM Cr (VI) expressed asa % of control cultures in the absence of Cr (VI). Week 1
Week 2 Week 3 Week 4 Results, corrected for pro-tein, are mean 6 SEM, n 5 3; *p < 0.05 ANOVA, followedby Dunnett’s multiple comparison test, compared withcontrol. A representative western blot of CapZb expressionin (A) FFC cell line exposed to chromium at the end of the1st week and (B) U937 cell line exposed to chromium atthe end of the 4th week is shown. Lane 1: cells exposed to0 lM Cr (VI); Lane 2: cells exposed to 0.05 lM Cr (VI);Lane 3: cells exposed to 0.1 lM Cr (VI); and Lane 4: cellsexposed to 0.5 lM Cr (VI). Control FFC cell optical den-sities are 0.623 6 0.045 (n 5 3; mean 6 SEM) and controlU937 cell optical densities are 0.707 6 0.035 (n 5 3; mean6 SEM).
CHRONIC TOXICITY OF Cr (VI): A PROTEOMIC APPROACH 621
Journal of Biomedical Materials Research Part A
a-oxoaldehyde, that could then interfere with cellproliferation, differentiation, and growth regula-tion.29,30 Methylglyoxal is a reactive metabolite thatcan be formed from the breakdown of the lipid per-oxidation product, malondialdehyde (MDA), in thepresence of phosphoglucose isomerase.31
Methylyglyoxal in the presence of H2O2 has beenfound to cause DNA damage.32 Inhibition of Glyoxa-lase I in Cr (VI)-treated cells may accentuate DNAdamage with increased generation of ROS. Thereduction of Cr (VI) generates ROS via Fenton orHaber-Weiss type reactions that can cause DNAdamage and lipid peroxidation. Cr (VI)-inducedDNA damage and its involvement in carcinogenesishas been extensively reviewed previously.9 Increasedincidence of chromosomal translocations and aneu-ploidy has been reported in peripheral blood lym-phocytes of patients undergoing revision arthro-
plasty of cobalt-chrome hip implants33; however, theexact causes for the changes observed in DNA ofpatients with metallic implants are not known.
Stress proteins
FFC cells, when subjected to chronic exposure toCr (VI), showed an early reduction (week 1) in theexpression of a mitochondrial heat shock protein 70(hsp70) family member (GRP75) by 21.8-fold com-pared with control osteoblasts. Expression increasedto þ1.7-fold at the end of the 3rd week. Another pro-tein of the hsp70 family, GRP78, an endoplasmicreticular heat shock protein, was found to beinduced (þ1.8-fold) at the end of the 1st week in Cr-treated U937 cells. hsp70 Proteins have previouslybeen found to be transcriptionally induced in
Figure 6. FFC cells exposed to (A) 0 lM Cr (VI), (B) 0.05 lM Cr (VI), (C) 0.1 lM Cr (VI), and (D) 0.5 lM Cr (VI) for1 week and stained with phalloidin-FITC. [Color figure can be viewed in the online issue, which is available at www.interscience.wiley.com.]
622 RAGHUNATHAN, GRANT, AND ELLIS
Journal of Biomedical Materials Research Part A

hypoxic and ischemic C2 myogenic cells,34 toenhance cell survival, and involved in the protectionof HeLa cells from stress-induced apoptosis by inhi-bition of the JNK pathway.35 In the present study,chaperonin (HSP60) was significantly downregulated(21.8-fold) at the end of the 3rd week. Inhibition ofthis stress protein may have an adverse effect onprotein folding/misfolding. Yu et al.36 have relatedthe induction of HSPs to increased production ofsuperoxide anion by heavy metals, and loss of via-bility to increased intracellular H2O2 level.
Another protein identified by 2DE as altered inCr-treated cells was Dj-1, a protein found to upregu-late GSH synthesis in oxidative stress in dopaminer-gic N27 cells.37 We have previously reported theover expression of the rate limiting (GSH) synthesiz-ing enzyme, GCL, in Cr (VI)-treated FFC and U937cells, and also observed a decline in GSH levels in0.5 lM Cr (VI)-treated U937 cells after 4 weeks.20
This, taken concurrently with the present findingwhere Dj-1 protein was downregulated in its expres-sion in U937 cells after 3 weeks, suggests that Dj-1may play a role in inhibition of GSH synthesis.However, the role of Dj-1 protein in regulating GCLexpression and activity in the synthesis of GSH isnot known at present.
Cytoskeletal-associated proteins
Stathmin, a cytoplasmic protein implicated in theregulation of the microtubule in cells,38 is progres-sively suppressed in Cr-treated U937 cells withincreasing time (21.6-fold at the end of 2nd weekand 22.6-fold at the end of the 3rd week). Stathminphosphorylation under extracellular stimulus39 wasassociated with intracellular signaling pathways40
and was found to be an important substrate formitogen-activated protein kinase (MAP Kinase)under heat shock and chemical stress.41
From the Western blot analysis, the presence of F-actin capping protein (CapZb) in Cr-treated cellswas confirmed and an initial over expression of theprotein was observed in Cr-treated FFC cells beforea progressive decline occurred. The F-actin cappingprotein (CapZb) did not appear to be present in thesame location of the 2DE gel in Cr-treated osteo-blasts as in the untreated cells (Fig. 4). This suggeststhat the reason for the absence of the spot in treatedcells may be due to a post-translational modification,rather than a change in protein expression levels.The rapid decline in the level of this protein asobserved in the 4th week in 0.5 lM Cr (VI)-treatedmonocytes could be attributed to a differentialincrease in protein degradation in the dying cells.Although we observed altered expression of CapZbin both U937 and FFC cells, we did not observe any
damage to F-actin in FFC cells exposed to 0.05, 0.1,and 0.5 lM Cr (VI) after a week. This is in contradic-tion to the result reported by Gunaratnam andGrant23 where damage to F-actin was observed afteracute exposure (24 h) of primary rat hepatocytes to 1lM Cr (VI). This difference may be due to the differ-ence in duration of exposure where chronic treat-ment may allow for an adaptive response followedby restoration of cytoskeletal structure. A differencein the response due to a difference in cell type canalso not be eliminated.
Other proteins identified by 2DE to be altered inCr-treated cells were a-2-HS-glycoprotein (a negativeacute-phase reactant), Rho GDP dissociation inhibi-tor b (a key protein that plays an important role incellular signaling; RhoGDIb), triosephosphate isom-erase 1 (glycolytic protein), phosphatidylethanol-amine binding protein (a G-protein dependent sig-naling protein, PEBP), Gelsolin-like capping protein(cytoskeletal protein), and Valosin containing protein(a key protein tyrosine kinase substrate).
CONCLUSIONS
The results of this study demonstrate that complexchanges are involved in both cell types studied afterchronic exposure to Cr (VI). Osteoblasts are continu-ously subjected to metal ions released from metallicimplants in patients and hence it is essential tounderstand the mechanisms that may be involved inaltering the regulation of bone formation andabsorption in vivo. Monocytes are essential to main-tain the balance in the immune system. Decreasednumbers of lymphocytes were observed in patientswith loosened prostheses who have elevated levelsof both cobalt and chromium in their blood42; how-ever, the mechanisms leading to their decreasednumber in circulation are not known. The resultsshown in the present article, illustrate for the firsttime, the complexities involved in the toxicity of Cr(VI) using a proteomic approach. This informationwill be useful for further analysis of cellularresponses in patients with metallic implants.
The authors would like to thank Dr. Richard Burchmore(Glasgow University) for the identification of protein spotsby mass spectrometry, and Mrs. Elizabeth Goldie for assis-tance and advice with the confocal microscopy. V.K.R. isthe recipient of an Overseas Research Studentship and apostgraduate award from the University of Strathclyde.
References
1. Sargeant A, Goswami T. Hip implants—Paper VI—Ion con-centrations. Mater Design 2007;28:155–171.
CHRONIC TOXICITY OF Cr (VI): A PROTEOMIC APPROACH 623
Journal of Biomedical Materials Research Part A

2. Merritt K, Brown SA. Release of hexavalent chromium fromcorrosion of stainless steel and cobalt-chromium alloys. JBiomed Mater Res 1995;29:627–633.
3. Bagchi D, Stohs SJ, Downs BW, Bagchi M, Preuss HG. Cyto-toxicity and oxidative mechanisms of different forms of chro-mium. Toxicology 2002;180:5–22.
4. Lalaouni A, Henderson C, Kupper C, Grant MH. The interac-tion of chromium (VI) with macrophages: Depletion of gluta-thione and inhibition of glutathione reductase. Toxicology2007;236:76–81.
5. Vandana S, Ram S, Ilavazhagan M, Kumar GD, Banerjee PK.Comparative cytoprotective activity of vitamin C. E and b-carotene against chromium induced oxidative stress in mu-rine macrophages. Biomed Pharmacother 2006;60:71–76.
6. Seiber HP, Rieker CB, Kottig P. Analysis of 118 second gener-ation metal on metal retrieved hip implants. J Bone JointSurg Br 1998;80:46–50.
7. Stea S, Visentin M, Granchi D, Cenni E, Ciapetti G. SudaneseA, Toni A. Apoptosis in peri-implant tissue. Biomaterials2000;21:1393–1398.
8. Shi X, Chiu A, Chen CT, Halliwell B, Castranova V, Vallya-than V. Reduction of chromium (VI) and its relationship tocarcinogenesis. J Toxicol Environ Health B Crit Rev1999;2:87–104.
9. Leonard SS, Bower JJ, Shi X. Metal-induced toxicity, carcino-genesis, mechanisms and cellular responses. Mol Cell Bio-chem 2004;255:3–10.
10. Asatiani N, Sapojnikova N, Abuladze M, Kartvelishvili T,Kulikova N, Kiziria E, Namchevadze E, Holman H. Effects ofCr (VI) long-term and low-dose action on mammalian antiox-idant enzymes (an in vitro study). J Inorg Biochem 2004;98:490–496.
11. O’Hara KA, Nemec AA, Alam J, Klei LR, Mossman BTBarchowsky A. Chromium (VI) inhibits heme oxygenase-1expression in vivo and in arsenic exposed human airway epi-thelial cells. J Cell Physiol 2006;209:113–121.
12. Rudolf E, Cervinka M, Cerman J, Schroterova L. Hexavalentchromium disrupts the actin cytoskeleton and induces mito-chondria-dependent apoptosis in human dermal fibroblasts.Toxciol In Vitro 2005;19:713–723.
13. Wang S, Leonard SS, Ye J, Ding M, Xhi S. The role ofhydroxyl radical as a messenger in Cr (VI)-induced p53 acti-vation. Am J Physiol Cell Physiol 2000;279:C868–C875.
14. Shi H, Hudson LG, Liu KJ. Oxidative stress and apoptosis inmetal ion induced carcinogenesis. Free Radic Biol Med2004;37:582–593.
15. Barchowsky A, O’Hara KA. Metal induced cell signalling andgene activation in lung diseases. Free Radic Biol Med2003;34:1130–1135.
16. Vasant C, Rajaram R, Ramasami T. Apoptosis of lymphocytesinduced by chromium (VI/V) is through ROS -mediated acti-vation of Src-family kinases and caspase-3. Free Radic BiolMed 2003;35:1082–1100.
17. Ning J, Grant MH. Chromium (VI)-induced cytotoxicity toosteoblast-derived cells. Toxicol In Vitro 1999;13:879–887.
18. Ning J, Henderson C, Grant MH. The cytotoxicity of chro-mium in osteoblasts: Effects on macromolecular synthesis.J Mater Sci: Mater Med 2002;13:47–52.
19. Ye J, Shi X. Gene expression profile in response to chromiuminduced cell stress in A549 cells. Mol Cell Biochem 2001;222:189–197.
20. Raghunathan VK, Tettey JNA, Ellis EM, Grant MH. Compar-ative chronic in-vitro toxicity of hexavalent chromium toosteoblasts and monocytes. J Biomed Mat Res A 2009;88:543–550.
21. Macnair R, Rodgers EH, MacDonald C, Wykman A, Goldie I,Grant MH. The cytotoxic response of primary rat and humanosteoblasts and an immortalised rat osteoblast cell line to
orthopaedic materials: Comparative sensitivity of several tox-icity indices. J Mater Sci: Mater Med 1997;8:105–111.
22. Bradford MM. A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principleof protein-dye binding. Anal Biochem 1976;72:248–254.
23. Gunaratnam M, Grant MH. Damage to F-actin and celldeath induced by chromium VI and nickel in primary mono-layer cultures of rat hepatocytes. Toxicol In Vitro 2004;18:245–253.
24. Seong JK, Kim DK, Choi KH, Oh SH, Kim KS, Lee S, Um H.Proteomic analysis of the cellular proteins induced by adapt-ive concentrations of hydrogen peroxide in human U937cells. Exp Mol Med 2002;34:374–378.
25. Iida H, Yahara I. Yeast heat-shock protein of Mr 48,000 is anisoprotein of enolase. Nature 1985;215:688–690.
26. Fratelli M, Demol H, Puype M, Casagrande S, Eberni I, Sal-mona M, Bonetto V, Mengozzi M, Duffieux F, Miclet E, BachiA, Vandekerckhove J, Gianzza E, Ghezzi P. Identification byredox proteomics of glutathionylated proteins in oxidativelystressed human T lymphocytes. Proc Natl Acad Sci USA2002;19:3505–3510.
27. Mitsumoto A, Kim KR, Oshima G, Kunimoto M, Okawa K,Iwamatsu A, Nakagawa Y. Glyoxalase I is a novel nitric-ox-ide-responsive protein. Biochem J 1999;344:837–844.
28. Van Herreweghe F, Mao J, Chaplen FWR, Grooten J, GevaertK, Vandekerckhove J, Vancompernolle K. Tumor necrosis fac-tor-induced modulation of glyoxalase I activities throughphosphorylation by PKA results in cell death and is accom-panied by the formation of a specific methylglyoxal-derivedAGE. Proc Natl Acad Sci USA 2002;99:949–954.
29. Choudhary D, Chandra D, Kale RK. Influence of methylgly-oxal on antioxidant enzymes and oxidative damage. ToxicolLett 1997;93:141–152.
30. Thornalley PJ. Advances in glyoxalase research. Glyoxalaseexpression in malignancy, anti-proliferative effects of methyl-glyoxal, glyoxalase I inhibitor diesters and S-D-lactoylgluta-thione, and methylglyoxal-modified protein binding andendocytosis by the advanced glycation endproduct receptor.Crit Rev Oncol Hematol 1995;20:99–128.
31. Agadjanyan ZS, Dugin SF, Dmitriev LF. Cumene peroxideand Fe2þ-ascorbate-induced lipid peroxidation and effectof phosphoglucose isomerase. Mol Cell Biochem 2006;289:49–53.
32. Tada A, Wakabayashi K, Totsuka Y, Sugimura T, Tsuji K,Nukaya H. 32P-Postlabeling analysis of a DNA adduct, anN2-acetyl derivative of guanine, formed in vitro by methyl-glyoxal and hydrogen peroxide in combination. Mutat Res1996;351:173–180.
33. Doherty AT, Howell RT, Ellis LA, Bisbinas I, Learmonth ID,Newson R, Case CP. Increased chromosome translocationsand aneuploidy in peripheral blood lymphocytes of patientshaving revision arthroplasty of the hip. J Bone Joint Surg Br2001;83:1075–1081.
34. Benjamin IJ, Horie S, Greenberg ML, Alpern RJ, Williams RS.Induction of stress proteins in cultured myogenic cell: Molec-ular signals for the activation of heat shock transcription fac-tor during ischemia. J Clin Invest 1992;89:1685–1689.
35. Stankiewicz AR, Lachapelle G, Foo CPZ, Radiconi SM,Mosser DD. Hsp70 inhibits heat-induced apoptosis upstreamof mitochondria by preventing bax translocation. J Biol Chem2005;280:38729–38739.
36. Yu Z, Yang X, Wang K. Metal ions induced heat shock pro-tein response by elevating superoxide anion level in HeLacells transformed by HSE-SEAP reporter gene. Toxicology2006;223:1–8.
37. Zhou W, Freed CR. DJ-1 up-regulates glutathione synthesisduring oxidative stress and inhibits A53T a-synuclein toxic-ity. J Biol Chem 2005;280:43150–43158.
624 RAGHUNATHAN, GRANT, AND ELLIS
Journal of Biomedical Materials Research Part A

38. Cassimeris L. The oncoprotein 18/stathmin family of micro-tubule destabilizers. Curr Opin Cell Biol 2002;14:18–24.
39. Sobel A, Tashijan AH. Distinct patterns of cytoplasmic proteinphophorylation related to regulation of synthesis and releaseof prolactin by GH cells. J Biol Chem 1983;258:10312–10324.
40. Gavet O, Ozon S, Manceau V, Lawler S, Curmi P, Sobel A.The stathmin phosphoprotein family: Intracellular localiza-tion and effects on the microtubule network. J Cell Sci1998;111:3333–3346.
41. Beretta L, Dubois M, Sobel A, Bensaude O. Stathmin is amajor substrate for mitogen-activated protein kinase duringheat shock and chemical stress in HeLa cells. Eur J Biochem1995;277:388–395.
42. Savarino L, Granchi D, Ciapetti G, Stea S, Donati ME,Zinghi G, Fontanesi G, Rotini R, Montanaro L. Effects ofmetal ions on white blood cells of patients with failedtotal joint arthroplasties. J Biomed Mater Res 1999;47:543–550.
CHRONIC TOXICITY OF Cr (VI): A PROTEOMIC APPROACH 625
Journal of Biomedical Materials Research Part A




















