
Predictors of short-term outcome in brain-injured patients with disorders of consciousness
Upload
independentCategory
view
2download
0
Proapoptotic Fas Ligand Is Expressed by Normal KidneyTubular Epithelium and Injured Glomeruli
CORINA LORZ, ALBERTO ORTIZ, PILAR JUSTO, SILVIA GONZALEZ-CUADRADO,NATALIA DUQUE, CARMEN GOMEZ-GUERRERO, and JESUS EGIDODivision of Nephrology, Fundacio´n Jimenez Dı´az, Universidad Autonoma de Madrid, Spain.
Abstract.Fas ligand (FasL) is a cell membrane cytokine thatcan promote apoptosis through activation of Fas receptors. Fasreceptor activation induces glomerular cell apoptosisin vivoand participates in tubular cell death during acute renal failure.However, there is little information on the expression of FasLin the kidney. This study reports that FasL mRNA and proteinare present in normal mouse and rat kidney.In situ hybridiza-tion and immunohistochemistry showed that proximal tubularepithelium is the main site of FasL expression in the normalkidney. In addition, increased total kidney FasL mRNA anddenovo FasL protein expression by glomerular cells were ob-served in two different models of glomerular injury: rat im-mune-complex proliferative glomerulonephritis and murine lu-pus nephritis. Both full-length and soluble FasL were increasedin the kidneys of mice with nephritis. Cultured murine proxi-mal tubular epithelial MCT cells and primary cultures of mu-
rine tubular epithelial cells expressed FasL mRNA and protein.Tubular epithelium-derived FasL induced apoptosis in Fas-sensitive lymphoid cell lines but not in Fas-resistant lymphoidcell lines. By contrast, MCT cells grown in the presence of thesurvival factors of serum were resistant to FasL, and onlybecame partially sensitive to apoptosis induced by high con-centrations (100 ng/ml) of FasL upon serum deprivation. How-ever, MCT cells stimulated with inflammatory mediators (tu-mor necrosis factor-a, interferon-g, and lipopolysaccharide)increased cell surface Fas expression and were sensitized toapoptosis induced by FasL (FasL 556 5% versuscontrol8.3 6 4.1% apoptotic cells at 24 h,P , 0.05). Cytokine-primed primary cultures of tubular epithelial cells also acquiredsensitivity to FasL-induced apoptosis. These results suggestthat FasL expression by intrinsic renal cells may play a role incell homeostasis in the normal kidney and during renal injury.
Fas ligand (FasL) is an extensively glycosylated, 36- to 40-kDtype II membrane protein whose main known function is toinduce apoptosis through cross-linking of the death-inducing re-ceptor Fas (1). Proteolytic cleavage of FasL yields a 26- to 29-kDsoluble form of the cytokine (1). FasL and Fas regulate theimmune response and have been implicated in peripheral deletionof autoimmune cells, activation-induced T cell death, and one ofthe two major cytolytic pathways mediated by CD81 cytolytic Tcells (1). In addition, Fas-induced apoptosis promotes parenchy-mal cell damage in glomerular injury, acute renal failure, liverdisease, and thyroiditis (2–6). FasL expression was originallythought to be restricted to activated T cells and natural killer cells(1). However, the earliest reports on this cytokine described thepresence of transcripts in several nonlymphoid organs, includingthe testis, small intestine, lung, and kidney (7). More recently,FasL transcripts have also been found in the eye, thymus, spleen,seminal vesicle, prostate, and uterus of adult mice, as well as inhuman tumor cells (8–10). The regulation of FasL-induced apo-ptosis is complex, and some cell types express FasL but are unableto promote apoptosis (11). Eye and testicular cells were shown to
express an active form of this cytokine that may have a role inimmune privilege in these organs (12,13). By contrast, there islittle information on the cells that express FasL in the kidney.Moreover, the biologic activity of FasL in renal cells has not beencompletely characterized. Agonistic anti-Fas antibodies kill mu-rine mesangial cells and renal fibroblasts (4,5,14), whereas cul-tured tubular epithelial renal cells have been reported to be lesssensitive to Fas-induced apoptosis (15,16).
Most studies on the biologic activity of FasL have relied on theusage of activating anti-Fas antibodies. It has recently becomeevident that the complex regulation of FasL-Fas apoptosis in-volves, among other factors, different biologic activities for sol-uble and membrane-bound FasL (17). Both soluble FasL andagonistic anti-Fas antibodies are less effective than membrane-bound FasL in inducing apoptosis (17). Thus, cells such as freshlyisolated human peripheral blood T lymphocytes, previouslythought to be resistant to FasL-induced cell death, have now beenfound to be sensitive to membrane-bound FasL (17).
We have now studied the site of FasL expression in normaland injured kidneysin vivo, the ability of renal tubular cells toexpress an active form of FasL, and the sensitivity of thesecells to recombinant FasL-induced apoptosis.
Materials and MethodsCells, Cytokines, and Antibodies
MCT murine proximal tubular epithelial cells were a generous giftfrom Eric G. Neilson (University of Pennsylvania, Philadelphia, PA)(18). They were cultured in RPMI 1640 (Life Technologies, Grand
Received March 23, 1999. Accepted October 23, 1999.Correspondence to Dr. Alberto Ortiz, Unidad de Dia´lisis, Fundacio´n JimenezDıaz, Avda Reyes Cato´licos 2, 28040 Madrid, Spain. Fax:134 915 494 764;E-mail: [email protected]
1046-6673/1106-1266Journal of the American Society of NephrologyCopyright © 2000 by the American Society of Nephrology
J Am Soc Nephrol 11: 1266–1277, 2000

Island, NY), 10% decomplemented fetal calf serum (FCS), 2 mMglutamine, 100 U/ml penicillin, and 100mg/ml streptomycin at 37°Cin 5% CO2. In some studies, cells were deprived of serum (0% FCS)for 24 to 48 h. Primary cultures of murine tubular epithelial cells wereperformed according to previously published techniques (19).
Fas-sensitive A20 and Fas-resistant A20R murine B lymphomacells were a kind gift from Ju¨rg Tschopp (Lausanne University,Switzerland) (9). L1210 (Fas-negative) and L1210Fas (Fas-trans-fected) murine leukemia cells were a kind gift from Alberto Anel(Universidad de Zaragoza, Spain) (20).
Polyclonal rabbit anti-FasL antibodies raised against a peptidecorresponding to amino acids 2 to 19 mapping at the amino terminusof FasL (13), and the control peptide were purchased from Santa CruzBiotechnology (Santa Cruz, CA). PE62 polyclonal rabbit anti-FasLantibody was a gift from J. Tschopp (9). The rat IgM monoclonal A11that binds to the FasL extracellular peptide 196-220 was from Alexis(Laufelfingen, Switzerland) (9), control rat IgM and biotin-anti-ratIgM was from ImmunoKontact (Frankfurt, Germany), and streptavi-din phycoerythrin was from Calbiochem (San Diego, CA). Neutral-izing anti-FasL antibodies (clone MFL3; Pharmingen, San Diego,CA) were used at a concentration of 10mg/ml (21). All of theseantibodies bind to murine FasL (9,13), and neutralizing MFL3 anti-bodies inhibit the cytotoxicity of FasL from all strains of mice tested,including SJL mice (21).
Recombinant human FasL (Alexis) was used in the presence of a10-fold excess of a cross-linking antibody, which by itself was devoidof lethal activity. Cross-linking of FasL restores the biologic activityof soluble FasL and simulates its presence on the cell membrane (22).Human FasL activates the murine Fas receptor (22).
Models of Renal InjuryInbred female Wistar rats and albino Swiss mice were obtained
from the Fundacio´n Jimenez Dıaz animal facilities. Studies wereconducted in accord with the National Institutes of Health Guide forthe Care and Use of Laboratory Animals. Immune complex-mediatedproliferative glomerulonephritis was induced in rats that were previ-ously immunized with ovalbumin (Sigma, St. Louis, MO), by dailyintraperitoneal administration of 10 mg of ovalbumin (23). Samplesfrom rats with nephritis (n 5 3) were obtained when glomerular injurywas fully developed (proteinuria.100 mg/d).
C57BL/6 (B6) and Balb/c mice were obtained from Iffa-CREDO(Barcelona, Spain). Four-week-old MRL-lpr/lpr mice without nephri-tis (n 5 3) and 4-mo-old MRL-lpr/lpr mice with nephritis (n 5 3)were a gift from Carlos Martinez-A (Centro Nacional de Biotecnolo-gıa, Madrid, Spain). Kidney samples were snap-frozen for RNA andprotein studies or fixed in 10% buffered formaldehyde and embeddedin paraffin for histologic studies.
Studies of Cell DeathTubular epithelial cells were cultured in 12-well plates. Cells were
cultured in serum-free media in the presence or absence of 30 ng/mlmurine tumor necrosis factor-a (TNF-a) (Immugenex, Los Angeles,CA), 300 U/ml human interferon-g (IFN-g) (Immugenex), and 10mg/ml bacterial lipopolysaccharide (LPS) (Sigma) for 48 h, with FasLadded for the last 24 h. When specified, some cells were also culturedin the presence of 10% FCS. Hypodiploid apoptotic cells were quan-tified by flow cytometry of permeabilized, propidium iodide-stainedcells, as described previously (14). For morphologic assessment ofapoptosis, cells were cultured in chamber slides, fixed with methanol:acetone (1:1), and stained with propidium iodide in the presence ofRNase A (24).
Reverse Transcription-PCR and NorthernHybridization
Kidney total RNA was isolated using the acid guanidinium-phenolchloroform method (25), and 40mg was separated in 1% agarose gelscontaining 2.3% formaldehyde. MCT poly(A) RNA was isolatedusing the FASTRACK mRNA kit (Invitrogen, San Diego, CA) and 5mg was loaded. RNA was transferred to nylon membranes (Gene-screen, New England Nuclear, Boston, MA) and prehybridized andhybridized as described previously (16).
Reverse transcription (RT)-PCR for mouse FasL detection wascarried out as described previously (16), using the following primers:59-CACAAATCTGTGGCTACCG, 39-GCCCATATCTGTCCAG-TAG. The resulting PCR product was cloned and sequenced to con-firm its identity. It expands several exons, thus excluding the possi-bility of amplification of genomic DNA. The following primers wereused for amplification of rat FasL: 59-CATAGAGCTGTGGCTACC,39-ATGGTCAGCAACGGTAAC. The PCR product had 534 bp.Semiquantitative PCR was performed in rat and mouse kidney sam-ples according to published techniques (26). FasL and G3PDH wereamplified in the presence of32P-dCTP (Amersham). The PCR prod-ucts were separated in 4% polyacrylamide/urea gels, and bands weremeasured by densitometry and corrected for G3PDH. The optimalnumber of amplification cycles was chosen on the basis of experi-ments that established the exponential range of the reaction.
The murine FasL probe has been described previously (16,27).Sense and antisense RNA probes forin situhybridization were labeledwith digoxigenin using a commercially available kit (DIG RNAlabeling kit; Boehringer Mannheim, Mannheim, Germany), followingthe manufacturer’s instructions.
Flow Cytometry Analysis of FasL and Fas ExpressionTo study cell surface, FasL cells were cultured for 3 h in the
presence of 5 mM ethylenediaminetetra-acetic acid (EDTA), whichinhibits metalloproteases and decreases the release of FasL from thecell membrane, thus increasing the availability of this cytokine at thecell surface (28). Cells were washed with phosphate-buffered saline(PBS) and resuspended in 0.2% bovine serum albumin/PBS. A total of5 3 105 cells was incubated with 20mg/ml monoclonal A11 anti-FasLantibody or control rat IgM in 5% FCS, 0.2% bovine serum albumin,in PBS for 30 min at 4°C, followed by incubation with 1:100 biotin-anti-rat IgM for 30 min at 4°C and by incubation with 1:50 strepta-vidin phycoerythrin. Cells were analyzed on a cytofluorograph, anddebris was excluded from analysis by selective gating based onanterior and right angle scatter. At least 10,000 events were collectedfor each sample, and data were displayed on a logarithmic scale ofincreasing fluorescence intensity (14). Mean cell fluorescence wascalculated using LYSIS II software.
To study cell surface Fas expression, cells were cultured in thepresence of control medium or cytokines for 48 h and stained withJo-2 anti-murine Fas antibody or control hamster IgG (Pharmingen),as described (14).
ImmunohistochemistryImmunohistochemistry was carried out as described previously
(29) in paraffin-embedded tissue sections 5mm thick. Primary anti-bodies were 20mg/ml A11 rat monoclonal anti-FasL (Alexis) or 1mg/ml rabbit polyclonal anti-FasL (Santa Cruz) (13). Sections werecounterstained with Carazzi’s hematoxylin. Some sections were sub-sequently incubated with the proximal tubule marker, fluorescein-conjugated,Tetranogolobus lotus(diluted 1:33) (Sigma) or the col-lecting tubule marker, fluorescein-conjugated,Dolichos biflorum
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1267

(diluted 1:100) (Sigma) (30). These sections were mounted in 90%glycerol/PBS and photographed immediately. Negative controls in-cluded incubation with a nonspecific Ig of the same isotype as theprimary antibody in both cases, and competition of the primaryantibody with a 10-fold excess of the immunogenic peptide in the caseof the rabbit polyclonal. Positive controls included testis (7) and eye(13).
In Situ HybridizationParaffin-embedded tissue sections, 5mm thick, were fixed in 1.5%
paraformaldehyde-1.5% glutaraldehyde for 10 min, treated with 5mM levamisole for 30 min, and deproteinized with 0.2N HCl for 20min at room temperature followed by digestion with 25mg/ml pro-teinase K in 0.1 M Tris, 0.005 M EDTA, 0.5% sodium dodecyl sulfatefor 30 min at 37°C. They were subsequently hybridized with 1 ng/mldenatured digoxigenin-11-UTP-labeled FasL riboprobe in hybridiza-tion solution (23 SSC, 13 Denhardt’s, 0.1 M sodium phosphate, pH6.5, 10% dextran sulfate, 40% deionized formamide, 24 mM vanadylribonuclease complex, and 0.5mg/ml yeast tRNA) at 42°C overnight,under sealed coverslips, in a humidified chamber. Slides were washedin 23 SSC for 5 min and 0.23 SSC for 3 min, and incubated with1:750 alkaline phosphatase-conjugated anti-digoxigenin antibody(Boehringer Mannheim) for 60 min at 37°C. Colorimetric detection ofmRNA was performed with nitroblue tetrazolium and 5-bromo-4-chloro-3-indolylphosphate (X-phosphate) in the dark for 10 to 30 min.Negative controls consisted of matched serial sections hybridizedwithout RNA probe or with sense probe or pretreated with 25mg/mlRNase A for 1 h (Sigma) before hybridization with the correspondingantisense probe. Testis samples were included as positive controls.
Western BlotTissue and cell samples were homogenized in lysis buffer (50 mM
Tris-HCl, 150 mM NaCl, 2 mM EDTA, 2 mM ethyleneglycol-bis(b-aminoethyl ether)-N,N9-tetra-acetic acid, 0.2% Triton X-100, 0,3%Nonidet-P40, 0.1 mM phenylmethylsulfonyl fluoride, and 1mg/mlpepstatin A) and then separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis under reducing conditions. Afterelectrophoresis, samples were transferred to polyvinylidene difluoridemembranes. The membranes were blocked with 5% skim milk in
PBS/0.5% vol/vol Tween 20 for 1 h, washed with PBS/Tween, andincubated with 40 ng/ml PE 62, a polyclonal anti-FasL antibody orpolyclonal anti-murine Fas (1:500) (Santa Cruz), in 5% milk PBS/Tween for 18 h at 4°C. The blots were washed with PBS/Tween andsubsequently incubated with horseradish peroxidase-conjugated anti-rabbit IgG (1:2500; Amersham). After washing with PBS/Tween, theblots were developed with the chemiluminescence method accordingto the manufacturer’s instructions (Amersham). FasL (a gift from J.Tschopp)-transfected 293 cell lysates were diluted 10-fold and used aspositive controls. As a negative control, the antibodies were preincu-bated with a 10-fold excess of the immunogenic peptide beforeWestern blotting. Similar results were obtained with a polyclonalanti-FasL antibody from Santa Cruz. Tubulin was used as loadingcontrol.
Cytosolic and membrane proteins were obtained by lysing the cellsin 2% Triton X-114 precondensed in PBS. After phase separation at37°C, the lower phase enriched in detergent contains the membraneproteins, and the upper aqueous phase contains the cytosolic proteins.Both phases were separated, precipitated with methanol/chloroform,and resuspended in sample buffer (22).
FasL BioactivityFasL killing activity was assessed by incubating 13 105 to 13 102
effector MCT cells, 6 h after seeding, with 13 105 bromodeoxyuri-dine-labeled target A20R and A20 cells (9) or L1210 and L1210Fascells (20) in 48-well plates. Higher effector:target (E:T) ratios wereused in preliminary experiments, but were abandoned because tubularcells became overconfluent at E:T ratios.1. The release of bromode-oxyuridine was then determined after a 14-h coincubation using acellular DNA fragmentation enzyme-linked immunosorbent assay(Boehringer Mannheim). As a positive control, MCT cells werereplaced by 10 ng/ml recombinant FasL. For the calculation of thepercentage of dead cells, cell death induced in Fas-sensitive cells byrecombinant FasL was considered to be 100%. Flow cytometry con-firmed more than 90% apoptosis among A20-Fas-sensitive cellstreated with FasL. Results are expressed as mean6 SEM of inde-pendent experiments each consisting of quadruplicate wells.
Figure 1.Fas ligand (FasL) expression in normal rodent kidney. (A) Reverse transcription (RT)-PCR. MWM, molecular weight markers. (B)Northern blot. Mouse kidney. Total RNA. (C) Western blot. Mouse kidney. Competition with the immunogenic peptide (control peptide)prevents binding of the antibody.
1268 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1266–1277, 2000

Statistical AnalysesResults are expressed as mean6 SEM. Significance at the 95%
level was established using ANOVA andt test. Bonferroni correctionfor multiple comparisons was used.
ResultsProximal Tubular Epithelium Is the Main Site of FasLExpression in Normal Murine Kidneys
FasL transcripts were detected by RT-PCR in normal mouseand rat kidney (Figure 1). The presence of a FasL transcriptwas confirmed by Northern blot in normal murine kidney(Figure 1). An approximately 36- to 40-kD FasL protein,corresponding to full-length transmembrane FasL, was themore abundant FasL form in normal mouse kidney (Figure 1).To determine whether intrinsic renal cells account for FasLexpression in the normal kidney, the site of FasL expression
was explored. FasL transcripts were detected in mouse tubularepithelium byin situ hybridization (Figure 2). Immunohisto-chemistry using two different anti-FasL antibodies confirmedthe expression of FasL protein by normal tubuli (Figure 3).Proximal tubules, identified by their ability to bind the proxi-mal tubule markerTetranogolobus lotuslectin (30), were themain source of FasL, while collecting ducts were negative. Bycontrast, FasL was absent from normal glomerular mesangialcells and vessels (Figures 2 and 3). Tubular expression of FasLwas noted in the different strains of mice tested (Swiss, Balb/c,B6, MRL lpr/lpr).
Increased Renal FasL and de Novo Glomerular FasLExpression during Glomerular Inflammation
We then addressed the question of whether FasL expressionchanged in the course of renal inflammation in rodent models.FasL mRNA, as assessed by semiquantitative RT-PCR, wasincreased in whole kidneys from rats with immune-complexproliferative glomerulonephritis and mice with lupus nephritis(Figure 4). In the course of murine lupus nephritis, both full-length FasL and an approximately 26-kD molecule correspond-ing to the size of soluble FasL were increased (Figure 4E).
The pattern of FasL expression also changed during glomer-ular and tubulointerstitial inflammation.De novoFasL expres-sion was noted in the glomerular mesangium, and FasL immu-noreactivity was also prominent in tubular cells and amonginfiltrating interstitial cells in MRL lpr/lpr mice with lupusproliferative glomerulonephritis and tubulointerstitial nephritis(Figure 5).
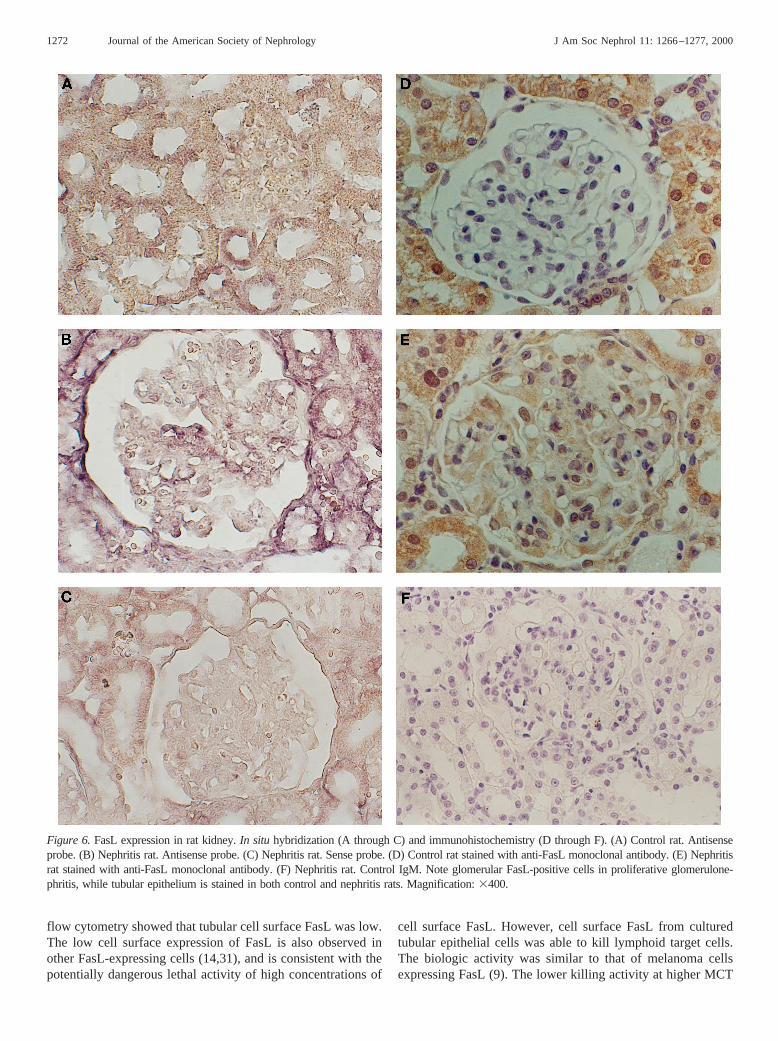
FasL mRNA and protein are also limited to tubular epithelialcells in normal rat kidney (Figure 6). As was the case duringmouse renal inflammation, in rats with immune complex pro-liferative glomerulonephritis, FasL mRNA and protein wereexpressedde novoby glomerular cells (Figure 6).
Figure 2. FasL mRNA expression in normal mouse kidney.In situhybridization. (A) Antisense probe. Note FasL expression in renaltubules, but its absence from glomeruli. (B) Sense probe. Magnifica-tion: 3200.
Figure 3.Localization of the tubular segment expressing FasL. Nor-mal mouse kidney (Balb/c) stained with both anti-FasL antibody (A)and the proximal tubule markerTetranogolobus lotus(B), or bothanti-FasL antibody (C) and the collecting tubule markerDolichosbiflorum (D). Magnification:3200. Arrows localize the same tubuliin A and B or C and D. Proximal tubules are the main source of renalFasL.
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1269

Murine Cultured Tubular Epithelial Cells ExpressFasL
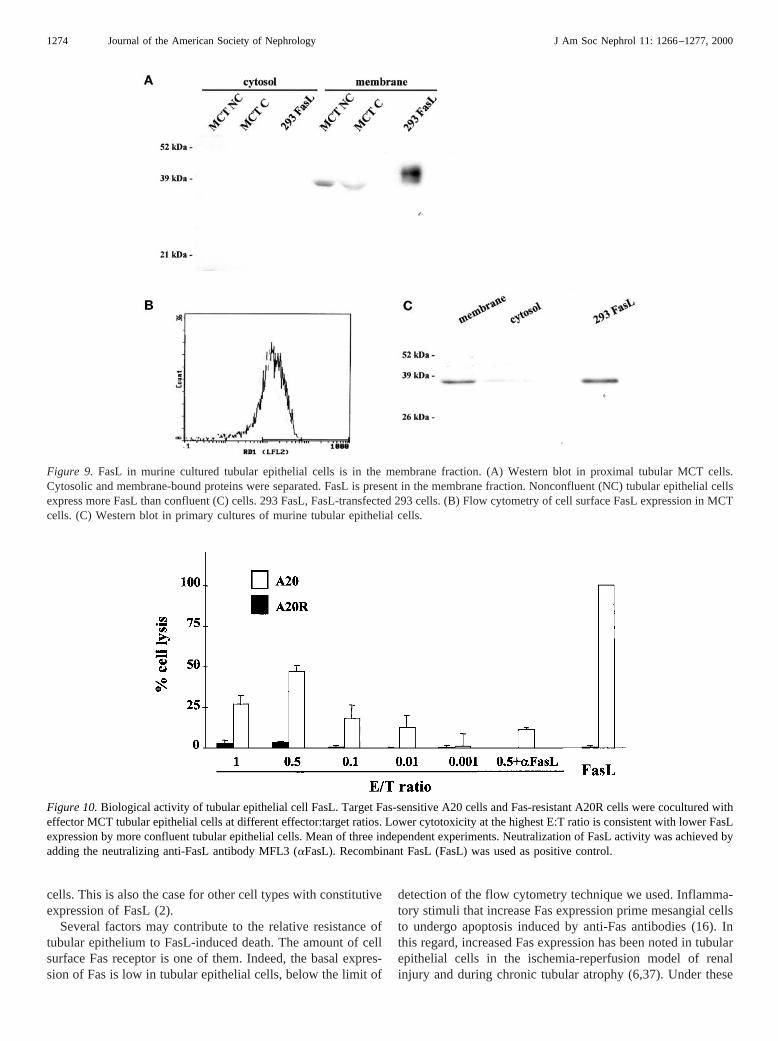
Proximal tubular epithelial cells were the main source ofrenal FasLin vivo. We then studied FasL expression in cul-tured tubular epithelial cells. FasL mRNA was detected incultured murine proximal tubular epithelial MCT cells byRT-PCR, Northern blot, andin situ hybridization (Figures 7and 8). RT-PCR also identified a FasL transcript in primarycultures of tubular epithelial cells (Figure 7B).In situ hybrid-ization showed that FasL mRNA expression was higher inMCT cells grown at low cell density than when cells wereconfluent (Figure 8). Western blot confirmed the presence of a36- to 40-kD FasL protein in these cells, which, after TritonX-114 extraction, appeared in the membrane fraction (Figure9A), as corresponds to the full-length transmembrane form ofFasL. As was the case for mRNA, nonconfluent cells expressedmore FasL protein than confluent cells (Figure 9A), indicatinga regulatory process that deserves further study. Flow cytom-etry showed that low levels of FasL protein are present in thecell surface, where they can be biologically active (mean cellfluorescence 30% higher than control isotype antibody-stainedcells) (Figure 9B). The low amount of FasL in the cell surfaceis consistent with data from other cell types (14,31), wheremost FasL is bound to intracellular membranes (31).
A 36- to 40-kD FasL protein was also present in the mem-brane fraction of primary cultures of mouse tubular epithelialcells (Figure 9C).
Tubular Epithelial Cell FasL Is Biologically ActiveNot all FasL-expressing cells are able to promote apoptosis
(11). Thus, we tested the ability of tubular epithelial MCT cell
FasL to promote apoptosis.In vitro assays using two differentsets of Fas-sensitive and Fas-resistant cells (A20 and A20R,and L1210Fas and L1210) demonstrated that tubular epithelialcell-derived FasL is able to induce cell death in lymphoid cells.A20R cells were generated by continuous culture in the pres-ence of FasL, and have downregulated Fas receptor expression(22). By contrast, L1210Fas were made sensitive to FasLapoptosis by transfecting them with a Fas expression vector(20). Results obtained in A20/A20R cells are shown in Figure10. An increased rate of cell death was noted among Fas-sensitive A20 cells cocultured with tubular epithelial cells,which was not present among Fas-resistant A20R cells underthe same culture conditions. The apoptotic nature of death wasconfirmed by morphologic evaluation of the cells. Similarresults were obtained with Fas-sensitive L1210Fas and Fas-resistant L1210 cells (not shown). Apoptosis induced by tubu-lar epithelial cells in A20 cells was reduced by 73% whenneutralizing anti-FasL antibodies were added to the coculture(Figure 10).
FasL Induces Apoptosis in Activated Tubular EpithelialCells
Tubular epithelial cells have been reported to be relativelyresistant to Fas-induced death (15,16). However, Fas defi-ciency protects tubular epithelial cells from apoptosis in thecourse of renal ischemia-reperfusion (6). This suggests thatsensitivity to FasL in tubular epithelial cells is a regulatedprocess. It has been observed that the survival factors presentin serum protect tubular epithelial cells from the lethal effect ofTNF-a (32). Indeed, tubular epithelial MCT cells were com-
Figure 4. Increased renal FasL during renal inflammation. (A) RT-PCR. Rat kidney. C, healthy control; N, rat with nephritis. Representativeof two experiments. (B) Mean densitometry values of FasL corrected for G3PDH of two gels similar to the one shown in A. (C) RT-PCR.Mouse kidney. C, healthy control; N, murine lupus nephritis. (D) Mean densitometry values of FasL corrected for G3PDH of two gels similarto the one shown in C. (E) Western blot. Kidneys from control (C) and lupus nephritis mice (N).
1270 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1266–1277, 2000

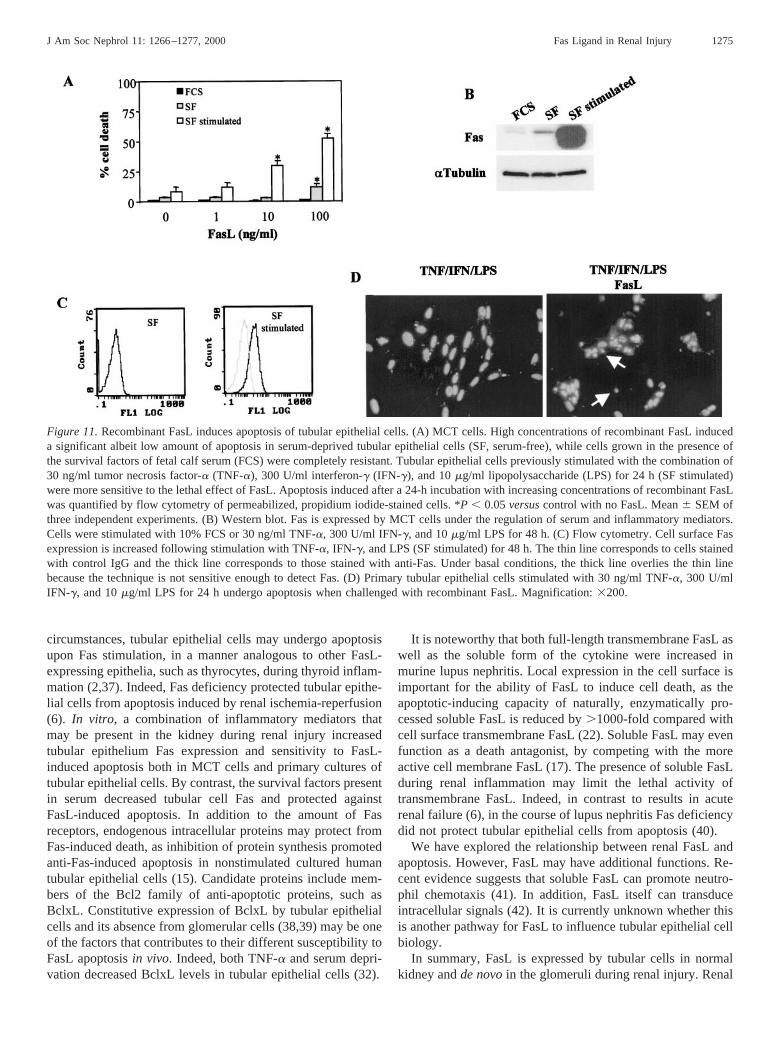
pletely resistant to FasL-induced apoptosis when grown in thepresence of serum (Figure 11A). In addition, nonstimulatedMCT cells grown in serum-free media were still relativelyresistant to FasL-induced apoptosis. Only the highest concen-tration of FasL induced a significant, albeit low, amount ofapoptosis (Figure 11A) (100 ng/ml FasL 12.86 2.4% versuscontrol 3.26 1.0% apoptotic cells at 24 h,P , 0.05). Underthese conditions, MCT cells were at least 100-fold more resis-tant to recombinant FasL than the Fas-sensitive lymphoid celllines used in our studies, as 1 ng/ml FasL already inducedsignificant apoptosis in these lymphoid cell lines. As expectedfrom these data, neutralizing anti-FasL antibodies did not mod-ify spontaneous apoptosis in tubular epithelial cells after 24 or48 h in culture in serum-free media (data not shown). However,during renal injury tubular epithelial cells are exposed to in-flammatory mediators. Thus, we studied their sensitivity toFasL in the presence of a cocktail of inflammatory mediators(30 ng/ml TNF-a, 300 U/ml IFN-g, and 10mg/ml bacterialLPS). Upon activation by inflammatory mediators, the sensi-tivity of MCT cells to apoptosis induced by FasL was in-creased: 10 ng/ml FasL already increased the rate of apoptosis,and the death rate of cells exposed to 100 ng/ml FasL was6.6-fold that of control cells (Figure 11A). The occurrence ofapoptosis was confirmed by morphologic criteria (fragmented,condensed, and pyknotic nuclei) of fixed, propidium iodide-stained cells (not shown). MCT cells activated by these inflam-matory mediators have an upregulated expression of Fas re-ceptor (Figure 11, B and C), which can contribute to theirincreased susceptibility to FasL-mediated apoptosis. Serumdeprivation also increased Fas expression (Figure 11B), as itdoes in other renal cells (14).
We then checked the sensitivity of primary cultures oftubular epithelial cells to FasL-induced apoptosis. Tubularcells were resistant to FasL under basal conditions; however,they were sensitized by prestimulation with 30 ng/ml TNF-a,300 U/ml IFN-g, and 10mg/ml bacterial LPS (Figure 11D).Similar results were obtained with freshly isolated mouse tu-bules (not shown).
DiscussionThis article shows that murine tubular epithelial cells are the
main renal source of local FasL in the normal kidney. Addi-tional sources of FasL, such as mesangial cells and infiltratingleukocytes, may be present in the glomeruli and interstitialspace during renal injury. Renal FasL may potentially regulatethe immune response, promote parenchymal cell death, and/orlimit inflammatory cell and fibroblast number by promotingapoptosis in leukocytes and renal fibroblasts (1,14).
FasL-induced apoptosis is a complex phenomenon. Not allFasL-expressing cells can kill FasL-sensitive cells (11,33). Inleukocytes, FasL is stored in the membranes of intracellulargranules and only becomes exposed in the cell surface, whereit has killing activity, upon cell activation (31,33,34). Culturedtubular epithelial cells contain full-length FasL in cell mem-branes, and the amount of FasL is subject to regulation by celldensity. Although additional studies are required to character-ize the regulation of FasL expression in tubular epithelium,
Figure 5. De novoglomerular FasL protein expression in mousekidney. Immunohistochemistry. (A) Normal mouse kidney from4-wk-old MRL lpr/lpr stained with anti-FasL monoclonal antibody.(B) Kidney from 4-mo-old MRL-lpr mouse with nephritis stainedwith anti-FasL monoclonal antibody. (C) Mouse nephritis kidneystained with control IgM antibody. Note tubular FasL staining withnegative glomeruli in normal kidney, and the presence of FasL-positive cells in glomerulus, tubuli, and interstitium during nephritis.Magnification:3400 in A and B;3100 in C.
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1271
flow cytometry showed that tubular cell surface FasL was low.The low cell surface expression of FasL is also observed inother FasL-expressing cells (14,31), and is consistent with thepotentially dangerous lethal activity of high concentrations of
cell surface FasL. However, cell surface FasL from culturedtubular epithelial cells was able to kill lymphoid target cells.The biologic activity was similar to that of melanoma cellsexpressing FasL (9). The lower killing activity at higher MCT
Figure 6.FasL expression in rat kidney.In situ hybridization (A through C) and immunohistochemistry (D through F). (A) Control rat. Antisenseprobe. (B) Nephritis rat. Antisense probe. (C) Nephritis rat. Sense probe. (D) Control rat stained with anti-FasL monoclonal antibody. (E) Nephritisrat stained with anti-FasL monoclonal antibody. (F) Nephritis rat. Control IgM. Note glomerular FasL-positive cells in proliferative glomerulone-phritis, while tubular epithelium is stained in both control and nephritis rats. Magnification:3400.
1272 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1266–1277, 2000

cell effector cell density could be attributed to decreased FasLexpression under these culture conditions. Expression of FasLby tumor cells is thought to limit the immune response againstthe tumor (9,10). We might hypothesize that tubular cell FasLkeeps in check the renal immune response. Indeed, local FasLexpression by epithelial cells has been implicated in immuneprivilege in the eye and testis (12,13). Nevertheless, this latterview has been called into question (35), and transgenic expres-sion of FasL by isletb cells did not protect them againstrejection (36).
Renal FasL was increased in two models of renal inflam-mation: murine lupus nephritis and rat immune complex-me-
diated proliferative glomerulonephritis. In addition to tubularepithelial cells, local sources of FasL during tubulointerstitialrenal injury include infiltrating leukocytes and interstitial fi-broblasts (16). Indeed, we noted that interstitial cells expressedFasL in lupus mice with combined glomerular and tubuloin-terstitial nephritis. Moreover,de novoglomerular FasL expres-sion was observed in immune complex-mediated proliferativeglomerulonephritis in mice and rats. The cellular origin ofglomerular FasL is not clear from our studies. Infiltrating Tcells or macrophages may be sources of FasL in this model(23). However, cultured glomerular mesangial cells also ex-press FasL transcripts, and inflammatory cytokines releasedduring glomerular injury, such as TNF-a, increase mesangialcell FasL mRNA levels (16).
During renal damage, FasL may promote apoptosis of renalparenchymal cells. In this sense, glomerular mesangial cellsundergo apoptosis when challenged with agonistic anti-Fasantibodies bothin vitro and in vivo (4,5). By contrast, non-stimulated tubular epithelial cells of human or murine originhave been reported to be resistant to cell death induced byagonistic anti-Fas antibodies (15,16). This feature is shared bymurine tubular epithelial cellsin vivo, as systemic injection ofagonistic anti-Fas antibodies induced apoptosis in glomerularcells but not in tubular epithelium (4). Recombinant FasL is amore potent inducer of apoptosis than agonistic anti-Fas anti-bodies (17), and cells that have been previously reported to beresistant to anti-Fas-induced apoptosis were later found to besensitive to FasL-induced apoptosis (17). Our present studiesconfirm that nonstimulated murine tubular epithelial cells arequite resistant not only to anti-Fas antibodies (16), but also torecombinant, cross-linked FasL when grown in the presence ofsurvival factors. Even when the survival factors present inserum were removed, only high concentrations of FasL in-creased significantly, although mildly, the rate of apoptosis inMCT tubular epithelial cells. This fact, together with the lowspontaneous apoptotic rate of cultured tubular epithelial cells,suggests that under basal conditions FasL does not act as anautocrine death factor. Indeed, neutralizing anti-FasL antibod-ies did not modify the rate of spontaneous apoptosis in these
Figure 7.FasL expression in murine cultured tubular epithelial cells. (A) RT-PCR. MCT, proximal tubular epithelial cell line. (B) RT-PCR.TEC, primary cultures of mouse tubular epithelial cells. (C) Northern blot, poly(A) mRNA (MCT cells). MWM, molecular weight markers.
Figure 8. In situhybridization for FasL in MCT cells. (A) Antisenseprobe. (B) Antisense probe. (C) Sense probe. (D) RNase-treated,antisense probe. Magnification:3200 in A and C;3400 in B and D.
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1273
cells. This is also the case for other cell types with constitutiveexpression of FasL (2).
Several factors may contribute to the relative resistance oftubular epithelium to FasL-induced death. The amount of cellsurface Fas receptor is one of them. Indeed, the basal expres-sion of Fas is low in tubular epithelial cells, below the limit of
detection of the flow cytometry technique we used. Inflamma-tory stimuli that increase Fas expression prime mesangial cellsto undergo apoptosis induced by anti-Fas antibodies (16). Inthis regard, increased Fas expression has been noted in tubularepithelial cells in the ischemia-reperfusion model of renalinjury and during chronic tubular atrophy (6,37). Under these
Figure 9. FasL in murine cultured tubular epithelial cells is in the membrane fraction. (A) Western blot in proximal tubular MCT cells.Cytosolic and membrane-bound proteins were separated. FasL is present in the membrane fraction. Nonconfluent (NC) tubular epithelial cellsexpress more FasL than confluent (C) cells. 293 FasL, FasL-transfected 293 cells. (B) Flow cytometry of cell surface FasL expression in MCTcells. (C) Western blot in primary cultures of murine tubular epithelial cells.
Figure 10.Biological activity of tubular epithelial cell FasL. Target Fas-sensitive A20 cells and Fas-resistant A20R cells were cocultured witheffector MCT tubular epithelial cells at different effector:target ratios. Lower cytotoxicity at the highest E:T ratio is consistent with lower FasLexpression by more confluent tubular epithelial cells. Mean of three independent experiments. Neutralization of FasL activity was achieved byadding the neutralizing anti-FasL antibody MFL3 (aFasL). Recombinant FasL (FasL) was used as positive control.
1274 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1266–1277, 2000
circumstances, tubular epithelial cells may undergo apoptosisupon Fas stimulation, in a manner analogous to other FasL-expressing epithelia, such as thyrocytes, during thyroid inflam-mation (2,37). Indeed, Fas deficiency protected tubular epithe-lial cells from apoptosis induced by renal ischemia-reperfusion(6). In vitro, a combination of inflammatory mediators thatmay be present in the kidney during renal injury increasedtubular epithelium Fas expression and sensitivity to FasL-induced apoptosis both in MCT cells and primary cultures oftubular epithelial cells. By contrast, the survival factors presentin serum decreased tubular cell Fas and protected againstFasL-induced apoptosis. In addition to the amount of Fasreceptors, endogenous intracellular proteins may protect fromFas-induced death, as inhibition of protein synthesis promotedanti-Fas-induced apoptosis in nonstimulated cultured humantubular epithelial cells (15). Candidate proteins include mem-bers of the Bcl2 family of anti-apoptotic proteins, such asBclxL. Constitutive expression of BclxL by tubular epithelialcells and its absence from glomerular cells (38,39) may be oneof the factors that contributes to their different susceptibility toFasL apoptosisin vivo. Indeed, both TNF-a and serum depri-vation decreased BclxL levels in tubular epithelial cells (32).
It is noteworthy that both full-length transmembrane FasL aswell as the soluble form of the cytokine were increased inmurine lupus nephritis. Local expression in the cell surface isimportant for the ability of FasL to induce cell death, as theapoptotic-inducing capacity of naturally, enzymatically pro-cessed soluble FasL is reduced by.1000-fold compared withcell surface transmembrane FasL (22). Soluble FasL may evenfunction as a death antagonist, by competing with the moreactive cell membrane FasL (17). The presence of soluble FasLduring renal inflammation may limit the lethal activity oftransmembrane FasL. Indeed, in contrast to results in acuterenal failure (6), in the course of lupus nephritis Fas deficiencydid not protect tubular epithelial cells from apoptosis (40).
We have explored the relationship between renal FasL andapoptosis. However, FasL may have additional functions. Re-cent evidence suggests that soluble FasL can promote neutro-phil chemotaxis (41). In addition, FasL itself can transduceintracellular signals (42). It is currently unknown whether thisis another pathway for FasL to influence tubular epithelial cellbiology.
In summary, FasL is expressed by tubular cells in normalkidney andde novoin the glomeruli during renal injury. Renal
Figure 11.Recombinant FasL induces apoptosis of tubular epithelial cells. (A) MCT cells. High concentrations of recombinant FasL induceda significant albeit low amount of apoptosis in serum-deprived tubular epithelial cells (SF, serum-free), while cells grown in the presence ofthe survival factors of fetal calf serum (FCS) were completely resistant. Tubular epithelial cells previously stimulated with the combination of30 ng/ml tumor necrosis factor-a (TNF-a), 300 U/ml interferon-g (IFN-g), and 10mg/ml lipopolysaccharide (LPS) for 24 h (SF stimulated)were more sensitive to the lethal effect of FasL. Apoptosis induced after a 24-h incubation with increasing concentrations of recombinant FasLwas quantified by flow cytometry of permeabilized, propidium iodide-stained cells. *P , 0.05versuscontrol with no FasL. Mean6 SEM ofthree independent experiments. (B) Western blot. Fas is expressed by MCT cells under the regulation of serum and inflammatory mediators.Cells were stimulated with 10% FCS or 30 ng/ml TNF-a, 300 U/ml IFN-g, and 10mg/ml LPS for 48 h. (C) Flow cytometry. Cell surface Fasexpression is increased following stimulation with TNF-a, IFN-g, and LPS (SF stimulated) for 48 h. The thin line corresponds to cells stainedwith control IgG and the thick line corresponds to those stained with anti-Fas. Under basal conditions, the thick line overlies the thin linebecause the technique is not sensitive enough to detect Fas. (D) Primary tubular epithelial cells stimulated with 30 ng/ml TNF-a, 300 U/mlIFN-g, and 10mg/ml LPS for 24 h undergo apoptosis when challenged with recombinant FasL. Magnification:3200.
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1275

cell FasL promotes apoptosis of lymphoid cells. It had beenpreviously demonstrated that Fas activation induced apoptosisof mesangial cellsin vitro andin vivo. We now show that whileunder basal conditions tubular epithelial cells are relativelyresistant, recombinant FasL induces apoptosis of tubular epi-thelial cells activated by inflammatory stimuli. Taken together,these results suggest that FasL and Fas play a role in normalkidney homeostasis and renal injury that deserves furtherstudy.
AcknowledgmentsThis work was supported by grants from Fondo de Investigaciones
Sanitarias de la Seguridad Social (98/0637), Ministerio de Educacio´ny Ciencia (PB 94/0211, PM 95/0093), Comision Interministerial deCiencia y Tecnologı´a (SAF 97/0071), Instituto Reina Sofia de Inves-tigaciones Nefrolo´gicas, and European Concerted Action GrantBMH4-CT98-3631 (DG12-SSMI). Dr. Lorz was supported by Min-isterio de Educacio´n y Ciencia.
References1. Nagata S, Golstein P: The Fas death factor.Science267: 1449–
1456, 19952. Giordano C, Stassi G, De Maria R, De Maria R, Todaro M,
Richiusa P, Papoff G, Ruberti G, Bagnasco M, Testi R, GalluzzoA: Potential involvement of Fas and its ligand in the pathogen-esis of Hashimoto’s thyroiditis.Science275: 960–963, 1997
3. Kondo T, Suda T, Fukuyama H, Adachi M, Nagata S: Essentialroles of the Fas ligand in the development of hepatitis.Nat Med3: 409–413, 1997
4. Gonza´lez-Cuadrado S, Lorz C, Garcı´a del Moral R, O’Valle F,Alonso C, Ramiro F, Ortiz-Gonza´lez A, Egido J, Ortiz A: Ago-nistic anti-Fas antibodies induce glomerular cell apoptosis inmice in vivo.Kidney Int51: 1739–1746, 1997
5. Gonza´lez-Cuadrado S, Lo´pez-Armada MJ, Go´mez-Guerrero C,Subira D, Ortiz-Gonza´lez A, Neilson EG, Egido J, Ortiz A:Anti-Fas antibodies induce cytolysis and apoptosis in culturedhuman mesangial cells.Kidney Int49: 1064–1070, 1996
6. Nogae S, Miyazaki M, Kobayashi N, Saito T, Abe K, Saito H,Nakane PK, Nakanishi Y, Koji T: Induction of apoptosis inischemia-reperfusion model of mouse kidney: Possible involve-ment of Fas.J Am Soc Nephrol9: 620–631, 1998
7. Suda T, Takahashi T, Golstein P, Nagata S: Molecular cloningand expression of the Fas ligand, a novel member of the tumornecrosis factor family.Cell 75: 1169–1178, 1993
8. French LE, Hahne M, Viard I, Radlgruber G, Zanone R, BeckerK, Muller C, Tschopp J: Fas and Fas ligand in embryos and adultmice: Ligand expression in several immune-privileged tissuesand coexpression in adult tissues characterized by apoptotic cellturnover.J Cell Biol 133: 335–343, 1996
9. Hahne M, Rimoldi D, Schroter M, Romero P, Schreier M, FrenchLE, Schneider P, Bornand T, Fontana A, Lienard D, Cerottini J,Tschopp J: Melanoma cell expression of Fas (Apo-1/CD95)ligand: Implications for tumor immune escape.Science274:1363–1366, 1996
10. Strand S, Hofmann WJ, Hug H, Muller M, Otto G, Strand D,Mariani SM, Stremmel W, Krammer PH, Galle PR: Lymphocyteapoptosis induced by CD95 (APO-1/Fas) ligand-expressing tu-mor cells: A mechanism of immune evasion?Nat Med2: 1361–1366, 1996
11. Bonfoco E, Stuart PM, Brunner T, Lin T, Griffith TS, Gao Y,
Nakajima H, Henkart PA, Ferguson TA, Green DR: Induciblenonlymphoid expression of Fas ligand is responsible for supe-rantigen-induced peripheral deletion of T cells.Immunity 9:711–720, 1998
12. Bellgrau D, Gold D, Selawry H, Moore J, Franzusoff A, DukeRC: A role for CD95 ligand in preventing graft rejection.Nature377: 630–632, 1995
13. Griffith TS, Brunner T, Fletcher M, Green DR, Ferguson TA:Fas ligand-induced apoptosis as a mechanism of immune privi-lege.Science270: 1189–1192, 1995
14. Ortiz A, Gonzalez-Cuadrado S, Lorz C, Garcia del Moral R,O’Valle F, Egido J: Cytokines and Fas regulate apoptosis inmurine renal interstitial fibroblasts.J Am Soc Nephrol8: 1845–1854, 1997
15. Boonstra JG, van der Woude FJ, Wever PC, Laterveer JC, DahaMR, van Kooten C: Expression and function of Fas (CD95) onhuman renal tubular epithelial cells.J Am Soc Nephrol8: 1517–1524, 1997
16. Ortiz-Arduan A, Danoff TM, Kalluri R, Gonza´lez-Cuadrado S,Karp SL, Elkon K, Egido J, Neilson EG: Regulation of Fasexpression in cultured murine renal cells and in the kidney duringendotoxemia.Am J Physiol40: F1193–F1201, 1996
17. Suda T, Hashimoto H, Tanaka M, Ochi T, Nagata S: MembraneFas ligand kills human peripheral blood T lymphocytes, andsoluble Fas ligand blocks the killing.J Exp Med186: 2045–2050,1997
18. Haverty TP, Kelly CJ, Hines WH, Amenta PS, Watanabe M,Harper RA, Kefalides NA, Neilson EG: Characterization of arenal tubular epithelial cell line which secretes the autologoustarget of autoimmune experimental interstitial nephritis.J CellBiol 107: 1359–1368, 1988
19. Garcı´a-Ocan˜a A, de Miguel F, Pen˜aranda C, Albar JP, Sarasa JL,Esbrit P: Parathyroid hormone-related protein is an autocrinemodulator of rabbit proximal tubule cell growth.J Bone MinerRes10: 1875–1884, 1995
20. Anel A, Gamen S, Alava MA, Schmitt-Verhulst AM, Pineiro A,Naval J: Inhibition of CPP32-like proteases prevents granzymeB- and Fas-, but not granzyme A-based cytotoxicity exerted byCTL clones.J Immunol158: 1999–2006, 1997
21. Kayagaki N, Yamaguchi N, Nagao F, Matsuo S, Maeda H,Okumura K, Yagita H: Polymorphism of murine Fas ligand thataffects the biological activity.Proc Natl Acad Sci USA94:3914–3919, 1997
22. Schneider P, Holler N, Bodmer JL, Hahne M, Frei K, Fontana A,Tschopp J: Conversion of membrane-bound Fas(CD95) ligand toits soluble form is associated with downregulation of its pro-apoptotic activity and loss of liver toxicityJ Exp Med187:1205–1213, 1998
23. Ortiz A, Alonso J, Gomez-Chiarri M, Lerma JL, Seron D, Con-dom E, Gonza´lez E, Egido J: Fibronectin decreases glomerularlesions and synthesis of TNF-a, PAF and fibronectin in prolif-erative glomerulonephritis.Clin Exp Immunol101: 334–340,1995
24. Ortiz A, Lorz C, Catala´n MP, Ortiz A, Egido J: Cyclosporine Ainduces apoptosis in murine tubular epithelial cells: Role ofcaspases.Kidney Int54[Suppl 68]: S25–S29, 1998
25. Chomczynski P, Sacchi N: Single-step method of RNA isolationby acid guanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem162: 156–159, 1987
26. Duque N, Gomez-Guerrero C, Egido J: Interaction of IgA withFc alpha receptors of human mesangial cells activates transcrip-tion factor nuclear factor-k B and induces expression and syn-
1276 Journal of the American Society of Nephrology J Am Soc Nephrol 11: 1266–1277, 2000

thesis of monocyte chemoattractant protein-1, IL-8, and IFN-inducible protein 10.J Immunol159: 3474–3482, 1997
27. Rodriguez-Tarduchy G, Sahuquillo AG, Alarcon B, Bragado R:Apoptosis but not other activation events is inhibited by a mu-tation in the transmembrane domain of T cell receptorb thatimpairs CD3j association.J Biol Chem271: 30417–30425, 1996
28. Mariani SM, Matiba B, Baumler C, Krammer PH: Regulation ofcell surface APO-1/Fas (CD95) ligand expression by metallo-proteases.Eur J Immunol25: 2303–2307, 1995
29. Largo R, Gomez-Garre D, Santos S, Pen˜aranda C, Blanco J,Esbrit P, Egido J: Renal expression of parathyroid hormone-related protein (PTHrP) and PTH/PTHrP receptor in a rat modelof tubulointerstitial damage.Kidney Int55: 82–90, 1999
30. D’Agati V, Trudel M: Lectin characterization of cystogenesis inthe SBM transgenic model of polycystic kidney disease.J AmSoc Nephrol3: 975–983, 1992
31. Bossi G, Griffiths GM: Degranulation plays an essential part inregulating cell surface expression of Fas ligand in T cells andnatural killer cells.Nat Med5: 90–96, 1999
32. Ortiz A, Lorz C, Catala´n MP, Danoff TM, Yamasaki Y, Egido J,Neilson EG: Expression of apoptosis regulatory proteins in tu-bular epithelium stressed in culture or following acute renalfailure. Kidney Int57: 969–981, 2000
33. Tsutsui H, Nakanishi K, Matsui K, Higashino K, Okamura H,Miyazawa Y, Kaneda K: IFN-gamma-inducing factor up-regu-lates Fas ligand-mediated cytotoxic activity of murine naturalkiller cell clones.J Immunol157: 3967–3973, 1996
34. Kiener PA, Davis PM, Rankin BM, Klebanoff SJ, Ledbetter JA,Starling GC, Liles WC: Human monocytic cells contain high
levels of intracellular Fas ligand: Rapid release following cellularactivation.J Immunol159: 1594–1598, 1997
35. Vaux DL: Immunology: Ways around rejection [Abstract].Na-ture 394: 133, 1998
36. Allison J, Georgiou HM, Strasser A, Vaux DL: Transgenicexpression of CD95 ligand on islet beta cells induces a granulo-cytic infiltration but does not confer immune privilege upon isletallografts.Proc Natl Acad Sci USA94: 3943–3947, 1997
37. Schelling JR, Nkemere N, Kopp JB, Cleveland RP: Fas-depen-dent fratricidal apoptosis is a mechanism of tubular epithelial celldeletion in chronic renal failure.Lab Invest78: 813–824, 1998
38. Krajewski S, Krajewska M, Shabaik A, Wang HG, Irie S, FongL, Reed JC: Immunohistochemical analysis of in vivo patterns ofBcl-X expression.Cancer Res54: 5501–5507, 1994
39. Ortiz A, Ziyadeh FN, Neilson EG: Expression of apoptosis-regulatory genes in renal proximal tubular epithelial cells ex-posed to high ambient glucose and in diabetic kidneys.J InvestMed 45: 50–56, 1997
40. Wada T, Schwarting A, Kinoshita K, Naito T, Griffiths RC,Coffman TM, Kelley VR: Fas on renal parenchymal cells doesnot promote autoimmune nephritis in MRL mice.Kidney Int55:841–851, 1999
41. Seino K, Iwabuchi K, Kayagaki N, Miyata R, Nagaoka I, Mat-suzawa A, Fukao K, Yagita H, Okumura K: Chemotactic activityof soluble Fas ligand against phagocytes.J Immunol161: 4484–4488, 1998
42. Desbarats J, Duke RC, Newell MK: Newly discovered role forFas ligand in the cell-cycle arrest of CD41 T cells.Nat Med4:1377–1382, 1998
J Am Soc Nephrol 11: 1266–1277, 2000 Fas Ligand in Renal Injury 1277
Copyright © 2022 FDOKUMEN




















