
Relations between cochlear histopathology and hearing loss in experimental cochlear implantation

lable at ScienceDirect
Hearing Research 315 (2014) 34e39
Contents lists avai
Hearing Research
journal homepage: www.elsevier .com/locate/heares
Research paper
Time course of cochlear injury discharge (excitotoxicity) determinedby ABR monitoring of contralateral cochlear events
Jaina Negandhi a, Adrienne L. Harrison a, Cullen Allemang a, Robert V. Harrison a, b, *
a Auditory Science Laboratory, Neuroscience and Mental Health Program, The Hospital for Sick Children, 555 University Ave., Toronto, M5G 1X8 Canadab Department of Otolaryngology-Head and Neck Surgery, Hospital for Sick Children and the University of Toronto, 190 Elizabeth St., Rm 3S-438,R. Fraser Elliott Building, Toronto, M5G 2N2 Canada
a r t i c l e i n f o
Article history:Received 2 April 2014Received in revised form9 June 2014Accepted 17 June 2014Available online 26 June 2014
* Corresponding author. Auditory Science LaboratoHealth Program, The Hospital for Sick Children, 555 U1X8 Canada. Tel.: þ1 416 813 6535.
E-mail address: [email protected] (R.V. Harrison).
http://dx.doi.org/10.1016/j.heares.2014.06.0020378-5955/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
The dynamics of cochlear excitotoxicity can be monitored from effects on the contralateral ear. Afterunilateral mechanical ablation of the cochlea (in a mouse model) we observed immediate elevations inauditory brainstem evoked response (ABR) thresholds in the contralateral ear. Threshold elevationspeaked at 2e3 h post ablation, and returned to baseline levels after 5e6 h. These contralateral effects areinitiated by cochlear afferent injury discharges most likely activating the olivocochlear efferent system.Six hours after cochlear injury, ABR thresholds were fully returned to pre-lesion baseline levels andremained normal for up to 10 days of monitoring. We have confirmed that our cochlear ablation pro-cedure increases short-term activity levels in the auditory brainstem and midbrain using c-fos labelling.The study provides insight into the dynamics of glutamate excitotoxicity, a pathological process directlyrelated to acute tinnitus after acoustic trauma, and more generally implicated in many types of braininjury and neuro-degenerative disease.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Glutamate is the predominant excitatory neurotransmitter inthe brain, typically involved in fast synaptic transmission andtemporal processing. In pathological conditions such synaptic sys-tems can have detrimental effects, for example in excitotoxicity,where overstimulation of glutamatergic receptors leads to neuro-toxicity and cell death (e.g. Mayer and Westbrook, 1987; Choi andRothman, 1990). Excitotoxicity has been implicated in many offorms of neuropathy, such as spinal cord injury, epilepsy, stroke,traumatic brain injury and hearing loss as well as in some neuro-degenerative diseases such as Alzheimer's, and Parkinson's (e.g.Matha et al., 2013). In most cases, excitotoxicity is involved insecondary cell death subsequent to some initial trigger lesion.Indeed, glutamate has emerged as one of the main chemicals thatmediate secondary cell death after stroke (Benveniste et al., 1984;Simon et al., 1984) and trauma (e.g. Faden and Simon, 1988).
In the cochlea, connections between inner haircells and spiralganglion neurons are glutamatergic, allowing for fast information
ry, Neuroscience and Mentalniversity Ave., Toronto, M5G
transmission to the auditory brainstem. Damage to cochlear hair-cells by noise trauma, ototoxic drugs or direct physical damage canlead to a surge of glutamate release at the inner haircell synapseand perhaps lead to neuronal death. (Puel et al., 1998, 1995; Pujoland Puel, 1999). The abnormal levels of glutamate and injury dis-charges during uncontrolled cochlear afferent depolarization willresult in an acute phase of peripheral tinnitus (e.g. Eggermont,1990; Lenarz et al., 1993; Salvi et al., 2000; Zacharek et al., 2002).Few studies have been designed to explore the dynamics of thisearly period of abnormal excitatory activity.
It is well established that unilateral cochlear activation willcause suppression of cochlear activity in the contralateral ear viaolivocochlear efferent pathways (Siegel and Kim, 1982; Liberman,1989; Warren and Liberman, 1989; Guinan, 2006). This system ismost commonly revealed in contralateral suppression of otoa-coustic emissions (e.g. Liberman et al., 1996; Giraud et al., 1997;Mott et al., 1989; Puel and Rebillard, 1990; Collet et al., 1990;Moulin et al., 1993; Maison et al., 2000 James et al., 2005), but itis also manifest by cochlear afferent threshold changes (Liberman,1989; Warren and Liberman, 1989). In the present mouse model,we have monitored ABRs initiated from one ear, during and aftermechanical ablation of contralateral cochlea. By recording thetiming and amplitude of ABR threshold changes we have indirectlydetermined the time-course of cochlear excitotoxicity. The

J. Negandhi et al. / Hearing Research 315 (2014) 34e39 35
increased cochlear afferent activity levels caused by ear ablationwere confirmed by observation of increases in c-fos immuno-labelling of cells in some auditory brainstem and midbrain regions.
2. Material and methods
The animal species of choice was the CBA/J mouse (Mus mus-culus); we used a total of 26 young male adults (6e10 weeks olds).Functional (ABR) studies were carried out in 7 experimental sub-jects and 7 control animals. C-fos immuno-labelling studies weredone in a separate set of 8 experimental and 4 control mice. Allprocedures were carried out under anaesthesia (Ketamine, 150 mg/kg; xylazine, 10 mg/kg administered intraperitoneally). After in-duction, 50% maintenance doses were given every hour as needed.All procedures were approved by the animal care committee of theResearch Institute of the Hospital for Sick Children (Toronto) andfollowed guidelines set by the Canadian Council for Animal Care(CCAC).
2.1. Cochlear ablation
Unilateral cochlear ablation was achieved inserting a fine (30gauge) syringe needle along the axis of the external ear canal,through the tympanic membrane and physically penetrating thecochlea. The typical cochlear puncture site is shown (arrow symbol)in Fig. 1A. The cochlea was then flushed with sterile water tocomplete the lesioning process; cellular components of the organ ofCorti are damaged by a combination of physical disruption andosmotic damage. We have previously reported on this method forlocal cochlear ablation (Harrison et al., 1997).
2.2. ABR recordings
In 7 subjects, ABR measurements were made before during andafter unilateral cochlear ablation, specifically at 30 and 15 minbefore cochlear ablation, and then at 30 min intervals for 5 h.Further recordings were made at 24, 48, 120, and 240 h (10 days).An identical regime of recordings was made in 7 non-ablatedcontrol animals. Skin needle recording electrodes were placed ina vertex-mastoid configuration. Signal amplification (�1000) andaveraging (512 sweeps) was done with the Smart eEP with highfrequency transducers (Intelligent Hearing Systems Miani, Fl. USA).ABRs were recorded to stimuli presented to the non-ablated ear.ABR threshold levels were determined to a 4 kHz or 32 kHz tone pipstimuli presented in closed field at levels ranging from 10 to 80 dBSPL. Example ABR raw data waveforms are illustrated in Fig. 2. Allsignals were calibrated (PCB Piezotronics 377B02 condensermicrophone; Larson Davis level meter #831, B&K model 4230
Fig. 1. Scanning electron image of needle puncture (arrow) in the mid-apical turn of th
reference). Threshold levels were determined from ABR (P3)amplitude vs. intensity plots. Sampled ABR threshold valuesshowed normal distributions; statistical comparisons were madeusing Student's t-test.
2.3. C-fos labelling studies
Screening ABR recordings were made in both experimental andsham control animals to verify normal auditory thresholds. Allanimals were maintained in a low noise environment for manyhours prior to use. Experimental mice (N ¼ 8) were subjected tounilateral cochlea ablation and then kept in a sound proof (>40 dBattenuation; IAC) booth for one hour before histological study.Control mice (N ¼ 4) were also kept in this booth for one hour priorto study. Anesthetised mice underwent transcardial perfusion withsaline and 4% paraformaldehyde. After fixation the brain wasremoved and kept in 4% paraformaldehyde at 4 �C over-night.Sections (60 mm) were cut using a Vibrotome and stored in 0.02%sodium azide solution. Sections were washed in 0.3% hydrogenperoxide, PBS and blocking solution, and then incubated in primaryanti-c-fos antibody (1:10,000) at 4 �C for 48 h. Sections were againrinsed in PBS before incubation in biotinylated secondary antibody(Biotin-SP-conjugated affinipure Goat anti-rabbit IgG; 1:1000) for2 h. Samples are then washed again in PBS before incubation inavidin-biotin complex (ABC) solution for 1e2 h at room tempera-ture. Sections were incubated in DAB (3,3-Diamino benzidine) so-lution for 8 min before 0.3% hydrogen peroxide treatment to revealbrown pigmented c-fos labelled cells. Sections were then washed(PBS) and slide mounted. Labelled cells were identified and quan-tified in a semi-automated system (Image J software) usingthreshold intensity, shape and size parameters (Rieux et al., 2002).We have previously used these techniques in studies of neural ac-tivity patterns in the mouse (Harrison and Negandhi, 2012).
3. Results
3.1. Cochlear lesions
Our method of cochlear damage by needle puncture is illus-trated in the scanning electronmicrographs of Fig.1. Image A showsdamage (indicated by arrow) to the left cochlea with the needlepiercing the apical region of the cochlea. Image B is of the un-damaged (right) cochlea.
3.2. Functional ABR measures
The experimental protocol and a sample of ABR waveformsmeasured from stimulation (4 kHz tone) of the ear contralateral to
e left cochlea in the mouse (A) compared with the un-damaged right cochlea (B).

Fig. 2. Schematic showing example ABR waveforms (4 kHz stimulus) measured before and at time intervals (0.5, 2, and 240 h) after contralateral cochlear ablation. ABR waveformsevoked by stimuli at levels 0e80 dB SPL are shown and thresholds are indicated. Maximum threshold elevation is at c. 2 hrs post cochlear lesion. Thresholds are back to normal inthe long-term.
J. Negandhi et al. / Hearing Research 315 (2014) 34e3936
the ablated cochlea are illustrated in Fig. 2. The upper ABR data set(A) was recorded before cochlear ablation and indicates a thresholdof 30 dB SPL. Immediately after left cochlear ablation (plot B), rightear ABR threshold is elevated by 20 dBe50 dB SPL and remains atthat level at 2 h post ablation (plot C). At the final recording session,240 h (10 days) following unilateral ablation, ABR thresholds havereturned to 28 dB SPL. The early peaks of the ABR waveforms fullyrecover to the pre-injury amplitudes, however (as can be noted bycomparing plot A and D in Fig. 2) the longer latency waveforms of
Fig. 3. The evolution of ABR threshold changes after contralateral cochlear ablation. Filled sbefore and after unilateral cochlea ablation of left ear. Open symbols show data from normelevation reaching a maximum within minutes. Other animals (panel B; n ¼ 3) showed aindicate standard deviations. Statistical comparisons (Student's t-test) between experimentindicated.
the ABR do not totally recover to their initial amplitudes. Thispossibly reflects some longer lasting effects on auditory areas morecentral than the cochlear nerve and nucleus.
The graphs of Fig. 3 plot average ABR threshold changes (4 kHzstimulus) from an initial level measured 30 min before, through to10 days post unilateral cochlear ablation. ABR threshold from twosets of experimental animals are shown (filled symbols) comparedto normal control subjects (followed for the same time with nointerventions; N ¼ 7; open symbols). We observe two patterns of
ymbols show changes in ABR thresholds measured with 4 kHz stimuli in the right earal control subjects (n ¼ 7). In panel A, subjects (n ¼ 4) showed a rapid ABR thresholdmore gradual change in ABR threshold after contralateral cochlear ablation. Error barsal and control subjects were made from averages of the data within the time windows
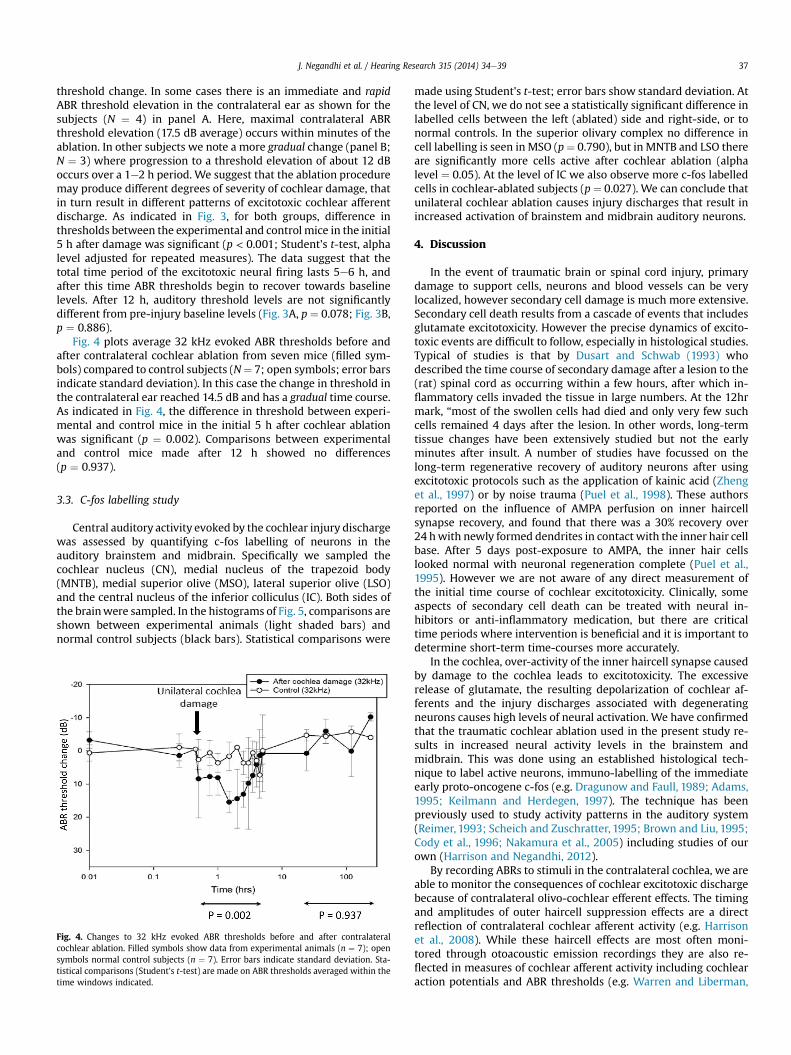
J. Negandhi et al. / Hearing Research 315 (2014) 34e39 37
threshold change. In some cases there is an immediate and rapidABR threshold elevation in the contralateral ear as shown for thesubjects (N ¼ 4) in panel A. Here, maximal contralateral ABRthreshold elevation (17.5 dB average) occurs within minutes of theablation. In other subjects we note a more gradual change (panel B;N ¼ 3) where progression to a threshold elevation of about 12 dBoccurs over a 1e2 h period. We suggest that the ablation proceduremay produce different degrees of severity of cochlear damage, thatin turn result in different patterns of excitotoxic cochlear afferentdischarge. As indicated in Fig. 3, for both groups, difference inthresholds between the experimental and control mice in the initial5 h after damage was significant (p < 0.001; Student's t-test, alphalevel adjusted for repeated measures). The data suggest that thetotal time period of the excitotoxic neural firing lasts 5e6 h, andafter this time ABR thresholds begin to recover towards baselinelevels. After 12 h, auditory threshold levels are not significantlydifferent from pre-injury baseline levels (Fig. 3A, p ¼ 0.078; Fig. 3B,p ¼ 0.886).
Fig. 4 plots average 32 kHz evoked ABR thresholds before andafter contralateral cochlear ablation from seven mice (filled sym-bols) compared to control subjects (N¼ 7; open symbols; error barsindicate standard deviation). In this case the change in threshold inthe contralateral ear reached 14.5 dB and has a gradual time course.As indicated in Fig. 4, the difference in threshold between experi-mental and control mice in the initial 5 h after cochlear ablationwas significant (p ¼ 0.002). Comparisons between experimentaland control mice made after 12 h showed no differences(p ¼ 0.937).
3.3. C-fos labelling study
Central auditory activity evoked by the cochlear injury dischargewas assessed by quantifying c-fos labelling of neurons in theauditory brainstem and midbrain. Specifically we sampled thecochlear nucleus (CN), medial nucleus of the trapezoid body(MNTB), medial superior olive (MSO), lateral superior olive (LSO)and the central nucleus of the inferior colliculus (IC). Both sides ofthe brainwere sampled. In the histograms of Fig. 5, comparisons areshown between experimental animals (light shaded bars) andnormal control subjects (black bars). Statistical comparisons were
Fig. 4. Changes to 32 kHz evoked ABR thresholds before and after contralateralcochlear ablation. Filled symbols show data from experimental animals (n ¼ 7); opensymbols normal control subjects (n ¼ 7). Error bars indicate standard deviation. Sta-tistical comparisons (Student's t-test) are made on ABR thresholds averaged within thetime windows indicated.
made using Student's t-test; error bars show standard deviation. Atthe level of CN, we do not see a statistically significant difference inlabelled cells between the left (ablated) side and right-side, or tonormal controls. In the superior olivary complex no difference incell labelling is seen in MSO (p¼ 0.790), but in MNTB and LSO thereare significantly more cells active after cochlear ablation (alphalevel ¼ 0.05). At the level of IC we also observe more c-fos labelledcells in cochlear-ablated subjects (p ¼ 0.027). We can conclude thatunilateral cochlear ablation causes injury discharges that result inincreased activation of brainstem and midbrain auditory neurons.
4. Discussion
In the event of traumatic brain or spinal cord injury, primarydamage to support cells, neurons and blood vessels can be verylocalized, however secondary cell damage is much more extensive.Secondary cell death results from a cascade of events that includesglutamate excitotoxicity. However the precise dynamics of excito-toxic events are difficult to follow, especially in histological studies.Typical of studies is that by Dusart and Schwab (1993) whodescribed the time course of secondary damage after a lesion to the(rat) spinal cord as occurring within a few hours, after which in-flammatory cells invaded the tissue in large numbers. At the 12hrmark, “most of the swollen cells had died and only very few suchcells remained 4 days after the lesion. In other words, long-termtissue changes have been extensively studied but not the earlyminutes after insult. A number of studies have focussed on thelong-term regenerative recovery of auditory neurons after usingexcitotoxic protocols such as the application of kainic acid (Zhenget al., 1997) or by noise trauma (Puel et al., 1998). These authorsreported on the influence of AMPA perfusion on inner haircellsynapse recovery, and found that there was a 30% recovery over24 hwith newly formed dendrites in contact with the inner hair cellbase. After 5 days post-exposure to AMPA, the inner hair cellslooked normal with neuronal regeneration complete (Puel et al.,1995). However we are not aware of any direct measurement ofthe initial time course of cochlear excitotoxicity. Clinically, someaspects of secondary cell death can be treated with neural in-hibitors or anti-inflammatory medication, but there are criticaltime periods where intervention is beneficial and it is important todetermine short-term time-courses more accurately.
In the cochlea, over-activity of the inner haircell synapse causedby damage to the cochlea leads to excitotoxicity. The excessiverelease of glutamate, the resulting depolarization of cochlear af-ferents and the injury discharges associated with degeneratingneurons causes high levels of neural activation. We have confirmedthat the traumatic cochlear ablation used in the present study re-sults in increased neural activity levels in the brainstem andmidbrain. This was done using an established histological tech-nique to label active neurons, immuno-labelling of the immediateearly proto-oncogene c-fos (e.g. Dragunow and Faull, 1989; Adams,1995; Keilmann and Herdegen, 1997). The technique has beenpreviously used to study activity patterns in the auditory system(Reimer, 1993; Scheich and Zuschratter, 1995; Brown and Liu, 1995;Cody et al., 1996; Nakamura et al., 2005) including studies of ourown (Harrison and Negandhi, 2012).
By recording ABRs to stimuli in the contralateral cochlea, we areable to monitor the consequences of cochlear excitotoxic dischargebecause of contralateral olivo-cochlear efferent effects. The timingand amplitudes of outer haircell suppression effects are a directreflection of contralateral cochlear afferent activity (e.g. Harrisonet al., 2008). While these haircell effects are most often moni-tored through otoacoustic emission recordings they are also re-flected in measures of cochlear afferent activity including cochlearaction potentials and ABR thresholds (e.g. Warren and Liberman,

Fig. 5. Graphs comparing the number of c-fos active cells between experimental animals after unilateral cochlear ablation and normal controls. Measures were made at the level ofthe cochlear nucleus (CN), medial nucleus of the trapezoid body (MNTB), medial superior olivary nucleus (MSO), lateral superior olivary nucleus (LSO) and the central nucleus ofinferior colliculus (IC). Error bars indicate standard deviation; p values (Student's t e test) are indicated.
J. Negandhi et al. / Hearing Research 315 (2014) 34e3938
1989; Liberman,1989). Larsen and Liberman (2010) using a conceptsimilar to our methodology here could not find evidence of acontralateral, olivocochlear mediated effect to monitor excitotox-icity. The main difference between their study and the presentwork is the timeline of acquiring ABR data. Larsen and Libermanmonitored contralateral effects at 2, 4, 8 and 16 days post exposure,whereas we monitor events over minutes/hours instead of days. Asshown in Figs. 3 and 4 we see ABR threshold changes immediatelyafter cochlear ablation. By 24 h, ABR thresholds have returned tocontrol levels and any subsequent monitoring post exposure willnot show changes in thresholds.
The contralateral effects that we observe after unilateralcochlear ablation could be related to changes in middle ear muscletone. We are well aware of this possibility, and have in anotherspecies (chinchilla) shownhow themiddle ear reflex can contributeto contralateral OAE recordings (Wolter et al., 2014). In the presentstudy we have not been able to control for middle ear effects;contributions from this source is a possibility.
We have described here the degree and time course of theexcitotoxic injury discharge. We note that the time course canreveal an immediate and rapid effect contralaterally (Fig. 3A) andwe suggest that this reflects a local and more severe cochlear lesionthan for subjects showing a more gradual ABR threshold change.Our method used to inflict unilateral cochlea damage results in
puncture of the low frequency mid-to-apical region of the cochlea(Fig. 1) where lower frequency regions are found. This may explainthe larger shifts and the immediate drop in 4 kHz evoked ABRthresholds compared to higher frequency (32 kHz) ABR thresholdchanges which are smaller and more gradual (Fig. 4). In regard tolonger-term effects of unilateral cochlear ablation on the oppositeear the ABR thresholds completely return to normal levels. We alsonote in Fig. 3A the differences between experimental and controlABR thresholds after 12 h, whilst not significant (p ¼ 0.078) show atrend towards improved thresholds.
By c-fos labelling, increases in active neurons in some areas ofthe auditory pathway could be determined after unilateral cochlearablation, confirming that ablation induces cochlear nerve injurydischarge. Whilst we observe significant changes (in c-fos labelledcells) in MNTB, LOS and IC there is no significant change at the levelof cochlear nucleus (Fig. 5). We suggest that under normal condi-tions there is already a high number of activated neurons becauseof high levels of spontaneous activity in primary cochlear afferents,and that additional injury discharge does not newly activate morecells.
Hyperactivity or increased spontaneous activity has previouslybeen noted in the dorsal cochlear nucleus of the rat and hamsterafter intense sound exposure and has been discussed in thatcontext as the basis for noise-induced tinnitus (Zhang and

J. Negandhi et al. / Hearing Research 315 (2014) 34e39 39
Kaltenbach, 1998; Zacharek et al., 2002). Whilst we see only a trendin CN activity increase, we do note increased cell activation inMNTB, LSO and IC immediately after cochlear ablation. It is parsi-monious to suggest that this general augmentation of auditoryneuron firing will give rise to persisting auditory percepts that, inhumans, would be called tinnitus. Such immediate, peripherallygenerated sensations are often reported as tinnitus after acoustictrauma, and generally subside within the time course of the exci-totoxic effects that we describe here. In general hyperactivity in theauditory pathway has also been linked to tinnitus, however theprimary region of generation is often debated. Some authorsbelieve the main source is at the peripheral level, others haveproposed a more central generation. The consensus is that imme-diate effects are peripheral, but these can lead at a much later stage,to central generation mechanisms (for discussion see: Eggermont,1990; Lenarz et al., 1993; Salvi et al., 2000; Zacharek et al., 2002;Guitton et al., 2003; Eggermont and Roberts, 2004; Mulders andRobertson, 2009; Eggermont, 2012).
In conclusion, in a mouse model we show that unilateralcochlear damage results in a temporary (5e6 h) effect on contra-lateral auditory thresholds. This reflects a cochlear excitotoxicityeffect that evokes a cochlear afferent (injury) discharge for this timeperiod. In some cases the injury discharge is initially very high, inother conditions the injury discharge appears to increase over a 2 htime period. We suggest that monitoring of contralateral effects,whilst indirect, provides an accurate evaluation of the dynamics ofcochlear excitotoxicity. We suggest that the excitotoxicity timecourse revealed in this mousemodel may have some bearing on theduration of acute tinnitus suffered in human subjects after acoustictrauma.
Acknowledgements
This research was funded by the Canadian Institutes of HealthResearch (CIHR).
References
Adams, J.C., 1995. Sound stimulation induces Fos-related antigens in cells withcommon morphological properties throughout the auditory brainstem. J. Comp.Neurol. 361, 645e668.
Benveniste, H., Drejer, J., Schousboe, A., Diemer, N.H., 1984. Elevation of the extra-cellular concentration of glutamate and aspartate in rat hippocampus duringtransient cerebral ischemia monitored by intracerebral microdialysis.J. Neurochem. 43, 1369e1374.
Brown, M.C., Liu, T.S., 1995. Fos-like immunoreactivity in central auditory neuronsof the mouse. J. Comp. Neurol. 357, 85e97.
Choi, D.W., Rothman, S.M., 1990. The role of glutamate neurotoxicity in hypo-xicischemic neuronal death. Annu. Rev. Neurosci. 13, 171e182.
Cody, A.R., Wilson, W., Leah, J., 1996. Acoustically activated c-fos expression inauditory nuclei of the anaesthetised guinea pig. Brain Res. 728, 72e78.
Collet, L., Kemp, D.T., Veuillet, E., Duclaux, R., Moulin, A., Morgon, A., 1990. Effect ofcontralateral auditory stimuli on active cochlear micromechanical properties inhuman subjects. Hear Res. 43, 251e261.
Dragunow, M., Faull, R., 1989. The use of c-fos as a metabolic marker in neuronalpathway tracing. J. Neurosci. Methods 29, 261e265.
Dusart, I., Schwab, M.E., 1993. Secondary cell death and the inflammatory reactionafter dorsal hemisection of the rat spinal cord. Eur. J. Neurosci. 6, 712e724.
Eggermont, J.J., 1990. On the pathophysiology of tinnitus; a review and a peripheralmodel. Hear Res. 48, 111e123.
Eggermont, J.J., 2012. The Neuroscience of Tinnitus. Oxford University Press, UK.Eggermont, J.J., Roberts, L.E., 2004. The neuroscience of tinnitus. Trends Neurosci.
27, 676e682.Faden, A.I., Simon, R.P., 1988. A potential role of excitotoxins in the pathophysiology
of spinal cord injury. Ann. Neurol. 23, 623e626.Giraud, A.L., Wable, J., Chays, A., Collet, L., Chery-Croze, S., 1997. Influence of
contralateral noise on distortion product latency in humans: is the medialolivocochlear efferent system involved? J. Acoust. Soc. Am. 102, 2219e2227.
Guinan Jr., J.J., 2006. Olivocochlear efferents: anatomy, physiology, function, and themeasurement of efferent effects in humans. Ear Hear. 27, 589e607.
Guitton, M.J., Caston, J., Ruel, J., Johnson, R.M., Pujol, R., Puel, J.L., 2003. Salicylateinduces tinnitus through activation of cochlea NMDA receptors. J. Neurosci. 23,3944e3952.
Harrison, R.V., Negandhi, J., 2012. Resting neural activity patterns in auditorybrainstem and midbrain in conductive hearing loss. Acta Otolaryngol. 132,409e414.
Harrison, R.V., Hirakawa, H., Mount, R.J., 1997. Total ablation of cochlear haircells byperilymphatic perfusion with water. Hear Res. 110, 229e233.
Harrison, R.V., Sharma, A., Brown, T., Jiwani, S., James, A.L., 2008. Amplitude mod-ulation of DPOAEs by acoustic stimulation of the contralateral ear. Acta Oto-laryngol. 128, 404e407.
James, A.L., Harrison, R.V., Pienkowski, M., Dajani, H.R., Mount, R.J., 2005. Dynamicsof real time DPOAE contralateral suppression in chinchillas and humans. Int. J.Audiol. 44, 118e129.
Keilmann, A., Herdegen, T., 1997. The c-Fos transcription factor in the auditorypathway of the juvenile rat: effects of acoustic deprivation and repetitivestimulation. Brain Res. 753, 291e298.
Larsen, E., Liberman, C.M., 2010. Contralateral cochlear effects of ipsilateral damage:no evidence for interaural coupling. Hear Res. 260 (1e2), 70. http://dx.doi.org/10.1016/j.heares.2009.11.011.
Lenarz, T., Schreiner, C., Snyder, R.L., Ernst, A., 1993. Neural mechanisms of tinnitus.Eur. Arch. Otorhinolaryngol. 249, 441e446.
Liberman, M.C., 1989. Rapid assessment of sound-evoked olivocochlear feedback:suppression of compound action potentials by contralateral sound. Hear Res.38, 47e56.
Liberman, M.C., Puria, S., Guinan Jr., J.J., 1996. The ipsilaterally evoked olivocochlearreflex causes rapid adaptation of the 2f1-f2 distortion product otoacousticemission. J. Acoust. Soc. Am. 99, 3572e3584.
Maison, S., Micheyl, C., Andeol, G., Gallego, S., Collet, L., 2000. Activation of medialolivocochlear efferent system in humans: influence of stimulus bandwidth.Hear Res. 140, 111e125.
Matha, A., Prabhakar, M., Kumar, P., Deshmukh, R., Sharma, P.L., 2013. Excitotoxicity:bridge to various triggers in neurodegenerative disorders. Eur. J. Pharmacol.698, 6e18.
Mayer, M.L., Westbrook, G.L., 1987. Cellular mechanisms underlaying excitotoxicity.Trends Neurosci. 10, 59e61.
Mott, J.B., Norton, S.J., Neely, S.T., Warr, W.B., 1989. Changes in spontaneous otoa-coustic emissions produced by acoustic stimulation of the contralateral ear.Hear Res. 38, 229e242.
Moulin, A., Collet, L., Duclaux, R., 1993. Contralateral auditory stimulation altersacoustic distortion products in humans. Hear Res. 65, 193e210.
Mulders, W., Robertson, D., 2009. Hyperactivity in the auditory midbrain afteracoustic trauma: dependence on cochlear activty. Neuroscience 164,33e746.
Nakamura, M., Rosahl, S.K., Alkahlout, E., Walter, G.F., Samii, M.M., 2005. Electricalstimulation of the cochlear nerve in rats: analysis of c-fos expression in audi-tory brainstem nuclei. Brain Res. 1031, 39e55.
Puel, J.L., Rebillard, G., 1990. Effect of contralateral sound stimulation on thedistortion product 2f1-f2: evidence that the medial efferent system is involved.J. Acoust. Soc. Am. 87, 1630e1635.
Puel, J.L., Safieddine, S., Gervais D'Aldin, C., Eybalin, M., Pujol, R., 1995. Synapticregeneration and functional recovery after excitotoxic injury in the guinea pigcochlea. CR Acad. Sci. 318, 67e75.
Puel, J.L., Ruel, J., Gervais D'Aldin, C., Pujol, R., 1998. Excitotoxicity and repair ofcochlear synapses after noise trauma. Neuro Rep. 9, 2109e2114.
Pujol, R., Puel, J.L., 1999. Excitotoxicity, synaptic repair and functional recovery inthe mammalian cochlea: a review of recent findings. Ann. N. Y. Acad. Sci. 884,249e254.
Reimer, K., 1993. Simultaneous demonstration of fos-like immunoreactivity and 2-deoxy-glucose uptake in the inferior colliculus of the mouse. Brain Res. 616,339e343.
Salvi, R.J., Wang, J., Ding, D., 2000. Auditory plasticity and hyperactivity followingcochlear damage. Hear Res. 147, 261e274.
Scheich, H., Zuschratter, W., 1995. Mapping of stimulus features and meaning ingerbil auditory cortex with 2-deoxyglucose and c-fos antibodies. Behav. BrainRes. 66, 195e205.
Siegel, J.H., Kim, D.O., 1982. Efferent neural control of cochlear mechanics? Olivo-cochlear bundle stimulation affects cochlear biomechanical nonlinearity. HearRes. 6, 171e182.
Simon, R.P., Swan, J.H., Griffiths, T., Meldrum, B.S., 1984. Blockade of N-methyl-D-aspartate receptors may protect against ischemic damage in the brain. Science226, 850e852.
Warren, E.H.D., Liberman, M.C., 1989. Effects of contralateral sound onauditory-nerve responses. I. Contributions of cochlear efferents. Hear Res.37, 89e104.
Wolter, N.E., Harrison, R.V., James, A.L., 2014. Separating the contributions of oli-vocochlear and middle ear muscle reflexes in modulation of distortion productotoacoustic emission levels. Audiol. Neurootol. 19, 41e48.
Zacharek, M.A., Kaltenbach, J.A., Mathog, T.A., Zang, J., 2002. Effects of cochlearablation on noise induced hyperactivity in the hamster dorsal cochlear nucleus:implications for the origin of noise induced tinnitus. Hear Res. 172, 137e143.
Zhang, J.S., Kaltenbach, J.A., 1998. Increases in spontaneous activity in the dorsalcochlear nucleus of the rat following exposure to high-intensity sound. Neu-rosci. Lett. 250, 197e200.
Zheng, X.Y., Henderson, D., Hu, B.H., McFadden, S.L., 1997. Reconvery of structureand function of inner ear afferent synapses following kainic acid excitotoxicity.Hear Res. 105, 65e76.
Copyright © 2022 FDOKUMEN




















