
Involvement ofaPlasmid inEscherichia coli Envelope Alterations
6
1983, 155(1):402. J. Bacteriol. Ghittoni and C Vásquez S B Rosas, A Calzolari, J L La Torre, N E alterations. Escherichia coli envelope Involvement of a plasmid in http://jb.asm.org/content/155/1/402 found at: Updated information and services can be These include: CONTENT ALERTS more» alerts (when new articles cite this article), Receive: RSS Feeds, eTOCs, free email http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on June 19, 2014 by guest http://jb.asm.org/ Downloaded from on June 19, 2014 by guest http://jb.asm.org/ Downloaded from
-
Upload
escrituracientifica -
Category
Documents
-
view
0 -
download
0
Transcript of Involvement ofaPlasmid inEscherichia coli Envelope Alterations

1983, 155(1):402. J. Bacteriol.
Ghittoni and C VásquezS B Rosas, A Calzolari, J L La Torre, N E alterations.Escherichia coli envelope Involvement of a plasmid in
http://jb.asm.org/content/155/1/402found at: Updated information and services can be
These include:
CONTENT ALERTS
more»alerts (when new articles cite this article), Receive: RSS Feeds, eTOCs, free email
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders:
http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
on June 19, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

Vol. 155, No. 1JOURNAL OF BACTERIOLOGY, July 1983, p. 402-4060021-9193/83/070402-05$02.00/0Copyright © 1983, American Society for Microbiology
Involvement of a Plasmid in Escherichia coli EnvelopeAlterations
SUSANA B. ROSAS,l* ALDO CALZOLARI,2 JOSE L. LA TORRE,2 NORA E. GHITTONI,1 ANDCESAR VASQUEZ2t
Departamento de Biologia Molecular, Universidad Nacional de Rio Cuarto, 5800 Rio Cuarto, C6rdoba,' andCentro de Virologia Animal, 1414 Buenos Aires,2 Argentina
Received 1 December 1982/Accepted 13 April 1983
A plasmid-containing wild-type Escherichia coli strain was treated with twoplasmid-curing agents, sodium dodecyl sulfate and ethidium bromide. Plasmidelimination was accompanied by drastic changes in the morphology of thecolonies. Analysis of the cured strain by scanning and transmission electronmicroscopy showed important alterations in size and morphology of the cells.Metabolic differences were also found between the wild-type and cured cells.
Morphogenesis and cell division in Escherich-ia coli are highly regulated events controlled bymany genes located around the bacterial chro-mosome (4). Understanding of these events hasbeen possible through studies of mutants carry-ing defects in different biosynthetic steps as wellas the assembly of cell envelope components.Some of these mutations produce alterations incolony morphology without affecting the struc-ture of individual cells (15). However, othermutations cause a variety of changes in cellmorphology such as the production of giant,rounded, or filamentous bacteria (1-3, 11). In afew cases, the metabolism of lipopolysaccha-rides and other components of the bacterial cellwall has been shown to be controlled by plas-mids (10, 12). Recently, Kopecko et al. (14, 18,19) reported that a plasmid controls the synthe-sis of the Shigella sonnei form I 0-side chains.Antigenic variation in S. sonnei due to plasmidloss was also associated with changes in themorphology of the colonies (14, 18).
In this paper, we report that plasmic curing inE. coli strain RC424 results in alterations incolony morphology and cell structure as well asin physiological behavior.
E. coli RC424 is a wild-type prototroph con-taining plasmid p424, which confers high-levelresistance to chloramphenicol and tetracycline.E. coli K-12 HB101 (pro lacY leu thi rpsL recA)has been described elsewhere (7). Plasmids ofknown molecular weight were obtained from E.Lederberg (Plasmid Reference Center, Stanford,Calif.). Bacteria were routinely grown in Luriabroth (LB) (17) with rotary shaking. When re-quired, chloramphenicol or tetracycline wasadded to a final concentration of 50 pug/ml. E.
t Deceased 19 April 1983. Dedicated to his memory.
coli RC424 and derivate cells were also grown inminimal medium containing, in grams per liter:KH2PO4, 3; Na2HPO4, 6; NH4Cl, 1; MgSO4,0.2; and glucose, 4 (pH 6.8).
Plasmid was eliminated from bacterium bytwo procedures. (i) A small inoculum was grownovernight at 37°C in LB containing a subinhibi-tory concentration of ethidium bromide (from 1,uM to 2.5 mM) (6). Cultures containing thehighest concentration of ethidium bromide inwhich growth was clearly visible (0.52 mM)were diluted and spread onto LB plates. Result-ing colonies were tested for drug sensitivity. (ii)An inoculum of E. coli RC424 was added to LBcontaining 10% sodium dodecyl sulfate (SDS)and incubated overnight at 37°C (20). The cul-ture was diluted, plated, and tested as above. Inselected cases, a verification of the relationshipbetween the loss of a resistance marker and theabsence of plasmid DNA was made by agarosegel electrophoresis (see below).
Plasmid DNA was prepared by the method ofBirnboim and Doly (5) with minor modifications.An overnight culture was centrifuged, and thebacterial pellet was washed with 40 mM Tris-hydrochloride, pH 8.0, and recentrifuged. Pel-lets were incubated with the lysis solution for 30min at 0°C. After the denaturation and renatur-ation steps, the lysate was incubated for 30 minat 0°C and centrifuged. The supernatant wastreated with RNase A at 20 ,ug/ml. DNA wasethanol precipitated, dried under vacuum, andresuspended in 10 mM Tris-hydrochloride-1mM EDTA, pH 8.0. Electrophoresis was carriedout in horizontal 0.7% agarose gels in 89 mMTris base-89 mM boric acid-1.5 mM Na2 EDTA(pH 8.1). The gels were stained with ethidiumbromide, soaked with distilled water, and photo-graphed under UV light (16). Plasmid DNA for
402
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 155, 1983
transformation was purified by CsCl-ethidiumbromide isopycnic gradient centrifugation as de-scribed previously (8). Cells were made compe-
tent and transformed by the procedure of pro-
longed incubation in CaCl2 of Dagert and Erlich(9).For microscope examination, cells were pre-
cipitated on 0.4-jim Nuclepore filters (Nucle-pore Corp., Pleasanton, Calif.) (21) or glasssquares of 0.8 mm, fixed wih glutaraldehyde,dehydrated, and rinsed. After rinsing, cells werecritical point dried and coated with gold-palladi-um. The coated cells were examined with a J5M-U3 JEOL scanning electron microscope. Cellswere also fixed in glutaraldehyde, postfixed inOS04, dehydrated, and embedded in Epon. Af-ter sectioning, the samples were mounted ongrids, stained, and observed with a JEM-1OOCJEOL transmission electron microscope.RC424 is a smooth-colony-forming strain of E.
coli which is able to grow in minimal medium.Strain characterization involved analysis of sug-
ar fermentation and gas production, degradationof organic acids and amino acids, and mobility(APY System). In addition, resistance to thefollowing antibiotics was tested: amikacin, am-picillin, cephalosporin, chloramphenicol, colis-tin, gentamicin, kanamycin, nalidixic acid,phosphonomycin, rifampin, tetracycline,tobramycin, and trimethroprim. This strainshowed a high level of resistance to chloram-phenicol (>1,000 ,ug/ml) and tetracycline (>500,ug/ml). Both resistances correlated with thepresence in E. coli RC424 of a single plasmid of45 + 5 megadaltons (Fig. 1). E. coli HB101 wasrendered resistant to both antibiotics whentransformed with the p424 DNA (data notshown).The effect of two plasmid-curing agents on the
stability of the p424 DNA in E. coli RC424 wastested. Bacteria were grown overnight at 37°C inLB with 10% SDS or 0.52 mM ethidium bromideand plated onto LB plates. More than 200 colo-nies were obtained after the first treatment, andmore than 100 colonies were obtained after thelater one. From each group, four randomlyselected representative colonies were subjectedto the following studies.Two main morphological changes of the cured
bacteria were observed: (i) an increase in thesize of the colonies, which were approximatelyfour times larger than those of the wild-typeafter 24 h of growth; (ii) cured bacteria formedrough-type colonies instead of the smooth ones
formed by the parent strain. An additional prop-erty was that, after 9 days of growth on agarpetri dishes, a single colony from the curedbacteria was able to cover a surface of about 60-cm diameter, whereas a wild-type colony onlycovered a 12-cm-diameter surface. Regardless of
B
FIG. 1. Agarose gel electrophoresis of plasmidDNA from wild-type RC424 (A), cured RC-7 (B), andtransformed RC-7(p424) (C) strains of E. coli. Themolecular mass standards were 112 and 34 megadal-tons for R27 and RP4, respectively. Chr, Chromo-somal DNA.
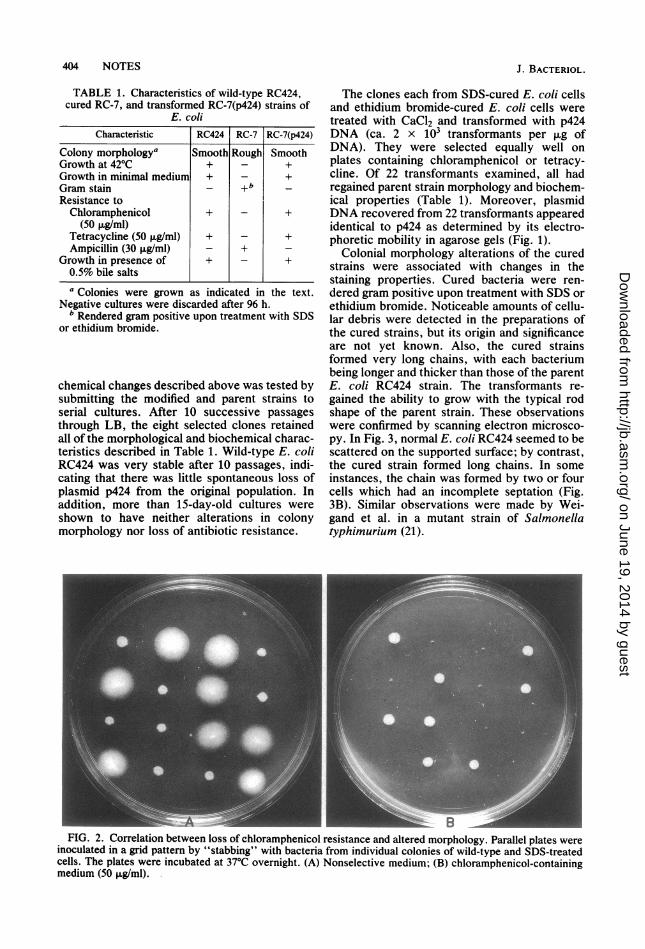
the morphological differences, the cured bacte-ria conserved most of the original properties ofthe parent strain: lysine decarboxylase, nega-tive; ornithine decarboxylase, negative; argininedehydrogenase, positive; gas from glucose, posi-tive; and motility in semisolid medium, negative.However, some changes were observed (Table1) such as the acquired resistance to ampicillin(>50 jiglml) and the inability to grow in minimalmedium and in the presence of detergents. Someof these properties are found to be present inmutants with alterations in the cell envelope (13)and could be an indication that the cured bacte-ria are carrying some modifications in the cellsurface components. There was a correlationbetween the loss of resistance to antibiotics andthe altered morphology of cured bacteria (Fig.2). Moreover, it was not possible to recoverplasmids from bacteria of either the four ran-domly selected SDS-cured colonies or the fourrandomly selected ethidium bromide-cured colo-nies. Figure 1 shows the agarose gel electropho-resis from one cured colony (RC-7).The stability of the morphological and bio-
NOTES 403
A C
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
404 NOTES
TABLE 1. Characteristics of wild-type RC424,cured RC-7, and transformed RC-7(p424) strains of
E. coliCharacteristic RC424 RC-7 RC-7(p424)
Colony morphologya Smooth Rough SmoothGrowth at 42°C + _ +Growth in minimal medium + - +Gram stain +bResistance toChloramphenicol + - +
(50 ,ug/ml)Tetracycline (50 ,g/ml) + - +Ampicillin (30 ,ug/ml) _ +
Growth in presence of + - +0.5% bile saltsa Colonies were grown as indicated in the text.
Negative cultures were discarded after 96 h.b Rendered gram positive upon treatment with SDS
or ethidium bromide.
chemical changes described above was tested bysubmitting the modified and parent strains toserial cultures. After 10 successive passagesthrough LB, the eight selected clones retainedall of the morphological and biochemical charac-teristics described in Table 1. Wild-type E. coliRC424 was very stable after 10 passages, indi-cating that there was little spontaneous loss ofplasmid p424 from the original population. Inaddition, more than 15-day-old cultures wereshown to have neither alterations in colonymorphology nor loss of antibiotic resistance.
The clones each from SDS-cured E. coli cellsand ethidium bromide-cured E. coli cells weretreated with CaCl2 and transformed with p424DNA (ca. 2 x 103 transformants per ,ug ofDNA). They were selected equally well onplates containing chloramphenicol or tetracy-cline. Of 22 transformants examined, all hadregained parent strain morphology and biochem-ical properties (Table 1). Moreover, plasmidDNA recovered from 22 transformants appearedidentical to p424 as determined by its electro-phoretic mobility in agarose gels (Fig. 1).
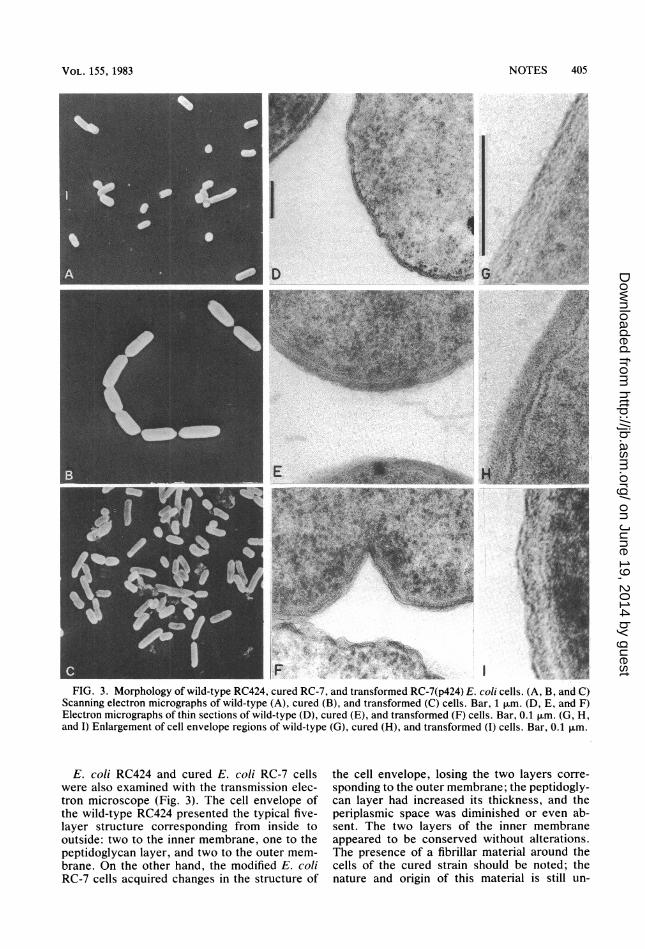
Colonial morphology alterations of the curedstrains were associated with changes in thestaining properties. Cured bacteria were ren-dered gram positive upon treatment with SDS orethidium bromide. Noticeable amounts of cellu-lar debris were detected in the preparations ofthe cured strains, but its origin and significanceare not yet known. Also, the cured strainsformed very long chains, with each bacteriumbeing longer and thicker than those of the parentE. coli RC424 strain. The transformants re-gained the ability to grow with the typical rodshape of the parent strain. These observationswere confirmed by scanning electron microsco-py. In Fig. 3, normal E. coli RC424 seemed to bescattered on the supported surface; by contrast,the cured strain formed long chains. In someinstances, the chain was formed by two or fourcells which had an incomplete septation (Fig.3B). Similar observations were made by Wei-gand et al. in a mutant strain of Salmonellatyphimurium (21).
_ : . A -. _ B i nFIG. 2. Correlation between loss of chloramphenicol resistance and altered morphology. Parallel plates were
inoculated in a grid pattern by "stabbing" with bacteria from individual colonies of wild-type and SDS-treatedcells. The plates were incubated at 37°C overnight. (A) Nonselective medium; (B) chloramphenicol-containingmedium (50 ,ug/ml).
J. BACTERIOL.
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from
NOTES 405
D
f. . .".
5S F5
.,;r. s-
, f } ^ -;5gr<W
}+, .u./' A; g
i.,.,s''..'S,.#gr J'
r_, ': >> S;-1b
E
l i ! 5- _*>>; ..
___ '' t' < +~~~~,fI~/iV
'_F * IX '.4X g xs t iFIG. 3. Morphology of wild-type RC424, cured RC-7, and transformed RC-7(p424) E. coli cells. (A. B, and CQ
Scanning electron micrographs of wild-type (A), cured (B), and transformed (C) cells. Bar, 1 ,um. (D, E, and F)Electron micrographs of thin sections of wild-type (D), cured (E), and transformed (F) cells. Bar, 0.1 pm. (G H,and I) Enlargement of cell envelope regions of wild-type (G), cured (H), and transformed (I) cells. Bar, 0.1 ,um.
E. coli RC424 and cured E. coli RC-7 cellswere also examined with the transmission elec-tron microscope (Fig. 3). The cell envelope ofthe wild-type RC424 presented the typical five-layer structure corresponding from inside tooutside: two to the inner membrane, one to thepeptidoglycan layer, and two to the outer mem-brane. On the other hand, the modified E. coliRC-7 cells acquired changes in the structure of
the cell envelope, losing the two layers corre-sponding to the outer membrane; the peptidogly-can layer had increased its thickness, and theperiplasmic space was diminished or even ab-sent. The two layers of the inner membraneappeared to be conserved without alterations.The presence of a fibrillar material around thecells of the cured strain should be noted; thenature and origin of this material is still un-
R-1.1.10.1...V. -1-i '.- "t". - . 4, :f " .., -.Atr, ",": I .6 , --,
VOL. 155, 1983
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from

406 NOTES
known. In contrast, transformed E. coli RC-7(p424) was similar in morphology to the originalRC424 strain.The results presented here show that treat-
ment of E. coli RC424 with either SDS orethidium bromide leads to loss of a plasmid,p424, which encodes resistance to chloramphen-icol and tetracycline among other properties.Plasmid loss was always correlated with bothaltered colony morphology and biochemicalmodifi'cations (Table 1). Stability of curedstrains was high since all biochemical and mor-phological changes observed were maintainedeven after 10 successive passages on LB plates.Under these culture conditions, no spontaneousrevertants to wild-type properties were ob-served. Detailed analysis of parent and curedstrains by both scanning and transmission elec-tron microscopy showed the existence of severalchanges in cellular morphology and variations inthe size of individual cells; alterations in septa-tion were detected as well. Plasmid loss in curedE. coli cells resulted in the disappearance of theouter membrane components and a concomitantchange in the thickness of the peptidoglycanlayer. Biochemical and genetic studies of themetabolism of the different cell envelope compo-nents are required to provide an adequate expla-nation of the intrinsic nature of the observedchanges. Restoration of plasmid p424 to curedcells by transformation rendered the resultantbacteria indistinguishable from the parent strain.It would be worthwhile to study the role thatplasmid p424 might play in the regulation ofstructure and behavior of E. coli cells.
This work was supported by grants from the ConsejoNacional de Investigaciones Cientificas y Tecnicas de laArgentina (CONICET) and the Fundaci6n para la Educaci6n,la Ciencia y la Cultura (FECIC).We thank Angela M. Suburu for excellent assistance in
electron microscopy procedures, N. De Vicenzo, G. Garbino,and J. Homman from the Servicio Nacional de MicroscopiaElectr6nica de Barrido and C. Ricarte for skillful technicalassistance, and Simon Silver for help in the English redactionof the paper.
LITERATURE CITED
1. Adler, H. I., and A. A. Hardigree. 1965. Growth anddivision of filamentous form of Escherichia coli. J. Bacte-riol. 90:223-226.
2. Allison, D. P. 1971. Giant cells of Escherichia coli: amorphological study. J. Bacteriol. 108:1390-1401.
3. Ant6n, D. 1979. Positive selection of mutants with cellenvelope defects of a Salmonella typhimurium strain
hypersensitive to the products of genes hisF and hisH. J.Bacteriol. 137:1271-1281.
4. Bachmann, B. J., and K. B. Low. 1980. Linkage map ofEscherichia coli K-12, edition VI. Microbiol. Rev. 44:1-56.
5. Birnboim, H., and J. Doly. 1979. A rapid alkaline extrac-tion procedure for screening recombinant plasmid DNA.Nucleic Acids Res. 7:1513-1523.
6. Bouanchaud, D., M. R. Scavizzi, and Y. A. Chabbert.1969. Elimination by ethidium bromide of antibiotic resist-ance in enterobacteria and staphylococci. J. Gen. Micro-biol. 54:417-425.
7. Boyer, H. W., and D. Roulland-Dussoix. 1969. A comple-mentation analysis of the restriction and modification ofDNA in Escherichia coli. J. Mol. Biol. 41:459-472.
8. Clewell, D. B., and D. R. Helinski. 1969. Super-coiledcircular DNA-protein complex in Escherichia coli. Purifi-cation and induced conversion to an open circular DNAform. Proc. Natl. Acad. Sci. U.S.A. 62:1159-1166.
9. Dagert, M., and S. D. Ehrlich. 1979. Prolonged incubationin calcium chloride improves the competence of Esche-richia coli cells. Gene 6:23-28.
10. Derylo, M., M. Glowacka, Z. Lorkiewicz, and R. Russa.1975. Plasmid-determined alterations of Salmonella typhi-murium lipopolysaccharides. Mol. Gen. Genet. 140:175-181.
11. Fletcher, G., C. Irwin, J. Henson, C. Fillingim, M. Ma-lone, and J. Walker. 1978. Identification of the Escherich-ia coli cell division gene sep and organization of the celldivision-cell envelope genes in the sep-mur-ftsA-envAcluster as determined with specialized transducing lambdabacteriophages. J. Bacteriol. 133:91-100.
12. Hesslewood, S. R., and J. T. Smith. 1974. Envelope alter-ations produced by R-factors in Proteus mirabilis. J. Gen.Microbiol. 85:146-152.
13. Inouye, M. (ed.). 1979. Bacterial outer membrane, p. 5.Wiley-Interscience Publications, New York.
14. Kopecko, D., 0. Washington, and S. Formal. 1980. Genet-ic and physical evidence for plasmid control of Shigellasonnei form I cell surface antigen. Infect. Immun. 29:207-214.
15. Meadow, P., P. Wells, M. Salkinoja-Salonen, and E.Nurmiahe. 1978. The effect of lipopolysaccharide compo-sition on the ultrastructure of Pseudomonas aeruginosa.J. Gen. Microbiol. 105:23-28.
16. Meyers, J. A., D. Sanchez, L. P. Elwell, and S. Falkow.1976. Simple agarose gel electrophoretic method for theidentification and characterization of plasmid deoxyribo-nucleic acid. J. Bacteriol. 127:1529-1537.
17. Miller, J. H. 1972. Experiments in molecular genetics.Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
18. Sansonetti, P., D. Kopecko, and S. Formal. 1981. Shigellasonnei plasmid: evidence that a large plasmid is necessaryfor virulence. Infect. Immun. 34:75-83.
19. Sansonetti, P., D. Kopecko, and S. Formal. 1982. Involve-ment of a plasmid in the invasive ability of Shigellaflexneri. Infect. Immun. 35:852-860.
20. Tomoeda, M., M. Inuzuka, N. Kubo, and S. Nakamura.1968. Effective elimination of drug resistance and sexfactors in Escherichia coli by sodium dodecyl sulfate. J.Bacteriol. 95:1078-1089.
21. Weigand, R., K. Vinci, and L. Rothfield. 1976. Morpho-genesis of the bacterial division septum: a new class ofseptation-defective mutants. Proc. Natl. Acad. Sci.U.S.A. 73:1882-1886.
J. BACTERIOL.
on June 19, 2014 by guesthttp://jb.asm
.org/D
ownloaded from




















