
Characterization of a platelet-activating factor acetylhydrolase secreted by the nematode parasite...
17
BIOLOGY OF REPRODUCTION 41, 587-603 (1989) Characterization of Platelet-Activating Factor Binding Sites on Uterine Membranes from Pregnant Rabbits1 GEORGE B. KUDOLO and MICHAEL J. K. HARPER2 Department of Obstetrics and Gynecology University of Texas Health Science Center at San Antonio San Antonio, Texas 78284-7836 ABSTRACT One of the earliest signs of endometrial preparation for blasiocyst implantation is a localized increase in capillary permeability, an event that is essentially inflammatory in character and thought to be a prerequisite for subsequent decidual tissue formation. Platelet-activating factor (PAF), chemically identified as 1 -0-alkyl- 2-acetyl-sn-glycero-3- phosphorylcholine, is a very potent vasoactive compound that recently has been implicated in the implantation process. In the present study, PAF binding, sites are characterized in the rabbit uterus. A specflc, reversible, saturable, and thermally labile binding of (-‘HJPAF to uterine membranes has been demon- strated, exhibiting multiple binding sites. The equilibrium dissociation constant (Kj) of the higher affinity binding site (type 1) was 3.6 ± 0.4 nM (mean ± SD) with a binding capacity (B,,,,) of 3.4 ± 1.6 pmollmg protein. The second (lower affinity)binding site (type 2) had an apparent Kd of 114.6 ± 13.5 nM and a of 164.3 ± 17.6 pmollmg membrane protein, under the conditions of maximal (3HJPAF binding, 25 V. 150 mm. Incubasions at 4 Vfor up to 3 h yielded only 30% of the observed at 25 V. In crude and pur(fied endometrial membrane preparations in which the PAP binding was predominantly located, the affinity of the binding for PAP was significantly higher than for the whole uterus, giving K(Js of 1.5 ± 0.8 and 0.8 ± 0.5 nM; these latter values were not significantly different. However, the B,, values of 3.9 ± 0.9 pmol/mg protein and 376.8 ± 1633 fmol/mg protein for the two endometrial preparations, respectively, did differ significantly. Kinetic analysis at 25 V re- sulted in a calculated Kd of328 ± 1.14 nM, which did not differ from the value for the whole uterus at the same temperature, but was greater than for the endomerrial preparations. Using 4 nM (3HJPAF to selectively label only the type 1 binding sites, the relative potencies of PAF and its antagonists in displacing (3H]PAF were lyso-PAF>CV3988>PAF>U66985>A02405>BN52021>U66982. The antagonists SRI 63,441 and L652,731 were ineffective in displacing (3HJPAF at upto 5000-fold molar excess of (3HJPAF. (3HjLyso-PAF binding at 4 nM was displaceable by PAP. All cations tested, i.e. Cafl, Mg2, K, Na, and Lit, inhibited (3H]PAF binding. Serine hydrolase inhibitors, diisopropylfluorophosphate (DFP) and phenylmethylsulfonyl fluoride (PMSF), inhibited binding, but bacitracin, leupeptin, and antipain stabilized it. Bacitracin was best and more consistent in combination with other enzyme inhibitors, allowing up to 20% more binding to be measured. Under the conditions of maximwn (3HJPAF binding, the label was metabolized to lyso- PAP (17%) and alkylacyl-glyceryl ether phosphoryicholine (GPC) (43%), with only 36% remaining intact. In the presence of the phospholipase A2 inhibitors, dibromoacetophenone (2p.M) and quinacrine (10 l.aM), and the receptor-stabilizing antibiotic bacitracin (100 p.M), metabolism of the (HJPAF was substantially controlled so that 60-70% of the added (3HJPAF remained unchanged, the level of conversion to alkylacyl-GPC was reduced to less than 6%, and the amount converted to lyso-PAF remained at less than 20%. Using both (3HJPAF and (3Hjlyso-PAF to trace the metabolic pathways, we found that (3HJPAF was metabolized first to lyso-PAP during the deacetylation step and then further degraded to alkylacyl-GPC. There was no evidence that the alkylacyl-GPC or lyso-PAF was converted back to PAP to any significant degree under these experimental conditions. These data suggest that the type 1 binding sites are the PAP receptors. The reversible nature of the binding between lyso-PAP, a biologically inactive metabolite, and PAP at the receptor site may serve under physiological conditions to desensitize the uterus during lyso-PAF occupation. The ability of PAP antagonists to bind to differ- eat degrees may help to elucidate the molecular characteristics of the PAP receptor sites and permit synthesis of antagonists that work specifically in the uterus. _____ INTRODUCTION Accepted May 16, 1989. Received February 13, 1989. A localized increase in endometrial vascular per- l’1j work was supported in part by NIH grants HD 14048, HD 25224, f th li t f and HI) 10202 (Hormone Receptor Core), and a Lalor Foundation Fellow- me tY is one o e ear es signs o astocyst ship to George B. Kudolo. implantation, and this response may be likened to an Obstethcs and Gynecology, University of Texas Health Science Center rnflammatory reaction elicited by vasoactive mediators San Antonio, 7703 Floyd Curl Drive, San Antonio, TX 78284-7836. (Psychoyos, 1973; Psychoyos and Casimiri, 1980). 587
Transcript of Characterization of a platelet-activating factor acetylhydrolase secreted by the nematode parasite...

BIOLOGY OF REPRODUCTION 41, 587-603 (1989)
Characterization of Platelet-Activating FactorBinding Sites on Uterine Membranes from Pregnant Rabbits1
GEORGE B. KUDOLO and MICHAEL J. K. HARPER2
Department of Obstetrics and Gynecology
University of Texas Health Science Center at San Antonio
San Antonio, Texas 78284-7836
ABSTRACT
One of the earliest signs of endometrial preparation for blasiocyst implantation is a localized increase incapillary permeability, an event that is essentially inflammatory in character and thought to be a prerequisite forsubsequent decidual tissue formation. Platelet-activating factor (PAF), chemically identified as 1 -0-alkyl-2-acetyl-sn-glycero-3- phosphorylcholine, is a very potent vasoactive compound that recently has been implicatedin the implantation process. In the present study, PAF binding, sites are characterized in the rabbit uterus. Aspec�flc, reversible, saturable, and thermally labile binding of (-‘HJPAF to uterine membranes has been demon-strated, exhibiting multiple binding sites. The equilibrium dissociation constant (K�j) of the higher affinity bindingsite (type 1) was 3.6 ± 0.4 nM (mean ± SD) with a binding capacity (B,,,�,) of 3.4 ± 1.6 pmollmg protein. Thesecond (lower affinity)binding site (type 2) had an apparent Kd of 114.6 ± 13.5 nM and a � of 164.3 ± 17.6pmollmg membrane protein, under the conditions of maximal (3HJPAF binding, 25 V. 150 mm. Incubasions at4 Vfor up to 3 h yielded only 30% of the � observed at 25 V. In crude and pur(fied endometrial membranepreparations in which the PAP binding was predominantly located, the affinity of the binding for PAP wassignificantly higher than for the whole uterus, giving K(Js of 1.5 ± 0.8 and 0.8 ± 0.5 nM; these latter values werenot significantly different. However, the B,,� values of 3.9 ± 0.9 pmol/mg protein and 376.8 ± 1633 fmol/mgprotein for the two endometrial preparations, respectively, did differ significantly. Kinetic analysis at 25 V re-sulted in a calculated Kd of328 ± 1.14 nM, which did not differ from the value for the whole uterus at the sametemperature, but was greater than for the endomerrial preparations.
Using 4 nM (3HJPAF to selectively label only the type 1 binding sites, the relative potencies of PAF and itsantagonists in displacing (3H]PAF were lyso-PAF>CV3988>PAF>U66985>A02405>BN52021>U66982. Theantagonists SRI 63,441 and L652,731 were ineffective in displacing (3HJPAF at upto 5000-fold molar excess of(3HJPAF. (3HjLyso-PAF binding at 4 nM was displaceable by PAP. All cations tested, i.e. Cafl, Mg2�, K�,Na�, and Lit, inhibited (3H]PAF binding. Serine hydrolase inhibitors, diisopropylfluorophosphate (DFP) andphenylmethylsulfonyl fluoride (PMSF), inhibited binding, but bacitracin, leupeptin, and antipain stabilized it.Bacitracin was best and more consistent in combination with other enzyme inhibitors, allowing up to 20% morebinding to be measured. Under the conditions of maximwn (3HJPAF binding, the label was metabolized to lyso-PAP (17%) and alkylacyl-glyceryl ether phosphoryicholine (GPC) (43%), with only 36% remaining intact. In thepresence of the phospholipase A2 inhibitors, dibromoacetophenone (2�p.M) and quinacrine (10 l.aM), and thereceptor-stabilizing antibiotic bacitracin (100 p.M), metabolism of the (�HJPAF was substantially controlled sothat 60-70% of the added (3HJPAF remained unchanged, the level of conversion to alkylacyl-GPC was reducedto less than 6%, and the amount converted to lyso-PAF remained at less than 20%. Using both (3HJPAF and(3Hjlyso-PAF to trace the metabolic pathways, we found that (3HJPAF was metabolized first to lyso-PAP duringthe deacetylation step and then further degraded to alkylacyl-GPC. There was no evidence that the alkylacyl-GPCor lyso-PAF was converted back to PAP to any significant degree under these experimental conditions.
These data suggest that the type 1 binding sites are the PAP receptors. The reversible nature of the bindingbetween lyso-PAP, a biologically inactive metabolite, and PAP at the receptor site may serve under physiologicalconditions to desensitize the uterus during lyso-PAF occupation. The ability of PAP antagonists to bind to differ-eat degrees may help to elucidate the molecular characteristics of the PAP receptor sites and permit synthesis ofantagonists that work specifically in the uterus.
_____ INTRODUCTIONAccepted May 16, 1989.Received February 13, 1989. A localized increase in endometrial vascular per-l’�1�j� work was supported in part by NIH grants HD 14048, HD 25224, f th li t f �
and HI) 10202 (Hormone Receptor Core), and a Lalor Foundation Fellow- me tY is one o e ear es signs o �astocystship to George B. Kudolo. implantation, and this response may be likened to an
Obstethcs and Gynecology, University of Texas Health Science Center � rnflammatory reaction elicited by vasoactive mediatorsSan Antonio, 7703 Floyd Curl Drive, San Antonio, TX 78284-7836. (Psychoyos, 1973; Psychoyos and Casimiri, 1980).
587

588 KUDOLO AND HARPER
There is persuasive evidence that prostaglandins (PGs)constitute one such group of vasoactive compounds(Lau et al., 1973; Saksena et a!., 1976; Biggers et al.,1978; Evans and Kennedy, 1978; Jones et a!., 1986).Much of this evidence has been derived from experi-ments in which the PG cyclooxygenase inhibitor indo-methacin has been used. In other experiments, an in-crease in PG synthesis has been concomitant with anincrease in another class of potent vasoactive mediators
collectively called platelet-activating factor (PAF)(Yousufzai and Abdel-Latif, 1985). It is now knownthat during the synthesis of PAF from its parent phos-pholipid, arachidonic acid (a precursor for PGs) isreleased. Consequently, any stimulus that affects phos-pholipid turnover may result in the induction of bothmediators. PAF, chemically identified as 1-O-alkyl-2-acetyl-sn-glycero-3-phosphorylcholine, has beenshown to modulate increased vascular permeability(McManus et aL, 1980, 1981; Pinckard et al., 1982;Snyder, 1985) and has a wide range of other biologicalactivities (Vargaftig et al., 1981; Braquet et al., 1987).Thus, it seems likely that PGs are not the only sub-stances that are able to alter uterine capillary per-meability.
A number of different cells are capable of producingPAF in response to various stimuli. Recently, it hasbeen shown that developing mouse and human zygotesproduce PAF (O’Neill and Saunders, 1984; O’Neill,1985a,b; O’Neill et al., 1985; Angle et al., 1988a) andthat the presence of fertilized zygotes in the maternalreproductive tract causes significant thrombocytopenia,
probably due to this PAF. The generation of embryo-derived PAF by human embryos in vitro correlates withtheir viability and pregnancy potential (O’Neill, 1987).Furthermore, PAF antagonists can interrupt early preg-nancy in mice (Spinks and O’Neill, 1987, 1988) andrats (Acker et a!., 1988).
There is evidence for uterine production of PAF inrabbits (Angle et a!., 1985, 1988b; Yasuda et a!., 1986)and rats (Yasuda et a!.. 1986; Nakayama et al., 1987)and in human endometrial stromal cells in culture
(Alecozay et al., 1988). Whether embryo-derived PAFor that derived from stromal cells actually modulates
the increased vascular permeability associated with theimplantation process has yet to be determined.
What has been confirmed is the involvement of
specific stereoreceptors for PAF. Initially, this assump-tion was based on the fact that only the naturally
occurring stereoisomer (R) stimulated PAF responses(Vargaftig and Benveniste, 1983). The existence of
PAF receptors has since been confirmed by biochemicalbinding studies for human polymorphonuclear leuko-cytes (PMNs) (Marquis et a!., 1988), human platelets(Korth and Benvemste, 1987), human lung membranes(Hwang and Lam, 1986), and rabbit platelet membranes(Hwang et a!., 1986a; Robaut et a!., 1987). In platelets,the binding was displaceable by several PAF antago-nists that evidently competed directly with receptor-binding. The potencies of these compounds and their
competitive inhibition of [3H]PAF binding correlatedhighly with a biological endpoint, the ability to antago-
nize PAF-induced platelet aggregation, hypotension,and bronchospasm (Braquet and Godfroid, 1986; God-
froid and Braquet, 1986). The affinity and bindingcapacities of these receptors displayed both tissue- andspecies-specificity, and the multiple molecular nature ofPAF itself has led to the suggestion that PAF receptorsubtypes may exist.
No published report is yet available on the presenceof PAF receptors in the female reproductive tract, and
the present study is directed to characterizing the PAFbinding sites in the rabbit uterus on Day 6 of pregnan-cy, and identifying which uterine tissue compartmentpossesses these receptors.
Materials
MATERIALS AND METHODS
Tritiated PAF (1 -O-[3H]alkyl-2-acetyl-sn-glyceryl-3-phosphorylcholine), a mixture of C16 and C18 alkyl
ethers (sp. act. 81 Ci/mmol), PAF,C18 (1-O-[3H]octa-decyl-2-acetyl-sn-glyceryl-3-phosphorylcholine: sp. act.132 Ci/mmol), and lyso-PAF (1-O-[3H]octadecyl-sn-glyceryl-3-phosphorylcholine; sp. act. 92 Cilmmol)were purchased from Amersham International, Arling-ton Heights, IL.
Unlabeled 1 -O-hexadecyl-2-acetyl-sn-glyceryl-3-phosphoryicholine (PAF,C16), 1 -O-octadecyl-2-acetyl-sn-glyceryl-phosphorylcholine (PAF,C18), and 1-0-
hexadecyl-sn-glyceryl-3-phosphorylcholine (lyso-PAF,C16) were obtained from Sigma Chemical Compa-ny, St. Louis, MO. CV3988 (rac-3-(N-n-octadecylcar-bamoyloxy)-2-methoxypropyl-2-thiazolioethyl phos-phate) was a gift from Dr. M Nishikawa, TakedaChemical Industries, Ltd., Osaka, Japan; U66982 (1-0-
octadecyl - 2 - acetyl - sn - glycero-3-phosphoric-acid-10’-trimethylammoniumdecyl ester) and U66985 (1-0-octadecyl-2-acetyl-sn-glycero-3-phosphoric acid-6’-trimethylammoniumhexyl ester) made by the Upjohn

ENDOMETRIAL PLATELET-ACTIVATING FACTOR RECEPTORS 589
Company were gifts from Dr. Donald Hanahan, Depart-ment of Biochemistry, University of Texas Health Sci-ence Center at San Antonio; L652,731 [trans-2,5-Bis(3,4,5-trimethoxypheny!) tetrahydrofuran] was a gift
from Dr. J. Chabala, Merck Sharp & Dobme, Rahway,NJ; Nova A02405 (1 -0-hexadecyl-2-acetyl-sn-glycero-3-phospho(N,N,N-trimethyl)hexanolamine) (this com-pound is the C16 homolog of U66985) was a gift fromNova Pharmaceuticals Corp., Baltimore, MD;SR163,441 (cis(±)-1 -[2-[hydroxy-[tetrahydro-5-[(oc-tadecylaminocarbonyl)oxy]methyl] furan-2-yl ]-methox-yphosphinyloxy ethyl] -quinolimum hydroxide, innersalt) was a gift from Sandoz Research Institute, Base!,Switzerland; and the ginkgolide B, BN52021 (3-t-butyl-hexahydro-4,7b, 1 1-trihydroxy-8-methyl-9H-1, 7a-ep-oxymethano-1H, 6a-H-cyclopenta[c]furol [2,3-b]furo [3’,2’:3,4] cyclopenta[ 1 ,2-d] furan-5,9, 12(4H)trionewas a gift from Dr. P. Braquet, Institut Henri Beaufort,Le Plessis-Robinson, France.
All reagents were of analytical grade. Bacitracin,diisopropylfluorophosphate (DFP), and dibromoaceto-phenone (BpB) were purchased from Aldrich Chemical
Company, Milwaukee, WI. Antipain, phenylmethylsul-fonylfluoride (PMSF), leupeptin, sucrose, sodium azide,tris(hydroxymethyl)aminomethane (Tris)-HC1, monothi-oglycerol, and quinacrine (dihydrochlonde) were ob-tained from Sigma Chemical Company. Ethylenedia-minetetraacetate (EDTA), disodium salt, was purchasedfrom Eastman Kodak Company, Rochester, NY, andbovine serum albumin (BSA, Fraction V) from Chemi-
cal Credential (ICN Immunobiochemicals), Lisle, IL.
Animals
Mature New Zealand White-Cambridge female rab-bits (BW>3.0 kg) were obtained from Penn AcresRanch, Wimberly, TX, and caged individually in acontrolled enviromnent with a 14L:1OD cycle, fed 170g rabbit pellets/day and provided with tap water adlibitum. On the day designated Day 0 of pregnancy,rabbits were inseminated with 0.5 ml mixed sperm
suspension collected, via an artificial vagina, from fer-tile bucks immediately before use. After insemination,donor animals were injected i.v. with 100 Hi humanchorionic gonadotropin (hCG, Ayerst Laboratories,New York, NY).
Tissue Preparation
On Day 2 or Day 6 of pregnancy, rabbits were killedwith an i.v. overdose of pentobarbital sodium. Uterine
horns were rapidly removed and the zygotes were re-moved by flushing (Day 2), or uterine horns wereopened along the line of mesometria! attachment and
blastocysts were removed (Day 6). The uterine hornswere then either cut into l-g portions for storage orseparated into endometrial and myometrial fractionsbefore storage at -137#{176}Cuntil required.
Preparation of Plasma Membranes
All steps were carried out at 0-4#{176}Cand the proce-dure for isolating the uterine/endometrial plasma mem-branes was based on that used by Lampelo et al. (1985)for purifying rabbit endometrial membranes. Fresh orfrozen uterine/endometrial tissues were rapidly cut intovery small pieces before homogenization in 20 volumesof TEM (10 mM Tns-HC1, 1 mM EDTA, 1 mMmonothioglycerol, pH 7.4) or TEBBQ (10 mM Tris-HC1, 1 mM EDTA, 0.1 M bacitracin, 2 i.LM dibromoac-etophenone, 10 �tM quunacrine, pH 7.4) using a Poly-tron homogenizer at speed setting 6 for 10 s. Thehomogenate was strained through a 350-�tm nylon meshand centrifuged at 1000 x g for 5 miii to isolate themembrane-rich supernatant, which was then centrifugedat 30,000 x g for 30 miii at 4#{176}Cto harvest a crudeplasma-membrane-rich pellet. The pellet was washedwith a 10 vols TEM buffer. The washed membranepreparation was then either stored at -137#{176}Cuntil re-quired for binding assays or resuspended in TEM/TEBBQ buffer in a Dounce glass tissue grinder using 3strokes of a teflon pestle (clearance 0.063-0.139 mm)for a binding assay.
Membrane Purification
Membranes from endometrial tissues were purifiedby suspending the 30,000 x g crude membrane pellet in10 ml 0.25 M sucrose in working buffer (TEM orTEBBQ). Two-milliliters of this suspension was loadedon 9 ml of 3 discontinuous gradients (3 ml of I M, 2M, and 3 M sucrose) and centrifuged for 30 mm at30,000 x g, producing two distinct bands of membranefractions (at the boundaries between 1 M and 2 M [Fl]and 2 M and 3 M sucrose [F2] concentrations) thatwere removed with a syringe and washed with workingbuffer, the membranes were repelleted by centrifuga-tion at 30,000 x g, for 45 mm before storage at -137#{176}C.Protein content of the membrane preparation was esti-mated by using a Bio-Rad kit (Bio-Rad Laboratories,Richmond, CA) with bovine gamma globulin as stan-dard reference protein.

590 KUDOLO AND HARPER
Enzyme Assays
Alkaline phosphalase enzyme activity was assayed todetermine the degree of enrichment of the purifiedmembrane fraction (Lampelo et al., 1985). The standardassay mixture consisted of 0.3 ml 100 mM Tris-HC1,containing 8.4 mM MgCl2, pH 9.5, 0.1 ml 5 mMphosphatase substrate (final I mM), and 0.1 ml sample.The assays were carried out at 37#{176}Cfor 30 miii and thereactions were terminated with 0.8 ml 0.1 M NaOH.Control values were determined by incubating the endo-metrial samples without substrate until after the30-mn incubation period and terminating the reactionwith 0.1 M NaOH. Enzyme activity was determinedafter 10 mm by measuring the absorbance at 410 nmfrom a standard curve constructed with p-nitrophenol.
Binding Assays
All binding assays were performed with uterineplasma membranes from Day 6 of pregnancy, with the
exception of the time course studies in which mem-branes isolated on Day 2 were used. The rabbits werebeing killed for zygote recovery in other experiments.The binding assays were initially conducted with TEM
containing 0.25% BSA, but since there was no increaseof specific binding compared to use of plain buffer,plain buffer was used in all subsequent assays. Theassays were routinely carried out in triplicate in clearpolystyrene tubes (12 x 75 mm) in a 0.4-mi reactionvolume (membrane suspension, 0.2 ml; [3H]PAF/lyso-PAF, 0.05 ml; radioinert competing ligand/buffer, 0.05ml; buffer containing inhibitors/cations, 0.1 ml). After
incubation of the reaction mixtures with ligand concen-trations and under conditions described below, the incu-
bates were rapidly transferred onto ice in batches of 9tubes; 1 ml of washing buffer was added, the sampleswere vortexed, and unbound tritiated ligands were re-moved by vacuum filtration through Whatman GF/Cglass fiber discs (Whatman International Ltd., Maid-stone, England) previously soaked in plain buffer or thewashing buffer (TEM or TEBBQ containing 150 mMNaC1, 0.5% NaN3 and 0.25% BSA). The filters werewashed with 20 ml ice-cold buffer, vacuumed dry, and
placed in mini-scintillation vials for counting of radio-activity after addition of 3.5 ml AquaMix (ICN Radio-chemicals, Irvine, CA).
Based on recent studies on [3H]PAF binding
(Hwang, 1987; Korth and Benveniste, 1987; Robaut eta!., 1987; Marquis et al., 1988), 100 �ig of uterine
plasma membrane protein/tube was used for the bindingassays.
Optimization of Specific (3HjPAF Binding:
Effect of Washing Buffer
Specific binding of [3H]PAF to 100 �.Lg of uterinemembranes was determined by using 4 nM [3H]PAFwith 20 �iM PAF,C18 as competitor at 25#{176}Cfor 150mm. After the incubation, 1 ml ice-cold TEM withoutBSA or with BSA, either at 0.25% or 1%, was addedand vortexed and placed on ice for 1-2 miii beforevacuum filtration. The filter discs were washed with 20ml of plain TEM or TEM containing 0.25% BSA. Theremaining set of tubes to which 1 ml 1% BSA bufferhad been added was washed with 10 ml 1% BSAfollowed by 10 ml 0.25% BSA washing buffer.
Competitive Inhibition Studies
Radiounert ligands were added at 10-10,000-fold mo-lar excess to compete for 4 nM [3H]PAF at 25#{176}Cfor150 mm. To assess the comparative potencies of somecompetitors to displace [3H]PAF and [3HIlyso-PAFfrom uterine binding sites, 4 nM of these radioligandswere incubated with 100 p�g uterine membranes in thepresence or absence of 20 IIM of the competing com-pounds at 4#{176}or 25#{176}Cfor 150 mm or at 4#{176}Cfor 18 h.Since the proportion of the PAF,C16 and PAF,C18 inthe [3H]PAF mixture was unspecified, the level ofnonspecific binding in purified endometrial membraneswas estimated with 20 i.LM PAF,C18, PAF,C16, or amixture of these ligands in the proportions (C18:C16),75:25, 50:50 and 25:75. Corresponding tubes containingthe same membranes and radioinert ligands were la-beled with [3HJPAF,C18.
Phospholipids also tend to form micelles, especiallyat high concentrations in aqueous solutions. Experi-ments were conducted to ascertain the effect of using20 �.tM of radioinert PAF,C18 on [3H]PAF binding.Endomeinal membranes were incubated with 1 or 4 nM[3H]PAF as follows: either (1) [3H]PAF was incubatedwith the membranes (100 p.g protein) for up to 30 miiiprior to addition of the radioinert PAF,C18 (5 or 20 j.LM,respectively), (2) the radiounert PAF,C18 was incubatedwith the membranes for up to 30 miii prior to additionof [3H]PAF, (3) both [3H]PAF and radioinert PAF,C18were incubated together for 30 miii prior to addition ofthe membranes, or (4) all reactants were mixed at thesame time. In all cases, the incubations were carried outfor 150 mm at 25#{176}C.

ENDOMETRIAL PLATELET-ACFIVATING FACTOR RECEPTORS 591
Time Course/Thermal Stability
of [3HJPAF Binding
A fmal concentration of 4 nM [3H]PAF (in theabsence and presence of 20 p.M radioinert PAF,C18)was used in incubations (4#{176}C,25#{176}C,and 60#{176}C)of wholeuterine membranes from very early pregnancy (Day 2).At 30-mn intervals for up to 3 h, aliquots were taken,and bound [3H]PAF was measured to determine thethermal stability of the [3H]PAF-uterine membranebinding.
Equilibrium Saturation Analyses
[3H]PAF equilibrium binding studies were performed
at 4#{176}Cor 25#{176}Cfor 150 miii with 0.125-16 nM of theradioligand for whole uterine plasma membranes and0.125-4 nM for both crude and purified endometrialmembranes. The nonspecific binding was assessed byusing 20 p.M PAF,C18.
Kinetic Analyses
Purified endometrial membrane suspensions (5 ml)were incubated at 25#{176}Cwith 4 nM [3H]PAF (finalconcentration) in two tubes: one for total binding in theabsence of radioinert PAF and the other for estimating
nonspecific binding in the presence of 20 p.M radioinertPAF,C18 (final concentration). To determine the associ-ation rate of [3HJPAF with the endometrial membranes,0.1-mi aliquots (100 p.g membrane protein) were takenat 5-min intervals for the first 30 mm (and thereafter at15-mn intervals) and added to I ml ice-cold 1% BSAin working buffer. This was vortexed quickly and themembrane-bound [3H]PAF was separated from the un-bound fraction by vacuum filtration. After 90-120 min,when equilibrium was achieved, 2 ml of each of theincubated membrane suspensions were taken, and ra-dioinert PAF,C18 was added to each to achieve a finalconcentration of 40 p.M (10,000-fold molar excess ofthe radioligand). Aliquots were taken at 5-min intervals
for the first 30 mm, and at 15-nun intervals thereafter,to determine the reversible nature of the [3H]PAF-membrane interaction and the rate at which the addedradioinert PAF,C18 displaced bound [3H]PAF. Concur-rently, the association kinetic studies continued, servingas controls during the dissociation kinetic studies.
Effect of Cations and Inhibitors
on [3H]PAF Binding
Specific [3HIPAF binding to 100 p.g uterine mem-branes was measured with 4 nM [3H]PAF in the pres-
ence of CaCl2 (2 mM), MgC12 (10 mM), LiC1 (10mM), KC1 (150 mM), NaC1 (150 mM), antipain (150p.M), leupeptin (150 I.LM), bacitracin (150 p.M), andPMSF (2 mM). Nonspecific binding was estimated byusing 20 p.M radioinert PAF,C18.
Localization of the [3H]PAF
Binding in Uterine Tissues
The endometrium was scraped from the uterine myo-metrium, and plasma membranes were prepared fromthe endometrium, residual myometrium, and the wholeuterus. Specific binding was measured in these fractionswith 100 jig of the membrane preparations labeled with4 nM [3H]PAF and nonspecific uptake of the radioli-gand estimated with 10 p.M PAF,C18.
Thin-Layer Chromatographic Analysis
of [3H]PAF Metabolic Products
Lipid extracts were made from uterine and endome-trial samples, previously incubated at 4#{176}Cand 25#{176}C for150 miii or 4#{176}Cfor 18 h and for which comparative[3H]PAF binding had been determined, by a modifiedprocedure of Bligh and Dyer (1959). Briefly, the reac-tion volume (0.4 ml) was made up to 1.6 ml with
distilled water (or 1.6 ml distilled water was added tovacuum-dried filter disks containing membrane-boundradioactivity), and then 6 ml chloroform 2% acetic acidin methanol (1:2) was added. This monophasic systemwas vortexed at 15-min intervals for an hour, and then4 ml distilled water:chloroform mixture (1:1) wasadded to effect separation of the aqueous and organic(chloroform) phases. The chloroform layer was re-moved by centrifugation, and the aqueous phase waswashed twice with the same volume of chloroform. Thepooled chloroform fraction was evaporated to drynessunder nitrogen gas and resuspended in 0.5 ml chloro-form; 0.05 ml was taken for scintillation counting andfour internal standards were added to the remainingextract: PAF,C18 (2 mg/ml), 0.005 ml; lyso-PAF,C16 (1mg/ml), 0.04 ml; sphingomyelin (1 mg/mi), 0.01 ml;phosphatidyichohne (1 mg/mI), 0.01 ml. The extractswere evaporated and resuspended in 0.05 ml chloro-form before they were loaded onto 20 x 20-cm silicaplates (Silica Gel G, 1000 jim, Analtech, Newark, DE).The silica plates were previously washed in a chioro-form:methanol:water (65:35:6) solvent system and heat-
activated for at least an hour. The plates were run inchlorofonn:methanol:acetic acid:water (50:25:8:4) and

592 KUDOLO AND HARPER
the lipids were visualized in iodine vapor. The zoneswere scraped and the lipid were extracted before count-
ing for radioactivity in 10 ml AquaMix, or the silicagels were counted directly in 10 ml BBOT scintillation
cocktail (2,5-bis (5-tert-butyl-2-benzoxazolyl) thio-phene, 15 g; naphthalene, 300 g; toluene, 2.5 liters;ethyleneglycolmonomethyl ether, 1.5 liters; distilled
water, 125 ml).
Effect of pH on (3H]PAF
Binding and Metabolism
Endometrial tissues were homogenized in TEBBQ
buffer adjusted to pH 6.0 , 7.4, and 9.0 at 4#{176}C,and theisolated plasma membranes were reconstituted in therespective buffers. [3H]PAF binding was carried out at4#{176}Cand 25#{176}Cfor 150 mm.
Analysis of Binding Data
Radioactivity was counted in a Beckman LiquidScintillation counter, Model L330 (Beckman Instru-
ments, Inc., Irvine, CA) at 38% efficiency. The resultsfrom the equilibrium saturation experiments were ana-lyzed by nonlinear regression analysis (Munson, 1987)and the Boeynaems and Swillens plot (Boeynaems andDumont, 1975). The parameters of the binding kineticswere calculated as described by Bennett and Yamamura
(1985). A plot of the in (BnJBm-Bt) against time t wasused to determine the observed association rate constant(k�), whereas the first order rate constant (k i) wasderived from a plot of In BtJBm against time, where Bmis the maximum specific [3H]PAF binding at equilib-rium and B� is the specific binding at time t. The secondorder association rate constant was derived from therelationship k � = (k�-k ...1)/F, where F is the concen-tration of free [3H]PAF used in the assay. The equilib-rium constant (lCd) was calculated from the equation Li
= k ...i/k +1�Binding data from the competition studies were nor-
malized in terms of percentage inhibition by assigningthe total binding in the absence of competitors as 100%.For construction of inhibition curves, the percentageinhibition at each competitor concentration was deter-mined by the formula BCIBT, where BC is the specificbinding (total minus nonspecific binding) at competitorconcentration C and BT is the total binding in theabsence of the competitor. Values of the major meta-bolic products of [3H]PAF/[3H]lyso-PAF were ex-pressed as percentages of the total radioactivity recov-
ered from the scraped silica gels, and the percentagerecovery for each experiment was the total radioactivityrecovered from silica gels/radioactivity loaded x 100.
All values quoted are expressed as mean ± SD for atleast three independent experiments using tissues fromthree rabbits.
RESULTS
Assay of (3H]PAF Binding
to Uterine Membranes and Separation of
Bound from Free Ligand
Previous reports of [3H]PAF binding studies in avariety of cells have shown the dependence of optimalmembrane-[3H]PAF interaction on serum albumin(Korth and Benveniste, 1987), implying that serumalbumin might be a necessary phospholipid carrier for[3H]PAF binding. In this study, BSA concentrations inexcess of 0.25% interfered with the measurable specific[3HJPAF binding. Unlike results reported by (Korth andBenveniste (1987), [3H]PAF binding was observedeven in the complete absence of BSA, and there was nosignificant difference in specific binding in the presenceor absence of 0.25% BSA. The studies reported here,therefore, were conducted without BSA in the incuba-tion medium. However, the presence of BSA in thewashing buffer (1% to terminate the reaction and 0.25%for washing) ensured a more thorough removal of un-bound [3H]PAF from the membranes. It was also ob-served that filters presoaked in 0.25% BSA retained asmuch as twice the radioactivity compared to thosesoaked in plain buffer, which was less than 0.4% ofadded radioligand in the presence or absence of radioin-ert competing compounds.
Estimation of Nonspecific Binding,
Time Course, and Thermal Stability
of (3HJPAF Binding
The lipophilic nature of PAF confers the ability toattach to a wide variety of membrane moieties-hencethe high level of nonspecific binding reported in almostall cells studied. In the present studies using 20 p.MPAF,C1g/100 jig of membrane protein to compete for[3H]PAF, which is a mixture of the 1-0-C16 and C18-alkyl-ethers, the level of nonspecific binding was
40-70%, depending on the radioligand concentration,membrane type (uterine/endometrial), its purity, and thetemperature of incubation. The proportion of the ethers

C-D
C
-oC)
‘4-
C-)
0.U)C0
z
.f-
.�
...
3.5
3.0
2.5
20
I.5
I.0
0.5
4#{176}C� 25#{176}C
�. 60#{149}C��J14rL4100:0 75:25 5Q:5Q 25:75 0:100
Relative % of PAF (C18:C16)
30 60 90 120 ISO 18o
Time (minutes)
IOO,�q/tube,4nM[3H]PAF ±20MM PAI�C18
FiG. 2. Time course of [3H]PAF binding in a whole uterine membranepreparation isolated at Day2 of pregnancy with 4 nM [3HIPAF and 20 pMPAF,C18 used to estimate nonspecific binding. Values represent the means ±SD of three independent experiments.
ENDOMETRIAL PLATELET-ACTIVA11NG FACTOR RECEPTORS 593
I [3H]PAF,CIS [I [3H]PAF,CI8IIG
FIG. 1. Comparative inhibition of the binding of [3H]PAF,C18 or[3H]PAF,C1�16 mixture by PAF,C18 and/or PAF,C16 in various combinations.The endomeirial membranes (100 �Lg/tube) were incubated at 25#{176}Cwith 4 nMof the labeled PAF for 150 mm. Nonspecific binding was estimated by additionof 20 pM of the competing ligands. Values are the means ± SD of three mdc-- c_.
in the labeled PAF was unspecified, so the ability of amixture of radioinert PAF,C16 and PAF,CI8 in varyingproportions (final concentration 20 jiM) to compete forboth the [3H]PAF mixture and the pure [3H]PAF,C18was compared. Figure 1 shows that the radioinertPAF,C18 was effective to the same extent in displacingeither the pure or mixed labeled PAF. These resultsvalidate the use of PAF,C18 for estimating nonspecificbinding of the mixed label used in the present studies.
Preliminary experiments were conducted to deter-mine the possible interference in the binding reactionscaused by micellar formation of the competing ligand at
high concentrations. The tune and order of addition ofthe 1 or 4 nM (3HIPAF and the 5 or 20 p.M concentra-tion of radioinert PAF as competitor were varied. Therewere no significant differences in the amount of spe-cific binding observed under these various conditions(data not shown). This indicates that (1) this 20-jAMconcentration did not reduce the amount of [3H]PAFable to interact with the membranes, (2) the interactionwas reversible, and (3) the results of the experimentsreported in this manuscript, where all reactants wereadded together, were measured under an equilibrium
steady state.
Time Course of �H]PAF 8inding mWhole Uterine Membranes, Day 2 Pregnancy
Figure 2 shows the time course of [3H]PAF bindingto uterine membranes isolated on Day 2 of pregnancyand indicates the presence of the binding entities at thistime, even though the rest of the study was conductedwith membranes isolated on Day 6, about 20 h beforeblastocyst implantation. At 4#{176}C,the uptake of [3HJPAFreached maximum binding after 90 mm and remained
stable for up to 3 Ii, but was completely lost after 18-24h (data not shown). Maximum binding was about30-40% of the highest equilibrium level achieved at25#{176}C(120-180 mm). This observation was similar inmembranes isolated on Day 6 (see Fig. 4). At 60#{176}C,maximum binding was achieved at 60 miii, representingabout 50-60% of the maximum binding at 25#{176}C,buthowever, the binding was highly unstable and wascompletely lost by 180 mm.
Estimation of the Affinity of
Binding Sites for (3HJPAF
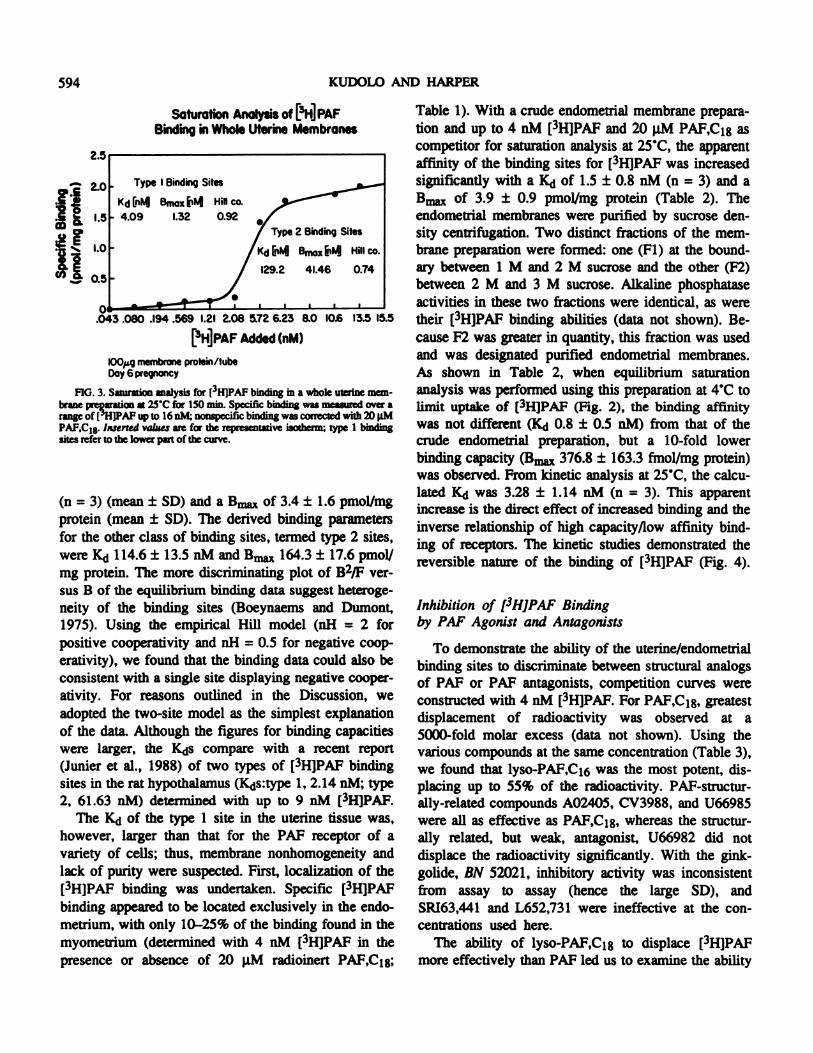
Saturation analysis was first conducted under optimalconditions for the measurement of maximum binding of[3H]PAF (25#{176}C,150 min), with up to 16 nM E3H]PAFand 20 p.M PAF,C18 used as competitor. Figure 3shows, for one experiment, a representative saturationisotherm exhibiting an apparent multiple-site bindingsystem. From a computer-assisted nonlinear regressionanalysis (Munson, 1987), we determined the simplestmultiple binding site system was two sites, with thehigher affinity binding site having a IQ of 3.6 ± 0.4 nM
2.5
� z.o Type I Binding Sites
1.5 Bn�ox�thfl Hill Co
.� Type 2 Bmding Sites
Bm�j�[nt�j
.043 .080 .194 .569 1.21 2.08 5�72 6.23 8.0 10.6 13.5 15.5
[3H]PAF Added (nM)
IOO�g membrane protein/tubeDoy6 pregnancy
FiG. 3. Saturation analysis for [3HJPAF binding in a whole uterine mem-brane preparation at 25#{176}Cfor 150 mm. Specific binding was measured over arange of (‘H)PAF up to l6nM; nonspecific binding was corrected with 20 pMPAF,C18. Inserted vahies are for the representative isotherm; type I bindingsites refer to the lower part of the curve.
594 KUDOLO AND HARPER
Saturation Analysis of [�H] PAFBinding in Whole Uterine Membranes
(n = 3) (mean ± SD) and a � of 3.4 ± 1.6 pmol/mgprotein (mean ± SD). The derived binding parameters
for the other class of binding sites, termed type 2 sites,were K<i 114.6 ± 13.5 nM and Bm� 164.3 ± 17.6 pmol/mg protein. The more discriminating plot of B2/F ver-sus B of the equilibrium binding data suggest heteroge-neity of the binding sites (Boeynaems and Dumont,1975). Using the empirical Hill model (nH = 2 forpositive cooperativity and nH = 0.5 for negative coop-erativity), we found that the binding data could also beconsistent with a single Site displaying negative cooper-ativity. For reasons outhned in the Discussion, weadopted the two-site model as the simplest explanationof the data. Although the figures for binding capacitieswere larger, the K(js compare with a recent report(Junier et al., 1988) of two types of [3H]PAF bindingsites in the rat hypothalamus (K4js:type 1, 2.14 nM; type2, 61.63 nM) determined with up to 9 nM [3H]PAF.
The K�j of the type 1 site in the uterine tissue was,however, larger than that for the PAF receptor of avariety of cells; thus, membrane nonhomogeneity andlack of purity were suspected. First, localization of the[3HIPAF binding was undertaken. Specific [3H]PAFbinding appeared to be located exclusively in the endo-metrium, with only 10-25% of the binding found in themyometrium (determined with 4 nM [3H]PAF in thepresence or absence of 20 p.M radioinert PAF,C18;
Table 1). With a crude endometrial membrane prepara-tion and up to 4 nM [3H]PAF and 20 p.M PAF,C18 ascompetitor for saturation analysis at 25#{176}C,the apparentaffinity of the binding sites for [3H]PAF was increasedsignificantly with a Li of 1.5 ± 0.8 nM (n = 3) and a� of 3.9 ± 0.9 pmol/mg protein (Table 2). Theendometrial membranes were purified by sucrose den-sity centrifugation. Two distinct fractions of the mem-brane preparation were formed: one (Fl) at the bound-ary between 1 M and 2 M sucrose and the other (F2)between 2 M and 3 M sucrose. Alkaline phosphatase
activities in these two fractions were identical, as weretheir [3H]PAF binding abilities (data not shown). Be-cause F2 was greater in quantity, this fraction was usedand was designated purified endometrial membranes.As shown in Table 2, when equilibrium saturationanalysis was performed using this preparation at 4#{176}Ctolimit uptake of [3H]PAF (Fig. 2), the binding affinitywas not different (K<j 0.8 ± 0.5 nM) from that of thecrude endometrial preparation, but a 10-fold lower
binding capacity (B� 376.8 ± 163.3 fmol/mg protein)was observed. From kinetic analysis at 25#{176}C,the calcu-lated Li was 3.28 ± 1.14 nM (n = 3). This apparentincrease is the direct effect of increased binding and theinverse relationship of high capacity/low affinity bind-ing of receptors. The kinetic studies demonstrated thereversible nature of the binding of [3H]PAF (Fig. 4).
Inhibition of [3HJPAF Binding
by PAF Agonist and Antagonists
To demonstrate the ability of the uterine/endometrialbinding sites to discriminate between structural analogsof PAF or PAF antagonists, competition curves wereconstructed with 4 nM [3HIPAF. For PAF,C18, greatestdisplacement of radioactivity was observed at a5000-fold molar excess (data not shown). Using thevarious compounds at the same concentration (Table 3),we found that 1yso-PAF,C1� was the most potent, dis-placing up to 55% of the radioactivity. PAF-structur-ally-related compounds A02405, CV3988, and U66985were all as effective as PAF,C18, whereas the structur-ally related, but weak, antagonist, U66982 did notdisplace the radioactivity significantly. With the gink-golide, BN 52021, inhibitory activity was inconsistentfrom assay to assay (hence the large SD), and
SR163,441 and L652,731 were ineffective at the con-
centrations used here.The ability of lyso-PAF,C18 to displace [3H]PAF
more effectively than PAP led us to examine the ability

Kinetic study of [3H] PAF-endometrial
membrane interaction
ENDOMETRIAL PLATELET-ACTIVATING FACTOR RECEPTORS
Association kinetic profile Dissociation kinetic profile
4-
�2.5
02E
5
� 0.5
595
/0
Comparative Disptacement of [�H] PAF and
[�H] Lyso-PAF in Uterine Membranes
‘C0E
C
-0.2
-0.4
-0.6
-0.8
�0 50 100 150 200
Time (minutes)0 10 20304050 60 70
Time (minutes)Competitors
C
lOO1�g/tube4nM label ±20MM PAI� C18
FIG. 5. Comparative ability of PAF,C18. lyso-PAF.C16. U66985. andU66982 (20 pM each) to displace 4 nM [3HIPAF0r [3}I]lyso-PAF from uterinemembranes (100 pg/tube) incubated at 25C for 150 min. The amount of mu-ated ligand displaced train the binding sites by the various competitors is ex-pressed as % bound total binding (no compctitoc)-noiispecific binding (pluscompetitor) + total binding x 100. Only PA1�Cj8 and lyso-PAF were able todisplace [‘H)lyso-PAF. U66985 displaced [311)PAF. but not (3H]tyso-PAF.while 1366982 had little activity against (3H)PAF and none against [3}l]lyso-PAF. Values are the means ± SD of at least three independent experiments.
4c�M?AF,CIe
550�
��:: (���200� o-?
0 50 100 ISO 200 250
Time (minutes)-0 - Dissoclotlon --Auoclatlon
FIG. 4. Kinetic study of (3H)PAF interaction with purified endometrialmembranes at 25#{176}C.The membranes were incubated with (3H]PAF(4 nM final
concentration) in the presence or absence of 20 pM radioinert PAF,C18. Axequilibrium (arrow), bound (3HJPAF was dissociated from the binding sateswith 40 1mM radioinert PAF.C18. Profiles shown are froni the satue repre-sentative experiment.
of PAF,C18 to displace [3H]lyso-PAF,C18. As shown inFigure 5, at equiinolar concentrations, [3H]PAF wasdisplaceable in this order of decreasing potency: lyso-PAF,C16, PAF,C18 = U66985, with U66982 showing nosignificant effect. [3H]Lyso-PAF,C18 was displaceableby both radioinert lyso-PAF,C16 and PAF,C18 but notby U66985 or U66982. Lyso-PAF,C16 was about 50%more effective than PAF,C18 in displacing [3H]lyso-PAF,C18.
Ionic Modulation of
(3H)PAF Binding
Ionic concentrations that might produce optimal
binding of [3H]PAF were examined. Whereas ��2+ and
Mg2� potentiated [3H]PAF binding in rabbit platelets,these divalent cations reduced binding by approxi-
mately 30% from control in the uterine membranes, andNa� reduced the binding by as much as 50% (Table 4).Li� and K� were even more detrimental to PAP bind-ing.
Of the protease inhibitors tested, only DFP andPMSF consistently inhibited labeling of the bindingsites. Bacitracin, antipain, and leupeptin all stabilizedand potentiated the binding, although, in isolated exper-
iments, [3H]PAF binding was inhibited-hence the largeSDs when expressed as percentage of control (Table 4).It therefore appeared that proteolysis of the binding sitewas not the only limiting factor for optimal binding.The other factor in the bimolecular second order reac-tion was the concentration of [3H]PAF during incuba-tion.
Metabolic Fate of (3H)PAF
during Incubation
Inevitably, the question of PAP metabolism underthe conditions of maximal [3H]PAF binding had to beaddressed. PAP is formed by the combined activities of
a phospholipase A2 and acetyltransferase (Vargaftig etal., 1981; Touqui et al., 1985). PAP inactivation in
596 KUDOLO AND HARPER
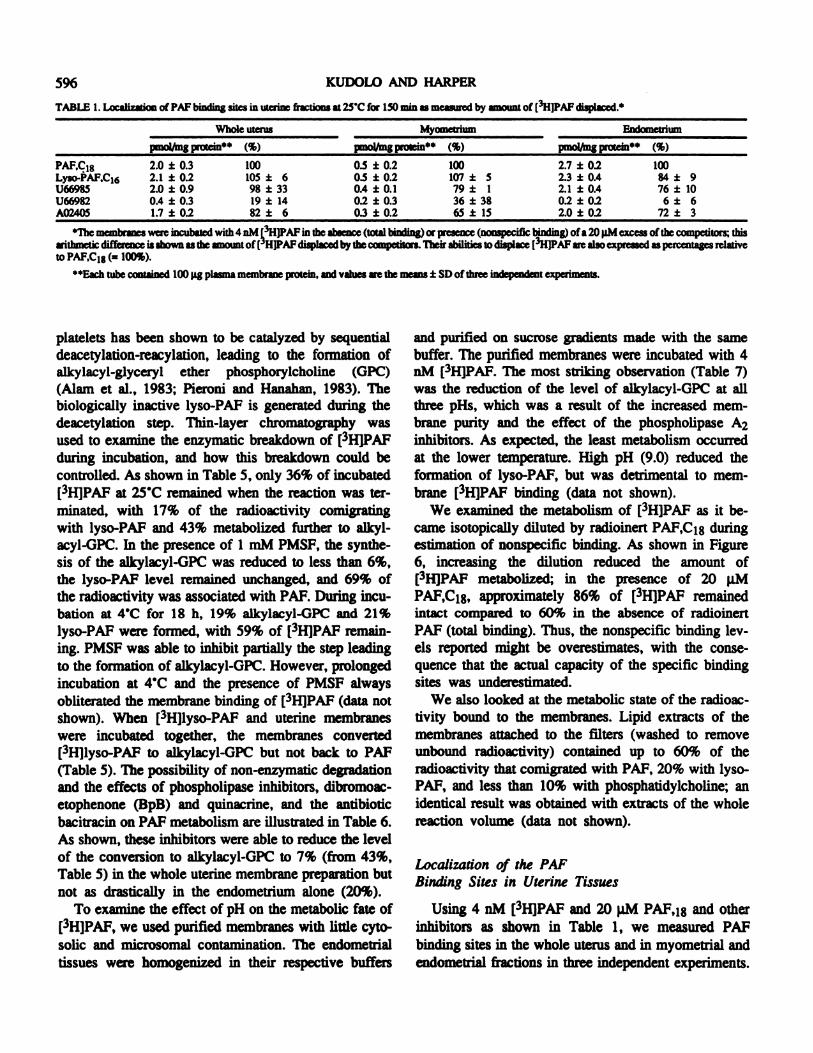
TABLE 1. Localization of PAF binding sites in uterine fractions at 25C for 150 mm as measured by amount of (3HWAF displaced.4
Whole uterus Myoinetnum Endomnettium
pmol/mg protein (%) pmo �/nmg pt�in44 (%) pm olftng prolein** (%)
PAF.C18Lyso-PAF,C161366985U66982A02405
2.02.12.00.41.7
± 0.3± 0.2± 0.9± 0.3± 0.2
100105981982
± 6± 33± 14± 6
0.50.50.40.20.3
± 0.2± 0.2± 0.1± 0.3± 0.2
100107 ±79 ±36 ±65 ±
51
3815
2.72.32.10.22.0
± 0.2± 0.4± 0.4± 0.2± 0.2
10084 ± 976 ± 10
6 ± 672 ± 3
4The membranes were incubated with 4 nM �H]PAFin the absence (total binding) or presence (nonspecific binding) of a 20 pM excess of the competitors; thisarithmetic difference is shown as the amount of( HJPAF displaced by the competitors. Their abilities to displace (3HIPAF are also expressed as percentages relativeto PAF,C18 (= 100%).
44Each tube contained 100 pg plasma membrane protein, and values are the means ± SD of three independent experiments
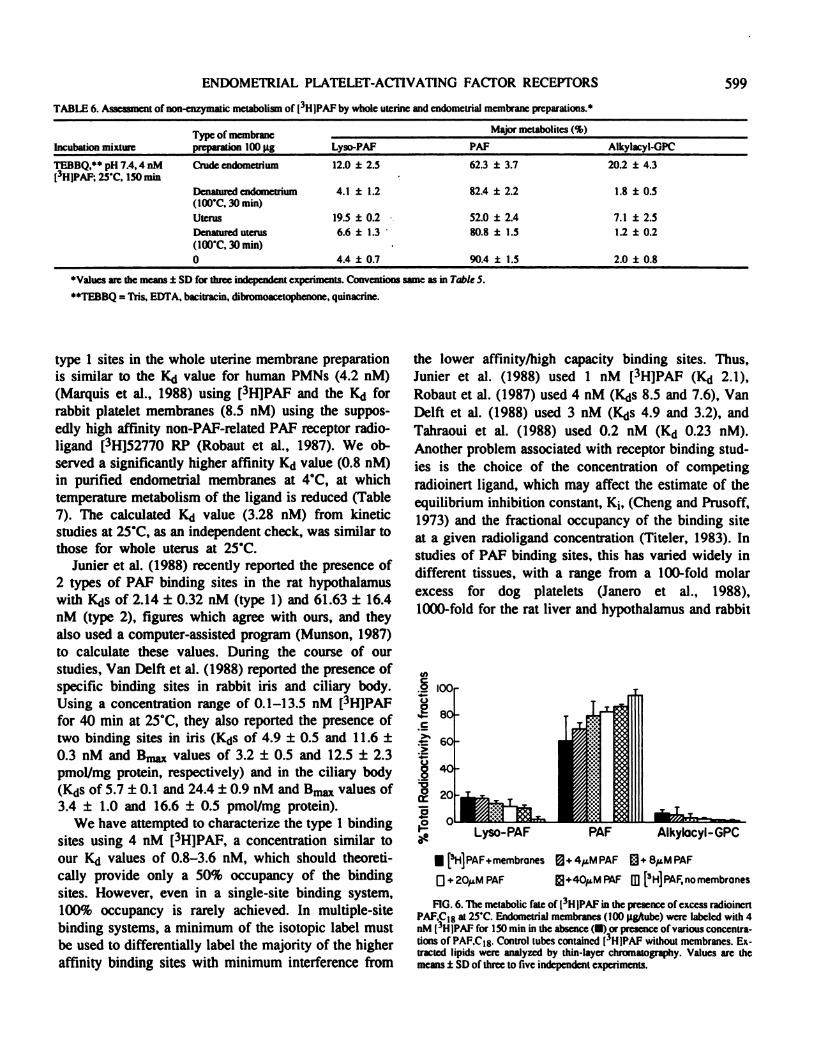
platelets has been shown to be catalyzed by sequentialdeacetylation-reacylation, leading to the fonnation ofalkylacyl-glyceryl ether phosphorylcholine (GPC)(Alam et al., 1983; Pieroni and Hanahan, 1983). Thebiologically inactive lyso-PAF is generated during thedeacetylation step. Thin-layer chromatography wasused to examine the enzymatic breakdown of [3HIPAFduring incubation, and how this breakdown could becontrolled. As shown in TableS, only 36% of incubated[3H]PAF at 25#{176}Cremained when the reaction was ter-
minated, with 17% of the radioactivity comigratingwith lyso-PAF and 43% metabolized further to alkyl-acyl-GPC. In the presence of 1 mM PMSF, the synthe-sis of the alkylacyl-GPC was reduced to less than 6%,the lyso-PAF level remained unchanged, and 69% ofthe radioactivity was associated with PAP. During incu-bation at 4#{176}Cfor 18 h, 19% alkylacyl-GPC and 21%lyso-PAP were formed, with 59% of [3H]PAF remain-ing. PMSF was able to inhibit partially the step leadingto the formation of alkylacyl-GPC. However, prolongedincubation at 4#{176}Cand the presence of PMSF alwaysobliterated the membrane binding of [3H}PAF (data notshown). When [3H]lyso-PAF and uterine membraneswere incubated together, the membranes converted[3H]lyso-PAF to alkylacyl-GPC but not back to PAP(Table 5). The possibility of non-enzymatic degradationand the effects of phospholipase inhibitors, dibromoac-etophenone (BpB) and quinacrine, and the antibioticbacitracm on PAP metabolism are illustrated in Table 6.As shown, these inhibitors were able to reduce the levelof the conversion to alkylacyl-GPC to 7% (from 43%,
Table 5) in the whole uterine membrane preparation butnot as drastically in the endometrium alone (20%).
To examine the effect of pH on the metabolic fate of[3H]PAF, we used purified membranes with little cyto-
solic and microsomal contamination. The endometrialtissues were homogenized in their respective buffers
and purified on sucrose gradients made with the samebuffer. The purified membranes were incubated with 4nM [3H]PAP. The most striking observation (Table 7)was the reduction of the level of alkylacyl-GPC at allthree pits, which was a result of the increased mem-brane purity and the effect of the phospholipase A2inhibitors. As expected, the least metabolism occurredat the lower temperature. High pH (9.0) reduced theformation of lyso-PAP, but was detrimental to mem-brane [3H]PAP binding (data not shown).
We examined the metabolism of [3HIPAF as it be-came isotopically diluted by radioinert PAF,C18 duringestimation of nonspecific binding. As shown in Figure6, increasing the dilution reduced the amount of[3H]PAF metabolized; in the presence of 20 p.MPAP,C18, approximately 86% of [3H]PAP remainedintact compared to 60% in the absence of radioinertPAP (total binding). Thus, the nonspecific binding lev-els reported might be overestimates, with the conse-quence that the actual capacity of the specific bindingsites was underestimated.
We also looked at the metabolic state of the radioac-tivity bound to the membranes. Lipid extracts of themembranes attached to the ifiters (washed to removeunbound radioactivity) contained up to 60% of theradioactivity that comigrated with PAP, 20% with lyso-PAP, and less than 10% with phosphatidylchollne; anidentical result was obtained with extracts of the wholereaction volume (data not shown).
Localization of the PAF
Binding Sites in Uterine Tissues
Using 4 nM [3H]PAP and 20 p�M PAP,18 and otherinhibitors as shown in Table 1, we measured PAP
binding sites in the whole uterus and in myometrial andendometrial fractions in three independent experiments.

ENDOMETRIAL PLATELET-ACTIVATING FACFOR RECEPTORS 597
TABLE 2. Effect of tern perature and membrane purity on the dissoc iation constant (K�J as determined by equilibrium saturation and kinetic analyses.*
Membrane Binding Lj (nM) B� (mean ± SD)preparation Method of analysis site (mean ± SD) pmolftng protein
Whole uterus Equilibrium saturation analysis[3HJPAF up to 16 nM; 25’C, 150 min
Type 1 3.6 ± 0.4’ 3.4 ± 1.61’
crude endomnetrium
Purified endometrium
Purified endometrium
Equilibrium saturation analysis[3H]PAFupto4nM; 25#{176}C,l5OminEquilibrium saturation analysis[3H]PAF up to 4 nM 4’C, 150 mmKinetic analysis�4 nM [3HJPAF; 25#{176}C,150 mm
Type 2Type!
Type 1
Type 1
114.6 ± 13.51.5 ± o.8�’
0.8 ± 0.5”
k�, (min�) = 0.03 ± 0.01k.4.1 (nM1.min1) = 4.14 ± 2.61 x 10�L1(min1)= 11.60 ± 2.60x103Kd(flM)(k_l/k+l)= 3.28 ± 1.14’
164.3 ± 17.63.9 ± 0.91’
0.377 ± 0#{149}163q
*Values are mean ± SI) of three independent experiments. Significance of differences between means was tested by analysis of variance and Student-Newman-
Keuls test (p<O.05) SAS Institute Inc., Cay, NC; letter superscripts indicate differences, a is different from b, andP from q�
“In this experiment, 40 pM radioinert PAF.C18 was used to dissociate the membrane-bound complexes formed at equilibrium with 4 nM [3HIPAF in the
absence or presence of 20 pM radioincrt PAF,C18.
Specific PAP binding appeared to be located solely inthe endometrium. In the endometrial membrane prepa-ration, lyso-PAP,C1� apparently competed for the samepopulation of sites as PAP,C18. This suggests that somelyso-PAP,C16 binding sites could be unique to themyometrium, and that in the endometrium, lyso-PAP,C16 actually binds only to PAP binding sites. ThePAP receptor antagonists, U66985 and A02405, bothcompeted for the PAP binding sites in the endometri-urn, but U66982 (a less active structural analog) did not(Table 1).
DISCUSSION
To our knowledge, these radioligand binding studies
document the first direct evidence of PAP binding sitesin the maternal reproductive tract. Endogenous PAP,whether embryo- or uterine-derived, may act by occu-pation of such sites. Indirect evidence to support recep-
tor-mediated actions of PAP may be inferred from theability of PAP antagonists to interrupt early pregnancyin rodents (Spinks and O’Neill, 1987, 1988; Acker etal., 1988).
PAP binding was observed in the uterus at both Day2 and Day 6 of pregnancy. In a pilot study, PAPbinding sites with similar affmities were demonstratedin uteri from estrous, Day 6 pseudopregnant, and Day 2and Day 6 pregnant animals (Kudolo and Harper,1988). In the present study, data were obtained predom-inantly from uteri on Day 6 of pregnancy, first, because
the tissues were collected on an opportunistic basis
after embryo collection for another study, and second,
because uterine PAP levels have been shown to de-
crease dramatically during Day 6 of pregnancy (Angleet al., 1988b), and thus there may be reduced occupa-tion of binding sites. In this study, similarities betweenDay 2 and Day 6 uterine/endornetrial PAP bindingparameters were limited to the concentration and integ-rity of the PAP binding sites and the optimal bindingconditions (time course) only. It is possible thatchanges in affinity and capacity of the binding sitesmay occur during the intervening period of gestation(Days 2-6), as may the activity of the PAP-metaboliz-ing enzymes. Consequently, similarities between theseattributes at different stages of pregnancy cannot beinferred.
Saturation isotherms and nonlinear regression analy-ses may not be adequate to distinguish between amultiple-site binding model and one with a single site
TABLE 3. The relative potency of PAF and related compounds at 20 pM ininhibiting 4 nM (3}IIPAF binding to uterine membranes after incubation at25’Cfor 150mm.
Compound % [3H] PAF displacement’
PAF,C18 44.1 ± 10.5 (n = 20)Lyso-PAF,C16 54.6 ± 10.3 (n = 13)A02405 38.7 ± 4.7 (n = 3)U66985 46.9 ± 9.8 (n = 7)U66982 9.1 ± 9.1 (n = 7)BN52021 12.7 ± 19.0 (n= 3)CV3988 51.4 ± 2.8 (n= 3)SR163.441 0 (n= 3)L652,731 0 (n= 3)
‘The membrane-bound [3H]PAF in the absence of competitor is normal-ized to 100%, and the binding in the presence of the competitors is expressedrelative to 100%; thus, figures greater than 44.1% (PAF,C18) are at least aseffective as PAF,C18 in displacing [314]PAF from its binding sites. Values arethe means ± SD and the number of independent experiments is shown in paren-theses.

598 KUDOLO AND HARPER
TABLE 4. Effect of cauons and protease inhibitors on (3HIPAF binding at 25C for 150 mm.’
pmo�gproteinT
% ofcontro I bound
TEM� 2.5 ± 1.0 100+ CaCI2 (2 mM) 1.8 ± 0.9 69.5 ± 8.1+MgCI2(IOmM) 1.7 ± 0.9 63.6 ± 8.3+ LiCI (10 mM) 0.9 ± 0.3 35.6 ± 2.7+NaCl (150 mM) 1.2 ± 0.2 53.2 ± 2.7+KCI (150 mM) 0.7 ± 0.1 30.2 ± 1.3+ PMSF(2 mM) 0.1 ± 0.04 5.9 ± 3.5+Baciuacin (150 pM) 2.7 ± 0.3 111.7 ± 27.0
+ Leupeptin (150 pM) 2.4 ± 0.02 102.8 ± 33.8
+ DFP (150 pM) 1.3 ± 0.4 51.8 ± 9.4
+Antipain (150 pM) 2.4 ± 0.3 107.7 ± 45.7
‘Values are the means ± SD of three independent experiments
tThM = Iris, EDTA, monothioglycerol buffer.
�The value is the amount of specifically bound (3HIPAF determined in the presence or absence of a 20 pM PAF,C18 when incubated with 4 nM I3HIPAF.
displaying cooperativity. In our system, both heteroge-
neity of binding and negative cooperativity can bepresumed (Teipel and Koshland, 1969). Because themacromolecular nature of the endometrial membranePAP binding sites has not been defined, we opted forthe two-site model because of its relative simplicity;however, we have not excluded the possibility that thetype 2 sites may be modified type I sites on a poly-meric macromolecule or a portion of loosely aggregatedmoieties. Heterogeneity in our system may also be dueto discrete lyso-PAF binding sites, which bind bothlyso-PAF and PAP. The saturation isotherm shown inFigure 3 supports the concept of negative cooperativity;if this model is adopted, the type 2 sites could be due toa conformational change of the membrane, with addi-
tional sites exposed but with PAP binding affinity de-creased. Resolution of the reason for the binding heter-ogeneity awaits further experimentation.
The uterine/endometrial plasma membrane bindingdata presented here meet several criteria for classifyingthe type I binding sites as specific, high-affinity recep-tors (Laduron, 1984). Through progressive membranepurification, the Kd was found to be comparable withthose reported for other PAP receptor systems such asrabbit platelets (Hwang et aL, 1986b; Homma et al.,1987), human platelets (Hwang and Lam, 1986), humanlung tissue (Hwang et al., 1985), human polymorphonu-clear neutrophils (Hwang, 1988), rat liver (Hwang,1987), and canine platelet membranes (Tahraoui et al.,1988). First, our reported K<j value (3.6 nM) for the
TABLE 5. Metabolism of I3HIPAF or I3HlIyso-PAF by uterine membranes. ‘
Membrane preparationand incubation mixture
Treatment
(1 mM)
Membrane �tein
content/tube (pg)
Major metaholites (%)t
Lyso-PAF PAF Alkylacyl-GPC
Whole uterus TEM.� pH 7.44nMI3HJPAF;25’C, 150mm
-PMSF#{247}PMSF
100
100
0
16.920.5
1.3
± 2.5± 2.2± 0.4
36.3 ± 0.869.4 ± 1.696.6 ± 0.6
42.6 ± 2.15.5 ± 1.20.8 ± 0.2
4’C, 18 h -PMSF+PMSF
100100
0
20.623.4
1.3
± 9.1± 1.8
± 0.4
58.6 ± 12.361.6 ± 1.8
96.1 ± 0.8
18.5 ± 3.29.4 ± 2.1
1.0 ± 0.4
Whole uterus TEM, pH 7.44nMl3HJIyso-PAF; 25’C. 150 mm
-PMSF+PMSF
100
1000
29.358.788.5
± 6.1±2.1± 2.2
3.1 ± 1.25.8 ± 0.9
6.9 ± 1.8
47.2 ± 9.4
10.7 ± 1.70.9 ± 0.1
4’C, 18 h -PMSF+PMSF
100100
0
44.850.690.9
± 4.3± 2.2± 1.2
2.9 ± 0.24.7 ± 1.45.0 ± 1.4
46.4 ± 14.2
29.7 ± 3.20.5 ± 0.2
‘Membranes were incubated as indicated. Values are the mean ± SD of three independent experiments.
tfl� major metabolites were separated by TLC and their percentages (corrected for recovery) were calculated relative to the total radioactivity loaded on the
silica plate prior to chromatography.
tTEM = Tris, EDTA. monothioglycerol.
Lyso-PAF PAF Alkylacyl-GPC
ENDOMETRIAL PLATELET-AC1’IVATING FACfOR RECEPTORS 599
TABLE 6. Assessment of non-enzymatic metabolism of L3HIPAF by whole uterine and endometnal membrane preparations.’
Incubation mixtureType of membranepreparation 100 pg
M ajor metabolites (%)
Lyso-PAF PAF Alkylacyl-GPC
TEBBQ,” pH 7.4,4 nM
(3HIPAF; 25�C, 150 mm
Crude endometrium 12.0 ± 2.5 62.3 ± 3.7 20.2 ± 4.3
Denatured endosnetrium(100’C, 30 mm)
Uterus
Denatured uterus(100’C, 30 mm)
0
4.1
19.5
6.6
4.4
± 1.2
± 0.2
± 1.3
± 0.7
82.4 ± 2.2
52.0 ± 2.4
80.8 ± 1.5
90.4 ± 1.5
1.8
7.1
1.2
2.0
± 0.5
± 2.5
± 0.2
± 0.8
‘Values are the means ± SD for three independent experiments. Conventions same as in Table 5.
“TEBBQ = Tris. EDTA, bacitracin, dibromoacetophenone, quinacrine.
type 1 sites in the whole uterine membrane preparationis similar to the K�J value for human PMNs (4.2 nM)(Marquis et al., 1988) using [3H]PAF and the K�J forrabbit platelet membranes (8.5 nM) using the suppos-edly high affinity non-PAP-related PAP receptor radio-ligand [3H]52770 RP (Robaut Ct al., 1987). We ob-served a significantly higher affinity Kd value (0.8 nM)in purified endometrial membranes at 4C, at whichtemperature metabolism of the ligand is reduced (Table7). The calculated K1j value (3.28 nM) from kineticstudies at 25C, as an independent check, was similar tothose for whole uterus at 25’C.
Junier et al. (1988) recently reported the presence of2 types of PAP binding sites in the rat hypothalamuswith K<js of 2.14 ± 0.32 nM (type I) and 61.63 ± 16.4nM (type 2), figures which agree with ours, and theyalso used a computer-assisted program (Munson, 1987)to calculate these values. During the course of ourstudies, Van Delft et al. (1988) reported the presence ofspecific binding sites in rabbit iris and ciliary body.Using a concentration range of 0.1-13.5 nM [3H]PAFfor 40 mm at 25’C, they also reported the presence oftwo binding sites in iris (KdS of 4.9 ± 0.5 and 11.6 ±0.3 nM and Bn� values of 3.2 ± 0.5 and 12.5 ± 2.3pmol/mg protein, respectively) and in the ciliary body(KdS of 5.7 ± 0.1 and 24.4 ± 0.9 nM and Bm� values of3.4 ± 1.0 and 16.6 ± 0.5 pmol/mg protein).
We have attempted to characterize the type I bindingsites using 4 nM [3H]PAF, a concentration similar toour Kd values of 0.8-3.6 nM, which should theoreti-cally provide only a 50% occupancy of the bindingsites. However, even in a single-site binding system,100% occupancy is rarely achieved. In multiple-sitebinding systems, a minimum of the isotopic label mustbe used to differentially label the majority of the higheraffinity binding sites with minimum interference from
the lower affinity/high capacity binding sites. Thus,Junier et al. (1988) used I nM [3H]PAF (Kd 2.1),Robaut et al. (1987) used 4 nM (K#{128}js8.5 and 7.6), VanDelft et al. (1988) used 3 nM (Kds 4.9 and 3.2), and
Tahraoui et al. (1988) used 0.2 nM (Kd 0.23 nM).Another problem associated with receptor binding stud-
ies is the choice of the concentration of competingradioinert ligand, which may affect the estimate of theequilibrium inhibition constant, K1, (Cheng and Prusoff,1973) and the fractional occupancy of the binding siteat a given radioligand concentration (Titeler, 1983). Instudies of PAP binding sites, this has varied widely indifferent tissues, with a range from a 100-fold molar
excess for dog platelets (Janero et al., 1988),1000-fold for the rat liver and hypothalamus and rabbit
U,C0
U
2C
>‘
>
U
80
0
U{�H]PAF+membrones D+4�MPAF D+8�MPAF
D+20,LM PAF �+4O�1MPAF m [3H]PAF,nomembrones
FIG. 6. The metabolic fate of (3H IPAF in the presence of excess radioincrt
PAF,C18 at 25’C. Endometrial membranes (100 pg/tube) were labeled with 4
nM (3HIPAF for 150 mm in the absence (#{149})or presence of various concentra-
tions of PAF,C18. Control tubes contained I3HIPAF without membranes. Ex-tracted lipids were analyzed by thin-layer chromatography. Values arc themeans ± SD of three to five independent experiments.

Incubation bufferand pH
TEBBQ.” 6.0
TEBBQ. 7.4
TEBBQ. 9.0
Temperature
�#{176}‘�‘�
(pg)
Major melabolites (%)
Lyso-PAF PAF Alkylacyl-GPC
4’C25�C
100100
14.7 ± 2.236.6 ± 0.6
79.8 ± 2.754.4 ± 1.3
2.63.5
± 1.3± 1.1
4’C25�C
5050
15.5 ± 2.237.9 ± 1.5
80.0 ± 2.254.3 ± 1.9
1.52.1.
± 0,2± 0.7
4C25t
00
3.9 ± 1.94.7 ± 1.2
89.6 ± 2.891.5 ± 2.7
1.51.4
± 0.4
± 0.64’C
25’C100100
14.6 ± 1.720.5 ± 2.3
79.3 ± 5.155.9 ± 8.6
2.47.0
± 1.1± 2.7
4�C25�C
5050
13.5 ± 1.520.9 ± 2.3
78.4 ± 2.861.0 ± 6.2
2.99.8
± 1.7± 2.6
4’C25C
00
4.1 ± 0.34.4 ± 0.1
89.5 ± 1.189.8 ± 2.4
2.63.1
± 0.5± 1.4
4C25C
100100
6.1 ± 2.8
11.6 ± 1.891.8 ± 2.972.7 ± 2.7
2.08.4
± 0.6± 0.9
4’C25’C
5050
3.4 ± 1.09.6 ± 1.7
87.6 ± 1.476.3 ± 1.8
2.57.7
± 13± 1.1
4C25C
00
2.6 ± 0.42.9 ± 0.9
94.5 ± 1.192.5 ± 2.6
1.83.2
± 0.6± 1.3
600 KUDOLO AND HARPER
TABLE 7. Effect of pH on 4 nM (3HJPAF metabolism by punfled endometrial membranes.
‘Values are the mean ± SD of three independent experiments conducted at the indicated temperatures for 150 nile. Conventions same as in Table 5.
“TEBBQ = Tris, EIYrA, bacitracin, dibromoacelophenone, quinacrmne.
iris and ciliary body (Hwang, 1987; Jurner et al., 1988;
Van Delft et al., 1988), 2500-fold for rabbit platelet(Robaut et al., 1987), to 50,000-fold in another canine
platelet membrane study (Tahraoui et al., 1988).In the present study, a 5000-fold molar excess was
required to displace up to 90% of the radioactivitybound to 100 j.tg of whole uterine membrane protein. Inpurified endometrial membrane preparations, however,a 1000-fold molar excess was adequate to achieve thesame level of [3H]PAP inhibition. These results con-cerning radioligand and competitor concentrations high-light problems that still exist with PAP binding systems
and indicate that PAP binding data from across species
and across tissues systems must be compared and/orinterpreted carefully, especially when species, specific-ity and tissue-specificity are involved. This point isexemplified by other lines of evidence. It would appear
that no single ion, mono- or divalent, has produced the
same effect in the PAP receptor systems studied to date
(Hwang et aL, 1986b; Hwang, 1987, 1988). In wholeuterine membranes, all of the cations tested reducedspecific binding of [3H]PAP. For example (Hwang,1988), Li� inhibited [3H]PAP binding to human PMNswith no potentiating or inhibiting effect on humanplatelet membranes; K’ potentiated [3H]PAP binding in
rabbit platelets (Hwang, 1988), but inhibited specific[3H]PAP binding by 70% in rabbit uterine membrane
(present study). Such tissue-specificity and species-
specificity were original reasons for postulating a multi-plicity of PAP receptor subtypes (Hwang and Lam,
1986; Lambrecht and Parnham, 1986), and results ofthe present study support this strongly.
Another criterion for assessing the specificity ofligand binding involves using analogs or antagonists tocompete for the binding sites. The proposed existenceof PAP receptor subtypes has also been based on thedifferent potency of the PAP antagonist, kadsurenone,in inhibiting PAP-induced chemiluminescence ofguinea pig peritoneal macrophages and of aggregation
of pig peripheral blood leukocytes (Lambrecht andPamham, 1986). Both L652,731 and kadsurenone canfully displace [3H}PAP in the rat liver membrane prepa-
ration (Hwang, 1987). In the rabbit uterine membranepreparation, kadsurenone displaced [3H]PAF only mar-ginally (data not shown) and L652,731 was ineffective.L652,731, however, can displace [3H]PAP in human
PMNs (Hwang, 1988), exhibiting species difference inpotency as reported earlier (Hwang and Lam, 1986).Similarly BN52021 is unable to displace [3H]PAP inthe uterine membrane as effectively as it does in humanplatelets (Braquet et al., 1985; Korth and Benveniste,
1987). Some antagonists, such as those structurally
related to PAP, like CV3988 and U66985, can displace[3H]PAP in both the rat liver (Hwang, 1987) and rabbit

ENDOMETRIAL PLATELET-ACTIVATING FACTOR RECEPTORS 601
platelet (Homma et al., 1987) membrane preparations,as they do in rabbit uterine membranes.
Specificity of ligand binding has also been evaluatedby the inability of biologically inactive enantiomers ormetabolites to displace [3H]PAP from the PAP bindingsites (Homma et al., 1987; Hwang, 1987). Contrary toall previous studies, we report that the biologicallyinactive metabolite of PAP, lyso-PAP, displaced[3H]PAF from the PAP binding sites in the uterinemembrane. We believe that the ability of lyso-PAP todisplace PAP from PAP binding sites in the uterus doesnot necessarily suggest that these sites are nonspecific.
To answer that question, uterine membrane preparationswere labeled with [3H]lyso-PAP, which was displace-able by radioinert PAF,C18 (80% compared to lyso-PAP,C18 = 100%); this implies that at least 80% of thelyso-PAF-displaceable sites, when occupied by[3HJPAF, may be authentic PAP binding sites. Further-more, even though U66985 is able to displace [3H]PAP,it is unable to displace [3H]lyso-PAF. This exemplifiesthe different levels of discrimination of the PAP bind-ing sites for ligands (i.e. the specificity of the bindingsites). The ability of these ligands to be displaced, oncebound, and the reversible nature of the binding betweenPAP and lyso-PAP in the uterus are attributes that mayhave potential physiological significance. Because lyso-PAP is biologically inactive, its ability to occupy theauthentic PAP sites may desensitize the uterus to PAPactivity. The validity of this suggestion remains to betested in a bioassay. However, a choline-containinglysoglycerophospholipid has recently been identified asone of the two endogenous inhibitors in the rat uterusthat inhibit PAP-induced aggregation of washed rabbitplatelets (Nakayama et al., 1987). It is possible that thislyso-compound acts by binding to the PAP receptor:this would support our suggestion. Tokumura et al.(1985) found that at equivalent concentrations (3 lLM)both CV3988 and lyso-PAF can prevent PAP-inducedplatelet aggregation. In uterine membranes, these twocompounds and U66985 displace [3H]PAF to an equalextent. CV3988 has been claimed to be a specific PAPreceptor antagonist in rat liver membranes (Hwang,1987) and U66985 in rabbit platelets (Homma et al.,1987). The ability of U66985 to displace [3H]PAP, butnot [3H]lyso-PAP, from the uterine membrane bindingsites may point to the molecular characteristics of thebinding sites themselves and the production of sterichindrance by bound ligands. The PAP binding sites,
however, do not significantly bind the less active ana-log U66982. This steric hindrance may, however, be
limited to substitutions on the sn carbon 3, rather thanOfl sn carbon 1, particularly the distance between thephosphate and choline moieties. In PAP itself, only 2methylenes are in the same position. Thus, U66982,with 10 methylenes, is far less effective than U66985,which has only 6. This supposition is also supported bythe similar abilities of PAF,C16, PAP,C18, or a mixtureto displace either pure [3H]PAP,C18 or the [3H]PAF,C1W16 mixture.
The incubation conditions at 25#{176}Cfor maximumspecific binding of [3H]PAF obviously led to metabolicdegradation of added ligands. We have highlighted theproblems associated with PAP metabolism at this tem-perature, especially since bioassays for PAP actions on,for example, effector enzymes may have to be con-ducted at elevated temperatures. We have not totallyovercome this problem, but up to 70% of added[3H]PAP remained unchanged at the end of the incuba-tion period and the concentration of the [3H]lyso-PAFproduct was only about 20% when phospholipase inhib-itors were used. We have also established that onlyminimal conversion of the generated alkylacyl-GPC toPAP took place. We have also tried to discern thenature of the radioactivity associated with the mem-brane. Our results show that the membrane-bound fil-ters had essentially the same proportion of the PAP andits metabolites as did the extracts of the whole reactionvolume. Thus, at least 50-60% of membrane-bound[3H]PAP remained unmetabolized, and it can be sur-mised that the phospholipid was metabolized after asso-ciation with the PAP binding sites. Further reduction ofthe [3H]PAP metabolism with increased concentration
of the enzyme inhibitors was not possible without sacri-ficing specific [3H]PAP binding. We have not exam-ined the effect of cations on [3H]PAF metabolism, since
they all inhibited membrane [3HIPAP binding. Onestriking feature observed during this study was that adecrease in the amount of alkylacyl-GPC formed was
paralleled by a reduction in the nonspecific uptake of[3H]PAF by the membranes (from 70% to 40-50%,
when this was estimated with 20 �.tM PAP,C18). Wealso found that a greater percentage of the labeledligand remained unmetabolized in the presence of thecompetitor; thus, the level of nonspecific binding wasoverestimated.
From the present data, we believe that the PAPbinding sites represent the PAP receptor, and that thesemay be located solely in the endometrium. In the morehomogeneous and purified preparation incubated at4#{176}C,the PAP receptor displayed a higher affinity (K�j =

602 KUDOLO AND HARPER
0.8 nM). The significance of the second class of bind-ing sites is not clear, but it seems that these sites maybe produced through an allosteric conformation of thetype 1 receptors in the membrane bilayer. The presenceof a putative second PAP receptor has also been re-ported in human PMNs, but no affinity binding datahave been presented (Hwang, 1988), and in rat hypo-thalamus (Junier et al., 1988) and rabbit iris and ciliarybody (Van Deift et al., 1988), with which there isagreement in the present studies for both type 1 and 2receptors. The K�j reported here for the higher affinityPAP binding sites is also Consistent with value reportedfor rabbit and human platelets and rat liver (Hwang andLam, 1986; Hwang, 1987; Korth and Benveniste, 1987;Robaut et al., 1987). It can be envisaged that as the type1 sites fill, membrane integrity may be compromised.This could then lead to exposure of adjacent, but hith-erto masked, binding sites. These sites may be involvedin either signal amplification or production of refractoryconditions. The high binding capacity probably ensuresthat this ceiling is not reached under normal physiologi-cal conditions.
As a further check on the specificity of the [3H]PAPbinding, we have also examined in preliminary experi-ments the ability of a biologically potent non-metabolizable tritiated methylcarbamyl analog of PAP(CPAF) (O’Flaherty et al., 1987) to bind to the endome-trial membranes. CPAF has been shown to competespecifically for PAP receptors in human PMNs with anapparent K�j of 1.1 nM (O’Flaherty et al., 1987). Withpurified endometrial membranes, we observed an ap-parent K�J for a single class of binding site of 0.49 nMand a B� value of 1.4 pmol/mg protein, when bindingstudies were conducted with up to 20 nM of [3H]CPAP.These values are in good agreement with those obtainedusing [3HJPAP. It should be noted that, since CPAP hasonly approximately one-fifth the activity of PAP, thesecond class of binding sites would not be revealedover the concentration range tested. We have also ob-served the displacement of [3H]CPAF by lyso-PAP,which confirms our suggestion that lyso-PAP may havea physiological role in the uterus. At this time, howev-er, we have no evidence to suggest the presence ofdistinct lyso-PAP binding sites.
In conclusion, a high affinity PAP binding site inrabbit endometrium has been described. Under optimal
conditions, less than 20% of the [3H]PAP is convertedto [3Hjlyso-PAF, and lyso-PAF binds to PAP bindingsites; therefore, we do not feel that the derived bindingparameters are overestimated. The tissue/species specif-
icily displayed by the PAP receptor may permit themolecular nature of the receptor to be studied. BecausePAP is produced by a variety of cells, a highly selectiveantagonist that can act only in the uterus must be foundin order to exploit the characteristics of the uterine PAPreceptor.
ACKNOWLEDGMENTS
Tntiazed 1-O-alkyl-2 - N - methylcarbamyl - glycerophosphocholine([3HJCPAF) was a gift from Dr. Robert Wykie, Department of Biochemistry,Wake Forest University Medical Center, Winston Salem, NC.
Thanks are due to Drs. Marjesie Jones, Donna S. Woodard. and Linda Mc-Mantis for helpful discussions and to Beverly Evans for typing the manuscript.
REFERENCES
Ackcr 0, Hccquct F, Etienne A, Braquet P. Mencia-Huena JM� 1988. Role of
platelet-activazing factor (PAP) in the ovoimplantalion in the rat: effect of
the specific PAF-acether antagonist, BN52021. Prostaglandins 35:233-41
Alam 1, Smith 10, Silver MJ, 1983. Metabolism of platelet activating factor byblood platelets and plasma. Lipids 18:534-38
Alecozay AA, DeLeon PD, Harper MIX, Nouchi 1, Hanahan Di, 1988. Plate-let-activating factor (PAP) inhuman endometrium. Biol Reprod 38 (Suppl1):78 (Abstr. 90)
Angle MJ, Byrd W, Johnston JM, 1988a. Embryonic production of platelet-activating factor in culture. Fertil Steel 44th Ann Meeting Program: S96(Absir. P-158)
Angle MJ, Jones MA, McManus LM� Pinckard RN, Harper MJK, 1988b. Plate-let-activating factor in the rabbit uterus during early pregnancy. 1 ReprodFertil 83:711-22
Angle MI, Jones MA, Piuckard RN, McManus LM, Harper MIX, 1985. Plate-let-activating factor (PAP) in the rabbit uterus during early pregnancy.Biol Reprod 32 (Suppl. 1):193 (Absir. 212)
Beencu ii’ Jr. Yamamura HI, 1985. Neuroirannniuer. hormone or drug recep-tor binding. In: Yamamura HI, Enna SJ, Kuhar MI (eds.). Neurotransmit-tee Receptor Bindin& New York: Raven Press, pp. 61-89
Biggers JD, Lconov BY. Baskar W, Fried J, 1978. Inhibition of hatching of
mouse blastocysts in vitro by prostaglandin antagonists. Biol Reprod 19:519-33
Bligh EG, Dyer WI, 1959. A rapid method of total lipid cztraction and purifica-tion. Can J Biochem Physiol 37911-17
Boeynaems 3M, Dumont SE, 1975. Quantitative analysis of the binding of theligands to their receptors. 3 Cyclic Nucleolide Real: 123-42
Braquet P. Etienne A, aesire F, 1985. Down-regulation of �-adrenergic re-ceptors by PAF-acether and its inhibition by the PAF-acether antagonistBN52(Y21. Proetaglandins 30:721 (Absir.)
Braquet P. Godfroid ii, 1986. PAF-accther specific binding sites: 2. Design ofspecific antagonists. Trends Pliaramacol Sci:7:397-403
Braquct P. Touqui L, Shen Y, Vargaftig BB, 1987. Perspectives in platelet-activating factor research. Phannacol Rev 39:97-145
(leng Y, PrusoffWH, 1973. Relationship between the inhibition constant (K�)and the concentration of inhibitor which causes 50 per cent inhibition(lC�) of an enzymatic reaction. Biochein Pharmacol 22:3099-3108
Domingo MT. Spinnewyn B, (labeler PE, Braquet P. 1989. Presence of spe-cific binding sites for platelet-activating factor (PAP) in brain. BiochemBiophys Rca Commun 151:730-36
Evans CA. Kennedy TO, 1978. The importance of peostaglandin synthesis forthe initiation of blastocyst implantation in the hamatee. I Reprod Fertil 54:255-61
Godfroid II, Braquet P. 1986. PAF-acether specific binding sites: 1. Quantita-tive SAR study of PAF-acether isoscres. Trends Pharmnacol Sd 7:368-96
Homma H, Tokumura A, Hanshm Di, 1987. Binding and internalization ofplatelet-activating factor l-O-alkyl-2-acctyl-sn-glycero-3-phosphocho-line in washed rabbit platelet. 3 Biol (lean 262:10582-87
Hwang S-B, 1981. Specific receptor sites for platelet activating factor on ratliver plasma membranes. Arch Biochim Biophys 257:339-44

ENDOMETRIAL PLATELET-ACTIVATING FACTOR RECEPTORS
Hwang S-B, 1988. Identification of a second putative receptor of platelet-acti-vating factor from human polymorphonuclear leukocytes. J Biol (1cm
263:3225-33
Hwang S-B, Lam M-H, 1986. Species difference in the specific receptors ofplatelet-activating factor. Biochem Pharmacol 35:4511-18
Hwang S-B, Lain M-H, (lang MN, 1986a. Specific binding of [3H]dihydro-kadsurenone to rabbit platelet membranes and its inhibition by the recep-tor agonists and antagonists of platelet-activating factor. I Biol (1cm 261:13720-26
Hwang S-B, Lam M-H, Pang SS, 19861,. Ionic and GTP regulation of binding ofplatelet-activating factor to receptors and platelet-activating factor-in-duced activation of GTPase in rabbit platelet membranes. I Biol Clew261:532-37
Hwang S-B, Lam M-H, Slim TY, 1985. Specific binding sites for platelet-activating factor in human lung tissue. Biochem Biophys Res Coanmun128:972-79
Janero D, Burghardt B, Burghardt C, 1988. Specific binding of 1-O-alkyl-2-acetyl-sn-glycero-3- phosphocholine (platelet-activating factor) to theintact canine platelet Thromb Rca 50:789402
Jones MA, Can Z-d, Anderson W, Norris C, Harper MJK, 1986. Capillarypermeability changes in the uteri of recipient rabbits after transfer of blast-ocysts from indomethacin-trealed donors. J Reprod Fenil 78:261-73
Jwiier MP, Tiberghicn C, Rougeot C, Fafeur V. Dray F, 1988. Inhibitory effectof platelet-activating factor (PAP) on luteinizing hormone-releasing hor-mone and somatostatin release from rat median eminence in vitro corre-lated with the characterization of specific PAP receptor sites in rat hypo-thalamus. Endocrinology 123:72-80
Korth R, Benveniste 1,1987. BN52021 displaces [3HJPAF-acether from, andinhibits its binding to intact human platelets. Eur 3 Pharmacol 142:331-41
Kudolo GB, Harper MIX, 1988. Binding parameters of rabbit uterine PAP-receptors: pilot study. Biol Reprod 38 (Suppl 1):153 (Abstr. 314)
Laduron PM, 1984. Criteria for receptor sites in binding studies. Biochem Thar-macol 33 :833-39
Lambrecht 0, Parnham MI, 1986. Kadsurenone distinguishes between differ-ent platelet-activating factor receptor subtypes on macrophages and poly-morphonuclear leukocytes. Br J Pharmacol 87:287-89
Lampelo SA, Rickeits AP, Bullock DW, 1985. Purification of rabbit endome-trial plasma membranes from receptive and nonrecepsive uteri. 3 ReprodFertil 75:475-84
Lau IF, Saksena SK, Chang MC, 1973. Pregnancy blockade by indomethacin,an inhibitor of proetaglandin synthesis: its reversal by prostaglalalins andprogesterone in mice. Prostaglandins 4:795-803
Marquis 0, Robaut C, Cavern I, 1988. (3H]52770 RP, a platelet-activating fac-tor receptor antagonist, and tritiated platelet-activating factor label a coin-mon specific binding site in human polymorphonuclear leukocytes. IPharmacol Exp Ther 244:709-15
McManus LM, Hm��h�n DJ, Demopoulos CA. Pinckard RN. 1980. Pathobio-logy of the intravenous infusion of acetyl glyceryl ether phoephorylcho-line (AG�’C), a synthetic platelet-activating factor (PAP), in the rabbit. JImmunol 124:2919-24
McManus LM, Pinckard RN, Fitzpatrick FA, O’Rowke RA, Crawford MH,Hanahan Di, 1981. Acetyl glyceryl ether phosphorylcholine. Intravascu-lee alterations following intravenous infusion into the baboon. Lab Invest45:303-07
Munson P1. 1987. A User’s Guide to Ligand: Data Analysis and Curve Fittingfor Ligand Binding Experiments. Bethesda, MD: National Institutes ofHealth
Nakayama R, Yasuda K, Saito K, 1987. Existence of endogenous inhibitors of
platelet-activating factor (PAP) with PAP in rat uterus. 3 Biol (1cm 262:13174-79
O’Flaherty IT, Redman JFJr, Schmitt ID, Ellis JM, Surles JR. Marx MN, Plan-tadosi C, Wykle RL, 1987. 1-O-alkyl-2-N-methylcarbamyl- glycerophos-phocholine: a biologically potent, non-metabolizable analog of platelet-activating factor. Biochem Biophys Res Commun 147:18-24
O’Neill C, 1985a. E,�min�tion of the causes of early pregnancy-associatedthrombocytopenia in mice. I Reprod Fertil 73:567-77
603
O’Neill C, 1985b. Partial characterization of the embryo-derived platelet-acti-vating factor in mice. 3 Reprod Fertil 75:375-80
O’Neill C, 1987. Embryo-derived platelet activating factor: a preimplantationembryo mediator of maternal recognition of pregnancy. Domnest AnimEndocrinol 4:69-85
O’Neill C, Pike IL, Porter RN, Gidley-Baird AA, Sinosich Mi, Saunders DM,1985. Maternal recognition of pregnancy prior to implantation: methodsfor monitoring embryonic viability in vitro and in vivo. Ann NY Acad Sci442:429-39
O’Neill C, Saunders DM. 1984. Assessment of embryo quality (letter]. Lancet2:1035
Pieroni 0. Hanahan Di, 1983. Metabolic behavior of acetyl glyceryl ether phos-phoryicholine on interaction with rabbit platelets. Arch Biochem Biophys224:485-93
Pinckard RN, McManus LM, Hanahan DI, 1982. Chemistry and biology ofacetyl glyceiyl ether phospbomylcholine (platelet-activating factor). AdvInflammation Rca 4:147-80
Psychoyos A, 1973. Endocrine control of egg implantation. In: Greep RO, Ast-wood EB, Geiger SR (eds.), Handbook of Physiology. Section 7, Endocri-nology, VoL 2, Part 2. Washington, DC: American Physiological Society,pp. 187-215
Psychoyos A, Casimiri V, 1980. Factors Involved in uterine receptivity and
refractoriness. Prog Reprod Biol 7:143-57Robaut C, Durand 0, James C, Lave D, Sedivy P. Floch A, Mondot S. Pacot D,
Cavern 1, Le Fur 0, 1987. PAP binding sites. Characterization by(3H)52770 RP, a pyrrolo(l,2-cjthiazole derivative, in rabbit platelets.Biochemn Pharmacol 36:3221-29
Saksena SK, Lau IF, (lang MC, 1976. Relationship between oestrogen, prosta-glandin F� and histamine in delayed implantation in the mouse. Acts
Endocrinol (Copenh) 81:801-07Snyder F, 1985. Otemnical and biochemical aspects of platelet activating factor:
a novel class of acetylated ether-linked choline-phospholipids. Med ResRev 5:107-40
Spinks NR, O’Neill C, 1987. Embryo-derived platelet-activating factor is es-sential for establishment of pregnancy in the mouse (letter]. Lancet 1:106-07
Spinks NR, O’Neill C, 1988. Antagonists of embryo-derived platelet activating
factor prevent implantation. I Reprod Fertil 84:89-98TSJIrSOW L, Floch A, Mondot 5, Cavern I, 1988. High affinity specific binding
sites for tritiated platelet-activating factor in canine platelet membranes:
counterparts of platelet-activating factor receptors mediating platelet ag-gregation. Mol Pharmacol 34:145-51
Teipel 3M, Koshland DE, 1969. The significance of intermediary plateau re-
gions In enzyme saturation curves. Biochemistry 111:4656-63Titeler M, 1983. Multiple Dopamine Receptors. New York: Marcel Dekker,
Inc., pp.9-36Tokumura A, Homnma H, Hanahan Di, 1985. Structural analogs of alkylacetyl-
glycerophosphocholine inhibitory behavior on platelet activation. 3 Biol(1cm 260:12710-14
Touqui L, Jacquemin C, Dumarey C, Vargaftig BB,1985. 1-O-alkyl-2-acyl-sn-
glyccro-3- phosphorylcholine Is the precursor of platelet-activating factorin stimulated rabbit platelets. Evidence for an alkylacetyl-glycerophos-phozylcholine cycle. Biochim Biophys Acta 833:111-18
Van Delft JL, van Haezingen Ni, Verbcij NLJ, Domingo HT, (labeler PE,BraquetP, 1988. Specific receptor sites for PAP in iris and ciliary body of
the rabbit eye. Current Eye Rca 7:1063-68Vargaftig BB, Benveniste 1, 1983. Platelet-activating factor today. Trends
Pharmacol Sci 4:341-43Vargaflig BB, Chignard M, Benveniste 1, Lefort I, Wal F, 1981. Background
and present status of research on platelet-activating factor (PAF-acether).Ann NY Acad Sci 370:119-37
Yasuda K, Salouchi K, Saito K, 1986. Platelet-activating factor in normal ratuterus. Biochem Biophys Rca Commun 138:1231-36
Yousufzai SYK, Abdel-Latif AA, 1985. Effects of platelet-activating factor onthe release of arachidonic acid and pro�gtan�1in� by rabbit iris smoothmuscle. Biochein J 228:697-706




















