
Advances in Microbiome-Derived Solutions and ... - MDPI
27
Citation: Gueniche, A.; Perin, O.; Bouslimani, A.; Landemaine, L.; Misra, N.; Cupferman, S.; Aguilar, L.; Clavaud, C.; Chopra, T.; Khodr, A. Advances in Microbiome-Derived Solutions and Methodologies Are Founding a New Era in Skin Health and Care. Pathogens 2022, 11, 121. https://doi.org/10.3390/ pathogens11020121 Academic Editor: María-Teresa Pérez-Gracia Received: 25 October 2021 Accepted: 12 January 2022 Published: 20 January 2022 Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affil- iations. Copyright: © 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https:// creativecommons.org/licenses/by/ 4.0/). pathogens Review Advances in Microbiome-Derived Solutions and Methodologies Are Founding a New Era in Skin Health and Care Audrey Gueniche 1 , Olivier Perin 1 , Amina Bouslimani 2 , Leslie Landemaine 1 , Namita Misra 1 , Sylvie Cupferman 1 , Luc Aguilar 1 ,Cécile Clavaud 1 , Tarun Chopra 3 and Ahmad Khodr 1, * 1 L’Oréal Research and Innovation, 93600 Paris, France; [email protected] (A.G.); [email protected] (O.P.); [email protected] (L.L.); [email protected] (N.M.); [email protected] (S.C.); [email protected] (L.A.); [email protected] (C.C.) 2 L’Oréal Research and Innovation, Clark, NJ 07066, USA; [email protected] 3 L’Oréal Research and Innovation, Singapore 048583, Singapore; [email protected] * Correspondence: [email protected]; Tel.: +33-1-49-79-56-57 Abstract: The microbiome, as a community of microorganisms and their structural elements, genomes, metabolites/signal molecules, has been shown to play an important role in human health, with significant beneficial applications for gut health. Skin microbiome has emerged as a new field with high potential to develop disruptive solutions to manage skin health and disease. Despite an incomplete toolbox for skin microbiome analyses, much progress has been made towards functional dissection of microbiomes and host-microbiome interactions. A standardized and robust investigation of the skin microbiome is necessary to provide accurate microbial information and set the base for a successful translation of innovations in the dermo-cosmetic field. This review provides an overview of how the landscape of skin microbiome research has evolved from method development (multi- omics/data-based analytical approaches) to the discovery and development of novel microbiome- derived ingredients. Moreover, it provides a summary of the latest findings on interactions between the microbiomes (gut and skin) and skin health/disease. Solutions derived from these two paths are used to develop novel microbiome-based ingredients or solutions acting on skin homeostasis are proposed. The most promising skin and gut-derived microbiome interventional strategies are presented, along with regulatory, safety, industrial, and technical challenges related to a successful translation of these microbiome-based concepts/technologies in the dermo-cosmetic industry. Keywords: skin health; microbiome; postbiotics; microbiome metabolites; cosmetic; microbiome data; methodology harmonization 1. Introduction Applications of Microbiome sciences are very large and have been proposed as a po- tential target solution for the 21st century socio-economic and environmental challenges [1]. For several decades, scientists have been interested in the microbiome and its impact on human health. A major focus was put on the gut microbiome, and its role in human health has been well established [2]. New knowledge on lung, oral cavity, and skin microbiome is beginning to emerge [3]. A deeper knowledge of the microbiome, specifically that of the skin, opens perspectives for a revolution in dermo-cosmetic development. These re- cent discoveries have changed our perception of the role of bacteria in skin health. For example, microbiome-derived and personalized dermo-cosmetic development would be possible, due to the advancement in skin microbiome analysis and diagnosis [4]. New products that respect, protect or rebalance the skin microbiome are a new trend in the dermo-cosmetic industry. The main aim of this review is to share a global view covering mechanistic knowl- edge regarding the interaction between the microbiome (gut and skin) and the skin. It Pathogens 2022, 11, 121. https://doi.org/10.3390/pathogens11020121 https://www.mdpi.com/journal/pathogens
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Advances in Microbiome-Derived Solutions and ... - MDPI

�����������������
Citation: Gueniche, A.; Perin, O.;
Bouslimani, A.; Landemaine, L.;
Misra, N.; Cupferman, S.; Aguilar, L.;
Clavaud, C.; Chopra, T.; Khodr, A.
Advances in Microbiome-Derived
Solutions and Methodologies Are
Founding a New Era in Skin Health
and Care. Pathogens 2022, 11, 121.
https://doi.org/10.3390/
pathogens11020121
Academic Editor: María-Teresa
Pérez-Gracia
Received: 25 October 2021
Accepted: 12 January 2022
Published: 20 January 2022
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2022 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
pathogens
Review
Advances in Microbiome-Derived Solutions and MethodologiesAre Founding a New Era in Skin Health and CareAudrey Gueniche 1, Olivier Perin 1, Amina Bouslimani 2, Leslie Landemaine 1, Namita Misra 1 ,Sylvie Cupferman 1 , Luc Aguilar 1, Cécile Clavaud 1 , Tarun Chopra 3 and Ahmad Khodr 1,*
1 L’Oréal Research and Innovation, 93600 Paris, France; [email protected] (A.G.);[email protected] (O.P.); [email protected] (L.L.); [email protected] (N.M.);[email protected] (S.C.); [email protected] (L.A.); [email protected] (C.C.)
2 L’Oréal Research and Innovation, Clark, NJ 07066, USA; [email protected] L’Oréal Research and Innovation, Singapore 048583, Singapore; [email protected]* Correspondence: [email protected]; Tel.: +33-1-49-79-56-57
Abstract: The microbiome, as a community of microorganisms and their structural elements, genomes,metabolites/signal molecules, has been shown to play an important role in human health, withsignificant beneficial applications for gut health. Skin microbiome has emerged as a new fieldwith high potential to develop disruptive solutions to manage skin health and disease. Despite anincomplete toolbox for skin microbiome analyses, much progress has been made towards functionaldissection of microbiomes and host-microbiome interactions. A standardized and robust investigationof the skin microbiome is necessary to provide accurate microbial information and set the base for asuccessful translation of innovations in the dermo-cosmetic field. This review provides an overviewof how the landscape of skin microbiome research has evolved from method development (multi-omics/data-based analytical approaches) to the discovery and development of novel microbiome-derived ingredients. Moreover, it provides a summary of the latest findings on interactions betweenthe microbiomes (gut and skin) and skin health/disease. Solutions derived from these two pathsare used to develop novel microbiome-based ingredients or solutions acting on skin homeostasisare proposed. The most promising skin and gut-derived microbiome interventional strategies arepresented, along with regulatory, safety, industrial, and technical challenges related to a successfultranslation of these microbiome-based concepts/technologies in the dermo-cosmetic industry.
Keywords: skin health; microbiome; postbiotics; microbiome metabolites; cosmetic; microbiomedata; methodology harmonization
1. Introduction
Applications of Microbiome sciences are very large and have been proposed as a po-tential target solution for the 21st century socio-economic and environmental challenges [1].For several decades, scientists have been interested in the microbiome and its impact onhuman health. A major focus was put on the gut microbiome, and its role in human healthhas been well established [2]. New knowledge on lung, oral cavity, and skin microbiomeis beginning to emerge [3]. A deeper knowledge of the microbiome, specifically that ofthe skin, opens perspectives for a revolution in dermo-cosmetic development. These re-cent discoveries have changed our perception of the role of bacteria in skin health. Forexample, microbiome-derived and personalized dermo-cosmetic development would bepossible, due to the advancement in skin microbiome analysis and diagnosis [4]. Newproducts that respect, protect or rebalance the skin microbiome are a new trend in thedermo-cosmetic industry.
The main aim of this review is to share a global view covering mechanistic knowl-edge regarding the interaction between the microbiome (gut and skin) and the skin. It
Pathogens 2022, 11, 121. https://doi.org/10.3390/pathogens11020121 https://www.mdpi.com/journal/pathogens

Pathogens 2022, 11, 121 2 of 27
also focuses on how this knowledge is translated into products/concepts in the dermo-cosmetic field. The latest methodological developments for skin microbiome analysis andthe specific regulatory/safety environment of microbiome-derived solutions used in thedermo-cosmetic industry are summarized. The translational challenges of the presentedmicrobiome-based concepts are also discussed.
2. The Skin and Its Microbiome
In adults, considering the appendages, the skin forms a large surface of 30 m2 [5]. Thisextensive surface constitutes an important protective barrier that is physically ensured byepithelial cells and is also ensured by the presence of a beneficial microbiome that interfereswith the three previously stated barrier properties.
The microbiome is an essential partner to our skin. It is a beneficial and invisibleecosystem of living microorganisms that is an integral part of our skin’s surface. It is anatural ecosystem of microbes that protects our skin from external damage and acts asa second genome, interacting with our bodies to ensure healthy functioning. It plays amajor role in our skin defense and regulates the exchanges between the body and theenvironment. Its balance is essential to our skin health and beauty.
The composition of the skin microbiome varies according to body sites that constitutediverse ecological/physicochemical niches. Briefly, the skin can be moist, dry, or sebaceous,and each of these classes has a distinct subset of microbial taxa that are particularly wellsuited to these conditions. Lipid content is a key factor driving the microbiome compositionthat drastically changes between dry and sebaceous sites [6,7]. On sebaceous sites lipophilicCutibacterium species (spp.) are the most abundant, whereas bacteria such as Staphylococcusand Corynebacterium spp. are preferentially abundant in moist areas, Cutibacterium spp.,Staphylococcus spp., and Streptococcus spp. are the most abundant bacteria on dry sites, [8,9].Malassezia spp. is the most abundant fungi across the body, except for the sites on the footwhich present greater diversity. Cutibacterium acnes (C. acnes) phage is the most representedvirus in different skin sites and eukaryotic viruses are more transient [9,10]. Cutibacteriumspp. and Staphylococcus spp. play important and multiple roles on the skin.
S. epidermidis is traditionally considered to be one of the major representative bacteriaof healthy skin Microbiome. It is among the most abundant species of the cultivablemicrobiome: up to 90% of the cultivable aerobic flora [11]. It is ubiquitous: present on thewhole body in dry, wet, and sebaceous areas [12]. Recent studies have shown its role inmaintaining an effective skin barrier in vitro [13,14], in wound healing [15]; in the fightagainst pathogens [16,17]; in modulating the immune system [15,18–21], and in preventingmelanoma [22].
C. acnes is also a highly represented bacterium in the human skin microbiome (>50%of bacterial species according to sequencing data), about 105 bacteria/cm2 on sebaceousareas (face, scalp, back; rich in sebum). C. acnes degrades long-chain fatty acids in sebum toshort-chain fatty acids (SCFA), including propionic acid (or propanoic acid) via its lipasesactivity [11]. Propionic acid is involved in skin odor, but more importantly, it maintainsthe pH of the skin and has antimicrobial properties [11]. C. acnes also participates inantimicrobial defenses through the secretion of bacteriocins or antimicrobial peptides likecutimycin [23,24]. It has been associated with and is believed to play a major role in skinhealth. It synthesizes free fatty acids such as vitamin B12 which deficiency can be associatedwith skin hyperpigmentation, vitiligo, peeling [25]. Vitamin B12 biosynthesis is decreasedin acne patients compared to healthy individuals [26]. Conjugated linoleic acid (anti-proliferative), riboflavin (vitamin B2, antioxidant [27] and folate (vitamin B9, play a role incell division and may protect the skin barrier especially following sun exposure [28–30] areother examples of the implication of C. acnes in skin health.
The interaction between S. epidermidis and C. acnes postbiotics/metabolites and skinhealth is detailed in the upcoming “effector molecules/metabolites” section.
Among the other bacteria, some have been described for their beneficial effects onthe skin, for example, Corynebacterium spp. which allows the regulation of the immune

Pathogens 2022, 11, 121 3 of 27
system [31], Micrococcus luteus degrade pollutants and isomerizes urocanic acid, which mayplay a role in UV protection [32,33].
Compared to other tissues, the skin microbiome shows the highest individual diversityand is less stable [34]. This is not surprising because of the exposure of the skin micro-biome to external factors (i.e., UV, humidity, pollution, environmental bacteria, cosmetics).The impact of external factors (exposome) on the microbiome has been comprehensivelyreviewed by Skowron et al. and is addressed hereafter [35].
Despite these extrinsic factors, samples and data generated from longitudinal studiesshow that the skin microbiome is, to some extent, stable at a strain level. Nevertheless,stable specific individual signatures at a strain or SNV level exist. This is the case forS. epidermidis on some specific sites [7,36]. This stability is not contradictory with theassumption that transient microbiome can enter the community from the environment(home, pets, other individuals).
Overall, the skin harbors a stable and diverse community of microorganisms thatinteract with the external environment, skin cells, and other microbial cells to maintainits homeostasis. Any dysbiosis driven by the overabundance of one of the commensalopportunistic microorganisms can be associated with skin diseases/conditions.
3. Skin/Scalp Conditions and Microbiome
The skin microbiome plays an important role in maintaining cutaneous health withthe skin microflora constantly adapting in response to intrinsic and extrinsic factors. Theenvironment, and therefore pollution exposure, has the potential to influence the skin mi-croflora and bacteria isolated from the human skin have been shown to degrade PolycyclicAromatic Hydrocarbons (PAHs) and related xenobiotic compounds [37].
3.1. Variability across Age
Today, a widely accepted assumption is that the environment of the fetus is sterile,and that colonization of the skin occurs at birth. This first microbiota varies, depending onthe mode of delivery: a vaginal birth allows colonization of the baby’s skin by the mother’svaginal microbiota (mainly Lactobacilli), while a C-section is associated with the colonizationby bacteria from the operating room environment and the mother’s skin (Staphylococcusspp., Corynebacterium spp., Cutibacterium spp.) [38]. The difference in microbiota acquired atbirth may have a longer-term impact on the microbiota that will develop in childhood [39].The low microbial diversity and the non-exposure to vaginal bacteria in children born bycesarean section could cause a delay in the development of their immunological functionswhich could partly explain their greater sensitivity to certain pathogens and allergies, andan increased risk of developing atopic dermatitis in childhood [40–43].
The microbiota changes gradually during the child’s first year. Unlike the adult indi-vidual, the skin microbiota of newborns is homogeneous throughout the body, regardlessof the type of delivery [44]. Primary colonization is characterized by a high proportion ofStaphylococci which will gradually decrease as the microbiota is enriched with new bacterialpopulations. The development of this diverse microbiota, from the first months of lifeonwards, with regards to the maturation of the skin, in particular its immune system,contributes to the establishment of skin homeostasis [45]. The diversity and composition ofthe microbiota remain stable during childhood.
At puberty, sebum secretion increases and promotes colonization by lipophilic bacteriaand in particular C. acnes, involved in acne [46]. Their population begins to increase at theage of 10 years, but it is especially between 15 and 25 years that studies have measured thestrongest expansion of these bacteria [47,48]. The increased secretion of sebum also affectspopulations of fungi. The fungal diversity observed in children decreases with age andlipophilic fungi are favored: the proportion of Malassezia increases and becomes dominant,in particular, M. restricta in sebaceous areas [49].
Between around 25 and 60 years of age, the microbiota is stable with equivalentenvironmental factors [7]. With age, from 55–70 years, depending on the population, the mi-
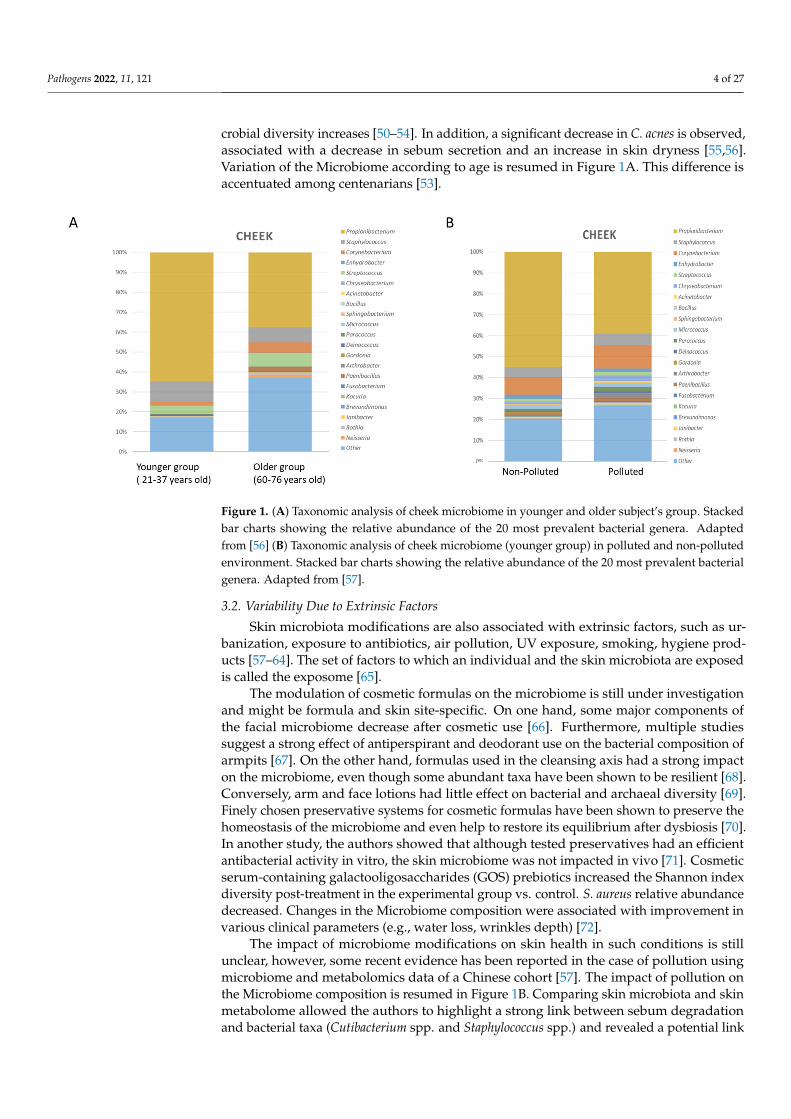
Pathogens 2022, 11, 121 4 of 27
crobial diversity increases [50–54]. In addition, a significant decrease in C. acnes is observed,associated with a decrease in sebum secretion and an increase in skin dryness [55,56].Variation of the Microbiome according to age is resumed in Figure 1A. This difference isaccentuated among centenarians [53].
Pathogens 2022, 11, x 4 of 28
At puberty, sebum secretion increases and promotes colonization by lipophilic bacteria and in particular C. acnes, involved in acne [46]. Their population begins to increase at the age of 10 years, but it is especially between 15 and 25 years that studies have measured the strongest expansion of these bacteria [47,48]. The increased secretion of sebum also affects populations of fungi. The fungal diversity observed in children decreases with age and lipophilic fungi are favored: the proportion of Malassezia increases and becomes dominant, in particular, M. restricta in sebaceous areas [49].
Between around 25 and 60 years of age, the microbiota is stable with equivalent environmental factors [7]. With age, from 55–70 years, depending on the population, the microbial diversity increases [50–54]. In addition, a significant decrease in C. acnes is observed, associated with a decrease in sebum secretion and an increase in skin dryness [55,56]. Variation of the Microbiome according to age is resumed in Figure 1A. This difference is accentuated among centenarians [53].
Figure 1. (A) Taxonomic analysis of cheek microbiome in younger and older subject’s group. Stacked bar charts showing the relative abundance of the 20 most prevalent bacterial genera. Adapted from [56] (B) Taxonomic analysis of cheek microbiome (younger group) in polluted and non-polluted environment. Stacked bar charts showing the relative abundance of the 20 most prevalent bacterial genera. Adapted from [57].
3.2. Variability Due to Extrinsic Factors Skin microbiota modifications are also associated with extrinsic factors, such as
urbanization, exposure to antibiotics, air pollution, UV exposure, smoking, hygiene products [57–64]. The set of factors to which an individual and the skin microbiota are exposed is called the exposome [65].
The modulation of cosmetic formulas on the microbiome is still under investigation and might be formula and skin site-specific. On one hand, some major components of the facial microbiome decrease after cosmetic use [66]. Furthermore, multiple studies suggest a strong effect of antiperspirant and deodorant use on the bacterial composition of armpits [67]. On the other hand, formulas used in the cleansing axis had a strong impact on the microbiome, even though some abundant taxa have been shown to be resilient [68]. Conversely, arm and face lotions had little effect on bacterial and archaeal diversity [69]. Finely chosen preservative systems for cosmetic formulas have been shown to preserve the homeostasis of the microbiome and even help to restore its equilibrium after dysbiosis [70]. In another study, the authors showed that although tested preservatives had an efficient antibacterial activity in vitro, the skin microbiome was not impacted in vivo [71]. Cosmetic serum-containing galactooligosaccharides (GOS) prebiotics increased the
Figure 1. (A) Taxonomic analysis of cheek microbiome in younger and older subject’s group. Stackedbar charts showing the relative abundance of the 20 most prevalent bacterial genera. Adaptedfrom [56] (B) Taxonomic analysis of cheek microbiome (younger group) in polluted and non-pollutedenvironment. Stacked bar charts showing the relative abundance of the 20 most prevalent bacterialgenera. Adapted from [57].
3.2. Variability Due to Extrinsic Factors
Skin microbiota modifications are also associated with extrinsic factors, such as ur-banization, exposure to antibiotics, air pollution, UV exposure, smoking, hygiene prod-ucts [57–64]. The set of factors to which an individual and the skin microbiota are exposedis called the exposome [65].
The modulation of cosmetic formulas on the microbiome is still under investigationand might be formula and skin site-specific. On one hand, some major components ofthe facial microbiome decrease after cosmetic use [66]. Furthermore, multiple studiessuggest a strong effect of antiperspirant and deodorant use on the bacterial composition ofarmpits [67]. On the other hand, formulas used in the cleansing axis had a strong impacton the microbiome, even though some abundant taxa have been shown to be resilient [68].Conversely, arm and face lotions had little effect on bacterial and archaeal diversity [69].Finely chosen preservative systems for cosmetic formulas have been shown to preserve thehomeostasis of the microbiome and even help to restore its equilibrium after dysbiosis [70].In another study, the authors showed that although tested preservatives had an efficientantibacterial activity in vitro, the skin microbiome was not impacted in vivo [71]. Cosmeticserum-containing galactooligosaccharides (GOS) prebiotics increased the Shannon indexdiversity post-treatment in the experimental group vs. control. S. aureus relative abundancedecreased. Changes in the Microbiome composition were associated with improvement invarious clinical parameters (e.g., water loss, wrinkles depth) [72].
The impact of microbiome modifications on skin health in such conditions is stillunclear, however, some recent evidence has been reported in the case of pollution usingmicrobiome and metabolomics data of a Chinese cohort [57]. The impact of pollution onthe Microbiome composition is resumed in Figure 1B. Comparing skin microbiota and skinmetabolome allowed the authors to highlight a strong link between sebum degradationand bacterial taxa (Cutibacterium spp. and Staphylococcus spp.) and revealed a potential link

Pathogens 2022, 11, 121 5 of 27
between these taxa and the modulation of metabolites such as carnitine, histamine, andphenyl-lactic acid (PLA) on the skin, which could represent new factors involved in thecommensal-host homeostasis [73].
3.3. Variability Associated with Skin Health and Disorders
The skin microbiome associated with skin conditions has been observed in manyskin disorders such as atopic dermatitis (increase in the proportion of S. epidermidis andS. aureus), psoriasis (increase in the proportion of S. aureus), acne (increase in the proportionof particular phylotypes of C. acnes), seborrheic dermatitis and dandruff (increase in theamount of Malassezia), vitiligo (decrease in microbial diversity and increase in the propor-tion of Firmicutes at the lesion level [74]. The association between the skin microbiomecomposition and skin health/conditions is resumed in Table 1.
Table 1. Major taxonomic modifications of the skin/scalp microbiome in skin diseases.
Skin Condition Microbiome Shift Reference
Atopic Dermatitis (AD)
Decreased microbial diversityIncrease in S. aureus associated with disease severity and
increase in S. epidermidis during flaresDecrease in C. acnes, lactobacilli, and Burkholderia spp.
Increase in M. sympodialis, (and secondly M. globosa, M.dermatis, M. restricta)
[75–81]
Psoriasis
Increase in C. simulans, C. kroppenstedtii, Finegoldia spp.,and Neisseriaceae spp.
Decrease in C. acnes, lactobacilli, and Burkholderia spp.No difference in the amount of Malassezia between
lesion area of psoriasis and healthy skin (measured byPCR) but there is more diversity of Malassezia species inpatients with psoriasis compared to healthy individuals
Increase in Brevibacterium, Kocuria palustris, Gordonia,and increase in M. restricta (back) and M. sympodialis
(elbow) increased in psoriatic lesions.
[76,79,82,83]
Acne
Increase in the proportion of C. acnes strains presentingwith virulence factors
Increase in S. epidermidis (secondary to C. acnes)Malassezia spp. could be involved in the development of
acne
[46,84–88]
Rosacea Increased in Demodex mites on the skin. [89,90]
Vitiligo Decrease in bacterial diversity [74]
Seborrheic dermatitis (SD) and Dandruff
Increase in S. epidermidis and decrease in C. acnes.Increase in the population of Malassezia restricta and
Malassezia globosa on the scalp.Malassezia spp. metabolize and oxidize sebum-derived
lipids into inflammatory compounds and produceindole derivatives (malassezin, indolocarbazole) which
may impact skin inflammation through arylhydrocarbon receptors.
[88,91–94]
The features of the skin microbiome in common inflammatory skin diseases have beenreviewed earlier [95]. Dysbiosis of the Microbiome is observed in many skin conditionssuch as atopic dermatitis, acne, seborrheic dermatitis and dandruff (increased amountof Malassezia), vitiligo (decrease in microbial diversity and increase in the proportion ofFirmicutes in the lesions) [74]. Minor species represent 5 to 20% of bacteria and fungiand up to 1000 different species have been identified. Their proportion and diversity varyaccording to the anatomical zones, the type of skin, and various factors (i.e., environmental,age, lifestyle hygiene). Their precise role remains to be elucidated [96].

Pathogens 2022, 11, 121 6 of 27
3.3.1. Role of S. epidermidis
In Atopic dermatitis, a dysbiosis of the skin microbiome is observed with an increase inthe proportion of S. aureus, and S. epidermidis during the atopic crisis [75,97,98]. S. aureus isconsidered as an opportunistic pathogen, as various virulence factors have been described(formation of biofilms, toxins, Phenol-soluble modulins (PSMs), proteases) in associa-tion with skin disorders [99–101]. It is a coagulase-positive bacteria, unlike the majorityof Staphylococci found on the skin, which are coagulase-negative Staphylococcus (CoNS:Coagulase-negative Staphylococci) [84].
Initially, the increase in S. epidermidis was described as a defense against S. aureusproliferation (via Esp protease which inhibits biofilm formation by S. aureus [102] andvia secretion of lantibiotics [103]), but new data on S. epidermidis/skin interaction in amouse model suggests that S. epidermidis may have a pro-inflammatory role and alterthe skin barrier in the context of atopic dermatitis and Netherton syndrome [104,105]. Ithas also been shown that excessive colonization of the skin by S. epidermidis could leadto an alteration of the cohesion of the epidermis following the lysis of desmosomes bythe bacterial proteases EcpA (Extracellular cysteine protease A) [104]. Further studies arerequired to unambiguously demonstrate the role of S. epidermidis in atopic dermatitis.
3.3.2. Role of C. acnes
The major microorganism of the pilosebaceous unit is C. acnes, representing up to 90%of the microbiome in sebum-rich sites such as the scalp, face, chest, and back. Numerouspaths have been proposed by which C. acnes exacerbates acne, including augmentationof lipogenesis, comedone formation, and host inflammation [106]. C. acnes promotescomedogenesis by generating oxidized squalene and free fatty acids, leading to a qualitativechange in sebum [107,108]. Moreover, C. acnes activates the IGF-1/IGF-1 receptor signalingpathway to upregulate filaggrin expression, which in turn increases integrin-α3, -α6, andvβ6 levels, thereby affecting keratinocyte proliferation and differentiation and resulting incomedone formation [109,110]. It has been shown to trigger sebum accumulation whenapplied to hamster auricles [111]. In addition, C. acnes induces and aggravates inflammation,by activating Toll-like receptors (TLR-2 and TLR-4) on keratinocytes, which leads to theactivation of the MAPK and NF-kB pathways [112,113]. Additionally, metabolites suchas porphyrins produced by C. acnes, are pro-inflammatory and are associated with acnedisorders [114]. Strain-level differences in porphyrin production and regulation in C. acneselucidate disorder associations [115]. It is suggested that indole compounds produced byMalassezia spp. downregulate the inflammatory response, thereby helping to establish theassociated pathology Pityriasis versicolor [116].
3.3.3. Role of Other Microorganisms
C. acnes is not the only microbial player in acne. Intriguingly, according to data fromBarnard et al. and Park et al., the abundance of the genus Cutibacterium is slightly higheron healthy skin compared to acne-affected skin [117,118]. Compared to healthy controls, anincrease of Cutibacterium granulosum, S. epidermidis, Proteobacteria, Firmicutes, Streptococcus(pre-adolescent) and Malassezia species associated with a decrease in Actinobacteria hasbeen also described. An increase in Bacteroides in the gut Microbiome has been alsoobserved [106]. In comedones and pustules, C. granulosum known for its higher lipaseactivity is highly abundant [109]. This high lipase activity (100 times higher than C. acnes)is also the major driver for the implication of M. restricta in acne pathology in young adultswith refractory acne [106]. In summary, the direct involvement of the microorganisms ortheir interaction with C. acnes still needs to be elucidated. Dissection of the role of C. acnes inacne is further complicated by recent findings that suggest that specific C. acnes phylotypescould play a major role in acne aggravation, while others could be beneficial for the skin.
In Psoriasis, Microbiome is affected. However, no clear conclusion is established thatconnects the Microbiome diversity to the disease status. Different studies show that there iseither an increase or a decrease or no change in the microbiome diversity [95]. Nevertheless,

Pathogens 2022, 11, 121 7 of 27
there is evidence that shows a higher S. aureus and a lower S. epidermis abundance inpsoriatic lesions [119,120].
More importantly than the connection between microbiome composition and pso-riasis or any other skin condition, it would be of interest to gain a better understandingof the functional relation between the Microbiome and these different skin conditions.Nonetheless, Microbiome modulation could be an attractive therapeutic approach. Thus,ingredients/products that modulate beneficially the Microbiome could help to improvethe clinical signs associated with each skin condition and is well covered in the reviewby Polak et al. [121]. Many topical or oral-formulas show improvement of atopic andacneic skin conditions after their application/ingestion. Topical formulas modulate theskin Microbiome mainly via their antibacterial activity against C. acnes and/or S. aureus thetwo major implicated microorganisms [121–123].
Different confounding factors could likely explain these controversial conclusionsabout Microbiome diversity among the different studies trying to establish a Microbiomesignature in each skin condition (cosmetic routines, exposome, skin site . . . ) but one of themost important factors explaining these disputed results could be the diverse approachesfor skin Microbiome analysis from wet lab to bioinformatics.
The study design for skin microbiome research is multifaceted and integral to alldownstream steps. Many published studies examined the biases introduced by the skinsampling methods and sample storage, controls and contamination sources, sequencingbiases, and possible quantitation [124–129]. The complexity of skin microbiome studies issummarized by Kong et al. [130].
Adapted methodology strategy dedicated to the skin microbiome and its metaboliteanalysis is key for a successful translation into microbiome-based concepts. Standardizationand robustness of the tools are of great importance, in addition to the use of a collection ofcutting-edge technologies which aim to reduce the bias (mock communities, primers choiceadapted for skin microbiome analysis, distinguishing between dead/alive bacteria for accurateestimation of diversity, platform methods and metadata validation/standardization . . . ).
4. Methods in Skin Microbiome Exploration
Over the last two decades, the development of the so-called Next Generation Se-quencing (NGS) technologies and their continuously decreasing costs have allowed thegeneration of a huge amount of metagenomic data. Today, these data represent a stronglever for microbiota characterization, whether in healthy conditions or disorders, includinginfectious diseases, cancers, and other different clinical applications [131–136]. In dermatol-ogy, for example, multi-omics have increased the knowledge of many skin disorders, suchas psoriasis, atopic dermatitis, or acne [76].
4.1. Perspectives for Skin Microbiome Metabolomics Analysis
An emerging “omics” tool to understand the function of the human microbiome ismetabolomics (i.e., the analysis of small molecules including sugars, amino acids, lipids,and nucleotides, in a system) [137]. In gut microbiome studies, metabolomic tools havebeen extensively applied to characterize microbial metabolites and to determine theirrole in mediating host-microbe crosstalk [138,139]. Particularly, untargeted metabolomics(i.e., metabolomic profiling to collect a chemical inventory from a sample) has led to thediscovery of many microbiome-derived biomarkers of host diseases, such as inflammatorybowel syndrome, Crohn’s disease, diabetes, and non-alcoholic fatty liver disease [140–143].These metabolomic applications have led to the identification of important bacterial-basedtargets for disease diagnosis and treatment, inspiring the use of metabolomic approachesto characterize the molecules associated with the skin microbiome and to explore their rolein influencing skin physiology [144].
In dermo-cosmetics, understanding the relationships between the chemistries involvedin host-microbiota-environment crosstalk is crucial for the development of innovativemicrobiome-based solutions to treat skin conditions. The development of new sampling

Pathogens 2022, 11, 121 8 of 27
methods for the collection of skin samples compatible with untargeted metabolomic work-flows has greatly improved the characterization of the chemical composition of the humanskin [144–147]. Although the identification of microbial metabolites from skin samplesremains challenging, mainly due to the underrepresentation of reference spectra collectedfrom microorganisms in public libraries, integrating metabolomics and microbiome datahas improved our ability to identify microbe-metabolite associations. Since its developmentin 2015, the 3D cartography approach has become a powerful tool to visualize and inte-grate large-scale microbiome and metabolomics data from the skin surface and to rapidlyscreen for omic spatial distributions associated with different phenotypes, through spatialcorrelations between molecular and bacterial distributions [147]. This tool has also enabledthe characterization of antimicrobial peptides (Human neutrophil Peptides—HNP-1 and2) that co-localize with bacteria including Provotella and Clostridium in the groin area, aswell as products of bacterial processing on the skin, such as free fatty acids resulting fromhydrolysis of triacylglycerides mediated by Cutibacterium. As the tool continues to evolve,future applications to skin conditions, such as atopic dermatitis and psoriasis, may revealbiogeographical microbe-metabolite correlations differentiating healthy skin from diseasedskin or lesional vs. non-lesional sites.
Recent advances in bioinformatics and computational pipelines have greatly facilitatedthe visualization and annotation of large-scale metabolomic data. Molecular networking,initially introduced in 2012, has become a powerful bioinformatics tool to organize, mine,and compare spectral data, as well as to connect related molecules by their spectral similar-ities [148]. This tool is part of an online infrastructure—GNPS (Global Natural ProductsSocial Molecular Networking http://gnps.ucsd.edu, accessed on 25 October 2021) andenables the processing of large-scale metabolomics data and molecular annotation throughthe interrogation of reference spectral libraries [149]. Furthermore, multivariate analysisand machine learning tools have recently been implemented in GNPS to compare molecu-lar profiles collected from different groups/types of samples [150]. Beyond its extensiveapplication in natural product discovery, clinical and environmental studies, applicationsin skin studies included the identification of molecular signatures associated with individ-ual lifestyles and further linked them to the built environment, investigating the impactof personal care products on the skin metabolomics and microbiome dynamics or evenmonitoring drugs and their metabolites directly on human skin [69,151–158]. Molecularnetworking has also proven to efficiently assign the origin of molecules detected on humanskin (skin cells, microbes, environment, and lifestyles) [147,156,159]. Since the developmentof molecular networking, several complementary bioinformatic tools have been developedto enhance quantitative metabolomics analysis, expand molecular annotations through achemical tree-based approach and in silico annotation tools, as well as to re-analyze publicmetabolomics data using formatted metadata to compare metabolites between groups ofsamples from different datasets [160–163].
Additional bioinformatic advances have improved functional interrogations of thehuman microbiome including genome mining tools, such as antiSMASH, SMURF, andPRISM that has greatly facilitated the prediction of biosynthetic gene clusters (BGCs). Fur-ther development of computational algorithms applied to metagenomic sequencing data ofthe human microbiome has led to the discovery of small molecule-producing BGCs, suchas Type II Polyketides that appear to be widely encoded in the oral, gut, and skin micro-biome [164–167]. Advances in algorithms that pair metagenomic and metabolomic datasets,such as NRPquest, NRPminer, and MetaMiner, have allowed more accurate identificationof bioactive metabolites produced by BGCs from different environments [168–170]. Exam-ples include the discovery of known and unknown post-translationally modified peptides(RiPPs) from lichen and human microbiomes, and the identification of Non-RibosomalPeptides (NRPs), such as lugdunin from Staphylococcus skin isolates and surugamides fromsoil datasets [168,169].

Pathogens 2022, 11, 121 9 of 27
4.2. Expectations from Metagenomic and Data Science
Metagenomics is of particular interest for the dermo-cosmetic industry, with themicrobiome representing a real natural reservoir for the discovery of new molecules ofinterest [171–173].
Recently, Liu et al. described a screening approach to analyze over 3000 humanskin isolates to evaluate bacterial competition within the human skin microbiota [174].The authors demonstrated that bacteriocin micrococcin P1 (MP1) from the Staphylococcushominis strain led to reduced Staphylococcus aureus infection and accelerated closure ofS. aureus-infected wounds, meaning that MP1 can be proposed as a candidate to developa new approach against S. aureus infections. More interestingly, their results show that,beyond the generation of metagenomic experiments to characterize microbial communitiesat the taxa level, it is also important to model the whole ecosystem of the microbiota better,including microbes-microbes but also host microbes’ interactions. The latter presents systembiology approaches, largely described as being the future of metagenomics and microbiomemodeling for biomarker discovery and new pharmacology applications [12,174,175].
4.3. Meta-Omics and System Biology Approach
In this field, challenges in bioinformatics and data science remain to develop newomics data integration and analytical strategies for microbiome analysis [176]. This taskis not easy, as an important amount of omics data can now be simply generated for bothhumans and microbes on different omics layers (e.g., genomics, transcriptomics, proteomics,and metabolomics). Researchers deal more and more with meta-omics approaches to assessthe potential functions encoded by microbial communities and quantify the metabolicactivities occurring within a complex microbiome.
In 2019, Zhang et al. discussed that functional meta-omics approaches could be oneof the most promising strategies to facilitate the identification of microbes’ metabolicpathways specific to clinical phenotypes [177]. In a second step, the authors suggestedthat these metabolic pathways could be modulated through either supplementation ofbeneficial species, engineered probiotics/commensals, prebiotics, bacteriophages, or highlyselective drugs to shift microbiome metabolic profiles closer to healthy conditions.
4.4. Network-Based Models’ Approach
Compared to the meta-omics approaches described above, which can be consideredas data-driven approaches, network-based models represent another category of systembiology approach for microbiome analysis. Network-based models are mainly representedby genome-scale metabolic models (GEMs), which have been the subject of many technicalpublications in recent years [178–183]. GEMs aim to reconstruct biological networks byassembling several biological pathways created using experimental data [184,185]. Areconstructed network describes gene-protein-reaction associations for all metabolic genesin an organism. Even if the reconstructed network approach was firstly described onthe level of a bacterium, the same approach has been applied to humans and Recon is awell-known published human metabolic map [184,186–188].
When combined, the human metabolic network and microbe networks can provide aninteractomic map between microbe metabolites and the human proteome. As an example,Magnúsdóttir et al. reconstructed networks for 773 members of the gut microbiota [189].This resource, called AGORA, is compatible with the human metabolic network Recon, thusrepresenting a powerful strategy to facilitate the study of host-microbiome interactions.Although these network-based models were mainly published to study gut microbiota, avery recent review described how the same approach can be transposed to skin microbiomefunctional analysis [180,189,190]. In addition, this metabolic-network approach can providemajor advantages for precision medicine and personalized cosmetic purposes. Indeed,in 2017, Jens Nielsen explained that multi-omics data coming from clinical trials can betranslated to GEMs to identify “reporter” metabolites or “sub-networks” that could bespecific to a cluster of patients [179].

Pathogens 2022, 11, 121 10 of 27
Multi-omics data analysis using a reconstructed network thus appears as a stronglever to understand microbiome functions more thoroughly, including microbes-microbesand host microbes interactions as well as the mechanisms of bacterial colonization that canlead to skin-specific diseases or disorders.
Improvements in these approaches are still widely expected by the scientific commu-nity as they are promising tools for the discovery of new biomarkers, as well as for thedevelopment of future active ingredients for both dermatology and cosmetics.
However, the amount of work to establish a network on the microorganism scale prop-erly remains laborious [178,191]. To avoid such a challenge, another method has been de-veloped to predict the metabolome using metagenomic sequencing data only [144,192–195].More recently, Yin et al. published a comparative analysis of their tool with two othermethods published in this domain [196]. To evaluate the performance of the three differentalgorithms, 900 microbiome-metabolome pairs of samples from six different studies ofhuman diseases have been used for occurrence prediction (presence vs. absence) andmetabolites considered as differentially expressed. As these prediction tools only usemetagenomic data as input, they may be considered as a good compromise and a serious al-ternative solution to the generation and analysis of meta-omics and network reconstructionsthat are known to be more expensive and time-consuming strategies.
In conclusion, associating multi-omics technologies, statistical and computationalanalyses, and more advanced 3D skin models, will offer a promising opportunity toestablish a microbiome-related skin condition causality and subsequently orient towardthe cosmetic solution/ingredient.
Moreover, the microbiome is nowadays a significant target of personalized technolo-gies by presenting interesting solutions for different skin conditions, such as dryness, agingsigns, reactive or irritated skin [197,198]. Applied microbiome research offers a functionalalternative path towards new solutions that might possess other levels of performance.
5. Microbiome-Based Cosmetic Solutions and Technologies
Many of the skin conditions described above are multifactorial, however, the micro-biome is a key factor in skin disorders. The interplay between the microbiome and the skinis key for its homeostasis health. Intervening and finely modulating the microbiome tocorrect skin conditions described above is a rising field of research. These interventions aremainly realized by prebiotics, postbiotics, and probiotics, as well as microbiota transplant.The latter is still in its infancy phase for the skin. In cosmetic/dermatology applications, afocus concentrates on the first three paths.
The microbiome has been extensively studied and reported in the field of nutri-tion [199]. Although some definitions exist on the World Health Organization level, thereare currently no available international guidelines regarding the definitions or terminolo-gies applicable for cosmetic ingredients that work with the skin’s microbiome. Currentdefinitions consider probiotics to be living microorganisms that must be ingested in asufficient amount to have a positive effect on health that is not limited to the nutritionaleffects alone [200–202].
Prebiotics are a food ingredient that results in specific changes in the compositionand/or activity of the gastrointestinal microbiota, thus conferring benefit(s) upon the host’shealth [199].
Very recently, the International Scientific Association of Probiotics and Prebiotics(ISAPP) defined the scope of postbiotics as a “preparation of inanimate microorganismsand/or their components that confers a health benefit on the host”.
Postbiotics could be intentionally inactivated microbial cells with or without metabo-lites, or cell components that contribute to establishing host health benefits.
The gut is not the only site of action of postbiotics. They could also be administeredon a host surface, such as in the oral cavity or on the skin [203].
The topic of the cosmetic microbiome was taken up in 2018 by the International Co-operation on Cosmetics Regulation (ICCR), a voluntary international group of cosmetic

Pathogens 2022, 11, 121 11 of 27
regulatory authorities and cosmetic industry trade associations from Brazil, Canada, Chi-nese Taipei, the European Union, Japan, the Republic of Korea, and the United States.They considered that new technologies exploring the relationship between the humanmicrobiome and healthy skin were an area of increasing interest and the safety, quality,regulation, and potential development of international guidelines for products arising fromthese technologies would be a worthwhile topic for the ICCR.
In 2020 they developed a set of categories and descriptors that could be used togroup and categorize microbiome-related products, their ingredients, and other relevantapproaches, in a cosmetic/skin-relevant context [204].
These ingredients were divided into two main categories based on viability: viable(live or dormant)—encompassing only probiotics (based on biological origin), and non-viable ingredients. The non-viable ingredients were further divided into two sub-categories;prebiotics (by their intended action on the skin microbiota) and postbiotics (based on theirbiological origin) (Table 2).
Table 2. Resuming the cosmetic description of prebiotic and postbiotic ingredients.
Postbiotic (Including Probiotic Fraction or Extract) Prebiotic
Non-viable ingredients comprised of inactivatedmicroorganisms and/or soluble factors (products or metabolicby-products) released by live or inactivated microorganisms,
added to a cosmetic product to achieve a cosmetic benefit at theapplication site, either directly or via an effect on the existing
microbiota.Categories: 1/Ferments, lysates, extracts, filtrates, 2/Non-viable
microorganisms (inactivated/heat-killed), 3/Metabolicproducts/by-products (isolated)
Non-viable ingredients are added to a cosmetic product to beactively used as nutrients by the microbiota of the application
site to achieve a cosmetic benefit.Examples: ingredients such as fibers, sugars, minerals, but also
complex biological mixtures/extracts, etc.
Postbiotic products/ingredients belong to the non-viable category. Based on theirbiological origin, postbiotic ingredients (ferments, extracts, lysates, filtrates) share a com-mon description: “Non-viable ingredients comprised of inactivated microorganisms and/or solublefactors (products or metabolic by-products) released by live or inactivated microorganisms, added toa cosmetic product to achieve a cosmetic benefit at the application site, either directly or via an effecton the existing microbiota” [204].
In cosmetics, postbiotics may be an alternative to the use of live whole microorganismsin probiotic form. To summarize the product entries, postbiotic ingredients were dividedinto three types:
“Ferments, lysates, extracts, filtrates or any combination of these ingredients that are notliving but which have been obtained by means of probiotic bacteria (Bacillus, Bifidobac-terium, Lactobacillus, Lactococcus, Vitreoscilla, Streptococcus thermophilus, Leuconostoc)or fungi used primarily as fermentation facilitators (Saccharomyces, Candida bombicola,Kloeckera, Hansenula-Pichia, Aspergillus)”:
“Non-viable microorganisms (inactivated/heat-killed), mostly lactic-acid forming bacte-ria: Enterococcus faecalis, Lactobacillus (paracasei, casei, acidophilus), Lactococcus, orVitreoscilla filiform”.
“Metabolic products/by-products (isolated) including bacteriocin extract, ectoin, succinic acid,lactic acid, hydrolyzed yogurt protein, sodium hyaluronate, and milk proteins” [204].
5.1. Postbiotics
While both prebiotics and probiotics are either used alone or in combination as sym-biotics focus on beneficial bacteria directly, postbiotics focus on downstream benefits tothe host and can be categorized into 3 major categories: 1. Inactivated microbial cells aslysates, including cell wall components and the cytosol, 2. Exopolysaccharides, 3. Secretedmolecules, including peptides, proteins (enzymes), and small metabolites.

Pathogens 2022, 11, 121 12 of 27
Postbiotics, also referred to as functional ingredients, impart their benefits through amyriad of bioactivities that can include anti-inflammatory, immunomodulatory, antioxidant,anti-microbial, pro-differentiation, etc. It is important to note that these activities areeffectuated through important host-microbiome interactions.
Classically, postbiotics have mainly been studied in the context of the gut microbiome,especially for their applications in early life nutrition, wherein an important role for shortchain fatty acids (SCFAs) and lipopolysaccharides has been suggested for health benefits.With increasing knowledge of the gut-brain-skin axis, further benefits for the skin havebeen suggested, although this warrants further investigation. Recent literature suggeststhat topical application of postbiotics may confer direct benefits for the skin. Of these,microbial lysates are widely characterized and are widely used in cosmetic products:Bifidobacterium/Lactobacillus/Vitreoscilla filiformis (Vf ) [205–210].
5.1.1. Probiotic Fractions
Oral probiotics have a positive effect on skin health, via the gut and the systemic route,by modulating the immune system [211].
For example, in addition to the ability to modulate both intestinal mucosal and sys-temic immune functions, the probiotic strain Lactobacillus paracasei NCC2461 (ST11) con-ferred benefits to the skin to reinforce the skin barrier function and decrease skin sensitivityand dandruff conditions [210,212–214]. The probiotic Lactobacillus johnsonii La1 has beenshown to protect the skin defenses by maintaining the number and function of Langerhanscells after UV exposure to the skin, as well as regulating skin inflammation [207,208,215].In addition, probiotics have been shown to prevent and reduce the severity of atopicdermatitis, acne vulgaris, dry skin, prevent signs of photo-aging, and facilitate woundhealing [211,216,217]. However, some published data raises safety concerns about the useof living probiotics in some specific contexts. A few weeks-gestation infants with abnor-malities in their intestinal tract presented cases of bacteremia following Lactobacillus GGsupplementation [218]. More recently, it has been demonstrated that Intensive Care Unitspatients that were administered with Lactobacillus rhamnosus as probiotics are at higherrisk of developing bacteremia compared to those not receiving probiotics [219]. Thus, inthe case of wounded or altered skin, special safety attention should be considered whencosmetics containing living probiotics are applied on the skin of people who might becritically ill.
Non-viable microorganisms extend the scope of probiotic concepts, for example, heat-killed and tyndallized probiotic lactic acid bacteria and bifidobacteria have key probioticeffects in gastrointestinal diseases [220]. Consumption of lactic acid bacteria extracts wasfound to have activity comparable to the live forms [221].
Health benefits on the skin can also be achieved by the topical application of inacti-vated probiotics [222,223]. The majority of topical bacterial extracts are ferments, fermentlysates, or ferment lysate filtrates generated after the cultivation and harvest of a probi-otic microorganism.
Some bacterial extracts (Bacillus coagulans, L. johnsonii, L. casei, L. plantarum, andL. acidophilus) have antimicrobial properties that may support skin healing [224].
Lactobacillus acidophilus extract has been shown in vitro to scavenge reactive oxygenspecies following UVB-induced oxidative stress in keratinocytes [225].
Lactobacillus plantarum and L. salivarius lysates accelerated re-epithelisation by inducingkeratinocyte migration [226].
Lysates of L. rhamnosus improved the skin barrier function in a reconstructed humanepidermis and S. thermophilus extracts were able to increase ceramide production andimprove skin hydration [227,228].
Vitreoscilla filiformis extract, via TLR2 activation, reinforced innate immunity and bar-rier function leading to a reduction of symptoms linked to atopic dermatitis and seborrheicdermatitis [205,206,229].

Pathogens 2022, 11, 121 13 of 27
In reactive skin conditions, B. longum extract decreased skin sensitivity and improvedresistance to physical aggression [209].
Topical inactivated probiotic fractions have also demonstrated efficacy in clinical trialsin symptoms linked to acne, atopic dermatitis, and rosacea.
5.1.2. Effector Molecules/Metabolites
The harmony of the microbiome ecosystem of the different human body sites isfirmly associated with the molecules that they produce [230]. The human microbiomemicroorganisms produce metabolites and molecules acting on a diverse set of targetsthat can modulate many physiological functions in the host, among them the immuneresponses. Several acts as antibacterials, but many other products have unknown targetsand effects on other commensals and the host. These molecules, playing as mediators ofthe microorganisms’ and microbes-host interactions, can have a non-local impact on tissuesor organs where the microbiome is not established. This is the case for the gut microbiome-derived metabolites that could reach the bloodstream and modulate the homeostasisof the skin and its associated conditions. However, the skin differs from the gut in itsphysicochemical properties. It is a dry, acidic, lipid-rich, high-salt environment withoutexogenous nutrient sources, and therefore has low microbial biomass [9,231]. Microbespresent on the skin—collectively referred to as the skin microbiota—are central to skinphysiology and immunity. They produce a very rich array of metabolites/molecules.
Regardless of the origin of these postbiotic molecules, their beneficial effect on theskin is mediated by two major mechanisms:
5.1.3. Modulation of the Skin Microbiome
Topical application of lactic acid-producing bacteria (LAB) lysates, in addition totheir direct positive impact on the skin (anti-inflammatory, keratinocyte proliferation re-epithelization), has a protective impact against infection by pathogen bacteria S. aureusand S. pyogenes. Secretion of lactic acid was also proposed as the mediator of the bene-ficial effect on the skin (moisture retention of the skin) induced by the augmentation ofS. epidermidis [232].
Another large family of postbiotic molecules, known for their protective property ofthe skin by restraining pathogenic bacterial infection, are bacteriocins [233].
Alpha-soluble modulins produced by S. epidermidis and free fatty acids (i.e., sapi-enic acid), which are the degradation products of lipids by bacteria, have antibacterialactivity against S. aureus and various Gram(+) bacteria [234,235]. S. epidermidis, isolatedfrom normal skin, unlike those from patients with AD, had antimicrobial activity againstS. aureus via the secretion of antimicrobial peptides (AMPs). In vitro, the production ofthese peptides, namely lantibiotics such as epidermine and Pep-5, is dependent on the ori-gin of the strain and inhibits S. aureus in a specific manner in synergy with LL-37 secretedby the keratinocytes [103]. S. epidermidis produces AMPs and PSMs which specificallyinhibit the growth of S. aureus and group A streptococci [217]. S. epidermidis is also able todegrade S. aureus biofilms using the serine protease EspA in vitro [102]. The synergisticand specific action of human AMP LL37 and unknown AMPs produced by coagulase-negative S. epidermidis and S. hominis has specific antibacterial activity against the pathogenS. aureus involved in atopy [236]. In a Toll-like receptor (TLR)-3-dependent healing mecha-nism, S. epidermidis lipoteichoic acid and the lipopeptide LP78 reduced the inflammatoryresponse to improve wound in a mouse model of skin injury. Additionally, some strains ofS. epidermidis can diminish S. aureus–induced neutrophil recruitment and pro-inflammatorycytokine [18,237,238].
Transplantation of a mixture of C. acnes was successful after a one-week interventionperiod. Strains SLST types H1 + A1 + D1 did not show any adverse effect on the skinand the clinical relevance was not addressed in the study from [239]. C. acnes is capa-ble of transforming glycerol into short-chain fatty acids, such as propionic acid and itsderivatives which, on the contrary, inhibit the growth of USA300, a methicillin-resistant S.

Pathogens 2022, 11, 121 14 of 27
aureus [240,241]. Propionic acid finally helps to maintain pH value, which inhibits coloniza-tion by pathogenic microbes such as S. aureus [242]. C. acnes is known to produce specificallyan enzyme called RoxP that possesses antioxidant activity and plays an important role inmaintaining redox homeostasis on human skin [243].
Microbe-microbe interactions play a major role in the microbial ecosystem equilibriumand thus skin homeostasis. S. epidermidis limits the proliferation of C. acnes via the secre-tion of succinic acid and regulates the inflammation induced by C. acnes in the contextof acne [244,245]. C. acnes, on the other hand, produce propionate, isobutyrate, and iso-valerate which inhibit the formation of biofilms by S. epidermidis and increase its sensitivityto antibiotics [246]. In addition, C. acnes produces bacteriocins against S. epidermidisin vitro [247].
5.1.4. Cross Talk with the Immune Response
The skin microbiota can induce and activate T lymphocytes, both in the basal state andduring infection. This interaction with the immune system helps to control skin homeostasisand protect the immune system against pathogens such as Leishmania major and Candidaalbicans [248]. It is well known that cellular mediators produced by S. epidermidis modulatethe production of various cytokine IL17+CD8+T Cells [249]. LTA lipoteichoic acid is a majorcomponent of the cell wall of gram positive bacteria such as S. epidermidis and also a ligandfor TLR2. Although mechanism-dependent on the latter, it has been identified to have ananti-inflammatory effect on keratinocytes and to stimulate the production of keratinocytestem cell factor (SCF). It is also critical for mast cell differentiation [18,250]. TLR2 and theproduction of pro-inflammatory cytokines induce the recruitment and maturation of Tlymphocytes in CD4 + and CD8 which protect against skin infections and favor healing.Finally, the activation of TLR2 (especially via lipoteichoic acid) suppresses the inflammationmediated by TLR3 [15,251,252].
SCFAs from C. acnes modulate cytokine expression and may influence both the localpilosebaceous unit as well as the surrounding skin. These SCFAs inhibit the activity ofkeratinocyte histone deacetylase, an enzyme involved in epigenetic control, resultingin enhanced sensitivity to TLR activation and cytokine expression by keratinocytes andsebocytes [253]. Certain clinical strains of C. acnes induce the expression of hBD2 byNHEK keratinocytes via the activation of the TLR2 and TLR4 receptors and the culturesupernatant of C. acnes (ATCC6919) induced expression of hBD2 and LL-37 mRNAs bykeratinocytes (HaCat) [254,255]. Similar results have been published for different speciesof Malassezia [256,257].
Another bacterial metabolite family of interest in the cross-talk between bacteria andthe host is indolic metabolites. The tryptophan-derived Indole 3 aldehyde was shown toreduce inflammation by binding to the Aryl hydrocarbon receptor AhR [258].
In many cases, metabolite-host interaction is modulated by the immune system. Thisis the case via the MAIT cell system, which is known to recognize riboflavin (B2) interme-diates which are metabolites produced specifically by the major microorganism generacomponents of the skin [259]. Therefore, MAIT cell-specific immunity through recogni-tion of riboflavin biosynthesis intermediates is likely to play an important role in skinbarrier health.
In addition, some skin bacterial metabolites may trigger specific actions on path-ways that do not trigger inflammation. This is the case of the nucleobase analog 6-N-hydroxyaminopurine (6-HAP) which is produced by S. epidermidis and is known to haveselective antiproliferative action against tumor cell lines in vitro [22].
Microbial effector metabolites also play a role in wound healing. Traditionally, treat-ment of wounds, ulcers, and burns has been paralleled with antibiotic treatment. However,in recent years, a paradigm shift has taken place following studies demonstrating betterwound healing in the presence of bacteria [260]. Pathogenic bacteria are thought to delayhealing [261,262] while interactions between the commensal microbiota and the skin canaid wound healing by regulating the immune response and promoting repair of the skin

Pathogens 2022, 11, 121 15 of 27
barrier [15,263]. For example, the interaction of lipoteichoic acid (LTA) from S. epidermidiswith TLR2 in keratinocytes inhibits inflammatory responses and therefore limits tissuedamage and promotes wound healing [18,264].
6. Future Implications/Outlook
Postbiotics, including probiotic fractions and effector molecules, are solutions alreadyused in the dermo-cosmetic field; however, the ambition for the cosmetic industry is to addlive probiotics to cosmetic formulas with the expectation of potentially higher performancethat would probably be driven by a dialogue between added living-microbes and host cells.
However, the use of probiotics as cosmetic ingredients raises many questions. From aformulation standpoint, the first challenge is to maintain these microorganisms alive in acosmetic formulation. Most cosmetics are water-based and pH neutral or slightly acidic,which can be considered as favorable conditions, but they also contain some ingredientsthat could affect probiotics ‘survival’: surfactants, chelating agents, glycols, preservatives,fragrance. Moreover, the preservation of cosmetic products from microbial contaminationand proliferation is a safety and regulatory requirement. Therefore, the challenge is to main-tain probiotics alive in cosmetic products while preventing the growth of microorganismsthat could adversely affect the health of the consumers. This can be achieved by differentmeans such as the encapsulation of probiotics, or the use of suitable packaging where theliving bacteria are kept separately and mixed with the formulation at the time of use.
From a regulatory standpoint, the ICCR report indicates that “There were no uniqueregulations governing cosmetic products or ingredients intended to work specifically withthe skin’s (or mucosal) commensal microbiome. Rather, such products are subject tothe applicable rules and regulations governing cosmetics in each respective jurisdiction,including those covering both safety and product representation (i.e., claims). Severaljurisdictions pointed out that while no distinct regulations exist specific to these productsthere are general quality standard requirements such as microbiological limits which applyto all cosmetic products, including those containing live or viable microorganisms” [204].
The microbiological limits for cosmetic products are given in an International Standard-ization Organization (ISO) standard [265]. This document states that although cosmeticsare not required to be sterile, microorganisms present in a product should not cause anadverse effect on consumer safety or product quality. Therefore, the manufacturer mustrespect the Good Manufacturing Practices and take the necessary precautions to limit theintroduction of microorganisms from raw materials, processing, and packaging.
In this standard, microorganisms are considered as contaminants that are unintention-ally introduced in the cosmetic product and microbiological limits are established to ensureproduct quality and consumer safety. Therefore, those limits should not apply to probioticswhich are well-characterized microorganisms intentionally introduced in cosmetic productsto achieve a cosmetic benefit. However, discussions are still ongoing, and some clarificationis needed to allow the use of living bacteria as cosmetic ingredients.
The use of live probiotics is one of the major future applications in the dermo-cosmeticindustry but not the only one. Each human has his or her own ‘microbial fingerprint’ thatis specific to his or her skin and this specific microbiome may influence its homeostasis.The microbiome is the path to an individualized skincare routine.
In this perspective, personalized microbiome-derived cosmetic solutions that wouldintervene specifically are the future paradigm for safe, effective, and successful skin/scalpcare products.
7. Conclusions
Research is at the dawn of a «new generation» cosmetic that will use the skin’s mi-crobiome to provide lasting products with new efficient performance. To bring to lightthis rising cosmetics category that harnesses the potential of the cutaneous microbiome,it is essential to dissect the dynamic interactions existing between microorganisms andthe interplay host/Microbiome. Researchers would also need to understand the regula-

Pathogens 2022, 11, 121 16 of 27
tory/safety framework to translate these innovations (Figure 2). However, only rigorousand unbiased experimental approaches considering the specificity of the skin-microbiomeenvironment can be applied. This discovery will be made possible by coupling multi-omicstechnologies, statistical data mining, and representative 3D skin models. These approachesmay provide the opportunity to establish microbiome/skin condition causality and, sub-sequently, cosmetic solutions. The consideration of subtle regulatory environments andcountry-specificities will also be of high concern.
Pathogens 2022, 11, x 17 of 28
Figure 2. Resuming the best strategy for a successful translation of Microbiome-based concepts into cosmetic products of the future. Combined approaches of multi-omics technologies, powerful data mining tools, and representative 3D vitro skin models associated with standardized and unbiased experimental approaches dedicated to skin Microbiome analysis are key. Harnessing recent scientific breakthroughs and deciphering the famous causality question allied to better characterization of the interaction between the microbiome, the immune system and skin cells in various skin conditions would accelerate the translation. Finally, consideration of regulatory and safety aspects related to these new/targeted Microbiome-derived technologies (postbiotics, phages, probiotics…) and how to leverage their performance in different formulation types is essential.
Author Contributions: Conceptualization, A.G., O.P., C.C., T.C. and A.K.; writing—original draft preparation, A.G., O.P., A.B., L.L., N.M., S.C., C.C., T.C. and A.K.; writing—review and editing, A.G., O.P., A.B., L.L., N.M., S.C., L.A., C.C., T.C. and A.K.; project administration, A.K. All authors have read and agreed to the published version of the manuscript.
Funding: This research received no external funding.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest: The authors have the following interests: A.G., O.P., A.B., L.L., N.M., S.C., L.A., C.C., T.C. and A.K. are employees of L’Oréal R&I. This article is purely for knowledge diffusion and authors agree on adhering with journal’s policy on sharing data and materials.
References 1. Blaser, M.J.; Cardon, Z.G.; Cho, M.K.; Dangl, J.L.; Donohue, T.J.; Green, J.L.; Knight, R.; Maxon, M.E.; Northen, T.R.; Pollard,
K.S.; et al. Toward a Predictive Understanding of Earth’s Microbiomes to Address 21st Century Challenges. mBio 2016, 7, e00714-16. https://doi.org/10.1128/mbio.00714-16.
2. Brody, H. The gut microbiome. Nature 2020, 577, S5. 3. Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–
506. https://doi.org/10.1038/s41422-020-0332-7.
Figure 2. Resuming the best strategy for a successful translation of Microbiome-based concepts intocosmetic products of the future. Combined approaches of multi-omics technologies, powerful datamining tools, and representative 3D vitro skin models associated with standardized and unbiasedexperimental approaches dedicated to skin Microbiome analysis are key. Harnessing recent scientificbreakthroughs and deciphering the famous causality question allied to better characterization of theinteraction between the microbiome, the immune system and skin cells in various skin conditionswould accelerate the translation. Finally, consideration of regulatory and safety aspects related tothese new/targeted Microbiome-derived technologies (postbiotics, phages, probiotics . . . ) and howto leverage their performance in different formulation types is essential.
Author Contributions: Conceptualization, A.G., O.P., C.C., T.C. and A.K.; writing—original draftpreparation, A.G., O.P., A.B., L.L., N.M., S.C., C.C., T.C. and A.K.; writing—review and editing, A.G.,O.P., A.B., L.L., N.M., S.C., L.A., C.C., T.C. and A.K.; project administration, A.K. All authors haveread and agreed to the published version of the manuscript.
Funding: This research received no external funding.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: No new data were created or analyzed in this study. Data sharing isnot applicable to this article.
Conflicts of Interest: The authors have the following interests: A.G., O.P., A.B., L.L., N.M., S.C., L.A.,C.C., T.C. and A.K. are employees of L’Oréal R&I. This article is purely for knowledge diffusion andauthors agree on adhering with journal’s policy on sharing data and materials.

Pathogens 2022, 11, 121 17 of 27
References1. Blaser, M.J.; Cardon, Z.G.; Cho, M.K.; Dangl, J.L.; Donohue, T.J.; Green, J.L.; Knight, R.; Maxon, M.E.; Northen, T.R.; Pollard, K.S.;
et al. Toward a Predictive Understanding of Earth’s Microbiomes to Address 21st Century Challenges. mBio 2016, 7, e00714-16.[CrossRef] [PubMed]
2. Brody, H. The gut microbiome. Nature 2020, 577, S5. [CrossRef]3. Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506.
[CrossRef] [PubMed]4. Grice, E.A. The skin microbiome: Potential for novel diagnostic and therapeutic approaches to cutaneous disease. Semin. Cutan.
Med. Surg. 2014, 33, 98–103. [CrossRef]5. Gallo, R.L. Human Skin Is the Largest Epithelial Surface for Interaction with Microbes. J. Investig. Dermatol. 2017, 137, 1213–1214.
[CrossRef]6. Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; NISC Comparative Sequencing Program; Bouffard, G.G.;
Blakesley, R.W.; Murray, P.R.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324,1190–1192. [CrossRef] [PubMed]
7. Oh, J.; Byrd, A.L.; Park, M.; Kong, H.H.; Segre, J.A.; NISC Comparative Sequencing Program. Temporal Stability of the HumanSkin Microbiome. Cell 2016, 165, 854–866. [CrossRef]
8. Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [CrossRef]9. Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [CrossRef]10. Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Kong, H.H.; et al.
Topographic diversity of fungal and bacterial communities in human skin. Nature 2013, 498, 367–370. [CrossRef]11. Cogen, A.L.; Nizet, V.; Gallo, R.L. Skin microbiota: A source of disease or defence? Br. J. Dermatol. 2008, 158, 442–455. [CrossRef]
[PubMed]12. Chen, Y.E.; Tsao, H. The skin microbiome: Current perspectives and future challenges. J. Am. Acad. Dermatol. 2013, 69, 143–155.e3.
[CrossRef] [PubMed]13. Ohnemus, U.; Kohrmeyer, K.; Houdek, P.; Rohde, H.; Wladykowski, E.; Vidal, S.; Horstkotte, M.A.; Aepfelbacher, M.; Kirschner, N.;
Behne, M.J.; et al. Regulation of Epidermal Tight-Junctions (TJ) during Infection with Exfoliative Toxin-Negative StaphylococcusStrains. J. Investig. Dermatol. 2008, 128, 906–916. [CrossRef]
14. Kuo, I.-H.; Carpenter-Mendini, A.; Yoshida, T.; McGirt, L.Y.; Ivanov, A.I.; Barnes, K.C.; Gallo, R.L.; Borkowski, A.W.; Yamasaki,K.; Leung, D.Y.; et al. Activation of Epidermal Toll-Like Receptor 2 Enhances Tight Junction Function: Implications for AtopicDermatitis and Skin Barrier Repair. J. Investig. Dermatol. 2013, 133, 988–998. [CrossRef] [PubMed]
15. Linehan, J.L.; Harrison, O.R.; Han, S.-J.; Byrd, A.L.; Vujkovic-Cvijin, I.; Villarino, A.V.; Sen, S.K.; Shaik, J.; Smelkinson, M.;Tamoutounour, S.; et al. Non-classical Immunity Controls Microbiota Impact on Skin Immunity and Tissue Repair. Cell 2018, 172,784–796.e18. [CrossRef]
16. Cogen, A.L.; Yamasaki, K.; Muto, J.; Sanchez, K.M.; Crotty Alexander, L.; Tanios, J.; Lai, Y.; Kim, J.E.; Nizet, V.; Gallo, R.L.Staphylococcus epidermidis antimicrobial delta-toxin (phenol-soluble modulin-gamma) cooperates with host antimicrobialpeptides to kill group A Streptococcus. PLoS ONE 2010, 5, e8557. [CrossRef]
17. Lai, Y.; Cogen, A.L.; Radek, K.A.; Park, H.J.; Macleod, D.T.; Leichtle, A.; Ryan, A.F.; di Nardo, A.; Gallo, R.L. Activation of TLR2by a small molecule produced by Staphylococcus epidermidis increases antimicrobial defense against bacterial skin in-fections. J.Investig. Dermatol. 2010, 130, 2211–2221.
18. Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.-R.; Hooper, L.V.; Schmidt, R.R.; Von Aulock, S.;et al. Commensal bacteria regulate Toll-like receptor 3–dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382.[CrossRef]
19. Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.;et al. Compartmentalized Control of Skin Immunity by Resident Commensals. Science 2012, 337, 1115–1119. [CrossRef] [PubMed]
20. Pastar, I.; O’Neill, K.; Padula, L.; Head, C.R.; Burgess, J.L.; Chen, V.; Garcia, D.; Stojadinovic, O.; Hower, S.; Plano, G.V.; et al.Staphylococcus epidermidis Boosts Innate Immune Response by Activation of Gamma Delta T Cells and Induction of Perforin-2in Human Skin. Front. Immunol. 2020, 11, 2253. [CrossRef]
21. Scharschmidt, T.C.; Vasquez, K.S.; Truong, H.-A.; Gearty, S.V.; Pauli, M.L.; Nosbaum, A.; Gratz, I.K.; Otto, M.; Moon, J.J.; Liese, J.;et al. A Wave of Regulatory T Cells into Neonatal Skin Mediates Tolerance to Commensal Microbes. Immunity 2015, 43, 1011–1021.[CrossRef]
22. Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.-J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. Acommensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2018, 4, eaao4502. [CrossRef]
23. Christensen, G.J.M.; Brüggemann, H. Bacterial skin commensals and their role as host guardians. Benef. Microbes 2014, 5, 201–215.[CrossRef] [PubMed]
24. Claesen, J.; Spagnolo, J.B.; Ramos, S.F.; Kurita, K.L.; Byrd, A.L.; Aksenov, A.A.; Melnik, A.V.; Wong, W.R.; Wang, S.; Hernandez,R.D.; et al. A Cutibacterium acnes antibiotic modulates human skin microbiota composition in hair follicles. Sci. Transl. Med.2020, 12. [CrossRef] [PubMed]
25. Kannan, R.; Ng, M.J.M. Cutaneous lesions and vitamin B12 deficiency. Can. Fam. Physician 2008, 54, 529–532. [PubMed]

Pathogens 2022, 11, 121 18 of 27
26. Kang, D.; Shi, B.; Erfe, M.C.; Craft, N.; Li, H. Vitamin B12 modulates the transcriptome of the skin microbiota in acne path-ogenesis.Sci. Transl. Med. 2015, 7, 293ra103. [CrossRef] [PubMed]
27. Mahabadi, N.; Bhusal, A.; Banks, S.W. Riboflavin Deficiency; StatPearls Publishing: Treasure Island, FL, USA, 2020.28. Williams, J.D.; Jacobson, E.L.; Kim, H.; Kim, M.; Jacobson, M.K. Folate in Skin Cancer Prevention. Subcell. Biochem. 2011, 56,
181–197. [CrossRef]29. Hasoun, L.Z.; Bailey, S.W.; Outlaw, K.K.; Ayling, J.E. Effect of serum folate status on total folate and 5-methyltetrahydrofolate in
human skin. Am. J. Clin. Nutr. 2013, 98, 42–48. [CrossRef]30. Rembe, J.D.; Fromm-Dornieden, C.; Stuermer, E.K. Effects of Vitamin B Complex and Vitamin C on Human Skin Cells: Is the
Perceived Effect Measurable? Adv. Skin Wound Care 2018, 31, 225–233. [CrossRef]31. Ridaura, V.K.; Bouladoux, N.; Claesen, J.; Chen, Y.E.; Byrd, A.L.; Constantinides, M.G.; Merrill, E.D.; Tamoutounour, S.; Fischbach,
M.A.; Belkaid, Y. Contextual control of skin immunity and inflammation by Corynebacterium. J. Exp. Med. 2018, 215, 785–799.[CrossRef]
32. Sowada, J.; Schmalenberger, A.; Ebner, I.; Luch, A.; Tralau, T. Degradation of benzo[a]pyrene by bacterial isolates from humanskin. FEMS Microbiol. Ecol. 2014, 88, 129–139. [CrossRef] [PubMed]
33. Patra, V.; Byrne, S.N.; Wolf, P. The Skin Microbiome: Is It Affected by UV-induced Immune Suppression? Front. Microbiol. 2016, 7, 1235.[CrossRef]
34. Flores, G.E.; Caporaso, J.G.; Henley, J.B.; Rideout, J.R.; Domogala, D.; Chase, J.; Leff, J.W.; Vázquez-Baeza, Y.; Gonzalez, A.; Knight,R.; et al. Temporal variability is a personalized feature of the human microbiome. Genome Biol. 2014, 15, 531. [CrossRef] [PubMed]
35. Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecinska-Piróg, J.;Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and ExtrinsicFactors on Skin Microbiota. Microorganisms 2021, 9, 543. [CrossRef]
36. Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; Kong, H.H.; Segre, J.A.; NISC Comparative Sequencing Program. Biogeography andindividuality shape function in the human skin metagenome. Nature 2014, 514, 59–64. [CrossRef] [PubMed]
37. Sowada, J.; Lemoine, L.; Schön, K.; Hutzler, C.; Luch, A.; Tralau, T. Toxification of polycyclic aromatic hydrocarbons by commensalbacteria from human skin. Arch. Toxicol. 2017, 91, 2331–2341. [CrossRef]
38. Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes theacquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107,11971–11975. [CrossRef]
39. Zhu, T.; Liu, X.; Kong, F.-Q.; Duan, Y.-Y.; Yee, A.L.; Kim, M.; Galzote, C.; Gilbert, J.A.; Quan, Z.-X. Age and Mothers: PotentInfluences of Children’s Skin Microbiota. J. Investig. Dermatol. 2019, 139, 2497–2505.e6. [CrossRef]
40. Tollanes, M.C.; Moster, D.; Daltveit, A.K.; Irgens, L.M. Cesarean section and risk of severe childhood asthma: A popula-tion-basedcohort study. J. Pediatr. 2008, 153, 112–116. [CrossRef]
41. Renz-Polster, H.; David, M.R.; Buist, A.S.; Vollmer, W.M.; O’Connor, E.A.; Frazier, E.A.; Wall, M.A. Caesarean section deliveryand the risk of allergic disorders in childhood. Clin. Exp. Allergy 2005, 35, 1466–1472. [CrossRef]
42. Kennedy, E.A.; Connolly, J.; Hourihane, J.O.; Fallon, P.G.; McLean, W.H.I.; Murray, D.; Jo, J.-H.; Segre, J.A.; Kong, H.H.; Irvine,A.D. Skin microbiome before development of atopic dermatitis: Early colonization with commensal staphylococci at 2 months isassociated with a lower risk of atopic dermatitis at 1 year. J. Allergy Clin. Immunol. 2016, 139, 166–172. [CrossRef] [PubMed]
43. Loo, E.X.L.; Sim, J.Z.T.; Loy, S.L.; Goh, A.; Chan, Y.H.; Tan, K.H.; Yap, F.; Gluckman, P.D.; Godfrey, K.M.; van Bever, H.; et al.Associations between caesarean delivery and allergic outcomes: Results from the GUSTO study. Ann. Allergy Asthma Immunol.2017, 118, 636–638. [CrossRef] [PubMed]
44. Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the infant microbiome com-munitystructure and function across multiple body sites and in relation to mode of delivery. Nat. Med. 2017, 23, 314–326. [CrossRef]
45. Capone, K.A.; Dowd, S.E.; Stamatas, G.N.; Nikolovski, J. Diversity of the Human Skin Microbiome Early in Life. J. Investig.Dermatol. 2011, 131, 2026–2032. [CrossRef] [PubMed]
46. Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) andacne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32 (Suppl. 2), 5–14. [CrossRef]
47. Mourelatos, K.; Eady, E.A.; Cunliffe, W.J.; Clark, S.M.; Cove, J.H. Temporal changes in sebum excretion and propionibacterialcolonization in preadolescent children with and without acne. Br. J. Dermatol. 2007, 156, 22–31. [CrossRef] [PubMed]
48. Shi, B.; Bangayan, N.J.; Curd, E.; Taylor, P.A.; Gallo, R.L.; Leung, D.Y.M.; Li, H. The skin microbiome is different in pediatricversus adult atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 1233–1236. [CrossRef]
49. Jo, J.-H.; Deming, C.; Kennedy, E.A.; Conlan, S.; Polley, E.C.; Ng, W.-I.; Segre, J.A.; Kong, H.H.; NISC Comparative SequencingProgram. Diverse Human Skin Fungal Communities in Children Converge in Adulthood. J. Investig. Dermatol. 2016, 136,2356–2363. [CrossRef] [PubMed]
50. Jugé, R.; Rouaud-Tinguely, P.; Breugnot, J.; Servaes, K.; Grimaldi, C.; Roth, M.-P.; Coppin, H.; Closs, B. Shift in skin microbiota ofWestern European women across aging. J. Appl. Microbiol. 2018, 125, 907–916. [CrossRef]
51. Zhai, W.; Huang, Y.; Zhang, X.; Fei, W.; Chang, Y.; Cheng, S.; Zhou, Y.; Gao, J.; Tang, X.; Zhang, X.; et al. Profile of the skinmicrobiota in a healthy Chinese population. J. Dermatol. 2018, 45, 1289–1300. [CrossRef]
52. Kim, H.-J.; Kim, J.J.; Myeong, N.R.; Kim, T.; Kim, D.; An, S.; Kim, H.; Park, T.; Jang, S.I.; Yeon, J.H.; et al. Segregation of age-relatedskin microbiome characteristics by functionality. Sci. Rep. 2019, 9, 16748. [CrossRef]

Pathogens 2022, 11, 121 19 of 27
53. Wu, L.; Zeng, T.; Deligios, M.; Milanesi, L.; Langille, M.G.I.; Zinellu, A.; Rubino, S.; Carru, C.; Kelvin, D.J. Age-Related Variationof Bacterial and Fungal Communities in Different Body Habitats across the Young, Elderly, and Centenarians in Sardinia. mSphere2020, 5. [CrossRef]
54. Li, Z.; Bai, X.; Peng, T.; Yi, X.; Luo, L.; Yang, J.; Liu, J.; Wang, Y.; He, T.; Wang, X.; et al. New Insights into the Skin MicrobialCommunities and Skin Aging. Front. Microbiol. 2020, 11, 565549. [CrossRef]
55. Leyden, J.J.; McGiley, K.J.; Mills, O.H.; Kligman, A.M. Age-Related Changes in The Resident Bacterial Flora Of The Human Face.J. Investig. Dermatol. 1975, 65, 379–381. [CrossRef] [PubMed]
56. Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.;et al. Aging-related changes in the diversity of women’s skin microbiomes associated with oral bacteria. Sci. Rep. 2017, 7, 10567.[CrossRef] [PubMed]
57. Leung, M.H.Y.; Tong, X.; Bastien, P.; Guinot, F.; Tenenhaus, A.; Appenzeller, B.M.R.; Betts, R.J.; Mezzache, S.; Li, J.; Bourokba, N.;et al. Changes of the human skin microbiota upon chronic exposure to polycyclic aromatic hydrocarbon pollutants. Microbiome2020, 8, 100. [CrossRef]
58. Rosenthal, M.; Goldberg, D.; Aiello, A.; Larson, E.; Foxman, B. Skin microbiota: Microbial community structure and its potentialassociation with health and disease. Infect. Genet. Evol. 2011, 11, 839–848. [CrossRef]
59. Dimitriu, P.A.; Iker, B.; Malik, K.; Leung, H.; Mohn, W.W.; Hillebrand, G.G. New Insights into the Intrinsic and Extrinsic FactorsThat Shape the Human Skin Microbiome. mBio 2019, 10, e00839-19. [CrossRef]
60. Kim, H.-J.; Kim, H.; Kim, J.J.; Myeong, N.R.; Kim, T.; Park, T.; Kim, E.; Choi, J.-Y.; Lee, J.; An, S.; et al. Fragile skin microbiomes inmegacities are assembled by a predominantly niche-based process. Sci. Adv. 2018, 4, e1701581. [CrossRef]
61. Clemente, J.C.; Pehrsson, E.C.; Blaser, M.J.; Sandhu, K.; Gao, Z.; Wang, B.; Magris, M.; Hidalgo, G.; Contreras, M.; Noya-Alarcón,Ó.; et al. The microbiome of uncontacted Amerindians. Sci. Adv. 2015, 1, e1500183. [CrossRef] [PubMed]
62. Araviiskaia, E.; Berardesca, E.; Bieber, T.; Gontijo, G.; Viera, M.S.; Marrot, L.; Chuberre, B.; Dreno, B. The impact of airbornepollution on skin. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1496–1505. [CrossRef]
63. Faergemann, J.; Larkö, O. The effect of UV-light on human skin microorganisms. Acta Derm. Venereol. 1987, 67, 69–72. [PubMed]64. Burns, E.M.; Ahmed, H.; Isedeh, P.N.; Kohli, I.; Van Der Pol, W.; Shaheen, A.; Muzaffar, A.F.; Al-Sadek, C.; Foy, T.M.; Abdel-
gawwad, M.S.; et al. Ultraviolet radiation, both UVA and UVB, influences the composition of the skin microbiome. Exp. Dermatol.2018, 28, 136–141. [CrossRef]
65. Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161.[CrossRef] [PubMed]
66. Lee, H.J.; Jeong, S.E.; Lee, S.; Kim, S.; Han, H.; Jeon, C.O. Effects of cosmetics on the skin microbiome of facial cheeks withdifferent hydration levels. Microbiol. Open 2017, 7, e00557. [CrossRef]
67. Urban, J.; Fergus, D.J.; Savage, A.M.; Ehlers, M.; Menninger, H.L.; Dunn, R.R.; Horvath, J.E. The effect of habitual and ex-perimental antiperspirant and deodorant product use on the armpit microbiome. PeerJ 2016, 4, e1605. [CrossRef] [PubMed]
68. Two, A.M.; Nakatsuji, T.; Kotol, P.F.; Arvanitidou, E.; Du-Thumm, L.; Hata, T.R.; Gallo, R.L. The Cutaneous Microbiome andAspects of Skin Antimicrobial Defense System Resist Acute Treatment with Topical Skin Cleansers. J. Investig. Dermatol. 2016, 136,1950–1954. [CrossRef]
69. Bouslimani, A.; da Silva, R.; Kosciolek, T.; Janssen, S.; Callewaert, C.; Amir, A.; Dorrestein, K.; Melnik, A.V.; Zaramela, L.S.; Kim,J.-N.; et al. The impact of skin care products on skin chemistry and microbiome dynamics. BMC Biol. 2019, 17, 47. [CrossRef][PubMed]
70. Pinto, D.; Ciardiello, T.; Franzoni, M.; Pasini, F.; Giuliani, G.; Rinaldi, F. Effect of commonly used cosmetic preservatives on skinresident microflora dynamics. Sci. Rep. 2021, 11, 8695. [CrossRef]
71. Murphy, B.; Hoptroff, M.; Arnold, D.; Eccles, R.; Campbell-Lee, S. In-vivo impact of common cosmetic preservative systems infull formulation on the skin microbiome. PLoS ONE 2021, 16, e0254172. [CrossRef]
72. Hong, K.-B.; Hong, Y.H.; Jung, E.Y.; Jo, K.; Suh, H.J. Changes in the Diversity of Human Skin Microbiota to Cosmetic SerumContaining Prebiotics: Results from a Randomized Controlled Trial. J. Pers. Med. 2020, 10, 91. [CrossRef] [PubMed]
73. Misra, N.; Clavaud, C.; Guinot, F.; Bourokba, N.; Nouveau, S.; Mezzache, S.; Palazzi, P.; Appenzeller, B.M.R.; Tenenhaus, A.;Leung, M.H.Y.; et al. Multi-omics analysis to decipher the molecular link between chronic exposure to pollution and human skindysfunction. Sci. Rep. 2021, 11, 18302. [CrossRef] [PubMed]
74. Ganju, P.; Nagpal, S.; Mohammed, M.H.; Kumar, P.N.; Pandey, R.; Natarajan, V.T.; Mande, S.S.; Gokhale, R.S. Microbial communityprofiling shows dysbiosis in the lesional skin of Vitiligo subjects. Sci. Rep. 2016, 6, srep18761. [CrossRef]
75. Nakatsuji, T.; Gallo, R.L. The role of the skin microbiome in atopic dermatitis. Ann. Allergy, Asthma Immunol. 2018, 122, 263–269.[CrossRef]
76. Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-host interplay in atopic dermatitis and psoriasis. Nat. Commun. 2019, 10, 4703.[CrossRef] [PubMed]
77. Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Murray, P.R.;et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis.Genome Res. 2012, 22, 850–859. [CrossRef]

Pathogens 2022, 11, 121 20 of 27
78. Seite, S.; Flores, G.E.; Henley, J.B.; Martin, R.; Zelenkova, H.; Aguilar, L.; Fierer, N. Microbiome of affected and unaffected skin ofpatients with atopic dermatitis before and after emollient treatment. J. Drugs Dermatol. 2014, 13, 1365–1372.
79. Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia genus in skin and systemic diseases. Clin.Microbiol. Rev. 2012, 25, 106–141. [CrossRef] [PubMed]
80. Theelen, B.; Cafarchia, C.; Gaitanis, G.; Bassukas, I.D.; Boekhout, T.; Dawson, T.L., Jr. Malassezia ecology, pathophysiology, andtreatment. Med. Mycol. 2018, 56 (Suppl. 1), S10–S25. [CrossRef]
81. Nowicka, D.; Nawrot, U. Contribution of Malassezia spp. to the development of atopic dermatitis. Mycoses 2019, 62, 588–596.[CrossRef] [PubMed]
82. Amaya, M.; Tajima, M.; Okubo, Y.; Sugita, T.; Nishikawa, A.; Tsuboi, R. Molecular analysis of Malassezia microflora in the lesionalskin of psoriasis patients. J. Dermatol. 2007, 34, 619–624. [CrossRef]
83. Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al.Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-occurrence of Fungal and Bacterial Communities. Front. Microbiol. 2019, 10, 438.[CrossRef]
84. Claudel, J.P.; Auffret, N.; Leccia, M.T.; Poli, F.; Corvec, S.; Dreno, B. Staphylococcus epidermidis: A Potential New Player in thePhysiopathology of Acne? Dermatology 2019, 235, 287–294. [CrossRef]
85. Bek-Thomsen, M.; Lomholt, H.B.; Kilian, M. Acne is Not Associated with Yet-Uncultured Bacteria. J. Clin. Microbiol. 2008, 46,3355–3360. [CrossRef] [PubMed]
86. Fitz-Gibbon, S.; Tomida, S.; Chiu, B.H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Pro-pionibacterium acnes strain populations in the human skin microbiome associated with acne. J. Investig. Dermatol. 2013, 133,2152–2160. [CrossRef] [PubMed]
87. Akaza, N.; Akamatsu, H.; Numata, S.; Yamada, S.; Yagami, A.; Nakata, S.; Matsunaga, K. Microorganisms inhabiting fol-licularcontents of facial acne are not only Propionibacterium but also Malassezia spp. J. Dermatol. 2016, 43, 906–911. [CrossRef] [PubMed]
88. Xu, H.; Li, H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am. J. Clin. Dermatol. 2019, 20, 335–344. [CrossRef] [PubMed]89. Yamasaki, K.; Gallo, R.L. The molecular pathology of rosacea. J. Dermatol. Sci. 2009, 55, 77–81. [CrossRef] [PubMed]90. Murillo, N.; Aubert, J.; Raoult, D. Microbiota of Demodex mites from rosacea patients and controls. Microb. Pathog. 2014, 71–72,
37–40. [CrossRef]91. Clavaud, C.; Jourdain, R.; Bar-Hen, A.; Tichit, M.; Bouchier, C.; Pouradier, F.; el Rawadi, C.; Guillot, J.; Menard-Szczebara, F.;
Breton, L.; et al. Dandruff is associated with disequilibrium in the proportion of the major bacterial and fungal populationscolonizing the scalp. PLoS ONE 2013, 8, e58203. [CrossRef]
92. An, Q.; Sun, M.; Qi, R.-Q.; Zhang, L.; Zhai, J.-L.; Hong, Y.-X.; Song, B.; Chen, H.-D.; Gao, X.-H. High Staphylococcus epidermidisColonization and Impaired Permeability Barrier in Facial Seborrheic Dermatitis. Chin. Med J. 2017, 130, 1662–1669. [CrossRef]
93. Wikramanayake, T.C.; Borda, L.J.; Miteva, M.; Paus, R. Seborrheic dermatitis—Looking beyondMalassezia. Exp. Dermatol. 2019,28, 991–1001. [CrossRef]
94. Massiot, P.; Clavaud, C.; Thomas, M.; Ott, A.; Guéniche, A.; Panhard, S.; Muller, B.; Michelin, C.; Kerob, D.; Bouloc, A.; et al.Continuous clinical improvement of mild-to-moderate seborrheic dermatitis and rebalancing of the scalp microbiome using aselenium disulfide–based shampoo after an initial treatment with ketoconazole. J. Cosmet. Dermatol. 2021. [CrossRef]
95. Fercek, I.; Lugovic-Mihic, L.; Tambic-Andraševic, A.; Cesic, D.; Grginic, A.G.; Bešlic, I.; Mravak-Stipetic, M.; Mihatov-Štefanovic,I.; Buntic, A.-M.; Civljak, R. Features of the Skin Microbiota in Common Inflammatory Skin Diseases. Life 2021, 11, 962. [CrossRef]
96. Grice, E.A.; Segre, J.A. The Human Microbiome: Our Second Genome. Annu. Rev. Genom. Hum. Genet. 2012, 13, 151–170.[CrossRef]
97. Byrd, A.L.; Deming, C.; Cassidy, S.K.B.; Harrison, O.J.; Ng, W.-I.; Conlan, S.; Belkaid, Y.; Segre, J.A.; Kong, H.H.; Program, N.C.S.Staphylococcus aureusandStaphylococcus epidermidisstrain diversity underlying pediatric atopic dermatitis. Sci. Transl. Med.2017, 9, eaal4651. [CrossRef]
98. Kong, H.H.; Segre, J.A. Skin Microbiome: Looking Back to Move Forward. J. Investig. Dermatol. 2012, 132, 933–939. [CrossRef][PubMed]
99. Moran, M.C.; Cahill, M.P.; Brewer, M.G.; Yoshida, T.; Knowlden, S.; Perez-Nazario, N.; Schlievert, P.M.; Beck, L.A. Staphy-lococcalVirulence Factors on the Skin of Atopic Dermatitis Patients. mSphere 2019, 4, e00616-19. [CrossRef] [PubMed]
100. Yamazaki, Y.; Nakamura, Y.; Núñez, G. Role of the microbiota in skin immunity and atopic dermatitis. Allergol. Int. 2017, 66,539–544. [CrossRef] [PubMed]
101. Bukowski, M.; Wladyka, B.; Dubin, G. Exfoliative Toxins of Staphylococcus aureus. Toxins 2010, 2, 1148–1165. [CrossRef][PubMed]
102. Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibitsStaphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [CrossRef]
103. Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al.Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis.Sci. Transl. Med. 2017, 9. [CrossRef]
104. Cau, L.; Williams, M.R.; Butcher, A.M.; Nakatsuji, T.; Kavanaugh, J.S.; Cheng, J.Y.; Shafiq, F.; Higbee, K.; Hata, T.R.; Horswill, A.R.;et al. Staphylococcus epidermidis protease EcpA can be a deleterious component of the skin microbiome in atopic der-matitis. J.Allergy Clin. Immunol. 2021, 147, 955–966.e16. [CrossRef]

Pathogens 2022, 11, 121 21 of 27
105. Williams, M.R.; Cau, L.; Wang, Y.; Kaul, D.; Sanford, J.A.; Zaramela, L.S.; Khalil, S.; Butcher, A.M.; Zengler, K.; Horswill, A.R.;et al. Interplay of Staphylococcal and Host Proteases Promotes Skin Barrier Disruption in Netherton Syndrome. Cell Rep. 2020, 30,2923–2933.e7. [CrossRef]
106. Lee, Y.B.; Byun, E.J.; Kim, H.S. Potential Role of the Microbiome in Acne: A Comprehensive Review. J. Clin. Med. 2019, 8, 987.[CrossRef] [PubMed]
107. Saint-Leger, D.; Bague, A.; Lefebvre, E.; Cohen, E.; Chivot, M. A possible role for squalene in the pathogenesis of acne. II. In vivostudy of squalene oxides in skin surface and intra-comedonal lipids of acne patients. Br. J. Dermatol. 1986, 114, 543–552. [CrossRef][PubMed]
108. Saint-Leger, D.; Bague, A.; Cohen, E.; Lchivot, M. A possible role for squalene in the pathogenesis of acne. I. In vitro study ofsqualene oxidation. Br. J. Dermatol. 1986, 114, 535–542. [CrossRef] [PubMed]
109. Jarrousse, V.; Castex-Rizzi, N.; Khammari, A.; Charveron, M.; Dréno, B. Modulation of integrins and filaggrin expression byPropionibacterium acnes extracts on keratinocytes. Arch. Dermatol. Res. 2007, 299, 441–447. [CrossRef]
110. Isard, O.; Knol, A.C.; Aries, M.F.; Nguyen, J.M.; Khammari, A.; Castex-Rizzi, N.; Dreno, B. Propionibacterium acnes activates theIGF-1/IGF-1R system in the epidermis and induces keratinocyte proliferation. J. Investig. Dermatol. 2011, 131, 59–66. [CrossRef]
111. Iinuma, K.; Sato, T.; Akimoto, N.; Noguchi, N.; Sasatsu, M.; Nishijima, S.; Kurokawa, I.; Ito, A. Involvement of Propioni-bacteriumacnes in the augmentation of lipogenesis in hamster sebaceous glands in vivo and in vitro. J. Investig. Dermatol. 2009, 129,2113–2119. [CrossRef]
112. Graham, G.M.; Farrar, M.D.; Cruse-Sawyer, J.E.; Holland, K.T.; Ingham, E. Proinflammatory cytokine production by humankeratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. Br. J. Dermatol. 2004, 150, 421–428. [CrossRef][PubMed]
113. Jugeau, S.; Tenaud, I.; Knol, A.C.; Jarrousse, V.; Quereux, G.; Khammari, A.; Dreno, B. Induction of toll-like receptors byPropionibacterium acnes. Br. J. Dermatol. 2005, 153, 1105–1113. [CrossRef]
114. Johnson, T.; Kang, D.; Barnard, E.; Li, H. Strain-Level Differences in Porphyrin Production and Regulation in Propionibacte-riumacnes Elucidate Disease Associations. mSphere 2016, 1, e00023-15. [CrossRef] [PubMed]
115. Barnard, E.; Johnson, T.; Ngo, T.; Arora, U.; Leuterio, G.; McDowell, A.; Li, H. Porphyrin Production and Regulation in Cu-taneousPropionibacteria. mSphere 2020, 5, e00793-19. [CrossRef]
116. Vlachos, C.; Schulte, B.M.; Magiatis, P.; Adema, G.J.; Gaitanis, G. Malassezia-derived indoles activate the aryl hydrocarbonreceptor and inhibit Toll-like receptor-induced maturation in monocyte-derived dendritic cells. Br. J. Dermatol. 2012, 167, 496–505.[CrossRef] [PubMed]
117. Barnard, E.; Shi, B.; Kang, D.; Craft, N.; Li, H. The balance of metagenomic elements shapes the skin microbiome in acne andhealth. Sci. Rep. 2016, 6, 39491. [CrossRef] [PubMed]
118. Park, Y.J.; Lee, H.K. The Role of Skin and Orogenital Microbiota in Protective Immunity and Chronic Immune-MediatedInflammatory Disease. Front. Immunol. 2017, 8, 1955. [CrossRef] [PubMed]
119. Liu, S.-H.; Yu, H.-Y.; Chang, Y.-C.; Hui, R.C.-Y.; Huang, Y.-C.; Huang, Y.-H. Host characteristics and dynamics of Staphylococcusaureus colonization in patients with moderate-to-severe psoriasis before and after treatment: A prospective cohort study. J. Am.Acad. Dermatol. 2018, 81, 605–607. [CrossRef]
120. Chen, L.; Li, J.; Zhu, W.; Kuang, Y.; Liu, T.; Zhang, W.; Chen, X.; Peng, C. Skin and Gut Microbiome in Psoriasis: Gaining Insightinto the Pathophysiology of It and Finding Novel Therapeutic Strategies. Front. Microbiol. 2020, 11, 3201. [CrossRef]
121. Polak, K.; Jobbágy, A.; Muszynski, T.; Wojciechowska, K.; Fratczak, A.; Bánvölgyi, A.; Bergler-Czop, B.; Kiss, N. MicrobiomeModulation as a Therapeutic Approach in Chronic Skin Diseases. Biomedicines 2021, 9, 1436. [CrossRef]
122. Dreno, B.; Martin, R.; Khammari, A.; Moyal, D.; Henley, J.B.; Seite, S. Skin Microbiome and Acne Vulgaris: Staphylococcus, a NewActor in Acne. Ski. J. Cutan. Med. 2017, 1, s54. [CrossRef]
123. Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes: Two MajorSentinels of Skin Microbiota and the Influence of Cosmetics. Microorganisms 2020, 8, 1752. [CrossRef] [PubMed]
124. Grice, E.A.; Kong, H.H.; Renaud, G.; Young, A.C.; Bouffard, G.G.; Blakesley, R.W.; Wolfsberg, T.G.; Turner, M.L.; Segre, J.A. Adiversity profile of the human skin microbiota. Genome Res. 2008, 18, 1043–1050. [CrossRef]
125. Lauber, C.L.; Zhou, N.; Gordon, J.I.; Knight, R.; Fierer, N. Effect of storage conditions on the assessment of bacterial communitystructure in soil and human-associated samples. FEMS Microbiol. Lett. 2010, 307, 80–86. [CrossRef] [PubMed]
126. Chng, K.R.; Tay, A.S.L.; Li, C.; Ng, A.H.Q.; Wang, J.; Suri, B.K.; Matta, S.A.; McGovern, N.; Janela, B.; Wong, X.F.C.C.; et al. Wholemetagenome profiling reveals skin microbiome-dependent susceptibility to atopic dermatitis flare. Nat. Microbiol. 2016, 1, 16106.[CrossRef]
127. Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W.Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87.[CrossRef]
128. Meisel, J.S.; Hannigan, G.D.; Tyldsley, A.S.; SanMiguel, A.J.; Hodkinson, B.P.; Zheng, Q.; Grice, E.A. Skin Microbiome SurveysAre Strongly Influenced by Experimental Design. J. Investig. Dermatol. 2016, 136, 947–956. [CrossRef]
129. Gao, Z.; Perez, G.P.; Chen, Y.; Blaser, M.J. Quantitation of Major Human Cutaneous Bacterial and Fungal Populations. J. Clin.Microbiol. 2010, 48, 3575–3581. [CrossRef]

Pathogens 2022, 11, 121 22 of 27
130. Kong, H.H.; Andersson, B.; Clavel, T.; Common, J.; Jackson, S.; Olson, N.; Segre, J.A.; Traidl-Hoffmann, C. Performing SkinMicrobiome Research: A Method to the Madness. J. Investig. Dermatol. 2017, 137, 561–568. [CrossRef] [PubMed]
131. Chan, J.Z.; Sergeant, M.J.; Lee, O.Y.; Minnikin, D.E.; Besra, G.S.; Pap, I.; Spigelman, M.; Donoghue, H.D.; Pallen, M.J. Meta-genomicanalysis of tuberculosis in a mummy. N. Engl. J. Med. 2013, 369, 289–290. [CrossRef]
132. Pallen, M.J. Diagnostic metagenomics: Potential applications to bacterial, viral and parasitic infections. Parasitology 2014, 141,1856–1862. [CrossRef] [PubMed]
133. Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [CrossRef]134. Bell, E.C.; Ravis, W.R.; Lloyd, K.B.; Stokes, T.J. Effects of St. John’s wort supplementation on ibuprofen pharmacokinetics. Ann.
Pharmacother. 2007, 41, 229–234. [CrossRef]135. Bhatt, A.P.; Redinbo, M.R.; Bultman, S.J. The role of the microbiome in cancer development and therapy. CA Cancer J. Clin. 2017,
67, 326–344. [CrossRef]136. Chiu, C.Y.; Miller, S.A. Clinical metagenomics. Nat. Rev. Genet. 2019, 20, 341–355. [CrossRef] [PubMed]137. Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol.
2016, 17, 451–459. [CrossRef]138. Chen, H.; Nwe, P.-K.; Yang, Y.; Rosen, C.E.; Bielecka, A.A.; Kuchroo, M.; Cline, G.W.; Kruse, A.C.; Ring, A.M.; Crawford, J.M.;
et al. A Forward Chemical Genetic Screen Reveals Gut Microbiota Metabolites That Modulate Host Physiology. Cell 2019, 177,1217–1231.e18. [CrossRef] [PubMed]
139. Zierer, J.; Jackson, M.A.; Kastenmüller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J.; et al.The fecal metabolome as a functional readout of the gut microbiome. Nat. Genet. 2018, 50, 790–795. [CrossRef]
140. Jeffery, I.B.; Das, A.; O’Herlihy, E.; Coughlan, S.; Cisek, K.; Moore, M.; Bradley, F.; Carty, T.; Pradhan, M.; Dwibedi, C.; et al.Differences in Fecal Microbiomes and Metabolomes of People With vs Without Irritable Bowel Syndrome and Bile Acid Mal-absorption. Gastroenterology 2020, 158, 1016–1028.e8. [CrossRef]
141. Dumas, M.-E.; Barton, R.H.; Toye, A.; Cloarec, O.; Blancher, C.; Rothwell, A.; Fearnside, J.; Tatoud, R.; Blanc, V.; Lindon, J.C.; et al.Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc. Natl. Acad. Sci.USA 2006, 103, 12511–12516. [CrossRef]
142. Metwaly, A.; Dunkel, A.; Waldschmitt, N.; Raj, A.C.D.; Lagkouvardos, I.; Corraliza, A.M.; Mayorgas, A.; Martinez-Medina,M.; Reiter, S.; Schloter, M.; et al. Integrated microbiota and metabolite profiles link Crohn’s disease to sulfur metabolism. Nat.Commun. 2020, 11, 4322. [CrossRef] [PubMed]
143. Zhou, W.; Sailani, M.R.; Contrepois, K.; Zhou, Y.; Ahadi, S.; Leopold, S.R.; Zhang, M.J.; Rao, V.; Avina, M.; Mishra, T.; et al.Longitudinal multi-omics of host–microbe dynamics in prediabetes. Nature 2019, 569, 663–671. [CrossRef] [PubMed]
144. Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.;McIver, L.J.; et al. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol. 2018, 4,293–305. [CrossRef] [PubMed]
145. Sadowski, T.; Klose, C.; Gerl, M.J.; Wójcik-Maciejewicz, A.; Herzog, R.; Simons, K.; Reich, A.; Surma, M.A. Large-scale humanskin lipidomics by quantitative, high-throughput shotgun mass spectrometry. Sci. Rep. 2017, 7, srep43761. [CrossRef]
146. Kuehne, A.; Hildebrand, J.; Soehle, J.; Wenck, H.; Terstegen, L.; Gallinat, S.; Knott, A.; Winnefeld, M.; Zamboni, N. An integrativemetabolomics and transcriptomics study to identify metabolic alterations in aged skin of humans in vivo. BMC Genom. 2017, 18, 169.[CrossRef]
147. Bouslimani, A.; Porto, C.; Rath, C.M.; Wang, M.; Guo, Y.; Gonzalez, A.; Berg-Lyon, D.; Ackermann, G.; Christensen, G.J.M.;Nakatsuji, T.; et al. Molecular cartography of the human skin surface in 3D. Proc. Natl. Acad. Sci. USA 2015, 112, E2120–E2129.[CrossRef]
148. Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers,J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752.[CrossRef]
149. Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan,T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking.Nat. Biotechnol. 2016, 34, 828–837. [CrossRef]
150. Leao, T.F.; Clark, C.M.; Bauermeister, A.; Elijah, E.O.; Gentry, E.C.; Husband, M.; Oliveira, M.F.; Bandeira, N.; Wang, M.;Dorrestein, P.C. Quick-start infrastructure for untargeted metabolomics analysis in GNPS. Nat. Metab. 2021, 3, 880–882. [CrossRef]
151. Jarmusch, A.K.; Vrbanac, A.; Momper, J.D.; Ma, J.D.; Alhaja, M.; Liyanage, M.; Knight, R.; Dorrestein, P.C.; Tsunoda, S.M.Enhanced Characterization of Drug Metabolism and the Influence of the Intestinal Microbiome: A Pharmacokinetic, Microbiome,and Untargeted Metabolomics Study. Clin. Transl. Sci. 2020, 13, 972–984. [CrossRef]
152. Garg, N.; Wang, M.; Hyde, E.; da Silva, R.R.; Melnik, A.V.; Protsyuk, I.; Bouslimani, A.; Lim, Y.W.; Wong, R.; Humphrey, G.; et al.Three-Dimensional Microbiome and Metabolome Cartography of a Diseased Human Lung. Cell Host Microbe 2017, 22, 705–716.e4.[CrossRef]
153. Ding, L.; Bar-Shalom, R.; Aharonovich, D.; Kurisawa, N.; Patial, G.; Li, S.; He, S.; Yan, X.; Iwasaki, A.; Suenaga, K.; et al.Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, aLipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells. Mar. Drugs 2021, 19, 397. [CrossRef] [PubMed]

Pathogens 2022, 11, 121 23 of 27
154. Luzzatto-Knaan, T.; Garg, N.; Wang, M.; Glukhov, E.; Peng, Y.; Ackermann, G.; Amir, A.; Duggan, B.M.; Ryazanov, S.; Gerwick, L.;et al. Digitizing mass spectrometry data to explore the chemical diversity and distribution of marine cyanobacteria and algae.eLife 2017, 6, e24214. [CrossRef] [PubMed]
155. Petras, D.; Minich, J.J.; Cancelada, L.B.; Torres, R.R.; Kunselman, E.; Wang, M.; White, M.E.; Allen, E.E.; Prather, K.A.; Aluwihare,L.I.; et al. Non-targeted tandem mass spectrometry enables the visualization of organic matter chemotype shifts in coastalseawater. Chemosphere 2021, 271, 129450. [CrossRef] [PubMed]
156. Kapono, C.A.; Morton, J.T.; Bouslimani, A.; Melnik, A.V.; Orlinsky, K.; Knaan, T.L.; Garg, N.; Vázquez-Baeza, Y.; Protsyuk, I.;Janssen, S.; et al. Creating a 3D microbial and chemical snapshot of a human habitat. Sci. Rep. 2018, 8, 3669. [CrossRef] [PubMed]
157. McCall, L.-I.; Callewaert, C.; Zhu, Q.; Song, S.J.; Bouslimani, A.; Minich, J.J.; Ernst, M.; Ruiz-Calderon, J.F.; Cavallin, H.; Pereira,H.S.; et al. Home chemical and microbial transitions across urbanization. Nat. Microbiol. 2019, 5, 108–115. [CrossRef]
158. Jarmusch, A.K.; Elijah, E.O.; Vargas, F.; Bouslimani, A.; da Silva, R.R.; Ernst, M.; Wang, M.; Del Rosario, K.K.; Dorrestein, P.C.;Tsunoda, S.M. Initial Development toward Non-Invasive Drug Monitoring via Untargeted Mass Spectrometric Analysis ofHuman Skin. Anal. Chem. 2019, 91, 8062–8069. [CrossRef] [PubMed]
159. Bouslimani, A.; Melnik, A.V.; Xu, Z.; Amir, A.; da Silva, R.R.; Wang, M.; Bandeira, N.; Alexandrov, T.; Knight, R.; Dorrestein, P.C.Lifestyle chemistries from phones for individual profiling. Proc. Natl. Acad. Sci. USA 2016, 113, E7645–E7654. [CrossRef]
160. Jarmusch, A.K.; Wang, M.; Aceves, C.M.; Advani, R.S.; Aguirre, S.; Aksenov, A.A.; Aleti, G.; Aron, A.T.; Bauermeister, A.;Bolleddu, S.; et al. ReDU: A framework to find and reanalyze public mass spectrometry data. Nat. Methods 2020, 17, 901–904.[CrossRef]
161. Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.;et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [CrossRef]
162. Tripathi, A.; Vázquez-Baeza, Y.; Gauglitz, J.M.; Wang, M.; Dührkop, K.; Nothias-Esposito, M.; Acharya, D.D.; Ernst, M.; van derHooft, J.J.J.; Zhu, Q.; et al. Chemically informed analyses of metabolomics mass spectrometry data with Qemistree. Nat. Chem.Biol. 2020, 17, 146–151. [CrossRef] [PubMed]
163. Da Silva, R.R.; Wang, M.; Nothias, L.-F.; Van Der Hooft, J.J.J.; Caraballo-Rodríguez, A.M.; Fox, E.; Balunas, M.J.; Klassen, J.L.;Lopes, N.P.; Dorrestein, P.C. Propagating annotations of molecular networks using in silico fragmentation. PLOS Comput. Biol.2018, 14, e1006089. [CrossRef] [PubMed]
164. Sugimoto, Y.; Camacho, F.R.; Wang, S.; Chankhamjon, P.; Odabas, A.; Biswas, A.; Jeffrey, P.D.; Donia, M.S. A metagenomic strategyfor harnessing the chemical repertoire of the human microbiome. Science 2019, 366, eaax9176. [CrossRef]
165. Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to thesecondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [CrossRef] [PubMed]
166. Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungalsecondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [CrossRef] [PubMed]
167. Skinnider, M.A.; Merwin, N.J.; Johnston, C.W.; Magarvey, N.A. PRISM 3: Expanded prediction of natural product chemicalstructures from microbial genomes. Nucleic Acids Res. 2017, 45, W49–W54. [CrossRef]
168. Cao, L.; Gurevich, A.; Alexander, K.L.; Naman, C.B.; Leão, T.; Glukhov, E.; Luzzatto-Knaan, T.; Vargas, F.; Quinn, R.; Bouslimani,A.; et al. MetaMiner: A Scalable Peptidogenomics Approach for Discovery of Ribosomal Peptide Natural Products with BlindModifications from Microbial Communities. Cell Syst. 2019, 9, 600–608.e4. [CrossRef]
169. Behsaz, B.; Bode, E.; Gurevich, A.; Shi, Y.N.; Grundmann, F.; Acharya, D.; Caraballo-Rodriguez, A.M.; Bouslimani, A.; Pa-nitchpakdi, M.; Linck, A.; et al. Integrating genomics and metabolomics for scalable non-ribosomal peptide discovery. Nat.Commun. 2021, 12, 3225. [CrossRef]
170. Mohimani, H.; Liu, W.-T.; Kersten, R.D.; Moore, B.S.; Dorrestein, P.C.; Pevzner, P.A. NRPquest: Coupling Mass Spectrometry andGenome Mining for Nonribosomal Peptide Discovery. J. Nat. Prod. 2014, 77, 1902–1909. [CrossRef]
171. Li, X.; Guo, J.; Dai, S.; Ouyang, Y.; Wu, H.; Sun, W.; Wang, G. Exploring and exploiting microbial diversity through meta-genomicsfor natural product drug discovery. Curr. Top. Med. Chem. 2009, 9, 1525–1535. [CrossRef]
172. Chavira, A.; Belda-Ferre, P.; Kosciolek, T.; Ali, F.; Dorrestein, P.C.; Knight, R. The Microbiome and Its Potential for Pharma-cology.Handb. Exp. Pharmacol. 2019, 260, 301–326.
173. Culligan, E.P.; Sleator, R.D.; Marchesi, J.R.; Hill, C. Metagenomics and novel gene discovery: Promise and potential for noveltherapeutics. Virulence 2014, 5, 399–412. [CrossRef]
174. Liu, Y.; Liu, Y.; Du, Z.; Zhang, L.; Chen, J.; Shen, Z.; Liu, Q.; Qin, J.; Lv, H.; Wang, H.; et al. Skin microbiota analysis-inspireddevelopment of novel anti-infectives. Microbiome 2020, 8, 85. [CrossRef]
175. Mathieu, A.; Vogel, T.M.; Simonet, P. The future of skin metagenomics. Res. Microbiol. 2014, 165, 69–76. [CrossRef] [PubMed]176. Van den Bogert, B.; Boekhorst, J.; Pirovano, W.; May, A. On the Role of Bioinformatics and Data Science in Industrial Microbiome
Applications. Front. Genet. 2019, 10, 721. [CrossRef] [PubMed]177. Zhang, X.; Li, L.; Butcher, J.; Stintzi, A.; Figeys, D. Advancing functional and translational microbiome research using meta-omics
approaches. Microbiome 2019, 7, 154. [CrossRef]178. Heinken, A.; Thiele, I. Systems biology of host-microbe metabolomics. Wiley Interdiscip. Rev. Syst. Biol. Med. 2015, 7, 195–219.
[CrossRef]179. Nielsen, J. Systems Biology of Metabolism: A Driver for Developing Personalized and Precision Medicine. Cell Metab. 2017, 25,
572–579. [CrossRef]

Pathogens 2022, 11, 121 24 of 27
180. Magnusdottir, S.; Thiele, I. Modeling metabolism of the human gut microbiome. Curr. Opin. Biotechnol. 2018, 51, 90–96. [CrossRef]181. Cakir, T.; Panagiotou, G.; Uddin, R.; Durmus, S. Novel Approaches for Systems Biology of Metabolism-Oriented Patho-gen-
Human Interactions: A Mini-Review. Front. Cell Infect. Microbiol. 2020, 10, 52. [CrossRef] [PubMed]182. Bernabé, B.P.; Cralle, L.; Gilbert, J.A. Systems biology of the human microbiome. Curr. Opin. Biotechnol. 2018, 51, 146–153.
[CrossRef]183. Son, J.W.; Shoaie, S.; Lee, S. Systems Biology: A Multi-Omics Integration Approach to Metabolism and the Microbiome. Endocrinol.
Metab. 2020, 35, 507–514. [CrossRef] [PubMed]184. Francke, C.; Siezen, R.J.; Teusink, B. Reconstructing the metabolic network of a bacterium from its genome. Trends Microbiol. 2005,
13, 550–558. [CrossRef] [PubMed]185. Oberhardt, M.A.; Palsson, B.O.; Papin, J.A. Applications of genome-scale metabolic reconstructions. Mol. Syst. Biol. 2009, 5, 320.
[CrossRef]186. Duarte, N.C.; Becker, S.A.; Jamshidi, N.; Thiele, I.; Mo, M.L.; Vo, T.D.; Srivas, R.; Palsson, B. Global reconstruction of the human
metabolic network based on genomic and bibliomic data. Proc. Natl. Acad. Sci. USA 2007, 104, 1777–1782. [CrossRef] [PubMed]187. Brunk, E.; Sahoo, S.; Zielinski, D.C.; Altunkaya, A.; Dräger, A.; Mih, N.; Gatto, F.; Nilsson, A.; Gonzalez, G.A.P.; Aurich, M.K.; et al.
Recon3D enables a three-dimensional view of gene variation in human metabolism. Nat. Biotechnol. 2018, 36, 272–281. [CrossRef]188. Haraldsdóttir, H.S.; Thiele, I.; Fleming, R.M. Comparative evaluation of open source software for mapping between metabolite
identifiers in metabolic network reconstructions: Application to Recon 2. J. Chemin- 2014, 6, 2. [CrossRef] [PubMed]189. Magnusdottir, S.; Heinken, A.; Kutt, L.; Ravcheev, D.A.; Bauer, E.; Noronha, A.; Greenhalgh, K.; Jäger, C.; Baginska, J.; Wilmes, P.;
et al. Generation of genome-scale metabolic reconstructions for 773 members of the human gut microbiota. Nat. Biotechnol. 2016,35, 81–89. [CrossRef] [PubMed]
190. Hussan, J.R.; Hunter, P.J. Our natural “makeup” reveals more than it hides: Modeling the skin and its microbiome. WIREs Mech.Dis. 2020, 13, e1497. [CrossRef]
191. Thiele, I.; Palsson, B.O. A protocol for generating a high-quality genome-scale metabolic reconstruction. Nat. Protoc. 2010, 5,93–121. [CrossRef]
192. E Larsen, P.; Collart, F.R.; Field, D.; Meyer, F.; Keegan, K.P.; Henry, C.S.; McGrath, J.; Quinn, J.; A Gilbert, J. Predicted RelativeMetabolomic Turnover (PRMT): Determining metabolic turnover from a coastal marine metagenomic dataset. Microb. InformaticsExp. 2011, 1, 4. [CrossRef] [PubMed]
193. Noecker, C.; Eng, A.; Srinivasan, S.; Theriot, C.M.; Young, V.B.; Jansson, J.K.; Fredricks, D.N.; Borenstein, E. Metabolic Mod-el-Based Integration of Microbiome Taxonomic and Metabolomic Profiles Elucidates Mechanistic Links between Ecological andMetabolic Variation. mSystems 2016, 1, e00013-15. [CrossRef]
194. Chang, Y.-L.; Rossetti, M.; Vlamakis, H.; Casero, D.; Sunga, G.; Harre, N.; Miller, S.; Humphries, R.; Stappenbeck, T.; Simpson,K.W.; et al. A screen of Crohn’s disease-associated microbial metabolites identifies ascorbate as a novel metabolic inhibitor ofactivated human T cells. Mucosal Immunol. 2018, 12, 457–467. [CrossRef] [PubMed]
195. Mallick, H.; Franzosa, E.A.; Mclver, L.J.; Banerjee, S.; Sirota-Madi, A.; Kostic, A.D.; Clish, C.B.; Vlamakis, H.; Xavier, R.J.;Huttenhower, C. Predictive metabolomic profiling of microbial communities using amplicon or metagenomic sequences. Nat.Commun. 2019, 10, 3136. [CrossRef]
196. Yin, X.; Altman, T.; Rutherford, E.; West, K.A.; Wu, Y.; Choi, J.; Beck, P.L.; Kaplan, G.G.; Dabbagh, K.; DeSantis, T.Z.; et al. A ComparativeEvaluation of Tools to Predict Metabolite Profiles From Microbiome Sequencing Data. Front. Microbiol. 2020, 11, 3132. [CrossRef][PubMed]
197. Zmora, N.; Soffer, E.; Elinav, E. Transforming medicine with the microbiome. Sci. Transl. Med. 2019, 11, eaaw1815. [CrossRef][PubMed]
198. Jobin, C. Precision medicine using microbiota. Science 2018, 359, 32–34. [CrossRef]199. Mills, S.; Stanton, C.; Lane, J.A.; Smith, G.J.; Ross, R.P. Precision Nutrition and the Microbiome, Part I: Current State of the Science.
Nutrients 2019, 11, 923. [CrossRef]200. Guarner, F.; Schaafsma, G.J. Probiotics. Int. J. Food Microbiol. 1998, 39, 237–238. [CrossRef]201. Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert
consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope andappropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [CrossRef]
202. Wieërs, G.; Belkhir, L.; Enaud, R.; Leclercq, S.; De Foy, J.-M.P.; Dequenne, I.; De Timary, P.; Cani, P.D. How Probiotics Affect theMicrobiota. Front. Cell. Infect. Microbiol. 2020, 9, 454. [CrossRef]
203. Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.;et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition andscope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [CrossRef] [PubMed]
204. Report of the ICCR: ICCR Microbiome and Cosmetics—Survey of Products Ingredients Terminologies and Regulatory Approaches.2021. Available online: https://www.iccr-cosmetics.org/topics-documents/14-microbiome (accessed on 25 November 2021).
205. Gueniche, A.; Cathelineau, A.C.; Bastien, P.; Esdaile, J.; Martin, R.; Queille Roussel, C.; Breton, L. Vitreoscilla filiformis biomassimproves seborrheic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 1014–1015. [CrossRef] [PubMed]

Pathogens 2022, 11, 121 25 of 27
206. Gueniche, A.; Knaudt, B.; Schuck, E.; Volz, T.; Bastien, P.; Martin, R.; Rocken, M.; Breton, L.; Biedermann, T. Effects of non-pathogenic gram-negative bacterium Vitreoscilla filiformis lysate on atopic dermatitis: A prospective, randomized, double-blind,placebo-controlled clinical study. Br. J. Dermatol. 2008, 159, 1357–1363. [CrossRef] [PubMed]
207. Peguet-Navarro, J.; Dezutter-Dambuyant, C.; Buetler, T.; LeClaire, J.; Smola, H.; Blum, S.; Bastien, P.; Breton, L.; Gueniche, A.Supplementation with oral probiotic bacteria protects human cutaneous immune homeostasis after UV exposure-double blind,randomized, placebo controlled clinical trial. Eur. J. Dermatol. 2008, 18. [CrossRef]
208. Gueniche, A.; Philippe, D.; Bastien, P.; Blum, S.; Buyukpamukcu, E.; Castiel-Higounenc, I. Probiotics for photoprotection.Dermatoendocrinology 2009, 1, 275–279. [CrossRef]
209. Guéniche, A.; Bastien, P.; Ovigne, J.M.; Kermici, M.; Courchay, G.; Chevalier, V.; Breton, L.; Castiel-Higounenc, I. Bifidobacteriumlongum lysate, a new ingredient for reactive skin. Exp. Dermatol. 2009, 19, e1–e8. [CrossRef]
210. Gueniche, A.; Benyacoub, J.; Philippe, D.; Bastien, P.; Kusy, N.; Breton, L.; Blum, S.; Castiel-Higounenc, I. Lactobacillus par-acaseiCNCM I-2116 (ST11) inhibits substance P-induced skin inflammation and accelerates skin barrier function recovery in vitro. Eur.J. Dermatol. 2010, 20, 731–737.
211. Li, L.; Han, Z.; Niu, X.; Zhang, G.; Jia, Y.; Zhang, S.; He, C. Probiotic Supplementation for Prevention of Atopic Dermatitis inInfants and Children: A Systematic Review and Meta-analysis. Am. J. Clin. Dermatol. 2018, 20, 367–377. [CrossRef]
212. Reygagne, P.; Bastien, P.; Couavoux, M.P.; Philippe, D.; Renouf, M.; Castiel-Higounenc, I.; Gueniche, A. The positive benefit ofLactobacillus paracasei NCC2461 ST11 in healthy volunteers with moderate to severe dandruff. Benef. Microbes 2017, 8, 671–680.[CrossRef]
213. Benyacoub, J.; Bosco, N.; Blanchard, C.; Demont, A.; Philippe, D.; Castiel-Higounenc, I.; Gueniche, A. Immune modulationproperty of Lactobacillus paracasei NCC2461 (ST11) strain and impact on skin defences. Benef. Microbes 2014, 5, 129–136.[CrossRef] [PubMed]
214. Gueniche, A.; Philippe, D.; Bastien, P.; Reuteler, G.; Blum, S.; Castiel-Higounenc, I.; Breton, L.; Benyacoub, J. Randomiseddouble-blind placebo-controlled study of the effect of Lactobacillus paracasei NCC 2461 on skin reactivity. Benef. Microbes 2014, 5,137–145. [CrossRef]
215. Bouilly-Gauthier, D.; Jeannes, C.; Maubert, Y.; Duteil, L.; Queille-Roussel, C.; Piccardi, N.; Montastier, C.; Manissier, P.; Pierard, G.;Ortonne, J.P. Clinical evidence of benefits of a dietary supplement containing probiotic and carotenoids on ultravio-let-inducedskin damage. Br. J. Dermatol. 2010, 163, 536–543. [CrossRef] [PubMed]
216. Kober, M.-M.; Bowe, W.P. The effect of probiotics on immune regulation, acne, and photoaging. Int. J. Women’s Dermatol. 2015, 1,85–89. [CrossRef]
217. Han, N.; Jia, L.; Guo, L.; Su, Y.; Luo, Z.; Du, J.; Mei, S.; Liu, Y. Balanced oral pathogenic bacteria and probiotics promoted woundhealing via maintaining mesenchymal stem cell homeostasis. Stem Cell Res. Ther. 2020, 11, 61. [CrossRef]
218. Kunz, A.N.; Noel, J.M.; Fairchok, M.P. Two Cases of Lactobacillus Bacteremia During Probiotic Treatment of Short Gut Syndrome.J. Pediatr. Gastroenterol. Nutr. 2004, 38, 457–458. [CrossRef]
219. Yelin, I.; Flett, K.B.; Merakou, C.; Mehrotra, P.; Stam, J.; Snesrud, E.; Hinkle, M.; Lesho, E.; McGann, P.; McAdam, A.J.; et al.Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat. Med. 2019,25, 1728–1732. [CrossRef]
220. Galdeano, C.M.; Perdigon, G. Role of viability of probiotic strains in their persistence in the gut and in mucosal immunestimulation. J. Appl. Microbiol. 2004, 97, 673–681. [CrossRef]
221. Sawada, J.; Morita, H.; Tanaka, A.; Salminen, S.; He, F.; Matsuda, H. Ingestion of heat-treated Lactobacillus rhamnosus GGprevents development of atopic dermatitis in NC/Nga mice. Clin. Exp. Allergy 2006, 37, 296–303. [CrossRef]
222. França, K. Topical Probiotics in Dermatological Therapy and Skincare: A Concise Review. Dermatol. Ther. 2020, 11, 71–77.[CrossRef] [PubMed]
223. Al-Ghazzewi, F.H.; Tester, R.F. Impact of prebiotics and probiotics on skin health. Benef. Microbes 2014, 5, 99–107. [CrossRef][PubMed]
224. Lopes, E.G.; Moreira, D.A.; Gullón, P.; Gullón, B.; Cardelle-Cobas, A.; Tavaria, F.K. Topical application of probiotics in skin:Adhesion, antimicrobial and antibiofilmin vitroassays. J. Appl. Microbiol. 2017, 122, 450–461. [CrossRef]
225. Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.-G.; Kim, T.-R.; Kim, H.; Lee, J.; et al. Antiwrinkle andAntimelanogenesis Effects of Tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [CrossRef][PubMed]
226. Brandi, J.; Cheri, S.; Manfredi, M.; Di Carlo, C.; Vanella, V.V.; Federici, F.; Bombiero, E.; Bazaj, A.; Rizzi, E.; Manna, L.; et al.Exploring the wound healing, anti-inflammatory, anti-pathogenic and proteomic effects of lactic acid bacteria on keratinocytes.Sci. Rep. 2020, 10, 11572. [CrossRef] [PubMed]
227. Jung, Y.-O.; Jeong, H.; Cho, Y.; Lee, E.-O.; Jang, H.-W.; Kim, J.; Nam, K.T.; Lim, K.-M. Lysates of a Probiotic, Lactobacillusrhamnosus, Can Improve Skin Barrier Function in a Reconstructed Human Epidermis Model. Int. J. Mol. Sci. 2019, 20, 4289.[CrossRef]
228. Dimarzio, L.; Cinque, B.; Cupelli, F.; De Simone, C.; Cifone, M.G.; Giuliani, M. Increase of Skin-Ceramide Levels in Aged Subjectsfollowing a Short-Term Topical Application of Bacterial Sphingomyelinase from Streptococcus Thermophilus. Int. J. Immunopathol.Pharmacol. 2008, 21, 137–143. [CrossRef]

Pathogens 2022, 11, 121 26 of 27
229. Volz, T.; Skabytska, Y.; Guenova, E.; Chen, K.M.; Frick, J.S.; Kirschning, C.J.; Kaesler, S.; Rocken, M.; Biedermann, T. Non-pathogenic bacteria alleviating atopic dermatitis inflammation induce IL-10-producing dendritic cells and regulatory Tr1 cells. J.Investig. Dermatol. 2014, 134, 96–104. [CrossRef]
230. Ding, T.; Schloss, P.D. Dynamics and associations of microbial community types across the human body. Nature 2014, 509, 357–360.[CrossRef]
231. Gallo, R.L.; Hooper, L.V. Epithelial antimicrobial defence of the skin and intestine. Nat. Rev. Immunol. 2012, 12, 503–516. [CrossRef]232. Nodake, Y.; Matsumoto, S.; Miura, R.; Honda, H.; Ishibashi, G.; Matsumoto, S.; Dekio, I.; Sakakibara, R. Pilot study on novel skin
care method by augmentation with Staphylococcus epidermidis, an autologous skin microbe—A blinded randomized clinicaltrial. J. Dermatol. Sci. 2015, 79, 119–126. [CrossRef]
233. Hols, P.; Ledesma-García, L.; Gabant, P.; Mignolet, J. Mobilization of Microbiota Commensals and Their Bacteriocins forTherapeutics. Trends Microbiol. 2019, 27, 690–702. [CrossRef]
234. Cogen, A.L.; Yamasaki, K.; Sanchez, K.M.; Dorschner, R.A.; Lai, Y.; MacLeod, D.T.; Torpey, J.W.; Otto, M.; Nizet, V.; Kim, J.E.; et al.Selective Antimicrobial Action Is Provided by Phenol-Soluble Modulins Derived from Staphylococcus epidermidis, a NormalResident of the Skin. J. Investig. Dermatol. 2010, 130, 192–200. [CrossRef]
235. Drake, D.R.; Brogden, K.A.; Dawson, D.V.; Wertz, P.W. Thematic Review Series: Skin Lipids. Antimicrobial lipids at the skinsurface. J. Lipid Res. 2008, 49, 4–11. [CrossRef]
236. Thiboutot, D.M. Inflammasome Activation by Propionibacterium acnes: The Story of IL-1 in Acne Continues to Unfold. J. Investig.Dermatol. 2014, 134, 595–597. [CrossRef]
237. Li, D.; Wang, W.; Wu, Y.; Ma, X.; Zhou, W.; Lai, Y. Lipopeptide 78 from Staphylococcus epidermidis Activates beta-Catenin toInhibit Skin Inflammation. J. Immunol. 2019, 202, 1219–1228. [CrossRef] [PubMed]
238. Bitschar, K.; Staudenmaier, L.; Klink, L.; Focken, J.; Sauer, B.; Fehrenbacher, B.; Herster, F.; Bittner, Z.; Bleul, L.; Schaller, M.; et al.Staphylococcus aureus Skin Colonization Is Enhanced by the Interaction of Neutrophil Extracellular Traps with Keratinocytes. J.Investig. Dermatol. 2019, 140, 1054–1065.e4. [CrossRef] [PubMed]
239. Paetzold, B.; Willis, J.R.; Pereira de Lima, J.; Knodlseder, N.; Bruggemann, H.; Quist, S.R.; Gabaldon, T.; Guell, M. Skin microbiomemodulation induced by probiotic solutions. Microbiome 2019, 7, 95. [CrossRef]
240. Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota-host interactions. Nature 2018, 553, 427–436. [CrossRef]241. Shu, M.; Wang, Y.; Yu, J.; Kuo, S.; Coda, A.; Jiang, Y.; Gallo, R.L.; Huang, C.-M. Fermentation of Propionibacterium acnes, a
Commensal Bacterium in the Human Skin Microbiome, as Skin Probiotics against Methicillin-Resistant Staphylococcus aureus.PLoS ONE 2013, 8, e55380. [CrossRef] [PubMed]
242. Youn, S.H.; Choi, C.W.; Choi, J.W.; Youn, S.W. The skin surface pH and its different influence on the development of acne lesionaccording to gender and age. Ski. Res. Technol. 2012, 19, 131–136. [CrossRef]
243. Allhorn, M.; Arve-Butler, S.; Brüggemann, H.; Lood, R. A novel enzyme with antioxidant capacity produced by the ubiquitousskin colonizer Propionibacterium acnes. Sci. Rep. 2016, 6, 36412. [CrossRef] [PubMed]
244. Dreno, B.; Bagatin, E.; Blume-Peytavi, U.; Rocha, M.; Gollnick, H. Female type of adult acne: Physiological and psychologicalconsiderations and management. J. der Dtsch. Dermatol. Ges. 2018, 16, 1185–1194. [CrossRef]
245. Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.-M. Staphylococcus epidermidis in the humanskin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris.Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [CrossRef] [PubMed]
246. Nakamura, K.; O’Neill, A.M.; Williams, M.R.; Cau, L.; Nakatsuji, T.; Horswill, A.R.; Gallo, R.L. Short chain fatty acids producedby Cutibacterium acnes inhibit biofilm formation by Staphylococcus epidermidis. Sci. Rep. 2020, 10, 21237. [CrossRef] [PubMed]
247. Christensen, G.J.M.; Scholz, C.F.P.; Enghild, J.; Rohde, H.; Kilian, M.; Thürmer, A.; Brzuszkiewicz, E.; Lomholt, H.B.; Brüggemann,H. Antagonism between Staphylococcus epidermidis and Propionibacterium acnes and its genomic basis. BMC Genom. 2016, 17, 152.[CrossRef]
248. Belkaid, Y.; Tamoutounour, S. The influence of skin microorganisms on cutaneous immunity. Nat. Rev. Immunol. 2016, 16, 353–366.[CrossRef]
249. Naik, S.; Bouladoux, N.; Linehan, J.L.; Han, S.-J.; Harrison, O.J.; Wilhelm, C.; Conlan, S.; Himmelfarb, S.; Byrd, A.L.; Deming, C.;et al. Commensal–dendritic-cell interaction specifies a unique protective skin immune signature. Nat. Cell Biol. 2015, 520, 104–108.[CrossRef]
250. Wang, Z.; Mascarenhas, N.; Eckmann, L.; Miyamoto, Y.; Sun, X.; Kawakami, T.; Di Nardo, A. Skin microbiome promotes mast cellmaturation by triggering stem cell factor production in keratinocytes. J. Allergy Clin. Immunol. 2016, 139, 1205–1216.e6. [CrossRef]
251. SanMiguel, A.; Grice, E.A. Interactions between host factors and the skin microbiome. Cell Mol. Life Sci. 2014, 72, 1499–1515.[CrossRef]
252. Williams, M.R.; Gallo, R.L. The Role of the Skin Microbiome in Atopic Dermatitis. Curr. Allergy Asthma Rep. 2015, 15, 65.[CrossRef]
253. Sanford, J.A.; Zhang, L.-J.; Williams, M.R.; Gangoiti, J.A.; Huang, C.-M.; Gallo, R.L. Inhibition of HDAC8 and HDAC9 bymicrobial short-chain fatty acids breaks immune tolerance of the epidermis to TLR ligands. Sci. Immunol. 2016, 1, eaah4609.[CrossRef] [PubMed]
254. Brandwein, M.; Bentwich, Z.; Steinberg, D. Endogenous Antimicrobial Peptide Expression in Response to Bacterial EpidermalColonization. Front. Immunol. 2017, 8, 1637. [CrossRef] [PubMed]

Pathogens 2022, 11, 121 27 of 27
255. Lee, S.E.; Kim, J.-M.; Jeong, S.K.; Jeon, J.E.; Yoon, H.-J.; Jeong, M.-K.; Lee, S.H. Protease-activated receptor-2 mediates theexpression of inflammatory cytokines, antimicrobial peptides, and matrix metalloproteinases in keratinocytes in response toPropionibacterium acnes. Arch. Dermatol. Res. 2010, 302, 745–756. [CrossRef] [PubMed]
256. Donnarumma, G.; Perfetto, B.; Paoletti, I.; Oliviero, G.; Clavaud, C.; Del Bufalo, A.; Guéniche, A.; Jourdain, R.; Tufano, M.A.;Breton, L. Analysis of the response of human keratinocytes to Malassezia globosa and restricta strains. Arch. Dermatol. Res. 2014,306, 763–768. [CrossRef]
257. Ishibashi, Y.; Sugita, T.; Nishikawa, A. Cytokine secretion profile of human keratinocytes exposed to Malassezia yeasts. FEMSImmunol. Med. Microbiol. 2006, 48, 400–409. [CrossRef] [PubMed]
258. Yu, J.; Luo, Y.; Zhu, Z.; Zhou, Y.; Sun, L.; Gao, J.; Sun, J.; Wang, G.; Yao, X.; Li, W. A tryptophan metabolite of the skin mi-crobiotaattenuates inflammation in patients with atopic dermatitis through the aryl hydrocarbon receptor. J. Allergy Clin. Immunol. 2019,143, 2108–2119.e12. [CrossRef]
259. Kjer-Nielsen, L.; Patel, O.; Corbett, A.J.; Le Nours, J.; Meehan, B.; Liu, L.; Bhati, M.; Chen, Z.; Kostenko, L.; Reantragoon, R.; et al.MR1 presents microbial vitamin B metabolites to MAIT cells. Nature 2012, 491, 717–723. [CrossRef]
260. Sams-Dodd, J.; Sams-Dodd, F. Time to Abandon Antimicrobial Approaches in Wound Healing: A Paradigm Shift. Wounds 2018,30, 345–352.
261. Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl.Med. 2014, 6, 265sr6. [CrossRef]
262. Tomic-Canic, M.; Burgess, J.L.; O’Neill, K.E.; Strbo, N.; Pastar, I. Skin Microbiota and its Interplay with Wound Healing. Am. J.Clin. Dermatol. 2020, 21 (Suppl. 1), 36–43. [CrossRef]
263. Zeeuwen, P.L.; Boekhorst, J.; van den Bogaard, E.H.; de Koning, H.D.; van de Kerkhof, P.M.; Saulnier, D.M.; van Swam, I.I.; vanHijum, S.A.; Kleerebezem, M.; Schalkwijk, J.; et al. Microbiome dynamics of human epidermis following skin barrier disruption.Genome Biol. 2012, 13, R101. [CrossRef] [PubMed]
264. Heath, W.R.; Carbone, F.R. The skin-resident and migratory immune system in steady state and memory: Innate lymphocytes,dendritic cells and T cells. Nat. Immunol. 2013, 14, 978–985. [CrossRef] [PubMed]
265. ISO. ISO 17516:2014 Cosmetics—Microbiology—Microbiological Limits; ISO: Geneva, Switzerland, 2014.




















