
Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells for the Production of...
25
85 M.A. Shiffman et al. (eds.), Stem Cells in Aesthetic Procedures, DOI 10.1007/978-3-642-45207-9_6, © Springer-Verlag Berlin Heidelberg 2014 T.C. Tran, M.D., Ph.D (*) Biomaterial Research Laboratory, Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants, Pham Ngoc Thach University of Medicine, 86/2 Thanh Thai St., 10th Dist., Ho Chi Minh City, Viet Nam Department of Histology – Anapathology, School of Medicine, Vietnam National University – Ho Chi Minh City, Quarter 6, Linh Trung Ward, Ho Chi Minh City, Thu Duc District, Viet Nam Department of Histology and Embryology, Faculty of Medicine, The University of Medicine and Pharmacy at Ho Chi Minh City, 217 Hong Bang St. 5th Dist., Ho Chi Minh City, Viet Nam e-mail: [email protected] C. Gargiulo, Ph.D., M.Sc. Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants, Pham Ngoc Thach University of Medicine, Ho Chi Minh City, Viet Nam University of Western Australia School of Anatomy and Human Biology, Perth, Australia e-mail: [email protected] T.D. Huynh, M.A. Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants, Pham Ngoc Thach University of Medicine, Ho Chi Minh City, Viet Nam e-mail: [email protected] 6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells for the Production of Compatible and Safe Biomaterial Crucial in Cosmetic, Reconstructive, and Regenerative Medicine Toai Cong Tran, Ciro Gargiulo, Thao Duy Huynh, Khanh Hong Thien Bui, Luis Filgueira, and Douglas M. Strong K.H.T. Bui, M.D., Ph.D Orthopaedic Department, University Medical Center, University of Medicine and Pharmacology, HCM City, 215–217 Hong Bang St., Dist.5, HCM City, Viet Nam e-mail: [email protected] L. Filgueira, M.D. School of Anatomy, Physiology and Human Biology, The University of Western Australia, 35 Stirling Highway, Perth, WA M309, Australia Anatomy Unit, Department of Medicine, University of Fribourg, CH-1700, Fribourg, Switzerland e-mail: luis.fi[email protected], lfi[email protected], luis.fi[email protected] D.M. Strong, Ph.D Department of Orthopeadics and Sports Medicine, University of Washington School of Medicine, 18624 94th Ave West, Edmonds, WA 98020, USA e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells for the Production of...

85M.A. Shiffman et al. (eds.), Stem Cells in Aesthetic Procedures,DOI 10.1007/978-3-642-45207-9_6, © Springer-Verlag Berlin Heidelberg 2014
T. C. Tran , M.D., Ph.D (*) Biomaterial Research Laboratory, Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants , Pham Ngoc Thach University of Medicine , 86/2 Thanh Thai St., 10th Dist. , Ho Chi Minh City , Viet Nam
Department of Histology – Anapathology , School of Medicine, Vietnam National University – Ho Chi Minh City , Quarter 6, Linh Trung Ward , Ho Chi Minh City, Thu Duc District , Viet Nam
Department of Histology and Embryology, Faculty of Medicine , The University of Medicine and Pharmacy at Ho Chi Minh City , 217 Hong Bang St. 5th Dist. , Ho Chi Minh City , Viet Nam e-mail: [email protected]
C. Gargiulo , Ph.D., M.Sc. Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants , Pham Ngoc Thach University of Medicine , Ho Chi Minh City , Viet Nam
University of Western Australia School of Anatomy and Human Biology , Perth , Australia e-mail: [email protected]
T. D. Huynh , M.A. Department of Histology, Embryology, Genetics and Biotechnology for Tissue Transplants , Pham Ngoc Thach University of Medicine , Ho Chi Minh City , Viet Nam e-mail: [email protected]
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells for the Production of Compatible and Safe Biomaterial Crucial in Cosmetic, Reconstructive, and Regenerative Medicine
Toai Cong Tran , Ciro Gargiulo , Thao Duy Huynh , Khanh Hong Thien Bui , Luis Filgueira , and Douglas M. Strong
K. H. T. Bui , M.D., Ph.D Orthopaedic Department , University Medical Center, University of Medicine and Pharmacology, HCM City , 215–217 Hong Bang St., Dist.5 , HCM City , Viet Nam e-mail: [email protected]
L. Filgueira , M.D. School of Anatomy, Physiology and Human Biology, The University of Western Australia , 35 Stirling Highway , Perth , WA M309 , Australia
Anatomy Unit, Department of Medicine , University of Fribourg , CH-1700 , Fribourg , Switzerland e-mail: luis.fi [email protected], lfi [email protected], luis.fi [email protected]
D. M. Strong , Ph.D Department of Orthopeadics and Sports Medicine , University of Washington School of Medicine , 18624 94th Ave West, Edmonds , WA 98020 , USA e-mail: [email protected]

86
6.1 Introduction
Several published works have confi rmed the pos-sibility to obtain a population of stem cells from adipose tissue that share similar features and morphogenetic profi le of MSCs from known sources such as BM, UCB, amniotic fl uid, scalp tissue, placenta, and synovial tissue [ 1 – 13 ]. A common pattern of all MSC cultures in vitro is a combination of specifi c and homogeneous traits including the formation of uniform colonies with a typical fi broblast spindle-like shape, self- renewal capacity, high viability, and multilineage ability such as osteoblasts, adipocytes, keratino-cytes, cardiomyocytes, chondrocytes, hepato-cytes, and tenocytes [ 4 – 6 , 14 – 18 ]. In general, they do not express defi nite markers of hemato-poietic cell surface phenotype and are positive for integrins and adhesion molecules, positive for matrix receptors, and positive for few very spe-cifi c markers such as CD13 (APN), CD28, CD166 (ALCAM), CD44, CD73, CD90, CD105 and CD166, vimentin, desmin, Runx2, osterix (OX), and HLA class I [ 4 , 10 , 11 , 14 , 15 , 19 , 20 ].
One of the most intriguing traits of MSCs is their immune-modulatory capacity that is of rel-evant clinical importance as they may be trans-planted without the need of HLA matching between donor and recipient which is strictly required in allogeneic adult stem cell procedures, in BM transfusion, or in organ transplants [ 3 , 21 – 26 ]. While each MSC subgroup shows an identical immune-phenotypic profi le, each set clearly expresses remarkable quantitative and qualitative differences, revealing a sort of hetero-geneity linked to a specifi c genetic set that con-trol different activities within the system [ 5 , 11 , 13 , 18 , 25 ]. The possibility to obtain MSCs from different sources other than the more classic ones like BM and UCB is of vital importance in the fi eld of regenerative medicine and may provide the opportunity to avoid unnecessary invasive procedures or to explore a better sustainable ther-apeutic strategy which is based on multiple pos-sible choices considered on a case-by-case basis [ 13 , 18 , 26 – 28 ]. Adipose tissue, like marrow, is of mesoderm origin; it starts to form during the last trimester of intrauterine life and contains a
variety of stromal cells that group into microvas-cular endothelial cells, smooth muscle cells, and MSCs that can be isolated by enzymatic diges-tion and centrifugation [ 18 , 24 , 29 , 30 ]. Adipose tissue is to be considered a complex structure that performs on multiple levels as an energy store and as an endocrine organ capable of producing and secreting a large number of molecules includ-ing leptin, adiponectin, and resistin which control endothelial functionality and may infl uence the vascular system [ 29 , 31 , 32 ]. Data from our study, in line with published research, have confi rmed that fat tissue compared to BM contains more MSCs. From 1 g of tissue, you may collect 5 × 10 4–5 MSCs which is 500-folds larger than 1 g of MSCs obtained from BM [ 24 , 33 , 34 ]. hATM-SCs, as their respective equivalent, have the abil-ity to differentiate into diverse cell lines such as myoblasts, chondroblasts, cardiomyocytes, hepa-tocytes, adipocytes, and osteoblasts maintaining a unique plasticity typical of all MSCs [ 13 , 19 , 24 , 30 , 35 , 36 ]. This study, in line with edited data, has demonstrated that hATMSCs, at least in vitro culture, behave in a similar manner and are able to home in a bone-like environment such as sea coral scaffold [ 10 , 19 , 27 ].
In line with other studies and with our previ-ously published work, the authors have tried to establish a method that allows the use of these cells in combination with bio-scaffolds to be used in the reconstruction of hard tissue such as bone and in scaffolds in repairing damaged soft tissue and semihard tissues such as cartilage and inter-vertebral disk bulbs [ 10 , 37 , 38 ]. Therefore, the fi rst task was to use hATMSCs and adipocytes and osteoblasts seeded onto marine coral Porites lutea to generate a hard structure bio-scaffold; the second task was to generate a soft scaffold using a fi brin gel seeded with human adipocyte cells to generate a soft structure scaffold. The fi brin gel has been generated by using autologous patient’s blood; from this sample, fi brinogen and thrombin have been isolated and then mixed in unique solution in a ratio of 1:1 [ 37 – 40 ].
However, despite the promising results from preliminary clinical trials, our concern is about the possible contribution of hATMSCs in vivo as an active player in the process of vascular
T.C. Tran et al.

87
calcifi cation and related infl ammatory condi-tions, as some authors assert, and it is well known that there are more specifi c differences in the metabolic properties and modalities of gene expression within the different fat deposits and these could be associated with different patho-genic conditions [ 29 ]. By now, a number of sci-entists have shown that there might be a correlation between fat tissue mass and the calci-fi cation of vascular epithelium and, in particular, the role that leptin, some of pro-infl ammatory cytokines such as IL6, TNFα, IL1-β, and mole-cules as glucocorticoids (such as dexametha-sone), vitamin D3, and homocysteine may eventually play, either in vitro or in vivo, in the differentiation mechanism of both local hATM-SCs, adipocytes, and endothelial cells into cal-cium matrix producer osteoblasts and in related infl ammatory processes [ 32 , 41 – 49 ]. The dyna-mism of the vascular calcifi cation recalls that one within the bone environment, with identical pro-cess, the same players, stimulation, and growth factors [ 46 , 48 ].
6.2 Human Adipose Tissue Mesenchymal Stem Cells
6.2.1 In Vitro Expansion of Human Adipose Tissue MSCs (Fig. 6.1 )
ATMSCs with multilineage capacity were isolated as earlier [ 18 ]. Fat tissues were originally collected from consented donors and tested for HIV, HBV, HCV, and VDRL. Samples were collected in apposite container including DMEM/F12 (Gibco, Grand Island NY, USA), FBS (10 %, Gibco, Grand Island NY, USA), and gentamicin (50 μg/ml). Tissues were successively transferred to the labo-ratory where they were placed into Petri dishes and washed with a PBS-streptomycin- penicillin solution three times; then samples were washed again in PBS with no antibiotic and immersed into a basic medium composed of DMEM/F12 (85.025 mL), FBS (5 %, 10 mL), HEPES (15 mM, 1.5 mL, 1 M), NaHCO 3 (14 nM, 0.35 mL, 4.10 3 nM), biotin (33 μL, 1.65 mL, 2.10 3 μM), d -panto (17 μM, 0.85 mL, 2.10 3 μM), gentamicin
(40.000 μg/mL, 0.125 mL), and penicillin- streptomycin (200×, 0.5 mL). Samples were selec-tively worked to remove foreign tissues, were fi nely chopped, and transferred into an enzymatic solution composed of dispase- collagenase solu-tion (3:1 ratio) and incubated for 90 min at 37 °C. Successively, the sample was centrifuged (Universal 32, Zentrifugen-Germany) for 5 min at a speed of 3,000 rpm, the fl oating materials were removed, and the sedimentation was collected and fi ltered by a cell strainer with diameter of 70 μM (BD FalconTM). This fi ltered solution was newly centrifuged at a speed of 3,000 rpm for 5 min, the fl oating suspension was removed, and the sedi-ment was collected by adding the basic medium; a small fraction of cells were collected and counted by Trypan Blue (Sigma). The rest of cells were successively cultured into T-25 cm 2 fl ask (Nunc, Wiesbaden, Germany) with a medium composed of DMEM/F12 (81.51 mL), FBS (10 %, 10 mL), HEPES (15 mM, 1.5 mL, 1 M), NaHCO 3 (14 nM, 0.35 mL, 4.10 3 nM), biotin (33 μL, 1.65 mL, 2.10 3 μM), d -panto (17 μM, 0.85 mL, 2.10 3 μM), human insulin (66 nM, 1.95 mL, 34.10 2 nM), triiodo- l -thyronine (1 nM, 65 μl, 0.065 mL, 1.5.10 3 nM), human transferrin (10 μg/mL, 2 mL, 500 μg/mL), and gentamicin (40.000 μg/mL, 0.125 mL). Cell cultures were incubated into an apposite incu-bator at 37 °C with 5 % CO 2 . The medium was replaced every 3 or 4 days. After a few days, a colony with the typical spindle-like shape started to form, and at 80–90 % of confl uence cells were harvested by enzymatic digestion by using a com-bination of Trypsin-EDTA (Gibco BRL, USA); suspended cells were centrifuged for 5 min at 3,000 rpm, and the pellet was removed and cul-tured in T-25 cm 2 fl ask with DMEM plus 10 % FBS. The average amount of adherent cells was 0.9 (±0.37) × 10 6 cells per ml of crushed adipose tissue. Cell viability at the time of passage was nearly 100 %. The MSCs were tested for specifi c CD markers to confi rm their phenotype identity; cells resulted negative for hematopoietic cell markers including CD14, CD45, and HLA-DR and were positive for exclusive markers such as CD13, CD44, CD73, CD90, CD105, and CD166. MSCs were subcultured several times and photographed.
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

88
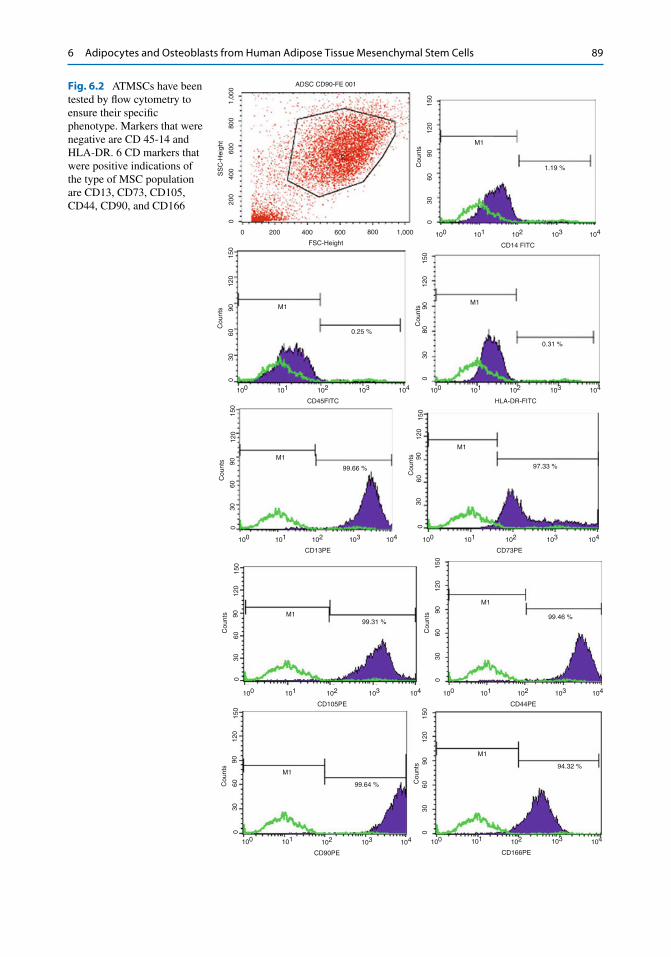
6.2.1.1 CD Markers Profi le (Fig. 6.2 ) A total of 9 cell cultures were harvested at the second generation and tested to corroborate their phenotypic characterization by fl ow cytometry procedures. hATMSCs were previ-ously tested to three specifi c hematopoietic markers CD14, CD45, and HLA-DR (negative) and subsequently tested for markers such as CD13, CD44, CD73, CD90, CD105, and CD166 (positive). By these results, it is clearly confi rmed that hATMSCs share similarities with their counterparts from hUCB or hBM as one can fi nd in other published works [ 16 , 18 , 19 , 50 ].
Second-generation hATMSCs were harvested at day 15 and stained with Giemsa stain in order to confi rm their morphology and profi le (Fig. 6.3 ). The hATMSCs show the unique clas-sic spindle- like shape of distinctive MSCs. These cells show an amazing survival grade and a high expansion and proliferative rate; we successfully obtained over six dozen cultures from a single fat tissue sample; from a mother cell culture, it was possible to reach up to the seventh to ninth generation with cultures able to maintain a con-stant rate growth with a very low number of losses.
6.2.2 In Vitro Differentiation of hATMSCs to Adipocytes
MSCs were harvested by enzymatic digestion by Trypsin-EDTA at the second passage and
were transferred in fl ask culture with a specifi c adipogenic medium containing DMEM/F12 (80.945 mL), HEPES (15 mM, 1.5 mL, 1 M), NaHCO 3 (14 nM, 0.35 mL, 4.10 3 nM), biotin (33 μL, 1.65 mL, 2.10 3 μM), d -panto (17 μM, 0.85 mL, 2.10 3 μM), human insulin (66 nM, 1.95 mL, 34.10 2 nM), triiodo- l -thyronine (1 nM, 65 μl, 0.065 mL/1.5.10 3 nM), human transferrin (10 μg/mL, 2 mL, 500 μg/mL), gen-tamicin (40.000 μg/mL, 0.125 mL), isobutyl- methylxanthine (0.005 μg/mL, 0.2 mL), hydrocortisone (100 nM, 0.365 mL, 365 μL), and dexamethasone (10 −8 M, 10 mL) (all from Sigma- Aldrich, USA). After a period of 31 days, cells appeared with the typical round shape containing small vesicles of fat; cells were tested positive for specifi c adipocyte stains, Oil Red and Nile red. Adipocytes were subcultured several times and photographed.
6.2.2.1 Cytochemical Stain: Oil Red At 90 % confl uence, second-generation hATM-SCs were harvested by enzymatic digestion and induced to differentiate into adipocytes, by using a specifi c adipogenic medium [ 17 , 19 ]. Cells stay in culture for almost a month and started to change shape at the second week to assume the complete mature adipocyte confor-mation at the end of 4th week showing the typi-cal fat droplets inside their cytoplasm. The adipocyte phenotype was confi rmed by using two specifi c cytochemical stains Nile Red and Oil Red (Fig. 6.4 ).
a b c
Fig. 6.1 Procedure of isolating MSCs from fresh human fat tissue sample by ( a ) enzymatic digestion and ( b ) centrifu-gation. ( c ) The suspended tissue has been collected and cultured in a proper medium
T.C. Tran et al.
89
M199.31 %
M1
99.46 %
100 101 102
CD105PE
103 104 100 101 102
CD44PE
103 104
Cou
nts
150
120
9060
300
Cou
nts
150
120
9060
300
M1
99.64 %
M1
94.32 %
100 101 102
CD90PE
103 104 100 101 102
CD166PE
103 104
Cou
nts
150
120
9060
300
Cou
nts
150
120
9060
300
M1
99.66 %
M1
97.33 %
100 101 102
CD13PE
103 104 100 101 102
CD73PE
103 104
Cou
nts
120
9060
300
Cou
nts
150
120
9060
300
150
100 101 102
CD45FITC
M1M1
0.25 %
0.31 %
103 104 100 101 102
HLA-DR-FITC
103 104
Cou
nts
Cou
nts
150
120
9060
300 0
3080
9012
015
0
ADSC CD90-FE 001
1,00
0
150
120
90
Cou
nts
6030
0
100 101 102
CD14 FITC
103 104
M1
1.19 %
800
600
400SS
C-H
eigh
t
200
00 200 400
FSC-Height
600 800 1,000
Fig. 6.2 ATMSCs have been tested by fl ow cytometry to ensure their specifi c phenotype. Markers that were negative are CD 45-14 and HLA-DR. 6 CD markers that were positive indications of the type of MSC population are CD13, CD73, CD105, CD44, CD90, and CD166
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

90
6.2.2.2 Nile Red by Fluorescence Electronic Microscope
At 90 % confl uence, second-generation hATM-SCs were harvested by enzymatic digestion and induced to differentiate into adipocytes, using a specifi c adipogenic medium [ 17 , 19 ]. Cells stay in culture for almost a month, they started to change shape at the second week to assume the complete mature adipocyte conformation at the end of 4th week showing the typical fat droplets inside their cytoplasm. At mature confl uence, cells were fi xed in 1 % paraformaldehyde in PBS and stained by Nile Red following the described procedure.
The reagents used are PBS 1 ml and permeabi-lization reagent 0.1 % Triton X in PBS for each specimen, containing 1 μl staining solution Nile Red fl uorescence. Slides were washed once in PBS in jar for 3 times for 5 min; once dried, it was added 100 μl of permeabilization reagent per slide to cover all sections for 1 min. Slides were washed in jar with PBS for other 3 min; once dried, it was added 1 μl of staining solution 1 per slide with 1 ml PBS to cover all sections for more than 10 min. Slides were dried, and it has been mounted the tissue by using Dako mounting medium, and cover slips sealed with nail varnish.
Fig. 6.3 ( Left and right ) Fibroblast spindle-like shape of ATMSCs, the morphology of these cells is typical of all MSCs population from BM, UCB, or placenta. ATMSCs are high proliferative cells with strong adherence ability (×200)
a b
Fig. 6.4 At 99 % confl uence, ATMSCs have been har-vested and differentiated into adipocytes. Cells remain in culture for 1 month showing ( a ) the typical round shape
with fat drops in their cytoplasm. ( b ) Mature adipocytes stained by Red Oil
T.C. Tran et al.

91
Abs
orba
nce
400 450 500 550
Wavelength (nm)
Flu
ores
cenc
e em
issi
on
600 650 700 750
Nile Red staining confi rmed that the cell cyto-plasm contains lipid particles and is very specifi c. This is the most effective method to identify cells as fat cells (Fig. 6.5 ).
6.2.3 In Vitro Differentiation of hATMSCs to Osteoblasts
MSCs from adipose tissue were harvested at the second passage and were induced to differentiate into osteoblasts using a specifi c osteogenic medium composed of IMDM (63.05 mL,
a b
Fig. 6.5 Results of Nile Red staining for adipocytes after differentiation. ( a ) Negative control samples, mesenchy-mal stem cells. ( b ) Adipocytes are differentiated after staining with Nile Red showing that the cytoplasm of
many cells appeared red - orange lipid particles, lipid par-ticles accounted for nearly all the cell cytoplasm, and the nucleus was push offset to one side of the cell
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

92
13.05 %), penicillin-streptomycin (8.70 ml), FBS (13.05 mL), dexamethasone (10.0 mL, 10 −7 M), ascorbic acid (1.00 mL, 0.05 mg/mL), β-glycerol phosphate (2.00 mL, 10 mM) (all from Sigma- Aldrich, USA), vitamin D2 (2.00 mL, 2.10 −8 M, Mekopharm-Vietnam), and FGF9 (0.20 mL, 10 ng/mL, Sigma-Aldrich, USA). After 3 weeks in osteogenic medium, cells tested positive for cytochemical stains AP, AR, and VK and for osteocalcin marker by RT-PCR; part of the osteo-blasts were harvested and cultured on coral scaf-fold Porites lutea and scanned by SEM and resulted positive for AP.
6.2.3.1 hATMSCs Undergo Osteoblastic Differentiation: Cytochemical Stains AR, AP, and VK (Fig. 6.6 )
This part of the project was to confi rm the hATM-SCs’ capacity for differentiation toward osteo-blast phenotype. The cells were harvested at the second passage by enzymatic digestion and cul-tured with a proper osteogenic medium [ 10 ]. New cell lines were collected after 3 weeks ca. and stained by classic cytochemical stains AR, AP, and VK to prove calcium matrix and hydroxyapa-tite nodules and fi lament deposits. Cells revealed a strong and consistent osteogenic activity.
6.2.4 In Vitro Transdifferentiation of Human Adipocyte Cells to Osteoblasts
Adipocytes from hATMSCs were directly obtained by adding into cell fl ask cultures at the second pas-sage, a specifi c osteogenic medium composed of
IMDM (63.05 mL, 13.05 %), penicillin- streptomycin (8.70 mL), FBS (13.05 mL), dexamethasone (10.0 mL, 10 −7 M), ascorbic acid (1.00 mL, 0.05 mg/mL), β-glycerol phosphate (2.00 mL, 10 mM) (all from Sigma- Aldrich, USA), vitamin D2 (2.00 mL, 2.10 −8 M), and FGF9 (0.20 mL, 10 ng/mL). After 3 weeks in osteogenic medium, cells tested positive for cytochemical stains AP, AR, and VK and for osteocalcin marker by RT-PCR; part of the of osteo-blasts were harvested and cultured on coral scaffold Porites lutea and scanned by SEM and resulted posi-tive for cytochemical stain Giemsa and AP.
6.2.4.1 Preparation of Fibrin Gel Fibrin gel was obtained by combining two com-ponents, fi brinogen and thrombin, extracted from peripheral blood of healthy and consented donors. Blood was tested negative for HIV, HBV, HCV, and VDRL.
6.2.4.2 Collection of Fibrinogen The blood was centrifuged at 3,000 rpm for 5 min. About 10 ml of serum was centrifuged at 3,000 rpm for 5 min. The solution was fi ltered using a fi lter with a diameter of 0.20 μm (Minisart Sartorius ® ) and placed into a new sterile tube and incubated at 4 °C for one hour and at −20 °C [ 19 ].
6.2.4.3 Collection of Thrombin Blood samples were centrifuged at 3,000 rpm for 5 min, and 10 mL of serum was collected. Serum was centrifuged at 3,000 rpm for 5 min. The solution was collected and fi ltered by a diameter 0.20 μm fi lter (Minisart Sartorius ® ) into a new sterile tube and incubated at 4 °C for one hour and overnight at −20 °C. Then the tube was
Fig. 6.6 To confi rm ATMSCs’ multipotent capacity, they were induced to differentiate into osteocells. After 21 days of culture, the cells were stained by classic cytochemical stains. ( Left ) AR, ( Middle ) AP, ( Right ) VK
T.C. Tran et al.

93
thawed at a temperature of 4 °C; PBS solution was added (ratio 1:9). Acetic acid was added 1 % to adjust the pH at 5.3. The solution was allowed to stand for 30 min to precipitate. Afterward it was centrifuged at 3,000 rpm for 5 min. The sedi-ment on the bottom was removed and collected. PBS was added (ratio 1:9). Na 2 CO 3 0.1 M was used to adjust to pH 7.0. The whole was placed in a thermostatic bath at 37 °C for 15 min, and CaCl 2 (0.01 M) was added to create clots. Clots were removed and discharged, while the remain-ing liquid that is thrombin was collected and pre-served at a temperature of −20 °C [ 40 ].
6.2.4.4 Seeding hFTMSCs onto Fibrin Gel
Fibrin gel is formed by the combination of fi brin-ogen and thrombin (ratio of 1:1). hFTMSCs, at second passage, were collected by enzymatic digestion, a solution of Trypsin-EDTA (0.25–0.02 %) (Fig. 6.7 ). Cells were centrifuged at 3,000 rpm for 5 min, and a fi brinogen solution was added to cells deposited at the bottom. The cell-fi brinogen solution was moved into 30 cm diameter Petri dishes, and additional thrombin was poured (ratio 1:1); the whole was manually mixed and rested for few minutes for it
to become a homogeneous compact formation. After one day, the cell culture medium was replaced with pre-adipocyte differentiation cul-ture medium. After 3 days, the culture medium was replaced by adipocyte differentiation culture medium. After 3 weeks of culture, the cells are tested by inverted microscope, Oil Red staining, and H&E staining for fi brin gel (Fig. 6.8 ).
When mixed, fi brinogen and thrombin (ratio of 1:1) are able to form a block of gel within 5 min; the fi nal result is a fi brin gel that is bright yellow in color and very tough and resilient in consistency. After 21 days culture, hFTMSCs, cultured on fi brin gel using an adipogenic culture medium, are completely differentiated into adult adipocytes.
Oil Red staining directly performed on cells cultured on fi brin gel showed that hFTMSCs are completely differentiated into adipocytes with the typical round shape and the very characteris-tic cytoplasm containing lipid particles captured by bright red with Oil Red dye (Fig. 6.9 ).
Fibrin gels containing adipocytes were fi xed in 10 % Neutral Formalin Buffer Saline (NBF) solution and stained with H&E. The results show that cells survive and grow well on fi brin gel; within the cell cytoplasm are visible lipid droplets (Fig. 6.10 ).
aa b
b
Fig. 6.7 The resulting fi brin gel. ( a ) Two components were isolated separately ( a ) fi brinogen and ( b ) thrombin. ( b ) The combination of fi brinogen and thrombin with a
1:1 volume within 1–5 min quickly forms a light yellow block of gel that seeded with adipocytes giving an elastic and supple tissue very close to human fat tissue
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells
94
a b c
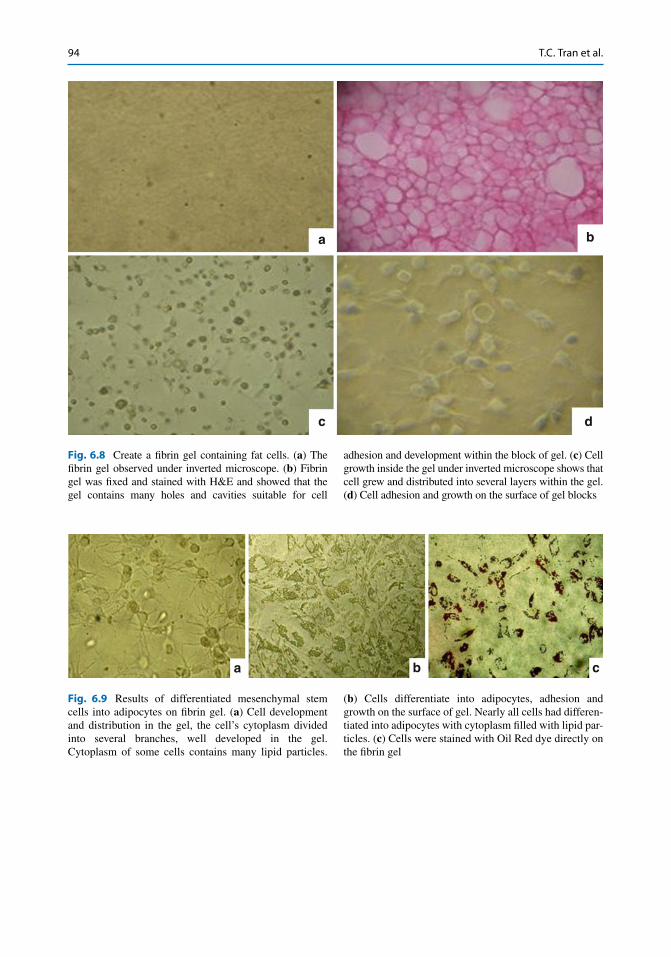
Fig. 6.9 Results of differentiated mesenchymal stem cells into adipocytes on fi brin gel. ( a ) Cell development and distribution in the gel, the cell’s cytoplasm divided into several branches, well developed in the gel. Cytoplasm of some cells contains many lipid particles.
( b ) Cells differentiate into adipocytes, adhesion and growth on the surface of gel. Nearly all cells had differen-tiated into adipocytes with cytoplasm fi lled with lipid par-ticles. ( c ) Cells were stained with Oil Red dye directly on the fi brin gel
a b
c d
Fig. 6.8 Create a fi brin gel containing fat cells. ( a ) The fi brin gel observed under inverted microscope. ( b ) Fibrin gel was fi xed and stained with H&E and showed that the gel contains many holes and cavities suitable for cell
adhesion and development within the block of gel. ( c ) Cell growth inside the gel under inverted microscope shows that cell grew and distributed into several layers within the gel. ( d ) Cell adhesion and growth on the surface of gel blocks
T.C. Tran et al.

95
6.2.4.5 Preparation and Structural Characteristics of Coral Scaffold Porites lutea (Fig. 6.11 and 6.12 )
Natural porous coral, Porites lutea (from the Oceanographic Institute of Nha Trang city VN), was washed in 0.9 % distilled water, lyophilized, and sterilized by gamma ray to 25KGy. Porites lutea shows to have ideal porosity and enough volume fraction with high surface area to carry a high number of cells within and the surround-ing host tissues allowing its ingrowth and vas-cularization. Data are presented as mean ± SE (Tables 6.1 , 6.2 , and 6.3 )
6.3 Seeding hATMSCs and Adipocytes onto Coral Scaffold
MSCs from hAT and adipocytes have been col-lected at second passage and seeded on coral scaf-fold and immersed in osteogenic medium; cells
were counted by Trypan Blue. Cells have been cen-trifuged together with the scaffold 5 times at 1,000 g per minute and incubated at 37 °C with 5 % CO 2 . Media have been changed every 3–4 days. Scaffold has been incubated for 7, 15, and 21 days and stained by Giemsa, H&E, AP, and fl uorescence analysis microscope and pictured by SEM.
The specimens are collected and fi xed by a fi x-ative buffer solution composed of glutaraldehyde- formaldehyde (0.7 μM/5 mL medium c.ca) and incubated at 37 °C with 0.5 % CO 2 . The fi xed cell tissue is dehydrated with organic solvents acetone or ethanol. Solvents are removed by car-bon dioxide at high pressure; once CO 2 is also removed, dried specimen is collected and placed on stub using an adhesive such as epoxy resin and sputter coated with gold alloy before examina-tion in the microscope.
Osteoblasts from both sources were seeded onto sea coral scaffold. At day 10 c.ca, osteocells started to produce a compact matrix that does not require enzymatic digestion; therefore, the tissue
a b
Fig. 6.10 Result of H&E staining for fi brin gel contain-ing adipocytes. ( a ) Gel containing adipocytes photo-graphed at 100 times magnifi cation with inverted microscope. ( b ) Gel containing adipocytes photographed
at 200 times magnifi cation with inverted microscope. The cells are arrested in purple , and around the nucleus of the cell are gaps that do not color. These were droplets of lipid within the cytoplasm of the cell
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells
96
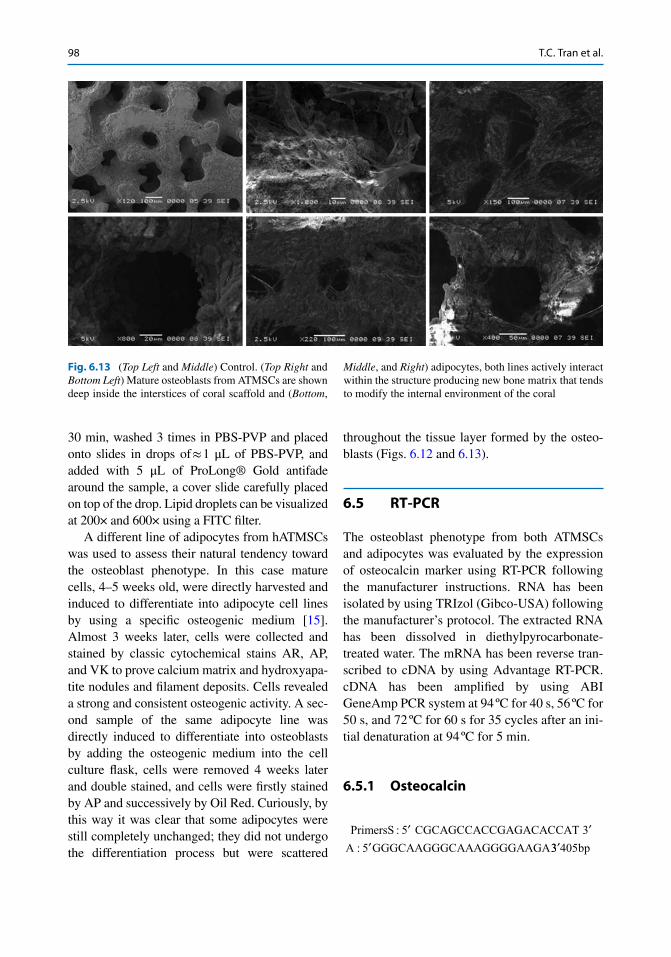
was manually removed, washed by PBS, and wrapped on corals and incubated for 14 days. SEM pictures clearly show that osteoblasts were able to home deep inside the coral interstices, growing and expanding, and able to interact with
their environment. The pictures show an elevated dense population and how osteoblasts have radi-cally changed the morphology of the scaffold where they homed (Fig. 6.11 ).
200
150
100
50
0% M0/V % M1/V
P. Lutea% M2/V
P. Australi% M3/V
M. Annuali% M4/V
Fig. 6.11 Water loading velocity: % when dried coral blocks were exposed in sterile 0.9 % sodium chloride at boiled temperature with low pressure
140
120
100
80
60
40
20
0% M0/V % M1/V % M2/V % M3/V % M4/V
P. Australi
P. Lutea
M. Annuali
Fig. 6.12 Water loading velocity: % when dried coral blocks were exposed in sterile 0.9 % sodium chloride at room temperature with pressure, chloride at room temperature with high pressure
Table 6.1 Density: Coral weight was measured by an electric scale (Sartorius-OSI-BP 124.78312). Their vol-ume was made by a technological rule
Density (cm 3 /kg) of dried coral rods (mean ± 2 SE)
Porites australiensis 1.3297 ± 0.0439 (60 dried coral blocks)
Porites lutea 1.0771 ± 0.0364 (60 dried coral blocks)
Montastrea annuligera 1.3752 ± 0.0462 (60 dried coral blocks)
Table 6.2 Porosity: The porous rates of coral were stud-ied indirectly by weighing both dried blocks and water saturated ones in Thermosi SR 3000, OSI, at 30–40 °C, 40–70 atm
Porosity (%) of coral blocks (mean ± 2 SE)
Porites australiensis 0.4291 ± 0.0167 (60 dried coral blocks)
Porites lutea 0.5538 ± 0.0218 (60 dried coral blocks)
Montastrea annuligera 0.3665 ± 0.0165 (60 dried coral blocks)
T.C. Tran et al.

97
6.4 Cytochemical Staining
Mineral matrix deposits and bone nodules of both groups of osteoblasts, from hATMSCs and adipo-cytes, were evaluated by staining cell cultures by AR, AP, and VK. Adipocyte phenotypes were confi rmed by using Nile Red and Red Oil stains.
6.4.1 Alizarin Red Stain Procedure
The presence of calcium deposits was detected by washing cells with cold PBS and fi xing them in 10 % NFB solution for 30 min in chemical hood. Cells were rinsed 3 times with distilled water and immerged in 2 % (w/v) solution of Alizarin red for 30 s to 5 min. Cells were rinsed again with distilled water 2–3 times and checked under inverse microscope and pictured.
6.4.2 Alkaline Phosphatase Stain Procedure
The presence of alkaline phosphates was detected by washing cells with cold PBS and fi xing them in 10 % NFB solution for 30 min in chemical hood and stained with solution naphthol As-MX-PO 4 (Sigma) and Fast Red Violet LB Salt (Sigma) for 45 min in dark at room temperature. Cells were rinsed three times with distilled water and checked by inverse microscope and pictured.
6.4.3 Von Kossa Stain Procedure
The presence of calcium deposits was detected by washing cells with cold PBS and fi xing them in 10 % NFB solution for 30 min in chemical hood and stained with 2.5 % silver nitrate (Merck- Germany) for 30 min in a dark place. Cells were rinsed 3 times with distilled water and checked by inverse microscope and pictured.
6.4.4 Oil Red for Adipocytes from hATMSC
At 90 % confl uence, second-generation hATM-SCs were harvested by enzymatic digestion and induced to differentiate into adipocytes, by using a specifi c adipogenic medium previously described. Cells stay in culture for almost a month; they started to change shape at the second week to assume the complete mature adipocyte conformation at the end of 4th week showing the typical fat droplets inside their cytoplasm.
6.4.5 Nile Red
At 90 % confl uence, second-generation hATMSCs were harvested by enzymatic digestion and induced to differentiate into adipocytes, by using a specifi c adipogenic medium previously described. Cells stay in culture for almost 30 days; they started to change shape at the second week to assume the complete mature adipocyte conforma-tion at the end of 4th week showing the typical fat droplets inside their cytoplasm. At mature confl u-ence, cells were fi xed in 1 % paraformaldehyde in PBS and stained by Nile Red following the described procedure; samples were removed from culture and washed 3 times in 100 μL PBS-PVP (2 min for each wash) and transferred to 100 μL of fi xative, then rinsed two times in fi xative and incu-bated for at least 15 min. The sample is transferred to a 100 μL of Nile Red working solution for
Table 6.3 Mechanical strengh:strengh was measured at the Nuclear Technology Center of HCMC by Strograph V10 - C Toyoseiki
Mechanical strength (Kg/cm 3 ) of dried coral blocks (mean ± 2SE)
Porites australiensis 68.6638 ± 10.5895 (60 dried coral blocks)
Porites lutea 53.6082 ± 10.3835 (60 dried coral blocks)
Montastrea annuligera 51.7504 ± 11.5625 (60 dried coral blocks)
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells
98
30 min, washed 3 times in PBS-PVP and placed onto slides in drops of ≈ 1 μL of PBS-PVP, and added with 5 μL of ProLong® Gold antifade around the sample, a cover slide carefully placed on top of the drop. Lipid droplets can be visualized at 200× and 600× using a FITC fi lter.
A different line of adipocytes from hATMSCs was used to assess their natural tendency toward the osteoblast phenotype. In this case mature cells, 4–5 weeks old, were directly harvested and induced to differentiate into adipocyte cell lines by using a specifi c osteogenic medium [ 15 ]. Almost 3 weeks later, cells were collected and stained by classic cytochemical stains AR, AP, and VK to prove calcium matrix and hydroxyapa-tite nodules and fi lament deposits. Cells revealed a strong and consistent osteogenic activity. A sec-ond sample of the same adipocyte line was directly induced to differentiate into osteoblasts by adding the osteogenic medium into the cell culture fl ask, cells were removed 4 weeks later and double stained, and cells were fi rstly stained by AP and successively by Oil Red. Curiously, by this way it was clear that some adipocytes were still completely unchanged; they did not undergo the differentiation process but were scattered
throughout the tissue layer formed by the osteo-blasts (Figs. 6.12 and 6.13 ).
6.5 RT-PCR
The osteoblast phenotype from both ATMSCs and adipocytes was evaluated by the expression of osteocalcin marker using RT-PCR following the manufacturer instructions. RNA has been isolated by using TRIzol (Gibco-USA) following the manufacturer’s protocol. The extracted RNA has been dissolved in diethylpyrocarbonate- treated water. The mRNA has been reverse tran-scribed to cDNA by using Advantage RT-PCR. cDNA has been amplifi ed by using ABI GeneAmp PCR system at 94 ºC for 40 s, 56 ºC for 50 s, and 72 ºC for 60 s for 35 cycles after an ini-tial denaturation at 94 ºC for 5 min.
6.5.1 Osteocalcin
PrimersS CGCAGCCACCGAGACACCAT
A GGGCAAGGGCAAAGGGGAAGA
:
:
5 3
5
′ ′′ 33 405′ bp
Fig. 6.13 ( Top Left and Middle ) Control. ( Top Right and Bottom Left ) Mature osteoblasts from ATMSCs are shown deep inside the interstices of coral scaffold and ( Bottom ,
Middle , and Right ) adipocytes, both lines actively interact within the structure producing new bone matrix that tends to modify the internal environment of the coral
T.C. Tran et al.

99
Osteoblasts from both lines were collected at day 21 and were tested for osteocalcin marker by RT-PCR. The osteogenic differentiation is induced by using a specifi c medium, as previ-ously described, that is, the same used in our edited studies to differentiate MSCs from hBM and hUCB into osteoblasts [ 10 ]. MSCs have been used as negative control and human bone as posi-tive control (Fig. 6.14 ).
6.6 Cell Surface Characterization Flow Cytometry
Cell surface phenotype of human ATMSCs is very similar to their counterpart from either hUCB or hBM. In this project, ATMSCs were tested for specifi c CD markers to confi rm their identity; cells resulted negative for classic hema-topoietic cell markers such as CD14, CD45, and HLA-DR and were positive for exclusive MSCs markers such as CD13, CD44, CD73, CD90, CD105, and CD166. Cells at passage 2 were har-vested by enzymatic digestion with Trypsin- EDTA and were stained in PBS containing FBS and incubated with both isotype control and anti-human monoclonal antibodies together with fl uo-rescein isothiocyanate (FITC) and phycoerythrin (PE) protein for 30 min at 4 °C. Cells were washed and analyzed by a FACSCalibur cytom-eter (Becton Dickinson). To detect the specifi c cell surface markers, cells were fi xed in 2 % para-formaldehyde for 5 min at room temperature and then permeabilized with 0.1 % Triton X-100
(Sigma) for 30 min at room temperature. Once this process was fi nished, cells were washed and blocked with 1 % BSA in PBS before incubation for 4 h at 4 °C with primary antibodies. After washing, the cells were incubated with fl uores-cein isothiocyanate (FITC)-conjugated second-ary antibodies for 30 min at room temperature. Next, cells were washed and analyzed as earlier presented.
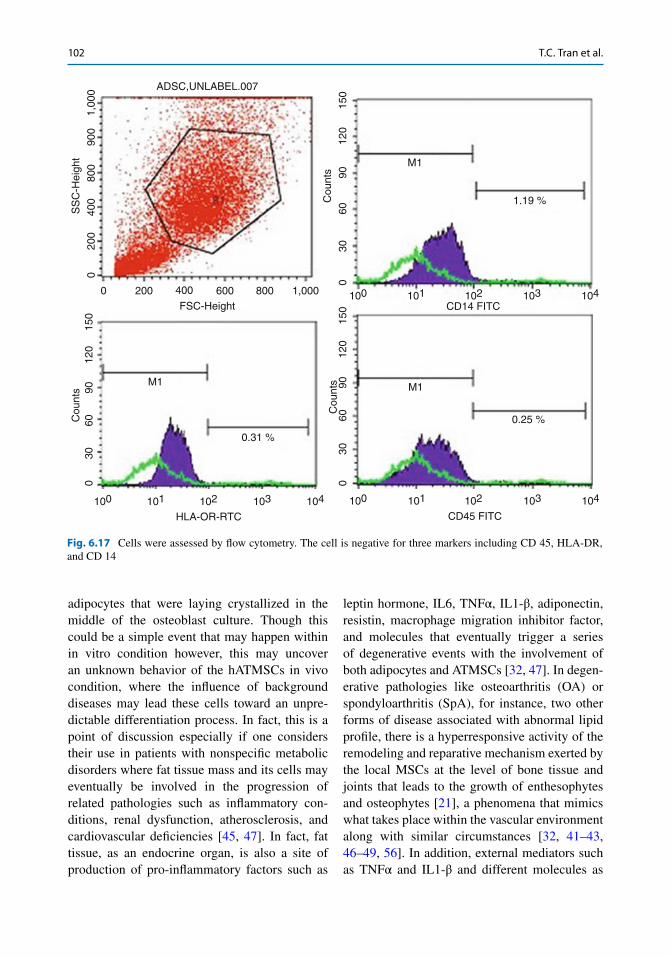
Results showed that cell lines are negative for the markers CD14, CD45, and HLA-DR; and positive for the markers CD13, CD44, CD73, CD90, CD105, and CD166. Thus, we have iso-lated and successfully cultured mesenchymal stem cells isolated from adipose tissue.
6.7 Discussion
Recently, scientists have started to consider that adipose tissue is not just a simple passive organ of energy storage but a real endocrine system and an active player in hormone regulation and homeostatic balance [ 31 , 32 ]. During the 1990s, it has been discovered that adipose tissue was the source of leptin hormone, and this was the foun-dation of a new period of investigation on adipo-cyte endocrinology [ 32 ]. In addition, it is now clear that adipose tissue has great potential in terms of providing stem cells and thus therapeu-tic opportunities [ 13 , 18 , 27 , 30 , 36 , 51 , 52 ]. These stem cells have been shown to belong to the large family of MSCs, sharing common traits and great potential that have made MSCs one of
Fig. 6.14 Mature adipocytes were induced to differenti-ate into osteoblast. At day 21 they were stained by cyto-chemical stains. ( Left ) AR, ( Middle ) VK, and ( Right ) AP.
The adipose cells started to change shape almost 10 days and became fully mature at 21 days
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

100
the biggest expectations in all the medical world, yet with unique distinctive features which is of importance in clinical use as well [ 13 , 18 , 19 , 27 , 30 , 36 , 51 , 52 ]. In addition, fat tissue is simple to obtain with a minimal invasive procedure and with minimal complications for the patient [ 13 , 18 , 24 , 27 , 30 , 36 , 51 , 52 ]. Every year the need for reconstructive surgical operations is increas-ing, along with the demand for repairing dam-aged tissues because of diseases, injuries, or congenital defects [ 27 , 28 , 33 , 34 , 51 ]. Nowadays, reconstructive strategies to repair tissue defects such as breast, cartilage, and bone are based on the use of implants and fi ller [ 27 , 51 , 53 , 54 ]. However, there is no single fi ller material or implant which may be sui for all different needs [ 51 ]. This is the main reason that has pushed medical scientists toward enormous efforts to fi nd new and alternative solutions to be used in reconstructive and regenerative medicine, tools that would combine the high feasibility of bioma-terials and the enormous ductility of stem cells [ 10 , 30 , 51 , 53 – 55 ]. As a result, we are witness-ing a large quantity of data coming from either in vitro or in vivo investigations of hATMSCs alone or in combination with biomaterials [ 13 , 18 , 24 , 28 , 30 , 33 – 36 , 51 , 52 ]. The clinical applications have seen the use of ATMSCs either in liver, heart, bone, and cartilage defects or as vectors in anticancer therapy, and they have been gaining
more and more consensus in the fi eld of cosmetic surgery [ 13 , 18 , 24 , 27 , 28 , 30 , 33 – 36 , 51 , 52 ].
This study has eventually confi rmed a few fi rm points regarding the nature and the in vitro behavior of hATMSCs. These cells show a great plasticity and transdifferentiation capacity, a strong adherence property, a high proliferative ability, and a natural tendency to home and grow in a biomaterial such as sea coral scaffold. The isolated primary cells started to adhere 2 days after being placed into a fl ask culture with a proper medium, after a week c.ca they started to assume the typical fi broblast spindle-like shape, and within 10 days/2 weeks, they reached a 90 % confl uence. Flow cytometry analysis confi rmed the cellular identity of these cells and their affi li-ation to the wider MSCs family group. At the second passage, cells were collected by an enzy-matic digestion process and were induced to dif-ferentiate to two different cell lines: adipocytes and osteoblasts.
The results were, respectively, assessed by Oil Red and Nile Red (fl uorescence microscopy) for the adipocyte lineage and by cytochemical stains AR, VK, and AP and RT-PCR (osteocalcin) for the osteocell lineage (Figs. 6.15 , 6.16 , 6.17 , and 6.18 ). Successively, adipocytes were collected and induced to transdifferentiate into osteoblasts and within 2 weeks c.ca of culture in proper medium solution, the majority of cells completely changed
Fig. 6.15 Part of osteoblasts from adipocytes were dou-ble stained with ( Left ) an AP fi rst and ( Right ) Oil Red later. It was noticed that on the surface of bone tissue formed by osteocells, there were some adipocytes that did
not undergo differentiation. This confi rmed that both ATMSCs and adipocytes may have a key role in ectopic formation of calcifi ed tissue (×400)
T.C. Tran et al.
101
their round shape with their typical intracellular fat droplets into cuboidal-like shape of osteoblast cells with the well-known presence of fi laments and hydroxyapatite nodules. Results were assessed by cytochemical stains AR, VK, and AP and by RT-PCR (osteocalcin). The next step was to obtain osteoblast cultures from either MSCs or adipo-cytes directly onto a coral scaffold, and results, by SEM analysis, showed an equal behavior of both group; osteoblasts were able to home into the deep interstices of the coral acting on the environment modifying it in the same manner and almost at the same pace. This indicates that a possible use of these type of cells together with a hard structure bio-scaffold as potential tool in the reconstruction of bone tissue defection. Moreover, little has been seen regarding the use of hFTMSCs together with a biomaterial, such as fi brin gel completely gener-ated by human donors. In line with these current results and other published studies, human fi brin
gel demonstrates an ideal environment where these cells may eventually switch into adipocytes generating a product with the consistency and characteristic certainly very similar to human fat tissue. Moreover, this fi nal biomaterial is abso-lutely safe and compatible since it was obtained from the patient’s own blood and fat tissue. To conclude, we are sure that it may eventually be of a great help in clinical fi eld, whether we think of reconstructive and regenerative procedures or cos-metic applications.
However, back to osteoblasts from adipo-cytes, a particular trait kept our attention. After AP stain has been attempted, a few black spots were visible under the microscope, and we theorized that few adipocytes probably did not undergo osteoblast differentiation. Therefore, it has been performed on the same culture a fur-ther Oil Red analysis, and the result provided evidence of the presence of a few unchanged
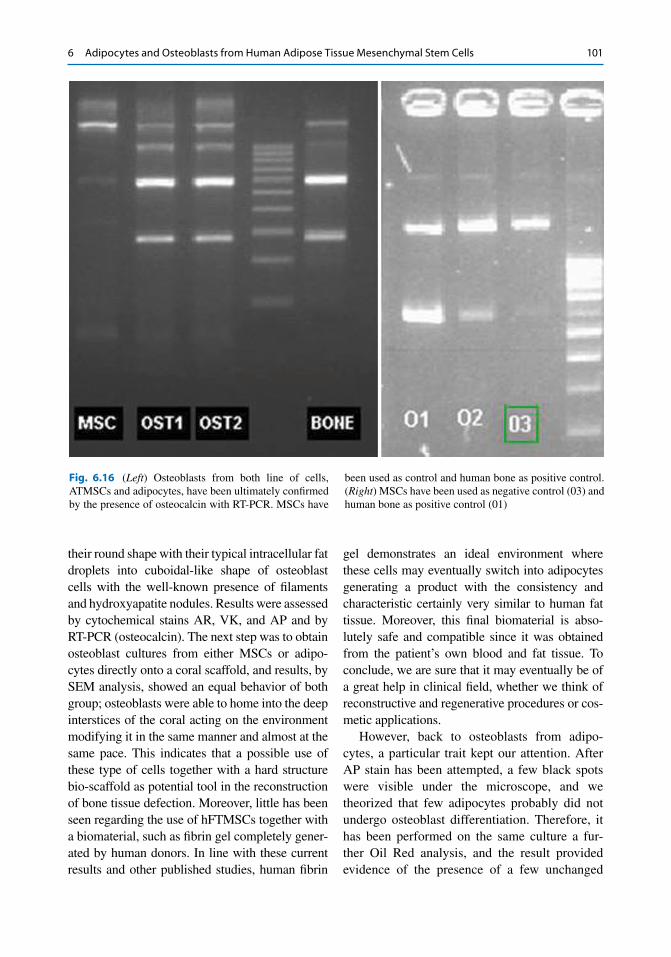
Fig. 6.16 ( Left ) Osteoblasts from both line of cells, ATMSCs and adipocytes, have been ultimately confi rmed by the presence of osteocalcin with RT-PCR. MSCs have
been used as control and human bone as positive control. ( Right ) MSCs have been used as negative control (03) and human bone as positive control (01)
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells
102
adipocytes that were laying crystallized in the middle of the osteoblast culture. Though this could be a simple event that may happen within in vitro condition however, this may uncover an unknown behavior of the hATMSCs in vivo condition, where the infl uence of background diseases may lead these cells toward an unpre-dictable differentiation process. In fact, this is a point of discussion especially if one considers their use in patients with nonspecifi c metabolic disorders where fat tissue mass and its cells may eventually be involved in the progression of related pathologies such as infl ammatory con-ditions, renal dysfunction, atherosclerosis, and cardiovascular defi ciencies [ 45 , 47 ]. In fact, fat tissue, as an endocrine organ, is also a site of production of pro-infl ammatory factors such as
leptin hormone, IL6, TNFα, IL1-β, adiponectin, resistin, macrophage migration inhibitor factor, and molecules that eventually trigger a series of degenerative events with the involvement of both adipocytes and ATMSCs [ 32 , 47 ]. In degen-erative pathologies like osteoarthritis (OA) or spondyloarthritis (SpA), for instance, two other forms of disease associated with abnormal lipid profi le, there is a hyperresponsive activity of the remodeling and reparative mechanism exerted by the local MSCs at the level of bone tissue and joints that leads to the growth of enthesophytes and osteophytes [ 21 ], a phenomena that mimics what takes place within the vascular environment along with similar circumstances [ 32 , 41 – 43 , 46 – 49 , 56 ]. In addition, external mediators such as TNFα and IL1-β and different molecules as
ADSC,UNLABEL.0071,
000
900
800
400
200
0
0 200 400 600 800 1,000 100 101 102CD14 FITCFSC-Height
M1
0.25 %
M1
1.19 %
M1
0.31 %
CD45 FITC
103 104
100 101 102 103 104
HLA-OR-RTC100 101 102 103 104
150
120
90C
ount
s
Cou
nts
SS
C-H
eigh
t
Cou
nts
6030
0
150
120
9060
300
150
120
9060
300
Fig. 6.17 Cells were assessed by fl ow cytometry. The cell is negative for three markers including CD 45, HLA-DR, and CD 14
T.C. Tran et al.

103
dexamethasone, vitamin D, C-reactive protein (CRP), and homocysteine may also have a direct role in endothelial cell activation and infl amma-tion of vascular tissue, incidents that promote the
progression and the evolvement of atherosclerotic tissues [ 32 , 41 – 43 , 45 – 49 , 56 , 57 ]. Therefore, atherosclerotic mechanism is not to be considered just as a simple inert concentration of calcium
M1
99.66 %
150
120
9060
300
100 101 102 103 104 100 101 102
CD44PECD13PE
M1
94.32 %
M1
99.46 %
M1
99.64 %
CD166PE
103 104
100 101 102 103 104CD90PE
100 101 102 103 104
150
120
90
Cou
nts
Cou
nts
Cou
nts
Cou
nts
6030
0
150
120
9060
300
150
120
9060
300
M1
97.33 %M1
99.31 %
CD73PE
100 101 102 103 104
CD105PE
100 101 102 103 104
150
120
90
Cou
nts
Cou
nts
6030
0
150
120
9060
300
Fig. 6.18 Cells were assessed by fl ow cytometry. The cell is positive for six markers including CD 13, CD 44, CD 90, CD 166, CD 73, and CD 105
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

104
Fig. 6.20 X-rays 28 months after treatment
Fig. 6.19 X-ray before treatment. Impression is bilateral osteophytes consistent with degenerative joint disease
matrix deposits dissolved in uremic serum and in the blood stream that sediment by the time, “but a precisely regulated—or perhaps precisely dereg-ulated—controlled process” which originates right within the fat mass environment [ 41 , 48 ]. And the event seems to be related to a precise cellular contributions that, under the infl uence of particular molecules and cytokines, eventu-ally induce the production of ectopic bone tissue formation, the same differentiation mechanism that takes place within a normal bone system [ 29 , 41 , 46 , 48 , 57 ]. Now, this occurrence may either indicate a direct involvement of vascular stem cells (VSC), which some author indicates as ATMSCs, or the involvement of local adipocytes generally located within the fat mass tissues [ 21 , 41 , 44 , 48 , 50 , 57 ]. For example, local ATMSCs can be recruited at the site of damaged tissue or infl amed areas by several proteins and com-pounds; the ability to cross the endothelial barrier is controlled by the expression of mediators such as matrix metalloproteinases and their inhibitors MMP-2, TIMP-2, and MT1 while once at the site, under particular stimulation of other differ-ent factors such as bone morphogenetic protein
2-a (BMP2-a), they may either over respond or start to differentiate into calcium matrix producer osteoblasts [ 21 , 41 , 44 , 45 , 48 ].
6.8 Case Report (Figs. 6.19 , 6.20 , 6.21 , and 6.22 )
This 55-year-old female had knee osteoarthri-tis for 3 years, and she was treated by mesen-chymal stem cell autograft from abdominal fat tissue. The knee injection was around 1–5 × 10 6 MSCs.
The patient was assessed by grading the Tegner Lysholm Knee Scoring Scale.
< − − >65 65 83 84 90 90: ; : ; : ; :Poor Fair Good Excellent
Lysholm Knee Scoring Scale before treatment: I. Limp
I have no limp when I walk. (5) I have a slight or periodical limp when I walk. (3) I have a severe and constant limp when I walk. (0)
T.C. Tran et al.

105
II. Using cane or crutches I do not use a cane or crutches. (5) I use a cane or crutches with some weight bearing. (2) Putting weight on my hurt leg is impossi-ble. (0)
III. Locking sensation in the knee I have no locking and no catching sensa-tions in my knee. (15) I have catching sensation but no locking sensation in my knee. (10) My knee locks occasionally. (6) My knee locks frequently. (2) My knee feels locked at this moment. (0)
IV. Giving way sensation from the knee My knee never gives way. (25) My knee rarely gives way, only dur-ing athletics or other vigorous activities. (20) My knee frequently gives way during ath-letics or other vigorous activities; in turn I am unable to participate in these activi-ties. (15) My knee occasionally gives way during daily activities. (10) My knee often gives way during daily activities. (5) My knee gives way every step I take. (0)
Fig. 6.21 MRI 28 months after treatment
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

106
V. Pain I have no pain in my knee. (25) I have intermittent or slight pain in my knee during vigorous activities. (20) I have marked pain in my knee during vig-orous activities. (15) I have marked pain in my knee during or after walking more than 1 mile. (10) I have marked pain in my knee during or after walking less than 1 mile. (5) I have constant pain in my knee. (0)
VI. Swelling I have no swelling in my knee. (10) I have swelling in my knee only after vig-orous activities. (6) I have swelling in my knee after ordinary activities. (2) I have swelling constantly in my knee. (0)
VII. Climbing stairs I have no problems climbing stairs. (10) I have slight problems climbing stairs. (6) I can climb stairs only one at a time. (2) Climbing stairs is impossible for me. (0)
VIII. Squatting I have no problems squatting. (5) I have slight problems squatting. (4) I cannot squat beyond a 90° bend in my knee. (2) Squatting is impossible because of my knee. (0) Patient total 30/100
Lysholm Knee Scoring Scale follow-up sur-vey at 28 months: I. Limp
I have no limp when I walk. (5) I have a slight or periodical limp when I walk. (3) I have a severe and constant limp when I walk. (0)
II. Using cane or crutches I do not use a cane or crutches. (5) I use a cane or crutches with some weight bearing. (2) Putting weight on my hurt leg is impossi-ble. (0)
III. Locking sensation in the knee I have no locking and no catching sensa-tions in my knee. (15) I have catching sensation but no locking sensation in my knee. (10) My knee locks occasionally. (6) My knee locks frequently. (2) My knee feels locked at this moment. (0)
IV. Giving way sensation from the knee My knee never gives way. (25) My knee rarely gives way, only during ath-letics or other vigorous activities. (20) My knee frequently gives way during athlet-ics or other vigorous activities; in turn I am unable to participate in these activities. (15) My knee occasionally gives way during daily activities. (10)
Fig. 6.22 Function 28 months after treatment
T.C. Tran et al.

107
My knee often gives way during daily activities. (5) My knee gives way every step I take. (0)
V. Pain I have no pain in my knee. (25) I have intermittent or slight pain in my knee during vigorous activities. (20) I have marked pain in my knee during vig-orous activities. (15) I have marked pain in my knee during or after walking more than 1 mile. (10) I have marked pain in my knee during or after walking less than 1 mile. (5) I have constant pain in my knee. (0)
VI. Swelling I have no swelling in my knee. (10) I have swelling in my knee only after vig-orous activities. (6) I have swelling in my knee after ordinary activities. (2) I have swelling constantly in my knee. (0)
VII. Climbing stairs I have no problems climbing stairs. (10) I have slight problems climbing stairs. (6) I can climb stairs only one at a time. (2) Climbing stairs is impossible for me. (0)
VIII. Squatting I have no problems squatting. (5) I have slight problems squatting. (4) I cannot squat beyond a 90° bend in my knee. (2) Squatting is impossible because of my knee. (0) Patient total 89/100
Conclusions
The authors are sure that ATMSCs may eventu-ally be of signifi cant applications in the clinical fi eld, whether we think of reconstructive and regenerative procedures or cosmetic applica-tion. However, we are aware that further analy-sis and deeper investigation should be performed on this area. Despite an initial favor-able position, some authors tend to be cautious of an unsystematic use of these cells, stressing their underlying paracrine effect which involves the secretion of several cytokines, growth fac-tors, and immunosuppressive capacity [ 28 ].
References
1. Chao NJ, Emerson SG, Weinberg KI. Stem cell trans-plant (cord blood transplants). Hematology Am Soc Hematol Educ Program. 2004;1:354–71.
2. Goodwin HS, Bicknese AR, Chien SN, Bogucki BD, Olivier DA, Qinn CO, Wall DA. Multilineage dif-ferentiation activity by cells isolated from umbili-cal cord blood: expression of bone, fat, and neural markers. Biol Blood Marrow Transplant. 2001;7(11):581–8.
3. Koc O, Lazarus HM. Mesenchymal stem cells: head-ing into the clinic. Bone Marrow Transplant. 2001;27(3):235–9.
4. Lee OK, Kuo TK, Chen WM, Lee KD, Hsien SL, Chen TH. Isolation of multipotent mesenchymal stem cells from umbilical cord blood. Blood. 2004;103(5):1669–75.
5. Liu TM, Martina M, Hutmacher DW, Po Hui JH, Lee EH, Lim B. Identifi cation of common pathways mediating differentiation of bone marrow and adipose tissue derived human mesenchymal stem cells into three mesenchymal lineages. Stem Cells. 2007;25(3):750–60.
6. Minguell JJ, Erices A, Conget P. Mesenchymal stem cells. Exp Biol Med (Maywood). 2001;226(6):507–20.
7. Parolini O, Alviano F, Bagnara GP, Bilic B, Uhring HJ, Evangelista M, Hennerbichler S, Liu B, Magatti M, Mao N, Miki T, Marongiu F, Nakajima H, Nikaido T, Portmann-Lanz ICB, Sankar V, Soncini M, Stadler G, Surbek JD, Takahashi TA, Redl H, Sakuragawa N, Wolbank S, Zeisberger S, Zisch A, Strom SC. Concise review: isolation and characterization of cells from human term placenta: outcome of the fi rst interna-tional Workshop on Placenta Derived Stem Cells. Stem Cells. 2008;26(2):300–11.
8. Rosada C, Justensen J, Melsvik D, Ebbesen PM, Kassem M. The human umbilical cord blood: a poten-tial source for osteoblast progenitor cells. Calcif Tissue Int. 2003;72(2):135–42.
9. Stocum DL. Regenerative biology and medicine. Philadelphia: Elsevier; 2006. p. 229–37.
10. Tran CT, Gargiulo C, Thao HD, Tuan HM, Filgueira L, Strong M. In vitro culture and differentiation of osteoblasts on coral scaffold from human bone mar-row mesenchymal stem cells. Cell Tissue Bank. 2010;12(4):247–61.
11. Tsai MS, Hwang SM, Chen KD, Lee YS, Hsu LW, Chang YJ, Wang CN, Peng HH, Chang YL, Chao AS, Chang SD, Lee KD, Wang TH, Wang HS, Soong YK. Functional network analysis on the transcriptomes of mesenchymal stem cells derived from amniotic fl uid, amniotic membrane, cord blood and bone marrow. Stem Cells. 2007;25(10):2511–23.
12. Xu W, Zhang X, Qian H, Zhu W, Sun X, Hu J, Zhou H, Chen Y. Mesenchymal stem cells from adult bone marrow differentiate into a cardiomyocyte phenotype in vitro. Soc Exp Biol Med. 2004;229(7):623–31.
13. Zhang DZ, Gai LY, Liu HW, Jin QH, Huang JH, Zhu XY. Transplantation of autologous adipose derived stem
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells

108
cells ameliorates cardiac function in rabbits with myo-cardial infarction. Chin Med J. 2007;120(4):300–7.
14. Bieback K, Kern S, Kutler H, Eichler H. Critical parameters for isolation of mesenchymal stem cells from umbilical cord. Stem Cells. 2004;22(4):625–34.
15. Boissy P, Malaval L, Jurdic P. Osteoblasts et osteo-clastes une cooperation exemplaire entre cellules mesenchymateuses et cellules hematopoietiques. Le Chercheur un Carrefour de l’Information; Revue Synthese 2007;1–18.
16. Park KS, Lee YS, Kang KS. In vitro neuronal and osteogenic differentiation of mesenchymal stem cells from human umbilical cord blood. J Vet Sci. 2006;7(4):343–8.
17. Sudo K, Kanno M, Miharada K, Ogawa S, Hiroyama T, Saijo K, Nakamura Y. Mesenchymal progenitors are able to differentiate into osteogenic, chondro-genic, and/or adipogenic cells in vitro are present in most primary fi broblast-like cell population. Stem Cells. 2007;25(7):1610–7.
18. Zuk PA, Zhu M, Asjian P, De Ugarte DA, Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P, Hedrick MH. Human adipose tissue is a source of multipotent stem cells. Mol Biol Cell. 2002;13(12):4279–95.
19. Sathishkumar S, Mohanashankar P, Boopalan PRJVC. Cell surface protein expression of stem cells from human adipose tissue at early passage with reference to mesenchymal stem cell phenotype. Intern J Med Med Sci. 2011;3(5):129–34.
20. Tuan RS, Chen FH. Cartilage. In: Botler A, Lear J, editors. Stem cells and gene based therapy. Berlin: Springer; 2006. p. 179–89.
21. De Bari C, Dell’Accio F. Mesenchymal stem cells in rheumatology: a regenerative approach to joint repair. Clin Sci (Lond). 2007;113(8):339–48.
22. Lin H, Xu R, Zhang Z, Chen L, Shi M, Wang FS. Implications of the immunoregulatory functions of mesenchymal stem cells in the treatment of human liver diseases. Cell Mol Immunol. 2011;8(1):19–22.
23. Niemeyer P, Krause U, Kasten P, Kreuz PC, Henle P, Südkamp NP, Mehlhorn A. Mesenchymal stem cell- based HLA-independent cell therapy for tissue engineering of bone and cartilage. Curr Stem Cell Res Ther. 2006;1(1):21–7.
24. Strem BM, Hicok KC, Zhu M, Wulur I, Alfonso Z, Schreiber RE, Fraser JK, Hedrick MH. Multipotential differentiation of adipose tissue-derived stem cells. Keio J Med. 2005;54(3):132–41.
25. Undale AH, Westendorf JJ, Yaszemski MJ, Khosla S. Mesenchymal stem cells for bone repair and metabolic bone diseases. Mayo Clin Proc. 2009;84(10):893–902.
26. Yanez R, Lamana ML, Castro JG, Colmenero I, Ramirez M, Bueren JA. Adipose tissue-derived mesenchymal stem cells have in vivo immunosuppres-sive properties applicable for the control of the graft-versus- host disease. Stem Cells. 2006;24(11):2582–91.
27. Ogawa R. The importance of adipose derived stem cells and vascularized tissue regeneration in the fi eld
of tissue transplantation. Curr Stem Cell Res Ther. 2006;1(1):13–20.
28. Tran TT, Kahn CR. Transplantation of adipose tissue and stem cells: role in metabolism and disease. Nat Rev Endocrinol. 2010;6(4):195–213.
29. Casteilla L, Planat-Benard V, Laharrague P, Cousin B. Adipose-derived stromal cells: their identity and uses in clinical trials, an update. World J Stem Cells. 2011;3(4):25–33.
30. Kucerova L, Altanerova V, Matuskova M, Tyciakova S, Altaner C. Adipose tissue derived human mesen-chymal stem cells mediated prodrug cancer gene therapy. Cancer Res. 2007;67(13):6304–13.
31. Lidong G, Shaoqing L, Yunfang W, Huimin Y, Daqing L, Lijuan H, Cixian B, Fang Y, Xue N, Shuangshuang S, Xuetao P. In vitro differentiation of human adipose derived mesenchymal stem cells into endothelial-like cells. Chin Sci Bull. 2006;51(15):1863–8.
32. Wisse BE. The infl ammatory syndrome: the role of adi-pose tissue cytokines in metabolic disorders linked to obesity. J Am Soc Nephrol. 2004;11(11):2792–800.
33. Mizuno H, Hyakusoku H. Mesengenic potential and future clinical perspective of human processed lipoaspi-rate cells. J Nippon Med Sch. 2003;70(4):300–6.
34. Mizuno H. Adipose derived stem cells for tissue repair and regeneration: ten years of research and lit-erature review. J Nippon Med Sch. 2009;76(2):56–66.
35. Banas A, Teratani T, Yamamoto YY, Takeshita F, Quinn G, Okochi H, Ochiya T. Adipose tissue derived mesenchymal stem cells as source of hepatocytes. Hepatology. 2007;46(1):219–28.
36. Wang Q, Steigelman MB, Walker JA, Chen S, Hornsby PJ, Bohnenblust ME, Wang HT. In vitro osteogenic differentiation of adipose stem cells after lentiviral transduction with green fl uorescent protein. J Craniofac Surg. 2009;20(6):2193–9.
37. Janmey PA, Winer JP, Weisel JW. Fibrin gels and their clinical and bioengineering applications. J R Soc Interface. 2009;6(30):1–10.
38. Ye Q, Zund G, Benedikt P, Jocknhoevel S, Hoerstrup SP, Sakyama S, Hubbell JA, Turina M. Fibrin gel as a three dimensional matrix in cardiovascular tissue engineering. Eur J Cardiothorac Surg. 2000;17(5):587–91.
39. Isaacson G, Herman JH. Autologous plasma fi brin glue: Rapid preparation. Am J Otolaryngol. 1996;17(2):92–4.
40. Quick AJ. Hemorrhagic diseases and thrombosis. 2nd ed. Philadelphia: Lee and Febiger; 1966. p. 442.
41. Abedin M, Tintut Y, Demer LL. Mesenchymal stem cells and the artery wall. Circ Res. 2004;95(7):671–7.
42. Efstratiadis G, Nikolaidou C, Vergoulas G. Leptin as a cardiovascular risk factor. Hippokratia. 2007;11(4):163–75.
43. Fantuzzi G, Mazzone T. Adipose tissue and athero-sclerosis: exploring the connection. Arterioscler Thromb Vasc Biol. 2007;27(5):996–1003.
T.C. Tran et al.

109
44. Jakob F, Limbert C, Schilling T, Benisch P, Seefried L, Ebert R. Biology of mesenchymal stem cells. Curr Rheumatol Rev. 2008;4:148–54.
45. Johnson RG, Leopold JA, Lo Scalzo J. Vascular calci-fi cation: pathobiological mechanisms and clinical implications. Circ Res. 2006;99(10):1044–59.
46. Jono S, Nishizawa Y, Shioi A, Morii H. 1,25-Dihydroxyvitamin D3 increases in vitro vascular calcifi cation by modulating secretion of endogenous parathyroid hormone-related peptide. Circulation. 1998;98(13):1302–6.
47. Reinders MEJ, Rabelink TJ. Adipose tissue derived stem cells: can impure cell preparation give pure results? Nephrol Dial Transplant. 2010;25(12):1–3.
48. Stompor T. An overview of the pathophysiology of vascular calcifi cation in chronic kidney disease. Perit Dial Int. 2007;27 Suppl 2:215–22.
49. Van Campenhout A, Moran CS, Parr A, Clancy P, Rush C, Jakubowski H, Golledge J. Role of homocys-teine in aortic calcifi cation and osteogenic cell differ-entiation. Atherosclerosis. 2009;202(2):557–66.
50. Pawitan JA. Prospect of adipose tissue derived mesen-chymal stem cells in regenerative medicine. Cell Tissue Transplant Ther. 2009;2:7–9.
51. Gomillion CT, Burg KJL. Stem cells and adipose tis-sue engineering. Biomaterials. 2006;27(36):6052–63.
52. Rebellatto CK, Aguiar AM, Moretao MP, Senegaglia AC, Hansen P, Barchiki F, Oliveira J, Martins J, Kuligowski C, Mansur F, Christofi s A, Amaral VF, Brofman PS, Goldbenberg S, Nakao LS, Correa A. Dissimilar differentiation of mesenchymal stem
cells from bone marrow, umbilical cord blood and adipose tissue. Exp Biol Med (Maywood). 2008;233(7):901–13.
53. Einhorn TA. Enhancement of fracture healing. J Bone Joint Surg Am. 1995;77(6):940–56.
54. O’Brien FJ, Farrell E, Waller MA, Connell I, O’Mahoney D, McGarry JP, Murphy BP, McHugh P, Campbell VA, Prendergast PJ. Scaffolds and cells: preliminary biomechanical analysis and results for the use of a collagen gag scaffold for bone tissue engi-neering. In: Prendergast PJ, McHugh PE, editors. Topics in bio-mechanical engineering. Dublin: Trinity Centre for Bio-Engineering (TCBE)/The National Centre for Biomedical Engineering Science (NCBES); 2004. p. 167–83.
55. Wu YC, Lee TM, Chiu KH, Shaw SY, Yang CY. A comparative study of the physical and mechanical properties of three natural corals based on the criteria for bone-tissue engineering scaffolds. J Mater Sci Mater Med. 2009;20(6):1273–80.
56. Georgiadis AN, Papavasiliou EC, Lourida ES, Alamanos Y, Kostara C, Tselepis AD, Drosos AA. Atherogenic lipid is a feature characteristic of patients with early rheumatoid arthritis: effect of early treat-ment a prospective, controlled study. Arthritis Res Ther. 2005;8(3):R82.
57. Mori K, Shioi A, Jono S, Nishizawa Y, Morii H. Dexamethasone enhances in vitro vascular calcifi ca-tion by promoting osteoblastic differentiation of vas-cular smooth muscle cells. Arterioscler Thromb Vasc Biol. 1999;19(2):2112–8.
6 Adipocytes and Osteoblasts from Human Adipose Tissue Mesenchymal Stem Cells




















