
Functional Expression of Chemokine Receptors in Osteoblasts: Role of Regulated upon Activation,...
12
Functional Expression of -Chemokine Receptors in Osteoblasts: Role of Regulated upon Activation, Normal T Cell Expressed and Secreted (RANTES) in Osteoblasts and Regulation of Its Secretion by Osteoblasts and Osteoclasts Shozo Yano, Romuald Mentaverri, Deepthi Kanuparthi, Sanghamitra Bandyopadhyay, Alicia Rivera, Edward M. Brown, and Naibedya Chattopadhyay Division of Endocrinology, Diabetes, and Hypertension (S.Y., R.M., D.K., E.M.B., N.C.), Department of Medicine and Membrane Biology Program, Brigham and Women’s Hospital and Department of Laboratory Medicine (A.R.), Children’s Hospital, Harvard Medical School, Boston, Massachusetts 02115; and Genetics and Aging Unit (S.B.), Department of Neurology, Massachusetts General Hospital, Harvard Medical School, Charlestown, Massachusetts 02129 The expression and functions of receptors for the -chemo- kine, regulated upon activation, normal T cell expressed, and secreted (RANTES)/CCL5, were investigated in osteoblasts. Both primary osteoblasts and the MC3T3-E1 osteoblast cell line express the RANTES receptors, CCR1, 3, 4, and 5 (by RT- PCR), which encode functional receptors in osteoblasts as shown by [ 125 I]-RANTES binding followed by Scatchard anal- ysis. Expression of all four RANTES receptor mRNAs in os- teoblast is in contrast to the reports of expression of CCR1 being the only RANTES receptor expressed by osteoclasts. Exogenous RANTES elicits chemotaxis of osteoblasts and pro- motes cell survival via phosphatidylinositol 3-kinase with at- tendant phosphorylation of Akt. Osteoclastic RANTES, ob- tained from the conditioned medium of receptor activator of nuclear factor-B ligand-differentiated RAW264.7 cells also induces chemotaxis of MC3T3-E1 cells. Incubating the condi- tioned medium with an anti-RANTES neutralizing antibody attenuated this effect. RANTES secretion from osteoblast is inhibited by differentiation promoting hormones, e.g. 1,25 (OH) 2 D 3 and dexamethasone, whereas macrophage inflamma- tory protein-1 (but not macrophage inflammatory protein- 1) and elevated calcium induce it. Elevated calcium also stimulated RANTES secretion by osteoclasts. Therefore, RANTES is an osteoblast chemoattractant and a survival-pro- moting molecule whose regulation in osteoblast is varied. Fur- thermore, RANTES secreted from osteoclasts induces osteo- blast chemotaxis. Therefore, expression of RANTES and its receptors in both osteoblasts and osteoclasts could enable this chemokine to act in autocrine/paracrine modes. (Endocrinol- ogy 146: 2324 –2335, 2005) I N OSTEOBLASTS, WHICH ARE derived from the mes- enchymal stromal cell lineage, cellular proliferation or differentiation is regulated by numerous factors, including mechanical stress, mineral intake, and age, through system- ic/local hormones and/or cytokines. Previous studies have shown that stromal cells and osteoblasts also have chemo- tactic potential in response to fragments of collagen or os- teocalcin (1); growth factors such as platelet-derived growth factor, vascular endothelial growth factor, IGFs, TGF, and bone morphogenetic proteins (2– 8); cytokines such as IL-4 or -13 (9); and extracellular divalent cations, including calcium (Ca 2 ) and zinc, possibly mediated by a calcium-sensing receptor (10 –12). The capacity to undergo chemotaxis is an important attribute of osteoblasts, which contributes impor- tantly to their migration to sites of bone resorption. Tradi- tionally, substances/factors released from bone matrix dur- ing bone resorption are thought to influence osteoblast chemotaxis, with scant reference to the possible production of chemoattractants by osteoclasts per se. Recently some chemokine receptors and their ligands have been identified in osteoblasts or osteoblast-like cells, espe- cially under inflammatory conditions (13–19). Emerging ev- idence suggests pivotal roles for chemokines in various cel- lular functions and human diseases beyond the realm of immune function (20, 21), such as their roles in angiogenesis or the development of the central nervous system (22, 23). Therefore, we hypothesized that chemokines might partici- pate in normal bone turnover. Chemokines are a family of chemotactic cytokines that regulate trafficking and homing of leukocytes and other cells at various differentiation stages (24). Receptors for chemo- kines are seven membrane-spanning, G protein-coupled re- ceptors, which are classified into four major groups, CC, CXC, CXXXC, and XC, according to the number and spacing of conserved N-terminal cysteine residues. Because most of these receptors share their ligands, targeted deletion of a single chemokine or chemokine receptor has not induced obvious phenotypes under normal circumstances, with the exception of CXCR4 and CXCR2 (25). In this study, we fo- First Published Online February 17, 2005 Abbreviations: Ca 2 , Calcium; CC, -chemokine; CCR, CC chemo- kine receptor; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; K d , dissociation constant; MIP, macrophage inflammatory protein; PI3K, phosphatidylinositol 3-kinase; RANKL, receptor activator of nuclear factor- ligand; RANTES, regulated upon activation normal T cell expressed and secreted; TRAP, tartrate-resistant acid phosphatase; TUNEL, terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end labeling. Endocrinology is published monthly by The Endocrine Society (http:// www.endo-society.org), the foremost professional society serving the endocrine community. 0013-7227/05/$15.00/0 Endocrinology 146(5):2324 –2335 Printed in U.S.A. Copyright © 2005 by The Endocrine Society doi: 10.1210/en.2005-0065 2324
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Functional Expression of Chemokine Receptors in Osteoblasts: Role of Regulated upon Activation,...

Functional Expression of �-Chemokine Receptors inOsteoblasts: Role of Regulated upon Activation, Normal TCell Expressed and Secreted (RANTES) in Osteoblasts andRegulation of Its Secretion by Osteoblasts and Osteoclasts
Shozo Yano, Romuald Mentaverri, Deepthi Kanuparthi, Sanghamitra Bandyopadhyay, Alicia Rivera,Edward M. Brown, and Naibedya Chattopadhyay
Division of Endocrinology, Diabetes, and Hypertension (S.Y., R.M., D.K., E.M.B., N.C.), Department of Medicine andMembrane Biology Program, Brigham and Women’s Hospital and Department of Laboratory Medicine (A.R.), Children’sHospital, Harvard Medical School, Boston, Massachusetts 02115; and Genetics and Aging Unit (S.B.), Department ofNeurology, Massachusetts General Hospital, Harvard Medical School, Charlestown, Massachusetts 02129
The expression and functions of receptors for the �-chemo-kine, regulated upon activation, normal T cell expressed, andsecreted (RANTES)/CCL5, were investigated in osteoblasts.Both primary osteoblasts and the MC3T3-E1 osteoblast cellline express the RANTES receptors, CCR1, 3, 4, and 5 (by RT-PCR), which encode functional receptors in osteoblasts asshown by [125I]-RANTES binding followed by Scatchard anal-ysis. Expression of all four RANTES receptor mRNAs in os-teoblast is in contrast to the reports of expression of CCR1being the only RANTES receptor expressed by osteoclasts.Exogenous RANTES elicits chemotaxis of osteoblasts and pro-motes cell survival via phosphatidylinositol 3-kinase with at-tendant phosphorylation of Akt. Osteoclastic RANTES, ob-tained from the conditioned medium of receptor activator ofnuclear factor-�B ligand-differentiated RAW264.7 cells also
induces chemotaxis of MC3T3-E1 cells. Incubating the condi-tioned medium with an anti-RANTES neutralizing antibodyattenuated this effect. RANTES secretion from osteoblast isinhibited by differentiation promoting hormones, e.g. 1,25(OH)2D3 and dexamethasone, whereas macrophage inflamma-tory protein-1� (but not macrophage inflammatory protein-1�) and elevated calcium induce it. Elevated calcium alsostimulated RANTES secretion by osteoclasts. Therefore,RANTES is an osteoblast chemoattractant and a survival-pro-moting molecule whose regulation in osteoblast is varied. Fur-thermore, RANTES secreted from osteoclasts induces osteo-blast chemotaxis. Therefore, expression of RANTES and itsreceptors in both osteoblasts and osteoclasts could enable thischemokine to act in autocrine/paracrine modes. (Endocrinol-ogy 146: 2324–2335, 2005)
IN OSTEOBLASTS, WHICH ARE derived from the mes-enchymal stromal cell lineage, cellular proliferation or
differentiation is regulated by numerous factors, includingmechanical stress, mineral intake, and age, through system-ic/local hormones and/or cytokines. Previous studies haveshown that stromal cells and osteoblasts also have chemo-tactic potential in response to fragments of collagen or os-teocalcin (1); growth factors such as platelet-derived growthfactor, vascular endothelial growth factor, IGFs, TGF�, andbone morphogenetic proteins (2–8); cytokines such as IL-4 or-13 (9); and extracellular divalent cations, including calcium(Ca2�) and zinc, possibly mediated by a calcium-sensingreceptor (10–12). The capacity to undergo chemotaxis is animportant attribute of osteoblasts, which contributes impor-
tantly to their migration to sites of bone resorption. Tradi-tionally, substances/factors released from bone matrix dur-ing bone resorption are thought to influence osteoblastchemotaxis, with scant reference to the possible productionof chemoattractants by osteoclasts per se.
Recently some chemokine receptors and their ligands havebeen identified in osteoblasts or osteoblast-like cells, espe-cially under inflammatory conditions (13–19). Emerging ev-idence suggests pivotal roles for chemokines in various cel-lular functions and human diseases beyond the realm ofimmune function (20, 21), such as their roles in angiogenesisor the development of the central nervous system (22, 23).Therefore, we hypothesized that chemokines might partici-pate in normal bone turnover.
Chemokines are a family of chemotactic cytokines thatregulate trafficking and homing of leukocytes and other cellsat various differentiation stages (24). Receptors for chemo-kines are seven membrane-spanning, G protein-coupled re-ceptors, which are classified into four major groups, CC,CXC, CXXXC, and XC, according to the number and spacingof conserved N-terminal cysteine residues. Because most ofthese receptors share their ligands, targeted deletion of asingle chemokine or chemokine receptor has not inducedobvious phenotypes under normal circumstances, with theexception of CXCR4 and CXCR2 (25). In this study, we fo-
First Published Online February 17, 2005Abbreviations: Ca2�, Calcium; CC, �-chemokine; CCR, CC chemo-
kine receptor; GAPDH, glyceraldehyde-3-phosphate dehydrogenase;Kd, dissociation constant; MIP, macrophage inflammatory protein; PI3K,phosphatidylinositol 3-kinase; RANKL, receptor activator of nuclearfactor-�� ligand; RANTES, regulated upon activation normal T cellexpressed and secreted; TRAP, tartrate-resistant acid phosphatase;TUNEL, terminal deoxynucleotidyl transferase-mediated deoxyuridinetriphosphate nick end labeling.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/05/$15.00/0 Endocrinology 146(5):2324–2335Printed in U.S.A. Copyright © 2005 by The Endocrine Society
doi: 10.1210/en.2005-0065
2324

cused on the functions of regulated upon activation, normalT cell expressed and secreted (RANTES) among the CC che-mokine (also called �-chemokine) receptors (CCR), such asthose for monocyte chemoattractant proteins and macro-phage inflammatory proteins (MIP)-1� and -�.
RANTES (also called CCL5) is a small protein of 68 aminoacids and is known as a proinflammatory chemokine. Al-though RANTES was originally identified as a T cell-specificgene, its expression has been detected in a variety of tissuesor cell lines, such as spleen, thymic cortex, tonsil, kidney,Wilm’s tumor, the rhabdomyosarcoma cell line RD, and theosteosarcoma cell line MG63 (26–28). RANTES is a chemoat-tractant for monocytes, memory T lymphocytes, and eosin-ophils but not for B cells or killer T cells (28, 29). Previouswork in bone suggests that RANTES is involved in the patho-logical progression of rheumatoid arthritis, osteoarthritis,osteomyelitis, and posttraumatic responses (15, 16). RANTES,along with other �-chemokines, has been shown to induceosteoclast chemotaxis (30). Here for the first time, we dem-onstrate that RANTES receptors are expressed in primarycalvarial osteoblasts and MC3T3-E1 cells, in which RANTESpromotes cell migration and survival. We also show that cellmigration can be induced by the RANTES secreted fromosteoclasts in a paracrine mode of action, although the com-ponents for an autocrine mode of action, e.g. expression ofboth the receptors and their ligands on the same cell, arepresent in both osteoblasts and osteoclasts. Collectively,these data strongly point toward RANTES being an impor-tant molecule for communication between osteoclasts andosteoblasts and shed new light on the functions of this che-mokines in osteoblast biology.
Materials and MethodsMaterials
Cell culture media and supplements were purchased from Invitrogen(Carlsbad, CA). Recombinant mouse RANTES, receptor activator ofnuclear factor-�� ligand (RANKL), MIP-1�, MIP-1�, an anti-RANTESneutralizing antibody, and a murine RANTES ELISA kit were purchasedfrom R&D Systems (Minneapolis, MN). 1,25 (OH)2D3 was from Biomol(Plymouth Meeting, PA) and dexamethasone from Sigma-Aldrich (St.Louis, MO). The phosphatidylinositol 3-kinase (PI3K) inhibitor,LY294002, and antibodies against total and phosphorylated (Ser473) Aktwere obtained from Cell Signaling Technology (Beverly, MA). A ter-minal deoxynucleotidyl transferase-mediated deoxyuridine triphos-phate nick end labeling (TUNEL) kit was purchased from Roche Diag-nostics Corp. (Indianapolis, IN). The enhanced chemiluminescence kit,Supersignal, was purchased from Pierce (Rockford, IL). Protease inhib-itors were from Boehringer-Ingelheim (Mannheim, Germany). All otherreagents were from Sigma Chemical Co. (St. Louis, MO).
Cell culture
The mouse calvarial osteoblast cell line, MC3T3-E1, was purchasedfrom American Type Culture Collection (Manassas, VA) and cultured in�MEM containing 10% fetal bovine serum and 1% penicillin-strepto-mycin. Rat calvarial osteoblasts were cultured from 21-d fetal rats(Sprague Dawley). Twenty-five to 30 calvariae were harvested at roomtemperature. Humane handling of rats was carried out according to theguidelines of The Center for Animal Resources and Comparative Med-icine at Harvard Medical School. Primary culture of calvarial osteoblastswas performed by a previously described method (31). Briefly, cells werereleased by repeated digestion of the calvariae with 0.05% trypsin and0.1% collagenase P. After cells from the first two digestions were dis-carded, cells from the next three digestions were pooled and cultured inDMEM containing 10% heat-inactivated fetal bovine serum and 1%
penicillin-streptomycin in 5% CO2 at 37 C. After 3 d of culture, cells wereserum-starved in DMEM (0.5 mm Ca2�, 4 mm l-glutamine, 1% penicillin-streptomycin, and 0.2% BSA) for 4 h before experimentation. All ex-periments were done within 7 d after beginning the culture.
The RAW 264.7 monocyte/macrophage mouse cell line was obtainedfrom American Type Culture Collection and maintained in culture ac-cording to their instructions. Cells were cultured routinely in �MEMcontaining 10% heat-inactivated fetal calf serum. To differentiate thecells to osteoclast-like cells, RAW cells were gently scraped and seededin either 100-mm2 plates or 24-well plates at a density of 250,000/plateor 3 � 103 cells/well, respectively, and cultured for 5–6 d in DMEMsupplemented with 10% fetal calf serum and 50 ng/ml recombinantmurine RANKL. Medium was replaced on the third day, and cells werecultured for 2 more days, at which point large numbers of multinucle-ated cells were observed. To purify these multinucleated cells fromcontaminating undifferentiated cells, which adhere less tightly to thesubstratum, we followed a previously published protocol (32). The pu-rity of the differentiated osteoclasts was confirmed by comparing theirexpression of tartrate-resistant acid phosphatase (TRAP) mRNA withthat of undifferentiated RAW cells using quantitative real-time PCR.Purity was also assessed in each experiment, using TRAP staining witha leukocyte acid phosphatase kit (387-A, Sigma). Only preparations inwhich the purity was close to 90% were used for further experiments.
[125I]-RANTES binding assay
The protocol used was a minor modification from the one previouslyused by our laboratory (33). MC3T3-E1 cells and primary calvarialosteoblasts were seeded at a density of 20,000 cells/well and wereallowed to grow to 85–90% confluency. The monolayers were thenwashed with HANK-isotonic solution (H1387, Sigma-Aldrich) contain-ing (grams per liter): 8 NaCl, 0.35 NaCO2, 0.4 KCl, 0.06 KHPO4, 0.05NaH2PO4, 0.09815 MgSO4, 0.14 CaCl2, 10 Tris-3[N-morpholino]propane-sulfonic acid (pH 7.4), and 0.02 mg/ml BSA at 4 C. Cells were thenincubated with various concentrations of radiolabeled RANTES (20–110pm, or 44 nCi to 242 nCi/ml) in the presence or absence of 400 nmunlabeled RANTES for 1 h at 4 C to avoid internalization of the radio-ligand. A 20-�l aliquot was removed to measure the total radioactivityin each well. Then cells were washed three times with six volumes ofHanks’ solution at 4 C to remove unbound, radiolabeled RANTES.Labeled cells were incubated with 0.5 ml phosphate buffer solutioncontaining 5 mm EGTA buffer and 0.01% Triton X-100 for 15 min at 4C to detach the cell monolayer. Cells were scraped and transferred into4-ml plastic tubes to measure cell-bound radioactivity in a �-counter. A30-�l aliquot was also removed to determine the protein content of eachwell using the BCA method (Pierce). The specific binding was calculatedfrom the total binding in the presence or absence of unlabeled RANTES.To determine the maximal binding capacity and dissociation constant(Kd) for the binding of RANTES, a Scatchard plot was performed asdescribed previously (34, 35). Similarly, an analysis of the displacementof radiolabeled RANTES by unlabeled RANTES was performed usingdifferent concentrations of unlabeled RANTES in the presence of aconstant concentration of 90 pm [125I]-RANTES as described above. Alllinear or nonlinear curve fittings were performed as described in thefigure legends using Sigmaplot (version 5, SPSS Science, Chicago, IL),unless otherwise stated.
Measurement of RANTES in cell culture supernatants
Cells prepared in 24-well plates were starved in serum-free mediumcontaining 0.2% BSA for 4 h and then incubated with this medium in thepresence of various mediators or vehicles. Supernatants collected after18 h were used for measurement of RANTES. A Quantikine colorimetricsandwich ELISA kit (R&D Systems) was used according to the manu-facturer’s instructions. To compare RANTES secretion from osteoblastsand osteoclasts, each sample was corrected using total protein levels.
RT-PCR
Total RNA was prepared using the TRIzol reagent (Invitrogen). Oneor 2 �g total RNA were employed for the synthesis of single-strandedcDNA (cDNA synthesis kit; Invitrogen). Primer pairs used for PCR aredescribed in Table 1. Hot start PCR protocol was used as described
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2325

before (31). The PCR products were fractionated on 1.5% agarose gelsand gel purified. Bidirectional sequencing of the PCR fragments wereperformed employing the same primer pairs used for PCR by means ofan automated sequencer (AB377; Applied Biosystems, Foster City, CA)and dideoxy terminator Taq technology in the DNA Sequence Facultyof the University of Maine (Orono, ME).
Quantitative real-time PCR
SYBR green chemistry was used to perform quantitative determina-tions of the mRNAs for CCRs, RANTES, and a housekeeping gene,glyceraldehyde-3-phosphate dehydrogenase (GAPDH), following anoptimized protocol (31). The design of sense and antisense oligonucle-otide primers was based on published cDNA sequences using the PrimerExpress (version 2.0.0, Applied Biosystems). Primer sequences are listedin Table 1, and cDNA was synthesized with the Omniscript reversetranscription kit (QIAGEN, Valencia, CA). For real-time PCR, the cDNAwas amplified using an ABI PRISM 7000 sequence detection system (PEApplied Biosystems). The double-stranded DNA-specific dye SYBRGreen I was incorporated into the PCR buffer provided in the Quanti-Tech SYBR PCR kit (QIAGEN) to allow for quantitative detection of thePCR product in a 30-�l reaction volume. The temperature profile of thereaction was 95 C for 15 min, 40 cycles of denaturation at 94 C for 15 sec,and annealing and extension at 60 C for 1 min. GAPDH was used to
normalize differences in RNA isolation, RNA degradation, and theefficiencies of the reverse transcription. The size of the PCR product wasfirst verified on a 2.0% agarose gel, and then it was subjected to melting-curve analysis.
Determination of chemotactic activity using achemotaxis bioassay
Chemotactic activity was assessed using a previously described pro-tocol from our laboratory (10). Briefly, cells incubated overnight in 3 mlof 0.2% BSA-containing medium were scraped, isolated by pipetting,and resuspended with 0.5 ml of serum-free medium. Preincubation ofcells with LY294002 (10 �m) was performed for 30 min before scraping.Chemotactic activity was determined in a Blind Well Boyden chambersystem (Neuroprobe Inc., Gaithersburg, MD) with a polyvinylpyrroli-done-free polycarbonate membrane (pore size 8 �m; Neuroprobe).Serum-free medium alone or that supplemented with RANTES wasplaced in the lower chamber, and cells were loaded in the upper chamberat a concentration of 1–5 � 104 cells in 100 �l. Cell viability was assessedby Trypan blue exclusion. The chamber was incubated at 37 C for 5 h,and the membrane was removed. After the cells on the upper side werescraped off, the membrane was fixed with 96% methanol and stainedwith Giemsa. Cells that had migrated to the lower side of the membranewere counted under a microscope in each of eight high-power fields.
TABLE 1. Primer pairs used for studying various genes
Accession no. Gene name Primers
NM_009912 mCCR1 CCCAGTGAGAAGAAGGTCAAAGC (1028–1050)TGAATCAGAAGCCACAGAGAGC (1412–1390)TGCAGGTGACTGAGGTGAATTG (1188–1205)a
TGAAACAG TGCCGAAGGTA (1290–1271)a
NM_009914 mCCR3 CAATGCTGATGCTCTCAAAGACC (1258–1290)GTTCCTTCCAAGATGCTCTCAATC (1723–1700)TGGCAATTTCTGACCTGCTCT (324–344)a
AACCCAGACAGCATTTTGCAC (412–432)a
X90862 mCCR4 AAGGCAAGGACCCTGACCTATG (574–595)CCCCGAGAAAGAAGTAAATGACG (1050–1038)CGTACAACGTGGTGCTTTTCC (905–925)a
TTCTGTAGCCTGGATGGCGTA (985–1005)a
NM_009917 mCCR5 TGACACACTGCT GCCTAAACCC (940–961)AAAGACTCGTTCTCC CTGCTCCTC (1331–1308)CTGGCAAAAAGCTGAAGAGCGT (253–271)a
GCAGCATAGTGAGCCCAGAAT (352–332)a
NM_020542 rCCR1 CCGAACTGTCACTTTTGGCAT (432–452)TGCTTGCTCTGCTCACACTGA (833–813)ACCTGTTCAACCTGGCTGTCTC (215–236)a
CATGGCATCACCAAAAACCC (315–296)a
NM_053958 rCCR3 TGCAGTATTGGCAGCATTGC (492–511)TTGATACAGCAGTGGGTGTGG (902–882)TGCCTTTGTTGGTGAGAGGTT (915–935)a
TCTCGCCAGGAAGGAATGAA (1015-996)a
NM_133532 rCCR4 GCAAAACCCAGTACTCGGTCA (560–580)GTTCTGAAGAGTTGGGCGATG (962-942)ATGGATGTACCTGGTGGGCTT (342–362)a
GCTCTCAAGGAAAACACTGCG (443-423)a
NM_053960 rCCR5 TTGGCTATTGTCCATGCTGTG (499–519)TCTCTGTCACCTGCATTGCCT (967-947)AACAAATTGCAGCCCAGCTC (194–213)a
TGCTCTTCAGCTTTTTGCAGC (307-287)a
NM_013653 mRANTES TGCCCACGTCAAGGAGTATTT (172–192)a
TCTCTGGGTTGGCACACACTT (252–272)a
NM_031116 rRANTES TGCCCACGTGAAGGAGTATTT (135)a
TTCTTCTCTGGGTTGGCACAC (239)a
NM_007388 mTRAP (ACP5) CACCCTGAGATTTGTGGCTGT (105–125)a
CGGTTC TGGCGATCTCTTTG (187–205)a
NM_008084 mGAPDH CCCAGAACATCATCCCTGCATa
TCAGATCCACGACGGACACATTa
NM_017008 rGAPDH TTCAATGGCACAGTCAAGGC (190–209)a
TCACCCCATTTGATGTTAGCG (290-270)a
m, Mouse; r, rat. Underlined primers, RT-PCR.a Quantitative PCR primers.
2326 Endocrinology, May 2005, 146(5):2324–2335 Yano et al. • RANTES Actions on Osteoblasts

To determine whether RANTES secreted from osteoclasts inducedchemotaxis of osteoblasts, RAW264.7 cells were differentiated and pu-rified by the method described earlier in 100-mm2 plates. After purifi-cation, the cells were cultured in 5 ml medium for 24 h in the presenceof 50 ng/ml RANKL. The conditioned medium was then concentratedin a Speed Vac concentrator to a 1 ml volume. Conditioned mediumprepared in this way generally contained approximately 15–20 ng/mlRANTES as determined by ELISA. Two hundred microliters of theconcentrated conditioned medium was incubated with 2.5 �g/ml ofeither mouse IgG or anti-RANTES neutralizing antibody (monoclonal)at room temperature for 2 h. One hundred microliters of these mediawere then used to study chemotaxis of MC3T3-E1 cells as describedabove.
Western blot analysis
Calvarial osteoblasts were incubated with 50 ng/ml RANTES forvarious durations after 4 h of serum deprivation. Cells were rinsed withice-cold PBS and scraped on ice into lysis buffer that contained 10 mmTris-HCl (pH 7.4), 1 mm EGTA, 1 mm EDTA, 0.25 m sucrose, 1% TritonX-100, 1 mm dithiothreitol, and a cocktail of protease inhibitors (10�g/ml each of aprotinin, leupeptin, and calpain inhibitor as well as 100�g/ml Pefabloc). The cell lysates were then sonicated for 30 sec. Nucleiand cell debris were removed by centrifugation (6000 � g for 10 min),and the resultant total cellular lysate in the supernatant was used forSDS-PAGE. Immunoblot analysis was performed as described previ-ously (31). The blots were incubated overnight at 4 C with gentle shakingwith a phospho-Akt antibody at a 1:1000 dilution. The blots were ex-tensively washed with PBS containing 1% Triton X-100 and 0.15% drymilk (washing solution) at room temperature and were further incu-bated with a 1:2000 dilution of horseradish peroxidase-coupled goatantimouse IgG (Sigma) in PBS containing 1% Triton X-100 for 1 h at roomtemperature. The blots were then washed, and the signal was visualizedby chemiluminescence according to the manufacturer’s protocol (Su-persignal, Pierce). After stripping of the blots, total Akt immunoreac-tivity was determined in the same membrane. National Institutes ofHealth (NIH) image software (version 1.62) was used to quantify thesignal intensity.
TUNEL assay
The TUNEL reaction was performed to detect apoptosis using apreviously published protocol from our laboratory (36). Rat primarycalvarial osteoblasts were cultured on coverslips to no more than 50–60% confluence. Cells were then washed twice with PBS and incubatedwith serum-free medium containing vehicle, 50 ng/ml RANTES, or 50ng/ml RANTES � 10 �m LY294002, for 24 h. This step was repeated afterthe first 24-h period for another 24 h. After a total of 48 h treatment inthis manner, apoptosis was assessed using the in situ cell death detectionkit (Roche Diagnostics), following the manufacturer’s recommenda-tions. Briefly, cells were washed with PBS once, fixed with 3.8% formalinfor 5 min, washed again with PBS, and permeabilized on ice with 0.1%Triton X-100 in 0.1% sodium citrate for 2 min. Slides were rinsed twicewith PBS and then incubated for 60 min at 37 C with terminal deoxy-nucleotidyl transferase enzyme in reaction buffer. The slides were rinsedthree times with PBS and mounted with Vectashield (Vector Laborato-ries, Burlingame, CA). Samples were analyzed by fluorescence micros-copy. TUNEL-positive nuclei were detected by the bright color in con-densed or ruptured nuclei. The rate of apoptosis was calculated as theratio of the number of apoptotic osteoblasts to the total number ofosteoblasts (both apoptotic and nonapoptotic cells).
Statistics
All data are presented as the mean � se of the indicated number ofexperiments. Data were analyzed by one-way ANOVA followed byFisher’s protected least significant difference post hoc test or Student’s ttest when appropriate. P � 0.05 was taken to indicate a statisticallysignificant difference.
ResultsExpression of CC chemokine receptors in the MC3T3-E1, mouse calvarial cell line, and rat primarycalvarial osteoblasts
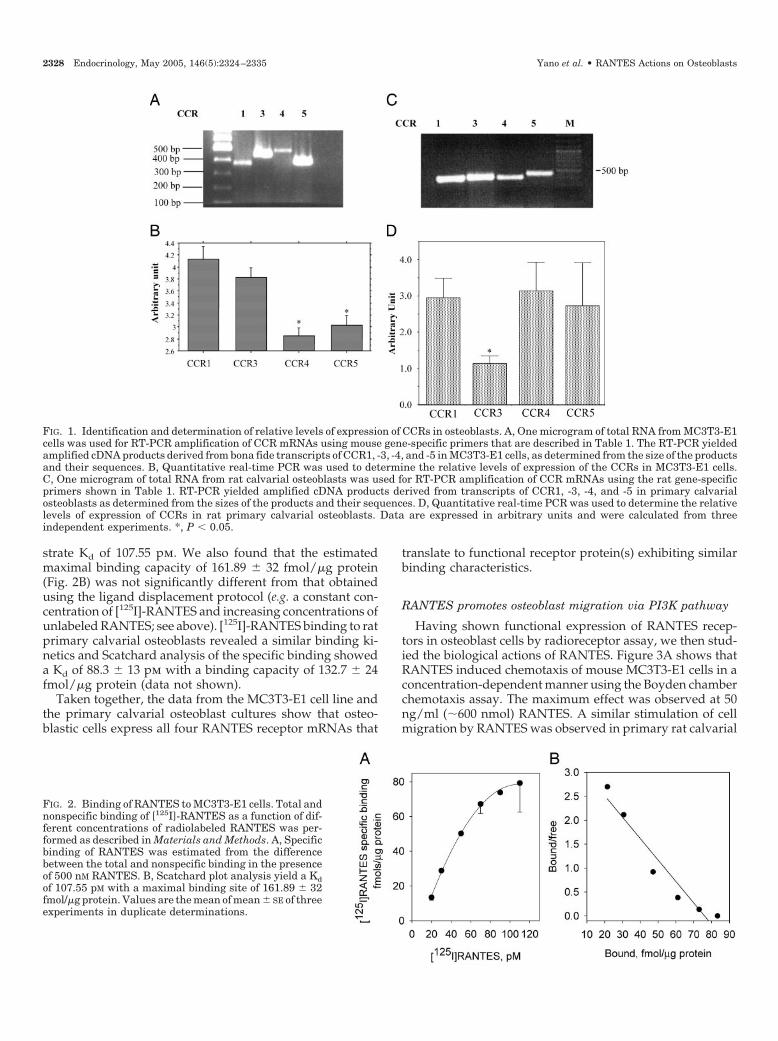
The 10 human CCRs identified to date have been classifiedinto CCR1–10. Previous studies have demonstrated thatRANTES binds to CCR1, -3, -4, and -5. Therefore, we initiallyevaluated the expression of CCR1, -3, -4, and -5 in theMC3T3-E1 cell line by RT-PCR using the primer pairs de-scribed in Table 1. Total RNA extracted from the cells wassubjected to DNase digestion to remove any contaminatingDNA before RT-PCR. Figure 1A shows amplification of theexpected mRNA fragments encompassed by the primer sets,which have the expected molecular sizes for the respectiveCCRs. No products were detected when the reverse tran-scription was omitted during synthesis of cDNA (data notshown). Bidirectional sequencing of these amplified prod-ucts using the primers employed for the respective PCRamplification revealed more than 99% homology with thecorresponding region of the cloned mRNAs. Therefore, theMC3T3-E1 mouse calvarial osteoblast cell line expressesthe mRNAs for CCR1, -3, -4, and -5. We next studied therelative abundance of the four receptors by real-time quan-titative PCR using the mouse-specific primers described inTable 1. Figure 1B shows that in MC3T3-E1 cells, the mRNAsfor CCRs 4 and -5 are expressed at modestly lower levels thanthose for CCR-1 and -3. Quantitative PCR data are expressedin arbitrary units after normalization with mouse GAPDHmRNA.
We then extended our study to primary osteoblastic cells.Rat calvarial osteoblasts were prepared following an opti-mized protocol (31), which yields more than 90% pure cul-tures as assessed by RUNX-2/cbfa-1 immunostaining. Fig-ure 1C shows that, similar to MC3T3-E1 cells, calvarialosteoblasts express mRNAs for CCR1, -3, -4, and -5. Thelevels of expression of the mRNAs for CCR1, -4, and -5 inthese cells were similar, whereas that for CCR3 mRNA was30–50% less than the others (Fig. 1D). Quantitative PCR dataare expressed in arbitrary units after normalization with ratGAPDH mRNA.
We next studied whether the expression of the mRNAs forthese four receptors known to bind RANTES is associatedwith functional binding sites using a radioreceptor assay. Totest whether RANTES specifically binds to a receptor, wemeasured [125I]-RANTES binding to cultured MC3T3-E1 os-teoblasts in the presence or absence of various concentrationsof unlabeled RANTES. The total [125I]-RANTES bound wasa saturable process in an hour at 4 C and was blocked morethan 60% with unlabeled RANTES, consistent with a specificreceptor interaction (data not shown). Analysis of the dis-placement data by unlabeled RANTES indicated an IC50 forRANTES receptor of 456 � 22 pm (Rosenthal plot) (35) witha maximal binding of 141.28 � 38 fmol/�g protein. To di-rectly measure the binding of RANTES to its receptor(s) inMC3T3-E1 cells, we measured the specific binding of [125I]-RANTES as a function of the added concentration of [125I]-RANTES at 4 C (to avoid internalization of the radioligand)(Fig. 2A). Scatchard analysis of the experimental points re-vealed the presence of a binding site with an intrinsic sub-
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2327
strate Kd of 107.55 pm. We also found that the estimatedmaximal binding capacity of 161.89 � 32 fmol/�g protein(Fig. 2B) was not significantly different from that obtainedusing the ligand displacement protocol (e.g. a constant con-centration of [125I]-RANTES and increasing concentrations ofunlabeled RANTES; see above). [125I]-RANTES binding to ratprimary calvarial osteoblasts revealed a similar binding ki-netics and Scatchard analysis of the specific binding showeda Kd of 88.3 � 13 pm with a binding capacity of 132.7 � 24fmol/�g protein (data not shown).
Taken together, the data from the MC3T3-E1 cell line andthe primary calvarial osteoblast cultures show that osteo-blastic cells express all four RANTES receptor mRNAs that
translate to functional receptor protein(s) exhibiting similarbinding characteristics.
RANTES promotes osteoblast migration via PI3K pathway
Having shown functional expression of RANTES recep-tors in osteoblast cells by radioreceptor assay, we then stud-ied the biological actions of RANTES. Figure 3A shows thatRANTES induced chemotaxis of mouse MC3T3-E1 cells in aconcentration-dependent manner using the Boyden chamberchemotaxis assay. The maximum effect was observed at 50ng/ml (�600 nmol) RANTES. A similar stimulation of cellmigration by RANTES was observed in primary rat calvarial
FIG. 2. Binding of RANTES to MC3T3-E1 cells. Total andnonspecific binding of [125I]-RANTES as a function of dif-ferent concentrations of radiolabeled RANTES was per-formed as described in Materials and Methods. A, Specificbinding of RANTES was estimated from the differencebetween the total and nonspecific binding in the presenceof 500 nM RANTES. B, Scatchard plot analysis yield a Kdof 107.55 pM with a maximal binding site of 161.89 � 32fmol/�g protein. Values are the mean of mean � SE of threeexperiments in duplicate determinations.
FIG. 1. Identification and determination of relative levels of expression of CCRs in osteoblasts. A, One microgram of total RNA from MC3T3-E1cells was used for RT-PCR amplification of CCR mRNAs using mouse gene-specific primers that are described in Table 1. The RT-PCR yieldedamplified cDNA products derived from bona fide transcripts of CCR1, -3, -4, and -5 in MC3T3-E1 cells, as determined from the size of the productsand their sequences. B, Quantitative real-time PCR was used to determine the relative levels of expression of the CCRs in MC3T3-E1 cells.C, One microgram of total RNA from rat calvarial osteoblasts was used for RT-PCR amplification of CCR mRNAs using the rat gene-specificprimers shown in Table 1. RT-PCR yielded amplified cDNA products derived from transcripts of CCR1, -3, -4, and -5 in primary calvarialosteoblasts as determined from the sizes of the products and their sequences. D, Quantitative real-time PCR was used to determine the relativelevels of expression of CCRs in rat primary calvarial osteoblasts. Data are expressed in arbitrary units and were calculated from threeindependent experiments. *, P � 0.05.
2328 Endocrinology, May 2005, 146(5):2324–2335 Yano et al. • RANTES Actions on Osteoblasts
osteoblasts (Fig. 3B). Because chemotaxis of preosteoblasts tosites of recent bone resorption is an important physiologicalfunction, our data show that RANTES is capable of inducingthis function.
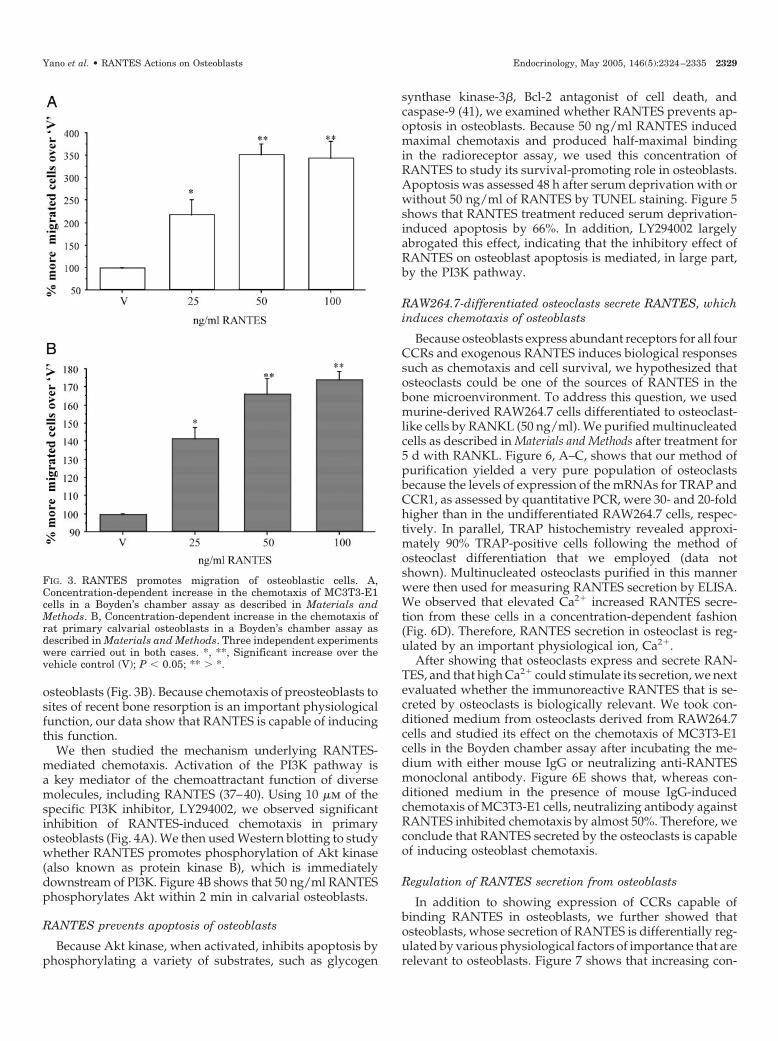
We then studied the mechanism underlying RANTES-mediated chemotaxis. Activation of the PI3K pathway isa key mediator of the chemoattractant function of diversemolecules, including RANTES (37–40). Using 10 �m of thespecific PI3K inhibitor, LY294002, we observed significantinhibition of RANTES-induced chemotaxis in primaryosteoblasts (Fig. 4A). We then used Western blotting to studywhether RANTES promotes phosphorylation of Akt kinase(also known as protein kinase B), which is immediatelydownstream of PI3K. Figure 4B shows that 50 ng/ml RANTESphosphorylates Akt within 2 min in calvarial osteoblasts.
RANTES prevents apoptosis of osteoblasts
Because Akt kinase, when activated, inhibits apoptosis byphosphorylating a variety of substrates, such as glycogen
synthase kinase-3�, Bcl-2 antagonist of cell death, andcaspase-9 (41), we examined whether RANTES prevents ap-optosis in osteoblasts. Because 50 ng/ml RANTES inducedmaximal chemotaxis and produced half-maximal bindingin the radioreceptor assay, we used this concentration ofRANTES to study its survival-promoting role in osteoblasts.Apoptosis was assessed 48 h after serum deprivation with orwithout 50 ng/ml of RANTES by TUNEL staining. Figure 5shows that RANTES treatment reduced serum deprivation-induced apoptosis by 66%. In addition, LY294002 largelyabrogated this effect, indicating that the inhibitory effect ofRANTES on osteoblast apoptosis is mediated, in large part,by the PI3K pathway.
RAW264.7-differentiated osteoclasts secrete RANTES, whichinduces chemotaxis of osteoblasts
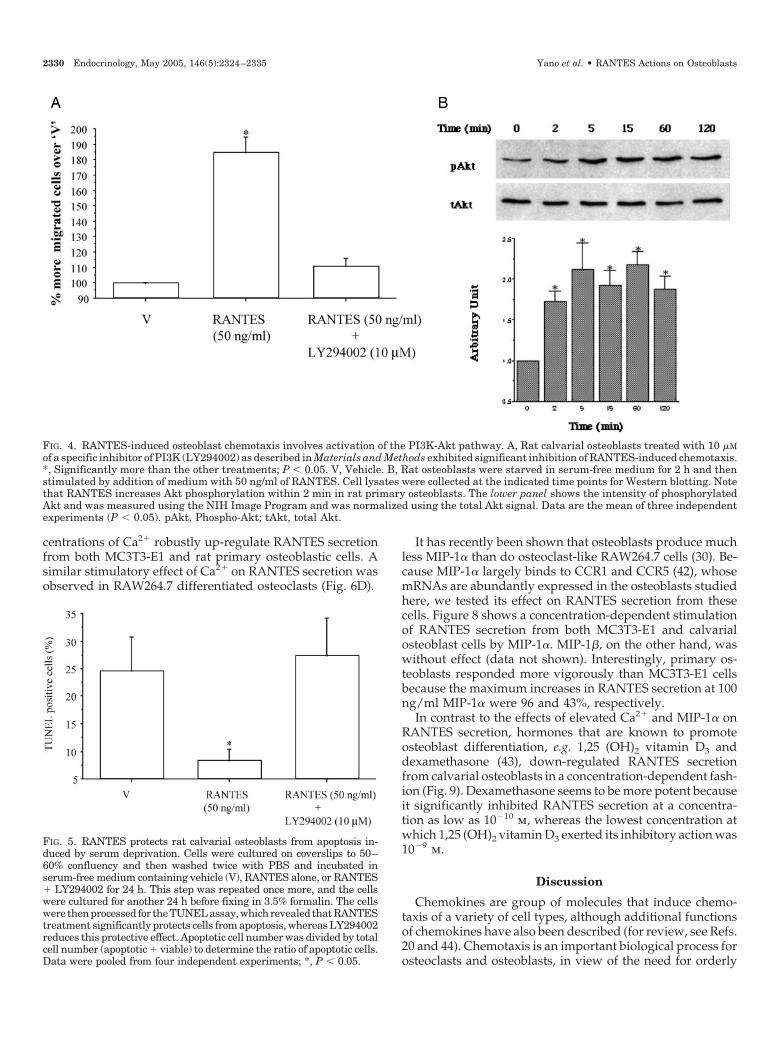
Because osteoblasts express abundant receptors for all fourCCRs and exogenous RANTES induces biological responsessuch as chemotaxis and cell survival, we hypothesized thatosteoclasts could be one of the sources of RANTES in thebone microenvironment. To address this question, we usedmurine-derived RAW264.7 cells differentiated to osteoclast-like cells by RANKL (50 ng/ml). We purified multinucleatedcells as described in Materials and Methods after treatment for5 d with RANKL. Figure 6, A–C, shows that our method ofpurification yielded a very pure population of osteoclastsbecause the levels of expression of the mRNAs for TRAP andCCR1, as assessed by quantitative PCR, were 30- and 20-foldhigher than in the undifferentiated RAW264.7 cells, respec-tively. In parallel, TRAP histochemistry revealed approxi-mately 90% TRAP-positive cells following the method ofosteoclast differentiation that we employed (data notshown). Multinucleated osteoclasts purified in this mannerwere then used for measuring RANTES secretion by ELISA.We observed that elevated Ca2� increased RANTES secre-tion from these cells in a concentration-dependent fashion(Fig. 6D). Therefore, RANTES secretion in osteoclast is reg-ulated by an important physiological ion, Ca2�.
After showing that osteoclasts express and secrete RAN-TES, and that high Ca2� could stimulate its secretion, we nextevaluated whether the immunoreactive RANTES that is se-creted by osteoclasts is biologically relevant. We took con-ditioned medium from osteoclasts derived from RAW264.7cells and studied its effect on the chemotaxis of MC3T3-E1cells in the Boyden chamber assay after incubating the me-dium with either mouse IgG or neutralizing anti-RANTESmonoclonal antibody. Figure 6E shows that, whereas con-ditioned medium in the presence of mouse IgG-inducedchemotaxis of MC3T3-E1 cells, neutralizing antibody againstRANTES inhibited chemotaxis by almost 50%. Therefore, weconclude that RANTES secreted by the osteoclasts is capableof inducing osteoblast chemotaxis.
Regulation of RANTES secretion from osteoblasts
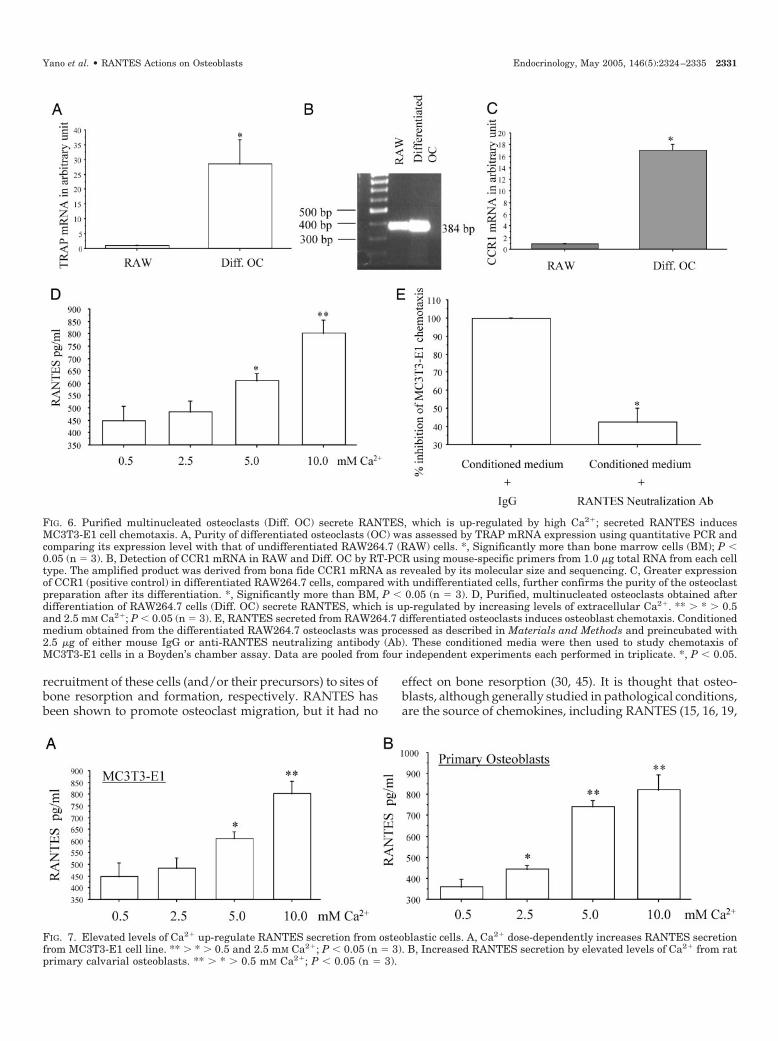
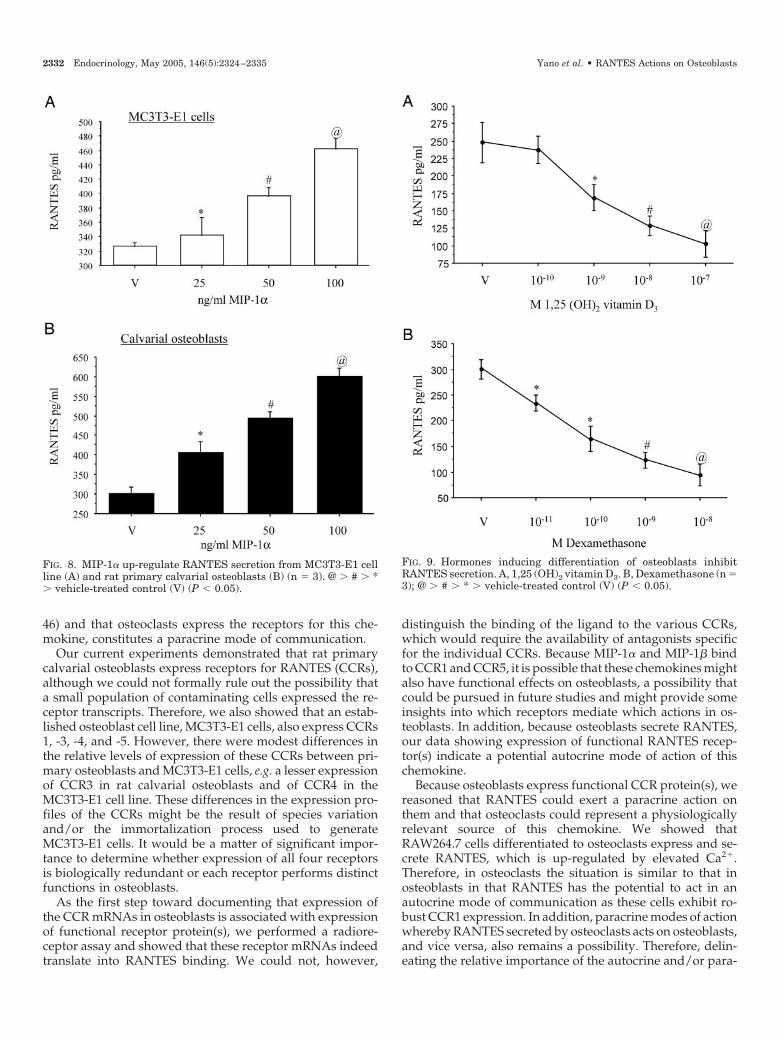
In addition to showing expression of CCRs capable ofbinding RANTES in osteoblasts, we further showed thatosteoblasts, whose secretion of RANTES is differentially reg-ulated by various physiological factors of importance that arerelevant to osteoblasts. Figure 7 shows that increasing con-
FIG. 3. RANTES promotes migration of osteoblastic cells. A,Concentration-dependent increase in the chemotaxis of MC3T3-E1cells in a Boyden’s chamber assay as described in Materials andMethods. B, Concentration-dependent increase in the chemotaxis ofrat primary calvarial osteoblasts in a Boyden’s chamber assay asdescribed in Materials and Methods. Three independent experimentswere carried out in both cases. *, **, Significant increase over thevehicle control (V); P � 0.05; ** � *.
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2329
centrations of Ca2� robustly up-regulate RANTES secretionfrom both MC3T3-E1 and rat primary osteoblastic cells. Asimilar stimulatory effect of Ca2� on RANTES secretion wasobserved in RAW264.7 differentiated osteoclasts (Fig. 6D).
It has recently been shown that osteoblasts produce muchless MIP-1� than do osteoclast-like RAW264.7 cells (30). Be-cause MIP-1� largely binds to CCR1 and CCR5 (42), whosemRNAs are abundantly expressed in the osteoblasts studiedhere, we tested its effect on RANTES secretion from thesecells. Figure 8 shows a concentration-dependent stimulationof RANTES secretion from both MC3T3-E1 and calvarialosteoblast cells by MIP-1�. MIP-1�, on the other hand, waswithout effect (data not shown). Interestingly, primary os-teoblasts responded more vigorously than MC3T3-E1 cellsbecause the maximum increases in RANTES secretion at 100ng/ml MIP-1� were 96 and 43%, respectively.
In contrast to the effects of elevated Ca2� and MIP-1� onRANTES secretion, hormones that are known to promoteosteoblast differentiation, e.g. 1,25 (OH)2 vitamin D3 anddexamethasone (43), down-regulated RANTES secretionfrom calvarial osteoblasts in a concentration-dependent fash-ion (Fig. 9). Dexamethasone seems to be more potent becauseit significantly inhibited RANTES secretion at a concentra-tion as low as 10�10 m, whereas the lowest concentration atwhich 1,25 (OH)2 vitamin D3 exerted its inhibitory action was10�9 m.
Discussion
Chemokines are group of molecules that induce chemo-taxis of a variety of cell types, although additional functionsof chemokines have also been described (for review, see Refs.20 and 44). Chemotaxis is an important biological process forosteoclasts and osteoblasts, in view of the need for orderly
FIG. 5. RANTES protects rat calvarial osteoblasts from apoptosis in-duced by serum deprivation. Cells were cultured on coverslips to 50–60% confluency and then washed twice with PBS and incubated inserum-free medium containing vehicle (V), RANTES alone, or RANTES� LY294002 for 24 h. This step was repeated once more, and the cellswere cultured for another 24 h before fixing in 3.5% formalin. The cellswere thenprocessed for theTUNELassay,whichrevealed thatRANTEStreatment significantly protects cells from apoptosis, whereas LY294002reduces this protective effect. Apoptotic cell number was divided by totalcell number (apoptotic � viable) to determine the ratio of apoptotic cells.Data were pooled from four independent experiments; *, P � 0.05.
FIG. 4. RANTES-induced osteoblast chemotaxis involves activation of the PI3K-Akt pathway. A, Rat calvarial osteoblasts treated with 10 �Mof a specific inhibitor of PI3K (LY294002) as described in Materials and Methods exhibited significant inhibition of RANTES-induced chemotaxis.*, Significantly more than the other treatments; P � 0.05. V, Vehicle. B, Rat osteoblasts were starved in serum-free medium for 2 h and thenstimulated by addition of medium with 50 ng/ml of RANTES. Cell lysates were collected at the indicated time points for Western blotting. Notethat RANTES increases Akt phosphorylation within 2 min in rat primary osteoblasts. The lower panel shows the intensity of phosphorylatedAkt and was measured using the NIH Image Program and was normalized using the total Akt signal. Data are the mean of three independentexperiments (P � 0.05). pAkt, Phospho-Akt; tAkt, total Akt.
2330 Endocrinology, May 2005, 146(5):2324–2335 Yano et al. • RANTES Actions on Osteoblasts
recruitment of these cells (and/or their precursors) to sites ofbone resorption and formation, respectively. RANTES hasbeen shown to promote osteoclast migration, but it had no
effect on bone resorption (30, 45). It is thought that osteo-blasts, although generally studied in pathological conditions,are the source of chemokines, including RANTES (15, 16, 19,
FIG. 6. Purified multinucleated osteoclasts (Diff. OC) secrete RANTES, which is up-regulated by high Ca2�; secreted RANTES inducesMC3T3-E1 cell chemotaxis. A, Purity of differentiated osteoclasts (OC) was assessed by TRAP mRNA expression using quantitative PCR andcomparing its expression level with that of undifferentiated RAW264.7 (RAW) cells. *, Significantly more than bone marrow cells (BM); P �0.05 (n 3). B, Detection of CCR1 mRNA in RAW and Diff. OC by RT-PCR using mouse-specific primers from 1.0 �g total RNA from each celltype. The amplified product was derived from bona fide CCR1 mRNA as revealed by its molecular size and sequencing. C, Greater expressionof CCR1 (positive control) in differentiated RAW264.7 cells, compared with undifferentiated cells, further confirms the purity of the osteoclastpreparation after its differentiation. *, Significantly more than BM, P � 0.05 (n 3). D, Purified, multinucleated osteoclasts obtained afterdifferentiation of RAW264.7 cells (Diff. OC) secrete RANTES, which is up-regulated by increasing levels of extracellular Ca2�. ** � * � 0.5and 2.5 mM Ca2�; P � 0.05 (n 3). E, RANTES secreted from RAW264.7 differentiated osteoclasts induces osteoblast chemotaxis. Conditionedmedium obtained from the differentiated RAW264.7 osteoclasts was processed as described in Materials and Methods and preincubated with2.5 �g of either mouse IgG or anti-RANTES neutralizing antibody (Ab). These conditioned media were then used to study chemotaxis ofMC3T3-E1 cells in a Boyden’s chamber assay. Data are pooled from four independent experiments each performed in triplicate. *, P � 0.05.
FIG. 7. Elevated levels of Ca2� up-regulate RANTES secretion from osteoblastic cells. A, Ca2� dose-dependently increases RANTES secretionfrom MC3T3-E1 cell line. ** � * � 0.5 and 2.5 mM Ca2�; P � 0.05 (n 3). B, Increased RANTES secretion by elevated levels of Ca2� from ratprimary calvarial osteoblasts. ** � * � 0.5 mM Ca2�; P � 0.05 (n 3).
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2331
46) and that osteoclasts express the receptors for this che-mokine, constitutes a paracrine mode of communication.
Our current experiments demonstrated that rat primarycalvarial osteoblasts express receptors for RANTES (CCRs),although we could not formally rule out the possibility thata small population of contaminating cells expressed the re-ceptor transcripts. Therefore, we also showed that an estab-lished osteoblast cell line, MC3T3-E1 cells, also express CCRs1, -3, -4, and -5. However, there were modest differences inthe relative levels of expression of these CCRs between pri-mary osteoblasts and MC3T3-E1 cells, e.g. a lesser expressionof CCR3 in rat calvarial osteoblasts and of CCR4 in theMC3T3-E1 cell line. These differences in the expression pro-files of the CCRs might be the result of species variationand/or the immortalization process used to generateMC3T3-E1 cells. It would be a matter of significant impor-tance to determine whether expression of all four receptorsis biologically redundant or each receptor performs distinctfunctions in osteoblasts.
As the first step toward documenting that expression ofthe CCR mRNAs in osteoblasts is associated with expressionof functional receptor protein(s), we performed a radiore-ceptor assay and showed that these receptor mRNAs indeedtranslate into RANTES binding. We could not, however,
distinguish the binding of the ligand to the various CCRs,which would require the availability of antagonists specificfor the individual CCRs. Because MIP-1� and MIP-1� bindto CCR1 and CCR5, it is possible that these chemokines mightalso have functional effects on osteoblasts, a possibility thatcould be pursued in future studies and might provide someinsights into which receptors mediate which actions in os-teoblasts. In addition, because osteoblasts secrete RANTES,our data showing expression of functional RANTES recep-tor(s) indicate a potential autocrine mode of action of thischemokine.
Because osteoblasts express functional CCR protein(s), wereasoned that RANTES could exert a paracrine action onthem and that osteoclasts could represent a physiologicallyrelevant source of this chemokine. We showed thatRAW264.7 cells differentiated to osteoclasts express and se-crete RANTES, which is up-regulated by elevated Ca2�.Therefore, in osteoclasts the situation is similar to that inosteoblasts in that RANTES has the potential to act in anautocrine mode of communication as these cells exhibit ro-bust CCR1 expression. In addition, paracrine modes of actionwhereby RANTES secreted by osteoclasts acts on osteoblasts,and vice versa, also remains a possibility. Therefore, delin-eating the relative importance of the autocrine and/or para-
FIG. 8. MIP-1� up-regulate RANTES secretion from MC3T3-E1 cellline (A) and rat primary calvarial osteoblasts (B) (n 3). @ � # � *� vehicle-treated control (V) (P � 0.05).
FIG. 9. Hormones inducing differentiation of osteoblasts inhibitRANTES secretion. A, 1,25 (OH)2 vitamin D3. B, Dexamethasone (n 3); @ � # � * � vehicle-treated control (V) (P � 0.05).
2332 Endocrinology, May 2005, 146(5):2324–2335 Yano et al. • RANTES Actions on Osteoblasts
crine actions of RANTES and other �-chemokines in osteo-blasts and osteoclasts is a much-needed subject of futureinvestigation. As a first approach to addressing this issue, weshow in this report that exogenous RANTES elicits severalbiological actions in osteoblasts.
Because RANTES is a chemokine, we first studied its effecton the chemotaxis of osteoblasts. Physiologically, preosteo-blast migration to the site of bone resorption during thereversal phase of the bone remodeling cycle is thought to beevoked by various factors released from the resorbed bonematrix (e.g. TGF� and bone morphogenetic proteins) (2, 3).However, there has been little work investigating the pos-sibility that factors released from osteoclasts per se also par-ticipate in promoting chemotaxis of preosteoblasts. Our datashow that both exogenous RANTES and conditioned me-dium derived from RAW264.7 cells differentiated to oste-oclasts elicit osteoblast chemotaxis. This action of RANTESis mediated by its cognate receptor(s), acting via the PI3Kpathway. Moreover, treating osteoblasts with RANTES re-sulted in phosphorylation of Akt, the kinase that is down-stream of PI3K, which would be expected to exert a prosur-vival effect. Indeed, we observed that RANTES preventsosteoblasts from undergoing apoptosis induced by serumstarvation. It is conceivable that, during the course of theirmigration to sites of recent bone resorption, there is a needfor protection from apoptotic cell death induced by the pleth-ora of cytokines and Ca2� released from the bone matrix.Involvement of the PI3K-Akt pathway has also been ob-served in platelet-derived growth factor-induced chemotaxisin an osteoblast cell line (4, 40). Therefore, migration of os-teoblasts in response to RANTES, along with the antiapop-
totic action of the latter, are functional evidence of the ex-pression of functional CCR(s) in osteoblasts, which could beactivated in a paracrine fashion, plausibly by RANTES re-leased by osteoclasts.
The paracrine mode of RANTES action in the bone mi-croenvironment has been hypothesized due to recent reportsincluding this report that osteoclast secretes RANTES, whichcould act on osteoblast. A report based on mRNA profilingdemonstrated that RANKL increases RANTES mRNA morethan 10-fold in both mouse and human osteoclast precursors(47). In addition, RANTES secretion was found to increase ina time-dependent fashion in RAW264.7 cells treated withRANKL, indicating that the process of differentiation couldup-regulate RANTES secretion in RAW264.7 cells (32). Weobserved expression of RANTES in addition to CCR1 mRNAin RAW264.7 differentiated osteoclasts, which suggests ex-istence of an autocrine mode of RANTES action in this cells.Osteoblast in contrast expresses all four RANTES receptorsand is a rich source of RANTES, suggesting a greater pos-sibility of autocrine mode of action. However, given theexpression of RANTES and its receptors in both cell typesalso raises the curious possibility of paracrine mode of com-munication between these two cell types. Therefore, we spec-ulate that RANTES secreted by the osteoclast could promoteosteoblast migration. We indeed have shown that RANTESsecreted from differentiated RAW264.7 cells induces chemo-taxis of MC3T3-E1 cells. Definitive evidence that the RANTESsecreted by osteoclasts was the cause of the osteoblastic che-motaxis was obtained by showing that the migration ofMC3T3-E1 cells by the conditioned medium was attenuatedby incubating it with neutralizing anti-RANTES antibody.
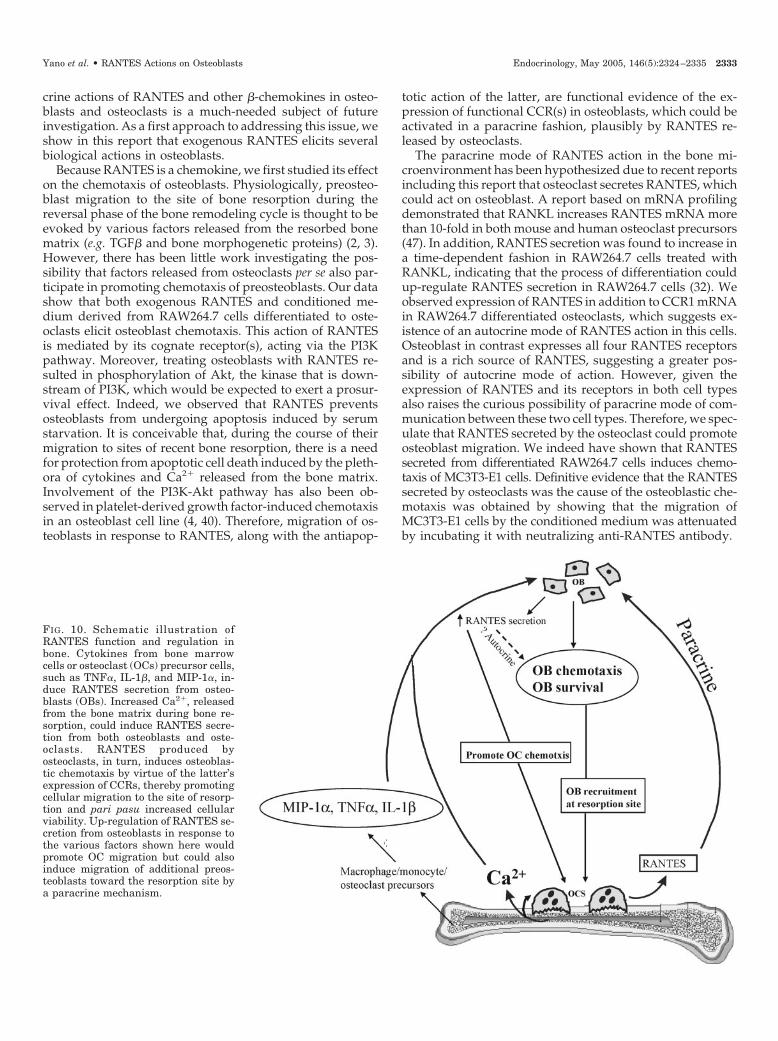
FIG. 10. Schematic illustration ofRANTES function and regulation inbone. Cytokines from bone marrowcells or osteoclast (OCs) precursor cells,such as TNF�, IL-1�, and MIP-1�, in-duce RANTES secretion from osteo-blasts (OBs). Increased Ca2�, releasedfrom the bone matrix during bone re-sorption, could induce RANTES secre-tion from both osteoblasts and oste-oclasts. RANTES produced byosteoclasts, in turn, induces osteoblas-tic chemotaxis by virtue of the latter’sexpression of CCRs, thereby promotingcellular migration to the site of resorp-tion and pari pasu increased cellularviability. Up-regulation of RANTES se-cretion from osteoblasts in response tothe various factors shown here wouldpromote OC migration but could alsoinduce migration of additional preos-teoblasts toward the resorption site bya paracrine mechanism.
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2333

RANTES secretion was up-regulated in both osteoblastsand osteoclasts by elevated extracellular Ca2�, which wouldbe encountered in the bone microenvironment during boneresorption. High Ca2� exerts a mitogenic effect on osteoblasts(31), and, therefore, it is conceivable that RANTES up-reg-ulation takes place as a function of the proliferation in thesecells. Induction of RANTES secretion by high Ca2� could bephysiologically important because the process of bone re-sorption by osteoclasts sets in motion changes in osteoblastfunction, such as chemotaxis, that culminate in bone forma-tion by differentiated osteoblasts at the site at which bonehad been resorbed. Therefore, it is possible that osteoclasts,in response to high local Ca2� generated as a result of theirresorptive activity, secrete more RANTES, which, in turn,induces osteoblast migration.
Furthermore, we observed that another �-chemokine,MIP-1�, which acts by binding to CCR1 and CCR5, inducedRANTES secretion from osteoblasts. MIP-1� secretion is highin osteoclasts and their precursor cells, compared with os-teoblasts, and we have shown expression of CCR1 and CCR5mRNAs in osteoblasts. Therefore, a concentration-depen-dent increase in RANTES by MIP-1� indicates that thesereceptors are functional in osteoblasts. Also, because RAN-TES regulates several important osteoclast functions andMIP-1� is secreted predominantly by osteoclasts, it is con-ceivable that communication between osteoclasts and osteo-blasts could be regulated by these two chemokines.
Differentiation-promoting factors, such as 1,25 (OH)2 vi-tamin D3 and dexamethasone, inhibit RANTES secretionfrom osteoblasts. The ability of dexamethasone to inhibit theup-regulation of RANTES mRNA as a result of infection withMycobacterium tuberculosis in the osteoblast-like MG63 osteo-sarcoma cell line suggested an inhibitory effect of glucocor-ticoid on RANTES expression (48). However, 10-fold higherconcentration of the hormone was required to maximallyinhibit RANTES secretion (e.g. 10�6 m vs. the 10�7 m observedin our study with normal osteoblasts). It is possible that M.tuberculosis infection up-regulates RANTES expression in os-teosarcoma cells to such a high level that a supraphysiologi-cal concentration of dexamethasone is required to inhibit it.
Although we have shown several functions of RANTES inosteoblasts that would be expected to favor bone formation,we need to understand in greater detail the importance ofthis chemokine, if any, for bone formation and resorption invivo. CCRs and their ligands are known for their redundancyof binding, and a similar situation may exist in osteoblastsbecause we observed the expression of multiple types ofCCRs. Therefore, it will be important to identify which, ifany, of these receptors participate in normal bone turnover.Gene targeting techniques, receptor-selective inhibitors, andneutralizing antibodies will be of use in addressing theseissues. Other classes of chemokines and chemokine receptorsmay also be involved in normal bone physiology. Furtherstudies are needed to clarify these issues.
Taken together, our data show for the first time that os-teoblasts respond to RANTES by virtue of their functionalexpression of CCRs. We also show that RANTES producedby osteoclasts can induce osteoblast chemotaxis and thatexogenous addition of RANTES protects these cells againstapoptosis. Osteoblasts and osteoclasts possess the capacity to
respond to both autocrine and paracrine modes of action ofRANTES owing to their expression of both RANTES and itsreceptors. As illustrated in Fig. 10, inflammatory/osteoclas-togenic cytokines, such as TNF� and IL-1� (30), elevatedCa2� (owing to increased osteoclastic activity), and MIP-1�from osteoclast precursor cells, will induce RANTES secre-tion by osteoblasts, which consequently will promote mi-gration of preosteoclasts to the site of future bone resorption(e.g. mediated by the osteoblast via RANKL). Elevated Ca2�
in the resorptive microenvironment will produce increasedRANTES production from osteoclasts, which, in turn, willinduce migration of preosteoblasts to sites of bone resorp-tion. It is also conceivable that increased RANTES secretionby the osteoblasts nearest the resorbing osteoclast as a resultof their exposure to higher concentrations of Ca2� and/orMIP-1� could then pass along the chemotactic signal(RANTES) to preosteoblasts farther away, a sort of an am-plification mechanism. This creates a possible scenario inwhich RANTES secreted by bone cells could participate bothin osteoclastogenesis and recruitment of osteoblasts and theirprotection against apoptosis during the resorptive and for-mative phases of bone turnover (49), respectively.
Acknowledgments
Received January 18, 2005. Accepted February 9, 2005.Address all correspondence and requests for reprints to: Naibedya
Chattopadhyay, Ph.D., Division of Endocrinology, Diabetes, and Hy-pertension, Brigham and Women’s Hospital and Harvard MedicalSchool, 221 Longwood Avenue, Boston, Massachusetts 02115. E-mail:[email protected].
This work was supported by National Institutes of Health GrantsAR02115 (to N.C.); HL67699 and DK069388 (to AR); and DK48330,DK52005, DK67111, and DK67155 (to E.M.B.).
References
1. Mundy GR, Poser JW 1983 Chemotactic activity of the gamma-carboxyglu-tamic acid containing protein in bone. Calcif Tissue Int 35:164–168
2. Lucas PA 1989 Chemotactic response of osteoblast-like cells to transforminggrowth factor �. Bone 10:459–463
3. Pfeilschifter J, Wolf O, Naumann A, Minne HW, Mundy GR, Ziegler R 1990Chemotactic response of osteoblastlike cells to transforming growth factor �.J Bone Miner Res 5:825–830
4. Tsukamoto T, Matsui T, Fukase M, Fujita T 1991 Platelet-derived growthfactor B chain homodimer enhances chemotaxis and DNA synthesis in normalosteoblast-like cells (MC3T3–E1). Biochem Biophys Res Commun 175:745–751
5. Panagakos FS 1993 Insulin-like growth factors-I and -II stimulate chemotaxisof osteoblasts isolated from fetal rat calvaria. Biochimie (Paris) 75:991–994
6. Godwin SL, Soltoff SP 1997 Extracellular calcium and platelet-derived growthfactor promote receptor-mediated chemotaxis in osteoblasts through differentsignaling pathways. J Biol Chem 272:11307–11312
7. Fiedler J, Roderer G, Gunther KP, Brenner RE 2002 BMP-2, BMP-4, andPDGF-BB stimulate chemotactic migration of primary human mesenchymalprogenitor cells. J Cell Biochem 87:305–312
8. Mayr-Wohlfart U, Waltenberger J, Hausser H, Kessler S, Gunther KP, DehioC, Puhl W, Brenner RE 2002 Vascular endothelial growth factor stimulateschemotactic migration of primary human osteoblasts. Bone 30:472–477
9. Lind M, Deleuran B, Yssel H, Fink-Eriksen E, Thestrup-Pedersen K 1995 IL-4and IL-13, but not IL-10, are chemotactic factors for human osteoblasts. Cy-tokine 7:78–82
10. Yamaguchi T, Chattopadhyay N, Kifor O, Butters Jr RR, Sugimoto T, BrownEM 1998 Mouse osteoblastic cell line (MC3T3–E1) expresses extracellular cal-cium (Ca2�
o)-sensing receptor and its agonists stimulate chemotaxis and pro-liferation of MC3T3–E1 cells. J Bone Miner Res 13:1530–1538
11. Kanno S, Anuradha CD, Hirano S 2001 Chemotactic responses of osteoblasticMC3T3–E1 cells toward zinc chloride. Biol Trace Elem Res 83:49–55
12. Godwin SL, Soltoff SP 2002 Calcium-sensing receptor-mediated activation ofphospholipase C-�1 is downstream of phospholipase C� and protein kinaseC in MC3T3–E1 osteoblasts. Bone 30:559–566
13. Lisignoli G, Toneguzzi S, Piacentini A, Cattini L, Lenti A, Tschon M, Cris-
2334 Endocrinology, May 2005, 146(5):2324–2335 Yano et al. • RANTES Actions on Osteoblasts

tino S, Grassi F, Facchini A 2003 Human osteoblasts express functional CXCchemokine receptors 3 and 5: activation by their ligands, CXCL10 and CXCL13,significantly induces alkaline phosphatase and �-N-acetylhexosaminidase re-lease. J Cell Physiol 194:71–79
14. Gasper NA, Petty CC, Schrum LW, Marriott I, Bost KL 2002 Bacterium-induced CXCL10 secretion by osteoblasts can be mediated in part throughtoll-like receptor 4. Infect Immun 70:4075–4082
15. Wright KM, Friedland JS 2002 Differential regulation of chemokine secretionin tuberculous and staphylococcal osteomyelitis. J Bone Miner Res 17:1680–1690
16. Lisignoli G, Toneguzzi S, Grassi F, Piacentini A, Tschon M, Cristino S,Gualtieri G, Facchini A 2002 Different chemokines are expressed in humanarthritic bone biopsies: IFN-� and IL-6 differently modulate IL-8, MCP-1 andRANTES production by arthritic osteoblasts. Cytokine 20:231–238
17. Graves DT, Jiang Y, Valente AJ 1999 The expression of monocyte chemoat-tractant protein-1 and other chemokines by osteoblasts. Front Biosci 4:D571–D580
18. Zhu JF, Valente AJ, Lorenzo JA, Carnes D, Graves DT 1994 Expression ofmonocyte chemoattractant protein 1 in human osteoblastic cells stimulated byproinflammatory mediators. J Bone Miner Res 9:1123–1130
19. Votta BJ, White JR, Dodds RA, James IE, Connor JR, Lee-Rykaczewski E,Eichman CF, Kumar S, Lark MW, Gowen M 2000 CK�-8 [CCL23], a novel CCchemokine, is chemotactic for human osteoclast precursors and is expressedin bone tissues. J Cell Physiol 183:196–207
20. Rossi D, Zlotnik A 2000 The biology of chemokines and their receptors. AnnuRev Immunol 18:217–242
21. Gerard C, Rollins BJ 2001 Chemokines and disease. Nat Immunol 2:108–11522. Strieter RM, Polverini PJ, Kunkel SL, Arenberg DA, Burdick MD, Kasper J,
Dzuiba J, Van Damme J, Walz A, Marriott D, Chan SY, Roczniak S, ShanafeltAB 1995 The functional role of the ELR motif in CXC chemokine-mediatedangiogenesis. J Biol Chem 270:27348–27357
23. Ma Q, Jones D, Borghesani PR, Segal RA, Nagasawa T, Kishimoto T, Bron-son RT, Springer TA 1998 Impaired B-lymphopoiesis, myelopoiesis, and de-railed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice. ProcNatl Acad Sci USA 95:9448–9453
24. Horuk R 2001 Chemokine receptors. Cytokine Growth Factor Rev 12:313–33525. Proudfoot AE, Power CA, Wells TN 2000 The strategy of blocking the che-
mokine system to combat disease. Immunol Rev 177:246–25626. Schall TJ, Jongstra J, Dyer BJ, Jorgensen J, Clayberger C, Davis MM, Krensky
AM 1988 A human T cell-specific molecule is a member of a new gene family.J Immunol 141:1018–1025
27. von Luettichau I, Nelson PJ, Pattison JM, van de Rijn M, Huie P, Warnke R,Wiedermann CJ, Stahl RA, Sibley RK, Krensky AM 1996 RANTES chemo-kine expression in diseased and normal human tissues. Cytokine 8:89–98
28. Schall TJ 1991 Biology of the RANTES/SIS cytokine family. Cytokine 3:165–183
29. Schall TJ, Bacon K, Toy KJ, Goeddel DV 1990 Selective attraction of mono-cytes and T lymphocytes of the memory phenotype by cytokine RANTES.Nature 347:669–671
30. Yu X, Huang Y, Collin-Osdoby P, Osdoby P 2004 CCR1 chemokines promotethe chemotactic recruitment, RANKL development, and motility of osteoclastsand are induced by inflammatory cytokines in osteoblasts. J Bone Miner Res19:2065–2077
31. Chattopadhyay N, Yano S, Tfelt-Hansen J, Rooney P, Kanuparthi D, Ban-
dyopadhyay S, Ren X, Terwilliger E, Brown EM 2004 Mitogenic action ofcalcium-sensing receptor on rat calvarial osteoblasts. Endocrinology 145:3451–3462
32. Okamatsu Y, Kim D, Battaglino R, Sasaki H, Spate U, Stashenko P 2004MIP-1� promotes receptor activator of NF-�B ligand-induced osteoclast for-mation and survival. J Immunol 173:2084–2090
33. Rivera A, Jarolim P, Brugnara C 2002 Modulation of Gardos channel activityby cytokines in sickle erythrocytes. Blood 99:357–363
34. Bylund DB 1986 Graphic presentation and analysis of inhibition data fromligand-binding experiments. Anal Biochem 159:50–57
35. Bylund DB, Toews ML 1993 Radioligand binding methods: practical guideand tips. Am J Physiol 265:L421–L429
36. Tfelt-Hansen J, Chattopadhyay N, Yano S, Kanuparthi D, Rooney P, SchwarzP, Brown EM 2004 Calcium-sensing receptor induces proliferation throughp38 mitogen-activated protein kinase and phosphatidylinositol 3-kinase butnot extracellularly regulated kinase in a model of humoral hypercalcemia ofmalignancy. Endocrinology 145:1211–1217
37. Parent CA, Devreotes PN 1999 A cell’s sense of direction. Science 284:765–77038. Firtel RA, Chung CY 2000 The molecular genetics of chemotaxis: sensing and
responding to chemoattractant gradients. Bioessays 22:603–61539. Hirsch E, Katanaev VL, Garlanda C, Azzolino O, Pirola L, Silengo L, Sozzani
S, Mantovani A, Altruda F, Wymann MP 2000 Central role for G protein-coupled phosphoinositide 3-kinase � in inflammation. Science 287:1049–1053
40. Fukuyama R, Fujita T, Azuma Y, Hirano A, Nakamuta H, Koida M, KomoriT 2004 Statins inhibit osteoblast migration by inhibiting Rac-Akt signaling.Biochem Biophys Res Commun 315:636–642
41. Mitsiades CS, Mitsiades N, Koutsilieris M 2004 The Akt pathway: moleculartargets for anti-cancer drug development. Curr Cancer Drug Targets 4:235–256
42. Murphy PM, Baggiolini M, Charo IF, Hebert CA, Horuk R, Matsushima K,Miller LH, Oppenheim JJ, Power CA 2000 International union of pharma-cology. XXII. Nomenclature for chemokine receptors. Pharmacol Rev. 52:145–176
43. Chen TL, Hauschka PV, Feldman D 1986 Dexamethasone increases 1,25-dihydroxyvitamin D3 receptor levels and augments bioresponses in rat os-teoblast-like cells. Endocrinology 118:1119–1126
44. Borish LC, Steinke JW 2003 Cytokines and chemokines. J Allergy Clin Im-munol 111:S460–S475
45. Fuller K, Owens JM, Chambers TJ 1995 Macrophage inflammatory protein-1�and IL-8 stimulate the motility but suppress the resorption of isolated ratosteoclasts. J Immunol 154:6065–6072
46. Lisignoli G, Toneguzzi S, Pozzi C, Piacentini A, Riccio M, Ferruzzi A,Gualtieri G, Facchini A 1999 Proinflammatory cytokines and chemokine pro-duction and expression by human osteoblasts isolated from patients withrheumatoid arthritis and osteoarthritis. J Rheumatol 26:791–799
47. Cappellen D, Luong-Nguyen NH, Bongiovanni S, Grenet O, Wanke C, SusaM 2002 Transcriptional program of mouse osteoclast differentiation governedby the macrophage colony-stimulating factor and the ligand for the receptoractivator of NF�B. J Biol Chem 277:21971–21982
48. Wright KM, Friedland JS 2003 Complex patterns of regulation of chemokinesecretion by Th2-cytokines, dexamethasone, and PGE2 in tuberculous osteo-myelitis. J Clin Immunol 23:184–193
49. Mundy GR 2003 Bone remodeling. In: Favus MJ, ed. Primer on the metabolicbone diseases and disorders of mineral metabolism. 5th ed. Philadelphia:Lippincott Williams & Wilkins; 46–58
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
Yano et al. • RANTES Actions on Osteoblasts Endocrinology, May 2005, 146(5):2324–2335 2335




















