
Nonlocal second-order geometric equations arising in tomographic reconstruction
Upload
independentCategory
view
0download
0
European Journal of Neuroscience, Vol. 4 , pp. 221 -234 @ 1992 European Neuroscience Association
Widespread Horizontal Connections Arising from Layer 5/6 Border Inverted Cells in Rabbit Visual Cortex
Concepcion Reblet, Antonio Lopez-Medina, Sonia M. Gbmez-Urquijo and Jose L. Bueno-Lopez Seccion de Anatomia, Departamento de Neurcciencias, Facultad de Medicina y Odontologia, Universidad del Pais Vasco, E48940 Leioa, Vizcaya, Spain Key words: Neural cells, neural networks, corticocortical, intrinsic, area 17, area 18, WGA - HRP, Fluoro-Gold
Abstract
Herein we describe the inverted cells [defined as those projection neurons having a major dendritic shaft abpially oriented (BueneL6pez et a/., Eur. J. Neurosci, 3, 415, 1991)l originating a unique set of cortical connections characterized by extraordinarily widespread horizontal distribution. Single and multiple injections of wheatgerm agglutinin - horseradish peroxidase were made in areas 17 and 18 and the resulting retrograde labelling in the cortex was analysed. The findings were assessed in independent control experiments in which FluoMold was used as retrograde tracer. Following single injections in area 17 several separate patches of labelled cells comprising layers 2-6 were consistently found in area 18. In addition to these associational cells a number of labelled cells appeared at the layer 516 border but were distributed over most of the tangential extent of the visual occipital cortex. This widespread pattern was particularly striking in brains after multiple injections. In these brains a conspicuous band of labelled cells at the 516 border radiated from the injection sites, making up an apparently continuous horizontal sheet that intersected the striate - extrastriate boundary and merged with the patches of labelled cells in area 18 and beyond. Most of the cells in the 516 border band were inverted cells (82V0; n = 2081). Injections in area 18 failed to produce such a widespread set of labelled cells in area 17. The functional significance of these connections furnished by the !Y6 border inverted cells remains to be determined, but their distribution would allow for convergent/divergent binding interactions both intra-areally (within area 17) and inter-areally (from area 18 to area 17).
Introduction
A general feature of cortical organization is that within a single cortical area each projection cell population providing a given target can have a specific morphology and laminar distribution. The characteristic dendritic distribution of each projection cell population allows sampling of a unique set of afferents, and thus different types of information can be relayed from a given cortical area to different targets (Katz, 1987; Hallman et al., 1988; Hubener and Bolz, 1988; De Lima et al., 1990). There is also evidence that among the cells of origin of a given projection a certain amount of heterogeneity in their morphology and laminar distribution can exist. This heterogeneity is particularly noticeable in corticocortical projections which can arise from various subtypes of pyramidal cells in layers 2-6, from spiny stellate cells in low layer 3 and layer 4, and from polymorphic cells in lnfragranular layers (Innocenti and Fiore, 1976; Meyer and Albus, 1981; Hedreen and Yin, 1981; Tigges et al., 1981; Lund, 1984; Schwartz and Goldman-Rakic, 1984; Winguth and Winer, 1986; Voigt et al., 1988; Einstein and Fitzpatrick, 1991). It is conceivable that the functional properties of the different anatomical types of corticocortical cells are distinct. A given corticocortical projection would thus be better viewed as a system of projections rather than a single efferent projection, allowing different information flows to be transferred in parallel from one cortical area
to another. Characterizing the projection patterns of the different cell types that furnish a given cortical target may well provide insights into the understanding of cortical function.
Among the polymorphic cells endowing corticocortical connections, a conspicuous cell class, the inverted cells, has recently been distinguished (Bueno-Lbpez et ul., 1991). Inverted cells, which are defined by a major dendritic system directed towards the white matter, are for the most part the so-called inverted pyramidal neurons (Cajal, 1899-1900, 1911; O’Leary and Bishop, 1938; Van der Loos, 1965; Globus and Scheibel, 1967; Valverde et al., 1989), but also include those among Cajal’s triangular and hiform cells with prominent descending dendritic systems (Cajal, 1899- 1900, 191 1). These inverted cells occur mainly in layers 5 and 6 and project chiefly to cortical areas. This projection is seen in rabbits (BuewGpez et al., 1988, 1991; Muller-Paschinger and Tombijl, 1989) and very probably across the mammalian series (Innocenti, 1980; Code and Winer, 1985; Games and Winer, 1988; De Lima etal . , 1990; Einstein and Fitzpatrick, 1991). Thus, the selective involvement of inverted cells in corticocortical connections and their particular laminar distribution and dendritic orientation suggests that these cells may play a specific role in cortical processing.
In the present study we sought to ascertain whether the pattern of
Correspondence to: J. L. BueneLbpez, as above
Received 30 July 1991. revised 23 August 1991. accepted 30 October I991

222 Distributed inverted cells
visuocortical connections arising from inverted cells is different from that arising from pyramidal cells. Cells were retrogradely labelled with single and multiple injections of retrograde tracers [either wheatgerm agglutinin - horseradish peroxidase (WGA -HRP) or Fluoro-Gold] into the visual occipital cortex, and the distribution of labelled inverted and labelled pyramidal cells in the visual cortex was analysed. We carried out the experiments on rabbit cortex because its non-convoluted nature allows the identification of cell morphology without the influence of distortions caused by cortical folding. Preliminary accounts of parts of this work have been presented to the European Neuroscience Association (Mpez-Medina ef al., 1988) and to the International Brain Research Organization (Reblet ef al. , 1991).
Materials and methods Animals, surgery and WGA - HRP experiments
The results were derived from 18 adult albino New Zealand rabbits of both sexes (2.5 -3 kg body weight) which had been raised in our breeding colony. Thirteen animals received WGA-HRP injections in area 17 and three animals in the extrastriate occipital cortex lateral to area 17 (area 18); as a control, the remaining two rabbits were injected with Fluoro-Gold in area 17 (Table I ) . All surgical procedures were done under aseptic conditions. Anaesthesia was achieved by an intraperitoneal injection of a mixture of ketamine hydrochloride (35 mg/kg) and xylazine (5 mg/kg) (White and Holmes, 1976; Vogt and Sikes, 1990). During surgery reflexes were tested periodically to assure a profound level of anaesthesia. Once anaesthetized, each animal was placed in a stereotaxic frame, the head stabilized, and the body temperature maintained at 37-38°C by a heating pad. The occipital cortex was exposed and single or multiple injections of 0.015-0.04 pI of 3% WGA-HRP (Sigma) in 0.5 M NaCl were given in different portions of areas 17 or 18 using a glass micropipette (tip diameter, 50 pm) attached to a I-pl Hamilton syringe. Multiple injections consisted of either rnediolaterally or rostrocaudally oriented rows of 4-5 regularly spaced sites, 0.015 -0.02 pl of WGA-HRP being injected into each site and the point-to-point inter-injection distances being 1 - 1.5 mm. After injection, the syringe was left in place for 5 min to minimize the backflow of the tracer along
TABLE 1. Details of experiments
Experiment Tracer Number of injections Survival time X (h)
RBI0 WGA-HRP 3% I XO.04 45 RB2 1 WGA-HRP 3% I X0.04 45 RB27 WGA-HRP 3% 5X0.02 43 RB30 WGA-HRP 3% 1 XO.04 50 RB34 WGA-HRP 3% 4X0.02 48 RB35 WGA-HRP 3% 4X0.015 40 RB37 WGA-HRP 3% 5X0.015 46 RB42 WGA-HRP 3% I x0.03 45 RB44 WGA-HRP 3% I x0.03 43 RB45 WGA-HRP 3% I x0.025 70 R&16 WGA-HRP 3% I XO.025 70 RB47 WGA-HRP 3% 1 ~ 0 . 0 2 5 7 0 RB48 WGA-HRP 3% 1 x0.03 48 RB49 WGA-HRP 3% I X0.015 48 RE353 WGA-HRP 3% 5X0.015 42 RB55 WGA-HRP 3% 5X0.015 43 T56 Fluoro-Gold 5 % 3 ~ 0 . 5 48 T63 Fluoro-Gold 5 % I X0.5 72
amount of tracer (PI)
the pipette tract. Recovery from anaesthesia was quick and complete. Following postinjection survival times of 40-70 h the rabbits were
anaesthetized with chloral hydrate (150 rnglkg) mixed with pentobarbital (30 mg/kg) [Chloropent, Fort Dodge Laboratories, Fort Dodge, IA, USA; 3.5 ml/kg i.p.; Vogt and Sikes (1990)l; they were then'perhsed through the cardiac left ventricle and ascending aorta with 750 ml of 0.9% NaCl (pH 7.4; 37"C), followed by 1ooO-1500 ml of 1 % paraformaldehyde and 2% glutaraldehyde in 0.12 M phosphate buffer (pH 7.4; room temperature) containing 3% sucrose. The brain was removed from the skull and stored in cold phosphate buffer (4°C) with 30% sucrose until it sank, then coronally sectioned on a freezing microtome at 80 pm. One of every two sections was processed for HRP histochemistry following the tetramethylbenzidine (TMB) procedure of Gibson et al. (1984). Incubated sections were washed in cold buffer, mounted onto gelatinized slides and air dried overnight; after that they were dehydrated, cleared and coverslipped. From each experiment alternate sections were counterstained with 1 % toluidine blue in distilled water to identify cytoarchitectural patterns. The sections were examined and the neurons drawn and classified within 2 weeks of the tracer injections.
Analysis All the specimens used in this study were selected with the tracer injection centred on layer 4 and no apparent invasion of the underlying white matter by the dark core of the tracer diffusion around the injection centre. The coronal sections were examined in the light microscope under bright- field and dark-field (polarized light) illumination for the presence of cells and axons labelled by TMB reaction product. The injection sites and the general distribution of the labelled cell and axonal fields were charted onto serial drawings of the sections to which vessel profiles and other landmarks were added in order to assign the labelling its proper position.
To investigate the laminar distribution and morphology of the labelled cells samples of sections were taken from relevant regions (see Results). The exact location of each labelled cell in these samples was plotted onto camera lucida drawings of the sections. In these drawings the radial dimension of the grey matter was divided into 20 equally thick horizontal strata, thus assigning each labelled cell to one of the strata. In turn, these strata were assigned to the cortical layers as cytoarchitectonically identified in the Nissl-stained sections. Only labelled cells showing good quality of labelling and having their soma and the labelled portions of their dendrites included in the 80 pm-thick sections were subsequently taken into account for morphological evaluation. These well-labelled cells were classified as inverted cells or as pyramidal cells upon their display of a prominent dendritic shaft oriented with <45" deviation from the radial cortical axis towards the white matter or the pia respectively. The remaining well-labelled cells were assigned to the category of 'unclassified cells'. The morphologies of the WGA-HRP labelled, inverted cells were compared with those of Golgi-impregnated inverted cells available from previous work (Bueno-Mpez er al., 1991).
Fluoro-Gold controls
To further assess the projection cell morphologies we decided to carry out control experiments in which the good quality of cell labelling yielded by the retrograde axonal transport of Fluoro-Gold was exploited. Typically, the procedure followed in these control experiments was- from anaesthesia to analysis-akin to that described above for the WGA -HRP experiments. It consisted of either a rostrocaudal triple injection (0.5 pl into each site, the sites being 1 mm apart) or a single injection (0.5 pl) of 5% Fluoro-Gold (Fluorochrome Inc., Englewood, CO, USA) in Ringer's solution. Both injections were attempted on area

Distributed inverted cells 223
17. Following the protocol of Schmued and Fallon (1986), 48-72 h after the Fluoro-Gold injection the animals were killed under deep anaesthesia and perfused through the left cardiac ventricle with successive solutions of phosphate-buffered saline (PBS) (0.1 M, pH 7.4; 37°C) and a fixative (0.1 M; pH 7.4; room temperature) of 4% paraformaldehyde in PBS containing 5 % sucrose. The brain was removed after 1 h and stored in PBS (0.1 M, pH 7.4; 4°C) with 20% sucrose until it sank; it was then frozen and sliced into 80-pm coronal sections, which were collected in PBS (0.1 M; pH 7.4). Immediately after that two sets of all alternate sections were mounted onto gelatin-coated slides
and air dried. One set was then xylene cleared, coverslipped with non- fluorescent DPX and stored in the refrigerator. The other set was counterstained with 0.05% methylene blue in distilled water prior to coverslipping (Sripanidkulchai and Wyss, 1986).
The counterstained sections were immediately examined in a Nikon Optiphot microscope equipped with an episcopic-fluorescence attachment (model EF-D) with a 100 W mercury lamp and a UV-1A filter cube. The location of labelling was plotted onto drawings of these sections following a procedure akin to that described above for WGA-HRP experiments. The morphological and quantitative analyses of the labelled
A
FIG. 1. Cytoarchitecture of rabbit visual cortex. (A) Drawing of a dorsolateral view of the rabbit brain showing the estimated location of areas 17 and 18 of the present study and the level at which the section shown in B and C was taken (bars). (B and C) Photomicrographs of a toluidine blue-stained coronal section through the visual cortex showing the cytoarchitectonic pattern of areas 29d. 17 and 18. Scale bars: in B. 500 pm; in C, 300 pm.
224 Distributed inverted cells
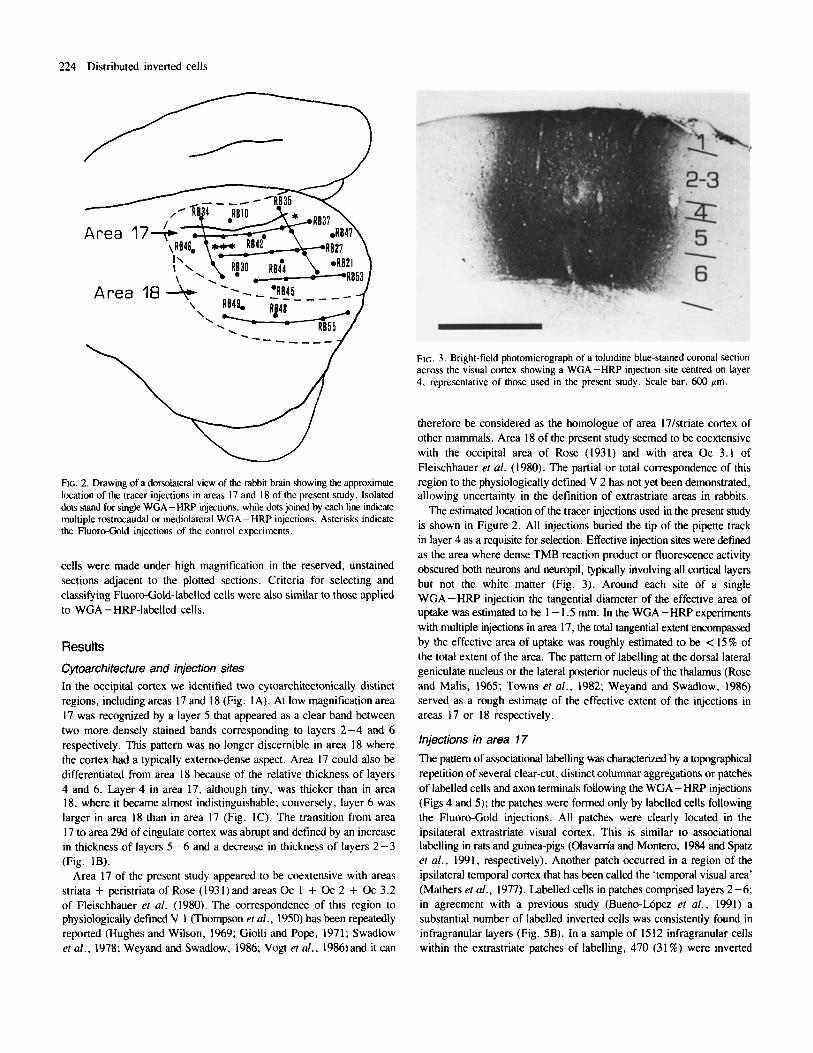
FIG. 2. Drawing of a dorsolated view of the rabbit brain showing the approximate location of the tracer injections in areas 17 and 18 of the present study. Isolated dots stand for single WGA-HRP injections, while dots joined by each line indicate multiple rostrocaudal or mediolateral WGA - HRP injections. Asterisks indicate the Fluoro-Gold injections of the control experiments.
cells were made under high magnification in the reserved, unstained sections adjacent to the plotted sections. Criteria for selecting and classifying Fluoro-Gold-labelled cells were also similar to those applied to WGA-HRP-labelled cells.
Results Cytoarchitecture and injection sites In the occipital cortex we identified two cytoarchitectonically distinct regions, including areas 17 and 18 (Fig. IA). At low magnification area 17 was recognized by a layer 5 that appeared as a clear band between two more densely stained bands corresponding to layers 2-4 and 6 respectively. This pattern was no longer discernible in area 18 where the cortex had a typically extemo-dense aspect. Area 17 could also be differentiated from area 18 because of the relative thickness of layers 4 and 6. Layer 4 in area 17, although tiny, was thicker than in area 18, where it became almost indistinguishable; conversely, layer 6 was larger in area 18 than in area 17 (Fig. IC). The transition from area I7 to area 29d of cingulate cortex was abrupt and defined by an increase in thickness of layers 5 -6 and a decrease in thickness of layers 2 -3 (Fig. IB).
Area 17 of the present study appeared to be coextensive with areas striata + peristriata of Rose (1931)and areas Oc 1 + Oc 2 + Oc 3.2 of Fleischhauer et al. (1980). The correspondence of this region to physiologically defined V 1 (Thompson et al., 1950) has been repeatedly reported (Hughes and Wilson, 1969; Giolli and Pope, 1971; Swadlow et al., 1978; Weyand and Swadlow, 1986; Vogt et al., 1986)and it can
FIG. 3. Bright-field photomicrograph of a toluidine blue-stained coronal section across the visual cortex showing a WGA-HRP injection site centred on layer 4. representative of those used in the present study. Scale bar, 600 pm.
therefore be considered as the homologue of area 17htriate cortex of other mammals. Area 18 of the present study seemed to be coextensive with the occipital area of Rose (1931) and with area Oc 3.1 of Fleischhauer et al. (1980). The partial or total correspondence of this region to the physiologically defined V 2 has not yet been demonstrated, allowing uncertainty in the definition of extrastriate areas in rabbits.
The estimated location of the tracer injections used in the present study is shown in Figure 2. All injections buried the tip of the pipette track in layer 4 as a requisite for selection. Effective injection sites were defined as the area where dense TMB reaction product or fluorescence activity obscured both neurons and neuropil, typically involving all cortical layers but not the white matter (Fig. 3). Around each site of a single WGA-HRP injection the tangential diameter of the effective area of uptake was estimated to be 1 - 1.5 mm. In the WGA - H W experiments with multiple injections in area 17, the total tangential extent encompassed by the effective area of uptake was roughly estimated to be < 15% of the total extent of the area. The pattern of labelling at the dorsal lateral geniculate nucleus or the lateral posterior nucleus of the thalamus (Rose and Malis, 1965; Towns et al., 1982; Weyand and Swadlow, 1986) served as a rough estimate of the effective extent of the injections in areas 17 or 18 respectively.
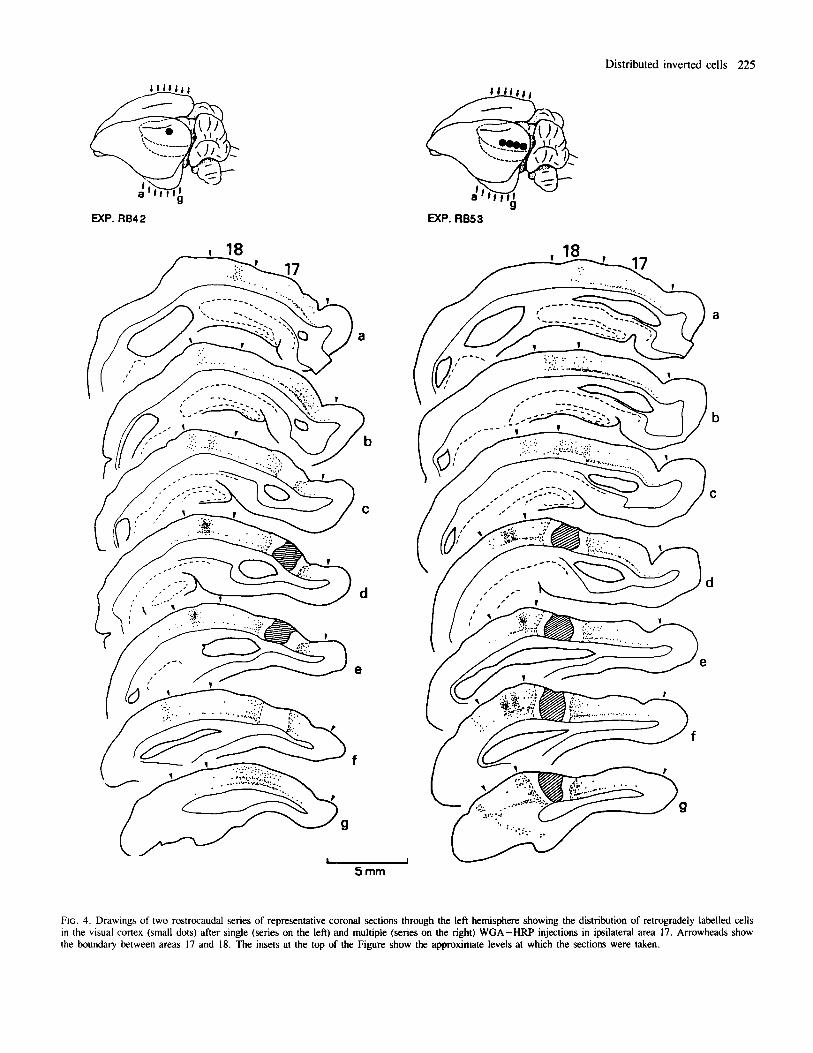
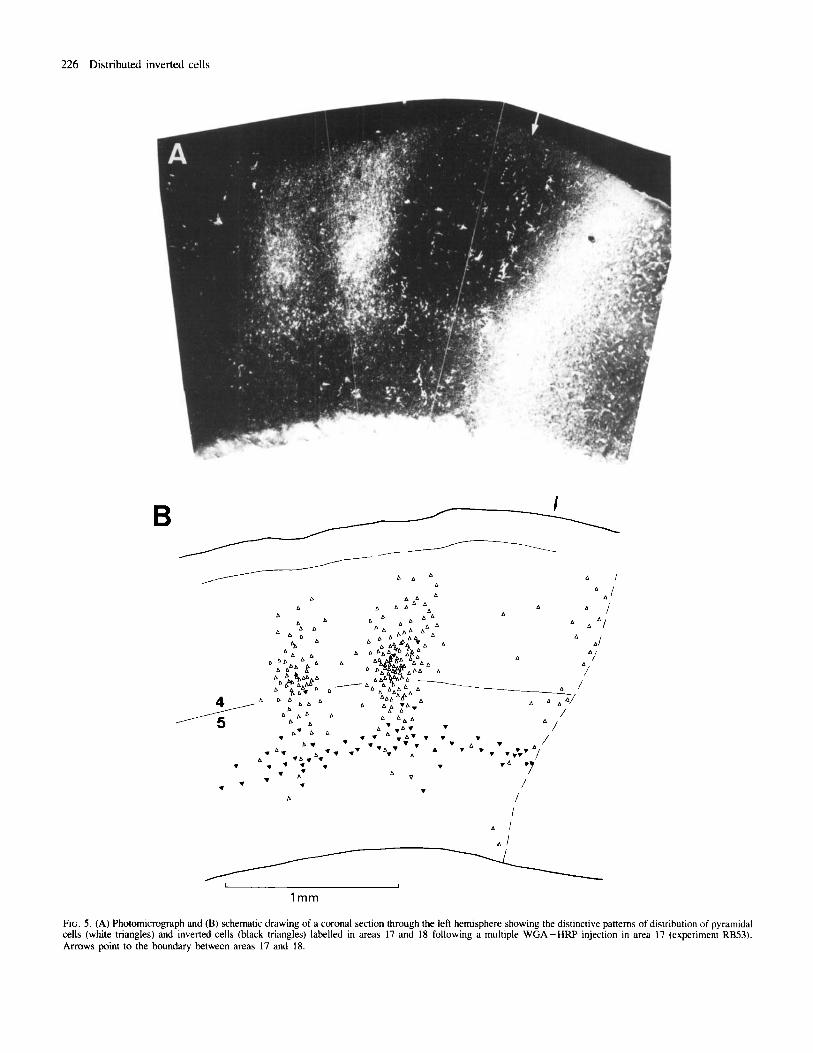
Injections in area 17 The pattern of associational labelling was characterized by a topographical repetition of several clear-cut, distinct columnar aggregations or patches of labelled cells and axon terminals following the WGA-HRP injections (Figs 4 and 5); the patches were formed only by labelled cells following the Fluoro-Gold injections. All patches were clearly located in the ipsilateral extrastriate visual cortex. This is similar to associational labelling in rats and guinea-pigs (Olavam’a and Montero, 1984 and Spatz et al., 1991, respectively). Another patch occurred in a region of the ipsilateral temporal cortex that has been called the ‘temporal visual area’ (Mathers et al., 1977). Labelled cells in patches comprised layers 2-6; in agreement with a previous study (Bueno-L6pez er al., 1991) a substantial number of labelled inverted cells was consistently found in infragranular layers (Fig. 5B). In a sample of 1512 infragranular cells within the extrastriate patches of labelling, 470 (31 %) were inverted
Distributed inverted cells 225
- MP. RB53
, , , . - . . .. . .... :
FIG. 4. Drawings of two rostrocaudal series of representative coronal sections through the left hemisphere showing the distribution of retrogradely labelled cells in the visual cortex (small dots) after single (series on the left) and multiple (series on the right) WGA-HRP injections in ipsilateral area 17. Arrowheads show the boundary between areas 17 and 18. The insets at the top of the Figure show the approximate levels at which the sections were taken.
226 Distributed inverted cells
. . . . b
v
A '
d A A l
A A " I
1 rnrn
FIG. 5 . (A) Photomicrograph and (B) schematic drawing of a coronal section through the left hemisphere showing the distinctive patterns of distribution of pyramidal cells (white triangles) and inverted cells (black triangles) labelled in areas 17 and 18 following a multiple WGA-HRP injection in area 17 (experiment RB53). Arrows point to the boundary between areas 17 and 18.
Distributed inverted cells 227
cells, 881 (58.5%) were pyramidal cells and 159 (10.5%) unclassified cells. This sample was taken from at least two coronal sections spaced 240 pm apart through each patch of extrastriate labelling of four animals (experiments RB30, RB42, RB44 and RB45). No percentage differences were apparent among sections, patches or experiments.
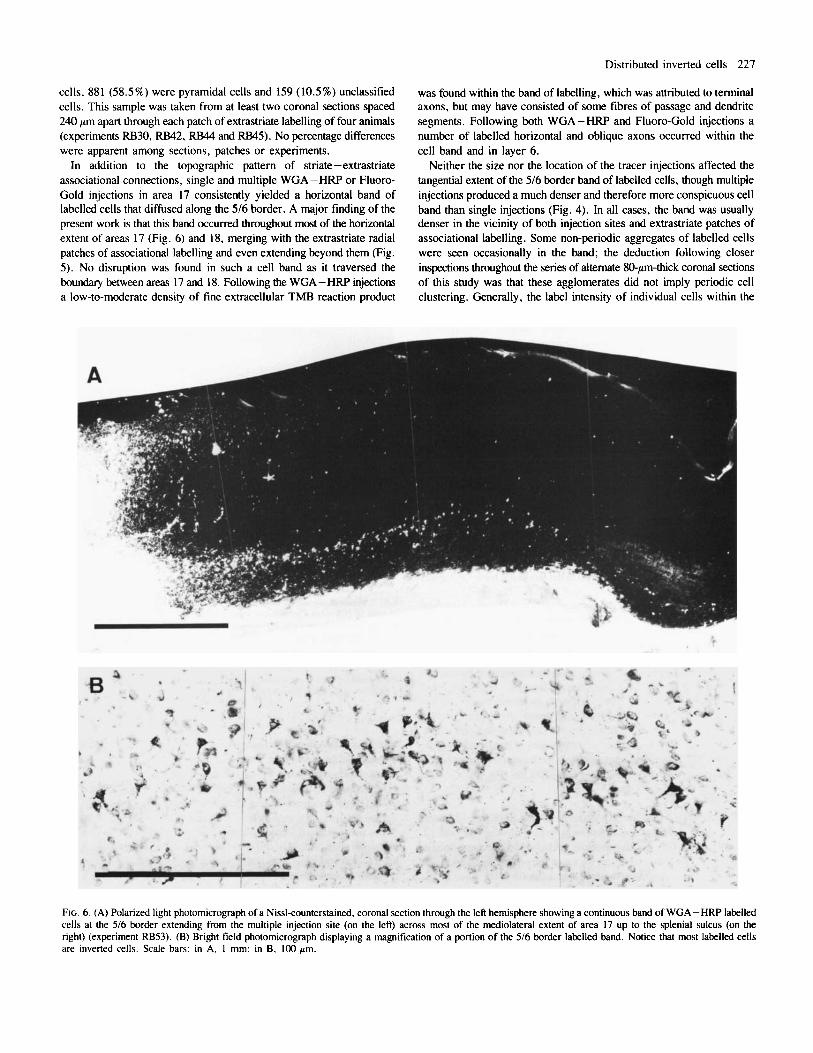
In addition to the topographic pattern of striate-extrastriate associational connections, single and multiple WGA -HW or Fluoro- Gold injections in area 17 consistently yielded a horizontal band of labelled cells that diffused along the 5/6 border. A major finding of the present work is that this band occurred throughout most of the horizontal extent of areas 17 (Fig. 6) and 18, merging with the extrastriate radial patches of associational labelling and even extending beyond them (Fig. 5). No disruption was found in such a cell band as it traversed the boundary between areas 17 and 18. Following the WGA-HRP injections a low-to-moderate density of fine extracellular TMB reaction product
was found within the band of labelling, which was attributed to terminal axons, but may have consisted of some fibres of passage and dendrite segments. Following both WGA -HRP and Fluoro-Gold injections a number of labelled horizontal and oblique axons occurred within the cell band and in layer 6.
Neither the size nor the location of the tracer injections affected the tangential extent of the 5/6 border band of labelled cells, though multiple injections produced a much denser and therefore more conspicuous cell band than single injections (Fig. 4). In all cases, the band was usually denser in the vicinity of both injection sites and extrastriate patches of associational labelling. Some non-periodic aggregates of labelled cells were seen occasionally in the band; the deduction following closer inspections throughout the series of alternate 80-pm-thick coronal sections of this study was that these agglomerates did not imply periodic cell clustering. Generally, the label intensity of individual cells within the
FIG. 6. (A) Polarized light photomicrograph of a Nissl-counterstained, coronal section through the left hemisphere showing a continuous band of WGA -HRP labelled cells at the 5/6 border extending from the multiple injection site (on the left) across most of the mediolateral extent of area 17 up to the splenial sulcus (on the right) (experiment RB53). (B) Bright field photomicrograph displaying a magnification of a portion of the 516 border labelled band. Notice that most labelled cells are inverted cells. Scale bars: in A, 1 mm; in B, 100 pm.

228 Distributed inverted cells
5/6 border band increased with the size of a given tracer injection; both lightly and heavily labelled cells could occasionally be found mixing with one another. This finding suggests that the cells were either in a different functional state at the perfusion time or that they have different patterns of axonal arborization.
Another major finding of this work was that most labelled cells at the 5/6 border band were inverted cells (Figs 5B and 6B). The quantitative analysis of labelled 5/6 border cells confirmed the impression gained from the qualitative observations. In a sample of 1574 WGA-HRP- labelled cells of the 5/6 border band, 1339 (85%) were inverted cells, 47 (3%) were pyramidal cells and the remaining 188 (12%) were unclassified cells. This sample was taken from a periodic, rostrocaudal series of coronal sections spaced 240 pm apart through the whole visual cortex of one animal (experiment RB53) and included all well-labelled cells found in infragranular layers, either in striate or extrastriate cortex but outside the area 18 patches and the tracer diffusion halo at the area 17 injection site. Later counts in similar samples of four other WGA - HRP experiments showed no significant differences (percentage test; P < 0.05).
The morphology and percentages of the inverted cells were independently assessed through use of Fluoro-Gold as a retrogradely transported tracer (Table 1 and Figs 2 and 7). Thus, when 243 well- labelled cells sampled at the 5/6 border band of one of the control experiments (experiment T56) were counted, 182 (75%) were inverted cells, 29 (12%) were pyramidal cells and 32 (13%) unclassified cells. Further, 264 well-labelled cells sampled at the 5/6 border band in the remaining control experiment were counted; of these cells, 188 (71 %) were inverted cells, 19 (7%) were pyramidal cells and 57 (22%) were unclassified cells. Additionally it should be noted that there were no apparent peaks of labelled inverted cells intermingling with peaks of other labelled cells in the cell band in any of the experiments. In the labelled samples of 5/6 border inverted cells of the present study, most cells appeared to be so-called inverted pyramids. The range of morphological variability of the inverted cells included in the samples is shown in Figures 7 and 8.
Besides the 5/6 border labelling across areas 17 and 18, other horizontal labelling extended from the tracer injections of area 17; this labelling, being part of layers 2-3 and 5a, was restricted to area 17. The total horizontal spread of this labelling from the halo of tracer diffusion could reach up to 1.5 mm, the layer 2-3 labelling usually being broader than the layer 5a labelling. The cell density within this labelling in layers 2-3 and 5a decreased with the distance from the injection site (Figs 4, 5 and 6). As in the 516 border band, a low to moderate density of fine extracellular TMB reaction product was found within this labelling following the WGA-HRP injections. Some periodicity was observed in this anterograde labelling of layer 2 - 3, the clusters of anterograde labelling b e i g thicker than the intervening labelling-free zones; the average centre- to-centre spacing of the clusters was 600-800 pm (Fig. 9).
Injections in area 18 To compare the distribution of neurons projecting from area 17 to area 18 with the distribution found following tracer injections in area 17, we made two single and one multiple WGA-HRP injections in area 18 (Fig. 2). These injections had the tip of the pipette centred in layer 4 and did not show invasion by the tracer diffusion of the white matter underlying the injection or the (area) 17/18 boundary medial to the injection. All three injections involved the intermediate portion (in the rostrocaudal direction) of area 18, and each produced a broad, clear- cut patch of associational labelling within area 17 but not widespread
FIG. 7. Photomicrograph of inverted cells retrogradely labelled at the 516 border of area 18 following Fluoro-Gold injections in area 17 (experiment T56). The findings of the Fluoro-Gold experiments fully confirmed those derived from the WGA-HRP experiments of the present study, concerning not only the widespread distribution of the cell band at the 5/6 border across areas 17 and 18 but also the morphology of the labelled cells. Note the axon emergence site in the two inverted cells looking like so-called inverted pyramids (arrows). Additionally. the top left cell is a Cajal's triangular cell, and the neighbouring cell resembles a fusiform cell. Scale bar. 50 pm.
horizontal connections diffusing across area 18 and/or area 17 (Fig. 10). To date we have not been able to perform satisfactory injections of Fluoro-Gold restricted to the grey matter of area 18, and this absence of extraordinarily widespread labelling at the 516 border following WGA-HRP injections in area 18 has thus not been adequately cross- examined and might therefore be a false negative finding.
The overwhelming majority of these patch neurons in area 17 were pyramidal cells, with only a few inverted cells occurring in infragranular layers. This is distinct from the reciprocal projection and corroborates previous results (Bueno-L6pez et al., 1991). Thus, of 604 labelled infragranular cells, 49 (8%) were inverted cells, 492 (81.5%) were pyramidal cells, and the remaining 63 (10.5%) were unclassified cells. This sample was taken from at least five Nissl-counterstained coronal sections spaced 240 pm apart through the labelled patch in area 17 in
Distributed inverted cells 229
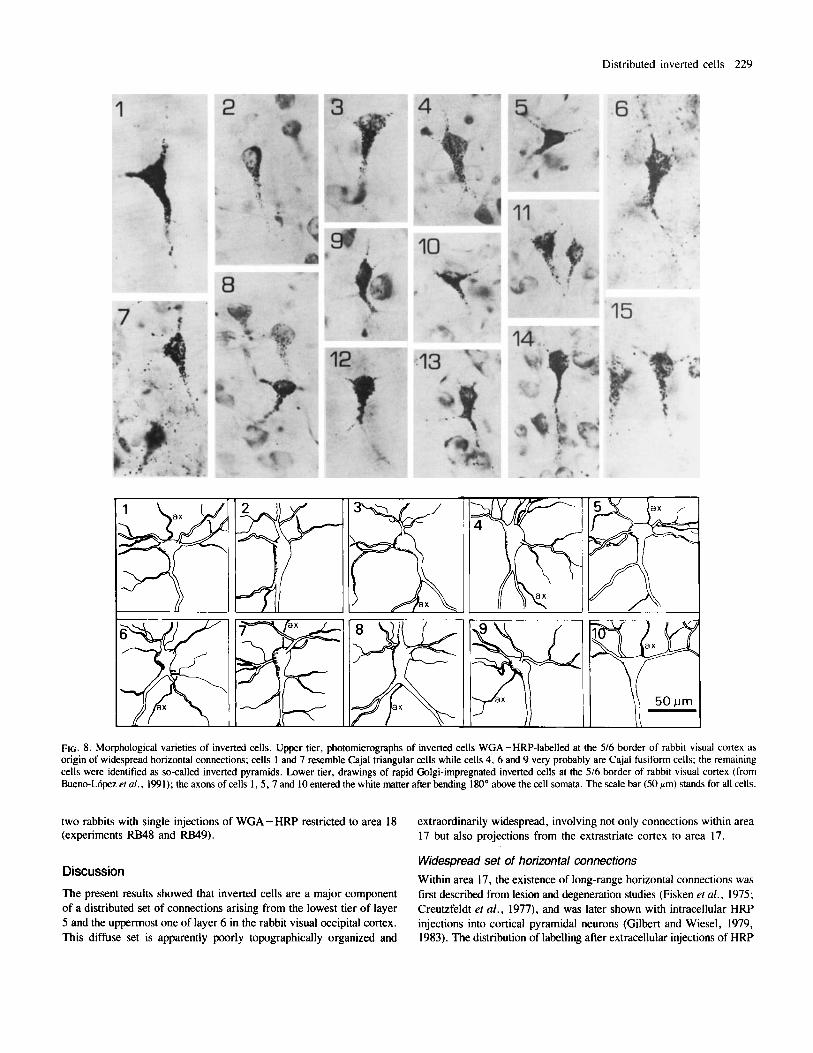
FIG. 8. Morphological varieties of inverted cells. Upper tier, photomicrographs of inverted cells WGA-HRP-labelled at the 5/6 border of rabbit visual cortex as origin of widespread horizontal connections; cells 1 and 7 resemble Cajal triangular cells while cells 4, 6 and 9 very probably are Cajal fusiform cells; the remaining cells were identified as so-called inverted pyramids. Lower tier, drawings of rapid Golgi-impregnated inverted cells at the 5/6 border of rabbit visual cortex (from Bueno-Lbpez el a/ . , 1991); the axons of cells I , 5 .7 and 10 entered the white matter after bending 180" above the cell somata. The scale bar (50 am) stands for all cells.
two rabbits with single injections of WGA-HRP restricted to area 18 (experiments RB48 and RB49).
extraordinarily widespread, involving not only connections within area 17 but also projections from the extrastriate cortex to area 17.
Widespread set of horizontal connections Within area 17, the existence of long-range horizontal connections was first described from lesion and degeneration studies (Fisken ef al., 1975; Creutzfeldt et al . , 1977), and was later shown with intracellular HRP injections into cortical pyramidal neurons (Gilbert and Wiesel, 1979, 1983). The distribution of labelling after extracellular injections of HRP
Discussion The present results showed that inverted cells are a major component of a distributed set of connections arising from the lowest tier of layer 5 and the uppermost one of layer 6 in the rabbit visual occipital cortex. This diffuse set is apparently poorly topographically organized and
230 Distributed inverted cells
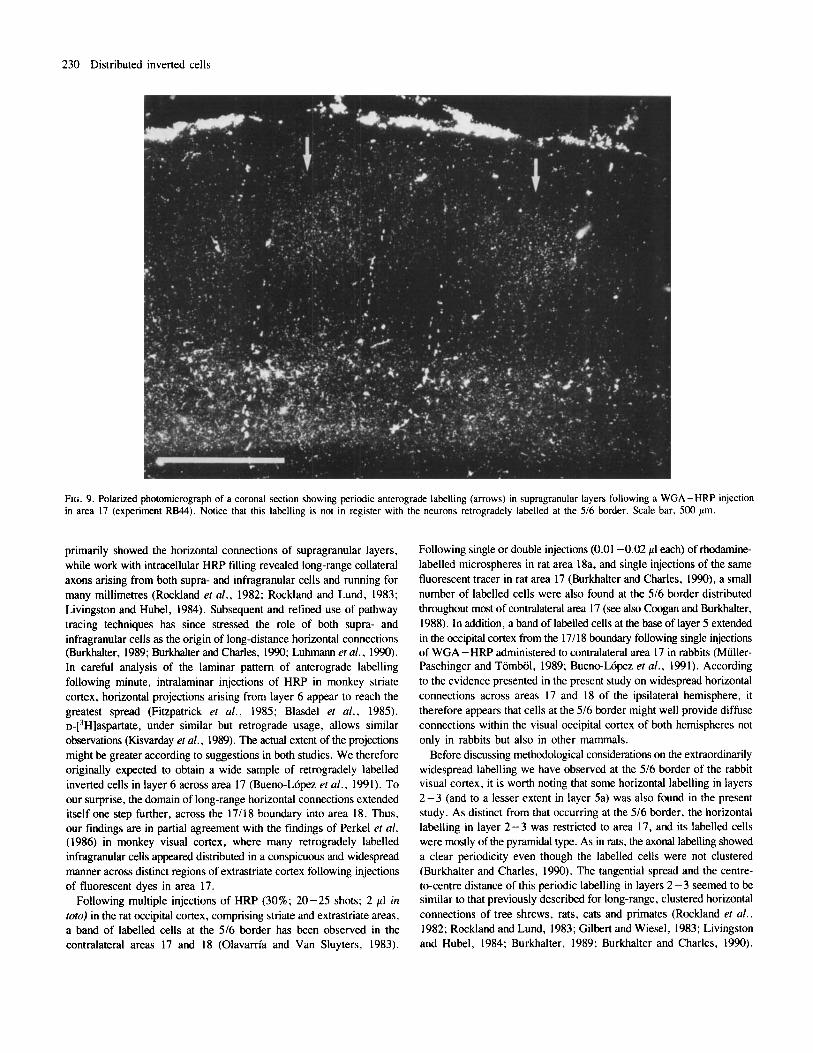
FIG. 9. Polarized photomicrograph of a coronal section showing periodic anterograde labelling (arrows) in supragranular layers following a WGA -HRP injection in area 17 (experiment RB44). Notice that this labelling is not in register with the neurons retrogradely labelled at the 516 border. Scale bar. 500 pm.
primarily showed the horizontal connections of supragranular layers, while work with intracellular HRP filling revealed long-range collateral axons arising from both supra- and infragranular cells and running for many millimetres (Rockland et al., 1982; Rockland and Lund, 1983; Livingston and Hubel, 1984). Subsequent and refined use of pathway tracing techniques has since stressed the role of both supra- and infragranular cells as the origin of long-distance horizontal connections (Burkhalter, 1989; Burkhalter and Charles, 1990; Luhann etal.. 1990). In careful analysis of the laminar pattern of anterograde labelling following minute, intralaminar injections of HRP in monkey striate cortex, horizontal projections arising from layer 6 appear to reach the greatest spread (Fitzpatrick et a / . , 1985; Blasdel et a / . , 1985). ~-[~H]aspartate, under similar but retrograde usage, allows similar observations (K~svarday et al., 1989). The actual extent of the projections might be greater according to suggestions in both studies. We therefore originally expected to obtain a wide sample of retrogradely labelled inverted cells in layer 6 across area 17 (Bueno-Lopez et a / . , 1991). To our surprise, the domain of long-range horizontal connections extended itself one step further, across the 17/18 boundary into area 18. Thus, our findings are in partial agreement with the findings of Perkel et al. (1986) in monkey visual cortex, where many retrogradely labelled infragranular cells appeared distributed in a conspicuous and widespread manner across distinct regions of extrastriate cortex following injections of fluorescent dyes in area 17.
Following multiple injections of HRP (30%; 20-25 shots; 2 pl in toto) in the rat occipital cortex, comprising striate and extrastriate areas, a band of labelled cells at the 5/6 border has been observed in the contralateral areas 17 and 18 (Olavarria and Van Sluyters, 1983).
Following single or double injections (0.01 -0.02 pl each) of rhcdamine- labelled microspheres in rat area 18a, and single injections of the same fluorescent tracer in rat area 17 (Burkhalter and Charles, 1990). a small number of labelled cells were also found at the 5/6 border distributed throughout most of contralateral a m 17 (see also Coogan and Burkhalter, 1988). In addition, a band of labelled cells at the base of layer 5 extended in the occipital cortex from the 17/18 boundary following single injections of WGA-HRP administered to contralateral area 17 in rabbits (Miiller- Paschinger and Tombal, 1989; BueneL6pez etal., 1991). According to the evidence presented in the present study on widespread horizontal connections across areas 17 and 18 of the ipsilateral hemisphere, it therefore appears that cells at the 5/6 border might well provide diffuse connections within the visual occipital cortex of both hemispheres not only in rabbits but also in other mammals.
Before discussing methodological considerations on the extraordinarily widespread labelling we have observed at the 5/6 border of the rabbit visual cortex, it is worth noting that some horizontal labelling in layers 2-3 (and to a lesser extent in layer 5a) was also found in the present study. As distinct from that occurring at the 5/6 border, the horizontal labelling in layer 2-3 was restricted to area 17, and its labelled cells were mostly of the pyramidal type. As in rats, the axonal labelling showed a clear periodicity even though the labelled cells were not clustered (Burkhalter and Charles, 1990). The tangential spread and the centre- to-centre distance of this periodic labelling in layers 2 -3 seemed to be similar to that previously described for long-range, clustered horizontal connections of tree shrews, rats, cats and primates (Rockland et a / . . 1982; Rockland and Lund, 1983; Gilbert and Wiesel, 1983; Livingston and Hubel, 1984; Burkhalter, 1989; Burkhalter and Charles, 1990).
Distributed inverted cells 23 1
- 5mm
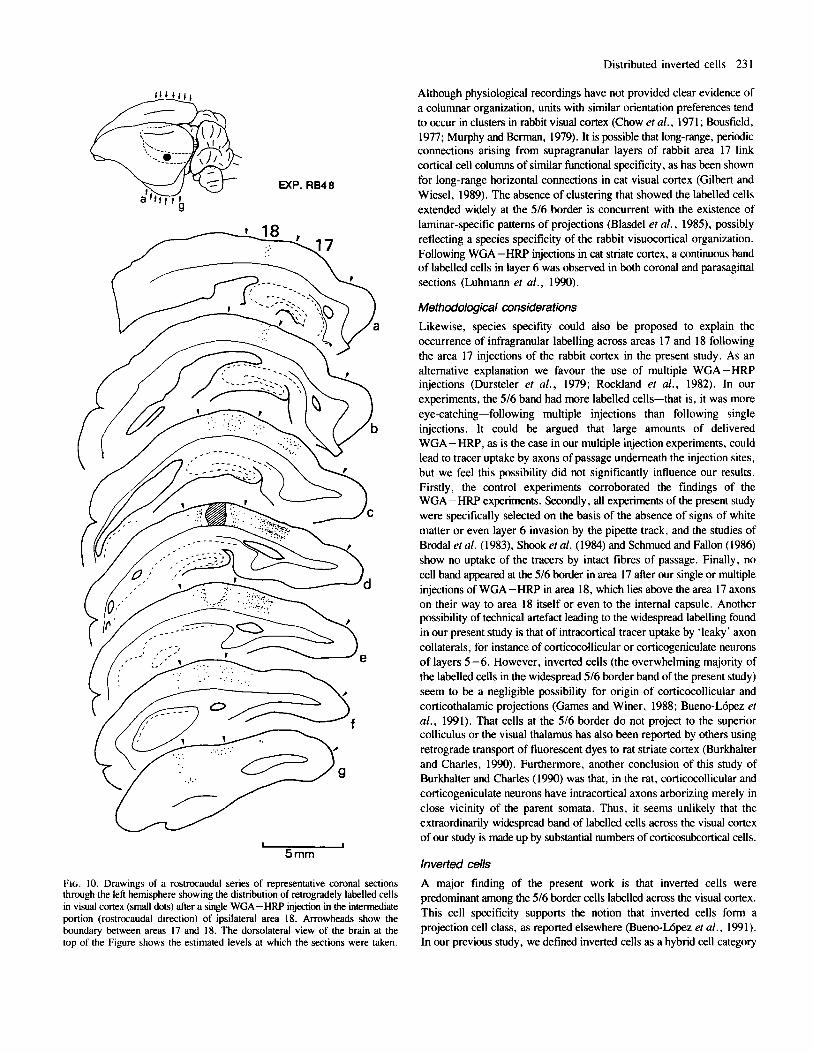
FIG. 10. Drawings of a rostrocaudal series of representative coronal sections through the left hemisphere showing the distribution of retrogradely labelled cells in visual cortex (small dots) after a single WGA-HRP injection in the intermediate portion (rostrocaudal direction) of ipsilateral area 18. Arrowheads show the boundary between areas 17 and 18. The dorsolateral view of the brain at the top of the Figure shows the estimated levels at which the sections were taken.
Although physiological recordings have not provided clear evidence of a columnar organization, units with similar orientation preferences tend to occur in clusters in rabbit visual cortex (Chow et al., 1971; busfield, 1977; Murphy and Berman, 1979). It is possible that long-range, periodic connections arising from supragranular layers of rabbit area 17 link cortical cell columns of similar functional specificity, as has been shown for long-range horizontal connections in cat visual cortex (Gilbert and Wiesel, 1989). The absence of clustering that showed the labelled cells extended widely at the 5/6 border is concurrent with the existence of laminar-specific patterns of projections (Blasdel et al., 1985), possibly reflecting a species specificity of the rabbit visuocortical organization. Following WGA-HRP injections in cat striate cortex, a continuous band of labelled cells in layer 6 was observed in both coronal and parasagittal sections (Luhmann et al., 1990).
Methodological considerations Likewise, species specifity could also be proposed to explain the occurrence of infragranular labelling across areas 17 and 18 following the area 17 injections of the rabbit cortex in the present study. As an alternative explanation we favour the use of multiple WGA-HRP injections (Dursteler et al., 1979; Rockland et al., 1982). In our experiments, the 5/6 band had more labelled cells-that is, it was more eye-catching-following multiple injections than following single injections. It could be argued that large amounts of delivered WGA-HRP, as is the case in our multiple injection experiments, could lead to tracer uptake by axons of passage underneath the injection sites, but we feel this possibility did not significantly influence our results. Firstly, the control experiments corroborated the findings of the WGA- HRP experiments. Secondly, aJl experiments of the present study were specifically selected on the basis of the absence of signs of white matter or even layer 6 invasion by the pipette track, and the studies of Brodal et al. (1983), Shook et al. (1984) and Schmued and Fallon (1986) show no uptake of the tracers by intact fibres of passage. Finally, no cell band appeared at the 5/6 border in area 17 after our single or multiple injections of WGA-HRP in area 18, which lies above the area 17 axons on their way to area 18 itself or even to the internal capsule. Another possibility of technical artefact leading to the widespread labelling found in our present study is that of intracortical tracer uptake by ‘leaky’ axon collaterals, for instance of corticocollicular or corticogeniculate neurons of layers 5 -6. However, inverted cells (the overwhelming majority of the labelled cells in the widespread 516 border band of the present study) seem to be a negligible possibility for origin of corticocollicular and corticothalamic projections (Games and Winer, 1988; Bueno-L6pez et al., 1991). That cells at the 5/6 border do not project to the superior colliculus or the visual thalamus has also been reported by others using retrograde transport of fluorescent dyes to rat striate cortex (Burkhalter and Charles, 1990). Furthermore, another conclusion of this study of Burkhalter and Charles (1990) was that, in the rat, corticocollicular and corticogeniculate neurons have intracortical axons arborizing merely in close vicinity of the parent somata. Thus, it seems unlikely that the extraordinarily widespread band of labelled cells across the visual cortex of our study is made up by substantial numbers of corticosubcortical cells.
Inverted cells A major finding of the present work is that inverted cells were predominant among the 5/6 border cells labelled across the visual cortex. This cell specificity supports the notion that inverted cells form a projection cell class, as reported elsewhere (Bueno-Upez et al., 1991). In our previous study, we defined inverted cells as a hybrid cell category

232 Distributed inverted cells
that (i) is made up mostly of secalled inverted pyramids but also of other polymorphic cells with their major dendritic shaft oriented in the direction of the white matter, (ii) occurs deep-seated in cortex, their number peaking at the 5/6 border, and (iii) preferentially originates corticocortical (and to a lesser extent other corticotelencephalic) projections.
Why the conspicuous inverted cells have not been detected as the cells of origin of the long-range horizontal connections arising from the 516 border in other mammals might be questioned. Less accurate techniques of cell observation used in previous rat and cat studies might partially explain this, and interspecies variations of cell morphology should be considered as well. We have observed that inverted cells tend to adopt a less conspicuous morphology in rat cortex than in rabbit cortex (J. L. Bueno-Upez, C. Reblet and A. Lbpez-Medina, unpublished). Inverted cells or cells that look like inverted cells are clearly distinguishable in the wall of cortical convolutions, but they apparently adopt the morphology of fusiform cells, at the top of the giri, or horizontal cells, at the bottom of the sulci (Tomkl, 1984; Ferrer et al., 1986a,b; Meyer, 1987; Voigt et al., 1988; Lubke and Albus, 1989; De Lima et al.. 1990). Nonetheless, cells that look like the inverted cells of the present study have been observed (depending on the pathways and the areas of origin) to be the majority of corticocortical infragranular cells not only in rabbits (Bueno-Upez et al., 1988, 1 9 9 1 ; Miiller-Paschinger and Tombiil, 1989) but also in rats and cats (Code and Winer, 1985; Games and Winer, 1988; Einstein and Fitzpahick, 1991). Layer 6 vertical fusiform cells and asymmetrical pyramids, which look like inverted cells as defined in the present study, form the majority of layer 5-6 cells projecting from inferior temporal g y m to prefrontal cortex in monkeys (De Lima et al . , 1990). It is possible that inverted cells lay down basic patterns of corticocortical connections that preferentially arise from infragranular layers throughout the mammalian series.
Axon disposition To explain the widespread set of inverted cells found retrogradely labelled at the 5/6 border in the present study, two axon and cell dispositions could be proposed. Firstly, a widely collateralized, isotropic axonal field occurring for every inverted cell of the 516 border band and covering most of the horizontal dimension of areas 17 and 18 (for area 18 cells) or area 17 (for area 17 cells). Alternatively, the axonal field of a 5/6 border band inverted cell could be anisotropic, extending itself only along a particular direction across the visual cortex and leaving terminals (or clusters of terminals) along its way. The anisotropic disposition must be favoured if we consider that the number of labelled cells at the 5/6 border increased following multiple tracer injections. On the other hand, long and narrow axonal fields, coherent with the proposed anisotropic axonal disposition for the 5/6 border inverted cells, have already been described for pyramidal cells originating long-range horizontal connections in the visual cortex (Gilbert and Wiesel, 1983; Livingston and Hubel, 1984; Gilbert, 1985, and references therein). These long and narrow axonal fields are not in contradiction with some degree of axonal dichotomization such as that shown by others (Martin, 1984; Bullier and Kennedy, 1987; Wilson, 1987; Burkhalter and Charles, 1990; Dreher et al., 1990). Projecting to several ipsilateral (and contralateral?) cortical sites that could be involved in performing a given task in the cortex is an intermediate alternative to the strictly isotropic and anisotropic models.
Functional considerations We will allow some speculations at present, although we have no functional role for the widespread connections that arise from the 516
border band of inverted cells described in this study to suggest. The widespread connections arising from the 516 border inverted cells might serve for axonal inputs received in low layer 5 and in layer 6 to uniquely converge and diverge within the visual cortex. The distributed set of horizontal connections described in the present study might well concur in ‘binding’ the activity of neurons spatially segregated within area 17 and across the various extrastriate visual areas and actively responding to the different aspects of a perceived object. Following experiments in cat visual cortex, it has been hypothesized that this binding could be achieved through temporarily correlated neuronal spike activity of the relevant neurons (Von der Malsburg and Schneider, 1986; Eckhorn et al., 1988; Gray er al., 1989). The horizontal span of the inverted cell connections, and their convergent/divergent disposition, could at least incorporate both intra- and inter-areal modulatory influences to the still unknown mechanism required for correlated firing of the relevant cortical neurons.
Acknowledgements This work was supported by the University of the Basque Country (UPV-20/89) and the Science Department of the Spanish Government (DGICyT-PM-890069). A. L.-M. and S. M. G.-U. were doctoral candidates subsidized by the Basque Country Regional Government. The authors thank Ann Maier and Denise Whitelam for correcting the English language.
Abbreviations HRP horseradish peroxidase PBS phosphate-buffered saline TMB tetramethylbenzidine WGA-HRP wheatgerm agglutinin-horseradish peroxidase
References Blasdel, G. G., Lund, J . S. and Fitzpatrick, D. (1985) Intrinsic connections of
macaque striate cortex: axonal projecfions of cells.outside lamina 4C. J . Neurosci., 5 , 3350-3369.
Bousfield, J . D. (1977) Columnar organization and the visual cortex of the rabbit. Brain Res., 136, 154- 158.
Brcdal, P., Dietrich, E., Bjaalie, J . G. , Nordby, T. and Walberg. F. (1983) Is lectin coupled horseradish peroxidase taken up and transported by undamaged as well as by damaged fibers in the central nervous system? Brain Res., 278,
Bueno, J . L., Grandes, P., Gondra, J . , Hennequet, L., L6pez-Medina. A. and Reblet, C. (1988) Inverted pyramidal neurons in rabbit telencephalic cortex: morphology, laminar location and axonal connectivity. Eur. J. Neurosci.. Suppl. 1 , 242.
Bueno-Mpez. J. L., Reblet, C . . Epez-Medina. A,, G6mez-Urquij0, S. M., Grandes, P. , Gondra, J. and Hennequet, L. (1991) Targets and laminar distribution of projection neurons with ‘inverted’ morphology in rabbit cortex. Eur. J . Neurosci., 3 , 415-430.
Bullier, J . and Kennedy, H. (1987) Axonal bifurcation in the visual system. Trendc Neurosci., 10, 205-210.
Burkhalter, A. (1989) Intrinsic connections of rat primary visual cortex: laminar organization of axonal projections. J . Comp. Neurol., 279, 171 - 186.
Burkhalter. A. and Charles, V . (1990) Organization of local axon collaterals of efferent projection neurons in rat visual cortex. J. Comp. Neurol., 302,
Cajal, S. Ram& y (1899- 1900) Studies on the human cerebral cortex 1-111 [I: Visual cortex, Revista Trimestral Micmgrajca, 4. 1-63. 1899. U: Structure of the motor cortex of man and higher mammals, Revista Trimestral Microgrdjca, 4, 117-200, 1899, and 5, 1 - 1 1 , 1900. 111: Structure of the acoustic cortex, Revista Trimestral MicrogrCjca, 5, 129- 183. 1900.1 In DeFelipe, J . and Jones, E. G. (eds and trans), Cajal On 7he Cerebral Conex: An Annotated Translntion of n e Complete Writings (History of Neuroscience,
1-9.
920 -934.

Distributed inverted cells 233
No. I ) (1988). Oxford University Press, New York. Oxford, pp. 147-288. Cajal, S. Ram& y (191 1) Hisrologie du Systkme Nervelcr de 1 'Homme er des
Vertkbres. Maloine, Paris (C.S.I.C. facsimile edition, Madrid, 1972). Chapters
Chow. K. L.. Masland, R. H. and Stewart, D. L. (1971) Receptive field characteristics of striate cortical neurons in the rabbit. Bmin Res., 33, 337-352.
Code, R. A. and Winer, J. A. (1985) Two systems of commissural projections in layer V of cat primary auditory cortex (AI). Anur. Rec., 212, 226-227.
Coogan, T. A. and Burkhalter, A. (1988) Sequential development of connections between striate and extrastriate visual cortical areas in the rat. J . Comp. Neurol.,
Creutzfeldt, 0. D., Garey, L. J., Kuroda, R. and Wolff, J.-R. (1977) The distribution of degenerating axons after small lesions in the intact and isolated visual cortex of the cat. Exp. Bruin Res., 27, 419-440.
De Lima, A. N., Voigt, T. and Morrison, J. H. (1990) Morphology of the cells within the inferior temporal gyrus that project to the prefrontal cortex in the macaque monkey. J . Comp. Neurol., 296, 159-172.
Dreher, B., Dehay, C. and Bullier, J. (1990) Bihemispheric collateralization of the cortical and subcortical afferents to the rat's visual cortex. Eur. J. Neurosci.,
Dursteler. M. R., Blakemore, C. and Garey, L. J. (1979) Projections to the visual cortex in the golden hamster. J . Comp. Neurol., 183, 185-204.
Eckhorn, R., Bauer, R., Jordan, W., Brosch, M., Kruse, W., Munk, M. and Reitboeck, H. J. (1988) Coherent oscillations: a mechanism of feature linking in the visual cortex? Biol. Cybem., 60, 121 -130.
Einstein, G. and Fitzpatrick, D. (1991) Distribution and morphology of area 17 neurons that project to the cat's extrastriate cortex. J. Comp. Neurol.. 303,
Ferrer, I., Fabregues. I. and Condom, E. (1986a) A Golgi study of the sixth layer of the cerebral cortex. I. The lissencephalic brain of Rodentia, Lagomorpha, Insectivora and Chiroptera. J. Anar., 145, 217-234.
Ferrer, I . , Fabregues, I. and Condom. E. (1986b) A Golgi study of the sixth layer of the cerebral cortex. 11. The gyrencephalic brain of Carnivora, Artiodactyla and Primates. J. Anat., 146. 87- 104.
Fisken, R. A. L., G a y , L. J. and Powell, T. P. S. (1975) The intrinsic association and commissural connections of area 17. Philos. Trans. R. SOC. Lond., Ser. B, 272, 487-536.
Fitzpatrick, K. A,, Lund, J. S. and Blasdel, G. G. (1985) Intrinsic connections of macaque striate cortex: afferent and efferent connections of lamina 4C. J. Neurosci.. 5 , 3329-3349.
Fleischhauer, K.. Zilles, K. and Schleicher, A. (1980) A revised cytoarchitectonic map of the neocortex of the rabbit (Orycrolugus cuniculus). Anat. Embryol. ,
Games, K. D. and Winer, J. A. (1988) Layer V in rat auditory cortex: projections to the inferior colliculus and contralateral cortex. Hear. Res., 34, 1-25.
Gibson, A. R., Hansma, D. I., Houk, J. C. and R o b i n , F. R. (1984) A sensitive low artifact TMB procedure for the demonstration of WGA-HRP in the CNS. Bruin Res., 298, 235-241.
Gilbert. C. D. (1985) Horizontal integration in the neocortex. Trends Neurosci.,
Gilbert, C. D. and Wiesel. T. N. (1979) Morphology and intracortical projections of functionally identified neurons in cat visual cortex. Nature, 280, 120- 125.
Gilbert, C. D. and Wiesel, T. N. (1983) Clustered intrinsic connections in cat visual cortex. J . Neurosci., 3, 11 16- 1133.
Gilbert, C. D. and Wiesel, T. N. (1989) Columnar specificity of intrinsic horizontal and corticocortical connections in cat visual cortex. J. Neurosci., 9,2432-2442.
Giolli, R. A. and Pope, J . E. (1971) The mode of innervation of the dorsal lateral geniculate nucleus and the pulvinar of the rabbit by axons arising from the visual cortex. J. Comp. Neurol.. 147, 129- 144.
Globus, A. and Scheibel, A. B. (l%7) Pattern and field in cortical structure: the rabbit. J . Comp. Neurol., 131, 155-172.
Gray. C. M., Konig, P., Engel. A. K. and Singer, W. (1989) Oscillatory responses in cat visual cortex exhibit intercolumnar synchronization which reflects global stimulus properties. Nature, 338, 334-337.
Hallman. L. R.. Schofield, B. R. and Lin, C . 4 . (1988) Dendritic morphology and axon collaterals of corticotectal, corticopontine and callosal neurons in layer V of primary visual cortex of the hooded rat. J. Comp. Neurol.. 272, 149- 160.
Hedreen, J. C. and Yin, T. C. T. (1981) Homotopic and heterotopic callosal afferents of caudal inferior parietal lobule in Macucu mulana. J. Comp. Neurol.,
Hiibener, M. and Bolz, J. (1988) Morphology of identified projection neurons
Hughes, A. and Wilson, M. E. (1969) Callosal terminations along the boundary
24-35.
278. 242 -252.
2, 317-331.
132- 149.
161. 121 - 143.
8, 160-165.
197, 605-621.
in layer 5 of rat visual cortex. Neurosci. Left., 94, 76-81.
between visual areas I and I1 in the rabbit. Bruin Res.. 12, 19-25. Innocenti, G. M. (1980) The primary visual pathway through the corpus callosum:
morphological and functional aspects in the cat. Arch. frul. Biol., 118, 124- 188. Innocenti, G. M. and Fiore, L. (1976) Morphological correlates of visual field
transformation in corpus callosum. Neurosci. Lert., 2, 245 -252. Katz, L. C. (1987) Local circuitry of identified projection neurons in cat visual
cortex brain slices. J. Neurosci., 7. 1223- 1249. Kisvarday, Z. F., Cowey, A,, Smith, A. D. and Somogyi, P. (1989) Interlaminar
and lateral excitatory amino acid connections in the striate cortex of monkey. J. Neurosci., 9, 667 -682.
Livingston, M. S. and Hubel, D. H. (1984) Anatomy and physiology of a color system in the primate visual cortex. J . Neurosci., 4, 309-356.
L6pez-Medina, A,, Reblet, C. and Bueno, J. L. (1988) Different patterns of corticocortical connections in the visual cortex of the rabbit. Eur. J . Neurosci., Suppl. I , 160 (abstract).
Lubke. J. and Albus, K. (1989) The mstnatal development of laver VI pyramidal ._ neurons in the cat striate cortex, as visualized by intracellular Lucifer yellow injections in aldehyde-fixed tissue. Dev. Bruin Res., 45, 29-38.
Luhmann, H. J., Singer, W. and Martinez-Millan, L. (1990) Horizontal interactions in cat striate cortex: I. Anatomical substrate and postnatal development. Eur. J . Neurosci., 2, 344-357.
Lund, J. S. (1984) Spiny stellate neurons. In Peters, A. and Jones, E. G. (eds), Cerebral Cortex, Vol. 1. Plenum Press, New York, pp. 255-308.
Martin, K. A. C. (1984) Neuronal circuits in cat striate cortex. In Peters, A. and Jones, E. G. (eds), Cerebral Cortex, Vol. 2. Plenum Press, New York, pp. 241-284.
Mathers, L. H., Douville, A. and Chow, K. L. (1977) Anatomical studies of a temporal visual area in the rabbit. S. Comp. Neurol., 171, 147-156.
Meyer, G. (1987) Forms and spatial arrangement of neurons in the primary motor cortex of man. J . Comp. Neurol., 262, 402-428.
Meyer, G. and Albus, K. (1981) Spiny stellates as cells of origin of association fibers from area 17 to area 18 in the cat's m o r t e x . Bruin Res., 210, 335-341.
Miiller-Paschinger, I.-B. and Tombijl, T. (1989) Corticocortical and subcortico- cortical afferent connections of the rabbit's primary visual cortex. Anur.
Murphy, E. H. and Berman, N. (1979) The rabbit and the cat: A comparison of some features of response properties of single cells in the primary visual cortex. J. Comp. Neurol., 188. 401-428.
Olavarria, J. and Montero, V. M. (1984) Relation of callosal and striate -extrastriate cortical connections in the rat: Morphological definition of extrastriate visual areas. Exp. Brain Res., 54, 240-252.
Olavarria, J. and Van Sluyters, R. (1983) Widespread callosal connections in infragranular visual cortex of the rat. Bruin Res.. 279, 233-237.
O'Leary, J. L. and Bishop, G. H. (1938) The optically excitable cortex of the rabbit. J . Comp. Neurol., 68, 423-478.
Perkel, D. J., Bullier, J. and Kennedy, H. (1986) Topography of the afferent connectivity of area 17 in the macaque monkey: a double-labelling study. J. Comp. Neurol., 253, 374-402.
Reblet, C., Bueno-L6pez. J. L. , L6pez-Medina. A,. G6mez-Urquijo. S. M. and Maseda, M. J. (1991) Extrinsic/intrinsic axonal projection system arising from inverted cells located at the layer 5/6 border in rabbit visual cortex. Iff fBR0 Congress (Absrrucrs), 396.
Rockland, K. S. and Lund, J. S. (1983) Intrinsic laminar lattice connections in primate visual cortex. J. Comp. Neurol., 216, 303-318.
Rockland, K. S., Lund, J. S. and Humphrey, A. L. (1982) Anatomical banding of intrinsic connections in striate cortex of the tree shrews (Tupaia glis). J. Comp. Neurol.. 209, 41 -58.
Rose, J. E. and Malis, L. I. (1965) Geniculo-striate connections in the rabbit. 11. Cytoarchitectonic structure of the striate region and of the dorsal lateral geniculate body; organization of the geniculo-striate projections. J. Comp. Neurol.. 125, 121 - 140.
Rose, M. (1931) Cytoarchitektonischer Atlas der Grosshirnrinde des Kaninchens. J. Psychol. Neurol. (Leiq ig) , 43. 353-440.
Schmued, L. C. and Fallon, J. H. (1986) Fluoro-Gold: a new fluorescent retrograde axonal tracer with numerous unique properties. Brain Res., 377,
Schwartz, M. L. and Goldman-Rakic, P. S. (1984) Callosal and intrahemispheric connectivity of the prefrontal association cortex in rhesus monkey: Relation between intraparietal and principal sulcal cortex. J. Comp. Neurol., 226, 406-420.
Shook, B. L., Abramson, B. P. and Chalupa, L. M. (1984) An analysis of the transport of WGA-HRP in the cat's visual system. J. Neurosci. Methods. 11, 65-77.
Embryo/., 180, 81-88.
147- 154.

234 Distributed inverted cells
Spatz, W. B., Vogt, D. M. and Illing, R.-B. (1991) Delineation of striate cortex, and the striate-peristriate projections in the guinea pig. Erp. Brain Res.. 84. 495-504.
Sripanidkulchai, K. and Wyss, M. (1986) Two rapid methods of counterstaining fluorescent dye tracer containing sections without reducing the fluorescence. Brain Res.. 39'7, 117-129.
Swadlow, H. A., Weyand, T . G. and Waxman. S. G. (1978) The cells of origin of the corpus callmum in rabbit visual cortex. Brain Res.. 156. 129- 134.
Thompson, J. M., Woolsey, C. N. and Talbot. S. A. (1950) Visual areas 1 and I1 of cerebral cortex of rabbit. J . Neurophysiol., 13, 277-288.
Tigges, J . , Tigges, M., Anschel, S. , Cross, N. A,, Letbeter. W. D. and McBride. R. D. (1981) Areal and laminar distribution of neurons interconnecting the central visual cortical areas 17, 18, 19, and MT in the squirrel monkey (Saimin). J . Comp. Neurol., 202, 539 - 560.
TomMI, T. (1984) Layer VI cells. In Peters. A. and Jones, E. G. (eds) Cerebral Conex, Vol. I . Plenum Press, New York. pp. 479-520.
Towns, L. C., Burton, S. L., Kimberly, C. J . and Fetterman, M. R. (1982) Projections of the dorsal lateral geniculate and lateral posterior nuclei to visual cortex in the rabbit. J . Comp. Neurol.. 210. 87-98.
Valverde, F., Facal-Valverde, M. V., Santacana. M. and Heredia, M. (1989) Development and differentiation of early generated cells of sublayer VIb in the somatosensory cortex of the rat: a correlated Golgi and autoradiographic study. J . Comp. Neurol.. 290, I 18- 140.
Van der Loos. H. (1965) The 'improperly' oriented pyramidal cell in the cerebral
cortex and its possible bearing on problems of neuronal growth and cell orientation. BUN. Johns Hopkins Hosp., 117, 228-250.
Vogt, B. A. and Sikes. R. W. (1990) Lateral magnocellular thalamic nucleus in rabbits: architecture and projections to cingulate cortex. J . Comp. Neurol.. 299. 64-74.
Vogt, B. A., Sikes. R. W., Swadlow. H. A. and Weyand, T. G . (1986) Rabbit cingulate cortex: cytoarchitecture. physiological border with visual cortex. and afferent cortical connections of visual, motor. postsubicular. and intracingulate origin. J . Comp. Neurol.. 248. 74-94.
Voigt, T.. LeVay. S. and Stamnes, M. A. (1988) Morphological and immunccytochemical observations on the visual callosal projections in the cat. J . Comp. Neurol.. 272, 450-460.
Von der Malsburg. C. and Schneider. V. (1986) A neural cocktail-party processor. Biol. Cvbem.. 54, 29-40.
Weyand. T . G . and Swadlow, H. A. (1986) Thalamic inputs to visual areas I and 2 in the rabbit. J . Comp. Neurol., 250. 521 -528.
White, G . L. and Holmes, D. D. (1976) A comparison of ketamine and the combination of ketamine-xylazine for effective surgical anesthesia in the rabbit. Lab. Anim. Sci., 26, 804-806.
Wilson, C. J . (1987) Morphology and synaptic connections of crossed corticostriatal neurons in the rat. J. Comp. Neurol.. 263. 567-580.
Winguth, S . D. and Winer. J. A. (1986) Corticocorticd connections of cat primary auditory cortex (At): Laminar organization and identification of supragranular neurons projecting to area 11. J . Comp. Neurol.. 248. 36-56.
Copyright © 2022 FDOKUMEN




















