
Use of sugarcane bagasse as biomaterial for cell immobilization for xylitol production
Upload
independentCategory
view
0download
0
1
This is a Preprint Version for scholarly purpose – appropriate bibliographic citation of this 1
article: SANTOS, D. T. ; SARROUH, B. F. ; RIVALDI, J. D. ; CONVERTI, A. ; SILVA, S. 2
S. . Use of Sugarcane Bagasse as Biomaterial for Cells Immobilization for Xylitol Production. 3
Journal of Food Engineering, v. 86, p. 542-548, 2008. 4
http://dx.doi.org/10.1016/j.jfoodeng.2007.11.004 5
The authors acknowledge Journal of Food Engineering for publisher value-added 6
contributions (formatting, copy editing and technical enhancement). 7
8
Use of Sugarcane Bagasse as Biomaterial for Cell 9
Immobilization for Xylitol Production 10
11
12
aDepartment of Biotechnology, School of Engineering of Lorena, University of São Paulo 13
(USP), Estrada Municipal do Campinho, s/nº, Cx. Postal 116, CEP: 12.602-810, Lorena, SP, 14
Brazil 15
16
bDepartment of Chemical and Process Engineering, Genoa University, Via Opera Pia 15, I-17
16145, Genoa, Italy 18
19
20
21
22
23
* Corresponding author. Tel.: +55 12 3159-5146; fax.: +55 12 3153-3165. 24
E-mail address: [email protected] 25

2
Abstract 1
This study provides a preliminary contribution to the development of a bioprocess for 2
the continuous production of xylitol from hemicellulosic hydrolyzate utilizing Candida 3
guilliermondii cells immobilized onto natural sugarcane bagasse fibers. To this purpose, cells 4
of this yeast were submitted to batch tests of “in situ” adsorption onto crushed and powdered 5
sugarcane bagasse after treatment with 0.5 M NaOH. The results obtained on a xylose-based 6
semi-synthetic medium were evaluated in terms of immobilization efficiency, cell retention 7
and specific growth rates of suspended, immobilized and total cells. The first two parameters 8
were shown to increase along the immobilization process, reached maximum values of 50.5% 9
and 0.31 g immobilized cells/g bagasse after 21 h and then sharply decreased. The specific 10
growth rate of suspended cells continuously increased during the immobilization tests, while 11
that of the immobilized ones, after an initial growth, exhibited decreasing values. Under the 12
conditions selected for cell immobilization, fermentation also took place with promising 13
results. The yields of xylitol and biomass on consumed xylose were 0.65 and 0.18 g/g, 14
respectively, xylitol and biomass productivities 0.66 and 0.13 g L-1
h-1
, and the efficiency of 15
xylose-to-xylitol bioconversion was 70.8 %. 16
17
Key Words: Sugarcane bagasse fibers, biomaterial, cell immobilization, bioconversion, xylitol 18
production. 19
20

3
1. Introduction 1
Xylitol is an alternative high added-value sweetener with anti-cariogenic properties of 2
great concern for both the food industry and the biomedical sector. It is in fact largely utilized 3
in the prevention and treatment of several pathologies, among which atopic dermatitis 4
(Masako, Hideyuki, Shigeyuki, & Zenro, 2005a; Masako et al., 2005b) pseudo-membranose 5
colitis (Naaber, Lehto, Salminen & Mikelsaar, 1996), medium acute otitis (Kontiokari, Uhari, 6
& Koskela, 1998), osteoporosis (Tischler et al., 1996), hemolytic anemia (Wang & Van Eys, 7
1981) and cystic fibrosis (Zabner et al., 2000). 8
Nowadays, xylitol is chemically produced by catalytic hydrogenation of D-xylose. 9
However, this process is too expensive owing to the need to use high pressure and 10
temperature as well as several steps of raw material purification (Silva, 1994; Winkelhausen 11
& Kusmanova, 1998). These reasons have been stimulating the search for alternative methods 12
for the production of such a polyol. To this purpose, the use of an optimized biotechnological 13
process could be a cheaper way to produce xylitol, because it would take place under mild 14
conditions and make use of unpurified hemicellulosic hydrolyzates. However, the 15
concentrations, yields and production rates obtained from fermentation media consisting in 16
lignocellulosic hydrolyzates are still the bottlenecks of a large-scale process, although they 17
can be improved by selecting the right fermentation system, operation mode and cultivation 18
conditions (Sarrouh, Santos, & Silva, 2007). 19
Hydrolyzates from different lignocellulosic residues, among which sugarcane bagasse 20
(Rodrigues et al., 2006; Santos, Carvalho, Silva, & Converti, 2003; Sarrouh et al., 2007) and 21
rice straw (Roberto et al., 1994; Silva, Mussatto, & Roberto, 2006), have been satisfactorily 22
used as alternative media. Hemicellulose hydrolyzates have been subject to different 23
treatments (Domínguez, Gong, & Tsao, 1996; Gurgel, Furlan, Martínez, & Mancilha, 1998) 24
and cultivation conditions (Santos, Converti, Carvalho, Mussatto, & Silva, 2005a; Sarrouh et 25
al., 2007), aimed at increasing process yield and productivity. 26

4
The use of metabolically engineered yeasts, Saccharomyces cerevisiae or Candida 1
spp. among others, has been investigated as a promising alternative for industrial production 2
of xylitol. However, the biotechnological method has not yet been able to overcome the 3
advantages of chemical dehydrogenation. The production of xylitol by Candida yeasts has 4
been extensively studied with regards to their ability to overproduce xylitol (Granström, 5
Izumori, & Leisola, 2007). Their main advantage over the metabolically engineered S. 6
cerevisiae strains is that they are natural D-xylose consumers, thus maintaining the reduction–7
oxidation balance during xylitol accumulation. Moreover, due to the opportunistic pathogenic 8
nature of some Candida spp., their application in the food sector is limited. Although high 9
xylitol yields from D-xylose have been frequently reported, a lot of research-work is devoted 10
to optimization of the conditions, with particular concern to the effect of the oxygen level 11
(Santos et al., 2005a). Recent efforts also include cell recycling (Granström, 2002; Kwon, 12
Park, & Oh, 2006) to prolong the production phase and thus increase the xylitol productivity 13
or screening for new strains (Guo et al., 2006; López, Delgado, Martínez, Spencer, & 14
Figueroa, 2004; Sampaio et al., 2005). 15
Immobilized cell systems have been traditionally considered as an alternative to 16
increase the process productivity and minimize the production costs, while offering 17
advantages over free cell fermentation systems (Carvalho et al., 2003; Santos et al., 2005a; 18
Sarrouh et al., 2007). Among the various matrices available for immobilization, Ca-alginate 19
beads stand out, because their manufacture does not require drastic conditions and, in regard 20
to food applications, utilize ingredients that are well accepted by the food industry (Mathew & 21
Abraham, 2005). 22
Sugarcane bagasse, the residue obtained after crushing the sugarcane to extract the 23
broth, is the most abundant lignocellulosic residue in Brazil, with an overall production of 24
about 103-million tons per year. Although most of bagasse has been employed in the own 25
sugarcane industry to generate energy, there is a surplus of this agro-industrial residue, and 26

5
several alternatives for its utilization have been evaluated, among which the production of 1
animal feed, single cell protein, ethanol, paper, enzymes, food additives such as vanillin 2
(Mathew & Abraham, 2005) and xylitol (Carvalho et al., 2003, 2005; Santos et al., 2003, 3
2005a). 4
For industrial purposes, an important choice criterion is the carrier cost, which, 5
combined with the interest in by-products recycling, has been leading to an increasing search 6
for cheap and available potential cell carriers. For this reason, the use of lignocellulosic 7
materials as cell carriers in fermentative processes has been explored. Pretreatments are 8
usually carried out aiming to increase the affinity of the biocatalyst to the lignocellulosic 9
materials (Brányik, Vicente, Cruz, & Teixeira, 2001). In this respect, sugarcane bagasse has 10
already been used with promising results as cell support in different bioprocesses (Sene, 11
Converti, Felipe, & Zilli, 2002). 12
The ability of sugarcane bagasse fibers as an alternative low cost biomaterial for the 13
immobilization of Candida guilliermondi cells has been evaluated in this work, aiming to 14
develop a new continuous or repeated-batch process of xylose-to-xylitol bioconversion from 15
sugarcane hemicellulosic hydrolyzate. 16
17
2. Materials and methods 18
19
2.1. Alkaline pretreatment of sugarcane bagasse 20
To increase the affinity of cells to the lignocellulosic material, sugarcane bagasse was 21
pretreated according to a variant of the methodology proposed by Brányik et al. (2001). It was 22
initially ground passing through a 14 mesh (1.41 mm) standard Tyler sieve and retained in a 23
35 mesh (0.5 mm) sieve. After separation and screening, the powdered sugarcane bagasse was 24
washed with distilled water and dried at 100°C up to constant weight. Aliquots of 0.5 g of 25
dried bagasse were sterilized at 121°C for 20 min in 125 mL-Erlenmeyer flasks, mixed with 26

6
8.3 mL of 0.5 M NaOH solution, then incubated in a rotary shaker at 120 rpm for 24 h, and 1
finally washed again with sterilized water (125 mL) and dried under the same conditions. 2
At the end of preliminary tests, the weight loss consequent to the alkaline treatment 3
was determined with the help of control tests and then taken into account for immobilized 4
biomass determination (see below). 5
6
2.2. Microorganism maintenance and inoculum preparation 7
Cells of the yeast Candida guilliermondii FTI 20037, belonging to the culture 8
collection of the Biotechnology Department of Lorena Engineering School of São Paulo 9
University, were maintained at 4°C in a medium containing agar malt extract (Merck, 10
Darmstadt, Germany). 11
To prepare the inoculum, yeast cells were cultured at 30°C and 200 rpm for 24 h in 12
250 mL-Erlenmeyer flasks containing 50 mL of a cultivation medium composed of 30 g/L 13
xylose, 3 g/ L (NH4)2SO4, 0.1 g/ L CaCl2.2H2O and 10% (w/v) rice bran extract, placed in an 14
incubator, model G25-KC (New Brunswick, Edison, NJ). After growth, cells were recovered 15
by centrifugation, model CU-500 (Damon/IEC, Needham, MA), at 2000 x g for 20 min, 16
washed and resuspended in isotonic solution (9 g/L NaCl in distilled water) in order to get a 17
highly concentrated cell suspension. 18
To obtain always the same inoculum (1.0 g/L wet weight), cell concentration in this 19
suspension was determined by optical density measurements (see below), and then 20
appropriate aliquots were added to the fermentation medium. 21
22
2.3. Cultivation conditions 23
All the fermentations were performed in duplicate in 125 mL-Erlenmeyer flasks 24
containing 50 mL of the same medium used for inculum preparation inoculated with 1.0 g/L 25
of cells (wet weight). The flasks were maintained in rotary shaker for 24 h at 30°C and 200 26

7
rpm. Aliquots of the medium of each fermentation were taken in duplicate after regular time 1
intervals of 3 h for sampling analysis. The standard deviations never exceeded 8%; therefore, 2
no additional statistical analysis was considered to be necessary. 3
4
2.4. Analytical determinations 5
The mass of cells adsorbed onto the support particles was quantified by gravimetry. 6
Sugarcane bagasse particles recovered from the cell-free fermentation media were dried and 7
used as a control to avoid any interference in weighing measurements. To this purpose, the 8
dry mass placed in the flask subtracted by the average weight loss was assumed as the starting 9
support amount. This was determined by weighing the sugarcane bagasse in an analytical 10
balance, model AB204 (Micronal S/A, São Paulo-SP, Brazil), after its alkaline pretreatment. 11
The support with adsorbed cells was screened through 60 mesh sieve to separate the liquid 12
phase, washed with 10 mL of distilled water and then dried at 100°C up to constant weight. 13
Cell concentration in the resulting suspension was determined by optical density (OD) 14
measurements at 640 nm, using a spectrophotometer, model DU 640B (Beckman Coulter, 15
Fullerton, CA). The control was the fermentation medium without cells and sugarcane 16
bagasse particles to avoid any interference of solids on the OD measurements. A previously 17
constructed calibration curve was used to relate the OD measurements to dry cell 18
concentration in samples of both this suspension and those used for inoculum. 19
Xylose and xylitol concentrations were measured by HPLC, model LC-10-AD 20
(Shimadzu, Tokyo, Japan), equipped with an Aminex HPX-87H (300 x 7.8 mm) column 21
(Bio-Rad, Hercules, CA) and a refractive index RID 6A detector. Samples were previously 22
filtered through a Sep Pak C18 filter and injected in the chromatograph under the following 23
conditions: injection volume of 20 µL, column temperature of 45°C, 0.01 N H2SO4 as the 24
mobile phase used at a flow rate of 0.6 mL/min. 25
26

8
2.5. Evaluation of immobilization and fermentation parameters 1
Cell retention onto the bagasse (R, g/g) was calculated as the ratio of mass of cells 2
immobilized in the carrier (g) to the carrier mass (g). The concentration of immobilized cells 3
(Xi, g/L) was calculated as the ratio of mass of cells immobilized in the carrier (g) to the 4
medium volume (L). The immobilization efficiency (Yi, %) was calculated as the ratio of Xi to 5
the concentration of total (suspended plus immobilized) cells (XT, g/L) and multiplying by 6
100. 7
The kinetic parameters and yields of batch fermentations were calculated at the end of 8
the immobilization process (24 h). 9
The xylitol volumetric productivity (QP, g L-1
h-1
) was calculated as the ratio of xylitol 10
concentration at the end of the run (Pf, g/L) to the fermentation time (t, h): 11
12
fP
t
PQ (1) 13
14
The yield of xylitol on consume xylose (YP/S, g/g) was defined as: 15
16
fo
f/P
SS
PY S
(2) 17
18
where So (g/L) and Sf (g/L) are the starting and the final xylose concentrations, respectively. 19
The efficiency of xylose conversion to xylitol (, %) has been estimated by the 20
relationship: 21
22
100 x th
/P
Y
Y S (3) 23
24

9
where Yth is the theoretical value of YP/S (0.917 g/g) proposed by Barbosa, Medeiros, 1
Mancilha, Schneider, & Lee (1988) on the basis of the knowledge on the metabolism of 2
Candida guilliermondii. 3
The yield of biomass con consumed xylose (YX/S, g/g) was defined as: 4
5
fo
T/X
SS
XY S
(4) 6
7
The volumetric rate of xylose consumption (QS, g L-1
h-1
) was calculated as: 8
9
fo
t
SSQS
(5) 10
11
12
3. Results and discussion 13
Numerous biotechnological processes were shown to be favored by cell 14
immobilization; therefore, different immobilization techniques and support materials have 15
been proposed. These techniques can be divided into four major categories based on the 16
physical mechanism employed: (a) attachment or adsorption on solid carrier surfaces, (b) 17
entrapment within a porous matrix, (c) self-aggregation by flocculation (natural) or with 18
cross-linking agents (artificially induced), and (d) cell containment behind barriers 19
(Pilkington, Margaritis, Mensour, & Russell, 1998). In general, cell immobilization allows 20
obtaining high cell concentration and productivity as a consequence of effective retention of 21
cells within the bioreactor, thus avoiding or at least remarkably reducing the washout 22
phenomenon (Ramakrishna & Prakasham, 1999). Besides, the possibility of recycling cells 23
for inoculum permits the fermentation to be profitably carried out in continuous or repeated-24
batch mode (Corcoran, 1985; Park & Chang, 2000). 25

10
It has been proposed that cell immobilization on a solid carrier is carried out by 1
physical adsorption due to electrostatic forces or by covalent binding between the cell 2
membrane and the carrier (Navarro & Durand, 1977; Norton & D’Amore, 1994). The 3
thickness of cell film usually ranges from one layer of cells to 1 mm or more. Systems using 4
immobilized cells on a surface are popular due to the relative easiness of carrying out this 5
type of immobilization. The strength of cell bound to the carrier as well as the biofilm depth 6
often varies and thus cannot be readily determined. As there are no barriers between the cells 7
and the solution, cell detachment and relocation is possible with possible occurrence of an 8
equilibrium between adsorbed and freely suspended cells. Examples of solid carriers used in 9
this type of immobilization are cellulosic materials (DEAE-cellulose, wood, sawdust, 10
delignified sawdust), inorganic materials (polygorskite, montmorilonite, hydromica, porous 11
porcelain, porous glass), etc. (Simeonova, 2004). Solid materials like glass or cellulose can 12
also be treated with polycations, chitosan or other chemicals (pre-formed carriers) to enhance 13
their adsorption power. The sugarcane bagasse is proposed and preliminarily investigated in 14
this study as an alternative, abundant, cheap immobilization support. 15
The yield of immobilization (Yi) and retention (R) of Candida guilliermondii cells by 16
pretreated sugarcane bagasse increased along the fermentation, reached maximum values of 17
50.5 % ± 1.41 and 0.31 ± 0.02 g immobilized cells/g bagasse after 21 h and then sharply 18
decreased (Figure 1). The time behavior of both parameters can be explained, according to 19
Veliky and McLean (1994), with the occurrence of two successive steps. In the former step, 20
characterized mainly by physico-chemical interactions, the cells were likely transferred and 21
effectively linked the solid support by surface attraction. The latter step would have been 22
characterized by the subsequent agglomeration of cells, thanks to their likely production of a 23
mucilaginous adhesive substance able to act agglutinating them to the solid surface (Veliky & 24
McLean, 1994). Santos, Mussatto Dragone, Converti, & Silva (2005b), who already reported 25
the formation of this substance during C. guilliermondii cells attachment onto porous glass 26

11
beads, proposed that the structure and composition of this matrix would vary among biofilms, 1
with polysaccharides and proteins usually accounting for up to 65% of its composition. 2
Moreover, it has been proposed that extra cellular polysaccharide glycocalyx (McLean, 3
Caldwell, & Costerton, 1994) has no other special function besides its participation in the cell 4
attachment process (Jones, Roth, & Sanders, 1969). However, to get further information on 5
this mechanism, additional work is needed to identify the components linked to the yeast cell 6
wall or those present in the extracellular matrix. 7
Several authors proposed that the process of cell adhesion by biosorption consists at 8
least of van der Waals forces and electrostatic interactions (Absolom, Zingg, & Neumam, 9
1985; Facchini, Neumam, & DiCosmo, 1988; Mozes & Rouxhet, 1985). These adhesion 10
forces are affected by variations in the medium composition and component concentrations, 11
because they can strongly influence the surface energy of the immobilization support. For 12
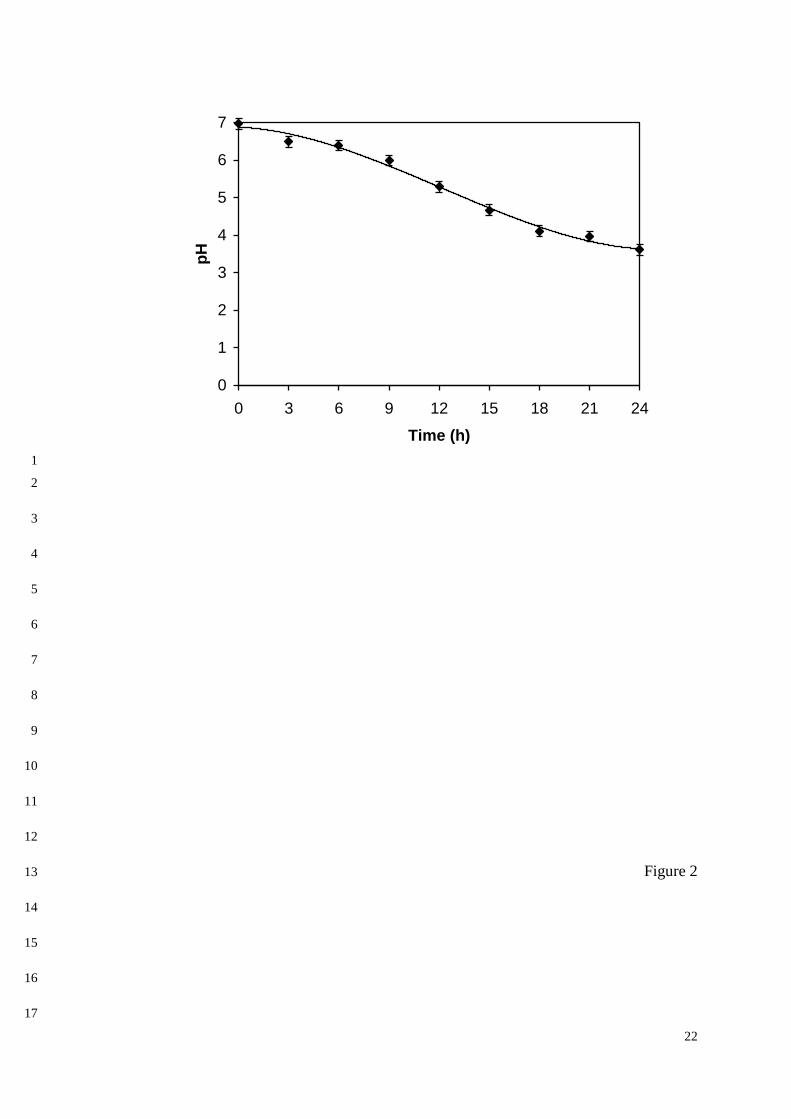
simplicity, these changes were followed in this study as pH variations along the time (Figure 13
2). 14
The observed pH behavior was indeed expected by the fact that cells were 15
metabolically active during the adhesion process and then consumed (NH4)2SO4 as the 16
nitrogen source and xylose as the carbon source. The former activity is fact responsible for 17
continuously release of H+ ions and the latter for CO2 development. As is well known, yeast 18
cells possess a large number of negative charges on the cell wall, because of the presence of 19
acidic (mainly carboxylic) groups that are negatively ionized under normal acidic conditions. 20
A pH decrease like that shown in Figure 2 certainly reduced the net surface charge of cells, 21
thus diminishing their electrostatic repulsion and favoring their adhesion to the support. 22
However, after about 21 h, the pH decreased to values lower than the pKa of carboxylic 23
groups (3.5-4.0); therefore, other alkaline groups present on the cell surface could have 24
become positively charged, hence newly promoting cell repulsion and their desorption from 25
the support to the medium. 26

12
As illustrated in Figure 3, the specific growth rate of suspended cells continuously 1
increased during the immobilization tests, while that of the immobilized ones, after an initial 2
growth, exhibited decreasing values. It is likely that a dynamic exchange took place between 3
immobilized and free cells, i.e., such a behavior could have been the result not only of simple 4
growth of the free cells but also of partial detachment of the immobilized ones and their 5
subsequent growth in the medium. Santos et al. (2005b) observed the same behavior working 6
with porous glass and zeolite as solid immobilization supports. However, only 50% of total 7
cells resulted to be entrapped in porous glass and less than 30% in zeolite beads at the end of 8
batch immobilization tests, likely due to the less porosity of the latter material. 9
The main fermentation parameters obtained during tests of cell immobilization onto 10
sugarcane bagasse pretreated by 0.5 M NaOH were YP/S = 0.65 g/g, YX/S = 0.18 g/g, QX = 0.13 11
g L-1
h-1
, QP = 0.66 g L-1
h-1
, QS = 1.01 g L-1
h-1
and = 70.8 %, which constitute quite 12
promising results taking into account that they were obtained under conditions selected to 13
exalt cell immobilization rather fermentation. Santos et al. (2005b) obtained YP/S = 0.52-0.53 14
g/g, QX = 0.10 g L-1
h-1
, QP = 0.32-0.33 g L-1
h-1
and = 56.7-57.8 % using zeolite and porous 15
glass as immobilization supports under comparable experimental conditions. The better 16
performance obtained with sugarcane bagasse in this work is probably the result of higher 17
biomass formation that allowed for higher xylitol yield and productivity. 18
Carvalho et al. (2003) reported YP/S = 0.54 g/g, QX = 0.08 g L-1
h-1
, QP = 0.44 g L-1
h-1
19
and = 58.9 % with Ca-alginate as an immobilization support. Those authors justified their 20
low productivity value as the result of limitation of oxygen diffusion into the beads, which 21
affected the growth of the immobilized cells. Compared with those results, the higher xylitol 22
efficiency and productivity observed in this work are probably due to better mass transfer in 23
the presence of sugarcane bagasse as a support, which would be linked to the particular 24
composition and the mechanical characteristics of this material. 25

13
To shed light on this phenomenon, the experimental data of the carbon sources 1
(glucose, xylose, arabinose and acetate) consumptions were used in the carbon material model 2
previously proposed by Cunha, Converti, Santos, & Silva (2006) to estimate the average 3
fractions of xylose progressively consumed by the anaerobic and semi-aerobic xylose-to-4
xylitol bioconversions, biomass synthesis from xylose, and xylose consumption by the 5
catabolic reaction (Figure 4). These results, which are qualitatively similar to those obtained 6
using polyvinyl alcohol hydrogel as cell matrix (Cunha et al., 2006), show that the fraction of 7
xylose consumed for the anaerobic xylitol formation progressively increased up to more than 8
43% of the overall uptake at the end of the bioconversion. This behaviour suggests that 9
oxygen was progressively depleted in the medium leading to oxygen-limited conditions inside 10
the matrix, which notoriously stimulate xylitol accumulation. 11
To confirm the occurrence of immobilization onto sugarcane bagasse particles as well 12
as the potential of this raw material as cell support, optical microphotographs were done at the 13
end of experiments. As is demonstrated by the uniform cell growth onto the whole surface of 14
bagasse fibers (Figure 5), cell immobilization was effective. This result suggests possible 15
recycling of cells in repeated-batch runs, also taking into account the fact that they were 16
shown to intensively grow even after the immobilization process (Figure 6). According to 17
Sarrouh et al. (2007), an increase in xylitol yield could be achieved by recycling the 18
immobilized cells through a repeated-batch fermentation system, thanks to the possible cell 19
adaptation to the toxic compounds present in the hydrolyzate (Carvalho et al., 2003, Santos et 20
al., 2005a). 21
22
4. Conclusions 23
The immobilization efficiency and cell retention were utilized as parameters to 24
evaluate and investigate the ability of sugarcane bagasse fibers to immobilize cells of Candida 25
guilliermondii to be used in fermentation processes. Using xylitol production as a model to 26

14
check this hypothesis, both parameters exhibited increasing values up to 21 h, corresponding 1
to about pH 4, and then decreased after this time, likely due to repulsive electrostatic 2
interactions resulting from significant acidification of the medium. Useful information about 3
the mechanism of cells attachment and detachment from support particles was also provided 4
by the behaviors of the specific growth rates of suspended and immobilized cells. Optical 5
microphotographs showed that cells colonized the sugarcane bagasse uniformly and 6
abundantly, thus demonstrating the promising ability of this raw material to immobilize them. 7
The next attempt will include an optimization study in shaker by techniques of 8
experimental planning aiming at efficient utilization of this raw material as cell support in 9
continuous or repeated-batch xylose-to-xylitol bioconversion from hemicellulosic 10
hydrolyzates in bioreactor. Among the sensitive parameters to be optimized, pH will certainly 11
be among those of utmost significance because of its profound effect demonstrated in this 12
study on the immobilization efficiency. 13
14
5. Acknowledgements 15
The authors gratefully acknowledge the financial support of FAPESP (Fundação de 16
Amparo à Pesquisa do Estado de São Paulo), CAPES (Coordenação de Aperfeiçoamento de 17
Pessoal de Nível Superior) and CNPq (Conselho Nacional de Desenvolvimento Científico e 18
Tecnológico), Brazil. 19
20
6. References 21
Absolom, S. R., Zingg, W., & Neumam, A.W. (1985). Surface thermodynamics of cellular 22
and protein interactions. In C. W. Robinson, & J. A. Howell (Eds.). Comprehensive 23
biotechnology (Vol. 4, pp. 433-446). Oxford, UK: Pergamon Press. 24
Barbosa, M. F. S., Medeiros, M. B., Mancilha, I. M., Schneider H., & Lee H. J. (1988). 25
Screening of yeasts for production of xylitol from D-xylose and same factors which affect 26

15
xylitol yield in Candida guilliermondii. Journal of Industrial Microbiology, 3(4), 241-1
251. 2
Brányik, T., Vicente, A. A., Cruz, J. M. M., & Teixeira, J. A. (2001). Spent grains – a new 3
support for brewing yeast immobilization. Biotechnology Letters, 23(13), 1073-1078. 4
Carvalho, W., Santos, J. C., Canilha, L., Silva, S. S., Perego, P., & Converti, A. (2005). 5
Xylitol production from sugarcane bagasse hydrolysate: Metabolic behaviour of Candida 6
guilliermondii cells entrapped in Ca-alginate. Biochemical Engineering Journal, 25(1), 7
25-31. 8
Carvalho, W., Silva, S. S., Santos, J. C., & Converti, A. (2003). Xylitol production by Ca-9
alginate entrapped cells: comparison of different fermentation systems. Enzyme and 10
Microbial Technology, 32(5), 553-559. 11
Corcoran, E. (1985). The production and use of immobilized living microbial cells. In A. 12
Wiseman (Ed.). Topics in enzyme and fermentation biotechnology (pp. 12-50). Chichester, 13
UK: Ellis Horwood. 14
Cunha, M. A. A., Converti, A., Santos, J. C., & Silva, S. S. (2006). Yeast immobilization in 15
Lentikats®: a new strategy for xylitol bioproduction from sugarcane bagasse. World 16
Journal of Microbiology and Biotechnology, 22(1), 65-72. 17
Domínguez, J. M., Gong, G. S., & Tsao, G. T. (1996). Pretreatment of sugar cane bagasse 18
hemicellulose hydrolysate for xylitol production by yeasts. Applied Biochemistry and 19
Biotechnology, 57/58, 49–56. 20
Facchini, P. J., Neumam, A. W., & DiCosmo, F. (1988). Thermodynamic aspects of cell 21
adhesion to polymer surfaces. Applied Microbiology and Biotechnology, 29(4), 346-355. 22
Granström, T. (2002). Biotechnological production of xylitol with Candida yeasts. Helsinky, 23
Finland: PhD thesis, Helsinki University of Technology. 24

16
Granström, T. B., Izumori, K., & Leisola, M. (2007). A rare sugar xylitol. Part II: 1
Biotechnological production and future applications of xylitol. Applied Microbiology and 2
Biotechnology, 74(2), 273-276. 3
Guo, C., Zhao, C., He, P., Lu, D., Shen, A., & Jiang, N. (2006). Screening and 4
characterization of yeasts for xylitol production. Journal of Applied Microbiology, 101(5), 5
1096–1104. 6
Gurgel, P. V., Furlan, S. A., Martínez, S. E. R., & Mancilha, I. M. (1998). Evaluation of 7
sugarcane bagasse acid hydrolyzate treatments for xylitol production. Brazilian Journal of 8
Chemical Engineering, 15(3), 309–312. 9
Jones, H. C., Roth, I. L., & Sanders, W. M. (1969). Electron microscopic study of a slime 10
layer. Journal of Bacteriology, 99(1), 316–325. 11
Kontiokari, T., Uhari, M., & Koskela, M. (1998). Antiadhesive effects on otopathogenic 12
bacteria. Journal of Antimicrobial Chemotherapy, 41(5), 563-565. 13
Kwon, S.-G., Park, S.-W., & Oh, D.-K. (2006). Increase of xylitol productivity by cell-recycle 14
fermentation of Candida tropicalis using submerged membrane bioreactor. Journal of 15
Bioscience and Bioengineering, 101(1), 13–18. 16
López, F., Delgado, O. D., Martínez, M. A., Spencer, J. F. T., & Figueroa, L. I. C. (2004). 17
Characterization of a new xylitol-producer Candida tropicalis strain. Antonie van 18
Leeuwenhoek, 85(4), 281–286. 19
Masako, K., Hideyuki, I., Shigeyuki, O., & Zenro, I. (2005a). A novel method to control the 20
balance of skin microflora. Part 1. Attack on biofilm of Staphylococcus aureus without 21
antibiotics. Journal of Dermatological Science, 38(3), 197-205. 22
Masako, K., Yusuke, K., Hideyuki, I., Atsuko, M., Yoshiki, M., Kayoko, M., & Makoto, K. 23
(2005b). A novel method of to control the balance of skin microflora. Part 2. A study to 24
assess the effect of a cream containing farnesol and xylitol on atopic dry skin. Journal of 25
Dermatological Science, 38(3), 207-213. 26

17
Mathew, S., & Abraham, T. E. (2005). Studies on the production of feruloyl esterase from 1
cereal brans and sugar cane bagasse by microbial fermentation. Enzyme and Microbial 2
Technology, 36(4), 565-570. 3
McLean, R. J. C., Caldwell, D. E., & Costerton, J. W. (1994). Biofilms, naturally occurring 4
communities of immobilized cells. In I. A. Veliky, & R. J. C McLean (Eds.). Immobilized 5
biosystems. Theory and practical applications (pp. 289–335). Glasgow, UK: Blackie 6
Academical and Professional. 7
Mozes, N., & Rouxhet, P. G. (1985). Metabolic activity of yeast immobilized as support 8
monolayer. Applied Microbiology and Biotechnology, 22(2), 92-97. 9
Naaber, P., Lehto, E., Salminen, S., & Mikelsaar, M. (1996). Inhibition of adhesion of 10
Clostridium difficile to Caco-2 cells. FEMS Immunology and Medical Microbiology, 11
14(4), 205-209. 12
Navarro, J. M., & Durand, G. (1977). Modification of yeast metabolism by immobilization 13
onto porous glass. European Journal of Applied Microbiology, 4(4), 243-254. 14
Norton, S., & D’Amore, T. (1994). Physiological effects of yeast cell immobilization: 15
applications for brewing. Enzyme and Microbial Technology, 16(5), 365-375. 16
Park, J. K., & Chang, H. N. (2000). Microencapsulation of microbial cells. Biotechnology 17
Advances, 18(4), 303-319. 18
Pilkington, P. H., Margaritis, A., Mensour, N. A., & Russell, I. (1998). Fundamentals of 19
immobilized yeast cells for continuous beer fermentation: a review. Journal of the 20
Institute of Brewing, 104, 19-31. 21
Ramakrishna, S. V., & Prakasham, R. S. (1999). Microbial fermentations with immobilized 22
cells. Current Science, 77(1), 87-100. 23
Roberto, I. C., Mancilha, I. M., Souza, C. A., Felipe, M. G. A., Sato, S., & Castro, H. F. 24
(1994). Evaluation of rice straw hemicellulose hydrolysate in the production of xylitol by 25
Candida guilliermondii. Biotechnology Letters, 16(11), 1211–1216. 26

18
Rodrigues, R. C. L. B., Sene, L., Matos, G. S., Roberto, I. C., Pessoa Jr, A., & Felipe, M. G. 1
A. (2006). Enhanced xylitol production by precultivation of Candida guilliermondii cells 2
in sugarcane bagasse hemicellulosic hydrolysate. Current Microbiology, 53(1), 53-59. 3
Sampaio, F. C., Mantovani, H. C., Passos, F. J. V., Moraes, C. A., Converti, A., & Passos, F. 4
M. (2005). Bioconversion of D-xylose to xylitol by Debaryomyces hansenii UFV-170: 5
product formation versus growth. Process Biochemistry, 40(11), 3600-3606. 6
Santos, J. C., Carvalho, W., Silva, S. S., & Converti, A. (2003). Xylitol production from 7
sugarcane bagasse hydrolyzate in fluidized bed reactor. Effect of air flowrate. 8
Biotechnology Progress, 19(4), 1210-1215. 9
Santos, J. C., Converti, A., Carvalho, W., Mussatto S. I., & Silva, S. S. (2005a). Influence of 10
aeration rate and carrier concentration on xylitol production from sugarcane bagasse 11
hydrolysate in immobilized-cell fluidized bed reactor. Process Biochemistry, 40(1), 113-12
118. 13
Santos, J. C., Mussatto S. I., Dragone, G., Converti, A., & Silva, S. S. (2005b) Evaluation of 14
porous glass and zeolite as cells carriers for xylitol production from sugarcane bagasse 15
hydrolysate. Biochemical Engineering Journal, 23(1), 1–9. 16
Sarrouh, B. F., Santos D. T., & Silva, S. S. (2007). Biotechnological production of xylitol in a 17
three phase fluidized bed bioreactor with immobilized yeast cells in Ca-alginate beads. 18
Biotechnology Journal, 2(6), 759-763. 19
Sene, L., Converti, A., Felipe, M. G. A., & Zilli, M. (2002). Sugarcane bagasse as alternative 20
packing material for biofiltration of benzene polluted gaseous streams. A preliminary 21
study. Bioresource Technology, 83(2), 153-157. 22
Silva, C. J. S. M., Mussatto, S. I., & Roberto, I. C. (2006). Study of xylitol production by 23
Candida guilliermondii on a batch bioreactor. Journal of Food Engineering, 75(1), 115-24
119. 25

19
Silva, S. S. (1994). Produção de xilitol por via biotecnológica: Estudo de sistemas de 1
biorreatores e parâmetros fermentativos. São Paulo, Brazil: Faculty of Pharmaceutical 2
Sciences, University of São Paulo, PhD Thesis, pp. 219. 3
Simeonova, D. D. (2004). Arsenic oxidation of Cenibacterium arsenoxidans: Potential 4
application in bioremediation of arsenic contaminated water. Strasburg, France: Louis 5
Pasteur University Strasburg I, PhD Thesis, pp. 155. 6
Tischler, A. S., Powers, J. F., Downing, J. C., Riseberg, J. C., Shahsavari, M., Ziar, J., & 7
McClain, R. M. (1996). Vitamin D3, lactose, and xylitol stimulate chromaffin cell 8
proliferation in rat adrenal medulla. Toxicology and Applied Pharmacology, 140(1), 115-9
123. 10
Veliky, I. A., & McLean, R. J. C. (1994). Immobilized biosystems theory and practical 11
applications (pp. 263-287). Glasgow, UK: Chapman & Hall. 12
Wang, Y. M., & Van Eys, J. (1981). Nutritional significance of fructose and sugar alcohols. 13
Annual Review of Nutrition, 1, 435-475. 14
Winkelhausen, E., & Kusmanova, S. (1998). Microbial conversion of D-xylose to xylitol. 15
Journal of Fermentation and Bioengineering, 86(1), 1-14. 16
Wiseman, A. (Ed). Topics in enzyme and fermentation biotechnology (pp. 12-50). Chichester, 17
UK: Ellis Horwood. 18
Zabner, J., Seiler, M. P., Launspach J. L., Karp, P. H., Kearney, W. R., Look, D. C., et al. 19
(2000). The osmolyte xylitol reduces the salt concentration of airway surface liquid and 20
may enhance bacterial killing. In Proceedings of the National Academy of Sciences of the 21
United States of America, 97(21), 11614-11619. 22
23

20
Figure Captions 1
2
Figure 1. Immobilization efficiency and retention of Candida guilliermondii cells onto 3
sugarcane bagasse as function of time. () Immobilization efficiency; () Cell retention. 4
5
Figure 2. Time variation of pH during the immobilization of Candida guilliermondii cells 6
onto sugarcane bagasse. 7
8
Figure 3. Time behavior of the specific growth rate of Candida guilliermondii cells during 9
immobilization onto sugarcane bagasse. () Total cells; () suspended cells; () 10
immobilized cells. 11
12
Figure 4. Time variations of the average fractions of xylose progressively consumed by 13
Candida guilliermondii cells during xylose-to-xylitol bioconversion in sugarcane 14
hemicellulose hydrolysate. Anaerobic xylose-to-xylitol conversion (); semi-aerobic xylose-15
to-xylitol conversion (); biomass synthesis from xylose (); catabolic reaction of xylose by 16
the TCA cycle (). 17
18
Figure 5. Optical microphotograph of Candida guilliermondii cells (400X) immobilized onto 19
sugarcane bagasse fiber done after 24 h of fermentation. 20
21
Figure 6. Optical microphotograph of Candida guilliermondii cells (400X) showing intensive 22
cell growth on sugarcane bagasse fiber even after 24 h. 23
24

21
0
10
20
30
40
50
60
0 3 6 9 12 15 18 21 24
Time (h)
Imm
ob
iliz
ati
on
Eff
icie
nc
y (
%)
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
Ce
ll R
ete
nti
on
(g
/g)
1
2
3
4
5
6
7
8
9
10
Figure 1 11
12
13
14
22
0
1
2
3
4
5
6
7
0 3 6 9 12 15 18 21 24
Time (h)
pH
1
2
3
4
5
6
7
8
9
10
11
12
Figure 2 13
14
15
16
17

23
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
Figure 3 23
24
25
0.0
0.1
0.2
0.3
0.4
0.5
0 3 6 9 12 15 18 21 24
Time (h)
Sp
ec
ific
gro
wth
ra
te (
h-1
)

24
0
10
20
30
40
50
60
6 12 18 24
Time (h)
Pe
rce
nta
ge
of
xy
los
e c
on
su
mp
tio
n (
%)
1
2
3
4
5
6
7
8
9
10
Figure 4 11
12
13
14
15
16

25
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Figure 5 22
23
24
25
26

26
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
Figure 6 20
21
Copyright © 2022 FDOKUMEN




















