
MicroRNA125a contributes to elevated inflammatory chemokine RANTES via targeting KLF13 in systemic...
11
ARTHRITIS & RHEUMATISM Vol. 62, No. 11, November 2010, pp 3425–3435 DOI 10.1002/art.27632 © 2010, American College of Rheumatology MicroRNA-125a Contributes to Elevated Inflammatory Chemokine RANTES Levels via Targeting KLF13 in Systemic Lupus Erythematosus Xia Zhao, Yuanjia Tang, Bo Qu, Huijuan Cui, Shujun Wang, Lijia Wang, Xiaobing Luo, Xinfang Huang, Jia Li, Shunle Chen, and Nan Shen Objective. MicroRNA (miRNA) have received in- creasing attention as posttranscriptional regulators that fine-tune the homeostasis of the inflammatory response. This study aimed to clarify whether miR-125a, which was identified in a pilot expression profiling step, is involved in the inflammatory chemokine pathway in systemic lupus erythematosus (SLE). Methods. Independent verification of miR-125a expression in amplified samples from SLE patients and normal controls was performed by TaqMan quantitative polymerase chain reaction (PCR) analysis. A combina- tion of 3 bioinformatic prediction techniques and re- porter gene assays was used to identify miR-125a tar- gets. In vitro systems of overexpression by transfection and inducible expression by stimulation were performed to investigate the function of miR-125a, which was followed by real-time quantitative PCR and enzyme- linked immunosorbent assay. Results. In SLE patients, the expression of miR- 125a was reduced and the expression of its predicted target gene, KLF13, was increased. Bioinformatics pre- dicted that miR-125a base-paired with sequences in the 3-untranslated region of KLF13. Overexpression of miR-125a led to a significant reduction in the expres- sion of RANTES and KLF13. MicroRNA-125a inhibited endogenous KLF13 expression in a dose-dependent manner, as determined using gain- and loss-of-function methods. A luciferase reporter system confirmed the miR-125a binding sites. Notably, miR-125a expression was induced in T cells in a dose- and time-dependent manner. Finally, the introduction of miR-125a into T cells from SLE patients alleviated the elevated RANTES expression. Conclusion. MicroRNA-125a negatively regulates RANTES expression by targeting KLF13 in activated T cells. The underexpression of miR-125a contributes to the elevated expression of RANTES in SLE. Our find- ings extend the role of miRNA in the pathogenesis of lupus and provide potential strategies for therapeutic intervention. MicroRNA (miRNA) are conserved noncoding RNAs of 22 nucleotides that regulate gene expression by binding to the 3-untranslated region (3-UTR) of target messenger RNAs (mRNA), leading to their trans- lational repression or degradation (1). Currently, over 700 miRNA have been identified in mammals (see the miRNA registry at www.sanger.ac.uk), which play piv- otal roles in diverse biologic processes, ranging from cell development and differentiation to apoptosis (2). The role of miRNA in the immune system has been high- lighted to maintain immune system homeostasis (3,4). Supported in part by the Knowledge Innovation Program of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences (grant 2007KIP305), the National Basic Research Program of China (973 Program, grant 2007CB947900), the National High Tech- nology Research and Development Program of China (863 Program, grant 2007AA02Z123), the National Natural Science Foundation of China (grants 30700734 and 30301026), the Program of the Shanghai Commission of Science and Technology (grants 06JC14050, 07ZR14130, and 08JC1414700), and the Program of Shanghai Subject Chief Scientist (grant 07XD14021). Xia Zhao, MD, Yuanjia Tang, PhD, Bo Qu, PhD, Huijuan Cui, MD, Shujun Wang, MD, Lijia Wang, MD, Xiaobing Luo, PhD, Xinfang Huang, MD, Jia Li, MD, Shunle Chen, MD, Nan Shen, MD: Joint Molecular Rheumatology Laboratory of Institute of Health Sciences and Shanghai Renji Hospital, Shanghai JiaoTong University School of Medicine, Shanghai Institutes for Biological Sciences, and Chinese Academy of Sciences, Shanghai, China. Drs. Zhao, Tang, and Qu contributed equally to this work. Address correspondence and reprint requests to Nan Shen, MD, Department of Rheumatology, Renji Hospital, Shanghai Jiao- Tong University School of Medicine, 145 Shan Dong Road (Central), Shanghai 200001, China. E-mail: nanshenisibs@gmail@com. Submitted for publication November 6, 2009; accepted in revised form June 22, 2010. 3425
Transcript of MicroRNA125a contributes to elevated inflammatory chemokine RANTES via targeting KLF13 in systemic...

ARTHRITIS & RHEUMATISMVol. 62, No. 11, November 2010, pp 3425–3435DOI 10.1002/art.27632© 2010, American College of Rheumatology
MicroRNA-125a Contributes toElevated Inflammatory Chemokine RANTES Levels via
Targeting KLF13 in Systemic Lupus Erythematosus
Xia Zhao, Yuanjia Tang, Bo Qu, Huijuan Cui, Shujun Wang, Lijia Wang, Xiaobing Luo,Xinfang Huang, Jia Li, Shunle Chen, and Nan Shen
Objective. MicroRNA (miRNA) have received in-creasing attention as posttranscriptional regulatorsthat fine-tune the homeostasis of the inflammatoryresponse. This study aimed to clarify whether miR-125a,which was identified in a pilot expression profiling step,is involved in the inflammatory chemokine pathway insystemic lupus erythematosus (SLE).
Methods. Independent verification of miR-125aexpression in amplified samples from SLE patients andnormal controls was performed by TaqMan quantitativepolymerase chain reaction (PCR) analysis. A combina-tion of 3 bioinformatic prediction techniques and re-porter gene assays was used to identify miR-125a tar-gets. In vitro systems of overexpression by transfectionand inducible expression by stimulation were performedto investigate the function of miR-125a, which was
followed by real-time quantitative PCR and enzyme-linked immunosorbent assay.
Results. In SLE patients, the expression of miR-125a was reduced and the expression of its predictedtarget gene, KLF13, was increased. Bioinformatics pre-dicted that miR-125a base-paired with sequences in the3�-untranslated region of KLF13. Overexpression ofmiR-125a led to a significant reduction in the expres-sion of RANTES and KLF13. MicroRNA-125a inhibitedendogenous KLF13 expression in a dose-dependentmanner, as determined using gain- and loss-of-functionmethods. A luciferase reporter system confirmed themiR-125a binding sites. Notably, miR-125a expressionwas induced in T cells in a dose- and time-dependentmanner. Finally, the introduction of miR-125a into Tcells from SLE patients alleviated the elevated RANTESexpression.
Conclusion. MicroRNA-125a negatively regulatesRANTES expression by targeting KLF13 in activated Tcells. The underexpression of miR-125a contributes tothe elevated expression of RANTES in SLE. Our find-ings extend the role of miRNA in the pathogenesis oflupus and provide potential strategies for therapeuticintervention.
MicroRNA (miRNA) are conserved noncodingRNAs of �22 nucleotides that regulate gene expressionby binding to the 3�-untranslated region (3�-UTR) oftarget messenger RNAs (mRNA), leading to their trans-lational repression or degradation (1). Currently, over700 miRNA have been identified in mammals (see themiRNA registry at www.sanger.ac.uk), which play piv-otal roles in diverse biologic processes, ranging from celldevelopment and differentiation to apoptosis (2). Therole of miRNA in the immune system has been high-lighted to maintain immune system homeostasis (3,4).
Supported in part by the Knowledge Innovation Program ofShanghai Institutes for Biological Sciences, Chinese Academy ofSciences (grant 2007KIP305), the National Basic Research Program ofChina (973 Program, grant 2007CB947900), the National High Tech-nology Research and Development Program of China (863 Program,grant 2007AA02Z123), the National Natural Science Foundation ofChina (grants 30700734 and 30301026), the Program of the ShanghaiCommission of Science and Technology (grants 06JC14050,07ZR14130, and 08JC1414700), and the Program of Shanghai SubjectChief Scientist (grant 07XD14021).
Xia Zhao, MD, Yuanjia Tang, PhD, Bo Qu, PhD, HuijuanCui, MD, Shujun Wang, MD, Lijia Wang, MD, Xiaobing Luo, PhD,Xinfang Huang, MD, Jia Li, MD, Shunle Chen, MD, Nan Shen, MD:Joint Molecular Rheumatology Laboratory of Institute of HealthSciences and Shanghai Renji Hospital, Shanghai JiaoTong UniversitySchool of Medicine, Shanghai Institutes for Biological Sciences, andChinese Academy of Sciences, Shanghai, China.
Drs. Zhao, Tang, and Qu contributed equally to this work.Address correspondence and reprint requests to Nan Shen,
MD, Department of Rheumatology, Renji Hospital, Shanghai Jiao-Tong University School of Medicine, 145 Shan Dong Road (Central),Shanghai 200001, China. E-mail: nanshenisibs@gmail@com.
Submitted for publication November 6, 2009; accepted inrevised form June 22, 2010.
3425

Accordingly, dysregulated levels of miRNA are associ-ated with a growing list of diseases, including auto-immune diseases (5), and unraveling their roles indisease pathogenesis has proved to be intriguing. Nota-bly, miRNA modulate the expression of target genes toan optimum level, rather than participating in on/offdecisions in the inflammatory response (6), providingunique roles in rheumatic diseases by regulating inflam-mation (7). Efforts to identify miRNA targets and tounderstand their effects on biologic processes havebecome active areas of research for their potentialdiagnostic and therapeutic roles in human diseases (8).
A fundamental question in rheumatic diseases ishow the abnormal inflammatory response works in ahighly reproducible way, triggered by pathogens orspecific antigens. Critical components in this process arethe inflammatory chemokines, which direct the circulat-ing leukocytes to sites of inflammation. This family ofsmall proteins is not only restricted merely to chemo-taxis, but it is also involved in the control of leukocyteactivation, angiogenesis, and the amplification of proin-flammatory cytokine production, Th cell polarization,etc. (9–12). Previous studies have shown that variousmechanisms of posttranscriptional control, such as nu-clear export, translation initiation, and mRNA decay,are critical in determining the amount of chemokines(13). Recently, emerging data have suggested thatmiRNA provide an additional layer of posttranscrip-tional regulation in many immunologic functions, suchas orchestrating inflammatory responses, especially bytargeting many central molecules or immunoregulatorycells (14). Relatively little is known about the role ofmiRNA regulation in inflammatory chemokines, how-ever. We hypothesized that the optimal regulation ofinflammatory chemokines probably involves the coordi-nated participation of multiple regulation systems, suchas an integrated network of transcription factors andmiRNA.
Systemic lupus erythematosus (SLE) is a chronicautoimmune inflammatory disease characterized by amyriad of immunoregulatory abnormalities that lead toinjuries of tissues and organs (15). Since inflammation isthe hallmark of the disease, considerable importance hasbeen attached to the aberrant production of the inflam-matory chemokines that regulate inflammation in SLE,which are promising drug targets (16,17). RANTES(also known as CCL5) is a typical chemokine that isknown for enhanced levels and detrimental effects ininflammatory processes such as arthritis, nephritis, andother inflammatory disorders (18,19). As a key chemo-kine for T cell recruitment to inflammatory tissues,
activated RANTES expression may be associated withpersistent or recurrent organ inflammation and can actas a biomarker for disease activity (20,21). Previousstudies have shown that RANTES initiates renal damagein MRL-Faslpr mice and is detectable in kidney tissuesbefore other signs of inflammation are apparent (22,23),whereas the pathologic mechanisms underlying elevatedRANTES levels in SLE have not been fully clarified.
In the present study, we identified the underex-pression of miR-125a in patients with SLE. By exploringthe function of miR-125a, we characterized a previouslyunrecognized role of miR-125a as a negative regulator inthe feedback loop of Kruppel-like factor 13 (KLF13)and RANTES production in activated T cells. Theunderexpression of miR-125a contributes to the elevatedexpression of RANTES by targeting KLF13 in lupus Tcells. These findings provide further insight into the roleof miRNA in the pathogenesis of lupus and are valuablefor developing novel therapeutic strategies.
PATIENTS AND METHODS
Patients and controls and handling of samples. AllSLE patients met at least 4 of the American College ofRheumatology 1982 revised criteria for SLE (24). Patients withconcurrent infection were excluded from the study. Healthydonors had no history of autoimmune diseases or treatmentwith immunosuppressive agents. (Additional clinical informa-tion is available upon request from the author.)
Whole blood samples (10 ml) were taken from allparticipants. Peripheral blood samples obtained from eachsubject were collected into tubes containing acid citrate dex-trose Formula A. Erythrocytes were immediately lysed, andleukocytes were kept in TRIzol Reagent (Invitrogen). Allpatient samples were obtained from patients attending theDepartment of Rheumatology of Renji Hospital in Shanghai.All healthy control samples were obtained from healthy vol-unteer donors. Samples of peripheral blood mononuclear cells(PBMCs) for isolating T cells and studying the function ofmiRNA were obtained from the Blood Center in Shanghai.
All study subjects gave their informed consent forparticipation. The study was approved by the Research EthicsBoard of Renji Hospital, Shanghai JiaoTong University Schoolof Medicine.
Isolation of CD3� T cells. PBMCs were separatedfrom heparinized whole blood by density-gradient centrifuga-tion on Lymphoprep Ficoll-Paque Plus (GE Healthcare).CD3� T cells were purified from fresh PBMCs by positiveselection using magnetic CD3 microbeads (Miltenyi Biotec)according to the manufacturer’s protocol. The purity of Tlymphocytes was �98%, as analyzed using a FACSCaliburinstrument (Becton Dickinson).
Cell culture. Purified T lymphocytes were cultured inRPMI 1640 medium supplemented with 10% fetal bovineserum (FBS), 100 units/ml of penicillin, and 100 units/ml ofstreptomycin, and were stimulated with phytohemagglutinin
3426 ZHAO ET AL

protein (PHA-P) (Sigma) for up to 7 days at 37°C in anatmosphere of 5% CO2. On days 3 and 5, fresh medium wasadded to maintain the cell density at 106/ml. HeLa cells weremaintained at 37°C under an atmosphere of 5% CO2 inDulbecco’s modified Eagle’s medium (DMEM) containing10% FBS and 100 units/ml of penicillin/streptomycin.
Plasmid construction. Two putative miR-125a targetsequences in the 3�-UTR of human KLF13 mRNA (theregions encompassing nucleotides –37 to �937 bases andnucleotides �1037 to �2014 bases in the KLF13 3�-UTR,selectively) were separately amplified from genomic DNA andcloned into psiCHECK-2 vector (Promega). These are re-ferred to as site 1 and site 2 constructs (site 1 and site 2 ofpsiCHECK-2-KLF13 3�-UTR). One fragment of the humanKLF13 3�-UTR that included no putative miR-125a bindingsite (region encompassing nucleotides �2000 to �3007 basesin the KLF13 3�-UTR) was also amplified from genomic DNA,and cloned into psiCHECK-2 vector downstream of the Renillaluciferase reporter gene. This is referred to as the site 3construct, and it was used as an unrelated fragment control.
The following primers were used to create the 3�-UTRluciferase reporter constructs: for the site 1 construct, 5�-CCGCTCGAGAGCAGCCCCACCATCAG-3� (forward) and5�-ATTTGCGGCCGCCGGGCAAACTCGGAACT-3� (re-verse); for the site 2 construct, 5�-CCGCTCGAGCTGTGTG-AGCGGCTGTG-3� (forward) and 5�-ATTTGCGGCCGCT-CCCACCACAGCAGAAAC-3� (reverse); and for the site 3construct, 5�-CCGCTCGAGTCTGCTGTGGTGGGAAC-T-3� (forward) and 5�-ATTTGCGGCCGCTGGAGGCTGTT-TCAGGAG-3� (reverse). The forward and reverse primerscreated Xho I and Not I sites, respectively, and the amplifiedfragments were inserted into the Xho I/Not I–digestedpsiCHECK-2 vector. All plasmids were verified by sequencing.
Site-directed mutagenesis. To generate KLF13 3�-UTR mutants, polymerase chain reaction (PCR)–based site-directed mutagenesis was performed to substitute 4 nucleo-tides in the seed sequences of the miR-125a binding sites in theabove constructs (site 1 and site 2), using KOD-Plus high-fidelity DNA polymerase (Toyobo). Primers for the mutationswere as follows: for the site 1 mutation, 5�-TGTTGAACCC-CCTTTCTCTCCCATGGACACGTTTC-3� (forward) and 5�-GAAACGTGTCCATGGGAGAGAAAGGGGGTTCAAC-A-3� (reverse); and for the site 2 mutation, 5�-GG-GGATTGGGTGGACTGTCCGTATCTTGCCAGAGA-3�(forward) and 5�-TCTCTGGCAAGATACGGACAGTCCA-CCCAATCCCC-3� (reverse). The mutated plasmids wereverified by sequencing.
Mimics and inhibitors of miRNA. MicroRNA oligo-nucleotides were obtained from Dharmacon. MicroRNA mim-ics are double-stranded RNA oligonucleotides, and miRNAhairpin inhibitors are single-stranded oligonucleotides. Thecombination of gain-of-function (mimic-induced down-regulation) and loss-of-function (inhibitor-induced up-regulation) experiments demonstrated the relationships be-tween the miRNA and their targets and allowed for thefunctional analysis of miRNA. Each negative control for theinhibitors and mimics is based on the sequences of miRNA inCaenorhabditis elegans (cel-miR-67), which has minimal se-quence identity with miRNA in humans, mice, or rats.
Transfection. The transfection of human primary Tcells was performed using INTERFERin transfection reagent
(Polyplus Transfection) according to the manufacturer’s pro-tocol. Six hours after transfection, T cells were added to freshcomplete medium (RPMI 1640 medium supplemented with10% FBS) and stimulated with PHA-P for up to 7 days at 37°Cin an atmosphere of 5% CO2. On days 3 and 5, fresh mediumwas added to maintain the cell density at 106/ml.
Real-time quantitative reverse transcription–PCR(RT-PCR). Total RNA was extracted using TRIzol reagent(Invitrogen) according to the manufacturer’s protocol. Toquantify miRNA, the expression of the 156 miRNA included inthe TaqMan MicroRNA Assays Human Panel (Early Access)kit (Applied Biosystems) was examined according to themanufacturer’s protocol. To validate the results, the TaqMankit specified that the quantification of miR-125a be used, withnormalization to the U6 small nuclear RNA (U6 snRNA).RNA (20 ng) from each sample was reverse transcribed usinga TaqMan Human MicroRNA Assays kit (Applied Biosys-tems). To quantify the mRNA, 500 ng of RNA from eachsample was reverse transcribed into complementary DNA(cDNA) using a PrimeScript RT Reagent kit (Takara, Shiga,Japan). The cDNA was amplified by real-time RT-PCR withSYBR Green (SYBR Premix Ex Taq RT-PCR kit; Takara).Ribosomal protein L13a (RPL13a) gene expression was usedas the internal control to normalize the amounts of cDNA.
The TaqMan and SYBR Green assays were performedin duplicate on a 7900HT Fast Real-Time PCR System (Ap-plied Biosystems). The relative expression levels were analyzedusing the comparative threshold cycle (Ct) method accordingto the manufacturer’s instructions. The primers used were asfollows: for KLF13, 5�-CCGCAGAGGAAGCACAA-3� (for-ward) and 5�-CTTCTTCTCGCCCGTGT-3� (reverse); forRANTES, 5�-AGTCGTCTTTGTCACCCGAAA-3� (forward)and 5�-AGCTCATCTCCAAAGAGTTGATGTAC-3� (re-verse); and for RPL13A, 5�-CCTGGAGGAGAAGAGGAA-AGAGA (forward) and 5�-TTGAGGACCTCTGTGTATTT-GTCAA (reverse).
Reporter gene assay. One day before transfection,HeLa cells were seeded at 2 � 104 cells/well in a 96-well plate.PsiCHECK-2 luciferase reporter plasmids (20 ng) containingeither the wild-type or the mutated KLF13 3�-UTR describedabove and miRNA oligonucleotides were cotransfected intoHeLa cells at 80–90% confluence using Lipofectamine 2000(Invitrogen) on the following day. The cells were harvested 24hours later, and luciferase activity was assessed using a Dual-Luciferase Reporter Assay System (Promega). Firefly lucif-erase was used to normalize the Renilla luciferase. All exper-iments were performed in triplicate.
Enzyme-linked immunosorbent assay (ELISA).RANTES protein secreted into the cell culture supernatantwas quantified using commercially available ELISA kits (XiTang Biology) according to the manufacturer’s protocol.
Western blotting. In a 6-well plate, T cells were seededat 5 � 106 cells/well and transfected with miRNA oligonucleo-tides to a 100 nM final concentration of miR-125a or controlper well. Twenty-four hours after transfection, T cells wereadded to fresh complete medium (RPMI 1640 medium sup-plemented with 10% FBS) and stimulated with PHA-P for upto 7 days at 37°C in an atmosphere of 5% CO2. On days 3 and5, fresh medium was added to maintain the cell density at106/ml. On day 7, cells were lysed, and proteins were extracted.Supernatants were then subjected to sodium dodecyl sulfate–
MicroRNA-125a AND ELEVATED RANTES LEVELS IN SLE 3427
polyacrylamide gel electrophoresis, blotted with the indicatedantibodies, and detected with Luminol/Enhancer Solution(Pierce). We carried out Western blotting of KLF13 and actinusing rabbit anti-KLF13 antibody (1:500 dilution; Abnova) andrabbit anti-actin antibody (1:5,000 dilution; Santa Cruz Bio-technology). Relative expression was quantified using QuantityOne software, version 4.52 (Bio-Rad).
Statistical analysis. Data were analyzed using Prism 4software, version 4.03 (GraphPad Software). The nonpara-metric Mann-Whitney U test was used to compare geneexpression between 2 groups, but an unpaired t-test was usedto compare reporter gene activity. P values (2-tailed) less than0.05 were considered statistically significant.
RESULTS
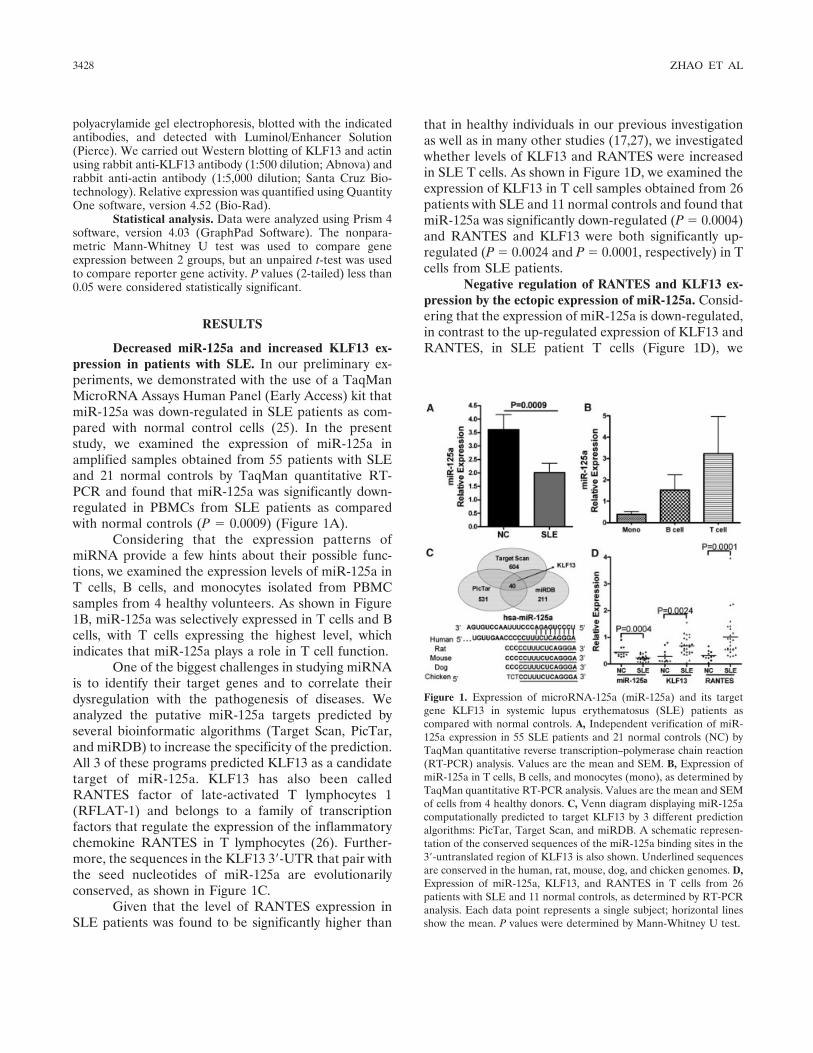
Decreased miR-125a and increased KLF13 ex-pression in patients with SLE. In our preliminary ex-periments, we demonstrated with the use of a TaqManMicroRNA Assays Human Panel (Early Access) kit thatmiR-125a was down-regulated in SLE patients as com-pared with normal control cells (25). In the presentstudy, we examined the expression of miR-125a inamplified samples obtained from 55 patients with SLEand 21 normal controls by TaqMan quantitative RT-PCR and found that miR-125a was significantly down-regulated in PBMCs from SLE patients as comparedwith normal controls (P � 0.0009) (Figure 1A).
Considering that the expression patterns ofmiRNA provide a few hints about their possible func-tions, we examined the expression levels of miR-125a inT cells, B cells, and monocytes isolated from PBMCsamples from 4 healthy volunteers. As shown in Figure1B, miR-125a was selectively expressed in T cells and Bcells, with T cells expressing the highest level, whichindicates that miR-125a plays a role in T cell function.
One of the biggest challenges in studying miRNAis to identify their target genes and to correlate theirdysregulation with the pathogenesis of diseases. Weanalyzed the putative miR-125a targets predicted byseveral bioinformatic algorithms (Target Scan, PicTar,and miRDB) to increase the specificity of the prediction.All 3 of these programs predicted KLF13 as a candidatetarget of miR-125a. KLF13 has also been calledRANTES factor of late-activated T lymphocytes 1(RFLAT-1) and belongs to a family of transcriptionfactors that regulate the expression of the inflammatorychemokine RANTES in T lymphocytes (26). Further-more, the sequences in the KLF13 3�-UTR that pair withthe seed nucleotides of miR-125a are evolutionarilyconserved, as shown in Figure 1C.
Given that the level of RANTES expression inSLE patients was found to be significantly higher than
that in healthy individuals in our previous investigationas well as in many other studies (17,27), we investigatedwhether levels of KLF13 and RANTES were increasedin SLE T cells. As shown in Figure 1D, we examined theexpression of KLF13 in T cell samples obtained from 26patients with SLE and 11 normal controls and found thatmiR-125a was significantly down-regulated (P � 0.0004)and RANTES and KLF13 were both significantly up-regulated (P � 0.0024 and P � 0.0001, respectively) in Tcells from SLE patients.
Negative regulation of RANTES and KLF13 ex-pression by the ectopic expression of miR-125a. Consid-ering that the expression of miR-125a is down-regulated,in contrast to the up-regulated expression of KLF13 andRANTES, in SLE patient T cells (Figure 1D), we
Figure 1. Expression of microRNA-125a (miR-125a) and its targetgene KLF13 in systemic lupus erythematosus (SLE) patients ascompared with normal controls. A, Independent verification of miR-125a expression in 55 SLE patients and 21 normal controls (NC) byTaqMan quantitative reverse transcription–polymerase chain reaction(RT-PCR) analysis. Values are the mean and SEM. B, Expression ofmiR-125a in T cells, B cells, and monocytes (mono), as determined byTaqMan quantitative RT-PCR analysis. Values are the mean and SEMof cells from 4 healthy donors. C, Venn diagram displaying miR-125acomputationally predicted to target KLF13 by 3 different predictionalgorithms: PicTar, Target Scan, and miRDB. A schematic represen-tation of the conserved sequences of the miR-125a binding sites in the3�-untranslated region of KLF13 is also shown. Underlined sequencesare conserved in the human, rat, mouse, dog, and chicken genomes. D,Expression of miR-125a, KLF13, and RANTES in T cells from 26patients with SLE and 11 normal controls, as determined by RT-PCRanalysis. Each data point represents a single subject; horizontal linesshow the mean. P values were determined by Mann-Whitney U test.
3428 ZHAO ET AL

hypothesized that miR-125a regulates RANTES expres-sion in T cells by targeting KLF13 and that deficientmiR-125a expression contributes to the elevatedRANTES and KLF13 expression observed in SLE pa-tients. To test this hypothesis, we used a mitogen(PHA-P)–activated T cell model in vitro to imitate theRANTES production process in T cells. After exposureto PHA-P, T cells undergo a 3–7-day maturation processinvolving activation, proliferation, and differentiation(28,29). Terminal differentiation is characterized by thegeneration of functional genes, including chemokineRANTES. As expected, our results showed thatRANTES secretion was induced after 3–5 days of stim-ulation with PHA-P, reached a peak on day 7, andincreased in a concentration-dependent manner withincreasing amounts of PHA-P. (Data on the kinetics ofPHA-P–induced RANTES in activated T lymphocytesare available upon request from the author.)
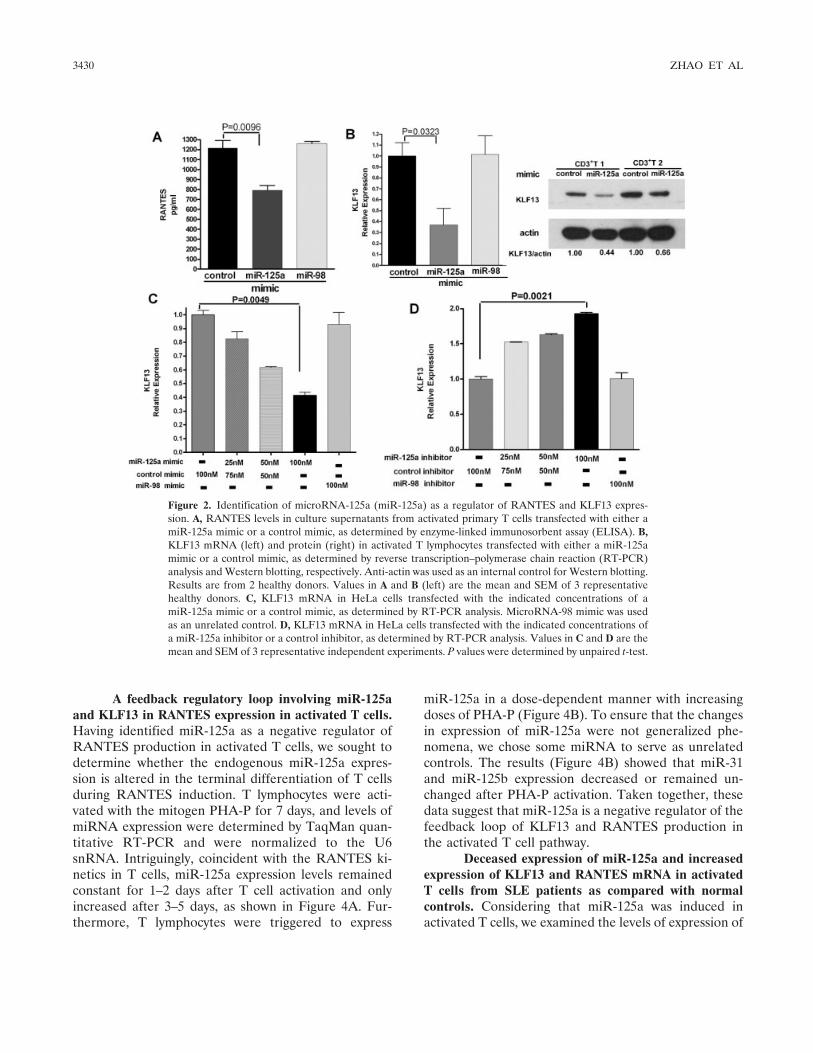
To determine whether miR-125a can alterRANTES expression, we transfected human primary Tcells with a miR-125a mimic or a control mimic and thenactivated those cells with PHA-P for 7 days, after whichRANTES levels in the cell supernatants were deter-mined by ELISA. The successful ectopic expression ofmiR-125a in primary T cells was verified by TaqManquantitative RT-PCR, which showed that the overex-pression of miR-125a following transfection resulted inan increase in miR-125a expression of �100-fold. (Dataon the expression of miR-125a before and after trans-fection in primary T cells are available upon requestfrom the author.) Notably, our results showed thatmiR-125a down-regulates RANTES expression as com-pared with that in control cells (Figure 2A). (Datashowing that overexpression of miR-125a repressed theexpression of RANTES mRNA in activated T cells areavailable upon request from the author.) Overexpressionof the unrelated control miR-98 had no effect on theRANTES expression. Transfection of T cells from dif-ferent individuals with a miR-125a mimic (100 nM)attenuated by �25%, on average, the amount ofRANTES released (Figure 2 and see below).
We next examined KLF13 expression by RT-PCR and Western blotting to further clarify the mech-anisms of the underlying miR-125a–mediated decreasedexpression of RANTES. We found that KLF13 expres-sion was also down-regulated in miR-125a mimic–transfected cells as compared with control mimic–transfected cells (Figure 2B). Overexpression of theunrelated control miR-98 had no effect on the expres-sion of mRNA for RANTES or KLF13.
HeLa cells were chosen as the cells in which to
examine whether miR-125a affects the stability ofKLF13 mRNA. HeLa cells produce moderate levels ofendogenous miR-125a and express mRNA for KLF13.By use of gain- and loss-of-function methods, weselectively transfected miR-125a mimic or inhibitor toinduce or repress the expression of miR-125a. Theresults of RT-PCR showed that overexpression ofmiR-125a inhibited KLF13 mRNA expression,whereas inhibition of miR-125a expression enhancedthe expression of KLF13 at the mRNA level (Figures2C and D). We used miR-98 as an unrelated control,and it had no effect on KLF13 expression. Whenincreasing concentrations of miR-125a mimic or in-hibitor were used, miR-125a regulated KLF13 expres-sion in a dose-dependent manner (Figures 2C and D).Taken together, our experiments indicate that miR-125a inhibits RANTES expression, possibly by target-ing the transcription factor KLF13.
Dose-dependent miRNA-125a targeting of the3�-UTR of KLF13. Bioinformatics analysis predicted thatthe KLF13 3�-UTR harbors at least 2 putative bindingsites for miR-125a, as depicted in Figure 3A. To deter-mine whether miR-125a directly regulates the 3�-UTRof KLF13, we cloned 2 genomic sequences from theKLF13 3�-UTR that contained the putative miR-125abinding sites, as well as another 2 sequences that con-tained a mutated version of the miR-125a binding sites(AGGG to UCCC). We also cloned 1 genomic sequencefrom the KLF13 3�-UTR that contained no miR-125abinding sites. We inserted these sequences separately,downstream of a Renilla luciferase reporter gene in thepsiCHECK-2 vector, as shown in Figure 3A. We thencotransfected HeLa cells with each luciferase reportervector and miRNA oligonucleotides and assessed theluciferase activity.
For luciferase reporter vectors that were fusedto the KLF13 3�-UTR fragment containing the miR-125a binding sites, increasing doses of a miR-125amimic reduced the luciferase activity, whereas in-creasing doses of miR-125a inhibitors increased theluciferase activity in a dose-dependent manner (Fig-ures 3B and C). The unrelated control miR-98 had noeffect on the luciferase activity of either luciferasereporter vector (P � 0.05). Mutation of the seedsequence of the predicted miR-125a binding siteabrogated this effect, as shown in Figure 3D. In addi-tion, miR-125a mimic also had no effect on the activity ofthe luciferase reporter vector–fused KLF13 3�-UTR frag-ment containing no miR-125a binding sites (Figure 3D).Taken together, these data suggest that miR-125a di-rectly targets the KLF13 3�-UTR.
MicroRNA-125a AND ELEVATED RANTES LEVELS IN SLE 3429
A feedback regulatory loop involving miR-125aand KLF13 in RANTES expression in activated T cells.Having identified miR-125a as a negative regulator ofRANTES production in activated T cells, we sought todetermine whether the endogenous miR-125a expres-sion is altered in the terminal differentiation of T cellsduring RANTES induction. T lymphocytes were acti-vated with the mitogen PHA-P for 7 days, and levels ofmiRNA expression were determined by TaqMan quan-titative RT-PCR and were normalized to the U6snRNA. Intriguingly, coincident with the RANTES ki-netics in T cells, miR-125a expression levels remainedconstant for 1–2 days after T cell activation and onlyincreased after 3–5 days, as shown in Figure 4A. Fur-thermore, T lymphocytes were triggered to express
miR-125a in a dose-dependent manner with increasingdoses of PHA-P (Figure 4B). To ensure that the changesin expression of miR-125a were not generalized phe-nomena, we chose some miRNA to serve as unrelatedcontrols. The results (Figure 4B) showed that miR-31and miR-125b expression decreased or remained un-changed after PHA-P activation. Taken together, thesedata suggest that miR-125a is a negative regulator of thefeedback loop of KLF13 and RANTES production inthe activated T cell pathway.
Deceased expression of miR-125a and increasedexpression of KLF13 and RANTES mRNA in activatedT cells from SLE patients as compared with normalcontrols. Considering that miR-125a was induced inactivated T cells, we examined the levels of expression of
Figure 2. Identification of microRNA-125a (miR-125a) as a regulator of RANTES and KLF13 expres-sion. A, RANTES levels in culture supernatants from activated primary T cells transfected with either amiR-125a mimic or a control mimic, as determined by enzyme-linked immunosorbent assay (ELISA). B,KLF13 mRNA (left) and protein (right) in activated T lymphocytes transfected with either a miR-125amimic or a control mimic, as determined by reverse transcription–polymerase chain reaction (RT-PCR)analysis and Western blotting, respectively. Anti-actin was used as an internal control for Western blotting.Results are from 2 healthy donors. Values in A and B (left) are the mean and SEM of 3 representativehealthy donors. C, KLF13 mRNA in HeLa cells transfected with the indicated concentrations of amiR-125a mimic or a control mimic, as determined by RT-PCR analysis. MicroRNA-98 mimic was usedas an unrelated control. D, KLF13 mRNA in HeLa cells transfected with the indicated concentrations ofa miR-125a inhibitor or a control inhibitor, as determined by RT-PCR analysis. Values in C and D are themean and SEM of 3 representative independent experiments. P values were determined by unpaired t-test.
3430 ZHAO ET AL

miR-125a, KLF13, and RANTES in activated T cellsisolated from SLE patients and normal controls. CD3�T cells were isolated by magnetic-activated cell sortingfrom the peripheral blood of 10 SLE patients and 8healthy donors. As shown in Figure 5, the results re-vealed that activated T cells from the SLE patients hadsignificantly reduced miR-125a levels and significantlyelevated RANTES and KLF13 mRNA levels as com-pared with activated T cells from the controls.
Reduction of elevated RANTES levels in T cellsfrom SLE patients by modification of miR-125a levels.The results described thus far demonstrated that miR-125a regulates the expression of RANTES in activated T
cells by targeting the critical transcription factor KLF13.We therefore explored whether manipulating the miR-125a levels could attenuate the elevated expression ofRANTES in T cells from SLE patients. We first exam-ined differences in RANTES protein expression in SLET cells as compared with normal control T cells, sincesuch an analysis had not previously been reported. Wefound that PHA-P–induced RANTES expression in SLET cells was higher than that observed in cells fromnormal controls (P � 0.0022) (Figure 6A). When Tlymphocytes were stimulated with the mitogen PHA-Pfor 7 days, the level of RANTES was 1,383 � 149.0pg/ml (mean � SEM) in T cells from normal controls
Figure 3. Confirmation of 2 specific microRNA-125a (miR-125a) target sites in the 3�-untranslatedregion (3�-UTR) of KLF13. A, A schematic representation of KLF13 mRNA shows 2 potential bindingsites in the 3�-UTR for miR-125a. The target sequences and an unrelated sequence in the 3�-UTR wereinserted into luciferase reporter plasmid and are indicated as site 1, site 2, and site 3. The mutants at eachpotential binding site were also generated. B, A miR-125a mimic suppressed the luciferase activity drivenby the site 1 construct, whereas a miR-125a inhibitor enhanced its activity in a dose-dependent manner.MicroRNA-98 was used as an unrelated control. C, A miR-125a mimic suppressed the luciferase activitydriven by the site 2 construct, whereas a miR-125a inhibitor enhanced its activity. D, A miR-125a mimichad no appreciable effect on luciferase expression driven by the site 1 or site 2 mutant constructs or by thesite 3 construct as compared with the effect of the control mimic (P � 0.05). Values in B–D are the meanand SEM fold change in relative luciferase activity from 3 independent experiments, each performed intriplicate. P values were determined by unpaired t-test.
MicroRNA-125a AND ELEVATED RANTES LEVELS IN SLE 3431

(n � 7), whereas in SLE T cells (n � 8), the level was1,967 � 83.04 pg/ml. In further analyses, T cells fromSLE patients were transfected with a miR-125a mimic ora control mimic, followed by PHA-P activation for 7days. As shown in Figure 6B, miR-125a transfectionreduced RANTES expression in activated T cells (n �6), from a mean � SEM of 1,711 � 158.2 pg/ml to1,306 � 155.4 pg/ml (P � 0.0411). This indicates thatmanipulation of the miR-125a levels could potentiallyprovide a therapeutic benefit for lupus patients.
DISCUSSION
A growing body of evidence indicates thatmiRNA are associated with the prognosis and progres-sion of several diseases and may serve as future targetsfor gene therapy. The role of miRNA in autoimmunity isalso beginning to be explored, and several recent studieshave unveiled the involvement of miRNA in rheumaticdiseases. For example, key components of the miRNApathway have been found to be targeted by disease-associated autoantibodies (30), and dysregulated expres-sion of miRNA has been linked to the pathogenesis ofdiseases (25,31). The results of our study establish thatmiR-125a is down-regulated, whereas its predicted tar-get gene, KLF13, is up-regulated, in patients with SLE.Further investigation provided insight into the mecha-nism and the role of miR-125a in controlling KLF13 andRANTES expression. Notably, miR-125a expression wasinduced in a time- and dose-dependent manner upon
PHA-P stimulation, which indicates that miR-125a actsas a negative regulator in the feedback loop of RANTESexpression in activated T cells.
Figure 4. Kinetics of phytohemagglutinin protein (PHA-P)–inducedmicroRNA-125a (miR-125a) in resting and activated T lymphocytes.A, T lymphocytes were triggered with either phosphate buffered saline(PBS) or mitogen (40 �g/ml of PHA-P) for the indicated time, usingPBS as the control. Levels of miR-125a expression were determined byreverse transcription–polymerase chain reaction (RT-PCR) analysisover a period of 7 days. Results were normalized to U6 small nuclearRNA (U6 snRNA). B, T lymphocytes were exposed to the mitogen(PHA-P) at the indicated concentrations for 3 days. Levels of miR-125a, miR-125b, and miR-31 expression were determined by TaqManquantitative RT-PCR analysis in PHA-P–stimulated T cells. Resultswere normalized to U6 snRNA. Values are the mean � SEM of 3representative experiments.
Figure 5. Expression of microRNA-125a (miR-125a), KLF13, andRANTES in activated T cells from systemic lupus erythematosus (SLE)patients as compared with normal controls. A, RANTES mRNA fromactivated T cells derived from 10 SLE patients and 8 normal controls wasdetected by reverse transcription–polymerase chain reaction (RT-PCR)analysis. B, KLF13 expression in the samples used in A was detected byRT-PCR analysis. C, Expression of miR-125a in the samples used in A wasdetected by TaqMan quantitative RT-PCR analysis. Horizontal linesshow the mean. P values were determined by Mann-Whitney U test.
Figure 6. Augmented RANTES protein production and its regulationby microRNA-125a (miR-125a) in phytohemagglutinin protein (PHA-P)–activated systemic lupus erythematosus (SLE) T cells. A, RANTESprotein from 8 SLE patients and 7 normal controls released into thesupernatant by activated T cells was determined by enzyme-linkedimmunosorbent assay (ELISA). Horizontal lines show the mean. B,Overexpression of miR-125a repressed the expression of RANTES inpaired samples of activated T cells from 6 SLE patients as comparedwith control conditions in the absence of miR-125a overexpression.RANTES levels in culture supernatants were detected by ELISA. Pvalues were determined by Mann-Whitney U test.
3432 ZHAO ET AL

RANTES is a typical CC (alpha) family chemo-kine, which plays an important role in the immunopatho-genesis of SLE and a myriad of disorders, includingacquired immunodeficiency syndrome (17,32). As previ-ously reported, RANTES is induced after the activationof T cells for 3–5 days with the mitogen PHA-P, which issimilar to the induction of proteins involved in T cellterminal differentiation, including perforin, granulysin,and granzymes A and B (19,33). Blanco et al (34)reported an increase in the expression of perforin andgranzyme B in activated CD8� T cells from SLEpatients. Our investigation showed that SLE T cellsproduced higher levels of RANTES, which allowed us tobetter delineate the altered behavior of these cells (35).
Since transcription factor KLF13 orchestrates theinduction of RANTES in activated T cells (26), whichhas been verified by the finding of reduced RANTESexpression in KLF13-deficient mice (36), we examinedKLF13 expression in SLE patients. We found thatKLF13 expression was significantly higher in SLE pa-tients than that in normal controls (Figure 1D), whichpartly explains the higher expression of RANTES inSLE patients. Interestingly, the expression of KLF13 inT cells is regulated by a posttranscriptional mechanismthrough its 5�-UTR, as reported previously (37). In thisstudy, we demonstrated a novel and evolutionarily con-served role of miRNA in the posttranscriptional regula-tion of KLF13 expression in T cells. Gain- and loss-of-function studies showed that miR-125a regulated KLF13expression. In addition, our reporter assays stronglysuggested that KLF13 was the direct target of miR-125a,with one or both of the binding sites being functional formiR-125 regulation.
MicroRNA have been identified as importantnegative regulators in the inflammatory response (38–40). Very recently, Toll-like receptor (TLR)–inducedmiRNA (e.g., miR-146 and miR-147) were demon-strated to prevent excessive TLR-stimulated inflamma-tory responses (38–39). Here, we found that miR-125aand a transcription factor interacted to form regulatorynetworks in the T cell activation process. (Illustration isavailable upon request from the author.) It is thereforepossible that the expression of miR-125a is alteredduring the T cell activation process. Interestingly, induc-tion of miR-125a in terminally differentiated T cells,which begins late after T cell activation by specificantigen or mitogen for 3–5 days, also occurs in a time-and dose-dependent manner, as does the induction ofKLF13 and RANTES. Thus, under physiologic condi-tions, it is well-established that PHA-P stimulation in-duces miR-125a to prevent excessive production of
RANTES and may act as a negative regulator in thefeedback loop of RANTES induction in activated T cellsby fine-tuning the expression of KLF13.
The deficient expression of miR-125a may partlyexplain the high levels of RANTES expression in SLEpatients and suggests that the introduction of miR-125amay help to reduce this high level of RANTES expres-sion in SLE, as was verified by our in vitro experiments.Due to the ability of miR-125a to potentially influencethe production of the inflammatory chemokineRANTES, therapeutic strategies based on the modula-tion of miR-125a activity hold great promise for thetreatment of patients with rheumatic diseases.
Initially identified as a tumor-related miRNA,miR-125a has been implicated in targeting the c-ErbB-2and c-ErbB-3 oncogenic proteins, and it has been shownto suppress the migration, invasiveness, and other criti-cal features of the malignant cell phenotype, as in cancer(41). MicroRNA-125a in hematopoiesis has been high-lighted as 1 of the top 2 underexpressed miRNA inhuman natural Treg cells (42), and miR-125a is one ofmost up-regulated miRNA during human monocyte-derived dendritic cell differentiation (43). To the best ofour knowledge, the functions of miR-125a in the hema-topoietic and immune systems are still unclear. It wouldbe interesting to determine the function of miR-125a inTreg cells and dendritic cells, as well as its contributionto the pathogenesis of lupus. Studies in knockout andtransgenic animal models would further identify the roleof miR-125a in rheumatic diseases.
To explore possible mechanisms of the deficientexpression of miR-125a in SLE, we characterized themiR-125a promoter region using FirstEF software(http://rulai.cshl.org/tools/FirstEF/), and we identified apotential CpG island in the predicted promoter ofmiR-125a using CpG Island Searcher software (http://cpgislands.usc.edu/). As we know, abnormal epigeneticalterations, including modifications of DNA methyl-ation, contribute to the etiology of SLE (44). It would beof interest to investigate the methylation status of themiR-125a gene in SLE.
In addition, since identifying the putative tran-scription factors that bind a promoter represents animportant step in understanding gene regulation, wesearched transcription factor binding sites in the miR-125a promoter using TFSearch software (http://mbs.cbrc.jp/research/db/TFSEARCH.html), whichshowed that the upstream region of miR-125a containsseveral putative Sp-1 motif–binding sites. As a eukary-otic Sp-1 like zinc-finger transcriptional protein, KLF13binds conservatively to GC-rich or GT-rich cis DNA
MicroRNA-125a AND ELEVATED RANTES LEVELS IN SLE 3433

elements and interacts with promoter-specific activatorsor repressors of transcription (45). Therefore, it may bepossible to identify transcription factors that bind to thepromoter region of miR-125a, which could clarify themechanism of miR-125a induction in activated T cells.
In conclusion, the findings of this study providenew insights into the role of miR-125a in lupus. Ourstudy shows that miR-125a is involved in a feedbackregulatory loop that controls RANTES expression interminally differentiated T cells. Our findings suggestthat miR-125a could serve as therapeutic targets for thetreatment of SLE via the regulation of the production ofinflammatory chemokines.
ACKNOWLEDGMENTS
We thank the patients, the healthy volunteer donors,and the rheumatologists at Shanghai Renji Hospital for theirparticipation in this study. We also thank Dr. Haibo Zhou andWen Pan for their helpful comments.
AUTHOR CONTRIBUTIONS
All authors were involved in drafting the article or revising itcritically for important intellectual content, and all authors approvedthe final version to be published. Dr. Shen had full access to all of thedata in the study and takes responsibility for the integrity of the dataand the accuracy of the data analysis.Study conception and design. Shen.Acquisition of data. Zhao, Tang, Qu, Cui, Shujun Wang, Lijia Wang,Li.Analysis and interpretation of data. Zhao, Tang, Qu, Cui, Luo, Huang,Chen, Shen.
REFERENCES
1. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, andfunction. Cell 2004;116:281–97.
2. Ambros V. The functions of animal microRNAs. Nature 2004;431:350–5.
3. Yu D, Rao S, Tsai LM, Lee SK, He Y, Sutcliffe EL, et al. Thetranscriptional repressor Bcl-6 directs T follicular helper celllineage commitment. Immunity 2009;31:457–68.
4. Liston A, Lu LF, O’Carroll D, Tarakhovsky A, Rudensky AY.Dicer-dependent microRNA pathway safeguards regulatory T cellfunction. J Exp Med 2008;205:1993–2004.
5. Pauley KM, Cha S, Chan EK. MicroRNA in autoimmunity andautoimmune diseases. J Autoimmun 2009;32:189–94.
6. Baltimore D, Boldin MP, O’Connell RM, Rao DS, Taganov KD.MicroRNAs: new regulators of immune cell development andfunction. Nat Immunol 2008;9:839–45.
7. Vinuesa CG, Rigby RJ, Yu D. Logic and extent of miRNA-mediated control of autoimmune gene expression. Int Rev Immu-nol 2009;28:112–38.
8. Czech MP. MicroRNAs as therapeutic targets. N Engl J Med2006;354:1194–5.
9. Sallusto F, Lenig D, Mackay CR, Lanzavecchia A. Flexibleprograms of chemokine receptor expression on human polarized Thelper 1 and 2 lymphocytes. J Exp Med 1998;187:875–83.
10. Moore BB, Keane MP, Addison CL, Arenberg DA, Strieter RM.
CXC chemokine modulation of angiogenesis: the importance ofbalance between angiogenic and angiostatic members of thefamily. J Investig Med 1998;46:113–20.
11. Bonecchi R, Bianchi G, Bordignon PP, D’Ambrosio D, Lang R,Borsatti A, et al. Differential expression of chemokine receptorsand chemotactic responsiveness of type 1 T helper cells (Th1s) andTh2s. J Exp Med 1998;187:129–34.
12. Charo IF, Ransohoff RM. The many roles of chemokines andchemokine receptors in inflammation. N Engl J Med 2006;354:610–21.
13. Hamilton TA, Novotny M, Datta S, Mandal P, Hartupee J, TeboJ, et al. Chemokine and chemoattractant receptor expression:post-transcriptional regulation. J Leukoc Biol 2007;82:213–9.
14. Pauley KM, Chan EK. MicroRNAs and their emerging roles inimmunology. Ann N Y Acad Sci 2008;1143:226–39.
15. Crow MK. Collaboration, genetic associations, and lupus erythem-atosus. N Engl J Med 2008;358:956–61.
16. Perez de Lema G, Maier H, Nieto E, Vielhauer V, Luckow B,Mampaso F, et al. Chemokine expression precedes inflammatorycell infiltration and chemokine receptor and cytokine expressionduring the initiation of murine lupus nephritis. J Am Soc Nephrol2001;12:1369–82.
17. Eriksson C, Eneslatt K, Ivanoff J, Rantapaa-Dahlqvist S,Sundqvist KG. Abnormal expression of chemokine receptors onT-cells from patients with systemic lupus erythematosus. Lupus2003;12:766–74.
18. Appay V, Rowland-Jones SL. RANTES: a versatile and contro-versial chemokine. Trends Immunol 2001;22:83–7.
19. Schall TJ, Jongstra J, Dyer BJ, Jorgensen J, Clayberger C, DavisMM, et al. A human T cell-specific molecule is a member of a newgene family. J Immunol 1988;141:1018–25.
20. Meller S, Winterberg F, Gilliet M, Muller A, Lauceviciute I,Rieker J, et al. Ultraviolet radiation–induced injury, chemokines,and leukocyte recruitment: an amplification cycle triggering cuta-neous lupus erythematosus. Arthritis Rheum 2005;52:1504–16.
21. Petrek M, Pantelidis P, Southcott AM, Lympany P, Safranek P,Black CM, et al. The source and role of RANTES in interstitiallung disease. Eur Respir J 1997;10:1207–16.
22. Moore KJ, Wada T, Barbee SD, Kelley VR. Gene transfer ofRANTES elicits autoimmune renal injury in MRL-Faslpr mice.Kidney Int 1998;53:1631–41.
23. Tian S, Li J, Wang L, Liu T, Liu H, Cheng G, et al. Urinary levelsof RANTES and M-CSF are predictors of lupus nephritis flare.Inflamm Res 2007;56:304–10.
24. Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, RothfieldNF, et al. The 1982 revised criteria for the classification of systemiclupus erythematosus. Arthritis Rheum 1982;25:1271–7.
25. Tang Y, Luo X, Cui H, Ni X, Yuan M, Guo Y, et al. MicroRNA-146A contributes to abnormal activation of the type I interferonpathway in human lupus by targeting the key signaling proteins.Arthritis Rheum 2009;60:1065–75.
26. Song A, Chen YF, Thamatrakoln K, Storm TA, Krensky AM.RFLAT-1: a new zinc finger transcription factor that activatesRANTES gene expression in T lymphocytes. Immunity 1999;10:93–103.
27. Fu Q, Chen X, Cui H, Guo Y, Chen J, Shen N, et al. Associationof elevated transcript levels of interferon-inducible chemokineswith disease activity and organ damage in systemic lupus erythem-atosus patients. Arthritis Res Ther 2008;10:R112.
28. Crabtree GR. Contingent genetic regulatory events in T lympho-cyte activation. Science 1989;243:355–61.
29. Ullman KS, Northrop JP, Verweij CL, Crabtree GR. Transmissionof signals from the T lymphocyte antigen receptor to the genesresponsible for cell proliferation and immune function: the missinglink. Annu Rev Immunol 1990;8:421–52.
30. Jakymiw A, Ikeda K, Fritzler MJ, Reeves WH, Satoh M, Chan EK.
3434 ZHAO ET AL

Autoimmune targeting of key components of RNA interference.Arthritis Res Ther 2006;8:R87.
31. Nakamachi Y, Kawano S, Takenokuchi M, Nishimura K, Sakai Y,Chin T, et al. MicroRNA-124a is a key regulator of proliferationand monocyte chemoattractant protein 1 secretion in fibroblast-like synoviocytes from patients with rheumatoid arthritis. ArthritisRheum 2009;60:1294–304.
32. Alkhatib G, Combadiere C, Broder CC, Feng Y, Kennedy PE,Murphy PM, et al. CC CKR5: a RANTES, MIP-1�, MIP-1�receptor as a fusion cofactor for macrophage-tropic HIV-1. Sci-ence 1996;272:1955–8.
33. Ortiz BD, Nelson PJ, Krensky AM. Switching gears during T-cellmaturation: RANTES and late transcription. Immunol Today1997;18:468–71.
34. Blanco P, Pitard V, Viallard JF, Taupin JL, Pellegrin JL, MoreauJF. Increase in activated CD8� T lymphocytes expressing perforinand granzyme B correlates with disease activity in patients withsystemic lupus erythematosus. Arthritis Rheum 2005;52:201–11.
35. Crispin JC, Tsokos GC. Novel molecular targets in the treatmentof systemic lupus erythematosus. Autoimmun Rev 2008;7:256–61.
36. Zhou M, McPherson L, Feng D, Song A, Dong C, Lyu SC, et al.Kruppel-like transcription factor 13 regulates T lymphocyte sur-vival in vivo. J Immunol 2007;178:5496–504.
37. Nikolcheva T, Pyronnet S, Chou SY, Sonenberg N, Song A,Clayberger C, et al. A translational rheostat for RFLAT-1 regu-lates RANTES expression in T lymphocytes. J Clin Invest 2002;110:119–26.
38. Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-�B-dependent induction of microRNA miR-146, an inhibitor targeted
to signaling proteins of innate immune responses. Proc Natl AcadSci U S A 2006;103:12481–6.
39. Liu G, Friggeri A, Yang Y, Park YJ, Tsuruta Y, Abraham E.miR-147, a microRNA that is induced upon Toll-like receptorstimulation, regulates murine macrophage inflammatory re-sponses. Proc Natl Acad Sci U S A 2009;106:15819–24.
40. O’Connell RM, Taganov KD, Boldin MP, Cheng G, Baltimore D.MicroRNA-155 is induced during the macrophage inflammatoryresponse. Proc Natl Acad Sci U S A 2007;104:1604–9.
41. Scott GK, Goga A, Bhaumik D, Berger CE, Sullivan CS, Benz CC.Coordinate suppression of ERBB2 and ERBB3 by enforcedexpression of micro-RNA miR-125a or miR-125b. J Biol Chem2007;282:1479–86.
42. Rouas R, Fayyad-Kazan H, El Zein N, Lewalle P, Rothe F, SimionA, et al. Human natural Treg microRNA signature: role ofmicroRNA-31 and microRNA-21 in FOXP3 expression. Eur J Im-munol 2009;39:1608–18.
43. Hashimi ST, Fulcher JA, Chang MH, Gov L, Wang S, Lee B.MicroRNA profiling identifies miR-34a and miR-21 and theirtarget genes JAG1 and WNT1 in the coordinate regulation ofdendritic cell differentiation. Blood 2009;114:404–14.
44. Huber LC, Stanczyk J, Jungel A, Gay S. Epigenetics in inflamma-tory rheumatic diseases [review]. Arthritis Rheum 2007;56:3523–31.
45. Scohy S, Gabant P, Van Reeth T, Hertveldt V, Dreze PL, VanVooren P, et al. Identification of KLF13 and KLF14 (SP6), novelmembers of the SP/XKLF transcription factor family. Genomics2000;70:93–101.
MicroRNA-125a AND ELEVATED RANTES LEVELS IN SLE 3435




















