
Preterm birth and structural brain alterations in early adulthood
Upload
independentCategory
view
2download
0
ORIGINAL ARTICLE
Alterations of lipid metabolism and gene expression in ratadipocytes during chronic olanzapine treatmentJ Minet-Ringuet1,2, PC Even2, P Valet3, C Carpene3, V Visentin3, D Prevot3, D Daviaud3,
A Quignard-Boulange4, D Tome2 and R de Beaurepaire1,5
1Laboratoire de Psychopharmacologie, Centre Hospitalier Paul Guiraud, Villejuif, France; 2INRA, UMR physiologie de lanutrition et du comportement alimentaire, Institut National Agronomique Paris-Grignon, Paris Cedex, France; 3INSERM, U586,Unite de recherche sur les Obesites, Universite Paul Sabatier, Institut Louis Bugnard IFR31, Toulouse, France; 4INSERM,U465, Faculte de medecine, rue de l’ecole de medecine, Paris, France and 5INSERM, U513, Creteil, France
Although antipsychotics are established drugs in schizophrenia treatment, they are admittedlyknown to induce side effects favoring the onset of obesity and worsening its complications.Despite potential involvement of histamine receptor antagonism, or of other neurotransmittersystems, the mechanism by which antipsychotic drugs increase body weight is not elucidated.The aim of the present study was to investigate whether chronic antipsychotic treatments candirectly alter the regulation of two main functions of white adipose tissue: lipolysis andglucose utilization. The influence of a classical antipsychotic (haloperidol) was compared tothat of two atypical antipsychotics, one known to favor weight gain (olanzapine), the other not(ziprasidone). Cell size, lipolytic capacity and glucose transport activity were determined inwhite adipocytes of rats subjected to 5-week oral treatment with these antipsychotics. Geneexpression of adipocyte proteins involved in glucose transport or fat storage and mobilization,such as glucose transporters (GLUT1 and GLUT4), leptin, matrix metallo-proteinase-9 (MMP9),hormone-sensitive lipase (HSL) and fatty acid synthase (FAS) was also evaluated. Adipocytesfrom chronic olanzapine-treated rats exhibited decreased lipolytic activity, lowered HSLexpression and increased FAS expression. These changes were concomitant to enlarged fatdeposition and adipocyte size. Alterations were observed in adipocytes from olanzapine-treated rats whereas the other antipsychotics did not induce any notable disorder. Our resultstherefore show evidence of an effect of chronic antipsychotic treatment on rat adipocytemetabolism. Thus, impairment of fat cell lipolysis should be considered as a side effect ofcertain antipsychotics, leading, along with the already documented hyperphagia, to theexcessive weight gain observed in patients under prolonged treatment.Molecular Psychiatry (2007) 12, 562–571; doi:10.1038/sj.mp.4001948; published online 9 January 2007
Keywords: antipsychotic; olanzapine; adiposity; weight gain; adipocyte; lipid metabolism
Introduction
Body weight gain is a side effect frequently observedduring antipsychotic treatments,1–3 also suspected ofincreasing insulin resistance and the likehood ofcardiovascular complications.4,5 It has been suggestedthat antipsychotics interact with the complex systemsof neurotransmitters, neuropeptides and other mod-ulators in brain neuronal circuits like hypothalamus,solitary tract or cortical structures, to provokedisturbances in energy homeostasis and body weightcontrol.6
The relative antagonist properties of atypical anti-psychotics for histamine H1 receptors have beenreported to be a robust correlate to the rate of weight
gain in treated schizophrenic patients.3 Centraladministration of H1 antagonists in rats does indeedincrease food intake, whereas H1 agonists decreasefood intake,7–9 indicating a role for histamine in thecontrol of feeding behavior. In addition, antipsycho-tics may alter sympathetic activity,10 and histidine,the precursor of histamine, has been reported tostimulate lipolysis in white adipose tissue via acti-vation of the sympathetic nervous system (SNS).11
Therefore, all these things considered, adipose tissuefunction appears to constitute a potential direct targetof antipsychotics, at least according to their relativehistamine H1 receptor antagonism. Furthermore,norepinephrine and epinephrine are key elementsby which the SNS regulates thermogenesis, lipolysisand lipid oxidation, as well as adipocyte proliferationand development, via a- and b-adrenoreceptor (AR)activation.12,13 Antipsychotics reported to increasebody weight gain have also been suspected ofdisrupting the SNS control of energy homeostasis bydirectly acting on ARs.6,14,15
Received 13 July 2006; revised 2 October 2006; accepted 26October 2006; published online 9 January 2007
Correspondence: Dr R de Beaurepaire, Laboratoire de Psycho-pharmacologic, Centre Hospitalier Paul Guiraud, 54 avenue de laRepublique, 94806 Villejuif, France.E-mail: [email protected]
Molecular Psychiatry (2007) 12, 562–571& 2007 Nature Publishing Group All rights reserved 1359-4184/07 $30.00
www.nature.com/mp

Many animal studies, using mainly female rats,have investigated the metabolic effects of antipsycho-tics,16–19 but only scarce information has been re-ported on adipocytes themselves during chronicantipsychotic treatments. Baptista et al.16 did reportthat there was no significant change in the adipocytenumber per mass unit of adipose tissue in female ratsafter repeated administration of the antipsychoticagents sulpiride or risperidone, but did not analyzethe lipolytic or lipogenic activities in adipocytes. Toour knowledge, the literature does not report anydescription of insulin sensitivity or of responsivenessto lipolytic agents in adipocytes isolated from anti-psychotic-treated animals or patients under antipsy-chotic treatment.
The goal of the present study was to investigate atthe cellular level the potential alteration of adiposecell size and of its lipolytic/lipogenic functionalbalance induced by chronic antipsychotic treatments.To that purpose, we used a rat model of antipsychotic-induced obesity that we recently developed,20 andwhich consists in feeding male rats with a diet (calledP14L) enriched in carbohydrates and impoverished inproteins to a level comparable to that observed in the‘human diet’ (31% fat energy, 56% glucid energy,14% protein energy, whereas rats spontaneously eathigh amounts of protein energy (35%) and smallamounts of carbohydrate energy (15%)). We measuredand compared the effect of a classical (haloperidol)and atypical antipsychotics (olanzapine and ziprasi-done) on lipolysis and glucose transport in whiteadipocytes. In addition, fat pad weights and adipo-cyte size were measured. Gene expression of severalkey factors involved in trophic or metabolic adipocyteresponses were also measured, such as glucosetransporters (GLUT1 and GLUT4), leptin, matrixmetallo-proteinase-9 (MMP9), hormone-sensitive li-pase (HSL) and fatty acid synthetase (FAS). Theresults indicate that chronic olanzapine treatment canalter the adipocyte lipolytic responses.
Materials and methods
AnimalsMale Sprague–Dawley rats (Harlan, Gannat, France)initially weighing 200–220 g were individuallyhoused in a room maintained at 24711C and at5575% humidity with an artificial 12:12-h light–darkcycle (lights on at 08:00). Food and water wereavailable ad libitum throughout the experiment. Allrats were fed with the P14-L diet provided by theFrench national institute of agronomical research(INRA, Jouy-en-Josas, France). It is composed ofwhole-milk protein (140 g/kg), cornstarch (538.1 g/kg), sucrose (87.6 g/kg) and soybean oil (137 g/kg)(metabolizable energy: 17.5 kJ/g). This diet wassupplemented with minerals and vitamins accordingto the AIN-93M requirements (mineral salts 35 g/kg;vitamins 10 g/kg; cellulose 50 g/kg; choline 2.3 g/kg).21
This food was administered in powder form, asdescribed previously.20
Antipsychotic solutionsAntipsychotics were administered by incorporationinto the food. Haloperidol (Sigma-Aldrich, L’Isled’Abeau Chesnes, France) (1 mg/kg of body weight)and olanzapine (olanzapine orally desintegratingtablets) (1 mg/kg of body weight) were solubilized in10% ethanol and ziprasidone (Pfizer inc., NY, USA)(10 mg/kg BW) was diluted in a 10% ethanol solution.Daily food intake was measured during a period ofadaptation, which made it possible to adjust thequantity of antipsychotic to be incorporated in thefood.
Experimental procedureAfter 1 week of adaptation to environmental condi-tions, rats were separated into four groups (controls,haloperidol-treated, olanzapine-treated, ziprasidone-treated) according to weight, and received the anti-psychotic solutions mixed with their food during 5weeks. At the end of the study, the rats were killed bydecapitation and the adipose pads dissected out for exvivo studies.
Preparation of rat adipocytesAdipocytes were prepared from intra-abdominal(INWAT: epididymal, perirenal, retroperitoneal fatpads) and subcutaneous inguinal white adiposetissues (SCWAT) as previously described.22,23 Briefly,after digestion with liberase at 371C, isolated fat cellswere filtered and washed with a Krebs–Ringer buffer,pH 7.4, containing 12.5 mM (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), 2 mM pyruvate and3.5% (w/v) bovine serum albumin (KRH buffer). Cellsuspension was immediately used for measurementsof lipolysis, glucose transport and adipocyte weightand size, or rapidly frozen for RNA extraction.
Lipolysis measurementsAdipocyte suspension in KRH buffer was distributedas 400ml portions in plastic vials and incubated at371C with ongoing gentle shaking in the absence(basal) or in the presence of isoprenaline (b-adrener-gic agonist), phenylisopropyladenosine (PIA), forsko-lin, 3-isobutyl-1-methylxanthine (IBMX), UK 14304(a2-adrenergic agonist), insulin or antipsychotics atthe indicated final concentrations. After 90 min, theincubation tubes were placed on ice and 150ml of cellmedium was removed for the enzymatic determina-tion of glycerol as previously described.24 Cell lipidswere gravimetrically evaluated after extraction usingthe Dole and Meinertz25 method. Results wereexpressed as mmol of glycerol released per 100 mg oflipid during 90 min.
Glucose transport measurementAfter preincubation for 45 min at 371C with theindicated final concentrations of insulin or otherwisementioned drugs, each vial (400 ml) received anisotopic dilution of 2-deoxy-D-[3H]glucose, providinga final concentration of 0.1 mM (equivalent to ap-proximately 1 000 000 dpm/vial). Assays were then
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
563
Molecular Psychiatry

incubated for a 10-min period and arrested with100ml of 100 mM cytochalasin B. The radioactivityincorporated into the fat cells was counted asdescribed by Olefsky,26 using dinonyl phthalate toseparate intact fat cells from medium by a pulse spinas reported previously.27
RNA preparation and real-time RT-PCRTotal RNA was extracted using RNA easy minikit(Qiagen, Courtaboeuf, France). cDNA was synthesizedfrom 1 mg of total RNA using random hexamers. Weverified that the generated fluorescence was notoverestimated by contaminations resulting from re-sidual genomic DNA amplification (by using controlswithout reverse transcriptase) and/or from primer/dimer formation (controls with neither DNA templatenor reverse transcriptase). Gene expression wasassessed with real-time PCR in an ABI PRISM 7000Sequence Detection System instrument (AppliedBiosystems, Warrington, UK) starting with 12.5 ngof reverse-transcripted total RNA as previouslydescribed.28 The gene expression of proteins wellrecognized to be key elements in adipocyte biology,such as glucose transporters, GLUT1 and GLUT4,leptin, the enzymes HSL, FAS and MMP9 have beenanalyzed by using the previously described sets ofprimers.28
Adipocyte size measurementAdipocyte suspensions were observed under a micro-scope connected to a camera. Three pictures fromeach preparation were saved and the diameter of eachadipocyte was measured using Perfect Image software(Numeris, Paris, France), allowing calculation of amean fat cell size and weight.
Statistical analysisResults are presented as means7s.e.m. For lipolysis,glucose transport and RT-PCR, the statistical signifi-cance of differences was analyzed with Student’st-test. For adipocytes sizing, comparison of meanvalues between groups was evaluated by one-wayanalysis of variance with Tukey’s multiple compar-ison post-test (GraphPad Sofware, San Diego, CA,
USA). Differences were considered significant atP < 0.05.
Results
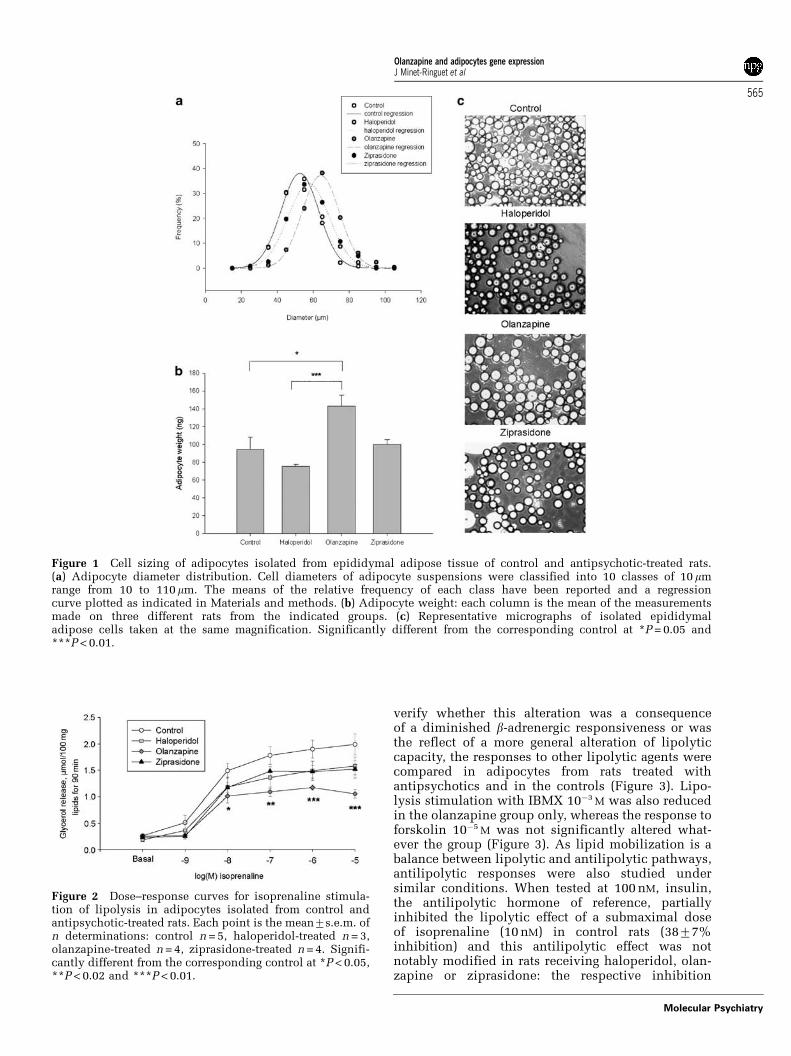
Adiposity at the end of treatmentsAlthough no difference in body weight was observedbetween the experimental groups, adiposity wassignificantly increased in olanzapine-treated rats asa result of an enlargement of both subcutaneous andvisceral fat pads (Table 1). In the epididymal fat padof control rats, the mean weight of fat cell wascalculated to average 94713 ng, and cell diametersmost frequently fell between 50 and 70mm (Figure 1).Mean adipocyte size and diameter distribution didnot differ in haloperidol- and ziprasidone-treated rats.However, adipocytes isolated from olanzapine-treatedgroup tended to be larger than in control, as shown inFigure 1c. A shift to the right was observed in thedistribution of diameter frequency of olanzapine-treated rats. Mean adipocyte weight of olanzapine-treated animals was larger than that of controls(olanzapine: 143712 ng, control: 94714 P = 0.05)(Figure 1). Food intake increased in olanzapine-treated animals without reaching significance (datanot shown); the increase in food intake was parallel tothe increase in body weight, similar to the resultsreported in our previously published experimentsusing the same technique.20
Lipolytic and antilipolytic activities in adipocytes fromcontrol and antipsychotic-treated ratsBasal lipolysis was unmodified in adipocytes isolatedfrom the INWAT of antipsychotic-treated rats whencompared to control. However, the dose-dependentstimulation of lipolysis induced by the b-adrenergicagonist isoprenaline was significantly weaker in theolanzapine-treated group than in the control group(Figure 2). When considering the fold increase overbasal lipolysis (set at 1), the higher dose of the b-adrenergic agonist (10�5 M) resulted in an activation of4.570.8-fold in the olanzapine-treated group, whichwas significantly lower than the corresponding8.371.3 observed in the control group (P < 0.05). To
Table 1 Body composition after 5 weeks of antipsychotic treatment
Group Control Haloperidol Olanzapine Ziprasidone
Body mass (g) 344.472.9 344.078.6 359.879.0 343.377.7INWAT mass (g) 5.970.6 7.672.0 8.470.8* 7.170.6SCWAT mass (g) 5.770.1 6.370.6 7.970.4*** 7.070.8Total WAT (%) 3.3870.19 4.1070.44 4.5270.28** 4.1370.30
Abbreviations: INWAT, intra-abdominal white adipose tissues; SCWAT, subcutaneous white adipose tissues; WAT, whiteadipose tissues.The mass of intra-abdominal (INWAT) or subcutaneous (SCWAT) white adipose tissues was obtained in the indicatedexperimental groups after 5-week treatment corresponding to a daily administration of 1 mg/kg body weight/day forhaloperidol and olanzapine, and 10 mg/kg/day for ziprasidone as detailed in Materials and methods. The total mass ofdissected WAT is expressed as percentage of body mass. Control n = 6, haloperidol n = 4, olanzapine n = 5, ziprasidone n = 4.Significantly different from control at *P < 0.05, **P < 0.02 and ***P < 0.01.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
564
Molecular Psychiatry
verify whether this alteration was a consequenceof a diminished b-adrenergic responsiveness or wasthe reflect of a more general alteration of lipolyticcapacity, the responses to other lipolytic agents werecompared in adipocytes from rats treated withantipsychotics and in the controls (Figure 3). Lipo-lysis stimulation with IBMX 10�3 M was also reducedin the olanzapine group only, whereas the response toforskolin 10�5 M was not significantly altered what-ever the group (Figure 3). As lipid mobilization is abalance between lipolytic and antilipolytic pathways,antilipolytic responses were also studied undersimilar conditions. When tested at 100 nM, insulin,the antilipolytic hormone of reference, partiallyinhibited the lipolytic effect of a submaximal doseof isoprenaline (10 nM) in control rats (3877%inhibition) and this antilipolytic effect was notnotably modified in rats receiving haloperidol, olan-zapine or ziprasidone: the respective inhibition
Figure 1 Cell sizing of adipocytes isolated from epididymal adipose tissue of control and antipsychotic-treated rats.(a) Adipocyte diameter distribution. Cell diameters of adipocyte suspensions were classified into 10 classes of 10 mmrange from 10 to 110mm. The means of the relative frequency of each class have been reported and a regressioncurve plotted as indicated in Materials and methods. (b) Adipocyte weight: each column is the mean of the measurementsmade on three different rats from the indicated groups. (c) Representative micrographs of isolated epididymaladipose cells taken at the same magnification. Significantly different from the corresponding control at *P = 0.05 and***P < 0.01.
Figure 2 Dose–response curves for isoprenaline stimula-tion of lipolysis in adipocytes isolated from control andantipsychotic-treated rats. Each point is the mean7s.e.m. ofn determinations: control n = 5, haloperidol-treated n = 3,olanzapine-treated n = 4, ziprasidone-treated n = 4. Signifi-cantly different from the corresponding control at *P < 0.05,**P < 0.02 and ***P < 0.01.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
565
Molecular Psychiatry

percentage being 4375, 5878 and 5577% (Table 2)(the use of a submaximal stimulation of lipolysis isthe best way to reveal potential antilipolytic effects).Similarly, the antilipolytic agent PIA (10�6 M) par-tially inhibited isoprenaline-induced stimulation oflipolysis in control as well as in antipsychotic-treatedrats (Table 2). The a2-adrenergic agonist UK 14304was much less efficient in counteracting the isoprena-line-induced lipolysis in control and its action wasnot modified in treated rats (not shown).
To further study the influence of antipsychotics onlipolysis regulation, it was tested whether they couldhave on their own an acute effect on adipocytes fromcontrol rats. As reported in Table 3, none of the testedantipsychotic drugs showed any lipolytic effect. Onthe contrary, ziprasidone 1 mM (an elevated dose)inhibited basal lipolysis, an effect which was not
mediated by its vehicle, 0.3% final dimethyl sulf-oxide. Neither the tested antipsychotics nor theirrespective vehicles were able to alter the lipolyticeffect of 10 nM isoprenaline (Table 3).
Glucose transport in adipocytes from control andantipsychotic-treated ratsBasal glucose uptake into adipocytes was not mod-ified in the antipsychotic-treated groups when com-pared to control. The dose–response curves forinsulin stimulation of hexose uptake were practicallysuperimposable for all groups (Figure 4). The max-imal activation was reached with 100 nM insulin andrepresented a eight- to 11-fold increase over basaluptake. Vanadate was also used to stimulate glucosetransport in order to test other agents than insulin.Vanadate (1 mM) partially reproduced the insulinstimulation in all groups (Table 4). The synergic
Figure 3 Adrenergic and nonadrenergic lipolysis stimula-tion in isolated adipocytes of control and antipsychotic-treated rats. Significantly different from control at *P < 0.05and ***P < 0.01. Each point is the mean7s.e.m. of fourdeterminations in control, olanzapine- and ziprasidone-treated and three in haloperidol-treated groups.
Table 2 Antilipolytic responses of adipocytes from controland antipsychotic-treated rats
Glycerol release (mmol/100 mg of lipids/90 min)
Control Haloperidol Olanzapine Ziprasidone
Basal 0.2570.04 0.1870.02 0.2570.04 0.2470.02Iso 1.5170.18 1.1870.19 1.0170.13 1.2570.04Isoþ Ins 1.0670.18 0.7470.11 0.5970.14 0.7670.05IsoþPIA 0.8870.19 0.8870.03 0.9270.17 0.9670.23
Abbreviations: INWAT, intra-abdominal white adiposetissues; PIA, phenylisopropyladenosine.Lipolysis was determined on adipocytes isolated fromINWAT of control and treated rats after a 90-min incubationwith submaximal dose of isoprenaline 10�8 M and PIA,(�)-phenylisopropyladenosine 10�6 M; Ins, insulin 10�7 M.Control n = 4, haloperidol n = 3, olanzapine n = 4, ziprasi-done n = 4. No significant difference between treated groupswas detected regarding inhibition of isoprenaline-inducedlipolysis.
Table 3 Short-term effects of antipsychotics on basal and isoprenaline-stimulated lipolysis in rat adipocytes
Glycerol release (mmol/100 mg lipids/90 min)
Addition to the medium None Isoprenaline 10 nM
Haloperidol Vehicle alone (1% ethanol) 0.3270.03 1.2870.240.1 mM 0.2970.02 1.1870.29
Olanzapine Vehicle alone (dist. water) 0.3370.02 1.3170.231 mM 0.2770.09 1.1970.21
Ziprasidone Vehicle alone (0.3% DMSO) 0.3370.04 1.3270.281 mM 0.0270.06** 1.0570.23
Abbreviation: DMSO, dimentyl sulfoxide.Data expressed as mean7s.e.m. of five determinations on adipocyte preparations of control rats. Final concentrations areindicated for all the added compounds, including vehicles. As drugs were added as a 4 ml portion in an incubation volume of400 ml, the one-hundredth dilution of distilled water in the adipocyte suspension corresponds to basal lipolysis. Nosignificant effect of the vehicles ethanol or DMSO detected when compared to basal or isoprenaline-stimulated lipolysis.Significantly different from the corresponding vehicle at **P < 0.01.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
566
Molecular Psychiatry
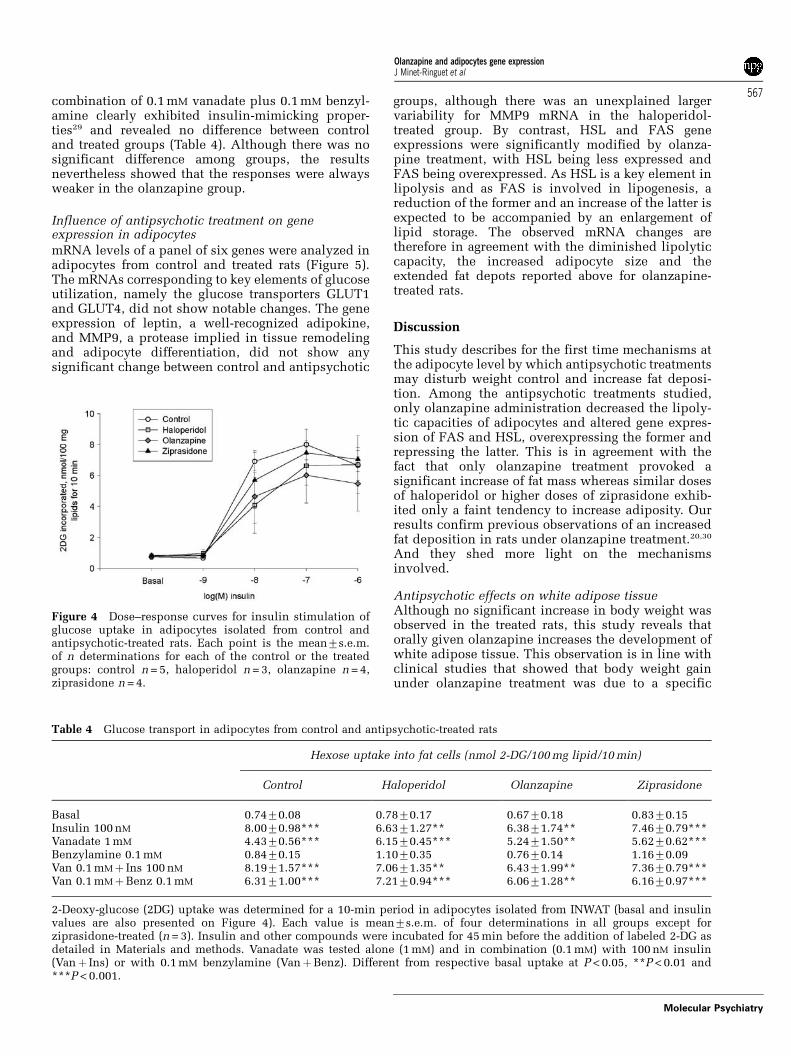
combination of 0.1 mM vanadate plus 0.1 mM benzyl-amine clearly exhibited insulin-mimicking proper-ties29 and revealed no difference between controland treated groups (Table 4). Although there was nosignificant difference among groups, the resultsnevertheless showed that the responses were alwaysweaker in the olanzapine group.
Influence of antipsychotic treatment on geneexpression in adipocytes
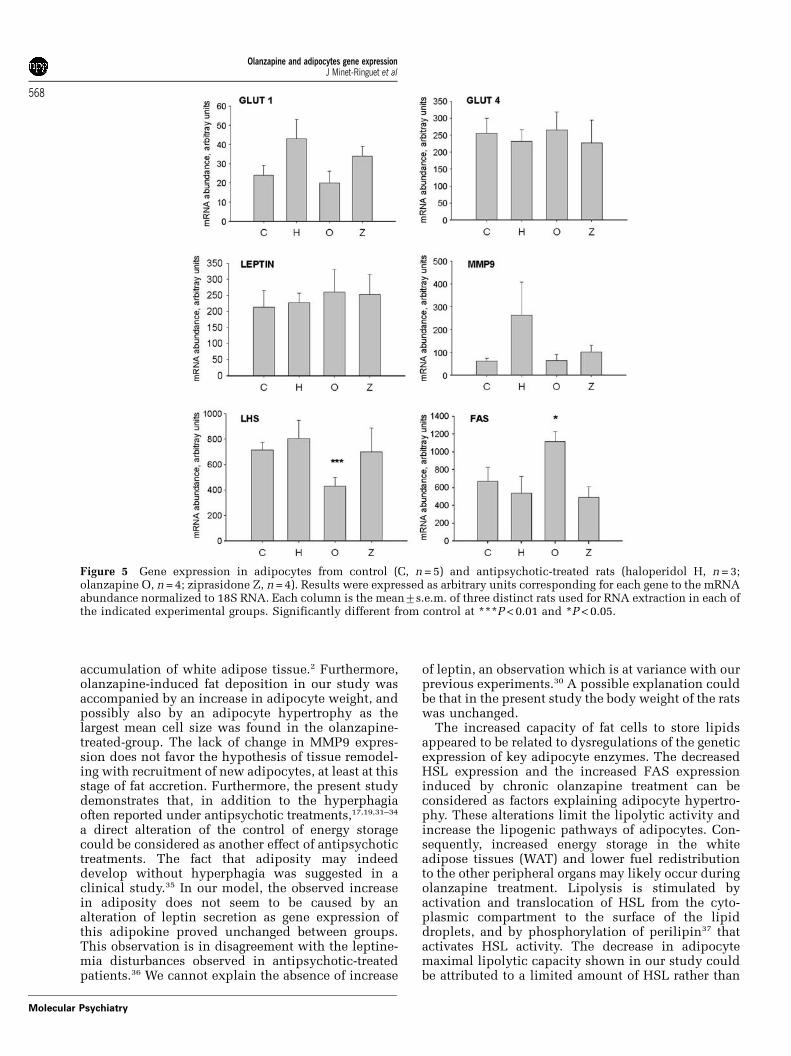
mRNA levels of a panel of six genes were analyzed inadipocytes from control and treated rats (Figure 5).The mRNAs corresponding to key elements of glucoseutilization, namely the glucose transporters GLUT1and GLUT4, did not show notable changes. The geneexpression of leptin, a well-recognized adipokine,and MMP9, a protease implied in tissue remodelingand adipocyte differentiation, did not show anysignificant change between control and antipsychotic
groups, although there was an unexplained largervariability for MMP9 mRNA in the haloperidol-treated group. By contrast, HSL and FAS geneexpressions were significantly modified by olanza-pine treatment, with HSL being less expressed andFAS being overexpressed. As HSL is a key element inlipolysis and as FAS is involved in lipogenesis, areduction of the former and an increase of the latter isexpected to be accompanied by an enlargement oflipid storage. The observed mRNA changes aretherefore in agreement with the diminished lipolyticcapacity, the increased adipocyte size and theextended fat depots reported above for olanzapine-treated rats.
Discussion
This study describes for the first time mechanisms atthe adipocyte level by which antipsychotic treatmentsmay disturb weight control and increase fat deposi-tion. Among the antipsychotic treatments studied,only olanzapine administration decreased the lipoly-tic capacities of adipocytes and altered gene expres-sion of FAS and HSL, overexpressing the former andrepressing the latter. This is in agreement with thefact that only olanzapine treatment provoked asignificant increase of fat mass whereas similar dosesof haloperidol or higher doses of ziprasidone exhib-ited only a faint tendency to increase adiposity. Ourresults confirm previous observations of an increasedfat deposition in rats under olanzapine treatment.20,30
And they shed more light on the mechanismsinvolved.
Antipsychotic effects on white adipose tissueAlthough no significant increase in body weight wasobserved in the treated rats, this study reveals thatorally given olanzapine increases the development ofwhite adipose tissue. This observation is in line withclinical studies that showed that body weight gainunder olanzapine treatment was due to a specific
Table 4 Glucose transport in adipocytes from control and antipsychotic-treated rats
Hexose uptake into fat cells (nmol 2-DG/100 mg lipid/10 min)
Control Haloperidol Olanzapine Ziprasidone
Basal 0.7470.08 0.7870.17 0.6770.18 0.8370.15Insulin 100 nM 8.0070.98*** 6.6371.27** 6.3871.74** 7.4670.79***Vanadate 1 mM 4.4370.56*** 6.1570.45*** 5.2471.50** 5.6270.62***Benzylamine 0.1 mM 0.8470.15 1.1070.35 0.7670.14 1.1670.09Van 0.1 mMþ Ins 100 nM 8.1971.57*** 7.0671.35** 6.4371.99** 7.3670.79***Van 0.1 mMþBenz 0.1 mM 6.3171.00*** 7.2170.94*** 6.0671.28** 6.1670.97***
2-Deoxy-glucose (2DG) uptake was determined for a 10-min period in adipocytes isolated from INWAT (basal and insulinvalues are also presented on Figure 4). Each value is mean7s.e.m. of four determinations in all groups except forziprasidone-treated (n = 3). Insulin and other compounds were incubated for 45 min before the addition of labeled 2-DG asdetailed in Materials and methods. Vanadate was tested alone (1 mM) and in combination (0.1 mM) with 100 nM insulin(Vanþ Ins) or with 0.1 mM benzylamine (VanþBenz). Different from respective basal uptake at P < 0.05, **P < 0.01 and***P < 0.001.
Figure 4 Dose–response curves for insulin stimulation ofglucose uptake in adipocytes isolated from control andantipsychotic-treated rats. Each point is the mean7s.e.m.of n determinations for each of the control or the treatedgroups: control n = 5, haloperidol n = 3, olanzapine n = 4,ziprasidone n = 4.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
567
Molecular Psychiatry
accumulation of white adipose tissue.2 Furthermore,olanzapine-induced fat deposition in our study wasaccompanied by an increase in adipocyte weight, andpossibly also by an adipocyte hypertrophy as thelargest mean cell size was found in the olanzapine-treated-group. The lack of change in MMP9 expres-sion does not favor the hypothesis of tissue remodel-ing with recruitment of new adipocytes, at least at thisstage of fat accretion. Furthermore, the present studydemonstrates that, in addition to the hyperphagiaoften reported under antipsychotic treatments,17,19,31–34
a direct alteration of the control of energy storagecould be considered as another effect of antipsychotictreatments. The fact that adiposity may indeeddevelop without hyperphagia was suggested in aclinical study.35 In our model, the observed increasein adiposity does not seem to be caused by analteration of leptin secretion as gene expression ofthis adipokine proved unchanged between groups.This observation is in disagreement with the leptine-mia disturbances observed in antipsychotic-treatedpatients.36 We cannot explain the absence of increase
of leptin, an observation which is at variance with ourprevious experiments.30 A possible explanation couldbe that in the present study the body weight of the ratswas unchanged.
The increased capacity of fat cells to store lipidsappeared to be related to dysregulations of the geneticexpression of key adipocyte enzymes. The decreasedHSL expression and the increased FAS expressioninduced by chronic olanzapine treatment can beconsidered as factors explaining adipocyte hypertro-phy. These alterations limit the lipolytic activity andincrease the lipogenic pathways of adipocytes. Con-sequently, increased energy storage in the whiteadipose tissues (WAT) and lower fuel redistributionto the other peripheral organs may likely occur duringolanzapine treatment. Lipolysis is stimulated byactivation and translocation of HSL from the cyto-plasmic compartment to the surface of the lipiddroplets, and by phosphorylation of perilipin37 thatactivates HSL activity. The decrease in adipocytemaximal lipolytic capacity shown in our study couldbe attributed to a limited amount of HSL rather than
Figure 5 Gene expression in adipocytes from control (C, n = 5) and antipsychotic-treated rats (haloperidol H, n = 3;olanzapine O, n = 4; ziprasidone Z, n = 4). Results were expressed as arbitrary units corresponding for each gene to the mRNAabundance normalized to 18S RNA. Each column is the mean7s.e.m. of three distinct rats used for RNA extraction in each ofthe indicated experimental groups. Significantly different from control at ***P < 0.01 and *P < 0.05.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
568
Molecular Psychiatry

to a dysregulation of HSL activation by perilipin,because the lack of perilipin tends to increase basallipolysis,38 and no difference was observed for basallipolysis between control and olanzapine-treatedgroups. The determinisms for the decrease in HSLand the increase in FAS expression remain unknownand deserve further investigations that may contributeto a better understanding of adipose tissue dysfunc-tion under antipsychotic challenge. As FAS is alsoexpressed in hepatocytes, it would also appearcritical to investigate whether liver can provideenergy to the whole organism whereas WAT is sparingits lipid stores.
Alteration of lipolytic and glucose transport activitiesin adipocytesDifferences in the pharmacological profile betweenziprasidone and olanzapine are suspected to be at theorigin of their widely recognized different effects onweight gain: very slight for the former and robust forthe latter.1 Similarly, the chronic effects of antipsy-chotics on adipocyte biology appear to depend on thenature of the antipsychotic agent. In this view,ziprasidone did not modify lipolytic responsivenessin the present model and did not alter HSL or FASexpression. Ziprasidone was also distinguishablefrom olanzapine in its tendency to inhibit basallipolysis. Whether these differences indicate that,despite its tendency to slightly increase fat stores,ziprasidone could be devoid of the deleterious effectsof olanzapine on glycemic control revealed in clinicalstudies5,6 remains to be established. No insulinresistance was detected in the adipocytes fromantipsychotic-treated animals whether on the antili-polytic or the glucose uptake responses to thepancreatic hormone. Similarly, the insulin mimickingagents, vanadate and benzylamine, were still activein adipocytes from antipsychotic-treated animals.Such lack of insulin action impairment by anti-psychotics was unexpected, as several of thesedrugs have been demonstrated elsewhere to developinsulin resistance, but might possibly be explainedby a reversible inhibitory action alleviated duringfat cell preparation. However, no effect of olanzapineon antilipolytic or glucose uptake responses wasdetected after chronic treatment or acute in vitroincubation.
According to Poyurovky et al.,39 the adrenergicsystem may be relevant to antipsychotic-inducedweight gain, as the activation of the adrenergic systemcan attenuate antipsychotic-induced weight gain. Inour study, the decrease in the lipolytic action ofisoprenaline in olanzapine-treated rats might have ledone to suppose that there was a reduction in the b-adrenergic receptivity of the adipocytes of olanza-pine-treated animals. However, the reduced responseto IBMX, a lipolytic agent with a mechanism of actionindependent from b-AR activation, together with thelack of direct inhibitory effect of olanzapine onisoprenaline-stimulated lipolysis, ruled out this hy-pothesis. Haloperidol, olanzapine and ziprasidone
have moderate a1- and weak a2-AR antagonist proper-ties.40 a1-AR are not involved in lipolysis in the rat.12
Therefore a1-antagonist properties of olanzapine areunlikely involved in its antilipolysis effects. Ourobservations showed that the a2-adrenergic antilipo-lysis was very weak in adipocytes of antipsychotic-treated animals, whatever be the experimental group.Thus, the a2-component is unlikely to represent themechanism of action of antipsychotic-induced im-pairment of lipid mobilization. The neurotransmittersystems or receptors involved in the antilipolyticactivity of olanzapine remain to be elucidated. Inparticular, the H1 blocking effect of olanzapine,which is not shared by haloperidol and ziprasidone,needs to be investigated.
Our study of glucose transport did not evidence anyclearcut alteration of insulin responsiveness at theadipocyte level. As glucose availability is critical forlipogenesis, such alteration would have impairedadipocyte replenishment and fat depot developmentunder antipsychotic treatment. A number of studieshave been recently devoted to the impairing effects ofolanzapine on glucose homeostasis. The results ofthese studies are not unequivocal. Engl et al.41 haveshown that olanzapine impairs glycogen synthesisand insulin signaling in skeletal muscle cells. On theother hand, Robinson et al.42 have observed thatolanzapine does not affect glucose transport in invitro preparations of adipocytes. These last resultscorrespond to our findings. However, Vestri et al.43
have shown that olanzapine (at a fairly high dose)decreases the maximal insulin-stimulated glucosetransport rate in adipocytes. Therefore, further workis needed to determine the effects of olanzapine onglucose transport and metabolism in adipocytes. It isalso possible that insulin resistance and metabolicsyndrome induced by antipsychotics5,6 are late dia-betic complications driven by exaggerated fat deposi-tion or generated by defects in pancreatic secretion oreven due to defects in insulin responsiveness inskeletal muscles or liver.
In conclusion, our study shows that the cellfunctions of a peripheral organ actively participatingin energy homeostasis are clearly disturbed afterolanzapine treatment. The adipocyte lipolysis isclassically known to be stimulated by the ortho-SNSand a disorder in this autonomic pathway might besecondary to an action of certain antipsychotics onthe central nervous system. However, direct long-termactions of antipsychotics on adipocyte metabolismcannot not be excluded. Although there are stillmissing links between olanzapine ingestion andalteration of adipocyte lipolysis, HSL and FAS geneexpression, our results show for the first time thatprimary alterations induced by olanzapine not onlyinclude energy intake disorders but also dysregula-tions of lipid accumulation and mobilization. Ourresults indicate that the adipose tissue may be a keytarget of antipsychotics in relation to their undesir-able side effects on the regulation of energy storageand expenditure.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
569
Molecular Psychiatry

Acknowledgments
This study was supported by a grant from Pfizer (NewYork). Julie Minet-Ringuet was supported by a grantfrom La Fondation pour la Recherche Medicale. Wethank Brooke Maddux for re-reading and correctingthe manuscript.
References
1 Allison DB, Mentore JL, Heo M, Chandler LP, Cappelleri JC,Infante MC et al. Antipsychotic-induced weight gain: a compre-hensive research synthesis. Am J Psychiatry 1999; 156: 1686–1696.
2 Eder U, Mangweth B, Ebenbichler C, Weiss E, Hofer A, Hummer Met al. Association of olanzapine-induced weight gain with anincrease in body fat. Am J Psychiatry 2001; 158: 1719–1722.
3 Wirshing DA, Wirshing WC, Kysar L, Berisford MA, Goldstein D,Pashdag J et al. Novel antipsychotics: comparison of weight gainliabilities. J Clin Psychiatry 1999; 60: 358–363.
4 Haupt DW, Newcomer JW. Hyperglycemia and antipsychoticmedications. J Clin Psychiatry 2001; 62: 15–26; discussion 40–41.
5 Wirshing DA, Boyd JA, Meng LR, Ballon JS, Marder SR, WirshingWC. The effects of novel antipsychotics on glucose and lipidlevels. J Clin Psychiatry 2002; 63: 856–865.
6 Basile VS, Masellis M, McIntyre RS, Meltzer HY, Lieberman JA,Kennedy JL. Genetic dissection of atypical antipsychotic-inducedweight gain: novel preliminary data on the pharmacogeneticpuzzle. J Clin Psychiatry 2001; 62: 45–66.
7 Itowi N, Nagai K, Nakagawa H, Watanabe T, Wada H. Changes inthe feeding behavior of rats elicited by histamine infusion. PhysiolBehav 1988; 44: 221–226.
8 Lecklin A, Etu-Seppala P, Stark H, Tuomisto L. Effects ofintracerebroventricularly infused histamine and selective H1, H2and H3 agonists on food and water intake and urine flow in Wistarrats. Brain Res 1998; 793: 279–288.
9 Morimoto T, Yamamoto Y, Yamatodani A. Brain histamine andfeeding behavior. Behav Brain Res 2001; 124: 145–150.
10 Kudoh A, Ishihara H, Matsuki A. Pituitary–adrenal and para-sympathetic function in chronic schizophrenic patients withpostoperative ileus or hypotension. Neuropsychobiology 1999;39: 125–130.
11 Yoshimatsu H, Tsuda K, Niijima A, Tatsukawa M, Chiba S, SakataT. Histidine induces lipolysis through sympathetic nerve in whiteadipose tissue. Eur J Clin Invest 2002; 32: 236–241.
12 Lafontan M, Bousquet-Melou A, Galitzky J, Barbe P, Carpene C,Langin D et al. Adrenergic receptors and fat cells: differentialrecruitment by physiological amines and homologous regulation.Obes Res 1995; 3: 507S–514S.
13 Cannon B, Nedergaard J. Brown adipose tissue: function andphysiological significance. Physiol Rev 2004; 84: 277–359.
14 Ashby Jr CR, Wang RY. Pharmacological actions of the atypicalantipsychotic drug clozapine: a review. Synapse 1996; 24:349–394.
15 Bymaster FP, Hemrick-Luecke SK, Perry KW, Fuller RW. Neuro-chemical evidence for antagonism by olanzapine of dopamine,serotonin, alpha 1-adrenergic and muscarinic receptors in vivo inrats. Psychopharmacology (Berlin) 1996; 124: 87–94.
16 Baptista T, de Baptista EA, Lalonde J, Plamondon J, Kin NM,Beaulieu S et al. Comparative effects of the antipsychoticssulpiride and risperidone in female rats on energy balance, bodycomposition, fat morphology and macronutrient selection. ProgNeuropsychopharmacol Biol Psychiatry 2004; 28: 1305–1311.
17 Baptista T, Araujo de Baptista E, Ying Kin NM, Beaulieu S, WalkerD, Joober R et al. Comparative effects of the antipsychoticssulpiride or risperidone in rats. I: bodyweight, food intake, bodycomposition, hormones and glucose tolerance. Brain Res 2002;957: 144–151.
18 Goudie AJ, Smith JA, Halford JC. Characterization of olanzapine-induced weight gain in rats. J Psychopharmacol 2002; 16:291–296.
19 Pouzet B, Mow T, Kreilgaard M, Velschow S. Chronic treatmentwith antipsychotics in rats as a model for antipsychotic-induced
weight gain in human. Pharmacol Biochem Behav 2003; 75:133–140.
20 Minet-Ringuet J, Even PC, Lacroix M, Tome D, de Beaurepaire R. Amodel for antipsychotic-induced obesity in the male rat. Psycho-pharmacology 2006; 187: 447–454.
21 Reeves PG, Nielsen FH, Fahey Jr GC. AIN-93 purified diets forlaboratory rodents: final report of the American Institute ofNutrition ad hoc writing committee on the reformulation of theAIN-76A rodent diet. J Nutr 1993; 123: 1939–1951.
22 Rodbell M. Localization of lipoprotein lipase in fat cells of ratadipose tissue. J Biol Chem 1964; 239: 753–755.
23 Honnor RC, Dhillon GS, Londos C. cAMP-dependent proteinkinase and lipolysis in rat adipocytes. I. Cell preparation,manipulation, and predictability in behavior. J Biol Chem 1985;260: 15122–15129.
24 Wieland O. Enzymic method for the determination of glycerin.Biochem Z 1957; 329: 313–319.
25 Dole VP, Meinertz H. Microdetermination of long-chain fatty acidsin plasma and tissues. J Biol Chem 1960; 235: 2595–2599.
26 Olefsky JM. Mechanisms of the ability of insulin to activate theglucose-transport system in rat adipocytes. Biochem J 1978; 172:137–145.
27 Carpene C, Chalaux E, Lizarbe M, Estrada A, Mora C, Palacin Met al. Beta 3-adrenergic receptors are responsible for the adrenergicinhibition of insulin-stimulated glucose transport in rat adipo-cytes. Biochem J 1993; 296: 99–105.
28 Le Lay S, Krief S, Farnier C, Lefrere I, Le Liepvre X, Bazin R et al.Cholesterol, a cell size-dependent signal that regulates glucosemetabolism and gene expression in adipocytes. J Biol Chem 2001;276: 16904–16910.
29 Enrique-Tarancon G, Marti L, Morin N, Lizcano JM, Unzeta M,Sevilla L et al. Role of semicarbazide-sensitive amine oxidase onglucose transport and GLUT4 recruitment to the cell surface inadipose cells. J Biol Chem 1998; 273: 8025–8032.
30 Minet-Ringuet J, Even PC, Guesdon B, Tome D, de Beaurepaire R.Effects of chronic neuroleptic treatments on nutrient selection,body weight, and body composition in the male rat under dietaryself-selection. Appetite 2006; 46: 254–262.
31 Baptista T, Contreras Q, Teneud L, Albornoz MA, Acosta A,Paez X et al. Mechanism of the neuroleptic-induced obesity infemale rats. Prog Neuropsychopharmacol Biol Psychiatry 1998; 22:187–198.
32 Arjona AA, Zhang SX, Adamson B, Wurtman RJ. An animal modelof antipsychotic-induced weight gain. Behav Brain Res 2004; 152:121–127.
33 Kaur G, Kulkarni SK. Studies on modulation of feeding behaviorby atypical antipsychotics in female mice. Prog Neuropsycho-pharmacol Biol Psychiatry 2002; 26: 277–285.
34 Thornton-Jones Z, Neill JC, Reynolds GP. The atypical antipsy-chotic olanzapine enhances ingestive behaviour in the rat: apreliminary study. J Psychopharmacol 2002; 16: 35–37.
35 Virkkunen M, Wahlbeck K, Rissanen A, Naukkarinen H, Franssila-Kallunki A. Decrease of energy expenditure causes weightincrease in olanzapine treatment – a case study. Pharmaco-psychiatry 2002; 35: 124–126.
36 Kraus T, Haack M, Schuld A, Hinze-Selch D, Kuhn M, Uhr M et al.Body weight and leptin plasma levels during treatment withantipsychotic drugs. Am J Psychiatry 1999; 156: 312–314.
37 Souza SC, Muliro KV, Liscum L, Lien P, Yamamoto MP, Schaffer JEet al. Modulation of hormone-sensitive lipase and protein kinaseA-mediated lipolysis by perilipin A in an adenoviral reconstitutedsystem. J Biol Chem 2002; 277: 8267–8272.
38 Martinez-Botas J, Anderson JB, Tessier D, Lapillonne A, Chang BH,Quast MJ et al. Absence of perilipin results in leannessand reverses obesity in Lepr(db/db) mice. Nat Genet 2000; 26:474–479.
39 Poyurovsky M, Isaacs I, Fuchs C, Schneidman M, Faragian S,Weizman R et al. Attenuation of olanzapine-induced weight gainwith reboxetine in patients with schizophrenia: a double-blind,placebo-controlled study. Am J Psychiatry 2003; 160: 297–302.
40 Duncan GE, Zorn S, Lieberman JA. Mechanisms of typical andatypical antipsychotic drug action in relation to dopamine andNMDA receptor hypofunction hypotheses in schizophrenia. MolPsychiatry 1999; 4: 418–428.
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
570
Molecular Psychiatry

41 Engl J, Laimer M, Niederwanger A, Kranebitter M, Starzinger M,Pedrini MT et al. Olanzapine impairs glycogen synthesis andinsulin signaling in L6 skeletal muscle cells. Mol Psychiatry 2005;10: 1089–1096.
42 Robinson KA, Yacoub Wasef SZ, Buse MG. At therapeuticconcentrations, olanzapine does not affect basal or insulin-
stimulated glucose transport in 3T3-L1 adipocytes. Prog Neuro-psychopharmacol Biol Psychiatry 2006; 30: 93–98.
43 Vestri HS, Maianu L, Moellering DR, Garvey WT. Atypicalantipsychotic drugs directly impair insulin action in adipocytes:effects on glucose transport, lipogenesis, and antilipolysis.Neuropsychopharmacology 2006 [E-pub ahead of print].
Olanzapine and adipocytes gene expressionJ Minet-Ringuet et al
571
Molecular Psychiatry
Copyright © 2022 FDOKUMEN




















