
ZM447439, a Novel Promising Aurora Kinase Inhibitor, Provokes Antiproliferative and Proapoptotic...
10
Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Original Paper Neuroendocrinology DOI: 10.1159/000258705 ZM447439, a Novel Promising Aurora Kinase Inhibitor, Provokes Antiproliferative and Proapoptotic Effects Alone and in Combination with Bio- and Chemotherapeutic Agents in Gastroenteropancreatic Neuroendocrine Tumor Cell Lines I. Georgieva a D. Koychev b Y. Wang a J. Holstein a W. Hopfenmüller c M. Zeitz a P. Grabowski a, d a Gastroenterologie/Infektiologie/Rheumatologie, Medizinische Klinik I, b Hämatologie/Onkologie und Transfusionmedizin, Medizinische Klinik III, und c Institut für Biometrie und Klinische Epidemiologie, Charité-Universitätsmedizin Berlin, Campus Benjamin Franklin, Berlin, und d Abteilung für Gastroenterologie/ Onkologie/Endokrinologie, Zentralklinik Bad Berka, Bad Berka, Germany proliferation of all three cell lines with IC 50 values in the nanomolar to low micromolar range. Moreover, aurora ki- nase inhibition by ZM447439 potently induced apoptosis, which was accompanied by DNA fragmentation and caspase 3 and 7 activation. Furthermore, we observed cell cycle ar- rest at G 0 /G 1 phase as well as a block in G 2 /M transition. In addition, combined treatment with the chemotherapeutic agents streptozocin and cisplatin augmented significantly the antiproliferative effects of those agents. Conclusion: Au- rora kinase inhibition by ZM447439 seems to be a promising new therapeutic approach in GEP-NETs, which should be evaluated in further clinical trials. Copyright © 2009 S. Karger AG, Basel Introduction Gastroenteropancreatic neuroendocrine tumors (GEP-NETs) represent a relatively rare and heterogeneous tumor entity. More than half of GEP-NETs is function- Key Words Apoptosis Aurora kinase inhibitor Cell cycle Gastroenteropancreatic neuroendocrine tumors Prognostic markers Proliferation ZM447439 Abstract Background: Therapeutic approaches to gastroenteropan- creatic neuroendocrine tumors (GEP-NETs) are still not satis- factory. A new direction in treatment options could be the novel aurora kinase inhibitor ZM447439, which was previ- ously reported to interfere with the mitotic spindle integrity checkpoint and chromosome segregation, but does not in- terfere with other kinases when used up to 5 M. Methods: We evaluated the antineoplastic effects of ZM447439 on growth and apoptosis of the GEP-NET cell lines BON, QGP-1 and MIP-101, representing the different malignant tumor types, using standard cell biological tests as crystal violet as- says, caspase activation, DNA fragmentation and cell cycle analysis. Results: ZM447439 dose-dependently inhibited Received: October 21, 2008 Accepted after revision: May 7, 2009 Published online: November 14, 2009 Patricia Grabowski Charité-Universitätsmedizin Berlin Campus Benjamin Franklin, Hindenburgdamm 30 DE–12200 Berlin (Germany) Tel. +49 30 8445 4579, Fax +49 30 8445 4481, E-Mail patricia.grabowski @ charite.de © 2009 S. Karger AG, Basel 0028–3835/09/0000–0000$26.00/0 Accessible online at: www.karger.com/nen
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of ZM447439, a Novel Promising Aurora Kinase Inhibitor, Provokes Antiproliferative and Proapoptotic...

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Neuroendocrinology DOI: 10.1159/000258705
ZM447439, a Novel Promising Aurora Kinase Inhibitor, Provokes Antiproliferative and Proapoptotic Effects Alone and in Combination with Bio- and Chemotherapeutic Agents in Gastroenteropancreatic Neuroendocrine Tumor Cell Lines
I. Georgieva a D. Koychev b Y. Wang a J. Holstein a W. Hopfenmüller c
M. Zeitz a P. Grabowski a, d
a Gastroenterologie/Infektiologie/Rheumatologie, Medizinische Klinik I, b Hämatologie/Onkologie und Transfusionmedizin, Medizinische Klinik III, und c Institut für Biometrie und Klinische Epidemiologie, Charité-Universitätsmedizin Berlin, Campus Benjamin Franklin, Berlin , und d Abteilung für Gastroenterologie/Onkologie/Endokrinologie, Zentralklinik Bad Berka, Bad Berka , Germany
proliferation of all three cell lines with IC 50 values in the nanomolar to low micromolar range. Moreover, aurora ki-nase inhibition by ZM447439 potently induced apoptosis, which was accompanied by DNA fragmentation and caspase 3 and 7 activation. Furthermore, we observed cell cycle ar-rest at G 0 /G 1 phase as well as a block in G 2 /M transition. In addition, combined treatment with the chemotherapeutic agents streptozocin and cisplatin augmented significantly the antiproliferative effects of those agents. Conclusion: Au-rora kinase inhibition by ZM447439 seems to be a promising new therapeutic approach in GEP-NETs, which should be evaluated in further clinical trials.
Copyright © 2009 S. Karger AG, Basel
Introduction
Gastroenteropancreatic neuroendocrine tumors (GEP-NETs) represent a relatively rare and heterogeneous tumor entity. More than half of GEP-NETs is function-
Key Words
Apoptosis � Aurora kinase inhibitor � Cell cycle � Gastroenteropancreatic neuroendocrine tumors � Prognostic markers � Proliferation � ZM447439
Abstract
Background: Therapeutic approaches to gastroenteropan-creatic neuroendocrine tumors (GEP-NETs) are still not satis-factory. A new direction in treatment options could be the novel aurora kinase inhibitor ZM447439, which was previ-ously reported to interfere with the mitotic spindle integrity checkpoint and chromosome segregation, but does not in-terfere with other kinases when used up to 5 � M . Methods: We evaluated the antineoplastic effects of ZM447439 on growth and apoptosis of the GEP-NET cell lines BON, QGP-1 and MIP-101, representing the different malignant tumor types, using standard cell biological tests as crystal violet as-says, caspase activation, DNA fragmentation and cell cycle analysis. Results: ZM447439 dose-dependently inhibited
Received: October 21, 2008 Accepted after revision: May 7, 2009 Published online: November 14, 2009
Patricia Grabowski Charité-Universitätsmedizin Berlin Campus Benjamin Franklin, Hindenburgdamm 30 DE–12200 Berlin (Germany) Tel. +49 30 8445 4579, Fax +49 30 8445 4481, E-Mail patricia.grabowski @ charite.de
© 2009 S. Karger AG, Basel0028–3835/09/0000–0000$26.00/0
Accessible online at:www.karger.com/nen

Georgieva /Koychev /Wang /Holstein /Hopfenmüller /Zeitz /Grabowski
Neuroendocrinology 2
ally inactive and usually diagnosed once signs and symp-toms of tumor metastases occur [1] . The other half of all metastasized GEP-NETs releases large quantities of bio-active amines and/or neurotransmitters and thereby causes hypersecretion syndromes that are characteristic of these tumors. These clinical symptoms can be well controlled generally by somatostatin analogues or inter-feron. However, therapeutic options to prevent growth and further dissemination of GEP-NETs are limited. Growth behavior of GEP-NETs differs widely, from very slow/moderately growing to very fast growing, aggressive tumors. The distinction between benign and malignant GEP-NETs is very important and has not yet been com-pletely resolved. Klöppel et al. [2] presented an updated World Health Organization system for the classification of GEP-NETs consisting of three main categories: well-differentiated endocrine tumors (WDETs), well-differen-tiated endocrine carcinomas (WDECs) and poorly dif-ferentiated endocrine carcinomas (PDECs) with the ob-jective to provide a prognostically relevant classification system that assessed tumors according to size, prolifera-tive activity, angioinvasion, organ invasion, metastases, hormone activity and clinical syndromes. This has some impact on therapeutic considerations: Patients with low-differentiated neuroendocrine cancers can profit from cytoreductive chemotherapy, whereas patients with slow-ly growing, well-differentiated endocrine small intestine tumors do not have a survival advantage by chemothera-py [3] . The therapeutic possibilities for this group of pa-tients are completely unsatisfactory and innovative treat-ment strategies, which are both effective and well toler-ated, have to be developed.
The aurora family of serine/threonine kinases plays an important role in chromosome alignment, segregation and cytokinesis during mitosis. The family consists of three members: aurora A, B and C. Aurora A is localized in the pericentrosomal region, where it recruits several important components on the forming mitotic spindle during mitosis [4, 5] . So far, aurora A is the only family member that has been reported to act as an oncogene [4, 6, 7] . Aurora B is a chromosomal passenger protein that localizes at centromeres during prometaphase and subse-quently relocates to midzone microtubules and midbod-ies during anaphase and telophase [4, 8] . It forms a com-plex with the proteins INCENP and survivin, which was shown to be a powerful novel prognostic marker for GEP-NET disease by our group [9] . Aurora B plays a role in chromosome alignment, kinetochore microtubule biori-entation, activation of the spindle assembly checkpoint and cytokinesis in association with phosphorylation of
histone H3 [10] . Recent studies described aberrant ex-pression of aurora A and B in a variety of solid tumors, including prostate [11] , colon [12] , pancreas [13] , breast [14] and thyroid cancers [15] . Aurora kinases A and B were reported as an attractive anti-cancer drug target [16–19], and aurora C was localized to the chromosome passenger complex [20] .
Usually, after treatment with DNA-damaging agents, cells pause at the G 2 /M checkpoint of the cell cycle to re-pair DNA damage before proceeding through mitosis. On the other hand, aurora B levels are increased in cells in G 2 /M phase – including those treated with DNA-dam-aging agents [21, 22] . Therefore, we hypothesized that ad-dition of an aurora kinase inhibitor may potentate the anti-proliferative effect of a cytotoxic drug. Previous studies with the novel, highly selective aurora kinase B inhibitor, ZM447439, have reported an accumulation of cells with 4N DNA content in a variety of human cancer cell lines (A549, MCF-7, DLD-1, HeLa and U2OS) [21] . ZM447439 has been proven to be an effective apoptosis-inducing agent in acute myeloid leukemia cell lines and Hep2 cancer cell lines [23, 24] .
Little is known about the functional role of aurora ki-nases in GEP-NETs. The aim of this study was therefore to determine the preclinical profile of the aurora kinase B inhibitor, ZM447439, as a candidate for anti-cancer therapy in this special tumor entity.
Materials and Methods
Cell Lines and Drugs Human pancreatic BON cells, which were established from a
human pancreatic carcinoid tumor, provide a useful model to study the biology of NETs in vitro [25] . BON cells were grown in a 1: 1 mixture of DMEM and Ham’s F-12 medium containing 10% FCS (Biochrom, Berlin, Germany) and 1% L -glutamine.
The human well-differentiated pancreatic carcinoma cell line, QGP-1 [26] , was cultured in RPMI 1640 (PAA, Cölbe, Germany) containing 10% fetal bovine serum, 100 U/ml penicillin and streptomycin and 1% L -glutamine. MIP-101 [27] , an undifferenti-ated colorectal carcinoma cell line, was kindly provided fromI. Tai (Vancouver, B.C., Canada). MIP-101 was grown in DMEM containing 10% fetal bovine serum. The same medium was used for culturing LCC-18, Colo205 and LS-174-T colon carcinoma cell lines. The colon carcinoma cell lines NCI-H716, NCI-H747 and Colo320 were cultured in RPMI 1640 (PAA) containing 10% FCS. The human colon adenocarcinoma cell line, HT 29, was used as negative control and was cultured in RPMI 1640 containing 10% fetal bovine serum. All cell lines were kept at 37 ° C in a humidi-fied atmosphere (5% CO 2 ).
ZM447439 was purchased from Tocris, Ellisville, Mo., USA. Cisplatin, 5-fluorouracil (5-FU), doxorubicin, streptozocin and octreotide were purchased from Sigma (Steinheim, Germany).

ZM447439 in Gastroenteropancreatic Neuroendocrine Tumors
Neuroendocrinology 3
SOM230 was kindly provided by Novartis Pharma (Nuremberg, Germany). Stock solutions were prepared in DMSO and stored at –20 ° C. Immediately before use, the drugs were diluted to the final concentration in fresh media. In all experiments, the final DMSO concentration was ! 0.5%, not affecting cell growth.
To evaluate the effects of ZM447439, cells were incubated with either control medium or medium containing rising concentra-tions of ZM447439. Media were changed every other day to ensure constant drug concentrations in the incubation medium.
Flow Cytometry The antibodies were purchased from Sigma if not mentioned
otherwise. Cells were trypsinized, washed twice with phosphate-buffered NaCl solution (PBS; 140 m M NaCl, 10 m M Na 2 HPO 4 , 2.6 m M KCl and 1.4 m M KH 2 PO 4 , pH 7.4). Cells were then fixed in 4% paraformaldehyde for 30 min at room temperature and washed once with PBS; the supernatant was removed, and 100 � l Fix and Perm Medium A (Caltag Laboratories, Vienna, Austria) were added. After a 15-min incubation, the cells were washed twice with 1 ml PBS. The supernatant was removed and the respective first antibody was added at the appropriate concentration: anti-synaptophysin (1: 1,000), anti-chromogranin A (CgA; 1: 100), anti-syntaxin (1: 1,000), anti-synaptobrevin (VAMP; 1: 100), SNAP25 (1: 1,000), mouse IgG-negative isotype control (Dako, Glostrup, Denmark, 1: 10) and incubated � 60 min at room temperature. Afterwards, the cells were washed twice in 1 ml PBS and 4 � g/ml secondary Alexa TM 488-labeled goat-anti mouse IgG antibody (Molecular Probes, Eugene, Oreg., USA) for 1 h in a dark place on ice. After the incubation, cells were washed twice with 1 ml of PBS. Finally, 300 � l of 1 ! PBS were added to the cells, and cells were transferred into FACS tubes and fluorescence was detected by flow cytometry on a FACSCalibur (BD, Heidelberg, Germany) and analyzed using CellQuest software (BD). The cell samples already prepared for flow cytometry were also used for immuno-fluorescence. The stained cells were transferred and pelleted onto coverslips using spin-down tubes after centrifugation in a swing-ing bucket rotor at 10,000 rpm for 10 min and then examined in a Leica fluorescent microscope.
Western Blot Analysis Whole cell extracts were prepared by harvesting and lysing the
cells with lysis buffer (0.1 SDS, 0.5% sodium deoxycholic acid, 1% Nonidet P-40, 0.1 m M PMSF, 1 � g/ml aprotinin and 1 � g/ml pep-statin A). The protein content of the lysate was determined using the BCA protein assay kit (Pierce, Rockford, Ill., USA). The cell lysate was mixed with gel loading buffer (62.5 m M Tris-HCl, 10% glycerol, 1% SDS and 2.5% � -mercaptoethanol). After boiling for 10 min, the lysates were subjected to SDS/polyacrylamide gel elec-trophoresis (40 � g protein/lane; gel: 12% polyacrylamide, 0.1% SDS and 25 m M Tris-HCl; running buffer: 25 m M Tris base, 50 m M glycine and 0.1% SDS). Proteins were transferred to PVDF membranes by electroblotting using transfer buffer (25 m M Tris base, 50 m M glycine and 20% methanol). Blots were blocked in 5% BSA and then incubated at 4 ° C overnight with the followingantibodies: survivin (1: 500, Biozol, Eching, Germany), AIM-1(1: 250, BD Biosciences, Franklin Lakes, N.J., USA) and � -actin(1: 5,000, Sigma-Aldrich). After washing with PBS containing 0.1% Tween-20 and incubating with horseradish peroxidase-cou-pled anti-IgG antibody (1: 10,000) at room temperature for 1 h, the blot was washed extensively and developed using enhanced che-
miluminescent detection (Amersham, Uppsala, Sweden). Blots were exposed to Hyperfilm ECL film (Amersham, Uppsala, Swe-den) for 1–30 min and analyzed densitometrically using the Im-age J software (National Institutes of Health, Bethesda, Md., USA). As a loading control, � -actin (mouse-anti-human � -actin antibody from Sigma, Deisenhofen, Germany) was detected on each blot.
Measurement of Growth Inhibition Cell number was evaluated by crystal violet staining, as de-
scribed [28] . In brief, cells in 96-well plates were fixed with 1% glutaraldehyde. Then cells were stained with 0.1% crystal violet. The unbound dye was removed by washing with water. Bound crystal violet was solubilized with 0.2% Triton X-100. Light ex-tinction which increases linearly with the cell number was ana-lyzed at 570 nm using an ELISA reader.
Drug Combination Studies To check for possible additive or synergistic effects of
ZM447439 with conventional biotherapeutic and cytotoxic drugs, combination treatment was performed. ZM447439 plus octreo-tide and SOM 230 in the BON cell line, ZM447439 plus strepto-zocin or doxorubicin in the QGP-1 cell line and ZM447439 plus cisplatin and 5-FU in the MIP-101 cell line were evaluated. The cytotoxic activities of the combinations were compared to those of each drug alone. For all experiments, cell number was evalu-ated by crystal violet staining, as described above.
Functional Detection of Apoptosis The activity of caspases 3 and 7 was calculated from the cleav-
age of fluorogenic substrates as described by the manufacturer (Apo-ONE Homogeneous Caspase-3/7 Assay; Promega, Mann-heim, Germany). Substrate cleavage was measured fluorometri-cally with a VersaFluor fluorometer (Bio-Rad, Munich, Germany) with optimal excitation wavelength detection at 499 nm.
DNA fragmentation was determined using a DNA laddering kit (Roche, Mannheim, Germany) according to the manufactur-er’s instructions. Briefly, after 24 h of incubation, cells were lysed in incubation buffer. The lysed samples were poured into cen-trifugation tubes containing glass fiber fleece which binds the apoptotic DNA in the presence of guanidine hydrochloride. After centrifugation and additional washing steps, apoptotic DNA was eluted by addition of warm elution buffer and collected by cen-trifugation. Purified DNA (2 � g/sample) was mixed with gel loading buffer and analyzed on agarose gel.
Cell Cycle Analysis After harvesting 1 ! 10 6 single cells in cold PBS, cells were
centrifuged twice for 5 min at 1,500 rpm. The pellet was resuspended in 1 ml of ice-cold 70% ethanol in
PBS (–20 ° C) and incubated for at least 2 h at –20 ° C. Then the cells were centrifuged and the pellet was resuspended in 1 ml PBS (room temperature) and again pelleted. The pellet was resuspend-ed in 1 ml propidium iodide/Triton X-100 staining solution with RNAse A and incubated 30 min at room temperature in the dark, and then the samples were analyzed flow cytometrically using ModFit software (BD).
Georgieva /Koychev /Wang /Holstein /Hopfenmüller /Zeitz /Grabowski
Neuroendocrinology 4
Statistical Analysis If not stated otherwise, data were expressed as means of at least
three independent experiments ( 8 SEM). Significance between controls and treated samples was calculated by Student’s two-sid-ed t test. Caspase 3 measurements were evaluated using the two-sided Welch t test. p ! 0.05 was considered statistically signifi-cant.
Results
Characteristics of GEP-NET Cell Lines and Aurora B Expression First, we searched for cancer cell lines possessing neu-
roendocrine characteristics using flow cytometry. In or-der to prove neuroendocrine differentiation of the cell lines, we looked for expression of at least two of the follow-ing markers: CgA, synaptophysin, syntaxin, SNAP25 and synaptobrevin ( table 1 ; fig. 1 ). From cell lines meeting these criteria, we chose for all further experiments cell lines which represent the different growth patterns of neu-roendocrine tumors. The BON cell line was used as a model for WDETs, whereas QGP-1 cells served as a mod-el for WDECs. So far, there is no established cancer cell line representing PDEC. In a number of experiments, we tried to establish cell lines as models for this special entity ( table 1 ). LCC-18 showed some neuroendocrine character-istics, although it was repetitively negative for syntaxin, SNAP25 and synaptobrevin. Caco2 and NCI-H716 also showed some neuroendocrine characteristics, but they were not sufficient to be included in this study. We report here that the poorly differentiated highly malignant colorectal cancer cell line MIP-101 also expresses neuro-endocrine markers. Since the cell lines BON, QGP-1 and MIP-101 represent the different growth patterns of neuro-endocrine tumors, they were used for further studies.
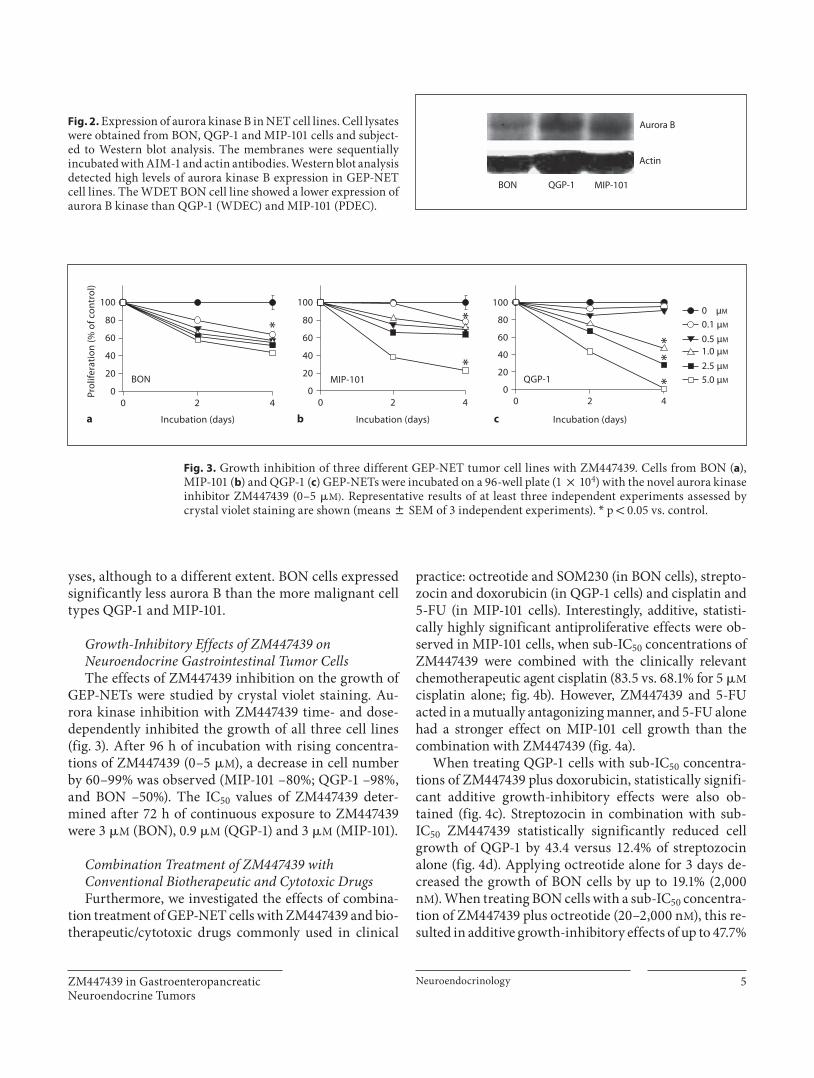
As shown in figure 2 , we detected in all three cell lines selected aurora kinase B expression by Western blot anal-
Table 1. Expression of neuroendocrine markers in GEP-NET cell lines
Cell lines Origin Synaptophysin CgA SNAP25 Synaptobrevin Syntaxin
HT 29 [35] colon carcinoma – – – – –BON [36] pancreas NET + + –/+ –/+ +Caco2 [37] colon carcinoma – – – –/+ –Colo205 [38] colon carcinoma – – – – –Colo320 [39] colon carcinoma – + – – –LCC-18 [40] colon carcinoma + + – – –LS-174-T [41] colon carcinoma – – – – –MIP-101 [27] colon carcinoma + – – –/+ +NCI-H716 [42] colon carcinoma + –/+ – –/+ +NCI-H747 [42] colon carcinoma – – – – –QGP-1 [26] pancreas NEC + + –/+ – +
Cells from 11 GEP-NET cell lines (1 ! 105) were washed, fixed, incubated with 1st and 2nd antibody and mean fluorescence was calculated at FACS against the isotype control. The HT 29 colon cancer cell line was used as a negative control. Here we show representative results of at least three independent experiments made in duplicate.
Fig. 1. Expression of synaptophysin in the cytoplasm of MIP-101 cells (marked with FITC-labeled antibody). The cells labeled with fluorescent antibodies were spun down onto coverslips using spin-down tubes and examined by a Leica fluorescent micro-scope. Here we show a representative picture of at least three in-dependent stainings.
ZM447439 in Gastroenteropancreatic Neuroendocrine Tumors
Neuroendocrinology 5
yses, although to a different extent. BON cells expressed significantly less aurora B than the more malignant cell types QGP-1 and MIP-101.
Growth-Inhibitory Effects of ZM447439 on Neuroendocrine Gastrointestinal Tumor Cells The effects of ZM447439 inhibition on the growth of
GEP-NETs were studied by crystal violet staining. Au-rora kinase inhibition with ZM447439 time- and dose-dependently inhibited the growth of all three cell lines ( fig. 3 ). After 96 h of incubation with rising concentra-tions of ZM447439 (0–5 � M ), a decrease in cell number by 60–99% was observed (MIP-101 –80%; QGP-1 –98%, and BON –50%). The IC 50 values of ZM447439 deter-mined after 72 h of continuous exposure to ZM447439 were 3 � M (BON), 0.9 � M (QGP-1) and 3 � M (MIP-101).
Combination Treatment of ZM447439 with Conventional Biotherapeutic and Cytotoxic Drugs Furthermore, we investigated the effects of combina-
tion treatment of GEP-NET cells with ZM447439 and bio-therapeutic/cytotoxic drugs commonly used in clinical
practice: octreotide and SOM230 (in BON cells), strepto-zocin and doxorubicin (in QGP-1 cells) and cisplatin and 5-FU (in MIP-101 cells). Interestingly, additive, statisti-cally highly significant antiproliferative effects were ob-served in MIP-101 cells, when sub-IC 50 concentrations of ZM447439 were combined with the clinically relevant chemotherapeutic agent cisplatin (83.5 vs. 68.1% for 5 � M cisplatin alone; fig. 4 b). However, ZM447439 and 5-FU acted in a mutually antagonizing manner, and 5-FU alone had a stronger effect on MIP-101 cell growth than the combination with ZM447439 ( fig. 4 a).
When treating QGP-1 cells with sub-IC 50 concentra-tions of ZM447439 plus doxorubicin, statistically signifi-cant additive growth-inhibitory effects were also ob-tained ( fig. 4 c). Streptozocin in combination with sub-IC 50 ZM447439 statistically significantly reduced cell growth of QGP-1 by 43.4 versus 12.4% of streptozocin alone ( fig. 4 d). Applying octreotide alone for 3 days de-creased the growth of BON cells by up to 19.1% (2,000 n M ). When treating BON cells with a sub-IC 50 concentra-tion of ZM447439 plus octreotide (20–2,000 n M ), this re-sulted in additive growth-inhibitory effects of up to 47.7%
Aurora B
Actin
BON QGP-1 MIP-101
Incubation (days) Incubation (days)Incubation (days)
0
20
40
60
80
100
MIP-101
0 2 4
*
*
0 2 40
20
40
60
80
100
QGP-1
**
*
0 μM
0.1 μM
0.5 μM
1.0 μM
2.5 μM
5.0 μM
0 2 4
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
BON
*
a b c
Fig. 3. Growth inhibition of three different GEP-NET tumor cell lines with ZM447439. Cells from BON ( a ), MIP-101 ( b ) and QGP-1 ( c ) GEP-NETs were incubated on a 96-well plate (1 ! 10 4 ) with the novel aurora kinase inhibitor ZM447439 (0–5 � M) . Representative results of at least three independent experiments assessed by crystal violet staining are shown (means 8 SEM of 3 independent experiments). * p ! 0.05 vs. control.
Fig. 2. Expression of aurora kinase B in NET cell lines. Cell lysates were obtained from BON, QGP-1 and MIP-101 cells and subject-ed to Western blot analysis. The membranes were sequentially incubated with AIM-1 and actin antibodies. Western blot analysis detected high levels of aurora kinase B expression in GEP-NET cell lines. The WDET BON cell line showed a lower expression of aurora B kinase than QGP-1 (WDEC) and MIP-101 (PDEC).

Georgieva /Koychev /Wang /Holstein /Hopfenmüller /Zeitz /Grabowski
Neuroendocrinology 6
Doxorubicin b
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
120
** *
*
+ZM447439
(0.5 μM)
–5
–10
–50
+5
+10
+50 nM
Cisplatin c
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
120
**
**
* *+
ZM447439(0.5 μM)
–5
–10
–25
+5
+10
+25 μM
Streptozocin d
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
120
* * *
+ZM447439
(0.5 μM)
–2.5
–10
–25
+2.5
+10
+25 μM
Octreotidee
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
120
*
*
+ZM447439
(2 μM)
–20
–200
–2,000
+20
+200
+2,000 nM
a
Prol
ifera
tion
(% o
f con
trol
)
0
20
40
60
80
100
**
+ZM447439
(5 μM)
–5
–10
–25
+5
+10
+25 μM
5-FU
Fig. 4. Antiproliferative effects of ZM447439-based combination treatment with established chemo- and biotherapy assessed by crystal violet staining. MIP-101 cells were incubated with increas-ing concentrations of 5-FU ( a ) and cisplatin ( b ); QGP-1 cells were incubated with doxorubicin ( c ) and streptozocin ( d ), and BON cells with octreotide ( e ). Percent inhibition was plotted and the concentration of each compound that induced 25, 50 and 75% growth inhibition (IC 25 , IC 50 and IC 75 ) was calculated (data not shown). Combination treatments of either octreotide, cisplatin or streptozocin with a sub-IC 50 concentration of ZM447439 were performed, and cell numbers were determined after 3 days of in-cubation. Data are given as percentage of untreated controls, which were set at 100% (means 8 SEM of 5 independent experi-ments). * p ! 0.05 vs. control.

ZM447439 in Gastroenteropancreatic Neuroendocrine Tumors
Neuroendocrinology 7
compared with the treatment with ZM447439 alone ( fig. 4 e). Interestingly, ZM447439 and SOM230 had an antagonizing effect on each other, and SOM230 alone had a stronger effect on BON cell growth than the com-bination with ZM447439 (data not shown).
ZM447439 and Cell Cycle Regulation To test whether induction of cell cycle arrest contrib-
uted to the antiproliferative potency of ZM447439 in GEP-NET cancer cells, we performed flow-cytometric cell cycle analysis. After 48 h of incubation, ZM447439 dose-dependently arrested GEP-NET cells in the G 0 /G 1 and G 2 /M phase of the cell cycle, thereby decreasing the proportion of cells in the S phase ( fig. 5 ). Moreover, es-pecially in BON cells, an apoptosis-specific increase in the sub-G 1 peak was observed at higher concentra-tions.
Proapoptotic Effects of ZM447439 To determine how ZM447439 affects caspase-depen-
dent apoptosis levels in GEP-NET cells, we evaluated the activation of caspases 3 and 7, key enzymes in the apop-totic signaling cascade.
Again, cells were treated with increasing concentra-tions of ZM447439, and caspase 3 and 7 activities were determined after 24 and 48 h, respectively. In BON cells, ZM447439 (0–5 � M ) induced a 50% increase in caspase 3 and 7 activity after 24 h of incubation. However, in MIP-101 and QGP-1 cells, ZM447439-induced caspase 3 and 7 activity showed a comparable dose dependency but a less marked increase in enzyme activity ( fig. 6 ).
To further substantiate the antiapoptotic effect of ZM447439 in GEP-NETs, we determined DNA fragmen-tation as a biochemical hallmark of apoptosis. The effect was evaluated after 48 h of incubation with ZM447439
0
20
40
60
80
100
0 2 5 0 2 5 0 2 5
QGP-1 MIP-101 BON
Cel
l pro
por
tion
(%)
*
G2/MSG0/G1
**
*
*
*
*
*
*
*
**
Fig. 5. ZM447439 induced cell cycle arrest in GEP-NET cell lines. BON, QGP-1 and MIP-101 cells (1 ! 10 6 /ml) were plated in 24-well plates and cultured with ZM447439. After 48 h, cells were harvested and cell cycle distribution was measured by flow cytometry. Results represent one of the experiments performed three times. ZM447439 dose-dependently arrested GEP-NET cells in the G 0 /G 1 and G 2 /M phase of the cell cycle, thereby decreasing the proportion of cells in the S phase. Moreover, especially in BON cells an apop-tosis-specific increase in the sub-G 1 -peak was observed at higher concentrations. * p ! 0.05 vs. control.
40
80
120
160
Cas
pas
e 3/
7 ac
tivi
ty (%
of c
ontr
ol)
MIP-101 BON
05 μM
*
2.5 μM
**
2.5 μM
*
*
5 μM
*
*24 h48 h
Fig. 6. Proapoptotic, dose- and time-de-pendent effects of ZM447439. Caspase 3 activity was evaluated in BON and MIP-101 cells after 48 h of continuous exposure to ZM447439. Caspase 3 and 7 activation is shown as percentage of untreated con-trols (means 8 SEM of 5 independent ex-periments for each cell line). * p ! 0.05 vs. controls, which were set at 100%.

Georgieva /Koychev /Wang /Holstein /Hopfenmüller /Zeitz /Grabowski
Neuroendocrinology 8
and was more pronounced in MIP-101 and QGP-1 than in BON cells ( fig. 7 ).
Effects of ZM447439 on Aurora Kinase B and Survivin Expression As previously mentioned, the experimental aurora ki-
nase inhibitor ZM447439 produced a dose-dependent in-hibition of aurora kinase B function when used at sub-IC 50 concentrations. However, blocking the function of aurora kinase B led to its compensatory overexpression in MIP-101 cells, which was followed by survivin overex-pression in this cell line. In BON and QGP-1 cell lines, the aurora kinase B inhibition led to a dose-dependent aurora kinase B protein downregulation followed by sur-vivin downregulation in both cases ( fig. 8 ).
Discussion
Treatment possibilities of GEP-NETs are still not satis-factory and the prognosis of these patients is also not evi-dent with the current diagnostic tools. Thus, novel thera-peutic options are needed. Previous studies found that au-rora kinases A and B were aberrantly expressed in a variety of solid tumors, including prostate [11] , colon [12] , pancreas [13] , breast [14] and thyroid cancers [15] . In ad-dition, increased levels of aurora kinases correlated with advanced clinical stage in patients with prostate cancer, as well as in patients with head and neck squamous cell car-cinoma [11, 29] . Recently, it has been shown that hemato-logical malignant cells, including those from acute my-
eloid leukemia, acute lymphoblastic leukemia and chron-ic myeloid leukemia, aberrantly expressed aurora kinases A and B [30] . Since aurora kinases form a complex with the antiapoptotic protein survivin, which we have shown previously to be of prognostic significance in GEP-NET disease, we hypothesized that an aurora kinase inhibitor would be a feasible approach in these tumors. Here, we provide evidence that ZM447439 may be a promising anti-cancer agent for GEP-NET monotherapy as well as in combination with conventional cytotoxic agents.
ZM447439 inhibited GEP-NET cell growth in a time- and dose-dependent manner in all three cell lines, QGP-1, MIP-101 and BON, but induced low levels of caspase 3 and 7 activation and DNA fragmentation, the last being consistent with the results from previous studies [31] . Surprisingly, apoptotic effects were also observed in the well-differentiated neuroendocrine cell lines (BON and QGP-1), probably due to pathway signals leading to apop-tosis after induction of cell cycle arrest. The antiprolif-erative effects observed were due to cell cycle arrest in S phase and accumulation of cells with 4N DNA content. Sub-IC 50 concentrations of ZM447439 enhanced the an-tiproliferative effect of cisplatin, doxorubicin and strep-tozocin. The most pronounced effect was observed in the QGP-1 cell line where the addition of ZM447439 to 2.5 � M of streptozocin reduced cell growth almost 90%. With the addition of ZM447439 to doxorubicin, the lack of response to doxorubicin was offset. This is very inter-esting because QGP-1, a proposed cell model of advanced WDEC, is often resistant to aggressive protocols, includ-ing streptozocin and doxorubicin. The somatostatin ana-
M A B DC
41 kDa aurora kinase B
16 kDa survivin
42 kDa �-actin
0.522 –––
BON MIP-101 QGP-1
ZM447439 (μM)
Fig. 7. ZM447439-induced DNA ragmentation in GEP-NET cell lines. Representative tracings of three independent experiments are shown. A = Negative control; B–D = MIP-101 (B), BON (C) and QGP-1 (D) treated with 5 � M of ZM447439; M = molecular weight marker (100 bp).
Fig. 8. Effects of ZM447439 on the aurora kinase B and survivin expression. BON, MIP-101 and QGP-1 cells were treated with ris-ing concentrations of ZM447439 (0–2 � M ) for 48 h. Changes in the expression of aurora kinase B and survivin were analyzed by Western blotting. Representative results of three independent ex-periments are shown.

ZM447439 in Gastroenteropancreatic Neuroendocrine Tumors
Neuroendocrinology 9
logue octreotide, which has only poor antiproliferative effects as monotherapy, inhibited cell growth in BON cells when combined with ZM447439. Interestingly, SOM230, a new somatostatin analogue which engages five somatostatin receptors, did not exert such effects in the BON cell line (data not shown). As expected, cisplat-in had a strong growth-inhibiting effect on the MIP-101 cell line, serving as a model of PDECs in this study, but addition of sub-IC 50 concentrations of ZM447439 inhib-ited cell growth even more effectively. This might be help-ful in clinical practice, facilitating the reduction of cis-platin doses and consequently side effects, too. The main side effect of other aurora kinase inhibitors used in clini-cal trials is a transient myelodepression. This may be avoided by administering lower doses of both drugs. In-triguing and surprising was the antagonizing effect on cell growth in MIP-101 cells of combination treatment with ZM447439 and 5-FU, a drug commonly used in the treatment of colorectal adenocarcinomas and proposed for the treatment of PDECs. A possible explanation may be the fact that 5-FU is a pyrimidine analogue which in-hibits thymidylate synthase and thus RNA synthesis, but does not induce effects on DNA molecules, whereas ZM447439 induces a G 2 /M cell cycle block inhibiting the function of aurora kinases A and B, which participate in DNA repair. The DNA-intercalating agents streptozocin, cisplatin and especially doxorubicin [32, 33] induce G 2 /M-phase arrest, a phase during which cells are overex-pressing aurora kinases A and B in order to overcome the G 2 /M block; thus the addition of ZM447439 might ac-count for the profound effect on tumor cell growth.
Blocking the function of aurora kinase B using ZM447439 led to its overexpression in MIP-101. In BON and QGP-1 cells, aurora kinase B inhibition led to dose-dependent aurora kinase B and survivin downregulation in both cases. Intriguingly, the last two cell lines possess no p53 mutations (p53 mutation database by Thierry Soussi, version 2007). In MIP-101, however, p53 muta-tions were established in exon 7, codon 241 [34] . Appar-ently, the mechanism for dynamic regulation of survivin may be the modulation of aurora kinase B, which might be additionally modulated by the p53 status [4] , being one possible explanation for the upregulation of aurora ki-nase B in MIP-101 cells in the absence of survivin upreg-ulation. Another explanation may be a possible ‘on/off ’ mechanism of the interaction between survivin and au-rora kinase B. However, further studies are needed to confirm these findings.
Existing cell models for WDEC include the rat RIM-1 and the mouse STC-1 cell lines, but the results from these
cell lines are not transferable to human GEP-NETs and, moreover, there is no cell model for PDEC. We propose that both MIP-101 and LCC-18 could be used as a model for PDEC, but we preferred MIP-101 for its faster growth and simple growth conditions. QGP-1 can be used as a WDEC model.
In conclusion, our study provides the first evidence that the aurora kinase B inhibitor ZM447439 potently in-hibits the growth of human gastrointestinal NET cells. Combined with the overexpression of aurora B in solid tumors shown in previous studies and the fact that the compound specifically blocks aurora kinases at nano- and micromolar concentrations, this in vitro model is well suited for combination treatment approaches and our results warrant further studies on aurora kinase in-hibitors in the treatment of GEP-NET. The combination of sub-IC 50 concentrations of ZM447439 and DNA-inter-acting cytotoxic agents could be extremely valuable in clinical practice, thus increasing the benefits of therapy while decreasing adverse reactions to either agent.
Acknowledgments
We thank J. Stehr for excellent technical assistance and Dr. K. Maaser for the critical revision of the paper. Furthermore, we thank Novartis Pharma for providing us SOM 230. This study was supported by grants from the Cancer Society of Berlin (Ernst von Leyden Fellowship to I.G.), the DAAD (German Academic Ex-change Service) and the Sonnenfeld foundation (grant to I.G.).
References 1 Arnold R: Endocrine tumours of the gastro-intestinal tract. Introduction: definition, historical aspects, classification, staging, prognosis and therapeutic options. Best Pract Res Clin Gastroenterol 2005; 19: 491–505.
2 Klöppel G: Oberndorfer and his successors: from carcinoid to neuroendocrine carcino-ma. Endocr Pathol 2007; 18: 141–144.
3 Kulke MH: Clinical presentation and man-agement of carcinoid tumors. Hematol On-col Clin North Am 2007; 21: 433–455.
4 Bolanos-Garcia VM: Aurora kinases. Int J Biochem Cell Biol 2005; 37: 1572–1577.
5 Tanaka E, Hashimoto Y, Ito T, Kondo K, Hi-gashiyama M, Tsunoda S, Ortiz C, Sakai Y, Inazawa J, Shimada Y: The suppression of aurora-A/STK15/BTAK expression enhanc-es chemosensitivity to docetaxel in human esophageal squamous cell carcinoma. Clin Cancer Res 2007; 13: 1331–1340.
6 Bischoff JR, Plowman GD: The Aurora/Ipl1p kinase family: regulators of chromosome segregation and cytokinesis. Trends Cell Biol 1999; 9: 454–459.

Georgieva /Koychev /Wang /Holstein /Hopfenmüller /Zeitz /Grabowski
Neuroendocrinology 10
7 Anand S, Penrhyn-Lowe S, Venkitaraman AR: Aurora-A amplification overrides the mitotic spindle assembly checkpoint, induc-ing resistance to Taxol. Cancer Cell 2003; 3: 51–62.
8 Parra MT, Viera A, Gomez R, Page J, Car-mena M, Earnshaw WC, Rufas JS, Suja JA: Dynamic relocalization of the chromosomal passenger complex proteins inner centro-mere protein (INCENP) and aurora-B kinase during male mouse meiosis. J Cell Sci 2003; 116: 961–974.
9 Grabowski P, Griss S, Arnold CN, Hörsch D, Göke R, Arnold R, Heine B, Stein H, Zeitz M, Scherübl H: Nuclear survivin is a powerful novel prognostic marker in gastroentero-pancreatic neuroendocrine tumor disease. Neuroendocrinology 2005; 81: 1–9.
10 Yang J, Ikezoe T, Nishioka C, Tasaka T, Tani-guchi A, Kuwayama Y, Komatsu N, Bando-bashi K, Togitani K, Koeffler HP, Taguchi H, Yokoyama A: AZD1152, a novel and selective aurora B kinase inhibitor, induces growth ar-rest, apoptosis, and sensitization for tubulin depolymerizing agent or topoisomerase II inhibitor in human acute leukemia cells in vi-tro and in vivo. Blood 2007; 110: 2034–2040.
11 Chieffi P, Cozzolino L, Kisslinger A, Liber-tini S, Staibano S, Mansueto G, De Rosa G, Villacci A, Vitale M, Linardopoulos S, Por-tella G, Tramontano D: Aurora B expression directly correlates with prostate cancer ma-lignancy and influence prostate cell prolif-eration. Prostate 2006; 66: 326–333.
12 Ewart-Toland A, Dai Q, Gao YT, Nagase H, Dunlop MG, Farrington SM, Barnetson RA, Anton-Culver H, Peel D, Ziogas A, Lin D, Miao X, Sun T, Ostrander EA, Stanford JL, Langlois M, Chan JM, Yuan J, Harris CC, Bowman ED, Clayman GL, Lippman SM, Lee JJ, Zheng W, Balmain A: Aurora-A/STK15 T+91A is a general low penetrance cancer susceptibility gene: a meta-analysis of multiple cancer types. Carcinogenesis 2005; 26: 1368–1373.
13 Li D, Zhu J, Firozi PF, Abbruzzese JL, Evans DB, Cleary K, Friess H, Sen S: Overexpres-sion of oncogenic STK15/BTAK/aurora A ki-nase in human pancreatic cancer. Clin Can-cer Res 2003; 9: 991–997.
14 Tanaka T, Kimura M, Matsunaga K, Fukada D, Mori H, Okano Y: Centrosomal kinase AIK1 is overexpressed in invasive ductal car-cinoma of the breast. Cancer Res 1999; 59: 2041–2044.
15 Ulisse S, Delcros JG, Baldini E, Toller M, Curcio F, Giacomelli L, Prigent C, Ambesi-Impiombato FS, D’Armiento M, Arlot-Bonnemains Y: Expression of aurora kinases in human thyroid carcinoma cell lines and tissues. Int J Cancer 2006; 119: 275–282.
16 Mountzios G, Terpos E, Dimopoulos MA: Aurora kinases as targets for cancer therapy. Cancer Treat Rev 2008; 34: 175–182.
17 Andrews PD: Aurora kinases: shining lights on the therapeutic horizon? Oncogene 2005; 24: 5005–5015.
18 Carvajal RD, Tse A, Schwartz GK: Aurora ki-nases: new targets for cancer therapy. Clin Cancer Res 2006; 12: 6869–6875.
19 Girdler F, Gascoigne KE, Eyers PA, Hart-muth S, Crafter C, Foote KM, Keen NJ, Tay-lor SS: Validating Aurora B as an anti-cancer drug target. J Cell Sci 2006; 119: 3664–3675.
20 Li X, Sakashita G, Matsuzaki H, Sugimoto K, Kimura K, Hanaoka F, Taniguchi H, Furu-kawa K, Urano T: Direct association with in-ner centromere protein (INCENP) activates the novel chromosomal passenger protein, Aurora-C. J Biol Chem 2004; 279: 47201–47211.
21 Ditchfield C, Johnson VL, Tighe A, Ellston R, Haworth C, Johnson T, Mortlock A, Keen N, Taylor SS: Aurora B couples chromosome alignment with anaphase by targeting BubR1, Mad2, and Cenp-E to kinetochores. J Cell Biol 2003; 161: 267–280.
22 Beardmore VA, Ahonen LJ, Gorbsky GJ, Kal-lio MJ: Survivin dynamics increases at cen-tromeres during G2/M phase transition and is regulated by microtubule-attachment and Aurora B kinase activity. J Cell Sci 2004; 117: 4033–4042.
23 Walsby E, Walsh V, Pepper C, Burnett A, Mills K: Effects of the aurora kinase inhibi-tors AZD1152-HQPA and ZM447439 on growth arrest and polyploidy in acute my-eloid leukemia cell lines and primary blasts. Haematologica 2008; 93: 662–669.
24 Long ZJ, Xu J, Yan M, Zhang JG, Guan Z, Xu DZ, Wang XR, Yao J, Zheng FM, Chu GL, Cao JX, Zeng YX, Liu Q: ZM 447439 inhibi-tion of aurora kinase induces Hep2 cancer cell apoptosis in three-dimensional culture. Cell Cycle 2008; 7: 1473–1479.
25 Evers BM, Ishizuka J, Townsend CM Jr, Thompson JC: The human carcinoid cell line, BON. A model system for the study of carcinoid tumors. Ann NY Acad Sci 1994; 733: 393–406.
26 Iguchi H, Hayashi I, Kono A: A somato-statin-secreting cell line established from a human pancreatic islet cell carcinoma(somatostatinoma): release experiment and immunohistochemical study. Cancer Res 1990; 50: 3691–3693.
27 Niles RM, Wilhelm SA, Steele GD Jr, Burke B, Christensen T, Dexter D, O’Brien MJ, Thomas P, Zamcheck N: Isolation and char-acterization of an undifferentiated human colon carcinoma cell line (MIP-101). Cancer Invest 1987; 5: 545–552.
28 Gillies RJ, Didier N, Denton M: Determina-tion of cell number in monolayer cultures. Anal Biochem 1986; 159: 109–113.
29 Reiter R, Gais P, Jutting U, Steuer-Vogt MK, Pickhard A, Bink K, Rauser S, Lassmann S, Hofler H, Werner M, Walch A: Aurora ki-nase A messenger RNA overexpression is correlated with tumor progression and shortened survival in head and neck squa-mous cell carcinoma. Clin Cancer Res 2006; 12: 5136–5141.
30 Ikezoe T, Yang J, Nishioka C, Tasaka T, Tani-guchi A, Kuwayama Y, Komatsu N, Bando-bashi K, Togitani K, Koeffler HP, Taguchi H: A novel treatment strategy targeting Aurora kinases in acute myelogenous leukemia. Mol Cancer Ther 2007; 6: 1851–1857.
31 Kim KW, Mutter RW, Willey CD, Subha-wong TK, Shinohara ET, Albert JM, Ling G, Cao C, Gi YJ, Lu B: Inhibition of survivin and aurora B kinase sensitizes mesothelioma cells by enhancing mitotic arrests. Int J Ra-diat Oncol Biol Phys 2007; 67: 1519–1525.
32 Minotti G, Menna P, Salvatorelli E, Cairo G, Gianni L: Anthracyclines: molecular ad-vances and pharmacologic developments in antitumor activity and cardiotoxicity. Phar-macol Rev 2004; 56: 185–229.
33 Rabik CA, Dolan ME: Molecular mecha-nisms of resistance and toxicity associated with platinating agents. Cancer Treat Rev 2007; 33: 9–23.
34 Kastrinakis WV, Ramchurren N, Rieger KM, Hess DT, Loda M, Steele G, Summer-hayes IC: Increased incidence of p53 muta-tions is associated with hepatic metastasis in colorectal neoplastic progression. Oncogene 1995; 11: 647–652.
35 Fogh J, Orfeo T, Tiso J, Sharkey FE: Estab-lishment of human colon carcinoma lines in nude mice. Exp Cell Biol 1979; 47: 136–144.
36 Evers BM, Townsend CM Jr, Upp JR, Allen E, Hurlbut SC, Kim SW, Rajaraman S, Singh P, Reubi JC, Thompson JC: Establishment and characterization of a human carcinoid in nude mice and effect of various agents on tu-mor growth. Gastroenterology 1991; 101: 303–311.
37 Jumarie C, Malo C: Caco-2 cells cultured in serum-free medium as a model for the study of enterocytic differentiation in vitro. J Cell Physiol 1991; 149: 24–33.
38 Bjork P, Jonsson U, Svedberg H, Larsson K, Lind P, Dillner J, Hedlund G, Dohlsten M, Kalland T: Isolation, partial characteriza-tion, and molecular cloning of a humancolon adenocarcinoma cell-surface glyco-protein recognized by the C215 mouse monoclonal antibody. J Biol Chem 1993; 268: 24232–24241.
39 Quinn LA, Moore GE, Morgan RT, Woods LK: Cell lines from human colon carcinoma with unusual cell products, double minutes, and homogeneously staining regions. Can-cer Res 1979; 39: 4914–4924.
40 Lundqvist M, Mark J, Funa K, Heldin NE, Morstyn G, Wedell B, Layton J, Oberg K: Characterisation of a cell line (LCC-18) from a cultured human neuroendocrine-differen-tiated colonic carcinoma. Eur J Cancer 1991; 27: 1663–1668.
41 Wolf B, Thompson J, Von Kleist S: Ultra-structural localization of carcino-embryon-ic antigen in a continuous human tumour cell line (LS 174 T) in relation to morphomet-ric investigations. Anticancer Res 1984; 4: 213–219.
42 Park JG, Oie HK, Sugarbaker PH, Henslee JG, Chen TR, Johnson BE, Gazdar A: Char-acteristics of cell lines established from hu-man colorectal carcinoma. Cancer Res 1987; 47: 6710–6718.




















