
Kepekaan Erythricium salmonicolor Penyebab Penyakit Jamur...
37
ISSN: 0215-7950 35 Volume 13, Nomor 2, Maret 2017 Halaman 35–42 DOI: 10.14692/jfi.13.2.35 *Alamat penulis korespondensi: Balai Penelitian Tanaman Rempah dan Obat. Jalan Tentara Pelajar No. 3. Bogor 16111 Tel: 0251-8321879, Faks: 0251-8327010, Surel: [email protected] Kepekaan Erythricium salmonicolor Penyebab Penyakit Jamur Upas pada Kemiri Sunan terhadap Suhu, Cahaya, dan Fungisida Sensitivity of Erythricium salmonicolor, the Pink Disease Causal Agent of Reutealis trisperma, to Temperature, Light, and Fungicides Dini Florina 1 *, Dono Wahyuno 1 , Siswanto 2 1 Balai Penelitian Tanaman Rempah dan Obat, Bogor 16111 2 Pusat Penelitian dan Pengembangan Perkebunan, Bogor 16111 ABSTRAK Gejala khas penyakit jamur upas ditemukan pada tanaman kemiri sunan (Reutealis trisperma) di Bogor. Penelitian ini bertujuan menentukan cendawan penyebab dan kepekaannya terhadap suhu, cahaya, dan fungisida. Cendawan diidentifikasi berdasarkan karakter morfologi. Inokulum diperoleh dari isolasi jaringan kulit batang terinfeksi. Uji patogenisitas dilakukan dengan menempelkan potongan cabang kemiri sunan yang sudah terkolonisasi pada bibit kemiri sunan dan diinkubasi di rumah kaca. Pengaruh suhu terhadap cendawan dilakukan dengan menanam koloni cendawan uji pada medium agar- agar dekstrosa kentang (ADK) dan diinkubasi pada suhu 25–34 °C. Kepekaan cendawan pada cahaya diuji dengan menanam koloni cendawan di dalam labu berisi medium kaldu dekstrosa kentang (KDK) pada shaker (60 rpm) yang terpapar cahaya (400 luks) atau pada kondisi gelap selama 15 hari pada suhu 25 °C. Bobot segar dan kering miselium cendawan yang diperoleh ditimbang. Efikasi tembaga hidroksida dan mankozeb diuji dengan menanam cendawan uji dimedium ADK yang diteteskan fungisida uji dengan konsentrasi 100, 200, 300, 400 dan 500 ppm. Cendawan penyebab jamur upas pada kemiri sunan diidentifikasi sebagai Erythricium salmonicolor berdasarkan karakteristik morfologinya. Pada uji patogenisitas, miselium E. salmonicolor tumbuh dan membentuk jaring laba-laba pada bibit 3 minggu setelah inokulasi. Cendawan uji dapat tumbuh pada semua kisaran suhu yang diuji, tetapi suhu optimum pertumbuhan E. salmonicolor adalah 28 °C. Cahaya tidak berpengaruh nyata terhadap pertumbuhan E. salmonicolor. Mankozeb menekan pertumbuhan koloni E. salmonicolor lebih baik dibanding tembaga hidroksida. Kata kunci: identifikasi morfologi, mankozeb, tembaga hidroksida, uji patogenisitas ABSTRACT A typical pink disease symptom appeared on stem of Reutealis trisperma in Bogor. The present study was aimed for confirming the causal pathogen and testing its sensitivity to temperature, light and fungicides. The fungus was identified based on its morphological characteristics. The fungal inoculum was obtained by tissue isolation of infected stem bark. For pathogenicity test, the fungus was inoculated onto sterilized pieces of woody R. trisperma branch, then fastened on the seedlings and incubated in the green house. The effect of temperatures on fungal growth was tested by planting the fungal colony onto potato dextrose agar (PDA) then incubated at 25–34 °C. The sensitivity of the fungus to light was tested by inoculating the fungus into flask with potato dextrose broth (PDB) medium on a shaker (60 rpm) exposing it to white light tube (400 luks) or in a dark condition for 15 days at 25 °C. The obtained
-
Upload
hoangkhanh -
Category
Documents
-
view
221 -
download
0
Transcript of Kepekaan Erythricium salmonicolor Penyebab Penyakit Jamur...

ISSN: 0215-7950
35
Volume 13, Nomor 2, Maret 2017Halaman 35–42
DOI: 10.14692/jfi.13.2.35
*Alamat penulis korespondensi: Balai Penelitian Tanaman Rempah dan Obat. Jalan Tentara Pelajar No. 3. Bogor 16111Tel: 0251-8321879, Faks: 0251-8327010, Surel: [email protected]
Kepekaan Erythricium salmonicolor Penyebab Penyakit Jamur Upas pada Kemiri Sunan terhadap Suhu, Cahaya, dan Fungisida
Sensitivity of Erythricium salmonicolor, the Pink Disease Causal Agent of Reutealis trisperma, to Temperature, Light, and Fungicides
Dini Florina1*, Dono Wahyuno1, Siswanto2
1Balai Penelitian Tanaman Rempah dan Obat, Bogor 161112Pusat Penelitian dan Pengembangan Perkebunan, Bogor 16111
ABSTRAK
Gejala khas penyakit jamur upas ditemukan pada tanaman kemiri sunan (Reutealis trisperma) di Bogor. Penelitian ini bertujuan menentukan cendawan penyebab dan kepekaannya terhadap suhu, cahaya, dan fungisida. Cendawan diidentifikasi berdasarkan karakter morfologi. Inokulum diperoleh dari isolasi jaringan kulit batang terinfeksi. Uji patogenisitas dilakukan dengan menempelkan potongan cabang kemiri sunan yang sudah terkolonisasi pada bibit kemiri sunan dan diinkubasi di rumah kaca. Pengaruh suhu terhadap cendawan dilakukan dengan menanam koloni cendawan uji pada medium agar-agar dekstrosa kentang (ADK) dan diinkubasi pada suhu 25–34 °C. Kepekaan cendawan pada cahaya diuji dengan menanam koloni cendawan di dalam labu berisi medium kaldu dekstrosa kentang (KDK) pada shaker (60 rpm) yang terpapar cahaya (400 luks) atau pada kondisi gelap selama 15 hari pada suhu 25 °C. Bobot segar dan kering miselium cendawan yang diperoleh ditimbang. Efikasi tembaga hidroksida dan mankozeb diuji dengan menanam cendawan uji dimedium ADK yang diteteskan fungisida uji dengan konsentrasi 100, 200, 300, 400 dan 500 ppm. Cendawan penyebab jamur upas pada kemiri sunan diidentifikasi sebagai Erythricium salmonicolor berdasarkan karakteristik morfologinya. Pada uji patogenisitas, miselium E. salmonicolor tumbuh dan membentuk jaring laba-laba pada bibit 3 minggu setelah inokulasi. Cendawan uji dapat tumbuh pada semua kisaran suhu yang diuji, tetapi suhu optimum pertumbuhan E. salmonicolor adalah 28 °C. Cahaya tidak berpengaruh nyata terhadap pertumbuhan E. salmonicolor. Mankozeb menekan pertumbuhan koloni E. salmonicolor lebih baik dibanding tembaga hidroksida.
Kata kunci: identifikasi morfologi, mankozeb, tembaga hidroksida, uji patogenisitas
ABSTRACT
A typical pink disease symptom appeared on stem of Reutealis trisperma in Bogor. The present study was aimed for confirming the causal pathogen and testing its sensitivity to temperature, light and fungicides. The fungus was identified based on its morphological characteristics. The fungal inoculum was obtained by tissue isolation of infected stem bark. For pathogenicity test, the fungus was inoculated onto sterilized pieces of woody R. trisperma branch, then fastened on the seedlings and incubated in the green house. The effect of temperatures on fungal growth was tested by planting the fungal colony onto potato dextrose agar (PDA) then incubated at 25–34 °C. The sensitivity of the fungus to light was tested by inoculating the fungus into flask with potato dextrose broth (PDB) medium on a shaker (60 rpm) exposing it to white light tube (400 luks) or in a dark condition for 15 days at 25 °C. The obtained

J Fitopatol Indones Florina et al
36
fungal colonies were weighed. Efficacy of copper hydroxide and mancozeb against the fungal colony growth was tested by inoculating the fungus onto PDA amended with 100, 200, 300, 400, and 500 ppm, respectively, of tested fungicides. The results showed that the causal pathogen was Erythricium salmonicolor. In the pathogenicity test, cobwebs of E. salmonicolor were formed on inoculated seedlings 3 weeks after inoculation. The optimum temperature for growth of E. salmonicolor was 28 °C. There was no significant effect of light on fungal growth. Mancozeb suppressed colony of E. salmonicolor growth better than copper hydroxide.
Keywords: morphological identification, mancozeb, copper hydroxide, pathogenicity test
PENDAHULUAN
Kemiri sunan (Reutealis trisperma; syn. Aleurites trisperma) merupakan tanaman asli Filipina yang telah beradaptasi luas di Indonesia (Aguilar dan Oyen 2002) dan mampu menghasilkan minyak nabati yang dapat diproses untuk sumber energi terbarukan. Koleksi tanaman kemiri sunan di petak koleksi Pusat Penelitian dan Pengembangan Perkebunan Cimanggu, Bogor menunjukkan gejala terserang jamur upas (pink disease) (Gambar 1a) yang dicirikan adanya tepi koloni cendawan berwarna putih terang. Koloni cendawan patogen yang menyerang batang utama (Gambar 1b) dan cabang (Gambar 1c) tanaman kemiri sunan terlihat tebal dan berwarna putih. Di Afrika Selatan jamur upas yang disebabkan oleh cendawan Erythricium salmonicolor (syn. Corticium salmonicolor) menjadi penyakit utama karena memiliki inang yang luas (Roux dan Coutzee 2005). Pada tanaman kemiri sunan, penyakit jamur upas belum pernah dilaporkan. Penelitian ini bertujuan memastikan patogen penyebab penyakit jamur upas pada tanaman kemiri sunan serta menentukan apakah suhu,cahaya, dan fungisida berpengaruh terhadap pertumbuhannya.
BAHAN DAN METODE
Insidensi, Isolasi, dan Identifikasi PatogenPengamatan penyakit tanaman dilakukan
di kebun koleksi Pusat Penelitian dan Pengembangan Perkebunan di Bogor dari bulan Maret sampai Agustus 2016. Populasi tanaman yang diamati berjumlah 18 tanaman berumur ± 5 tahun, jarak tanam 6 m × 6 m.
Insidensi penyakit (IP) dan keparahan penyakit (KP) diukur menggunakan rumus:
IP = nN × 100%, dengan
n, jumlah tanaman yang terserang; dan N, jumlah tanaman yang diamati.
N × VKP = × 100%, dengan(ni × vi)∑
i
i=0
ni, jumlah tanaman dengan skor ke-i; vi, nilai skor penyakit dari I = 0,1,2 sampai i t-skor tertinggi; N, jumlah tanaman yang diamati; dan V, skor tertinggi.
Kategori kerusakan menggunakan kriteria Griffin et al. (2003), yaitu: 0, tanaman sehat; 1, koloni pada cabang primer, kanopi masih 75%; 2, koloni pada batang utama, terdapat retakan pada kulit, kanopi 25–75%; 3, koloni pada batang utama dan cabang primer, meranting, kanopi 25%; dan 4, tanaman mati.
Isolasi patogen dilakukan dengan merendam potongan kulit pohon terinfeksi ke dalam alkohol 70% selama 2–3 menit, dibilas dengan air steril dan dikeringanginkan (Akrofi et al. 2014), kemudian diinokulasikan pada medium agar-agar dekstrosa kentang (ADK) dan diinkubasikan pada suhu 25–27 °C selama 15 hari. Koloni berwarna putih sampai oranye dimurnikan pada medium ADK dan diberi kode EK-01. Ada tidaknya spora seksual dan atau aseksual diamati secara mikroskopi. Selain itu, morfologi cendawan juga diamati dari miselium yang diambil dari permukaan kulit batang tanaman terinfeksi dan membuat preparat menggunakan asam laktat yang diberi pewarna biru kapas sebagai mounting (Gambar 1d). Karakter morfologi (seksual dan aseksual) cendawan patogen EK-01 yang tumbuh di permukaan kulit batang kemiri

J Fitopatol Indones Florina et al
37
sunan dan miselium yang diambil dari biakan ADK didentifikasi berdasarkan kriteria Jülich (1975).
Cendawan EK-01 ditumbuhkan pada medium ADK berisi potongan ranting kemiri sunan steril (± 5 cm panjang; berdiameter ± 1 cm), diinkubasikan pada suhu 28 °C selama 3 minggu. Potongan ranting yang sudah terkolonisasi ditempelkan pada batang bibit kemiri sunan berumur ± 1 tahun dengan jarak ± 10 cm dari permukaan tanah (Akrofi et al. 2014). Perlakuan diulang sebanyak 3 kali, masing-masing terdiri atas 3 tanaman. Sebagai kontrol digunakan potongan ranting yang tidak diinokulasi cendawan uji sehingga total tanaman yang digunakan adalah 12 tanaman.
Pengujian Suhu, Cahaya, dan FungisidaPengujian pengaruh suhu dilakukan
dengan menumbuhkan cendawan EK-01 pada medium ADK selama 5 hari pada suhu kamar (25–29 °C). Bagian tepi koloni cendawan diambil (± 5 mm), ditanam pada medium ADK di cawan petri, kemudian diinkubasi pada suhu 25, 28, 31, dan 34 °C serta diulang 5 kali. Peubah yang diamati ialah pertumbuhan koloni dengan mengukur diameter koloni secara tegak lurus untuk setiap cawan petri kemudian dirata-ratakan.
Pengaruh cahaya terhadap kecepatan tumbuh koloni diuji dengan meletakkan cawan petri yang berisi koloni EK-01 di bawah lampu (± 400 luks) atau diletakkan pada tempat gelap untuk perlakuan tanpa pencahayaan pada suhu inkubasi 28 °C. Setiap perlakuan diulang 5 kali. Peubah yang diamati ialah diameter koloni seperti pada uji suhu. Pengaruh cahaya terhadap bobot koloni dilakukan dalam labu dengan medium kaldu dekstrosa kentang (KDK) dengan kecepatan kocok 60 rpm. Koloni cendawan dipanen pada hari ke-15, disaring, dikeringkan dan ditimbang.
Pengujian fungisida berbahan aktif tembaga hidroksida dan mankozeb dilakukan dengan metode peracunan makan (Valarmathi et al. 2013), yaitu menumbuhkan potongan koloni cendawan uji (diameter ± 5 mm) pada medium ADK di cawan petri yang sudah
ditetesi fungisida terlebih dahulu, kemudian diinkubasi pada suhu 28 °C. Konsentrasi bahan aktif fungisida ialah 0 (kontrol), 100, 200, 300, 400 dan 500 ppm. Setiap perlakuan diulang 4 kali dan peubah yang diamati ialah diameter koloni.
HASIL
Pengamatan lapanganInsidensi penyakit tanaman kemiri sunan
terinfeksi jamur upas pada awal bulan Maret ialah 27.7% dan nilainya stabil hingga pengamatan pada bulan Agustus 2016. Tingkat kerusakannya bervariasi mulai skala 0 sampai 3.Keparahan penyakit pada bulan Maret 2016 ialah 9.72% dan berkembang hingga 18.06% pada bulan Agustus 2016. Kondisi kanopi sudah saling bersentuhan dan rapat. Tanaman kemiri sunan yang termasuk kedalam skala 3 daunnya menguning, rontok dan meranggas.
Pada himenium (Gambar 1e) yang diambil dari permukaan kulit batang terinfeksi ditemukan basidium berbentuk gada, hialin dengan 4 sterigma di atasnya. Basidiospora berbentuk bulat sampai lonjong dengan apiculus di ujungnya dan jernih (Gambar 1f dan 1g). Pada koloni yang tebal dan tua, ditemukan konidium yang terbentuk dari fragmentasi hifa (Gambar 1h).
Koloni yang tumbuh pada medium ADK berwarna putih tipis di bagian tepi, di bagian tengah tebal berwarna putih-oranye terang (Gambar 1i). Diameter koloni mencapai 73–77 mm setelah diinkubasi selama 5 hari pada 28 °C.
Pada uji patogenisitas, miselium cendawan dari subtrat kayu tumbuh di permukaan batang bibit kemiri sunan berwarna putih tebal hingga oranye setelah diinkubasi selama 3 minggu (Gambar 1j).
Berdasarkan karakteristik warna koloni,miselium yang membentuk jalinan himenium, basidium yang berbentuk gada, dan basidio-spora yang berbentuk bulat sampai lonjong dengan apiculus di ujungnya, patogen penyebab penyakit jamur upas diidentifikasi sebagai cendawan Erythricium salmonicolor.

J Fitopatol Indones Florina et al
38
Pertumbuhan E. salmonicolor pada Perlakuan Suhu, Cahaya dan Fungisida
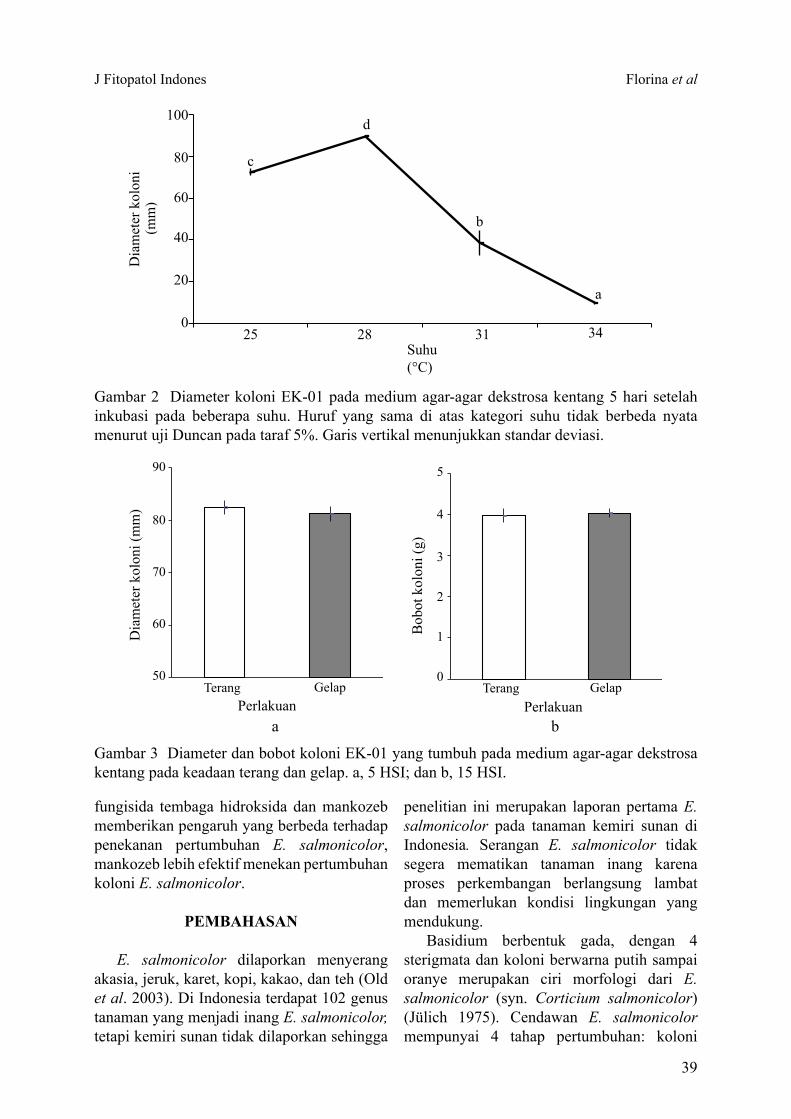
Miselium tumbuh pada kisaran suhu 25-34 °C. Suhu 28 °C merupakan suhu optimum bagi pertumbuhan miselium E. salmonicolor (Gambar 2). Cendawan yang tumbuh pada suhu 25 °C dan 28 °C tumbuh tebal seperti kapas, tetapi tidak menghasilkan basidium maupun konidium pada 21 hari setelah inokulasi.
Pemberian cahaya tidak menyebabkan koloni E. salmonicolor tumbuh lebih cepat dan lebih subur dibandingkan pada kondisi gelap (Gambar 3a dan 3b). Koloni yang diberi perlakuan cahaya menghasilkan warna koloni yang lebih terang dibandingkan yang tidak terpapar cahaya.
Kedua fungisida yang diuji (mankozeb dan tembaga hidroksida) efektif menekan E. salmonicolor. Mankozeb menekan pertumbuhan koloni E. salmonicolor lebih tinggi dibandingkan dengan tembaga hidroksida (Gambar 4). Konsentrasi mankozeb 100 ppm lebih efektif menekan pertumbuhan koloni E. salmonicolor dibandingkan dengan 500 ppm tembaga hidroksida.
Analisa regresi menunjukkan perbedaan yang nyata pada setiap kenaikan konsentrasi 100 ppm tembaga hidroksida terhadap daya hambat pertumbuhan E. salmonicolor. Sebaliknya, mankozeb tidak menunjukkan perbedaan yang nyata diantara konsentrasi yang diuji pada E. salmonicolor (Gambar 5).Uji homogenitas pada kedua persamaan
Gambar 1. Erythricium salmonicolor pada tanaman kemiri sunan. a, Gejala dan tanda tanaman terserang; b, Koloni pada batang; c, Koloni pada cabang; d, Miselium yang membentuk titik-titik warna terang (pustule); e, Hymenium; f, Basidium (→); g, Basidiospora; h, Konidium (→); i, Koloni (EK-01) pada medium ADK; dan j, Miselium tumbuh pada permukaan batang saat inokulasi buatan.
a b c
d e f
5 μm
g h i j
5 μm 5 μm
J Fitopatol Indones Florina et al
39
Gambar 2 Diameter koloni EK-01 pada medium agar-agar dekstrosa kentang 5 hari setelah inkubasi pada beberapa suhu. Huruf yang sama di atas kategori suhu tidak berbeda nyata menurut uji Duncan pada taraf 5%. Garis vertikal menunjukkan standar deviasi.
0
20
40
60
80
100
25 28 31 34
Dia
met
er k
olon
i(m
m)
Suhu(°C)
100
80
60
40
20
025 28 31 34
a
b
c
d
Gambar 3 Diameter dan bobot koloni EK-01 yang tumbuh pada medium agar-agar dekstrosa kentang pada keadaan terang dan gelap. a, 5 HSI; dan b, 15 HSI.
50
60
70
80
90
Terang Gelap
Dia
met
er k
olon
i (m
m)
PerlakuanTerang Gelap
90
80
70
60
50 0
1
2
3
4
5
Terang Gelap
Bob
ot k
olon
i (g)
Perlakuan
5
4
3
2
1
0Terang Gelap
fungisida tembaga hidroksida dan mankozeb memberikan pengaruh yang berbeda terhadap penekanan pertumbuhan E. salmonicolor, mankozeb lebih efektif menekan pertumbuhan koloni E. salmonicolor.
PEMBAHASAN
E. salmonicolor dilaporkan menyerang akasia, jeruk, karet, kopi, kakao, dan teh (Old et al. 2003). Di Indonesia terdapat 102 genus tanaman yang menjadi inang E. salmonicolor, tetapi kemiri sunan tidak dilaporkan sehingga
penelitian ini merupakan laporan pertama E. salmonicolor pada tanaman kemiri sunan di Indonesia. Serangan E. salmonicolor tidak segera mematikan tanaman inang karena proses perkembangan berlangsung lambat dan memerlukan kondisi lingkungan yang mendukung.
Basidium berbentuk gada, dengan 4 sterigmata dan koloni berwarna putih sampai oranye merupakan ciri morfologi dari E. salmonicolor (syn. Corticium salmonicolor) (Jülich 1975). Cendawan E. salmonicolor mempunyai 4 tahap pertumbuhan: koloni
a b

J Fitopatol Indones Florina et al
40
tumbuh cepat dan tipis menyerupai sarang laba-laba, fase infeksi ditandai adanya miselium yang mengumpul, tebal dan sporadis (pustule), terjadinya kerak atau pecah pada kulit batang terinfeksi (pink incrustation), dan fase pembentukan konidium (necator) (Old et al. 2003).
Cendawan E. salmonicolor (EK-01) mempunyai suhu pertumbuhan optimum 28 °C. Hal serupa juga dilaporkan oleh
Shamsuri et al. (1997) pada tanaman karet, sedangkan kisaran suhu untuk pertumbuhan ialah 5–40 °C. Di Jepang, miselium E. salmonicolor pada sembilan inang yang berbeda menunjukkan pola pertumbuhan yang sama, yaitu tumbuh optimum pada 25 °C,lebih rendah dibandingkan dengan E. salmonicolor yang tumbuh di daerah tropis (Nomi dan Kobayashi 2000).
Gambar 5 Kepekaan Erythricium salmonicolor pada jenis dan konsentrasi fungisida. , Mankozeb; dan , Tembaga hidroksida.
0
10
20
30
40
50
60
70
80
90
0 100 200 300 400 500
Dia
met
er k
olon
i (m
m)
90
80
70
60
50
40
30
20
10
0
Konsentrasi (ppm)0 100 200 300 400 500
Y = -0.097x + 70.363R2 = 0.9164
Y = -0.0983x + 48.792R2 = 0.4858
Gambar 4 Diameter koloni EK-01 yang tumbuh pada medium agar-agar dekstrosa kentang yang sudah diberi perlakuan fungisida dan diinkubasi selama 5 hari. Huruf yang sama pada tiap grafik batang tidak berbeda nyata menurut uji Duncan pada taraf 5%. , Kontrol; , Fungisida mankozeb; dan , Tembaga hidroksida pada konsentrasi 100–500 ppm.
0
10
20
30
40
50
60
70
80
90
0 100 200 300 400 500 100 200 300 400 500
Dia
met
er k
olon
i (m
m)
90
80
70
60
50
40
30
20
10
0
Konsentrasi (ppm)0 100 200 300 400 500 100 200 300 400 500
g
f
e
dc c
abb
a a a

J Fitopatol Indones Florina et al
41
Cahaya dengan intensitas 400 luks tidak berpengaruh nyata terhadap pertumbuhan dan bobot koloni E. salmonicolor. Di awal pengamatan lapangan, koloni E. salmonicolor tumbuh lebih panjang dengan warna lebih terang pada sisi timur batang primer kemiri sunan. Kondisi lingkungan yang lembap dan curah hujan tinggi pada bulan Maret, serta kanopi tanaman yang lebat tidak menyebabkan terjadinya kenaikan suhu yang tinggi pada permukaan batang primer yang terserang. Sebaliknya pada musim kemarau, koloni E. salmonicolor pada ranting di bagian atas, cenderung tumbuh di bagian bawah ranting yang terlindung dari sinar matahari. Hal ini mengindikasikan, pengaruh cahaya lebih besar pada kenaikan suhu medium sehingga menghambat pertumbuhan koloni, dibandingkan dengan pengaruhnya secara langsung pada cendawan. Alam et al. (2001) berpendapat bahwa perlakuan cahaya tidak berpengaruh nyata terhadap kecepatan tumbuh koloni tetapi berpengaruh terhadap daya sporulasi Botryodiplodia theobromae. Respons Cerrena unicolor, Pycnoposrus sanguineus dan Phlebialindtneri (Polyporales) untuk menghasilkan enzim-enzim pendegradasi kayu, tidak sama terhadap panjang gelombang cahaya yang diberikan. C. unicolor dan P. sanguineus membutuhkan warna biru untuk meningkatkan sintesis enzim lakase, tetapi P lindtneri membutuhkan warna hijau (Janusz et al. 2016).
Fungisida berbahan aktif tembaga hidroksida dan mankozeb banyak digunakan untuk mengendalikan penyakit tanaman yang disebabkan oleh cendawan. Perbedaan bahan aktif fungisida menyebabkan mankozeb lebih efektif menekan pertumbuhan E. salmonicolor dibandingkan dengan tembaga hidroksida. Pada konsentrasi 1000 ppm, fungisida mankozeb lebih efektif daripada fungisida berbahan aktif tembaga dalam melindungi tanaman buah-buahan dari Erwinia amylovora, Pseudomonas syringae dan Agrobacterium tumefaciens di rumah kaca (Mikicinski et al. 2012).
Tembaga hidroksida relatif tidak berbahaya untuk agens hayati (2500 ppm untuk Trichoderma) sehingga dalam penggunaannya
di lapangan dapat dikombinasikan (Valarmathi et al. 2013). Ion tembaga merupakan racun kontak, mempunyai afinitas pada berbagai kelompok senyawa protein dan enzim (Okorski et al. 2015). Fungisida tembaga efektif mengendalikan E. salmonicolor pada pohon karet dan efektifitasnya meningkat dalam ukuran nano (Cao et al. 2014).
Mankozeb merupakan racun kontak yang menghambat perkecambahan spora berbagai kelas cendawan (Gullino et al. 2010) dan efektif sebagai protektan atau sebelum terjadi infeksi (Wong dan Wilcox 2001). Setelah terjadi kontak dengan air, mankozeb melepas ethylen bisisothiocyanate sulfidae (EBIS), terurai lagi menjadi ethylen bisisothiocyanate (EBI) yang beracun dan merusak sistem enzim di dalam sitoplasma dan mitokondria sel cendawan (Gullino et al. 2010).
Cendawan E. salmonicolor umum ditemukan di area bercurah hujan tinggi, tersebar melalui basidiospora maupun konidium dan menginfeksi jaringan kulit tanaman (Lee 2003). Musim hujan yang panjang di Bogor pada tahun ini menciptakan kondisi ideal bagi E. salmonicolor untuk berkembang. Tindakan budi daya berupa pemangkasan, sanitasi, dan pengaturan jarak tanam kemiri sunan serta aplikasi fungisida apabila diperlukan berguna dalam menekan serangan E. salmonicolor.
DAFTAR PUSTAKA
Alam MS, Begum MF, Sarkar MA, Islam MR, Alam MS. 2001. Effect of temperature, light and media on growth, sporulation, formation of pigments and pycnidia of Botryodiplodia theobromae Pat. Pakistan J Biol Sci. 4(10):1224-1227. DOI: https://doi.org/10.3923/pjbs.2001.1224.1227.
Aguilar NO, Oyen LPA. 2002. Reutealis trisperma (Blanco). Di dalam: Van der Vossen HAM, Umali BE, editor. Plant Resources of South-East Asia No. 14. Vegetable Oils and Fats. Bogor (ID): Prosea Bogor Indonesia. hlm 112–115.
Akrofi AY, Attal A, Assuah M, Kumi-Asare E. 2014. Pink disease caused by Erythricium

J Fitopatol Indones Florina et al
42
salmonicolor (Berk & Broome) Burdsall: an epidemiological assessment of its potential effect on cocoa production in Ghana. J Plant Pathol Microb. 5:215–220 DOI: https://doi.org/10.4172/2157-7471.1000215.
Cao VD, Nguyen PP, Khuong VQ, Nguyen CK, Nguyen XC, Dang CH, Tran NQ. 2014. Ultrafine copper nanoparticles exhibiting a powerful antifungal/killing activity against Corticium salmonicolor. Bull Korean Chem Soc. 35(9):2645–2648. DOI: https://doi.org/10.5012/bkcs.2014.35.9.2645.
Griffin JM, Lovett GM, Arthur MA, Weathers KC. 2003. The distribution and severity of beech bark disease in the Catskill Mountains, NY. Can J Forest Res. 33:1754–1760. DOI: https://doi.org/10.1139/x03-093.
Gullino ML, Tinivella F, Garibaldi A, Kemmit GM, Bacci L, Sheppard B. 2010. Mancozeb past, present and future. Plant Dis. 94(9):1076–1087. DOI: https://doi.org/10.1094/PDIS-94-9-1076.
Janusz G, Sulej J, Jaszek M, Jaroszuk MO. 2016. Effect of different wavelengths of light on laccase, cellobiose dehydrogenase, and proteases produced by Cerrena unicolor, Pycnoporus sanguineus and Phlebia lindtneri. Acta Biochimica Polinica. 63(2):22–228. DOI: 10.18388/abp.2015_1235. DOI: https://doi.org/10.18388/abp.2015_1235.
Jülich W. 1975. Studies in resupinate Basidiomycetes-III. Persoonia. 8(3):291–305.
Lee S. 2003. Pathology of tropical hardwood plantations in South-East Asia. New Zealand J Forestry Sci. 33(3):321–335.
Mikicinski A, Sobiczewski P, Berczynski S. 2012. Efficacy of fungicides and essential
oils against bacterial disease of fruit trees. J Plant Protec Res. 52(4):467–471. DOI: https://doi.org/10.2478/v10045-012-0075-7.
Nomi CS, H Kobayashi, T. 2000. Occurence and life-history of pink disease caused by Erythricium salmonicolor in Northern Kanto, Honshu, Japan. J Agric Sci Tokyo Nogyo Daigaku. 44(4):252–262.
Old KM, Wingfield MJ, Yuan ZQ. 2003. A Manual of Diseases of Eucalypts in South-East Asia. Bogor (ID): ACIAR CIFOR.
Okorski A, Pszczolkowska A, Oszaco T, Nowakowska JA. 2015. Current possibilities and prospects of using fungicides in forestry. Forest Res Papers. 76(2):191–206. DOI: https://doi.org/10.1515/frp-2015-0019.
Roux J, Coetzee MPA. 2005. First report of pink disease on native trees in South Africa and phylogenetic placement of Erythricium salmonicolor in the homobasidiomycetes. Plant Dis. 89(11):1158–1163. DOI: https://doi.org/10.1094/pd-89-1158.
Shamsuri MH, Omar M, Napi D. 1997. Studies on epidemiology of pink disease and the effect of temperature on mycelial growth of Corticium salmonicolor of Hevea rubber. J Nat Rubb Res. 12(1):58–66.
Valarmathi P, Kumar PS, Vanaraj P, Ramalingam R, Gopal C. 2013. Compatibility of copper hydroxide (Kocide 3000) with biocontrol agents. IOSR-JAVS. 3(6):28–31. DOI: https://doi.org/10.9790/2380-0362831.
Wong FP, Wilcox WF. 2001. Comparative physical modes of action of azoxystrobin, mancozeb, and metalaxyl against Plasmopora viticola (grapevine downey mildew). Plant Dis. 85(6):649–656. DOI: https://doi.org/10.1094/PDIS.2001.85.6.649.

ISSN: 0215-7950
43
Volume 13, Nomor 2, Maret 2017Halaman 43–50
DOI: 10.14692/jfi.13.2.43ISSN: 0215-7950
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Jalan Kamper, Kampus IPB, Dramaga, Bogor 16680 Tel: 0251-7533525, Faks : 0251-8629364, Surel: [email protected]
Cara Preservasi Fitoplasma dari Jaringan Kacang Tanah Bergejala Sapu untuk Deteksi DNA dengan Teknik PCR
Phytoplasma Preservation Methods of Symptomatic Peanut Witches‘ broom for DNA Detection Using PCR Technique
Siska Irhamnawati Pulogu, Kikin Hamzah Mutaqin*, GiyantoInstitut Pertanian Bogor, Bogor 16680
ABSTRAK
Penyakit sapu (witches' broom) oleh fitoplasma pada kacang tanah adalah penyakit umum di Indonesia. Fitoplasma dapat dideteksi dengan teknik polymerase chain reaction (PCR). Salah satu faktor penentu keberhasilan deteksi fitoplasma ialah penyediaan DNA dari contoh segar. Penelitian bertujuan mengevaluasi beberapa cara preservasi sampel tanaman terinfeksi fitoplasma. Aspek yang dievaluasi meliputi waktu (1, 2, 3, dan 4 minggu), suhu (-20 °C, 4 °C, dan 25 °C), dan medium preservasi (bufer PGB 1X, NaCl 3 M, bufer CTAB, Alkohol 70%, tanpa medium, dan kertas FTA) untuk mempertahankan jaringan tanaman terinfeksi supaya tetap segar. Cara preservasi yang baik akan mengoptimalkan deteksi DNA fitoplasma dengan teknik PCR standar yang dilanjutkan dengan teknik nested-PCR. Penyimpanan pada suhu -20 °C, 4 °C dan 25 °C dalam medium CTAB dapat mempertahankan jaringan tetap segar selama 4 minggu dengan kualitas dan kuantitas DNA yang cukup untuk deteksi dengan teknik PCR. Teknik PCR standar dengan pasangan primer P1/P7 menunjukkan bahwa tidak semua DNA fitoplasma dari hasil preservasi contoh terdeteksi positif. Namun, PCR standar yang dilanjutkan dengan nested-PCR menggunakan primer fU5/rU5 mampu meningkatkan pendeteksian fitoplasma yang berasal dari preservasi contoh pada berbagai medium selama 4 minggu dengan memberikan hasil positif dari contoh yang terdeteksi negatif pada teknik PCR standar.
Kata kunci: nested-PCR, PCR standar, penyimpanan, sapu kacang tanah
ABSTRACT
Witches‘ broom of peanut caused by phytoplasma is a common disease found in Indonesia. Phytoplasma is able to be detected using polymerase chain reaction (PCR) technique. One of important factor which determine the successful of phytoplasma amplification is the DNA availability from fresh tissues. The research was aimed to evaluate some preservation methods of phytoplasma from infected plant samples. The aspects to be evaluated consisted of time (1, 2, 3, and 4 weeks), temperature (-20 °C, 4 °C, and 25 °C), and preservation medium (1X PGB buffer, 3 M NaCl, CTAB buffer, 70% ethanol, non medium, and FTA-card) for storing the fresh phytoplasma infected samples. Good preservation method will optimize the phytoplasma DNA amplification using PCR standard technique followed by nested-PCR. The results showed that preservation of samples at -20 °C, 4 °C, and 25 °C in CTAB buffer was able to maintain the tissue freshness for 4 weeks and was able to provide the DNA of either quality or quantity sufficiently for PCR detection. PCR standard using a primer pair P1/P7 showed that not all of the preserved DNA of phytoplasma were amplified positively. However, standard PCR followed by nested-PCR using primer pair fU5/rU3 was able to increase the DNA detectability. Preserved samples

J Fitopatol Indones Pulogu et al
44
derived from various medium and stored for 4 weeks gave positive results. This results were in contrary with previous same samples which were detected negatively by standard PCR technique.
Keywords: nested-PCR, preservation, peanut witches’ broom, standard PCR
PENDAHULUANDi Indonesia fitoplasma telah menginfeksi
beberapa tanaman penting seperti kacang tanah yang dapat menurunkan bobot polong sebesar 41-100% (Nugroho et al. 2000). Selain itu, fitoplasma terdeteksi menyebabkan penyakit daun putih rumput bermuda, penyakit kuning bambu, penyakit daun kecil ubi jalar, dan penyakit sapu/proliferasi mosaik kaktus (Mutaqin et al. 2003).
Polymerase Chain Reaction (PCR) ialah teknik molekuler yang umum digunakan dalam deteksi dan identifikasi fitoplasma. Penyediaan contoh tanaman yang tetap segar sangat penting untuk isolasi DNA total. Faktor jarak jauh atau waktu yang lama dalam pengiriman contoh segar menjadi kendala dalam isolasi DNA karena deteriorasi jaringan dapat terjadi secara cepat dan munculnya senyawa inhibitor yang dapat memengaruhi kualitas dan kuantitas DNA dalam syarat teknik PCR (Nejat dan Vadamalai 2013). Titer fitoplasma dalam jaringan sangat rendah sehingga DNA yang diisolasi seringkali belum cukup dalam PCR standar untuk menghasilkan amplikon yang terlihat jelas. Oleh karena itu, modifikasi PCR untuk meningkatkan kemampuan deteksinya dengan PCR bersarang (nested-PCR) menggunakan pasangan primer untuk reamplifikasi DNA sasaran secara internal dalam wilayah sasaran PCR standar diperlukan (Gundersen dan Lee 1996). Penelitian bertujuan mengevaluasi cara preservasi jaringan tanaman bergejala sapu sehingga tetap diperoleh DNA yang layak dalam deteksi fitoplasma dengan PCR standar yang dilanjutkan dengan nested-PCR.
BAHAN DAN METODE
Penyediaan dan Preservasi Tanaman SakitTanaman kacang tanah bergejala penyakit
sapu diperoleh di Desa Cikarawang, Kecamatan Dramaga, Kabupaten Bogor. Tanaman tersebut
diuji dengan PCR untuk memastikan terinfeksi oleh fitoplasma. Sebanyak 1 g contoh tanaman sakit berupa potongan batang muda, tangkai, dan tulang daun tanpa nekrosis disimpan dalam botol. Preservasi dilakukan dalam tiga aspek, yaitu lama penyimpanan (1, 2, 3, 4 minggu), suhu penyimpanan (-20, 4, dan 25 °C), dan medium penyimpanan (bufer PGB1X [47.4 mM K2HPO4.3H2O; 15 mM KH2PO4; Sukrosa 5%; Polyvinylpyrrolidone-10 1%], NaCl 3 M, bufer CTAB [CTAB 2%; 1.4 MNaCl; 100 mM Tris; 20 mM EDTA; Polyvinylpyrrolidone-40 1%], Alkohol 70%,tanpa medium, dan kertas Flinders Technology Associates (FTA) sebagai pembanding. Preservasi dengan FTA-card (Whatman) dilakukan melalui blotting jaringan contoh sebanyak 0.1 g pada permukaan FTA-card, diinkubasi selama 5 menit pada suhu ruang dan selanjutnya disimpan pada suhu -20 °C, 4 °C, dan 25 °C.
Isolasi DNA dari Jaringan Tanaman Sakit dan Pengukuran DNA
Isolasi DNA secara total dari jaringan tanaman sakit dilakukan dengan metode Dellaporta et al. (1983). DNA total hasil ekstraksi maupun preservasi pada FTA-card diukur pada absorbansi 260/280 menggunakan spektrofotometer Nanodrop 2000 (Thermo Scientific) di IPB Culture Collection, Departemen Biologi.
Amplifikasi DNA dengan PCR dan Nested-PCR
Deteksi fitoplasma dengan PCR standar menggunakan pasangan primer P1/P7. Amplikon DNA hasil PCR standar (pengenceran 1:29) selanjutnya digunakan sebagai DNA templat dalam nested-PCR menggunakan pasangan primer fU5 (5’-CGG CAA TGG AGG AAA CT-3’)/rU3 (5’-TTC AGC TAC TCT TTG TAA CA-3’) (Lorenz et al. 1995). Reaksi PCR pada volume total

J Fitopatol Indones Pulogu et al
45
25 µL terdiri atas DreamTaq Green PCR master mix 2X (Thermo Scientific) 12.5 µL, primer forward dan reverse 1 µL (5 pmol), ddH2O 9.5 µL, dan DNA templat 1 µL (1–2000 ng µL-1).Aplikasi DNA dengan teknik PCR menggunakan mesin GeneAmp PCR System 9700 dengan kondisi denaturasi awal 92 °C, 1 menit; denaturasi 94 °C, 1 menit; aneling 55 °C, 1 menit; ekstensi 72 °C, 1.5 menit; ekstensi akhir 72 °C, 10 menit. PCR dilakukan sebanyak 35 siklus. Nested-PCR menggunakan primer fU5/rU5 dan kondisinya hanya berbeda pada suhu aneling primer, yakni 57 °C (Duduk et al. 2013). DNA hasil amplifikasi PCR dielektroforesis menggunakan gel agarosa 1%yang mengandung EtBr pada tegangan 75 Volt DC selama 30 menit. Hasil elektroforesis divisualisasi pada transilluminator UV untuk mengamati dan mendokumentasi pita DNA.
HASIL
Deteksi Awal Penyakit Sapu pada Tanaman Kacang Tanah
Tanaman kacang tanah yang terinfeksi fitoplasma umumnya akan menunjukkan gejala setelah berumur di atas 45 hari berupa daun kecil pada batang utama dan cabang. Pada batang utama terjadi pembentukan tunas samping yang terdiri atas daun kecil yang sangat banyak. Tanaman yang menunjukkan gejala tersebut selanjutnya dideteksi dengan
teknik PCR standar. Hasil amplifikasi DNA dengan teknik PCR standar menunjukkan tanaman positif terinfeksi fitoplasma dengan terbentuknya pita DNA berukuran ±1800 pb (Gambar 1).
Tanaman Kacang Tanah Terinfeksi Fitoplasma pada Berbagai Kondisi Preservasi
Preservasi contoh tanaman terinfeksi pada suhu -20 °C dalam berbagai medium selama 4 minggu secara keseluruhan tidak mengalami perubahan warna. Pada suhu 4 °C tanaman mengalami perubahan warna untuk beberapa contoh seperti perubahan warna dari warna hijau muda menjadi hijau kecokelatan, cokelat muda, dan cokelat tua pada medium NaCl, alkohol, dan tanpa medium. Preservasi tanaman contoh pada suhu ruang (25 °C) dalam berbagai medium menunjukkan sebagian besar contoh mengalami perubahan warna hingga mengalami kerusakan/deteriorasi, kecuali pada medium CTAB. Contoh yang disimpan dalam medium CTAB selama 4 minggu tidak mengalami perubahan bentuk dan warna. Adapun preservasi DNA tanaman contoh dalam kertas FTA pada semua suhu mengalami kerusakan sejak minggu ke-1 (Tabel 1).
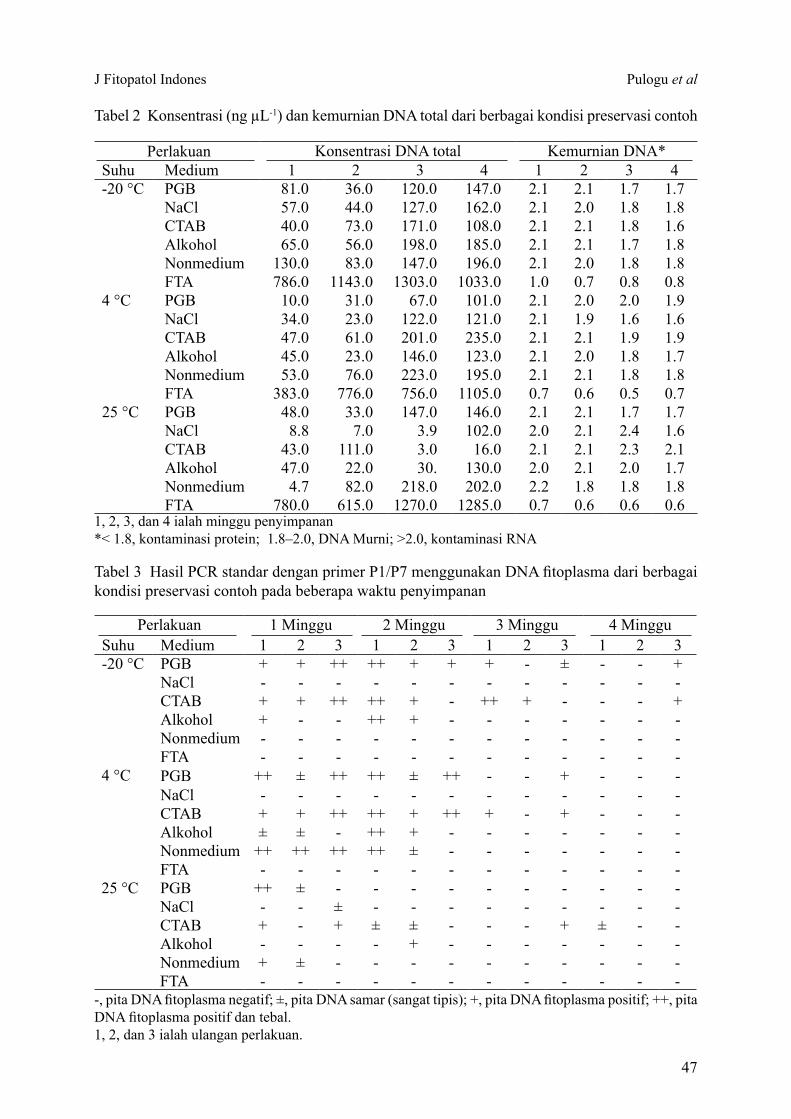
Konsentrasi dan kemurnian DNA total dari hasil preservasi contoh tanaman pada berbagai kondisi menunjukkan nilai yang beragam. Konsentrasi DNA berkisar 3–1303 ng µL-1
Gambar 2 Visualisasi fragmen DNA fitoplasma menggunakan primer P1/P7 hasil deteksi awal penyakit sapu tanaman kacang tanah. M, Penanda 1 Kb; KP, kontrol positif; KT–1 s/d KT–10, tanaman kacang tanah terinfeksi fitoplasma.
1800 pb
M KP KT-1 KT-2 KT-3 KT-4 KT-5 KT-6 KT-7 KT-8 KT-9 KT-10

J Fitopatol Indones Pulogu et al
46
dari berbagai kondisi preservasi contoh. Nilai kemurnian DNA yang diperoleh berkisar antara 0.5 dan 2.4 (Tabel 2).
Pendeteksian Fitoplasma dari Preservasi Jaringan
Amplifikasi DNA dengan teknik PCR standar menggunakan sepasang primer P1/P7menunjukkan amplikon yang positif fitoplasma bervariasi pada masing-masing contoh tanaman (Tabel 3). Contoh tanaman yang dipreservasi dalam bufer PGB dan CTAB konsisten menghasilkan amplikon DNA positif fitoplasma pada suhu 4 °C selama 3 minggu dan suhu -20 °C hingga minggu ke-4.Pada suhu 25 °C, DNA positif fitoplasma teramplifikasi dari contoh yang dipreservasi hanya terdapat pada medium CTAB selama 4 minggu.
Amplifikasi DNA dengan teknik nested-PCR menggunakan sepasang primer fU5/rU3 mengamplifikasi fragmen DNA fitoplasma berukuran ± 890 pb. Semua DNA fitoplasma yang awalnya terdeteksi negatif dari contoh tanaman yang dipreservasi pada berbagai
kondisi berbeda-beda selama minggu 1 dan 4 menunjukkan semuanya berhasil teramplifikasi dengan teknik nested-PCR tersebut (Gambar 2).
PEMBAHASAN
Infeksi fitoplasma pada tanaman dapat menyebabkan gangguan keseimbangan hormon seperti peningkatan sepuluh kali lipat indole-3acetic acid (IAA). Selanjutnya fitoplasma mepengaruhi fungsi jaringan floem dalam mengangkut hasil fotosintesis. Hasil fotosintesis menumpuk pada kloroplas dan terjadi peningkatan konsentrasi gula pada daun sehingga menyebabkan gangguan fisiologi pada tanaman yang ditandai dengan gejala–gejala khas. Gejala khas pada tanaman yang terinfeksi fitoplasma meliputi proliferasi tunas kecil pada ketiak cabang, phyllody (pembentukan daun dari struktur bunga), kerdil, dan pemanjangan ruas batang yang abnormal (Bertaccini et al. 2014).
Preservasi jaringan tanaman kacang tanah bergejala penyakit sapu pada berbagai kondisi
Tabel 1 Tingkat deteriorasi contoh terinfeksi fitoplasma pada berbagai perlakuan preservasi
N, normal; R, rusak; H, hijau; HC, hijau kecokelatan; CM, Cokelat muda; C, Cokelat ; CT, Cokelat tua.
Perlakuan Perubahan contoh pada berbagai kondisi1 minggu 2 minggu 3 minggu 4 minggu
Suhu Medium Bentuk Warna Bentuk Warna Bentuk Warna Bentuk Warna-20 °C PGB N H N H N H N H
NaCl N H N H N H N HCTAB N H N H N H N HAlkohol N HC N HC N HC N HCN.medium N H N H N H N HFTA R HC R HC R HC R HC
4 °C PGB N H N H N H N HNaCl N H N HC N HC N HCCTAB N H N H N H N HAlkohol N CM N CM N CM N CMN.medium N H N HC N HC N CTFTA R HC R HC R HC R HC
25 °C PGB R CM R CM R CM R CMNaCl N HC N C N C N CCTAB N H N H N H N HAlkohol N CM N CM N CM N CMN.medium N CM N CT N CT N CTFTA R HC R HC R HC R HC
J Fitopatol Indones Pulogu et al
47
Tabel 2 Konsentrasi (ng µL-1) dan kemurnian DNA total dari berbagai kondisi preservasi contoh
1, 2, 3, dan 4 ialah minggu penyimpanan*< 1.8, kontaminasi protein; 1.8–2.0, DNA Murni; >2.0, kontaminasi RNA
Perlakuan Konsentrasi DNA total Kemurnian DNA*Suhu Medium 1 2 3 4 1 2 3 4-20 °C PGB 81.0 36.0 120.0 147.0 2.1 2.1 1.7 1.7
NaCl 57.0 44.0 127.0 162.0 2.1 2.0 1.8 1.8CTAB 40.0 73.0 171.0 108.0 2.1 2.1 1.8 1.6Alkohol 65.0 56.0 198.0 185.0 2.1 2.1 1.7 1.8Nonmedium 130.0 83.0 147.0 196.0 2.1 2.0 1.8 1.8FTA 786.0 1143.0 1303.0 1033.0 1.0 0.7 0.8 0.8
4 °C PGB 10.0 31.0 67.0 101.0 2.1 2.0 2.0 1.9NaCl 34.0 23.0 122.0 121.0 2.1 1.9 1.6 1.6CTAB 47.0 61.0 201.0 235.0 2.1 2.1 1.9 1.9Alkohol 45.0 23.0 146.0 123.0 2.1 2.0 1.8 1.7Nonmedium 53.0 76.0 223.0 195.0 2.1 2.1 1.8 1.8FTA 383.0 776.0 756.0 1105.0 0.7 0.6 0.5 0.7
25 °C PGB 48.0 33.0 147.0 146.0 2.1 2.1 1.7 1.7NaCl 8.8 7.0 3.9 102.0 2.0 2.1 2.4 1.6CTAB 43.0 111.0 3.0 16.0 2.1 2.1 2.3 2.1Alkohol 47.0 22.0 30. 130.0 2.0 2.1 2.0 1.7Nonmedium 4.7 82.0 218.0 202.0 2.2 1.8 1.8 1.8FTA 780.0 615.0 1270.0 1285.0 0.7 0.6 0.6 0.6
Tabel 3 Hasil PCR standar dengan primer P1/P7 menggunakan DNA fitoplasma dari berbagai kondisi preservasi contoh pada beberapa waktu penyimpanan
-, pita DNA fitoplasma negatif; ±, pita DNA samar (sangat tipis); +, pita DNA fitoplasma positif; ++, pita DNA fitoplasma positif dan tebal.1, 2, dan 3 ialah ulangan perlakuan.
Perlakuan 1 Minggu 2 Minggu 3 Minggu 4 MingguSuhu Medium 1 2 3 1 2 3 1 2 3 1 2 3-20 °C PGB + + ++ ++ + + + - ± - - +
NaCl - - - - - - - - - - - -CTAB + + ++ ++ + - ++ + - - - +Alkohol + - - ++ + - - - - - - -Nonmedium - - - - - - - - - - - -FTA - - - - - - - - - - - -
4 °C PGB ++ ± ++ ++ ± ++ - - + - - -NaCl - - - - - - - - - - - -CTAB + + ++ ++ + ++ + - + - - -Alkohol ± ± - ++ + - - - - - - -Nonmedium ++ ++ ++ ++ ± - - - - - - -FTA - - - - - - - - - - - -
25 °C PGB ++ ± - - - - - - - - - -NaCl - - ± - - - - - - - - -CTAB + - + ± ± - - - + ± - -Alkohol - - - - + - - - - - - -Nonmedium + ± - - - - - - - - - -FTA - - - - - - - - - - - -

J Fitopatol Indones Pulogu et al
48
waktu dipengaruhi oleh suhu dan medium. Namun tidak semua kombinasi suhu dan medium mampu mempertahankan keawetan jaringan tanaman. Hanya beberapa medium tertentu seperti medium PGB pada suhu rendah dan medium CTAB pada suhu yang tinggi dapat menjaga keawetan jaringan tanaman. Suhu diduga merupakan faktor utama yang dapat memengaruhi perubahan jaringan tanaman. Hal ini dibuktikan dengan semakin tinggi suhu maka semakin cepat proses kerusakan jaringan tanaman yang disimpan. Begitupun sebaliknya, semakin rendah suhu maka semakin lama proses terjadinya kerusakan sehingga contoh masih terlihat segar selama waktu tertentu. Suhu yang sangat rendah dapat secara efektif menghentikan pertumbuhan dan perkembangan biologi dalam sel sehingga keutuhan sel-sel terjaga dalam jangka panjang (Zeliang dan Pattanayak 2012). Suhu yang rendah menimbulkan sel-sel dalam jaringan membeku sehingga proses metabolisme semua sel hidup terhenti. Jaringan daun terinfeksi fitoplasma yang disimpan selama 32 hari pada suhu -20 °C masih dalam keadaan segar. Adapun penyimpanan contoh pada suhu 4 °C dalam waktu lama dapat mengakibatkan perubahan jaringan tanaman dan meningkatkan infeksi berbagai patogen sehingga sulit digunakan untuk deteksi DNA fitoplasma (Wongwarat et al. 2011).
Medium penyimpanan bufer PGB pada suhu rendah (-20 °C dan 4 °C) dan bufer CTAB pada suhu standar (25 °C) cukup efektif mempertahankan keawetan contoh selama proses penyimpanan. Bufer PGB yang mengandung senyawa polyvinylpyrrolidone (PVP), asam askorbat (AA), dan bovine serum albumin (BSA) serta sukrosa diduga dapat mencegah munculnya senyawa-senyawa kontaminan perusak sel dalam jaringan tanaman dan menstabilkan DNA. Adapun bufer CTAB mengandung senyawa-senyawa yang dapat mencegah munculnya senyawa kontaminan perusak DNA. Menurut Hodkinson et al (2007) penyimpanan contoh menggunakan bufer CTAB dapat mencegah terjadinya antioksidan dengan merusak enzim yang menghasilkan senyawa kontaminan dalam jaringan sehingga kehomogenan jaringan tanaman terjaga dan tidak mengalami kerusakan.
Perbedaan konsentrasi DNA dipengaruhi adanya sisa larutan-larutan hasil penyimpanan contoh yang ikut terbawa DNA dalam proses ekstraksi. Kemurnian yang sesuai nisbah menandakan bahwa hasil DNA cukup murni dengan kontaminan protein maupun RNA yang minimum. Nilai kemurnian DNA >2.0 diduga terkontaminasi RNA yang tidak terpecah dalam proses ekstraksi sehingga mengganggu tingkat kemurnian DNA. DNA yang murni
Gambar 2 Visualisasi fragmen DNA fitoplasma hasil nested-PCR menggunakan primer P1/P7 dilanjutkan dengan primer fU5/rU3 dari penyimpanan tanaman kacang tanah bergejala sapu. M, Penanda DNA 1 Kb; P, PGB; N, NaCl; C, CTAB; A, Alkohol; K, Non Media; F, FTA.
890 pb
M P N C A K F P N C A K F P N C A K F
890 pb
-20 °C 4 °C 25 °C
1 minggu
4 minggu

J Fitopatol Indones Pulogu et al
49
dengan jumlah yang relatif cukup banyak dapat digunakan untuk proses amplifikasi DNA hingga beberapa kali.
Contoh yang dipreservasi dalam bufer PGB pada suhu rendah (-20 °C dan 4 °C) dan bufer CTAB pada suhu standar (25 °C) lebih baik untuk menjaga keutuhan DNA fitoplasma dalam jaringan tanaman. PGB mengandung senyawa PVP yang dapat mengurangi kandungan fenol pada jaringan tanaman. Bufer CTAB yang mengandung senyawa PVP, mercaptoethanol, dan NaCl yang dapat menghambat senyawa kontaminan seperti fenolik, enzim DNAse, dan polisakarida. Oleh karena itu, ketika proses preservasi dan ekstraksi, DNA fitoplasma terlindungi oleh senyawa-senyawa dalam bufer PGB dan CTAB hingga proses amplifikasi menghasilkan pita DNA positif dan tebal pada gel agarosa.
Adanya pita DNA fitoplasma yang tidak terlihat bukan berarti selalu negatif, tetapi dapat diduga karena konsentrasi DNA rendah ataupun konsentrasi yang terlalu tinggi dan adanya inhibitor yang terbawa bersama DNA templat sehingga mengganggu reaksi amplifikasi dengan PCR standar. Inhibitor berupa polisakarida, polifenol maupun konsentrasi garam yang tinggi pada proses amplifikasi PCR mampu menghambat laju aktivitas Taq polymerase dalam mensintesis DNA (Mullis dan Faloona 1987).
Penelitian ini menunjukkan bahwa preservasi contoh hingga minggu ke-4 jaringan tanaman kacang tanah bergejala penyakit sapu pada berbagai suhu dan medium tetap efektif untuk mendeteksi fitoplasma melalui pengujian nested-PCR. Pengujian nested-PCR dirancang untuk meningkatkan sensitivitas dan kespesifikan, khususnya amplifikasi fitoplasma dari contoh dengan konsentrasi DNA yang rendah sehingga dapat mengganggu keberhasilan PCR. Selain itu, amplikon DNA hasil PCR pertama yang telah terencerkan dan selanjutnya menjadi templat pada PCR kedua (nested-PCR) mampu mengurangi konsentrasi inhibitor pada reaksi PCR kedua sehingga hasil amplifikasi DNA fitoplasma lebih optimal (Prasetyo 2012).
DAFTAR PUSTAKA
Bertaccini A, Duduk B, Paltrinieri S, Contaldo N. 2014. Phytoplasmas and phytoplasma diseases: a severe threat to agriculture. Amer J Sci Res. 5:1763–1788. DOI: https://doi.org/10.4236/ajps.2014.512191.
Dellaporta SL, Wood J, Hicks JB.1983. A plant DNA minipreparation: version II. Plant Mol Biol Rep. 1:19–21. DOI: https://doi.org/10.1007/BF02712670.
Duduk B, Paltrinieri S, Lee IM, Bertaccini A. 2013. Nested PCR and RFLP Analysis Based on the 16S rRNA Gene. Di dalam Dickinson M, Hodgetts J, editor. Phytoplasma methods and protocols. New York (US): Humana Press. hlm 159–171.
Gundersen DE, Lee IM. 1996. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol Mediterr. 35:144–151.
Hodkinson TR, Waldren S, Parnell JAN, Kelleher CT, Salamin K, Salamin N. 2007. DNA banking for plant breeding, biotechnology and biodiversity evaluation. J Plant Res. 120:17–29. DOI: https://doi.org/10.1007/s10265-006-0059-7.
Lorenz KH, Schneider B, Ahrens U, Seemuller E. 1995. Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and non-ribosomal DNA. Phytopathology. 85:771–776. DOI: https://doi.org/10.1094/Phyto-85-771.
Mullis KB, Fallona FA. 1987. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Didalam: Methods in Enzymology 155. Academic Press, Inc. hlm 335–350.
Mutaqin KH, Suseno R, Tjahjono B, Hidayat P. 2003. Deteksi molekuler dan uji penularan fitoplasma asal rumput bermuda. Hayati. 10(2):66–70.
Nejat N, Vadamalai G. 2013. Diagnostic techniques for detection of phytoplasma diseases: past and present. J Plant Dis Prot. 120(1):16–25. DOI: https://doi.org/10.1007/BF03356449.

J Fitopatol Indones Pulogu et al
50
Nugroho S, Suseno R, Hidayat SH, Hidayat P. 2000. Evaluasi ketahanan beberapa varietas kacang tanah terhadap fitoplasma. Bul HPT. 12(2):48–52.
Prasetyo AE. 2012. Deteksi dan identifikasi fitoplasma yang berasosiasi dengan penyakit layu kelapa di Pulau Derawan Kalimantan Timur [tesis]. Bogor (ID): Institut Pertanian Bogor.
Wongwarat T, Sakuanrungsirikul S, Theerakulpisut P. 2011. Effective methods of preserving SCWL-diseased sugarcane leaves for genomic DNA extraction and molecular detection of phytoplasma.
Afr J Biotech. 10(53):10871–10876. DOI: https://doi.org/10.5897/AJB11.1570.
Zeliang PK, Pattanayak A. 2012. Fundamental crybiology and basic physical, thermodynamical and chemical aspects of plant tissue cryopreservation. Di dalam Abdurakhmonov I, editor. Biochemistry, Genetics and Molecular Biology: Plant Breeding. https://www.intechopen.com/books/plant-breeding/fundamental-c r y o b i o l o g y - a n d - b a s i c - p h y s i c a l -thermodynamical-and-chemical-aspects-of-plant-tissue-cryoInTech.

51
ISSN: 0215-7950
Volume 13, Nomor 2, Maret 2017Halaman 51–58
DOI: 10.14692/jfi.13.2.51
*Alamat penulis korespondensi: Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian, Universitas Lambung Mangkurat. Jalan Jendral Ahmad Yani. KM 36, Banjarbaru 70714. Kotak Pos 1028.Tel: 0511-4777392, Faks : 0511-4777392, Surel: [email protected]
Ketahanan Kultivar Pisang Lokal Kalimantan Selatan terhadap Penyakit Bercak Sigatoka (Mycosphaerella sp.)
Resistance of South Kalimantan Local Banana Cultivars Against Sigatoka Disease
Mariana*, Rodinah, Ismed Setya BudiUniversitas Lambung Mangkurat, Banjarbaru 70714
ABSTRAK
Sigatoka yang disebabkan oleh Mycosphaerella sp. merupakan penyakit penting yang menyebabkan rendahnya kualitas dan kuantitas produksi pisang. Penanaman kultivar pisang yang tahan dianggap sebagai pendekatan yang menjanjikan untuk mengurangi kerugian akibat penyakit ini. Penelitian bertujuan menentukan ketahanan kultivar pisang lokal asal Kalimantan Selatan terhadap penyakit sigatoka. Cendawan diisolasi dari daun pisang kultivar Kapas dengan gejala sigatoka yang parah menggunakan medium agar-agar dekstrosa kentang. Isolat dikonfirmasi menggunakan postulat Koch. Isolat diidentifikasi sebagai Mycosphaerella sp. dan digunakan untuk menguji 11 kultivar pisang lokal yang tumbuh di Kalimantan Selatan. Tingkat ketahanan ditentukan berdasarkan perkembangan gejala. Kerapatan stomata masing-masing kultivar diukur. Secara umum gejala muncul 2 sampai 8 hari setelah inokulasi. Hasil penelitian menunjukkan terdapat perbedaan tingkat ketahanan, yaitu rentan (‘Awak’, ‘Jaranang’, ‘Kapas’, ‘Mas’, dan ‘Paikat’), agak tahan (‘Ambon’, ‘Kepok’, ‘Mas Bantan’, ‘Mauli’, dan ‘Talas’), dan tahan (‘Tarati’). Masa inkubasi penyakit yang semakin panjang berhubungan dengan tingkat ketahanan yang tinggi. Intensitas penyakit berkorelasi negatif terhadap masa inkubasi, tetapi tidak berhubungan secara nyata dengan kerapatan stomata.
Kata kunci: intensitas penyakit, masa inkubasi, postulat Koch
ABSTRACT
Sigatoka of banana caused by Mycospherella sp. is an important disease, since it causes yield losses. Resistant variety is considered a promising approach for sigatoka disease management. This study was aimed to evaluate the resistance of local banana cultivars from South Kalimantan against sigatoka disease. The fungus was isolated from leaves of Kapas cultivar showing severe symptoms of sigatoka disease, onto potato dextrose agar medium. The isolate was then confirmed as Mycosphaerella sp. following Koch’s postulate. Level of resistance is determined based on symptoms development. Resistance of local banana cultivars can be differentiated into susceptible (‘Awak’, ‘Jaranang’, ‘Kapas’, ‘Mas’, and ‘Paikat’), medium resistant (‘Ambon’, ‘Kepok’, ‘Mas Bantan’, ‘Mauli’, and ‘Talas’), and resistant (‘Tarati’). In general, the symptom was visible 2 to 8 days after inoculation. The longer incubation period the more resistant response of banana varieties. Incubation period was negatively correlated to the disease intensity, but it is not correlated with stomatal density.
Keyword: disease intensity, incubation period, Koch postulate

J Fitopatol Indones Mariana et al
52
PENDAHULUAN
Cendawan Mycosphaerella musicola (anamorf: Pseudocercospora musicola) merupakan penyebab gejala Sigatoka Kuning sedangkan M. Fijiensis (Anamorf: Pseudocercospora fijiensis) penyebab gejala Sigatoka Hitam (Gomes et al. 2013). Penyakit ini dapat menyebabkan penurunan kecepatan fotosintesis dan transpirasi. Infeksi M. fijiensis juga menurunkan kandungan klorofil, sukrosa, pati, dan gula pada daun pisang yang mengakibatkan gangguan pertumbuhan dan produktivitas tanaman (Rodriguez-Gaviria dan Cayon 2008). Penyakit sigatoka menyebabkan daun meranggas, buah yang berukuran kecil-kecil, menyebabkan bakal buah rontok, menurunkan kualitas buah, pematangan buah lebih awal, hingga produksi pisang menurun sampai 50% (Ploetz 2007). Penyakit ini mematikan tanaman dengan lambat, tetapi jika tidak dikendalikan akan menimbulkan kerugian yang besar. Pada tahun 1995, di Costa Rica biaya untuk pengendalian penyakit sigatoka mencapai US$900–1500 ha-1
setiap tahunnya (Bennett dan Arneson 2003; Ploetz 2007). Taiwan Banana Research Institute mengendalikan penyakit sigatoka hitam dengan introduksi kultivar pisang yang tahan (ICDF 2013).
Terdapat beberapa jenis pisang yang banyak tumbuh subur dan banyak diusahakan di Kalimantan Selatan. Hal tersebut memberi peluang untuk mendapat pisang yang membawa gen tahan untuk merakit kultivar pisang yang lebih tahan penyakit sigatoka. Penelitian ini dilakukan untuk menguji tingkat ketahanan 11 kultivar pisang lokal Kalimantan Selatan terhadap penyakit sigatoka.
BAHAN DAN METODE
Sumber InokulumSumber inokulum merupakan isolat
patogen penyebab sigatoka yang telah di uji dengan postulat Koch. Dimulai dengan isolasi dari tanaman pisang kultivar Kapas yang bergejala bercak sigatoka hitam di Banjarbaru. Isolasi dilakukan dengan menumbuhkan
cendawan pada medium agar-agar dekstrosa kentang (ADK) dan diinkubasi pada ± 26 °Cselama 3–5 hari. Inokulasi potongan hifa miselium dilakukan pada daun pisang kultivar Kapas (kultivar rentan) dengan cara mengoleskan suspensi potongan (fragmen) miselium sebanyak 0.05 mL (12 mg mL-1 hifa dalam tween 20 0.05%) pada permukaan bawah daun seluas 6 × 6 cm2 menggunakan kuas halus (Donzelli dan Churchill 2007). Bagian ini ditutup dengan plastik transparan dan dijaga kelembapannya. Isolat cendawan yang menghasilkan gejala positif sigatoka pada daun pisang yang diinokulasi (Gambar 1b), dimurnikan, diidentifikasi dan disimpan sebagai biakan murni. Daun yang menunjukkan gejala penyakit sigatoka tersebut, diisolasi kembali dan dimurnikan serta diidentifikasi berdasarkan karakteristik morfologinya (Crous dan Mourichon 2002; Crous et al. 2007; Arzanlou et al. 2008; PaDIL 2011). Isolat dengan karakter morfologi makro dan mikro yang sama dengan isolat hasil isolasi awal, digunakan sebagai sumber inokulum. Isolat tersebut adalah isolate BS 1.
Uji Ketahanan PisangPisang uji yang digunakan berasal
dari Banjarbaru, Kalimantan Selatan dan sekitarnya. Pisang terdiri atas 11 kultivar, yaitu pisang kultivar Awak, Ambon, Jaranang Habang, Kapas, Kepok, Mauli, Mas, Mas Bantan, Paikat, Talas, dan Tarati. Umur tanaman berkisar 3–4 bulan yang ditanam pada kantong plastik berukuran 19.5 cm ×19.5 cm × 40 cm dengan medium campuran tanah dan sekam (1:1 v/v). Setiap kultivar diulang 2 kali. Inokulasi patogen isolat BS 1 dilakukan seperti pada uji postulat Koch. Inokulasi dilakukan pada 5 bidang permukaan daun yang paling muda (sudah terbuka sempurna).
PengamatanPengamatan dilakukan sejak gejala mulai
terlihat. Peubah yang diamati ialah masa inkubasi patogen (hari inokulasi sampai timbulnya gejala pertama), intensitas penyakit, tingkat ketahanan varietas, dan kerapatan stomata tiap kultivar pisang. Intensitas

J Fitopatol Indones Mariana et al
53
penyakit diukur menggunakan rumus indeks infeksi modifikasi Gauhl (Carlier et al. 2002):Indeks infeksi =S n × b
(N-1) × T × 100%, dengan
n, jumlah daun dari tiap kategori serangan. b, nilai skala tiap kategori serangan. N,jumlah skor dalam skala ini (7). T, jumlah daun yang diskor.
Kategori skor yang digunakan untuk pengukuran intensitas penyakit sebagai berikut: Skor 0, tidak terserang; skor 1, < 1% daun terserang; skor 2, 1–5% daun terserang; skor 3, 5–15% daun terserang; skor 4, 15–33% daun terserang; skor 5, 33–50% daun terserang; dan skor 6, >50% daun terserang.
Tingkat ketahanan diklasifikasi ber-dasarkan pada tingkat perkembangan gejala sebagai berikut: tahan, tingkat perkembangan, gejala 0–1; agak tahan, tingkat perkembangan gejala 2–3; dan rentan, tingkat perkembangan gejala 4–5 (Fullerton dan Olsen 1995).
Setiap tingkat perkembangan gejala dideskripsikan mengikuti Fullerton dan Olsen (1995), yaitu: tingkat 0, tidak terdapat gejala; tingkat 1, bercak berwarna kemerahan di bawah permukaan daun dan tidak terdapat gejala di atas permukaan daun; tingkat 2, bercak berwarna kemerahan, beraturan atau tidak beraturan pada bawah permukaan daun; tingkat 3, bagian bercak terlihat pada permukaan daun bagian atas; tingkat 4, bercak hitam atau cokelat, disertai halo kuning atau klorosis, kadang terdapat bagian bercak berwarna hijau; dan tingkat 5, bercak hitam dengan bagian tengah berwarna abu-abu, daun mengalami nekrosis dan terkadang gugur.
HASIL
Hasil uji postulat Koch dan pengamatan morfologi patogen, menunjukkan cendawan penyebab bercak daun diidentifikasi dari genus Mycosphaerella, dengan anamorf Pseudocercospora. Pengamatan pada daun pisang kultivar Kapas dengan gejala sigatoka yang berasal dari lapang (Banjarbaru) (Gambar 1a), dan pada daun pisang hasil inokulasi (Gambar 1b), ditemukan struktur reproduksi patogen, saat 20 hari setelah inokulasi. Struktur reproduksi aseksual berupa sporodokium (konidioma) dan konidium (Gambar 1c). Konidium bervariasi dari soliter hingga bergerombol dalam sporodokium. Konidiumnnya tidak terlepas secara aktif, berwarna cokelat muda, lurus atau melengkung dengan ujung runcing, dan bersekat (3–7). Konidium juga ditemukan pada biakan di medium ADK (Gambar 1e). Struktur reproduksi seksual yang terdeteksi adalah askokarp (berupa peritesium) dan askospora (Gambar 1d). Berdasarkan karakter karakter tersebut, cendawan patogen diidentifikasi sebagai Mycosphaerella sp. (Crous dan Mourichon 2002; Arzanlou et al. 2008).
Pada pengamatan pertama intensitas penyakit tertinggi ialah pada pisang kultivar Mauli, yakni 20%. Pada akhir pengamatan intensitas penyakit tertinggi ialah pada kultivar Kapas, Mauli, Mas, dan Jaranang berturut-turut 71.67%, 56.67 %, 51.67 % dan 51.67% (Tabel 1). Pada kultivar Ambon, Awak, Mas Bantan, Talas, dan Tarati peningkatan
Gambar 1 Gejala penyakit sigatoka dan struktur morfologi Mycosphaerella sp. a, Gejala sigatoka di lapangan; b, Gejala hasil inokulasi; c, Struktur reproduksi aseksual Pseudocercospora; d, Struktur reproduksi seksual Mycosphaerella; dan e, Koloni cendawan. 1, Sporodokium; 2, Konidium; 3, Askokarp; 4, Askospora.
a b c d e
1
2
4
3

J Fitopatol Indones Mariana et al
54
Kultivar pisang Masa inkubasi (hari)
Intentas penyakit (%)
Kerapatan stomata (stomata mm-2)
Tingkat ketahanan
Ambon 5 20.00 139 Agak tahanAwak 7 31.67 158 RentanJaranang Habang 2 51.67 111 RentanKapas 3 71.67 111 RentanKepok 5 28.33 178 Agak tahanMauli 2 56.67 127 Agak tahanMas 4 51.67 175 RentanMas bantan 4 28.33 94 Agak tahanPaikat 2 31.67 151 RentanTalas 5 15.00 139 Agak tahanTarati 8 28.33 138 Tahan
Tabel 1 Masa Inkubasi, Intensitas Penyakit, Kerapatan Stomata dan Tingkat Ketahanan beberapa kultivar pisang di Kalimantan Selatan
Gambar 2 Perkembangan intensitas penyakit sigatoka pada 11 kultivar pisang lokal Kalimantan Selatan yang diinokulasikan dengan suspensi hifa. Pengamatan ke-1 ialah hari mulai munculnya gejala pada masing masing kultivar, pengamatan selanjutnya interval 3 hari. Jenis kultivar pisang: , Ambon;
, Awak; , Jaranang; , Kapas; , Kepok; , Mas; , Mas Batan; , Mauli; , Paikat; , Talas; , Tarati.
80
70
60
50
40
30
20
10
01 42 5 763
Pengamatan ke-
Inte
nsita
s pen
yaki
t (%
)
intensitas tertinggi terjadi pada pengamatan ke-4 dan di pengamatan ke-2 untuk kultivar lainnya. Namun, perkembangannya melambat pada pengamatan selanjutnya (Gambar 2).
Masa inkubasi terpanjang dijumpai pada pisang kultivar Tarati, yaitu 8 hari setelah inokulasi yang dikategorikan sebagai kultivar tahan. Pisang kultivar lain masa inkubasinya lebih singkat sehingga tingkat ketahanannya dikategorikan lebih rendah (Tabel 1). Korelasi antara masa inkubasi dengan intensitas penyakit di akhir pengamatan menunjukkan nilai negatif, artinya semakin panjang masa
inkubasi maka intensitas serangan akan semakin rendah (Gambar 3). Hubungan kedua faktor tersebut kuat, yakni 52.61% (R= -0.52611). Namun, faktor masa inkubasi hanya mampu menjelaskan faktor intensitas serangan sebesar 27.68% (R2 = 0.2768).
Kerapatan stomata dan intensitas penyakit berkorelasi negatif artinya intensitas serangan semakin rendah apabila kerapatan stomata meningkat. Jika dilihat dari nilai R yang kurang dari 0.5 (R = -0.23664) maka hubungan kedua faktor tersebut dikategorikan lemah. Kerapatan stomata hanya mampu menjelaskan

J Fitopatol Indones Mariana et al
55
faktor intensitas serangan sebesar 5.60 % (R2= 0.056) (Gambar 4).
PEMBAHASAN
Lama masa inkubasi pada penelitian ini lebih singkat sekitar 2–8 hari dibandingkan dengan hasil penelitian Taylor (2005) yang melaporkan sekitar 8–10 hari setelah inokulasi. Donzelli dan Churchill (2007) melaporkan masa inkubasi M. fijiensis di lapangan berkisar 10–14 hari setelah inokulasi dan 35 hari pada iklim kering, serta berkisar 12–21 hari apabila diletakkan di rumah kaca. Lama masa inkubasi dipengaruhi oleh tingkat ketahanan tanaman
inang dan virulensi patogen. Adanya korelasi yang kuat antara masa inkubasi dan intensitas penyakit menunjukkan bahwa masa inkubasi merupakan komponen dari ketahanan tanaman terhadap penyakit sigatoka (Leiva-Mora et al. 2015). Masa inkubasi memiliki keterkaitan yang erat dengan tingkat ketahanan. Apabila tingkat ketahanan semakin tinggi maka masa inkubasi akan semakin panjang, begitu juga sebaliknya.
Kerapatan stomata merupakan faktor yang memengaruhi ketahanan tanaman terutama untuk patogen yang penetrasinya melalui stomata. M. musicola masuk melalui stomata yang terbuka (Kannan dan Prakasam 2012). Lemahnya korelasi antara kerapatan stomata
Gambar 4 Hubungan kerapatan stomata dengan intensitas penyakit bercak sigatoka.
80
70
60
50
40
30
20
10
0170 180130 140110 120 150 160 19090 100
Kerapatan stomota (stomata mm-2)
Inte
nsita
s pen
yaki
t (%
)
Gambar 3 Hubungan masa inkubasi dengan intensitas penyakit bercak sigatoka.
80
70
60
50
40
30
20
10
00 21 3 8 96 74 5
Y= -4.9895x + 58.138R = 0.526117858R² = 0.2768
Masa inkubasi (hari)
Inte
nsita
s pen
yaki
t (%
)
Y = -0.1695x + 60.251R = 0. 236643 R² = 0.056

J Fitopatol Indones Mariana et al
56
dan intensitas penyakit pada penelitian ini menunjukkan ada faktor ketahanan lain yang berperan lebih dominan. Faktor fisik lainnya yang berperan terhadap ketahanan ialah ketebalan dinding sel epidermis. Kultivar pisang tahan memiliki kutikula yang lebih tebal daripada yang rentan (Aliah et al 2015).
Pada kultivar pisang tahan seperti Tarati memiliki gen tahan, yaitu gen bsr (Black Sigatoka Resistant) (Orluchukwu dan Ogburia, 2014) yang menyebabkan reaksi hipersensitif segera terjadi setelah infeksi sehingga mampu membatasi aktivitas patogen (Sánchez et al. 2016). Hal ini juga berkaitan dengan toksin yang dihasilkan oleh M. fijiensis yang terlibat dalam inisiasi infeksi, maupun dalam reaksi hipersensitif. Pada kultivar agak tahan seperti Kepok, Mauli, Mas Bantan dan Talas, toksin ini berkontribusi terhadap perluasan lesio (nekrosis) (Harelimana et al. 1997). Toksin (5-hydroxy-1,4-naphthoquinone) dari M. fijiensis berperan penting dalam perkembangan gejala bercak daun karena menghambat fungsi kloroplas dalam transfer elektron (Busogoro et al. 2004); fitotoksin bersifat tidak spesifik inang (Cruz-Cruz et al. 2011). Lepoivre et al. (2003) menjelaskan pada interaksi tanaman pisang dengan M. fijiensis yang tidak kompatibel (reaksi tahan) karena kematian sel penjaga stomata terjadi diawal, dan terjadi penumpukan senyawa dengan elektron yang padat mengelilingi tempat infeksi. Kurangnya peran kerapatan stomata terhadap intensitas penyakit disebabkan karena tabung kecambah cendawan Mycosphaerella sp. bisa menyeberangi stomata tanpa melakukan penetrasi (Kema et al. 1996).
Gen ketahanan terhadap penyakit bercak sigatoka bersifat resesif, sedangkan gen rentan bersifat dominan (Etebu dan Young-Harry 2011). Oleh karena itu, meskipun pada tanaman rentan terdapat gen tahan, tetapi ekspresi gen ini tertutupi oleh gen rentan sehingga tanaman menjadi rentan. Hal tersebut menyebabkan kultivar yang dikategorikan rentan dalam penelitian ini lebih banyak dibandingkan dengan kultivar tahan.
DAFTAR PUSTAKA
Aliah NU, Sulistyowati L, Muhibbudin A. 2015. Hubungan ketebalan lapisan epidermis daun terhadap serangan jamur (Mycosphaerella musicola) penyebab penyakit bercak daun sigatoka pada sepuluh kultivar pisang. JHPT Tropika. 3(1):35–43.
Arzanlou M, Groenewald JZ, Fullerton RA, Abeln ECA, Carlier J, Zapater M-F, Buddenhagen IW, Viljoen A, Crous PW. 2008. Multiple gene genealogies and phenotypic characters differentiate several novel species of Mycosphaerella and related anamorphs on Banana. Persoonia. 20:19–37. DOI: https://doi.org/10.3767/003158508X302212.
Busogoro JP, Etamé JJ, Lognay G, Messiaen J, van Cutsem P, Lepoivre P. 2004. Analysis of the mechanisms of action of Mycosphaerella fijiensis toxins during the development of black leaf streak disease. http://www.fao.org/docrep/007/ae216e/ae216e0g.htm (diakses 28 Juni 2016).
Bennett RS, Arneson PA. 2003. Black Sigatoka of bananas and plantains. http://www.apsnet.org/edcenter/intropp/lessons/fungi/ascomycetes/Pages/BlackSigatoka.aspx (diakses 12 Juli 2015). DOI: https://doi.org/10.1094/PHI-I-2003-0905-01.
Carlier J, De Waele D, Escalant JV. 2002. Global evaluation of Musa germplasm for resistance to Fusarium wilt, Mycosphaerella leaf spot diseases and nematodes: In-dept Evaluation Vol. 6. Montpellier:(FR).Biodiversity International. hlm 45–46
Crous PW, Braun U, Groenewald JZ. 2007. Mycosphaerella is polyphyletic. Studies in Mycology. 58:1–32. DOI: https://doi.org/10.3114/sim.2007.58.01.
Crous PW, Mourichon X. 2002. Mycosphaerella eumusae and its anamorph Pseudocercospora eumusae spp nov: causal agent of eumusae leaf spot disease of banana. Sydowia. 54(1):35–43.
Cruz-Cruz CA, Garcıa-Sosa K, Escalante-Erosa F, Pena-Rodrıguez LM. 2011.

J Fitopatol Indones Mariana et al
57
Physiological effects of the hydrophilic phytotoxins produced by Mycosphaerella fijiensis, the causal agent of black sigatokain banana plants. J Gen Plant Pathol. 77:93–100. DOI: https://doi.org/10.1007/s10327-010-0288-4.
Donzelli BGG, Churchill ACL. 2007. A quantitative assay using mycelial fragments to assess virulence of Mycosphaerella fijiensis. Phytopathology. 97(8):916–929. DOI: https://doi.org/10.1094/PHYTO-97-8-0916.
Etebu E, Young-Harry W. 2011. Control of black sigatoka disease: challenges and prospects. Afr J Agric Res. 6(3):508–514.
Fullerton RA, Olsen TL. 1995. Pathogenic variability in Mycosphaerella fijiensis Morelet, cause of black sigatoka in banana and plantain. J Crop Hort Sci. 23:39–48. DOI: https://doi.org/10.1080/01140671.1995.9513866.
Gomes LIS, Douhan GW, Bibiano LBJ, Maffia LA, Mizubuti ESG. 2013. Mycosphaerella musicola identified as the only pathogen of the sigatoka disease complex present in Minas Gerais State, Brazil. Plant Disease. 97(12):1537–1543. DOI: https://doi.org/10.1094/PDIS-12-12-1212-RE.
Harelimana G, Lepoivre P, Jijakli H, Mourichon X. 1997. Use of Mycosphaerella fijiensis toxins for the selection of banana cultivars resistant to black leaf streak. Euphytica. 96(1):125–128. DOI: https://doi.org/10.1023/A:1002960902950.
ICDF (International Cooperationand Development Fund). 2013. Banana Black Sigatoka Disease Prevention and Treatment Project (St. Lucia) http://www.icdf.org.tw/ct.asp?xItem=18907&CtNode=29823&mp=2(diakses 23 November 2015.
Kannan C, Prakasam V. 2012. Ultra structural studies on the infection process of Mycosphaerella musicola-causal agent of yellow sigatoka on banana. Indian Phytopath. 65(2):192–195.
Kema GHJ, Yu D, Rijkenberg FHJ, Shaw MW and Baayen RP. 1996. Histology of pathogenesis of Mycosphaerella
graminicola in wheat. Phytopathology 86(7):777–786. DOI: https://doi.org/10.1094/Phyto-86-777.
Leiva-Mora M, Capó YA, Suárez MA, Martín MC, Roque B, Méndez EM. 2015. Components of resistance to assess black sigatoka response in artificially inoculated Musa genotypes. Revista de Protección Vegetal. 30(1):60–69.
Lepoivre P, Busogoro JP, Etame JJ, El Hadrami A, Carlier J, Harelimana G, Mourichon X, Panis B, Stella RA, Sallé G, Strosse H, Swennen R. 2003. Banana Mycosphaerella fijiensis Interactions. Di dalam: Proceedings of the 2nd International workshop on Mycosphaerella leaf spot diseases International Network for the Improvement of Banana and Plantain, 20–23 May 2002. MontpellierFranch. International Network for the Improvement of Banana and Plantain (INIBAP). hlm 151–159
Orluchukwu JA, Ogburia MN. 2014. Introgression of bsr gene from wild 2n accessions and derivative hybrids to cultivated 3n landraces of plantains (Musa sp.). Inter J Plant Res. 4(1):1–4.
PaDIL (Pest and Disease Image Library). 2009. Diagnostic Methods for Black Sigatoka Mycosphaerella fijiensis. http://www.padil.gov.au/pbt (diakses 13 januari 2015).
Ploetz RC. 2007. Diseases of tropical perennial crops: challenging problems in diverse environments. Plant Dis. 91(6):644-663. https://doi.org/10.1094/PDIS-91-6-0644.
Rodriguez-Gaviria PA, Cayon G. 2008. Physiological effect of Mycosphaerella fijiensis in banana leaves. Agron Colomb. 26(2):256–265.
Sánchez TE, Hidalgo PL, Pacheco CR, Chávez NT, NavarreteVO, Santos OE. 2016. Identification of differentially-expressed genes in response to Mycosphaerella fijiensis in the resistant musa accession ‘Calcutta-4’ using suppression subtractive hybridization. PLoS ONE. 11(8):1–17. DOI: https://doi.org/10.1371/journal.pone.0160083.

J Fitopatol Indones Mariana et al
58
Taylor MK. 2005. Characterization of potential fungal disease resistance genes in banana [tesis]. Quensland University of Technology.

ISSN: 0215-7950
59
Volume 13, Nomor 2, Maret 2017Halaman 59–66
DOI: 10.14692/jfi.13.2.59
*Alamat penulis korespondensi: Laboratorium Bakteriologi Departemen Proteksi Tanaman, Fakultas Pertanian,Institut Pertanian Bogor, Kampus IPB Dramaga, Bogor 16680Tel: 0251-8376268, Faks : 0251-8629364, Surel: [email protected]
Galur Isogenik Xanthomonas oryzae pv. oryzae Hasil Penyinaran Ultraviolet dan Potensinya Sebagai Penginduksi Resistensi Padi
terhadap Penyakit Hawar Daun Bakteri
Isogenic Xanthomonas oryzae pv. oryzae Strain Resulted from Ultraviolet Irradiation and Their Potency as Plant Resistance Inducer
Against Rice Bacterial Leaf Blight
Fitria Yuliani, Giyanto*, Kikin Hamzah MutaqinInstitut Pertanian Bogor, Bogor 16680
ABSTRAK
Penyakit hawar daun bakteri oleh Xanthomonas oryzae pv. oryzae merupakan salah satu penyakit penting pada tanaman padi dengan tingkat kerusakan mencapai 50%. Salah satu upaya yang dilakukan untuk mengendalikan penyakit ini ialah menggunakan galur bakteri nonpatogen isogenik yang dapat menginduksi ketahanan tanaman. Penelitian ini bertujuan mendapatkan X. oryzae pv. oryzae isogenik menggunakan penyinaran ultraviolet dan mengevaluasi kemampuan isolat dalam menginduksi ketahanan tanaman padi. Biakan X. oryzae pv. oryzae dipapar dengan sinar UV selama 3, 5, 10 dan 15 menit. Sebanyak 99 koloni yang tumbuh diseleksi patogenisitasnya, selanjutnya dikonfirmasi identitasnya menggunakan primer XOR-F dan XOR-R2 sehingga didapat 16 galur isogenik. Produksi eksopolisakarida X. oryzae pv. oryzae isogenik berkisar antara 1.4 dan 9.8 mg mL-1, sedangkan tipe liarnya 6.0 mg mL-1. Lima galur terpilih (M313, M57, M101, M1513 dan M1515) menunjukkan penurunan virulensi 92.8–98.6%. Aplikasi kelima galur tersebut pada bibit padi memperlihatkan masa inkubasi 6.6–5.4 hari setelah inokulasi (hsi), lebih pendek dibandingkan dengan tipe liar, yaitu 4.8 hsi.Perlakuan X. oryzae pv. oryzae isogenik juga mengakibatkan penekanan perkembangan penyakit dengan nilai AUDPC lebih kecil dibandingkan dengan tipe liar dan kontrol. Aplikasi X. oryzae pv. oryzae isogenik tidak memengaruhi pertumbuhan, tetapi meningkatkan produksi tanaman. Galur M101 memiliki masa inkubasi 5.7 hsi, nilai AUDPC paling rendah (615), dan bobot kering gabah (2.8 g per 100 bulir) lebih tinggi dibandingkan dengan tipe liarnya (2.0 g per 100 bulir).
Kata kunci: AUDPC, induksi ketahanan, masa inkubasi, patogenisitas
ABSTRACTBacterial leaf blight (BLB) caused by Xanthomonas oryzae pv. oryzae is an important rice disease
and may cause 50% plant damage. One approach to manage this disease is using isogenic non-pathogenic strain that can induce plant resistance. This study was conducted to obtain isogenic strains of X. oryzae pv. oryzae through ultraviolet irradiation and evaluate ability of the isolates in inducing plant resistance. Bacteria X. oryzae pv. oryzae were exposed to UV for 3, 5, 10 and 15 min. A total of 99 colonies obtained were selected through pathogenicity tests, followed by confirmation using XOR-F and XOR-R2 primers. As much as 16 isogenic strain isolates were obtained with the ability to produce exopolysaccharides (EPS) ranges between 1.4–9.8 mg mL-1 compared to wild-type 6.0 mg mL-1. Five isogenic strains isolates (M313, M57, M101, M1513 and M1515) caused virulence reduction of 92.8–98.6%. Application of those five isogenic isolates prolonged incubation period of X. oryzae pv. Oryzae,

J Fitopatol Indones Yuliani et al
60
i.e. 6.6–5.4 days compared to the wild-type i.e. 4.8 days after inoculation. Suppression of disease progression was also observed with AUDPC value 615.0-827.5, which was lower than those caused by wild-type and control, i.e. 863.3 and 682.5, respectively. Application of isogenic strain isolates did not affect plant growth but it increased plant productivity. Isogenic strain M101 was the best isolate with an incubation period of 5.7 days after inoculation, having the lowest AUDPC value (615.0), and yielded dry weight of 100 grains higher (2.8 g) than those of the wild-type (2.0 g).
Keywords: AUDPC, incubation period, pathogenicity, resistance inducer
PENDAHULUAN
Penyakit hawar daun bakteri (HDB) oleh Xanthomonas oryzae pv. oryzae merupakan salah satu penyakit utama tanaman padi. Bakteri ini menyerang pada fase pertumbuhan vegetatif dan generatif dan menimbulkan gejala berupa hawar berwarna cokelat keabuan pada daun. Luas serangan penyakit HDB di Indonesia selama periode Oktober 2011–Maret 2012 mencapai 98% (Ditjen TP 2012). X. oryzae pv. oryzae patotipe IV merupakan galur yang ganas dan banyak ditemukan di lapangan (Harfiah et al. 2015).
Salah satu upaya pengendalian penyakit tanaman ialah pemanfaatan bakteri nonpatogen isogenik (Rukayadi et al. 2000). Bakteri inimerupakan bakteri yang kehilangan ke-mampuan dalam menimbulkan penyakit akibat perubahan gen yang mengodekan virulensi yang dapat dihasilkan melalui mutasi.
Bakteri nonpatogen isogenik sebagai agens biokontrol lebih menguntungkan karena mutan akan berperilaku sama dengan tipe liarnya dalam merespons perubahan lingkungan (Wahyudi et al. 2011). Gangguan pada gen pembentukan eksopolisakarida menjadi salah satu target mutasi untuk menghasilkan galur isogenik. Eksopolisakarida berupa lendir terdapat pada permukaan sel dan sering dikaitkan dengan virulensi patogen (Lee et al. 2005). Gangguan pada gen pengatur produksi eksopolisakarida menyebabkan penurunan kemampuan galur bakteri isogenik menginfeksi inang sehingga diharapkan mampu berkompetisi dengan tipe liarnya dan mengaktifkan mekanisme ketahanan tanaman. Penelitian ini bertujuan mendapatkan galur X. oryzae pv. oryzae isogenik yang memiliki virulensi rendah melalui penyinaran ultraviolet
dan mengevaluasi kemampuan galur isogenik untuk mengendalikan penyakit hawar daun bakteri pada tanaman padi.
BAHAN DAN METODE
Induksi Penyinaran Ultraviolet X. oryzae pv. oryzae
X. oryzae pv. oryzae patotipe IV (tipe liar) merupakan koleksi Balai Besar Penelitian Tanaman Padi, Sukamandi. Bakteri ini diperbanyak dalam 50 mL medium cair sukrosa kentang dan digojog ± 22 jam sampai kerapatan sel 108 cfu mL-1 (OD600 = 0.2). Sebanyak 100 μL suspensi bakteri disebar pada medium agar-agar sukrosa kentang (ASK) menggunakan glass bead selanjutnya dipapar sinar UV-C (100-280 nm) pada ESCO laminar model EQU/03-EHC dengan waktu yang berbeda, yaitu 3, 5, 10, dan 15 menit. Selanjutnya biakan bakteri diinkubasi pada suhu 28 °C di tempat gelap selama 48 jam. Koloni yang tumbuh dipindahkan pada medium ASK dan selanjutnya dipapar sinar UV dengan perlakuan yang sama dengan sebelumnya (Thein dan Prathuangwong 2010).
Seleksi Galur X. oryzae pv. oryzae IsogenikUji Patogenisitas. Suspensi galur X.
oryzae pv. oryzae isogenik dan tipe liar (108 cfu mL-1) masing-masing diinokulasikan pada daun padi berumur 30 hari setelah tanam (HST). Gunting dicelupkan ke dalam suspensi bakteri, selanjutnya digunakan untuk menggunting daun. Pengamatan panjang hawar dilakukan pada hari ke-14 setelah inokulasi.
Konfirmasi Galur X. oryzae pv. oryzae Isogenik. Konfirmasi X. oryzae pv. oryzae isogenik dilakukan dengan teknik PCR menggunakan

J Fitopatol Indones Yuliani et al
61
pasangan primer spesifik XOR-F(5'-GCATGACGTCATCGTCCTG-3') dan XOR-R2(5'-CTCGGAGCTATATGCCGTGC-3'). Proses PCR dilakukan dengan tahapan denaturasi awal 95 °C selama 2 menit; diikuti 29 siklus terdiri atas tahapan denaturasi 95 °Cselama 30 detik, aneling pada suhu 63 °C selama 30 detik, dan pemanjangan pada suhu 72 °C selama 1 menit; serta ekstensi akhir 72 °C selama 7 menit (Keshavarz et al. 2011).
Produksi Eksopolisakarida. Sebanyak 16 galur X. oryzae pv. oryzae isogenik diremajakan di dalam 10 mL medium cair sukrosa kentang dan digojog 24 jam. Sebanyak 1 mL suspensi bakteri dipindahkan ke tabung mikro 2 mL selanjutnya disentrifugasi pada suhu 4 °C dengan kecepatan 6000 rpm selama 20 menit. Supernatan dipindahkan ke tabung mikro 2 mL dan ditambahkan etanol dingin (96%) sebanyak 2 kali volume supernatan dan didiamkan satu malam. Selanjutnya tabung mikro disentrifugasi pada suhu 4 °C kecepatan 6000 rpm selama 20 menit. Pelet dikeringkan di dalam oven (55 °C) dan ditimbang bobot keringnya (Nudyanto dan Zubaidah 2015).
Induksi Ketahanan Tanaman Padi Menggunakan X. oryzae pv. oryzae Isogenik
Percobaan ini disusun dalam rancangan acak lengkap. Perlakuan terdiri atas inokulasi 5 galur X. oryzae pv. oryzae isogenik terpilih, kontrol positif (X. oryzae pv. oryzae patotipe IV), dan kontrol negatif (akuades steril) dengan 8 ulangan, masing-masing terdiri atas 1 unit tanaman (Tabel 1). Percobaan dilakukan di lahan percobaan Departemen Proteksi Tanaman IPB Bogor (6o 30’ LS, 106o 30’ BT) dengan ketinggian tempat 150 m dpl.
Aplikasi X. oryzae pv. oryzae isogenik tahap I dilakukan pada 25 benih padi IR-64 dengan merendamnya di dalam suspensi sel bakteri (108 cfu mL-1) selama 48 jam. Selanjutnya benih padi disemai pada tanah steril. Bibit berumur 14 hari dipindahkan ke ember plastik yang berisi tanah sawah steril. Aplikasi tahap II dilakukan pada tanaman padi berumur 35 HST dengan penyiraman suspensi sel bakteri sebanyak 20 mL tanaman-1 di sekitar perakaran tanaman.
Inokulasi X. oryzae pv. oryzae patotipe IV dilakukan pada daun bendera (85 HST) dengan metode pengguntingan daun menggunakan gunting steril yang telah dicelupkan pada biakan cair X. oryzae pv. oryzae patotipe IV (108 cfu mL-1). Tanaman yang telah diinokulasi disungkup dengan plastik bening selama 3 hari untuk menjaga kelembapan mikro di sekitar tanaman. Pengamatan Penyakit Hawar Daun Bakteri
Pengamatan masa inkubasi dimulai dari hari pertama setelah inokulasi sampai gejala penyakit terlihat pada tanaman. Keparahan penyakit (KP) merupakan persentase luasnya jaringan tanaman terserang patogen dari total luasan yang diamati, yang dinyatakan dalam rumus berikut:
KP =(ni × vi)Z × N × 100 %, dengan
n
i=1∑
KP, keparahan penyakit; ni, jumlah jaringan terserang pada setiap skala; vi, skala serangan; Z, skala serangan tertinggi; dan N, total jaringan yang diamati. Skala perhitungan berdasarkan standard evolution system (IRRI 2014). Pengamatan dilakukan setiap 3 hari dimulai 3 hari setelah inokulasi dan dihentikan setelah gejala hawar sampai pangkal daun.
Area Under Disease Progress Curve (AUDPC) adalah jumlah penyakit pada setiap perlakuan dari pengamatan pertama hingga pengamatan terakhir. Penghitungan AUDPC menggunakan rumus:AUDPC =
nƩi=0
yi + yi+1
2 × (t i+1– t i), dengan[ ]Yi , insidensi penyakit pada waktu ke-i; ti, waktu pengamatan ke-i.
Pengukuran tinggi dan jumlah anakan padi dilakukan setiap minggu sampai minggu ke-10
Tabel 1 Galur X. oryzae pv. oryzae isogenik terpilih dan lama penyinaran UV-C (100–280 nm)
Lama penyinaran UV (menit)
Kode galurX. oryzae pv. oryzae
0 Patotipe IV (tipe liar) 3 M313 5 M5710 M10115 M1513 dan M1515

J Fitopatol Indones Yuliani et al
62
setelah tanam. Jumlah anakan produktif dihitung menggunakan rumus:
Anakan produktif = × 100 %, denganAB
A, jumlah anakan yang menghasilkan malai; dan B, jumlah anakan keseluruhan. Bobot kering gabah ditentukan dengan menghitung bobot malai yang telah dikeringkan. Pengamatan juga dilakukan terhadap bobot kering 100 bulir padi.
Analisis DataData penyakit hawar daun dianalisis
menggunakan program statistical analisys system (SAS) versi 9.0 dan diuji lanjut menggunakan DMRT pada taraf 5%.
HASIL
Induksi penyinaran UV menghasilkan 99 galur isogenik. Jumlah koloni yang tumbuh semakin sedikit dengan semakin lamanya waktu pemaparan UV (Tabel 2). Galur M101, M1513, dan M1515 memperlihatkan perubahan bentuk, demikian juga warna menjadi kuning pucat dan lendir lebih sedikit dibandingkan dengan tipe liarnya (Gambar 1).
Dari 99 galur bakteri yang diuji dengan teknik PCR diperoleh 16 galur yang positif sama dengan tipe liarnya. Uji patogenisitas 26 galur hasil penyinaran UV menunjukkan penurunan virulensi 25.4–98.6%. Galur M1513 dan M101 memperlihatkan produksi eksopolisakarida paling rendah, yaitu 1.4 mg mL-1 dan 1.6 mg mL-1 berbeda nyata dengan tipe liar 6.0 mg mL-1. Hasil seleksi didapat 5 galur terpilih (M313, M57, M101,
M1513, dan M1515) yang menunjukkan penurunan virulensi sebesar 92.8–98.6% dan penurunan produksi eksopolisakarida dibandingkan tipe liarnya (Tabel 3).
Induksi Ketahanan Galur X. oryzae pv. oryzae Isogenik
Aplikasi galur isogenik mampu memperpanjang masa inkubasi X. oryzae pv. oryzae dan memperlihatkan intensitas penyakit yang lebih rendah dibandingkan dengan aplikasi tipe liarnya. Hasil pengamatan menunjukkan lama masa inkubasi tidak menggambarkan rendahnya intensitas pe-nyakit (Tabel 4). Masa inkubasi paling lama terdapat pada perlakuan galur M1515, yaitu 6.6 HSI sedangkan tipe liar 4.9 HSI, tetapi menunjukkan intensitas penyakit yang tidak berbeda nyata dengan aplikasi tipe liar. Galur M101 mempunyai masa inkubasi 5.7 HSI dan memperlihatkan intesitas penyakit terendah dari semua perlakuan, yaitu 66.1% berbeda nyata dengan aplikasi tipe liar (85.5%). Galur M101 juga memiliki nilai AUDPC yang paling rendah (615.0) sedangkan tipe liar 863.3. Nilai AUDPC yang rendah menunjukkan perkembangan penyakit lebih lambat.
Aplikasi galur isogenik tidak berpengaruh nyata terhadap pertumbuhan, tetapi ber-pengaruh pada produksi tanaman (Tabel 5). Sebanyak 5 galur isogenik menunjukkan kenaikan bobot kering bulir per malai yang nyata daripada tipe liarnya. Bobot bulir per malai berkisar antara 1.1 g dan 1.4 g, sedangkan tipe liarnya hanya 0.5 g dan kontrol 1.1 g. Galur M101 memperlihatkan bobot kering 100 bulir tertinggi dan berbeda nyata dengan tipe liarnya.
PEMBAHASAN
Radiasi ultraviolet mampu melakukan penetrasi ke dinding sel yang mengakibatkan perubahan komposisi asam nukleatnya. Terbentuknya timina dimer memengaruhi proses replikasi menyebabkan terjadinya mutasi. Mutasi dengan penyinaran ultraviolet berakibat pada kesalahan menyandi protein jika tidak bersifat letal sehingga menimbulkan
Tabel 2 Jumlah koloni bakteri X. oryzae pv. oryzae isogenik pada lama penyinaran UV-C (100–280 nm) yang berbeda
Lama penyinaran UV(menit)
Jumlah koloni yang tumbuh
3 50 5 2310 1815 8Total isolat 99

J Fitopatol Indones Yuliani et al
63
penampakan fenotip yang berbeda dari keadaan normalnya dan dapat diwariskan pada keturunannya (Rajeshwar et al. 2002). Mutasi yang dihasilkan berupa salah baca kode genetik (Hockbeger 2002). Absorbsi radiasi ultraviolet menyebabkan terbentuknya ikatan kovalen dua molekul timin yang saling berdekatan (timina dimer) pada suatu urutan DNA (Rastogi et al. 2010).
Pemaparan UV pada penelitian ini menghasilkan perubahan morfologi pada beberapa galur, penurunan kemampuan galur dalam menginfeksi inang dan penurunan produksi eksopolisakarida. Mutan Streptomyces fradiae NRRL-2702 mengalami perubahan morfologi dari tipe liarnya setelah diberi pemaparan UV (Khaliq et al. 2007). Uji patogenisitas mutan X. oryzae pv. oryzae
Tabel 3 Karakteristik fenotip galur Xanthomonas oryzae pv. oryzae isogenik
Lama Penyinaran UV
(menit)
Kode galurX. oryzae pv.
oryzae
Uji PCR Panjang hawar(cm)
Penurunan virulensi
Produksi EPS
(mg mL-1)0 Patotipe 4
(tipe liar)+ 13.8 a 0.0 6.0 c
3
M36 + 7.9 c 42.8 8.4 bM313 + 1.0 e 92.8 2.6 ghM323 + 4.0 d 71.0 3.4 efM331 + 6.4 c 53.6 2.0 ijM334 + 3.1 d 77.5 2.8 ghM338 + 6.9 c 50.0 3.8 e
5M57 + 0.3 e 97.8 5.8 cM511 + 7.8 c 43.5 3.2 fgM514 + 6.9 c 50.0 9.8 aM517 + 10.3 b 25.4 6.2 c
10M101 + 0.5 e 96.4 1.6 jM102 + 0.5 e 96.4 3.6 efM105 + 0.7 e 94.9 2.4 hi
15M153 + 1.4 e 89.9 4.6 dM1513 + 0.2 e 98.6 1.4 jM1515 + 0.2 e 98.6 2.2 hi
+, berdasarkan pada uji PCR isolat berkerabat dengan X. oryzae pv. oryzae.Angka dalam satu kolom diikuti huruf yang sama tidak berbeda nyata pada Uji DMRT α 5%.
Gambar 1 Morfologi Xanthomonas oryzae pv. oryzae M101 berubah dibandingkan dengan tipe liarnya pada medium agar-agar sukrosa kentang (5 hsi), perubahan terlihat pada bentuk, warna, pinggiran, dan lendir pada permukaan koloni. a, Koloni patotipe IV; b, Galur M101.
a b

J Fitopatol Indones Yuliani et al
64
Hrp-(M5) menunjukkan mutan masih menimbulkan gejala penyakit pada padi, namun lebih rendah daripada tipe liarnya. Wahyudi (2011) menduga karena adanya keterlibatan gen-gen lain yang menentukan sifat virulen X. oryzae pv. oryzae.
Eksopolisakarida (EPS) merupakan polimer dari gula dengan bobot molekul tinggi yang disekresikan oleh mikroorganisme ke lingkungan eksternalnya. EPS dari bakteri berperan dalam menghambat translokasi unsur hara dan air, melindung bakteri dalam keadaan ekstrem, dan menetralisir senyawa yang dikeluarkan oleh tanaman (Lee et al. 2005). Pada penelitian ini penurunan produksi EPS menjadi indikator terjadinya gangguan genetik pada faktor pembentukan EPS. Thein dan Prathuangwong (2010) menyatakan penurunan produksi EPS mutan X. oryzae pv. oryzae hasil penyinaran UV dilihat dari ukuran
koloni mutan yang lebih kecil dibandingkan dengan tipe liarnya pada medium yang mengandung sukrosa.
Galur isogenik diduga mampu menekan pertumbuhan tipe liarnya dengan menginduksi ketahanan tanaman. Berdasarkan hasil penelitian terlihat masa inkubasi bakteri tipe liar lebih panjang pada perlakuan galur isogenik. Thein dan Prathuangwong (2010) melaporkan galur M-690 hasil mutasi UV bakteri X. oryzae pv. oryzae mampu menginduksi ketahanan tanaman terhadap penyakit HBD dengan intensitas penyakit 36.7% dibandingkan dengan tipe liarnya 46.7%.
Aplikasi galur isogenik tidak berpengaruh terhadap pertumbuhan, tetapi berpengaruh terhadap hasil tanaman yang ditunjukkan dengan adanya peningkatan dibandingkan dengan aplikasi tipe liarnya. Peningkatan
Tabel 5 Aplikasi galur Xanthomonas oryzae pv. oryzae isogenik terhadap pertumbuhan dan hasil tanaman padi
Angka dalam satu kolom diikuti huruf yang sama tidak berbeda nyata pada Uji DMRT α 5%.
Galur isogenikX. oryzae pv. orzae
Tinggi tanaman
(cm)
Jumlah anakan
Anakan produktif
(%)
Bobot bulir per malai
(g)
Bobot 100 bulir
(g)M313 93.2 a 26.2 a 69.0 a 1.1 ab 2.4 dM57 92.4 a 21.5 a 78.4 a 1.0 ab 2.5 cdM101 94.4 a 25.2 a 72.3 a 1.1 ab 2.8 aM1513 93.1 a 21.5 a 76.7 a 1.4 a 2.6 bcM1515 92.9 a 25.0 a 69.2 a 1.3 ab 2.6 bcPatotipe IV (tipe liar) 95.1 a 26.4 a 69.5 a 0.5 c 2.0 eKontrol 92.8 a 24.0 a 74.9 a 1.0 b 2.6 ab
Tabel 4 Aplikasi galur Xanthomonas oryzae pv. oryzae isogenik terhadap perkembangan penyakit hawar daun bakteri
Angka dalam satu kolom diikuti huruf yang sama tidak berbeda nyata pada Uji DMRT α 5%. HSI, hari setelah inokulasi; AUDPC, Area Under Disease Progress Curve.
Galur isogenik X. oryzae pv. orzae
Masa inkubasi(HSI)
Keparahan penyakit(%)
Nilai AUDPC
M313 5.7 b 71.7 b 617.5M57 5.4 bc 76.7 ab 827.5M101 5.7 b 66.1 b 615.0M1513 6.5 a 74.4 ab 624.2M1515 6.6 a 70.3 b 632.1Patotipe IV (tipe liar) 4.9 c 85.6 a 863.3Kontrol 5.6 b 72.2 b 682.5

J Fitopatol Indones Yuliani et al
65
produksi tanaman diduga akibat aktifnya mekanisme ketahanan tanaman. Mulsanti et al. (2014) melaporkan bahwa bobot kering 100 bulir padi IR-64 berkisar antara 2.4 g dan 2.57 g.
Dari hasil yang didapat terlihat bahwa rendahnya produksi EPS tidak diiringi dengan penurunan virulensi galur isogenik. EPS merupakan salah satu faktor virulensi X. oryzae pv. oryzae sehingga penurunan produksi EPS tidak menyebabkan hilangnya seluruh kemampuan patogen dalam menimbulkan penyakit. Hal ini disebabkan adanya keterlibatan gen-gen lain yang menentukan sifat virulen X. oryzae pv. oryzae (Wahyudi et al. 2011). Penurunan virulensi dan produksi EPS pada galur isogenik terpilih yang diaplikasikan pada padi mampu menginduksi ketahanan tanaman. Hal ini terlihat dari perlambatan perkembangan X. oryzae pv. oryzae berdasarkan nilai AUDPC dengan intensitas penyakit yang tidak berbeda nyata dengan kontrol. Keberadaan galur isogenik tidak memberikan dampak negatif terhadap pertumbuhan dan hasil tanaman.
Galur isogenik yang didapat dari penelitian ini masih berada pada tahap penelitian dasar sehingga masih diperlukan pendalaman untuk menentukan potensinya sebagai agens hayati seperti kestabilan virulensi, respons beragam varietas padi, dan dinamika populasinya di dalam jaringan tanaman padi.
DAFTAR PUSTAKA
[Ditjen TP] Direktorat Jendral Tanaman Pangan. 2012. Evaluasi prakiraan serangan OPT utama tanaman padi dan jagung [Internet]. [diunduh 2016 Januari 24]. Tersedia pada: http://tanamanpangan.deptan.go.id/index.php/.
Harfiah W, Abadi AL, Qurata’aini L. 2015. Ketahanan lima galur padi (Oryza sativa L.) terhadap dua isolat Xanthomonas oryzae pv. oryzae penyebab penyakit hawar daun bakteri pada tanaman padi. J HPT. 3(2): 2338-4336.
Hockberger PE. 2002. A history of ultraviolet photobiology for humans, animals and
microorganisms. Photobiol. 76: 561–579. DOI: https://doi.org/10.1562/0031-8655(2002)0760561AHOUPF2.0.CO2.
[IRRI] International Rice Research Institute. 2014. Standart evaluation system for rice (5th ed). Philippines (PH): Agricultural Research Los Banos University.
Keshavarz K, Sijam K, Ahmad M, Abidin Z, Hashim H, Nazerian E. 2011. Rapid identification and differention of Xanthomonas oryzae pv. oryzae strain with primer 16S-23S rDNA from rice fields in Peninsular Malaysia. Asian J Plant Pathol. 5(2): 93-99. DOI: https://doi.org/10.3923/ajppaj.2011.93.99.
Khaliq S, Kalsoom A, Muhammad AG, Ruqia I, Ahmad MK,Muhammad M. 2007. Change incolony morphology and kinetic softy losin production after UV and gammairradiation mutagenesis of Streptomycesfradiae NRRL-2702. Microbiol Res. 164: 469-477. DOI: https://doi.org/10.1016/j.micres.2007.02.005.
Lee BM, Park DS, Kang JG, Kim ES, Song IC, Park UH, Yoon JH, Hahn BS, Koo GB, Koh JS. 2005. The genome sequence of Xanthomonas oryzae pathovar oryzae KACC10331, the bacterial blight pathogen of rice. Nucleic Acids Res. 33(2):577-586. DOI: https://doi.org/10.1093/nar/gki206.
Mulsanti IW, Wahyuni S, Sembiring H. 2014. Hasil Padi dari empat kelas benih yang berbeda. Penelitian Pertanian Tanaman Pangan. 33(3): 169-176.
Nudyanto A, Zubaidah E. 2015. Isolasi bakteri asam laktat penghasil eksopolisakarida dari kimchi. J Pangan Agroindustri. 3(2): 743-748.
Rukayadi Y, Suwanto A, Tjahjono, Harling R. 2000. Survival and epiphytic fitness of a nonpathogenic mutant of Xathomonas campestris pv. glycines. Appl Environ Microbiol. 66(3): 1183-1189. DOI: https://doi.org/10.1128/AEM.66.3.1183-1189.2000.
Rajeshwar PS, Donat PH. 2002. UV-induced DNA damage and repair. Photochem Photobiol Sci. 1: 225-236. DOI: https://doi.org/10.1039/b201230h.

J Fitopatol Indones Yuliani et al
66
Rastogi RP, Richa, Kumar A, Tyagi MB, Rajeshwar PS. 2010. Molecular mechanisms of ultraviolet radiation-induced DNA damage and repair. J Nucleic Acids. doi:10.4061/2010/592980. DOI: https://doi.org/10.4061/2010/592980.
Thein A, Prathuangwong S. 2010. Novel strains of Xanthomonas oryzae pv. oryzae uv mutated induce systemic resistance in
rice against bacterial leaf blight disease. Kasetsart J. 44:1026–1043.
Wahyudi AT, Meliah S, Nawangsih AA. 2011. Xanthomonas oryzae pv. oryzae bakteri penyebab hawar daun pada padi: isolasi, karakterisasi, dan telaah mutagenesis dengan transposon. Makara Sains. 15(1):89-96.

ISSN: 0215-7950
67
Volume 13, Nomor 2, Maret 2017Halaman 67–71
DOI: 10.14692/jfi.13.2.67ISSN: 0215-7950
*Alamat penulis korespondensi: Jurusan Proteksi Tanaman, Fakultas Pertanian Universitas Lampung. Jalan Prof. Dr. Soemantri Brodjonegoro No. 1 Bandar Lampung, 35142.Tel: 0721-787029, Faks: 0721-787029, Surel: [email protected]
KOMUNIKASI SINGKAT
Penyakit Busuk Akar dan Pangkal Batang Tebu di Sumatera Selatan
Sugarcane Root and Basal Stem Rot Disease in South Sumatera
Tri Maryono*, Ani Widiastuti, Achmadi PriyatmojoUniversitas Gadjah Mada, Yogyakarta 55281
ABSTRAK
Penyakit busuk akar dan pangkal batang tebu yang disebabkan oleh Xylaria sp. merupakan salah satu penyakit baru diperkebunan tebu Indonesia. Sampai saat ini, penyakit busuk akar dan pangkal batang tebu hanya dilaporkan ada di Sumatera dan informasi mengenai penyakit ini masih terbatas. Penelitian dilakukan untuk mendeskripsikan gejala, tanda, dan pola sebaran penyakit di lapangan yang dapat digunakan sebagai dasar untuk menentukan keberadaan penyakit di lapangan. Survei gejala, tanda, dan pola sebaran penyakit dilakukan di perkebunan tebu yang terdapat di Sumatera Selatan (3° 26’ 16.6” LS dan 104° 40’ 09.8” BT). Gejala khas serangan Xyalaria sp. ialah tanaman tebu mengering seluruh daunnya dan mati, akar dan pangkal batang busuk kering, dan ratoon tidak tumbuh. Gejala tanaman tebu mati, tanda penyakit berupa stroma pada pangkal batang atau di tanah sekitar tanaman sakit, dan pola sebaran penyakit mengelompok merupakan indikator yang kuat mengenai keberadaan Xylaria di perkebunan tebu.
Kata kunci: pola sebaran, tanda penyakit, ratoon, Xylaria
ABSTRACT
Root and basal stem rot disease caused by Xylaria sp. is a new diseases in Indonesian sugarcane plantation. The disease has been reported only occurred in sugarcane plantations in Sumatera. Limited information about the disease has slowed down disease diagnosis in the field. The research was aimed to describe symptoms, signs, and spatial distribution of the disease caused by Xylaria. A survey was conducted at a sugarcane plantation in South Sumatera (3° 26’ 16.6” S dan 104° 40’ 09.8” E). Typical symptoms of the disease involved drying of all leaves and plant death, dry rot of root and basal stem, and retarded growth of ratoon cane were recorded. Stroma was found on surface of diseased stems and on soil surface of diseased plants, and spatial aggregation of diseased plants was observed in all locations. The distribution of the diseased plant aggregately and the presence of stroma on diseased plants are important characteristics of the disease diagnosis in sugarcane plantations.
Keywords: disease distribution, disease sign, ratoon, Xylaria

J Fitopatol Indones Maryono et al
68
Penyakit busuk akar dan pangkal batang atau disebut penyakit lapuk akar dan pangkal batang atau juga disebut penyakit xylaria merupakan penyakit yang relatif baru di perkebunan tebu Indonesia. Penyakit tersebut pertama ditemukan pada tahun 1993 di perkebunan tebu PT Gunung Madu Plantation, Provinsi Lampung (Hersanti dan Sitepu 2005; Sitepu et al. 2010). Sampai saat ini penyakit hanya dilaporkan pada perkebunan tebu di Sumatera (Achadian et al. 2012). Selain di Indonesia, penyakit tersebut juga terdapat di Taiwan, Amerika Serikat, dan Puerto Rico (Fang dan Lee 2000). Penyakit xylaria yang ada di Lampung disebabkan oleh Xylaria cf warburgii (Sitepu et al. 2010), sama seperti yang di Taiwan (Fang et al. 1986). Namun, di Amerika Serikat dan Puerto Rico penyakit tersebut sebabkan oleh Xylaria arbuscula (Fang dan Lee 2000).
Penyakit busuk akar dan pangkal batang tebu memiliki arti ekonomi yang penting pada budi daya tebu karena dapat mematikan tanaman, menurunkan bobot batang, dan menurunkan rendemen. Selain itu juga mengakibatkan tanaman keprasan (ratoon cane) gagal tumbuh karena tanaman induknya mati. Kerugian tersebut akan lebih tinggi pada tanaman ratoon dibandingkan dengan tanaman dari bibit (plant cane) dan akan terus meningkat pada generasi tanaman ratoon berikutnya. Kerugian lainnya ialah apabila pada tanaman ratoon pertama serangannya tinggi maka harus dilakukan penanaman ulang (replanting) untuk menghindari kerugian yang lebih besar. Di Lampung keparahan penyakit 26% dapat menyebabkan penurunan hasil sampai 15% (Sitepu et al. 2010).
Sampai saat ini, informasi mengenai penyakit xylaria masih terbatas karena belum banyak dikaji khususnya di Indonesia. Di sisi lain keberadaan penyakit xylaria relatif sulit untuk ditentukan terutama pada tanaman induk (Hersanti dan Sitepu 2005).
Survei penyakit xylaria dilakukan dari Juni–Agustus 2016 di perkebunan tebu di Sumatera Selatan (3° 26' 16.6" LS dan 104°
40' 09.8" BT). Tanaman tebu yang diamati ialah tanaman ratoon varietas PS 882 berumur
9 bulan. Pengamatan dilakukan pada tiga petak yang berbeda dengan luas petak 10, 12.5, dan 16 Ha. Pengamatan gejala penyakit dilakukan secara visual meliputi gejala pada tajuk tanaman, gejala pada akar, dan gejala pada jaringan pangkal batang. Pengamatan gejala pada bagian akar dilakukan dengan mencabut tanaman sakit, sedangkan pengamatan gejala pada jaringan pangkal batang dilakukan dengan memotong secara membujur pangkal batang tanaman sakit. Pengamatan tanda penyakit juga dilakukan secara visual pada tanaman sakit dan sekitar tanaman sakit. Pengamatan pola sebaran penyakit dilakukan pada satu musim tanam dengan mengamati sebaran gejala kerusakan tanaman tebu di lapangan berupa tanaman ratoon yang tidak tumbuh. Pengamatan dilakukan pada dua titik pada masing-masing petak pengamatan. Untuk memastikan bahwa ratoon yang tidak tumbuh disebabkan oleh Xylaria maka keberadaan stroma Xylaria pada sekitar titik tersebut dijadikan dasarnya. Hasil pengamatan sebaran penyakit xylaria yang diperoleh dibandingkan dengan model pola sebaran penyakit yang dikemukakan Brown (1997).
Kegiatan survei yang dilakukan berhasil menemukan gejala penyakit xylaria pada tanaman tebu. Gejala yang ditemukan berupa gejala awal dan gejala akhir, yaitu matinya tanaman. Gejala awal penyakit ialah daun menguning dan mengering dari ujung daun (Gambar 1a). Seiring berjalannya waktu, semua daun akan mengering yang menandakan perakaran dan pangkal batang sudah rusak parah. Pada serangan berat, rumpun tanaman akan mengering dan mati (Gambar 1b). Tanaman yang mati mudah dicabut karena akar-akarnya juga mati. Akar tanaman sakit tampak membusuk (busuk kering) dan berwarna hitam (Gambar 1d). Apabila pangkal batang tebu sakit dibelah maka jaringan pangkal batang tebu terlihat cokelat muda dan kemerahan, serta terdapat garis berwarna hitam (Gambar 1d) yang merupakan ciri khas serangan Xylaria. Apabila tanaman induk mengalami kerusakan yang berat atau mati maka tanaman ratoon tidak akan tumbuh sehingga akan tampak kosong dan dipenuhi gulma.

J Fitopatol Indones Maryono et al
69
a b c d
Gambar 1 Gejala penyakit xylaria pada tanaman tebu. a, Daun menguning dan mengering dari ujung daun; b, Rumpun tanaman tebu mati; c, Akar tanaman sakit yang tampak menghitam; dan d, Penampang membujur pangkal batang tebu sakit dan massa hifa. Ciri khas serangan Xylaria spditunjukkan dengan tanda panah.
Tanda penyakit berupa stroma berhasil ditemukan di lapangan pada batang tebu sakit dan tanah di sekitar tanaman tebu sakit (Gambar 2). Ada dua bentuk stroma yang berhasil ditemukan. Stroma yang pertama berwarna hitam, berkelompok dan bercabang banyak, dan bagian ujungnya berwarna putih (Gambar 2a). Stroma yang kedua berbentuk batang silindris dengan semakin meruncing keujungnya, tunggal, bertangkai pendek, berwarna hitam atau kecoklatan, dan tidak bercabang atau bercabang pada pangkalnya (Gambar 2b dan 2c). Adanya stroma merupakan salah satu ciri khas Xylaria (Ma et al. 2011; Srihanant et al. 2015; Huang et al. 2015)
Sebaran tanaman ratoon yang tidak tumbuh terlihat berkelompok pada semua
titik pengamatan. Pada satu baris bisa terdapat banyak tanaman ratoon yang tidak tumbuh sedangkan pada baris sebelahnya hanya ada beberapa tanaman ratoon tidak tumbuh atau tumbuh semua. Penyebaran secara berkelompok mencirikan bahwa penyebab penyakit merupakan cendawan tular tanah, seperti yang dilaporkan oleh Stewart et al. (2014).
Hasil isolasi penyebab penyakit dari bagian tanaman sakit, setelah ditumbuhkan pada medium agar-agar dektrosa kentang, didapatkan cendawan yang awalnya berwarna putih yang akan berubah menjadi gelap yang dimulai dari bagian tengahnya (Gambar 3a). Pada biakan yang berumur lebih dari dua minggu akan tumbuh stroma yang bercabang banyak, yang awalnya berwarna putih, namun
Gambar 2 Stroma Xylaria sp. pada tanaman tebu. a, Stroma pada pangkal tanaman tebu sakit; b, Stroma yang keluar dari tanah di sekitar tanaman sakit; dan c, Stroma yang tumbuh dari sisa-sisa tanaman sakit.
a b c

J Fitopatol Indones Maryono et al
70
Gambar 3 Karakter morfologi penyebab Xylaria sp. pada tanaman tebu. a, Biakan cendawan Xylaria sp. pada medium agar-agar dekstrosa kentang; b, Stroma yang terbentuk pada medium agar-agar miring; dan c, Askospora (perbesaran 100×).
a b c
pada akhirnya berwarna hitam (Gambar 3b). Spora yang didapat dari stroma berbentuk lonjong dan berwarna hitam (Gambar 3c).Berdasar karakter morfologi tersebut, cendawan yang didapat adalah Xylaria sp. Secara umum Xylaria hidup sebagai saprob atau sebagai parasit lemah dan sedikit yang menjadi patogen (Whalley 1996), bersimbiosis dengan rayap (Rogers et al. 2005; Ju dan Hsieh 2007), dan sebagai endofit (Ibrahim et al. 2014).
Hasil survei ini menujukkan bahwa penyakit busuk akar dan pangkal batang tebu telah berkembang di perkebunan tebu di Sumatera Selatan. Adanya tanaman tebu yang mengering dan mati, adanya tanaman ratoon yang tidak tumbuh yang tersebar secara berkelompok dan adanya stroma pada pangkal batang tanaman sakit atau di tanah sekitar tanaman sakit dapat digunakan sebagai petunjuk adanya penyakit xylaria pada suatu perkebunan tebu.
DAFTAR PUSTAKA
Achadian E, Kristini A, Magarey R, Sallam N, Samson P, Goebel FR, Lonie K. 2012. Sugarcane pests and diseases–Field guide–Indonesian version. Indonesian Sugar Research Institute, BSES Limited, Australian Centre for International Research. Montpellier (FR): CIRAD.
Brown J. 1997. Survival and dispersal of plant parasites: general concepts. Di dalam: Brown JF, Ogle HJ, editor. Plant
Pathogens and Plant Diseases. New South Wales (AU): Rockvale Publication. Hlm 195–206.
Fang JG, Hsieh WH, Hu CH, Lee CS. 1986. Survival of Xylaria sp. in the soil and the effect of green manure application on disease incidence. Report of the Taiwan Sugar Research Institute. 112:21–26.
Fang JG, Lee CS. 2000. Root and basal stem rot. Di dalam: Rott P, Bailey RA, Comstock JC, Croft BJ, Saumtally AS, editor. A Guide to Sugarcane Diseases. Montpellier (FR): CIRAD-ISSCT. Hlm 170–173.
Hersanti, Sitepu R. 2005. Identifikasi penyebab penyakit lapuk akar dan pangkal batang (LAPB) tebu di PT Gunung Madu Plantations Lampung Tengah. Biotika. 4(1):24–27.
Huang G, Wang R, Guo L, Liu N. 2015. Three new species of Xylaria from China. Mycotaxon. 130:299–304. DOI: https://doi.org/10.5248/130.299.
Ibrahim A, Sørensen D, Jenkins HA, McCarry BE, Sumarah, MW. 2014. New diplosporin and agistatine derivatives produced by the fungal endophyte Xylaria sp. isolated from Vitis labrusca. Phytochemistry Letters. 9:179–183. DOI: https://doi.org/10.1016/j.phytol.2014.06.011.
Ju YM, Hsieh HM. 2007. Xylaria species associated with nests of Odontotermes formosanusin Taiwan. Mycologia. 99:936–957. DOI: http://dx.doi.org/10.1080/15572536.2007.11832525.

J Fitopatol Indones Maryono et al
71
Ma HX, Vasilyeva L, Li Y. 2011. A new species of Xylaria from China. Mycotaxon. 116:151–155. DOI: https://doi.org/10.5248/116.151.
Rogers J, Ju Y-M, Lehmann J. 2005. Some Xylariaon termite nests. Mycologia. 97:914–923. DOI: http://dx.doi.org/10.1080/15572536.2006.11832783.
Sitepu R, Sunaryo, Widyatmoko K, Purwoko H. 2010. Root and basal stem rot disease of sugarcane in Lampung, Indonesia. Di dalam: Prosiding Kongress XXVII International Society of Sugar Cane Technologists; 2010 Mar 7–11; Veracruz (MX): ISSCT. hlm 1–7.
Srihanant N, Petcharat V, Vasilyeva LN. 2015. Xylaria thailandica – a new species from Southern Thailand. Mycotaxon. 130:227–231. DOI 10.5248/130.227.
Stewart LR, Morrison ML, Appel DN, Wilkins RN. 2014. Spatial and temporal distribution of Oak wilt in Golden-Cheeked Warbler habitat. Wildlife Soc Bull. 38(2):288–296. DOI: https://doi.org/10.1002/wsb.395.
Whalley AJS. 1996. The xylariaceous way of life. Mycol Res.100:897–922. DOI: https://doi.org/10.1016/S0953-7562(96)80042-6.




















