
Predicting Genetic Interactions in Caenorhabditis elegans ...
Upload
khangminh22Category
view
1download
0
C. elegans Secreted Lipid-Bi
Current Biology 22, 1276–1284, July 24, 2012 ª2012 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2012.06.004
Articlending
Protein NRF-5 Mediates PS Appearanceon Phagocytes for Cell Corpse Engulfment
Yan Zhang,1,3 Haibin Wang,2,3 Eriko Kage-Nakadai,4
Shohei Mitani,4 and Xiaochen Wang3,*1Graduate Program in Chinese Academy of Medical Sciencesand Peking Union Medical College, Beijing 100730, China2College of Biological Sciences, China Agriculture University,Beijing 100094, China3National Institute of Biological Sciences, No. 7 Science ParkRoad, Zhongguancun Life Science Park, Beijing 102206, China4Department of Physiology, Tokyo Women’s MedicalUniversity, School of Medicine and Institute for IntegratedMedical Sciences, 8-1, Kawada-cho, Shinjuku-ku, Tokyo162-8666, Japan
Summary
Background: During programmed cell death, apoptotic cellsare rapidly removed by phagocytes. How dying cells arerecognized remains poorly understood.Results: Here we identify a secreted lipid transfer/LPS-binding family protein, NRF-5, which is required for efficientclearance of cell corpses. We observed that phosphatidylser-ine (PS), which is externalized to the outer leaflet of plasmamembranes in apoptotic cells, is also detected on the surfaceof engulfing cells. Loss of NRF-5 function completely blocksPS appearance on engulfing cells but causes accumulationof PS on apoptotic cells, a phenotype observed in bothced-7(lf) and ttr-52(lf)mutants. TheNRF-5 protein is expressedin and secreted from body wall muscle cells and clustersaround apoptotic cells in a CED-7-dependent manner. NRF-5associates with TTR-52, binds PS, and displays lipid transferactivity in vitro.Conclusion:Our data suggest that NRF-5 may act with CED-7and TTR-52 to mediate PS transfer from apoptotic cells to en-gulfing cells and thus promotes engulfment by phagocytes.
Introduction
Apoptotic cells generated by programmed cell death arerapidly removed to prevent inflammatory and autoimmuneresponses and maintain tissue homeostasis [1, 2]. Phosphati-dylserine (PS), which is normally restricted to the inner leafletof plasma membranes, is exposed on the surface of apoptoticcells, thus distinguishing them from normal living cells andserving as an ‘‘eat me’’ signal for engulfment [3, 4]. Surfaceexpression of PS also occurs in nonapoptotic macrophagesand differentiating monocytes and appears to be related tothe acquisition of the ability to engulf PS-positive target cells[5–7]. However, it is unknown whether this is characteristic ofprofessional phagocytes like macrophages or a general andconserved feature of all phagocytes. It is also unclear howPS expression on phagocytes is regulated and contributes toengulfment.
PS on apoptotic cell surfaces can be recognized directly byphagocyte receptors or indirectly through extracellular
*Correspondence: [email protected]
bridging molecules that cross-link apoptotic cells to phago-cytes [8]. Although the latter mechanism was only found tobe important for apoptotic cells cleared by remote phagocytes[9, 10], we recently identifiedC. elegans TTR-52 as an extracel-lular bridging molecule that links PS on apoptotic cells withreceptors on neighboring phagocytes [11].TTR-52 functions in the ced-1/6/7 pathway, which is one of
two partially redundant signaling pathways that control cellcorpse engulfment in C. elegans (Figure 1E). In the parallelpathway, multiple receptors recognize apoptotic cells andtransduce the signal to the conserved intracellular moleculesCED-2(CRKII), CED-5(DOCK180), and CED-12(ELMO), whichactivate the small guanosine triphosphatase (GTPase) CED-10(RAC1), leading to the cytoskeleton rearrangement requiredfor engulfment [12–15]. In the ced-1/6/7 pathway, TTR-52, anextracellular protein secreted from nonapoptotic intestinecells, mediates in part the recognition of apoptotic cells bybinding to both surface-exposed PS on cell corpses and theextracellular domain of the phagocytic receptor CED-1[11, 16]. ced-7, which encodes an ATP-binding cassette(ABC) transporter, is also involved in cell corpse recognitionand functions in both apoptotic and engulfing cells [16, 17].It has been proposed that CED-7 is required for exposure ofPS in apoptotic cells, whereas ABCA1, the mammalian coun-terpart of CED-7, is thought to play a role in exposing PS inapoptotic cells and macrophages [7, 18, 19]. However, recentstudies in cells from ABCA12/2 mice and individuals withTangier disease or in ABCA1-expressing HeLa cells suggestthat ABCA1 is dispensable for the transbilayer movement ofPS in macrophages and apoptotic cells [20]. Therefore, therole of the ABC transporter in apoptotic cell recognitionremains elusive. It also remains to be determined whetheradditional extracellular proteins are involved in cell corpseengulfment.Here we identify a secreted lipid transfer/LPS (lipopolysac-
charide)-binding family protein, NRF-5, which acts withCED-7 and TTR-52 to mediate PS appearance on engulfingcells, thus promoting phagocytic engulfment.
Results
nrf-5 Is Required for Cell Corpse Clearance
In a forward genetic screen for regulators of cell corpseengulfment, we isolated a recessive mutant qx16, which accu-mulated significantly more embryonic cell corpses than wild-type (WT) (Figure 1A). No excessive cell death occurred inqx16 mutants, but cell corpses persisted significantly longerin qx16 embryos than in WT (53 versus 28 min on average),indicating that cell corpse clearance is defective (Figures 1Band 1C).The gene affected in qx16 mutants is a previously identi-
fied gene, nrf-5, loss of which causes a defect in nosemuscle contraction induced by the antidepressant Fluoxe-tine [21] (see Figure S1A available online). NRF-5 is homolo-gous to proteins of the LPS-binding/lipid transfer protein(LTP) family including human bactericidal/permeabilityincreasing protein (BPI), LPS-binding protein (LBP), choles-teryl ester transfer protein (CETP), and phospholipid transfer

Figure 1. nrf-5 Is Required for Cell Corpse Clearance
(A) Time-course analysis of cell corpses during embryonic development was performed in wild-type (N2, open bar), nrf-5(qx16) (black bar), or nrf-5(sa513)
(gray bar). At least 15 embryos were scored at every stage. Data are shown as mean6 SEM. Data derived from WT and nrf-5(qx16) or WT and nrf-5(sa513)
were compared by unpaired t test. **p < 0.0001; all other points had p > 0.05.
(B) Embryonic cell deaths were followed inWT (N2, n = 3) and nrf-5(qx16) (n = 3) embryos. The average number of total cell deaths (6SEM) is shown in paren-
thesis.
(C) The duration of 33 corpses fromWT (N2, open bar, n = 3) and 61 corpses from nrf-5(qx16) (black bar, n = 4) wasmonitored. The average duration (6SEM)
is in parenthesis.
(D) nrf-5(sa513) does not enhance the engulfment defect in ttr-52 and ced-7mutants. Cell corpses were scored in the head region of 4-fold embryos. At least
15 animals were scored in each strain. Data are shown as mean 6 SEM. The number of cell corpses in WT was compared with that in ttr-52(tm2078) or nrf-
5(sa513). For other points, single ced mutants were compared with ced;nrf-5 double mutants, and ttr-52;nrf-5 double mutants were compared with nrf-5
single mutants. **p < 0.0001; all other points had p > 0.05.
(E) Schematic diagram of the cell corpse engulfment pathway.
See also Figure S1.
NRF-5 Regulates Cell Corpse Engulfment1277
protein (PLTP) [21] (Figure S1B). nrf-5(sa513), a previouslyisolated Fluoxetine-resistant mutant with a stop mutationat Ser159, showed a persistent cell corpse phenotypesimilar to qx16, whereas an incompletely penetrant embry-onic lethality and yolk trafficking defect reported previouslyin nrf-5(sa513) mutants was also observed in qx16 worms[21, 22] (Figure 1A; Figures S1D–S1H). Sequencing of nrf-5in qx16 mutants revealed two G to A transitions that resultedin replacement of Val446 by Met and of Trp476 by a prema-ture stop codon (Figures S1A and S1B). Overexpression ofNRF-5(V446M) but not NRF-5(W476-stop) rescued qx16phenotypes, indicating that the W476-stop mutation causesloss of nrf-5 function (Figure S1A).
NRF-5 Is a Secretory Protein that Clusters around CellCorpses
Like other LTP/LPS-binding family proteins, NRF-5 has anN-terminal secretion signal (Figure S1B) [21]. To test whether
NRF-5 is secreted, we expressed a NRF-5::mCHERRY fusion,driven by heat-shock promoters, or a ssGFP::mNRF-5 fusion,driven by either heat-shock promoters or the engulfingcell-specific ced-1 promoter. In ssGFP::mNRF-5, GFP wasfused to the amino terminus of mature NRF-5 (NRF-5[21–551]) and the secretion signal was added to the aminoterminus of GFP. Apoptotic cells were specifically labeledby both NRF-5 fusion proteins, which formed ring-likestructures surrounding cell corpses and accumulated withinthe extra-embryonic cavity (Figures 2A, 2A0, 2D, and 2D0).Because heat-shock promoters induce global gene expres-sion in C. elegans, these results suggest that NRF-5 is asecretory protein that can recognize apoptotic cells. Re-porters lacking secretion signals (GFP::mNRF-5 or NRF-5[21–551]::mCHERRY), displayed diffuse or punctate stainingpatterns but failed to surround cell corpses or accumulate inthe extra-embryonic cavity (Figures 2B, 2B0, 2E, and 2E0).Expression of secreted but not nonsecretory NRF-5 fully

Figure 2. NRF-5 Is a Secretory Protein fromBody
Wall Muscle Cells and Clusters around Apoptotic
Cells
(A–F0) Differential interference contrast (DIC)
and fluorescent images of WT (A–B0, D–E0) or
ced-3(n717) embryos (C, C0, F, F0) that carry
PhspNRF-5::mCHERRY (A, A0, C, C0), PhspNRF-
5(21-551)::mCHERRY (B, B0), Pced-1ssGFP::
mNRF-5 (D, D0, F, F0), or Pced-1GFP::mNRF-5
(E, E0). Arrows indicate apoptotic cells, and
arrowheads point to accumulated fluorescent
proteins in the extraembryonic cavity. Cell corp-
ses indicated by yellow arrows are shown at 33
magnification in the insets.
(G–H00 0) DIC and fluorescent images of WT
animals transgenic for both Pnrf-5GFP and Pmyo-3
mCHERRY.
Scale bars in all panels represent 5 mm. See also
Figure S2.
Current Biology Vol 22 No 141278
rescued the persistent cell corpse, embryonic lethality, anddefective yolk trafficking phenotypes of nrf-5(qx16) mutants,indicating that NRF-5 must be secreted to function (Fig-ure S1A). Cell corpse labeling by NRF-5::mCHERRY orssGFP::mNRF-5 was abolished by a ced-3 loss-of-functionmutation that blocks almost all apoptosis in worms, confirmingthat the cells labeledbyNRF-5were apoptotic (Figures 2C, 2C0,2F, and 2F0).
Using transcriptional reporters, we found that the nrf-5promoter (Pnrf-5) drove expression in body wall muscle cells,in a pattern similar to the myo-3 promoter (Pmyo-3) (Figures2G–2H00 0) [23], but distinct from the ttr-52 and ges-1promoters, which are active only in the intestine (Fig-ures S2A–S2D00 0) [11, 24]. We also generated NRF-5 trans-lational fusions with or without the secretion signal(Pnrf-5ssGFP::mNRF-5 and Pnrf-5GFP::mNRF-5). SecretedGFP::mNRF-5 (ssGFP::mNRF-5) displayed a diffuse stainingpattern and associated with the basal surface of pharyngealcells and the basal-lateral membranes of the intestine,whereas nonsecretory GFP::mNRF-5 labeled body wallmuscle cells but not the intestine (Figures S2E–S2H00 0).These results suggest that NRF-5 is synthesized in andsecreted from muscle cells to recognize apoptotic cellsand associate with the basal surface of pharyngeal andintestine cells.
NRF-5 and TTR-52 Recognize Apoptotic Cells in Different
MannersNRF-5 behaves very like TTR-52, a secretory protein fromintestine cells which specifically recognizes apoptotic cellsby binding to surface-exposed PS [11]. In tat-1(qx30)mutants, which ectopically expose PS on the surface of livingcells due to loss of TAT-1 P4-ATPase function, TTR-52 stainsvirtually all cells (Figures 3J and 3J0) [11]. In contrast,ssGFP::mNRF-5 was still enriched on apoptotic cells in
tat-1(qx30) embryos, indicating thatcell corpse recognition by NRF-5 isunaffected by ectopic surface expo-sure of PS by living cells (Figures 3E,3E0, 3K, and 3L). Loss of function ofced-1, ced-6, ced-2, ced-12, or ced-5did not disrupt clustering of NRF-5around apoptotic cells (Figures 3B,3B0, 3K, and 3L). However, labeling of
cell corpses by ssGFP::mNRF-5 was significantly reducedin ced-7 mutants, suggesting that ced-7 is required forcell corpse recognition by NRF-5 (Figures 3C, 3C0, 3K,and 3L). None of the above ced mutants, including ced-7(lf), affected binding of apoptotic cells by TTR-52 (Figures3F–3H0, 3M, and 3N). These results suggest that TTR-52binds apoptotic cells through surface-exposed PS, whereasrecognition of cell corpses by NRF-5 requires CED-7. Weobserved slightly increased labeling of cell corpses by TTR-52 in nrf-5(qx16) mutants and enhanced apoptotic celllabeling by NRF-5 in ttr-52(lf) embryos (Figures 3K–3N;Figure S1C).
nrf-5 Is Required for the Appearance of PS on the Engulfing
Cell SurfaceExposure of PS on apoptotic cells distinguishes them fromnormal living cells and serves as an ‘‘eat me’’ signal for clear-ance. We also detected PS on the surface of living cells adja-cent to cell corpses. In WT embryos expressing the secretedPS-binding protein Annexin V::GFP, >50% of GFP-positivecell corpses were surrounded by neighboring living cells,which were also labeled by Annexin V::GFP, albeit moreweakly (Figures 4A, 4A0, and 4I). No GFP signal was detectedon the surface of living cells that did not directly contactdying cells. Similarly, TTR-52::mCHERRY, which bindsspecifically to PS, labeled the surfaces of both the apoptoticcell and the surrounding living cell (Figure 6C). These resultssuggest that PS appears on the surface of both dying cellsand neighboring engulfing cells. Loss of function of ced-6,ced-5, ced-12, or ced-2 did not significantly affect theappearance of PS on engulfing cell surfaces (Figure 4I). Bycontrast, in nrf-5(lf) animals, only 2% of cell corpses weresurrounded by Annexin V::GFP-positive neighboring cells(Figures 4B, 4B0, and 4I). Annexin V::GFP staining of engulfingcells was also completely blocked by loss of function of

Figure 3. ced-7 Is Required for the Clustering of NRF-5 around Cell Corpses
(A–J0) DIC and fluorescent images of WT (A, A0, F, F0), ced-1(e1735) (B, B0, G, G0), ced-7(n2094) (C, C0, H, H0), ttr-52(tm2078) (D, D0), nrf-5(qx16) (I, I0), or tat-1(qx30) (E, E0, J, J0) embryos expressing ssGFP::mNRF-5 (A–E0) or TTR-52::mCHERRY (F–J0). Arrows indicate apoptotic cells. Cell corpses indicated by
yellow arrows are shown at 33 magnification in the insets. Scale bars represent 5 mm.
(K–N) Thepercentageof cell corpses labeledbyssGFP::mNRF-5 (K) orTTR-52::mCHERRY (M)wasquantifiedat the1.5-fold (K) or comma (M) stage in thestrains
indicated. The ratio of fluorescence intensity on apoptotic cells versus their living neighbors (X-fold) was determined at the 1.5-fold (L) or comma (N) stage in
indicated strains expressing ssGFP::mNRF-5 (L) or TTR-52::mCHERRY (N). At least 15 embryos (K, M) or 30 cell corpses (L, N) were quantified in each strain,
anddata are shownasmean6SEM.Dataderived fromdifferentmutant backgroundswerecompared toWT. **p< 0.0001, *p< 0.05; all other points hadp>0.05.
NRF-5 Regulates Cell Corpse Engulfment1279
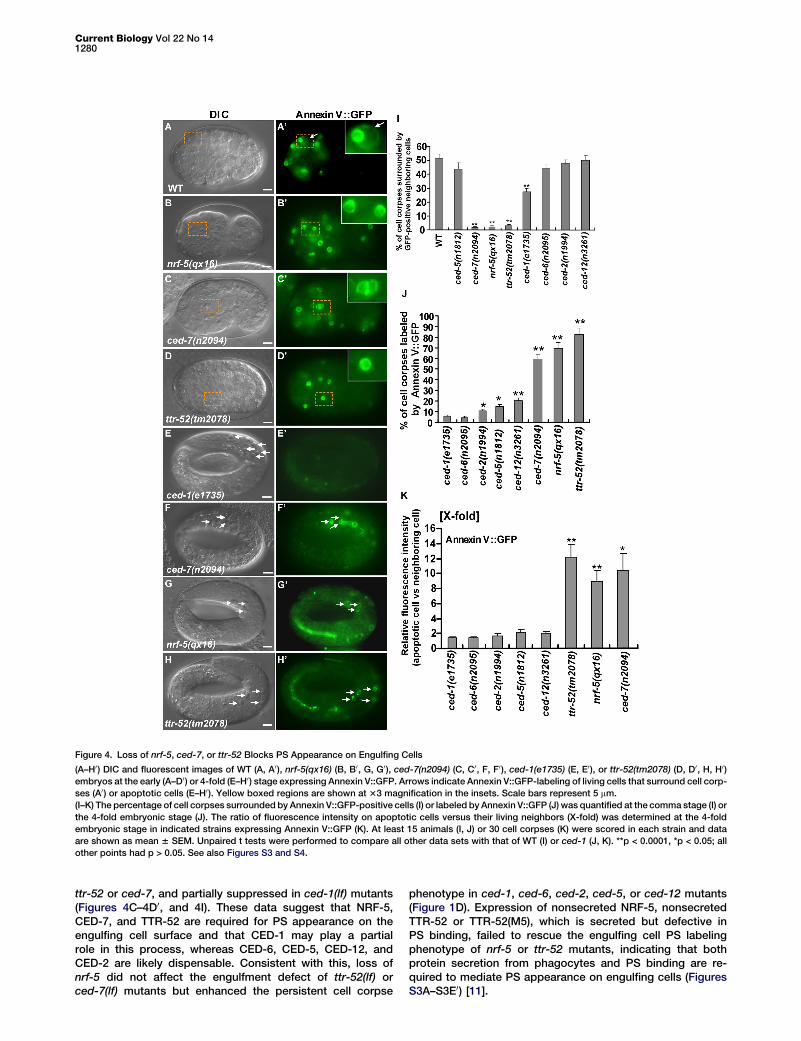
Figure 4. Loss of nrf-5, ced-7, or ttr-52 Blocks PS Appearance on Engulfing Cells
(A–H0) DIC and fluorescent images of WT (A, A0), nrf-5(qx16) (B, B0, G, G0), ced-7(n2094) (C, C0, F, F0), ced-1(e1735) (E, E0), or ttr-52(tm2078) (D, D0, H, H0)embryos at the early (A–D0) or 4-fold (E–H0) stage expressing Annexin V::GFP. Arrows indicate Annexin V::GFP-labeling of living cells that surround cell corp-
ses (A0) or apoptotic cells (E–H0). Yellow boxed regions are shown at 33 magnification in the insets. Scale bars represent 5 mm.
(I–K) The percentage of cell corpses surrounded by Annexin V::GFP-positive cells (I) or labeled by Annexin V::GFP (J) was quantified at the comma stage (I) or
the 4-fold embryonic stage (J). The ratio of fluorescence intensity on apoptotic cells versus their living neighbors (X-fold) was determined at the 4-fold
embryonic stage in indicated strains expressing Annexin V::GFP (K). At least 15 animals (I, J) or 30 cell corpses (K) were scored in each strain and data
are shown as mean 6 SEM. Unpaired t tests were performed to compare all other data sets with that of WT (I) or ced-1 (J, K). **p < 0.0001, *p < 0.05; all
other points had p > 0.05. See also Figures S3 and S4.
Current Biology Vol 22 No 141280
ttr-52 or ced-7, and partially suppressed in ced-1(lf) mutants(Figures 4C–4D0, and 4I). These data suggest that NRF-5,CED-7, and TTR-52 are required for PS appearance on theengulfing cell surface and that CED-1 may play a partialrole in this process, whereas CED-6, CED-5, CED-12, andCED-2 are likely dispensable. Consistent with this, loss ofnrf-5 did not affect the engulfment defect of ttr-52(lf) orced-7(lf) mutants but enhanced the persistent cell corpse
phenotype in ced-1, ced-6, ced-2, ced-5, or ced-12 mutants(Figure 1D). Expression of nonsecreted NRF-5, nonsecretedTTR-52 or TTR-52(M5), which is secreted but defective inPS binding, failed to rescue the engulfing cell PS labelingphenotype of nrf-5 or ttr-52 mutants, indicating that bothprotein secretion from phagocytes and PS binding are re-quired to mediate PS appearance on engulfing cells (FiguresS3A–S3E0) [11].

Figure 5. Loss of nrf-5, ced-7, or ttr-52 Causes
PS Accumulation on Apoptotic Cells
(A–G0) DIC and fluorescent images of WT (A, A0),tat-1(qx30) (B, B0, F, F0), ced-1(e1735);tat-
1(qx30) (C, C0), ced-7(n2094)tat-1(qx30) (D, D0),tat-1(qx30);nrf-5(sa513) (F, F0), or tat-1(qx30)ttr-
52(tm2078) (G, G0) embryos expressing TTR-
52::mCHERRY (A–E0) or GFP::Lact-C2 (F–G0).Arrows indicate apoptotic cells. Scale bars repre-
sent 5 mm.
(H) The ratio of fluorescence intensity on
apoptotic cells versus their living neighbors
(X-fold) was determined in indicated strains ex-
pressing TTR-52::mCHERRY (left) or GFP::Lact-
C2 (right). At least 30 cell corpses were quantified
in each strain. Data are shown as mean 6 SEM.
Unpaired t tests were performed to compare all
other data sets with that of tat-1(qx30). **p <
0.0001; all other points had p > 0.05.
See also Figure S3.
NRF-5 Regulates Cell Corpse Engulfment1281
nrf-5 Mutants Accumulate PS on Apoptotic CellsWe next examined whether loss of nrf-5, ced-7, or ttr-52affected PS appearance on apoptotic cells. In earlyembryos, almost all cell corpses were positive for AnnexinV::GFP in nrf-5(lf), ced-7(lf), or ttr-52(lf) mutants as in WTor other ced mutants defective in cell corpse engulfment,indicating that PS exposure on apoptotic cells is unaffected(Figure S4A). The intensity of Annexin V::GFP labeling onapoptotic cells in nrf-5(lf), ced-7(lf), or ttr-52(lf) mutantswas greater than in WT or other ced mutants at both 1.5-and 2.5-fold stages (Figures S4B and S4C), suggestingthat PS may accumulate on apoptotic cells in these mutants.We next examined Annexin V labeling at the 4-fold embry-onic stage when cell deaths have long been completedand cell corpses are all cleared in WT but persist in engulf-ment-defective mutants. PS, indicated by Annexin V::GFPlabeling, was absent from most cell corpses in ced-1,ced-6, ced-2, ced-5, or ced-12 embryos; a few apoptoticcells (4.6% to 20%) were weakly positive (Figures 4E, 4E0,4J, and 4K). In ced-7 (60%), ttr-52 (83%), or nrf-5 (70%)embryos, more corpses were Annexin V::GFP-positive andthe signal was much stronger, suggesting that PS accumu-lates on apoptotic cell surfaces in these mutants (Figures4F–4K). In tat-1(lf) mutants, inward movement of PS isblocked, and living cells have ectopic surface exposure ofPS, which can be labeled by the PS binding proteins Lact-C2 or TTR-52 (Figures 5B0 and 5F0) [11, 25]. We found thatTTR-52::mCHERRY or GFP::Lact-C2, which is distributedmore evenly between apoptotic and living cells in tat-1(lf)mutants, became re-enriched on apoptotic cells defectivefor nrf-5, ced-7, or ttr-52 but not ced-1 or ced-5 (Figures5C–5H). Loss of nrf-5, ced-7, or ttr-52 did not alter the cellsurface labeling by TTR-52::mCHERRY or GFP::Lact-C2 intat-1(lf);ced-3(lf) embryos in which cell death is blocked(Figure S3F–S3L). This suggests that NRF-5, CED-7, and
TTR-52 do not affect PS externaliza-tion in living cells. Therefore, re-enrich-ment of PS-binding proteins on cellcorpses in tat-1(lf) embryos defectivein ced-7, nrf-5, or ttr-52 is likelycaused by increased PS on apoptoticcell surfaces. Collectively, these dataindicate that loss of nrf-5, ced-7, or
ttr-52 blocks PS appearance on engulfing cell surfaces whilecausing PS accumulation on apoptotic cells.
NRF-5 Binds PS and Possesses Lipid Transfer Activity
NRF-5 belongs to the LPS-binding/lipid transfer protein (LTP)family, members of which bind and transfer various lipidsbetween lipoprotein particles or lipid bilayers. EGFP-NRF-5-Flag bound to PS but not other phospholipids, as previouslyobserved for TTR-52, suggesting that NRF-5 is a PS-bindingprotein. (Figure 6A) [11]. NRF-5 is required for trafficking ofthe lipoprotein VIT-2 that associates with cholesterol (FiguresS1D–S1G0) [22, 26], so we examined cholesterol transferbetween NRF-5 and liposomes. We found that [3H]cholesterolcan be transferred fromNRF-5 to liposomes or from liposomestoNRF-5, suggesting that NRF-5 has lipid transfer activity (Fig-ure 6B). Because NRF-5, TTR-52, and the ABC transporterCED-7 all mediate PS appearance on engulfing cells, we exam-ined whether they associate with each other. Myc-taggedNRF-5 (albeit expressed at a low level), but not the intracellularpart of CED-1 (CED-1[intra]), was specifically coimmunopreci-pitated with TTR-52-flag, indicating that NRF-5 associateswith TTR-52 (Figure S5B). Although recognition of apoptoticcells by NRF-5 requires CED-7 function, no NRF-5/CED-7interaction was detected by coimmunoprecipitation or cross-linking. Their association may involve specific modification(s)acquired during apoptosis.
Cell Corpses Are Internalized by PS-PositiveNeighboring Cells
C. elegans cell corpses are surrounded by multiple living cellsbut are engulfed by only one of them [16, 27]. How phagocytesare specified is not known. We found that PS, as indicated byTTR-52::mCHERRY, first appeared on the apoptotic cellsurface, forming a bright ring-like structure (Figure 6C). In about2 min, PS was detected on the surface of one living neighbor in

Figure 6. NRF-5 Binds PS and Possesses Lipid Transfer
Activity In Vitro
(A) NRF-5 binds PS in vitro. Affinity-purified EGFP-NRF-
5-Flag but not EGFP-Flag bound PS spotted on
a membrane strip.
(B) Transfer of [3H]cholesterol from donor proteins to
acceptor liposomes (left) and from donor liposomes to
acceptor proteins (right) was examined as described in
Experimental Procedures. At least two independent
experiments were performed.
(C) Cell corpses are internalized by the PS-positive
neighbor. The clearance of apoptotic cells is followed
in WT (a, b) and nrf-5(qx16) embryos (c, d) carrying Pttr-52
TTR-52::mCHERRY. The apoptotic cells (white arrow-
heads) are quickly internalized and cleared by the PS-
positive neighbor (yellow arrowheads) in WT but persist
much longer in nrf-5(qx16) embryos, which lack PS
labeling on the neighboring cell. Scale bar represents
1 mm.
See also Figure S5 and Movies S1, S2, S3, and S4.
Current Biology Vol 22 No 141282
WT but not nrf-5(qx16) embryos and was gradually enriched asthis neighbor internalized the cell corpse (Figure 6C, a and b;Movies S1 and S2). PS always appeared on apoptotic cellsbefore engulfing cells (n = 106), and most cell corpses (94/106)were eventually engulfed by the PS-positive neighbor. Becauseall neighboring cells are capable of engulfing dying cells [16, 27,28], the appearance of PS may promote engulfment by thePS-positive neighbor, allowing it to ‘‘outplay’’ other neighboringcells. In support of this, cell corpses persisted significantlylonger in nrf-5(lf)mutants, inwhich PSdid not appear on engulf-ing cells (Figures 1C, 4I, 6C, c and d; Movies S3 and S4).
Discussion
NRF-5 Is a Secreted PS-Binding Protein
Required for Cell Corpse EngulfmentNRF-5 and TTR-52 are the two secreted PS-binding proteins involved in cell corpse engulf-ment. NRF-5 is recruited to apoptotic cellsprimarily by CED-7, whereas TTR-52 bindssurface-exposed PS, cross-linking it withthe phagocytic receptor CED-1. Both TTR-52and CED-1 are required for removal of normalcells in tat-1(lf) mutants, in which PS is ectop-ically exposed on all living cells [11]. However,loss of nrf-5 does not significantly affectCED-1 binding to apoptotic cells. Neither isNRF-5 required for the ‘‘missing cell’’ pheno-type in tat-1(lf) mutants, where a proportionof nonapoptotic cells are absent, presumablyhaving been engulfed after ectopically ex-pressing surface PS (data not shown) [29].Therefore, NRF-5 does not seem to act simplyas a bridge between surface-exposed PS onapoptotic cells and the phagocytic receptorCED-1 like TTR-52.
CED-7, NRF-5, and TTR-52 May Act
Together to Mediate PS Transfer fromApoptotic Cells to Engulfing Cells
We observed PS appearance on the surfaceof engulfing cells, a phenomenon observedpreviously in macrophages, suggesting thatthis is a conserved event during phagocytosis
of apoptotic cells. Our data show that loss of the ABC trans-porter CED-7, or either of the two extracellular PS-bindingproteins NRF-5 and TTR-52, completely blocks PS appear-ance on engulfing cells and causes accumulation of PS onapoptotic cells without affecting PS externalization in eitherliving or dying cells. These results are consistent with aproposed model in which CED-7, NRF-5, and TTR-52 maycooperate to transfer PS from apoptotic to engulfing cells (Fig-ure S5C). NRF-5 belongs to a secreted LPS binding/LTP familyincluding CETP and PLTP, which regulate lipoprotein metabo-lism by exchanging neutral lipids like cholesteryl ester and

NRF-5 Regulates Cell Corpse Engulfment1283
phospholipids between lipoproteins [30], and LBP and BPI,which mediate innate immunity by transferring LPS to themacrophage surface for cell activation [31]. Our finding thatNRF-5 can transfer cholesterol to or extract it from liposomesindicates that NRF-5 has lipid transfer activity, consistent withits role in yolk protein trafficking and PS transfer duringengulfment.
Apoptotic cell recognition by NRF-5 requires CED-7, anABC transporter previously thought to promote PS external-ization in apoptotic cells [18]. We observed that loss ofced-7 caused increased PS on apoptotic cell surfaces anddid not affect ectopic exposure of PS on living cells causedby tat-1 mutation, indicating that ced-7 is not responsiblefor initial exposure of PS on apoptotic cells or living cells.These results are consistent with the finding that ABCA1,the mammalian counterpart of CED-7, is dispensable fortransbilayer PS movement in apoptotic cells or macro-phages [20]. ABCA1 loads cholesterol and phospholipidsonto apoA1, a process promoted by PLTP, which bindsABCA1 on the cell surface [32]. We found that CED-7recruits NRF-5 specifically to dying cells despite beingpresent in both dying and engulfing cells, suggesting theinvolvement of other factors or modifications of CED-7 inapoptotic cells [17]. Because PS is exposed on apoptoticbut not living cells, it is possible that CED-7, when itreceives PS on the apoptotic cell surface, recruits andloads NRF-5 with PS, which is then transferred to engulfingcells (Figure S5C). The requirement of PS and/or modifica-tions of CED-7 acquired during apoptosis may explainwhy we failed to detect associations between CED-7 andNRF-5.
The extracellular PS-binding protein TTR-52 is also requiredfor mediating PS appearance on engulfing cells. TTR-52 asso-ciates with NRF-5 and may act like soluble CD14 (sCD14)(which receives LPS or phospholipids from LBP and transferthem to target cells or HDL particles [33–35]) to relay PSfrom the NRF-5-PS complex to engulfing cells.
PS-Positive Neighbors Outplay the Others to Engulf Cell
CorpsesMammalian apoptotic cells are cleared mainly by mobileprofessional phagocytes like macrophages. In C. elegans, allthe cells neighboring a somatic cell corpse can recognizedying cells and initiate engulfment, but only one completesinternalization [16, 27, 28]. We found that most cell corpseswere internalized by the PS-positive neighbor, suggestingthat PS may help to specify engulfing cell fate. Appearanceof PS on one neighboring cell may promote engulfment bythis cell, thereby excluding other neighbors from completingengulfment. Consistent with this, loss of nrf-5 blocks PSappearance on engulfing cells, severely delaying engulfment.Macrophage surface expression of PS is important for phago-cytosis of PS-positive targets, suggesting that PS appearanceon phagocytes is a conserved mechanism for removingapoptotic cells [5, 6]. How PS appearance on phagocytespromotes engulfment requires further investigation. The pres-ence of PS on both dying and engulfing cells may stabilize thepairing between the eater and the prey; alternatively, PS maymodify the engulfing cell membrane so that it is in a morefavorable state for internalization. In addition, TTR-52 maytransfer PS to cell surface proteins to further activate engulf-ment, like the sCD14-LPS complex, which induces cell activa-tion of macrophages or neutrophils [34] (Figure S5C). CED-1 isa potential candidate to test because loss of ced-1 partially
blocked PS appearance on engulfing cells but did not causePS accumulation on cell corpses, suggesting that CED-1 isnot involved in transferring PS from dying cells but may havea role in anchoring PS on the engulfing cell surface.
Experimental Procedures
Genetic Analysis
Strains of C. elegans were cultured using standard protocols [36]. The WT
strain was N2 Bristol. Details of strain usage, mapping, and cloning are
provided in the Supplemental Experimental Procedures.
Quantification of Cell Corpses, Cell Corpse Duration, PS Appearance,
and PS Accumulation
Cell corpses were counted in the head region of living embryos either at
the six different embryonic stages for a time course analysis or at the
4-fold embryonic stage as described before [37]. Embryonic cell deaths
and cell corpse duration were examined as described previously
[14, 38]. Detailed procedures for quantifying PS appearance and accu-
mulation are provided in the Supplemental Experimental Procedures.
In Vitro Lipid Binding and Lipid Transfer Assay
EGFP-NRF-5-Flag and EGFP-Flag expressed in and secreted from 293T
cells were affinity purified from the culture medium and used for in vitro lipid
binding and transfer assays as described in the Supplemental Experimental
Procedures.
Statistical Analysis
The SEMwas used as y error bars for bar charts plotted from themean value
of the data. Data derived from different genetic backgrounds were
compared by Student’s two-way unpaired t test. Data were considered
statistically different at p < 0.05. p < 0.0001 is indicatedwith double asterisks
and p < 0.05 with single asterisks.
Supplemental Information
Supplemental Information includes five figures, Supplemental Experimental
Procedures, and four movies and can be found with this article online at
doi:10.1016/j.cub.2012.06.004.
Acknowledgments
We thank P. Liu, T. Wei, and K. Zhao for help with the lipid transfer assay, D.
Xue for providing the Annexin V::GFP reporter, and the Caenorhabditis
Genetic Center (CGC) for strains.We thank Jing Lin,Mingxing Lu, andDong-
feng Zhao for technical support and Isabel Hanson for editing services. This
work was supported by a grant from theMinistry of Science and Technology
to X.W. (2010CB835201).
Received: January 9, 2012
Revised: April 10, 2012
Accepted: June 1, 2012
Published online: June 21, 2012
References
1. Savill, J., and Fadok, V. (2000). Corpse clearance defines themeaning of
cell death. Nature 407, 784–788.
2. Savill, J., Dransfield, I., Gregory, C., and Haslett, C. (2002). A blast from
the past: clearance of apoptotic cells regulates immune responses. Nat.
Rev. Immunol. 2, 965–975.
3. Fadok, V.A., Voelker, D.R., Campbell, P.A., Cohen, J.J., Bratton, D.L.,
and Henson, P.M. (1992). Exposure of phosphatidylserine on the
surface of apoptotic lymphocytes triggers specific recognition and
removal by macrophages. J. Immunol. 148, 2207–2216.
4. Krahling, S., Callahan, M.K., Williamson, P., and Schlegel, R.A. (1999).
Exposure of phosphatidylserine is a general feature in the phagocytosis
of apoptotic lymphocytes by macrophages. Cell Death Differ. 6,
183–189.
5. Callahan, M.K., Williamson, P., and Schlegel, R.A. (2000). Surface
expression of phosphatidylserine on macrophages is required for
phagocytosis of apoptotic thymocytes. Cell Death Differ. 7, 645–653.

Current Biology Vol 22 No 141284
6. Callahan, M.K., Halleck, M.S., Krahling, S., Henderson, A.J., Williamson,
P., and Schlegel, R.A. (2003). Phosphatidylserine expression and
phagocytosis of apoptotic thymocytes during differentiation of mono-
cytic cells. J. Leukoc. Biol. 74, 846–856.
7. Marguet, D., Luciani, M.F., Moynault, A., Williamson, P., and Chimini, G.
(1999). Engulfment of apoptotic cells involves the redistribution of
membrane phosphatidylserine on phagocyte and prey. Nat. Cell Biol.
1, 454–456.
8. Fadeel, B. (2003). Programmed cell clearance. Cell. Mol. Life Sci. 60,
2575–2585.
9. Hanayama, R., Tanaka, M., Miwa, K., Shinohara, A., Iwamatsu, A., and
Nagata, S. (2002). Identification of a factor that links apoptotic cells to
phagocytes. Nature 417, 182–187.
10. Ishimoto, Y., Ohashi, K., Mizuno, K., and Nakano, T. (2000). Promotion of
the uptake of PS liposomes and apoptotic cells by a product of growth
arrest-specific gene, gas6. J. Biochem. 127, 411–417.
11. Wang, X., Li, W., Zhao, D., Liu, B., Shi, Y., Chen, B., Yang, H., Guo, P.,
Geng, X., Shang, Z., et al. (2010). Caenorhabditis elegans transthyre-
tin-like protein TTR-52 mediates recognition of apoptotic cells by the
CED-1 phagocyte receptor. Nat. Cell Biol. 12, 655–664.
12. Hsu, T.Y., and Wu, Y.C. (2010). Engulfment of apoptotic cells in C.
elegans is mediated by integrin alpha/SRC signaling. Curr. Biol. 20,
477–486.
13. Reddien, P.W., and Horvitz, H.R. (2004). The engulfment process of pro-
grammed cell death in caenorhabditis elegans. Annu. Rev. Cell Dev.
Biol. 20, 193–221.
14. Wang, X., Wu, Y.C., Fadok, V.A., Lee, M.C., Gengyo-Ando, K., Cheng,
L.C., Ledwich, D., Hsu, P.K., Chen, J.Y., Chou, B.K., et al. (2003). Cell
corpse engulfmentmediated by C. elegans phosphatidylserine receptor
through CED-5 and CED-12. Science 302, 1563–1566.
15. Cabello, J., Neukomm, L.J., Gunesdogan, U., Burkart, K., Charette, S.J.,
Lochnit, G., Hengartner, M.O., and Schnabel, R. (2010). The Wnt
pathway controls cell death engulfment, spindle orientation, and migra-
tion through CED-10/Rac. PLoS Biol. 8, e1000297.
16. Zhou, Z., Hartwieg, E., and Horvitz, H.R. (2001). CED-1 is a transmem-
brane receptor that mediates cell corpse engulfment in C. elegans.
Cell 104, 43–56.
17. Wu, Y.C., and Horvitz, H.R. (1998). The C. elegans cell corpse engulf-
ment gene ced-7 encodes a protein similar to ABC transporters. Cell
93, 951–960.
18. Venegas, V., and Zhou, Z. (2007). Two alternativemechanisms that regu-
late the presentation of apoptotic cell engulfment signal in
Caenorhabditis elegans. Mol. Biol. Cell 18, 3180–3192.
19. Hamon, Y., Broccardo, C., Chambenoit, O., Luciani, M.F., Toti, F.,
Chaslin, S., Freyssinet, J.M., Devaux, P.F., McNeish, J., Marguet, D.,
and Chimini, G. (2000). ABC1 promotes engulfment of apoptotic cells
and transbilayer redistribution of phosphatidylserine. Nat. Cell Biol. 2,
399–406.
20. Williamson, P., Halleck, M.S., Malowitz, J., Ng, S., Fan, X., Krahling, S.,
Remaley, A.T., and Schlegel, R.A. (2007). Transbilayer phospholipid
movements in ABCA1-deficient cells. PLoS ONE 2, e729.
21. Choy, R.K., Kemner, J.M., and Thomas, J.H. (2006). Fluoxetine-resis-
tance genes in Caenorhabditis elegans function in the intestine and
may act in drug transport. Genetics 172, 885–892.
22. Choy, R.K., and Thomas, J.H. (1999). Fluoxetine-resistant mutants in C.
elegans define a novel family of transmembrane proteins. Mol. Cell 4,
143–152.
23. Fire, A., andWaterston, R.H. (1989). Proper expression of myosin genes
in transgenic nematodes. EMBO J. 8, 3419–3428.
24. Kennedy, B.P., Aamodt, E.J., Allen, F.L., Chung, M.A., Heschl, M.F., and
McGhee, J.D. (1993). The gut esterase gene (ges-1) from the nematodes
Caenorhabditis elegans and Caenorhabditis briggsae. J. Mol. Biol. 229,
890–908.
25. Chen, B., Jiang, Y., Zeng, S., Yan, J., Li, X., Zhang, Y., Zou, W., and
Wang, X. (2010). Endocytic sorting and recycling require membrane
phosphatidylserine asymmetry maintained by TAT-1/CHAT-1. PLoS
Genet. 6, e1001235.
26. Matyash, V., Geier, C., Henske, A., Mukherjee, S., Hirsh, D., Thiele, C.,
Grant, B., Maxfield, F.R., and Kurzchalia, T.V. (2001). Distribution and
transport of cholesterol in Caenorhabditis elegans. Mol. Biol. Cell 12,
1725–1736.
27. Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983).
The embryonic cell lineage of the nematode Caenorhabditis elegans.
Dev. Biol. 100, 64–119.
28. Hoeppner, D.J., Hengartner, M.O., and Schnabel, R. (2001). Engulfment
genes cooperate with ced-3 to promote cell death in Caenorhabditis
elegans. Nature 412, 202–206.
29. Darland-Ransom, M., Wang, X., Sun, C.L., Mapes, J., Gengyo-Ando, K.,
Mitani, S., and Xue, D. (2008). Role of C. elegans TAT-1 protein in main-
taining plasmamembrane phosphatidylserine asymmetry. Science 320,
528–531.
30. Tall, A. (1995). Plasma lipid transfer proteins. Annu. Rev. Biochem. 64,
235–257.
31. Schultz, H., and Weiss, J.P. (2007). The bactericidal/permeability-
increasing protein (BPI) in infection and inflammatory disease. Clin.
Chim. Acta 384, 12–23.
32. Oram, J.F., Wolfbauer, G., Vaughan, A.M., Tang, C., and Albers, J.J.
(2003). Phospholipid transfer protein interacts with and stabilizes
ATP-binding cassette transporter A1 and enhances cholesterol efflux
from cells. J. Biol. Chem. 278, 52379–52385.
33. Wurfel, M.M., Hailman, E., andWright, S.D. (1995). Soluble CD14 acts as
a shuttle in the neutralization of lipopolysaccharide (LPS) by LPS-
binding protein and reconstituted high density lipoprotein. J. Exp.
Med. 181, 1743–1754.
34. Hailman, E., Vasselon, T., Kelley, M., Busse, L.A., Hu, M.C., Lichenstein,
H.S., Detmers, P.A., and Wright, S.D. (1996). Stimulation of macro-
phages and neutrophils by complexes of lipopolysaccharide and
soluble CD14. J. Immunol. 156, 4384–4390.
35. Yu, B., Hailman, E., andWright, S.D. (1997). Lipopolysaccharide binding
protein and soluble CD14 catalyze exchange of phospholipids. J. Clin.
Invest. 99, 315–324.
36. Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics
77, 71–94.
37. Parrish, J., Li, L., Klotz, K., Ledwich, D., Wang, X., and Xue, D. (2001).
Mitochondrial endonuclease G is important for apoptosis in C. elegans.
Nature 412, 90–94.
38. Guo, P., Hu, T., Zhang, J., Jiang, S., and Wang, X. (2010). Sequential
action of Caenorhabditis elegans Rab GTPases regulates phagolyso-
some formation during apoptotic cell degradation. Proc. Natl. Acad.
Sci. USA 107, 18016–18021.
Copyright © 2022 FDOKUMEN




















