
Measurement of Fecal Testosterone Metabolites in Mice - MDPI
Upload
independentCategory
view
1download
0
International Journal of Andrology, 1988, 11, pages 339-348
Nutrition-related alterations in liver microsomal testosterone hydroxylases
L. G I L * , M Y R I A M O R E L L A N A , H . V A S Q U E Z and M A R C E L A SILVA Medicine, University of Chile, PO Box 70086, Santiago, Chile
Department of Biochemistry, Faculty of
Summary The oxidation products of testosterone formed by liver microsomes from normal- fed and protein-energy malnourished male rats have been analysed by HPLC. Microsomes from normal-fed rats oxidized testosterone at a rate of 4.52 nmol/min/ mg protein. The major products formed were: 6p-, 7a- and l6a-hydroxytestos- terone; these three metabolites represented 65% of the total testosterone meta- bolism. Microsomes from protein-energy malnourished rats oxidized testosterone at a reduced rate of 2.03 nmol/min/mg protein. The major product formed was 7a- hydroxytestosterone, which accounted for 43% of total testosterone oxidation. Microsomes from protein-energy malnourished rats showed a CO-reduced cyto- chrome P-450 spectra with a maxima at 452 nm, and a 38% decrease in the total content of cytochrome P-450. Some testosterone hydroxylases were drastically affected by protein-energy malnutrition but others, such as 7a-hydroxylase, remained unchanged. The present results suggest that nutritional status can modify the relative amounts of individual cytochrome P-450 isozymes, thus explaining the observed changes in several testosterone hydroxylases. Protein-energy malnutri- tion seems to be an excellent tool with which to obtain a microsomal fraction containing predominantly P-450 isozymes, which are probably involved in key mono-oxygenations of physiological substrates.
Keywords: testosterone hydroxylases, cytochrome P-450, nutritional regulation.
Introduction Cytochrome P-450 is a member of a group of enzymes known as mono-oxygenases, which catalyse the addition of oxygen to a substrate via molecular oxygen activation (White & Coon, 1980). In the rat, liver microsomal P-450 represents an unknown number of haemoproteins capable of metabolizing endogenous compounds such as steroids, fatty acids and prostaglandins, as well as exogenous chemicals, including numerous drugs and carcinogens (Lu & West, 1980). It is generally accepted that the wide versatility of these haemoprotein groups in oxidizing different structures results from the presence of multiple forms of isozymes possessing a broad sub-
Abbreviations: HPLC, high-performance liquid chromatography; Tris-HC1, tris (hydroxymethyl) aminomethane hydrochloride; SDS-PAGE, sodium dodecyl sulphate polyacrylamide gel electro- phoresis; OHA, hydroxyandrostenedione; OHT, hydroxytestosterone. *To whom correspondence should be addressed.
339

340 L. Gif et al.
strate specificity. The relative amounts of the individual cytochrome P-450 iso- zymes are markedly dependent on the sex, species, age, hormonal and genetic constitution, as well as on prior exposure to haemoprotein inducers (Thomas et a f . , 1981; Kato & Kamataki, 1982; Waxman, 1984; Waxman, Dannan & Guengerich, 1985).
Although much less studied, the oxidative activity of the mono-oxygenase system is also regulated by the nutritional status of the animal (Wiebel & Gelboin, 1975; Hietanen, 1980; Hasumura et a f . , 1983). We have demonstrated that rat liver benzo[a]pyrene hydroxylase is affected drastically during protein-energy malnutri- tion (Salazar, Litvak & Gil, 1983). The enzyme levels were almost undetectable in malnourished rats, but were increased rapidly to normal levels by feeding mal- nourished rats with a high-quality protein diet. It was concluded that nutritional status drastically changes the levels of some P-450 isozymes, providing an explan- ation for the observed changes in aryl hydrocarbon hydroxylase activity. More recently, Gil et a f . (1988) have purified to homogeneity and characterized a cytochrome P-450 isozyme from protein-energy malnourished rats induced with (3-naphthoflavone.
Many steroid hormones are hydroxylated in the liver by the mono-oxygenase system, leading to the formation of inactive or less active hormones. These com- pounds can be excreted in bile or urine, or may contribute to the formation of sterols having specific physiological functions, some not yet recognized. Testos- terone and androstenedione, high affinity natural substrates for the liver mono- oxygenase system, are hydroxylated stereo-selectively at multiple positions around the steroid ring (Gustafsson, 1978; Sheets & Estabrook, 1985; Sheets et a f . , 1986). The main products formed by liver microsomes from untreated rats are: andro- stenedione, 2a-, 6p-, 7a- and 16a-hydroxytestosterone (Shiverick & Neims, 1979; Wood et a f . , 1983). Hydroxylation at position 2, however, has been reported to be 2 a instead of 28 (Nakamura & Ueda, 1980). Traces of 16(3-hydroxytestosterone have been detected in uninduced rats, although this product is induced drastically by phenobarbital (van der Hoeven, 1981; Wood et a f . , 1983).
The purpose of the present study was to demonstrate the influence of the nutritional status of the animal on male liver microsomal P-450 and testosterone hydroxylation. We describe an experimental model of protein-energy malnutrition obtained after a long period of protein deprivation, which modifies the content of some liver cytochrome P-450 isozymes resulting in a lower capability of the mono- oxygenase system to hydroxylate testosterone. We propose that another isozyme, which is not affected by this extremely drastic nutritional stress, might play an important physiological role in the metabolism of testosterone.
Materials and methods
Animals and diets Male Wistar rats were used in all experiments. The following groups of animals were studied:
Malnourished group. Protein-energy malnutrition was provoked with the follow- ing experimental model: after birth, 16 offspring from the same litter were nursed

Nutritional alterations in testosterone hydroxylases 341
by one mother for 21 days and then, for 14 days, were offered water ad libitum and a protein-free diet, as described by Salazar et al. (1983). Animals from this group are referred to as ‘malnourished rats’.
Normal group. After parturition, eight offspring from the same litter were nursed by one mother for 21 days and then, for 14 days, were offered water ad libitum and a commercial balanced-diet pellet purchased from Alimentos Cisternas, S.A. Animals from this group are referred to as ‘normal-fed rats’. Normal-fed and malnourished rats were killed at 35 days of age. In each experiment three to five normal-fed or six to eight malnourished rats were used.
Preparation of microsomes Animals were killed by decapitation and exsanguinated. Livers were removed, pooled, weighed, minced and rinsed several times with 1.14% KCI-10 mM EDTA, pH 7.5. They were then homogenized in four volumes 0.1 M Tris-HC1-5 mM EDTA, pH 7.4. Microsomes were obtained by differential centrifugation as described by Salazar et al. (1983), but were washed with 0.25 M Tris, pH 7.4, and stored at -70°C. No change in activity was detected after 1 week of storage. Pellets were resuspended in 0.25 M Tris-HCI, pH 7.2, for measurement of hydroxylase activities.
Cytochrome P-450 Concentrations of cytochrome P-450 were measured in microsomal suspensions containing 0.5-1.0 mg protein by the method of Omura & Sat0 (1964), and the protein content estimated by the method of Lowry et al. (1951) using bovine serum albumin as standard.
Testosterone hydroxylation The incubation mixture for testosterone hydroxylation provided linear response with respect to both time and protein concentration for all metabolites assayed (2a-, 68-, 7a- and 16a-hydroxytestosterones, androstenedione and 66-hydroxy- androstenedione). Each incubation contained in 1 ml: 17 pmol Tris-HCI pH 7.2; 60 pmol KCl; 5 pmol MgCI,; 138 nmol testosterone (added in 20 p1 of methanol) and 1 mg of microsomal protein. After a 5-min pre-incubation at 37”C, the reaction was started by the addition of 1 pmol NADPH. Incubations were carried out for 10 min at 37°C and the reaction stopped by the addition of 6 ml dichloromethane with vigorous mixing. Blanks to which the substrate was added after stopping the reaction were carried through the same procedure. In order to separate the phases, the samples were cooled on ice and centrifuged at 10 000 X g for 5 min. The upper layer (aqueous phase) was discarded by aspiration and the organic phase evapor- ated under a stream of nitrogen. The residue was resuspended in 100 pl n-hexane : 2-propanol (91 : 9; v/v) and a 10 p1 aliquot analysed by HPLC.
High-performance liquid chromatography separation and quantitation was per- formed as described previously (Mancilla & Gil, 1984) on a prepacked stainless steel Lichrosorb Diol column (250 mm length, 4.6 mm internal diameter, 5 pm particle size, purchased from H. Knauer, FRG), using a solvent mixture of

342 L. Gil et al.
n-hexane : 2-propanol (91 : 9; v/v) at a flow rate of 1.0 ml/min. Column effluents were monitored at 254 nm, at 2 2 T , room temperature. Metabolites were generally quantitated by comparison of their peak heights with those of authentic standards. When [4-I4C] testosterone was used as substrate, 0.2-0.4 ml fractions were col- lected and radioactive metabolites were quantitated by scintillation counting. All analyses were performed with a Waters Model 6000 liquid chromatograph equipped with a second Waters Model 6000-A pump, a U6K injector, a model 440 high sensitive detector and a dual Omniscribe recorder (Houston Instruments, U.S.A.).
All data were analysed by Student’s t-test for differences between the means (Steel & Torrie, 1980).
Chemicals Testosterone and [4-14C] testosterone (specific activity 56.9 mCi/mmol) were obtained from Sigma (St Louis, MO, U.S.A.) and the Radiochemical Centre (Amersham, U.K.) , respectively; androstenedione, 68- and 16a-hydroxytestos- terone were purchased from Steraloids (Wilton, NH, U.S.A.); 7a-hydroxy- testosterone, 2cc-hydroxytestosterone, 2(3-hydroxytestosterone and 68-hydroxy- androstenedione were kindly supplied by Professor D. N. Kirk, from the Steroid Reference Collection, MRC, London, U.K. All biochemicals were obtained from Sigma and HPLC organic solvents from E. Merck, Darmstadt, and Mallinckrodt, Inc., Kentucky, U.S.A.
Results The experimental condition chosen for this study is one in which protein-energy malnutrition is provoked in the immature male rat. After weaning, the malnour- ished group was fed for 14 days with a protein-free diet. This diet contained 3.75 kcal/g, while the average intake for a 35-day-old malnourished rat was 0.11 g/g rat/day, thus providing only 0.4 kcal/g rat/day, whereas the intake for the normal group was 0.7 kcalig rat/day.
Body and liver weight, microsomal protein and cytochrome P-450 content Table 1 presents the changes in body and liver weights, microsomal protein content and in cytochrome P-450 content in normal and malnourished male rats. At 35 days of age the body weight in the malnourished group was 22% and liver weight 18% of that in normal rats. Liver microsomal protein had decreased by 29%. The content of hepatic cytochrome P-450 was diminished by 38% and the maximal wavelength for the carbon monoxide reduced cytochrome P-450 was shifted from 450 nm to 452 nm.
Nutrition-induced alteration in liver microsomal testosterone mono-oxygenation The HPLC profile for testosterone and its oxidation products using microsomes from normal and malnourished rats is shown in Fig. 1. In Fig. l a the correlation between peak absorbance at 254 nm and retention time in microsomes from normal-fed rats is illustrated. The following metabolites were identified by their
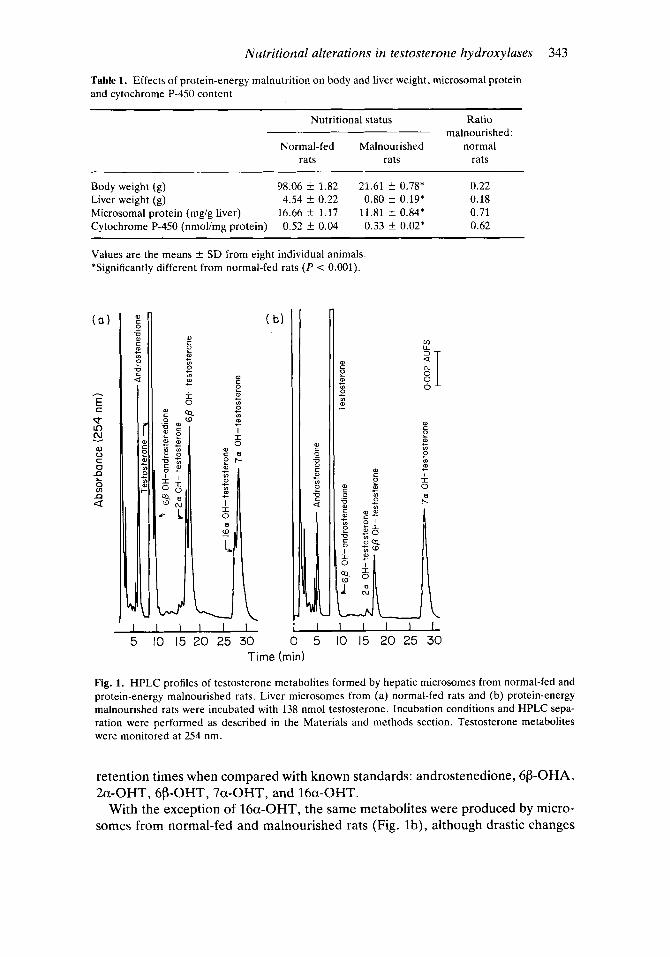
Nutritional alterations in testosterone hydroxylases 343
Table 1. Effects of protein-energy malnutrition on body and liver weight, microsomal protein and cytochrome P-450 content
Nutritional status Ratio
Normal-fed Malnourished normal malnourished:
rats rats rats
Body weight (8) 98.06 ?i 1.82 21.61 f 0.78* 0.22 Liver weight (8) 4.54 f 0.22 0.80 f 0.19* 0.18 Microsomal protein (mg/g liver) 16.66 f 1.17 11.81 k 0.84* 0.71 Cytochrome P-450 (nmolimg protein) 0.52 f 0.04 0.33 2 0.02* 0.62
Values are the means f SD from eight individual animals *Significantly different from normal-fed rats ( P < 0.001).
01 c
v c 0 c y1
v
0
e
m 0, + m
+ m m
I I 0
c 0
e + Lo
m 0) t
t
I
- Time (mid
Fig. 1. HPLC profiles of testosterone metabolites formed by hepatic microsomes from normal-fed and protein-energy malnourished rats. Liver microsomes from (a) normal-fed rats and (b) protein-energy malnourished rats were incubated with 138 nmol testosterone. Incubation conditions and HPLC sepa- ration were performed as described in the Materials and methods section. Testosterone metabolites were monitored at 254 nm.
retention times when compared with known standards: androstenedione, 6P-OHA, 2a-OHT, 6P-OHT7 7a-OHT7 and 16a-OHT.
With the exception of 16a-OHT, the same metabolites were produced by micro- somes from normal-fed and malnourished rats (Fig. lb), although drastic changes
344 L. Gil et al.
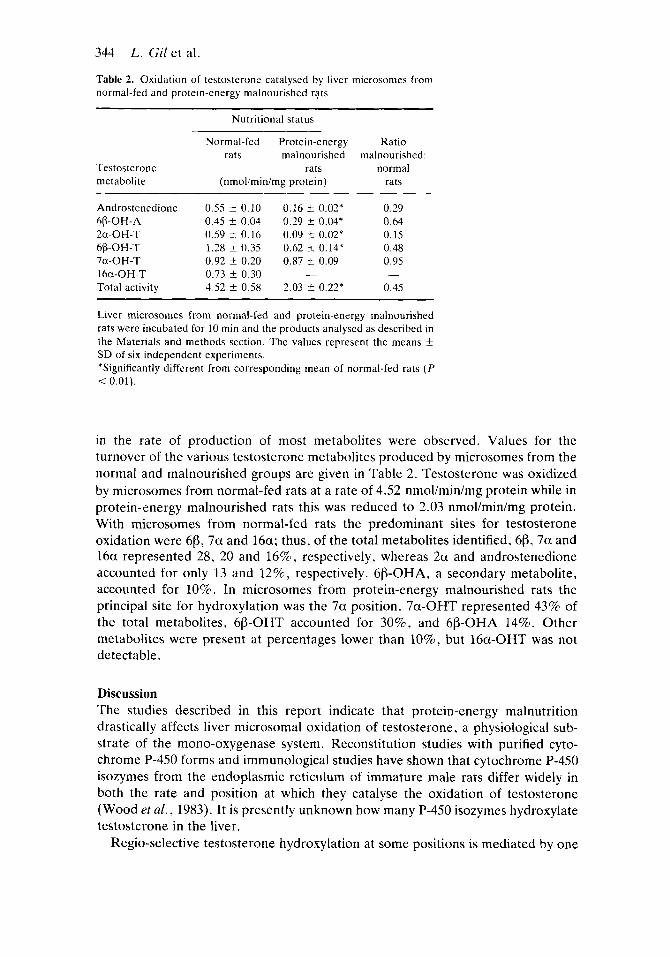
Table 2. Oxidation of testosterone catalysed by liver microsomes from normal-fed and protein-energy malnourished rats
~~~~
Nutritional status
Testosterone metabolite
Normal-fed Protein-energy rats malnourished
rats (nmoliminimg protein)
Ratio malnourished:
normal rats
Androstenedione
2a-OH-T
7a-OH-T 16a-OH-T Total activity
68-OH-A
66-OH-T
0.55 -t 0.10 0.16 f 0.02* 0.45 k 0.04 0.29 5 0.04* 0.59 f 0.16 0.09 f 0 . 0 2 * 1.28 i 0.35 0.62 k 0.14* 0.92 f 0.20 0.87 t 0.09 0.73 f 0.30 - 4.52 f 0.58 2.03 5 0.22*
0.29 0.64 0.15 0.48 0.95
0.45 -
Liver microsomes from normal-fed and protein-energy malnourished rats were incubated for 10 min and the products analysed as described in the Materials and methods section. The values represent the means -t SD of six independent experiments. *Significantly different from corresponding mean of normal-fed rats ( P < 0.01).
in the rate of production of most metabolites were observed. Values for the turnover of the various testosterone metabolites produced by microsomes from the normal and malnourished groups are given in Table 2. Testosterone was oxidized by microsomes from normal-fed rats at a rate of 4.52 nmol/min/mg protein while in protein-energy malnourished rats this was reduced to 2.03 nmol/min/mg protein. With microsomes from normal-fed rats the predominant sites for testosterone oxidation were 613, 7a and 16a; thus, of the total metabolites identified, 68, 7a and 16a represented 28, 20 and 16%, respectively, whereas 2a and androstenedione accounted for only 13 and 12%, respectively. 6p-OHA, a secondary metabolite, accounted for 10%. In microsomes from protein-energy malnourished rats the principal site for hydroxylation was the 7a position. 7a-OHT represented 43% of the total metabolites, 6p-OHT accounted for 30%, and 613-OHA 14%. Other metabolites were present at percentages lower than lo%, but 16a-OHT was not detectable.
Discussion The studies described in this report indicate that protein-energy malnutrition drastically affects liver microsomal oxidation of testosterone, a physiological sub- strate of the mono-oxygenase system. Reconstitution studies with purified cyto- chrome P-450 forms and immunological studies have shown that cytochrome P-450 isozymes from the endoplasmic reticiilum of immature male rats differ widely in both the rate and position at which they catalyse the oxidation of testosterone (Wood et al . , 1983). It is presently unknown how many P-450 isozymes hydroxylate testosterone in the liver.
Regio-selective testosterone hydroxylation at some positions is mediated by one

Nutritional alterations in testosterone hydroxylases 345
P-450 isozyme, and at other positions is catalysed by several P-450 isozymes. Only one P-450 isozyme is known to hydroxylate testosterone at position 2. This isozyme has been purified in three different laboratories and has been termed P-450h, P-450 PB2c and P-450 RLM5 (Waxman, KO & Walsh, 1983; Cheng & Schenkman, 1983; Ryan et al., 1984). Similarly, hydroxylation at position 7a is catalysed only by P-450a (alternative designations P-450 UT-F and P-450 PB3) (Wood et al., 1983). Conversely, hydroxylation at position 60 is mediated by cytochrome P-450c, P-450d and P-45Og (Wood et a f . , 1983; Ryan et al., 1984), and by P-450 RLM3 (Cheng & Schenkman, 1983), which is identical to P-45Og. Furthermore, at least four P-450 isozymes function as 16a hydroxylators of testosterone: P-450b, P-450e, P-450f and P-450h (Wood et a[., 1983; Ryan et al., 1984). Other positions such as 168 and 170 are also hydroxylated by several isozymes to generate androstenedione. Therefore, total liver microsomal hydroxylation of testosterone is a combination of the activi- ties of many P-450 isozymes. This is confirmed by studies showing that factors involved in the expression of different P-450 isozymes, such as age, sex, hormonal imprinting and xenobiotic inducibility, affect the catalytic activities of several hormone hydroxylases, reflecting the contributions of several isozymes to the overall microsomal metabolism (Waxman et al., 1985).
Our results suggest that protein-energy malnutrition modulates the expression of several testosterone hydroxylases. Complete suppression of 16a-hydroxylase activ- ity and a significant reduction of 2a- (85%) and 68- (52%) testosterone hydroxy- lases were observed in protein-energy malnourished rats. In addition, 60-OHA was diminished by 36%. The activity of 2a- and 16a-testosterone hydroxylase has been identified mainly with P-450 PB2c (P-450h = P-450 RLMS), whereas 6P-androste- nedione hydroxylase is associated mainly with P-450 2a/PCN-E (Waxman et al., 1985). These results enable us to conclude tentatively that the levels of these hormonally regulated P-450 isozymes (Waxman et al., 1985) are decreased drasti- cally as a result of protein malnourishment.
It should be noted that the dramatic loss of activity of 2a-, 16a- and 60- hydroxylases was obtained by protein malnourishment over a time period when these activities are responsive to changing developmental and hormonal influences (i.e. 3-5 weeks of age). Furthermore, the 170-hydroxylase activity, which yields androstenedione, was decreased by 71%. This activity has also been associated with P-450 2c and P-450b (PB-4 = PB-B), suggesting that the latter isozyme might also be regulated nutritionally. In contrast to other isozymes, 7a-hydroxylase was apparently unaffected by the change in nutrition. The total testosterone oxidative activity was decreased by 55% in malnourished rats, but cytochrome P-450 was diminished by only 38%, suggesting that P-450 isozymes related to testosterone oxidation are affected more by malnutrition than are other P-450 forms not involved in this process.
SDS-PAGE of microsomes from protein-energy malnourished rats showed a decline in band intensity of several proteins in the region 46 000-60 000 daltons, compared with microsomes from normal-fed rats (results not shown). Thus, a deficit of protein in the diet for a long period of time seems to elicit drastic changes in cytochrome P-450 isozyme patterns, which might explain the 2-nm shift in the different CO-reduced cytochrome P-450 spectra of microsomes from malnourished rats.

346 L. Gil et al.
It is possible that under these extreme nutritional conditions some (but not all) isozymes related to testosterone oxidation are either not synthesized or are synthe- sized at a lower rate. Another possibility is that the rate of degradation of these forms is enhanced.
In the malnourished rat, 43% of testosterone oxidative activity is catalysed by 7a-hydroxylase, which has been related to P-450a. P-450a is indistinguishable catalytically and electrophoretically from P-450 PB3 (Waxman et al., 1983) and from P-450 UT-F (Guengerich et al., 1982). The spectra of P-450a showed a CO- reduced peak at 452 nm (Ryan et al., 1979). Similarly, microsomes from protein- energy malnourished rats also showed a peak at 452 nm in the CO-difference spectra, suggesting that P-450a might be one of the predominant isozymes present in this preparation. Immunological studies are in progress in our laboratory to explore this possibility. Other isozymes, cytochrome P-450j, induced by isoniazide (Ryan et al., 1985) and P-452, induced by clofibrate (Gibson, Orton & Tamburini, 1982; Tamburini et al., 1984), also present a Soret maximum at 452 nm in CO spectra, but these forms showed almost undetectable testosterone hydroxylase activity. P-450a has shown very high rates of turnover for the hydroxylation of testosterone and androstenedione in the 7a position, and exceedingly poor mono- oxygenase catalytic activity with exogenous substrates (Ryan et al., 1979; Guenger- ich et al., 1982; Wood et al., 1983; Waxman et al., 1983). This isozyme of narrow regio-selectivity to hydroxylate steroids is also not subject to significant induction by a variety of xenobiotics (Guengerich et al., 1982; Dannan et al. , 1983). However, two- to five-fold induction of this protein has been reported (Thomas et al., 1981; Parkinson et al., 1983).
The fact that 7a-testosterone hydroxylase is not diminished by malnutrition might indicate that this isozyme might play a key physiological role, and that 7a- hydroxytestosterone, the product of its catalytic activity, might be an important metabolic intermediate. This is supported by the observation that 7a-hydroxytes- tosterone inhibited the transformation in-vitro of pregnenolone to progesterone in a rat testicular microsomal fraction, indicating a possible role in intracellular regulation of testosterone production (Inano & Tamaoki, 1971).
The activity of testosterone 7a-hydroxylase is known to be regulated develop- mentally (Jacobson & Kuntzman, 1969) and it is unaffected by sex and hormonal status (Waxman et al. , 1985). Our results clearly establish that this hydroxylase is not modulated by the nutritional status. The present results suggest that protein- energy malnutrition can be an interesting tool with which to modify the P-450 isozymic profile, but with the main advantage over other methods (i.e. chemical induction) that the microsomal fraction contains predominantly P-450 isozymes probably involved in key mono-oxygenation of physiological substrates. In a sub- sequent report we will demonstrate that feeding malnourished rats with a high protein diet is able to return the activity of some, but not all, testosterone hydroxylases to normal.
Acknowledgments We thank Dr Ramiro Martinez from the Pan American Health Organization and

Nutritional alterations in testosterone hydroxylases 347
Dr Catherine Connelly for their assistance, also Dr N. Kirk for providing sterol standards.
This study was supported by grants from the University of Chile (B 1970), Fondecyt and the Organization of American States.
References Cheng, K. C . & Schenkman, J . B. (1983) Testosterone metabolism by cytochrome P-450 isozymes
RLM3 and RLM5 and by microsomes. Journal of Biological Chemistry, 258, 11738-11744. Dannan, G . A., Guengerich, F. P., Kaminsky, L. S . & Aust, S. D. (1983) Regulation of cytochrome
P-450. Immunochemical quantitation of eight isozymes in liver microsomes of rats treated with polybrominated biphenyl congeners. Journal of Biological Chemistry, 258, 1282-1288.
Gibson, G . G . , Orton, T. C. & Tamburini, P. P. (1982) Cytochrome P-450 induction by clofibrate. Purification and properties of a hepatic cytochrome P-450 relatively specific for the 12- and 11- hydroxylation of dodecanoic acid (lauric acid). Biochemical Journal, 203, 161-168.
Gil, L., Vasquez, H., Orellana, M., Selkirk, J . , Wold, F. & Strobel, H . (1988) Purification and characterization of liver cytochrome P-446 isolated from protein-energy malnourished rats. Molecular and Cell Biochemistry, in press.
Guengerich, F. P., Dannan, G. A , , Wright, S. T., Martin, M. V. & Kaminsky, L. S. (1982) Purification and characterization of liver microsomal cytocromes P-450. Electrophoretic, spectral catalytic and immunochemical properties and inducibility of eight isozymes isolated from rats treated with phenebarbital or p-naftoflavone. Biochemistry, 21, 6019-6030.
Gustafsson, J . A. (1978) Steroid hydroxylations catalized by cytochrome P-450. In: Methods in Enzymo- logy 52 (eds S. Fleischer and L. Packer), pp. 377-388. Academic Press, New York.
Hasumura, S . , Endou, H., Kakuno, K. & Sakai, F. (1983) Effects of starvation on microsomal cytochrome P-450 and laurate-omega-hydroxylation of rat kidney and liver. Japanese Journal of Pharmacology, 33,999-1006.
Hietanen, E. (1980) Modification of hepatic drug metabolizing enzyme activities and their induction by dietary protein. General Pharmacology, 11, 443-450.
van der Hoeven, Th. (1981) Testosterone oxidation by rat liver microsomes: effects of phenobarbital pretreatment and the detection of seven metabolites by HPLC. Biochemical and Biophysical Research Communications, 100, 1285-1291.
Inano, H. & Tamaoki, B. (1971) Regulation of testosterone biosynthesis in rat testes by 7a-hydroxy- lated C 19-steroids. Biochimica et Biophysica Acta, 239, 482493.
Jacobson, M. & Kuntzman, R. (1969) Studies on testosterone hydroxylation and phenobarbital oxi- dation by rat liver microsomes: selective effects of age, sex, castration and testosterone propio- nate treatment. Steroids, 15, 327-341.
Kato, R. & Kamataki, T. (1982) Cytochrome P-450 as a determinant of sex difference of drug metabolism in the rat. Xenobiotica, 12, 787-800.
Lowry, 0. H., Rosebrough, N. J . , Farr, A. L. & Randall, R. J . (1951) Protein measurements with folin phenol reagent. Journal of Biological Chemistry, 193,265-275.
Lu, A. Y . H. & West, S. B. (1980) Multiplicity of mammalian microsomal cytochrome P-450. Pharmacological Reviews, 31,277-295.
Mancilla, J. & Gil, L. (1984) A simple, reliable and rapid HPLC method to separate and quantify androstenedione and hydroxy testosterone. Analytical Letters, 17(B9), 873-886.
Nakamura, Y. & Ueda, S. (1980) Induction of testosterone 16a-hydroxylase in rat liver microsomes by phenobarbital pretreatment. Biochemical and Biophysical Research Communications, 93, 10141019.
Omura, T. & Sato, R. (1964) The carbon monoxide binding pigment of liver microsomes solubilization, purification and properties. Journal of Biological Chemistry, 239, 2379-2385.
Parkinson, A . , Safe, S. H., Robertson, L. W., Thomas, P. E., Ryan, D . E., Reik, L. M. & Levin, W. (1983) Immunochemical quantitation of cytochrome P-450 isozymes and epoxide hydrolase in liver microsomes from polychlorinated or polybrominated biphenyl-treated rats. A study of structure-activity relationships. Journal of Biological Chemistry, 258, 5967-5976.
Ryan, D. E., Iida, S. , Wood, A. W., Thomas, P. E., Lieber, C. & Levin, W. (1984) Characterization of three highly purified cytochrorne P-450 from hepatic microsomes of adult male rats. Journal of Biological Chemistry, 259, 1239-1250.

348 L. Gil et al.
Ryan, D. E., Ramanathan, L., Iida, S. , Thomas, P. E., Haniu, M., Shively, J . , Lieber, C. S. & Levin, W. (1985) Characterization of a major form of rat hepatic microsomal cytochrome P-450 induced by isoniazid. Journal of Biological Chemistry, 260, 6385-6393.
Ryan, D. E., Thomas, P. E., Korzeniowski, D. & Levin, W. (1979) Separation and characterization of highly purified forms of liver microsomal cytochrome P-450 from rats treated with polychlori- nated biphenyls, phenobarbital, and 3-methylcholanthrene. Journal of Biological Chemistry, 254,
Salazar, I . , Litvak, S. & Gil, L. (1983) Modulation of rat liver aryl hydrocarbon benzo(a)pyrene hydroxylase activity by nutritional effects. Journal of Toxicology and Environmental Health, 11,
Sheets, J . J . & Estabrook, R. (1985) Multiple sites of steroids hydroxylation by the liver microsomal cytochrome P-450 system: primary and secondary metabolism of androstenedione. Biochemistry, 24, 6591-6597.
Sheets, J . J . , Mason, I., Wise, C. A. & Estabrook, R. W. (1986) Inhibition of rat liver microsomal cytochrome P-450 steroid hydroxylase reactions by imidazole antimycotic agents. Biochemical Pharmacology, 35, 487-491.
Shiverick, K. L. & Neims, A. H. (1979) Multiplicity of testosterone hydroxylases in a reconstituted hepatic cytochrome P-450 system from uninduced male rats. Drug Metabolism and Disposition, 7 , 29CL295.
Steel, R. G . D. & Torrie, J . H. (eds) (1980) Comparisons involving two sample means. In: Principles and Procedures of Statistics. A Biometrical Approach, pp. 86100. McGraw Hill, New York.
Tamburini, P. P., Masson, H. A, , Bains, S. K., Makowski, R. J. , Morris, B. & Gibson, G. G. (1984) Multiple forms of hepatic cytochrome P-450. Purification, characterization and comparison of a novel clofibrate-induced isozyme with other major forms of cytochrome P-450. European Journal of Biochemistry, 159, 235-246.
Thomas, P. E., Deik, L. M., Ryan, D. E. & Levin, W. (1981) Regulation of three forms of cytochrome P-450 and epoxide hydrolase in rat liver microsomes. Journal of Biological Chemistry, 256, 1044-1 052.
Waxman, D. J. (1984) Rat hepatic P-450 isoenzyme 2c. Identification as a male-specific developmentally steroid 16a-hydroxylase and comparison to a female-specific cytochrome P-450 isoenzyme. Journal of Biological Chemistry, 259, 15481-15490.
Waxman, D. J . , Dannan, G. A. & Guengerich, F. P. (1985) Regulation of rat hepatic cytochrome P-450: age-dependent expression, hormonal imprinting, and xenobiotic inducibility of sex- specific isoenzymes. Biochemistry, 24, 440S4417.
Waxman, D. J. , KO, A. & Walsh, C. (1983) Regioselectivity and stereoselectivity of androgen hydroxylations catalyzed by cytochrome P-450 isozymes purified from phenobarbital-induced rat liver. Journal of Biological Chemistry, 258, 11937-11947.
White, R . E. & Coon, M. J . (1980) Oxygen activation by cytochrome P-450. Annual Review of Biochemistry, 49, 315-356.
Wiebel, F. J . & Gelboin, H. V. (1975) Aryl hydrocarbon (Benzo(a)pyrene) hydroxylases in liver from rats of different age, sex and nutritional status. Biochemical Pharmacology, 24, 151 1-1515.
Wood, A. W., Ryan, D. E . , Thomas, P. E . & Levin, W . (1983) Regio and stereoselective metabolism of two C 19 steroids by 5 highly purified and reconstituted rat hepatic cytochrome P-450 isoenzymes. Journal of Biological Chemistry, 258, 8839-8847.
1365-1374.
5 19-533,
Received 26 June 1987; accepted 26 October 1987
Copyright © 2022 FDOKUMEN




















