
The_abundance_and_diversity_of.pdf - UCL Discovery
267
The abundance and diversity of desert invertebrates in Abu Dhabi and their roie in the diet of the houbara bustard Chlamydotis undulata macqueeni Barbara Jane Tigar Doctor of Philosophy (PhD) University of London 1998 BIBL [LOUDON] UNIV.
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of The_abundance_and_diversity_of.pdf - UCL Discovery

The abundance and diversity of desert invertebrates in Abu Dhabi and their roie in the diet
of the houbara bustard Chlamydotis undulatamacqueeni
Barbara Jane Tigar
Doctor of Philosophy (PhD) University of London 1998
BIBL [LOUDON]
UNIV.

ProQuest Number: 10014434
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest 10014434
Published by ProQuest LLC(2016). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code.
Microform Edition © ProQuest LLC.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

A b str a c t
Invertebrate abundance and diversity were monitored for two years at five sites in
Abu Dhabi, and houbara bustard faecal and gizzard contents were examined to
establish the role of invertebrates in the diet.
Literature on houbara diet was used to calculate a relative citation index for known
foods. Invertebrates, particularly Coleoptera, Orthoptera and Formicidae, scored
highest; Tenebrionidae were especially important. All houbara subspecies ate plants
from the families: Gramineae, Leguminosae, Cruciferae, Compositae and
Solanaceae. African and Arabian houbara also ate Zygophyllaceae and
Chenopodiaceae. Canary Island birds may have a narrower diet than mainland
houbara.
Overnight pitfall and light-traps captured 143,397 invertebrates, including 196 new
records and ten new species. Pitfalls frequently contained: Formicidae (75.4%),
Thysanura (12%), Coleoptera (8.4%), Araneae (1.5%) and Scorpiones (1.1%). Light
traps had high catches of Lepidoptera (56.4%) and Coleoptera (19.1%); Coleoptera
contributed the highest biomass (42.6%). Fewer invertebrates were caught in winter
than in summer, and catches were positively correlated with temperature. There
were significant differences in community composition between sites and
substrates. Isoptera, Tettigoniidae and Carabidae were rarely captured inland, but
more taxa occurred near the coast. Pitfalls captured higher numbers of more
diverse predaceous arthropods at new moons than at full moons, suggesting
differences in invertebrate predation risk or visual awareness of traps.
Similar food remains were recovered from houbara faeces and gizzards. Trials
established a calibration for the recovery of prey remains in faeces. Formicidae
were the most frequently consumed prey, but Tenebrionidae formed the highest
biomass. Plants made a low but frequent contribution to the diet. Estimated Active
Metabolic Rate suggested that invertebrates contributed 89-94.6% to houbara
energy needs, and the frequency of invertebrates in faeces showed good rank
agreement with pitfall catches.
Desert invertebrates showed spatial and temporal variation and over-wintering
houbara appeared to be non-selective, consuming mainly locally available, ground- dwelling invertebrates.

T a b le o f C o n te n ts
PageAbstract 2Table of contents 3List of tables 7List of figures 10List of plates 11List of appendices 12Acknowledgements 13
C h a p te r 1 - General introduction 15
Synopsis of Chapter 1 151.1 Identifying the problem: rationale behind the research 161.2 Abu Dhabi and the United Arab Emirates 17
Introduction 17Geology 19Climate 20Ecology 21Invertebrates 22
1.3 Introduction to the bustards - family Otididae 261.4 The houbara bustard 27
Subspecies and their ranges 27Houbara habitat 29Ecology of Chlamydotis undulata macqueeni 29Conservation and status of the houbara 30
C h a p t e r 2 -The diet of the houbara bustard - a review 32
Synopsis of Chapter 2 322.1 Introduction 332.2 The review of the diet 33
2.2.1 The Canary Island houbara: Chlamydotis undulata fuertaventurae 33General descriptions of the diet 33Animal foods 45Plant foods 45Studies based on faecal analysis 45Feeding behaviour of C.u.fuertaventurae 47
2.2.2 The North African houbara: Chlamydotis undulata undulata 47General descriptions of the diet 47Gizzard content analysis 48
2.2.3 MacQueen’s bustard: Chlamydotis undulata macqueeni 49Middle Eastern population 49
Studies based on faecal analysis 50Pakistani and Indian populations 50
General description of the diet 50Plant foods 51Studies based on gizzard contents 52
Central Asian population 53General descriptions of the diet 53Plant foods 54Studies based on gizzard contents 54Studies based on faecal analysis 54

C h a p te r 2 -The diet of the houbara bustard - a review (continued)
2.2 The review of the diet (continued)2.2.4 Vagrant houbara 562.2.5 Gizzard stones 572.2.6 The diet of semi-captive houbara 572.2.7 Diets for captive houbara 57
2.3 Discussion 602.3.1 Relative merits of techniques used to study the diet 602.3.2 A comparison of the diet of the three houbara subspecies 61
Citation indices for plant foods 64Citation indices for animal foods 65
2.3.3 Seasonal trends in the houbara's diet 66Seasonal consumption of plant foods 66Seasonal consumption of animal foods 67
2.3.4 Implications for the management of habitat and captive birds 682.4 Conclusion 69
C h a p te r 3 - Pilot studies for monitoring desert invertebrates 70
Synopsis of Chapter 3 703.1 General Introduction 713.2 Pitfall trapping 71
3.2.1 Introduction to pitfall trials 713.2.2 Methods 72
The trapping-time trial 72The transect length trial 72
3.2.3 Results 73The trapping-time trial 73The transect length trial 74Species-effort curve 76
3.2.4 Discussion 783.3 The use of refuge boards 80
3.3.1 Introduction 803.3.2 Methods 803.3.3 Results 803.3.4 Discussion 81
3.4 Sweep-netting and beating trays 813.4.1 Introduction and methods 813.4.2 Results 823.4.3 Discussion 83
3.5 Walking transects for Orthoptera 833.5.1 Introduction and methods 833.5.2 Results 833.5.3 Discussion 84
3.6 Flight interception traps 843.6.1 Introduction and methods 843.6.2 Results 843.6.3 Discussion 84
3.7 The use of light traps for the capture of nocturnal insects. 863.8 General discussion and recommendations for trapping methods 86

C h a p te r 4 - The diversity and abundance of invertebrates in Abu Dhabi's 88 deserts
Synopsis of Chapter 4 884.1 Patterns of abundance and diversity of ground dwelling arthropods 89
4.1.1 Introduction 894.1.2 Methods 90
Trapping sites 90Pitfall trapping 90Climatic data 93Data analysis 93
4.1.3 Results 95The arthropod community 95Temporal variation in total catch 95Spatial variation in total catch 98Spatial variation in taxonomic diversity 100
4.1.4 Discussion 102Community composition 102Temporal variation 104Spatial-temporal variation 104Spatial variation 105Conclusion 106
4.2 Does the phase of the moon influence invertebrate trap catch? 1074.2.1 Introduction 1074.2.2 Methods 107
Invertebrate trapping 107Data analysis 107
4.2.3 Results 109The arthropod community 109Variation in arthropod abundance 111Variation in taxonomic diversity 111Species composition according to the phase of the moon and 113substrate
4.2.4 Discussion 1164.3 Patterns of available biomass and diversity of flying invertebrates 119
4.3.1 Introduction 1194.3.2 Methods 119
The light trap 119Data analysis 119
4.3.3 Results 120Variations in the number of flying insects 121Variations in insect diversity between sites 124
4.3.4 Discussion 1264.4 General discussion on Abu Dhabi’s invertebrates 129
The invertebrate community 129Temporal patterns 130Spatial patterns 131Implications for biodiversity studies 131

C h a p te r 5 - The diet of the houbara bustard in Abu Dhabi 133
Synopsis of Chapter 5 1335.1 Introduction 1345.2 Quantifying the diet via faecal analysis 134
5.2.1 Introduction 1345.2.2 Methods 135
Calibration experiments with animal prey 135Faecal sample collection from wild birds 137Faecal analysis 137Dietary calculations 138
5.2.3 Results 139Calibration experiments with animal foods 139Faecal analysis 142
5.2.4 Discussion 148The relative merits of different dietary calculations and indices 148Identification of key fragments 151
5.3 Analysis of houbara gizzards from Abu Dhabi 1535.3.1 Introduction 1535.3.2 Methods 1535.3.3 Results 1535.3.4 Discussion 155
5.4 General discussion of the diet 1575.4.1 Description of houbara diet in Abu Dhabi 1575.4.2 Comparison with other studies on houbara diet 1585.4.3 Measurement of the relative proportions of food types 1595.4.4 Nutritional and energetic implications 160
C h a p t e r 6 - General discussion 162
Synopsis of Chapter 6 1626.1 The ecology of arid-zone invertebrates 163
Seasonal abundance of invertebrates 163Distribution of desert invertebrates 164Diversity 165
6.2 The desert food web 1676.3 Studying houbara and other avian diets 1706.4 Houbara diet in DAE 1726.5 Conservation of houbara and wildlife in UAE 1746.6 Implications for captive breeding programmes 1756.7 Conclusions 176
Epilogue 177
R e f e r e n c e s 178

L ist o f Ta b le s
Table 2.1 List of plants eaten by the three subspecies of houbara as 34
vegetative material
Table 2.2 List of flowers, fruit and seeds eaten by the three subspecies 37
of houbara
Table 2.3 List of invertebrates eaten by the three subspecies of 40
houbara
Table 2.4 List of vertebrates eaten by the three subspecies of houbara 44
Table 2.5 Summary of hogbara gizzard contents from Central Asia 55
Table 2.6 Summary of foods eaten by vagrant houbara 56
Table 2.7 Summary of observational and other data on the diet of 58
captive and semi-captive houbara
Table 2.8 Summary of diets given to captive houbara 59
Table 2.9 The number of citations and citation index of plant foods for 62
the three houbara subspecies
Table 2.10 The number of citations and citation index of animal foods for 63
the three houbara subspecies
Table 3.1 Numbers of invertebrates and species caught by three pitfall 73
transects in place for 14, 24 and 32 hours
Table 3.2 Percentage recovery of refuge boards after 28 days in the 81
desert
Table 3.3 Number of invertebrates and taxa caught along twelve 200 m 82sweep-net transects
Table 3.4 Insects caught by a Malaise trap at Sweihan Research 85
Station
Table 4.1 Site characteristics and summary statistics for the arthropod 92
communities
Table 4.2 Composition of the 53,396 arthropods caught by pitfall- 94
trapping over two years in Abu Dhabi
Table 4.3 ANOVA F statistics for differences in the log catch of various 96
arthropod taxa at different sites, on different substrates and
for different sampling periods

L is t o f T a b le s (continued)
Table 4.4 Pearson correlations between the minimum soil temperature 98
and log catch of various arthropod groups at five desert
locations
Table 4.5 Arthropod community statistics for the five desert sites split 101by substrate
Table 4.6 Percentage composition of the sand and gravel communities 103at the five desert sites, reduced to 14 taxa for simplicity
Table 4.7 Summary of the number of arthropod taxa captured in pitfall 109
trap, grouped by phase of the moon and by substrate
Table 4.8 Total list of taxa caught in pitfall traps around full and new 110moons, shown according to biological groups
Table 4.9 F statistics for differences in log total invertebrates, 111predaceous arthropods, macroarthropod detritivores,
herbivorous insects and social insects captured in pitfall traps
at different phases of the moon, substrates and their
interaction
Table 4.10 F statistics for differences in log number of taxa for total 112invertebrates, predaceous arthropods, macroarthropod
detritivores, herbivorous insects and social insects captured
in pitfall traps at different phases of the moon, substrates and
their interaction
Table 4.11 Varimax rotated component loadings from the PCA, 114calculated from the covariance matrix
Table 4.12 Summary of total light trap catch by number and biomass 121
over two years
Table 4.13 F statistics for differences in log total invertebrates caught in 123a light trap at different sites and at different minimum air
temperatures
Table 4.14 F statistics for differences in number of orders of insects and 125the number of coleopteran families caught in a light trap at
different sites
Table 5.1 Functional groups of potential animal prey of houbara 136bustards in UAE

L is t o f T a b le s (continued)
Table 5.2 Mean and standard error (SE) of the recovery rate for prey 140
remains in faeces with key fragments in bold
Table 5.3 The correction factor and recovery rate for fragments 141
recovered for group 6 (large nocturnal tenebrionids)
Table 5.4 Estimated percentage of invertebrate matter of houbara 142
faeces from Abu Dhabi
Table 5.5 Plant remains identified from wild houbara faeces, listed as 143
percentage frequency by faeces and by track
Table 5.6 Percentage frequency of each prey group by faeces and by 144
track with prey identity where known
Table 5.7 Comparison of three methods of faecal analysis, with 145
estimates of the percentage contribution to biomass, fat and
protein
Table 5.8 Calorific value of invertebrate diet corrected for metabolizable 146
energy content per average faecal sample for wild houbara
over-wintering in Abu Dhabi
Table 5.9 Calculated values for typical daily diet, percentage frequency 147
of prey in pitfall traps and the estimate daily foraging distance
for houbara
Table 5.10 List of animal and plant foods from houbara faeces and 154
gizzards
Table 5.11 The percentage contribution of animal prey by group from 155
gizzard samples and estimated energy value with
comparative values for faecal samples
Table 5.12 Score for percentage invertebrate material and identity of 155
plant material present in gizzard contents
Table 6.1 Trophic relationships in Abu Dhabi (excluding houbara) 168

10
L ist o f F ig u r es
Figure 1.1 Map of Abu Dhabi and the United Arab Emirates showing the 18
five study sites
Figure 3.1 Mean catch of total invertebrates, Coleoptera and 75
Hymenoptera caught during 50 simulations of trap catch
Figure 3.2 The overall mean catch of beetles for line 1 against number 76
of traps
Figure 3.3 The cumulative number of species caught along three lines of 77
60 pitfall traps
Figure 4.1 Relationship between the average catches at the sites and 97
climatic conditions
Figure 4.2 Differences in log catch at the five sites (Fisher’s LSD test) 99
Figure 4.3 Cluster analysis of the sand and gravel communities at five 101
desert sites
Figure 4.4 The relationship between log number of taxa of predaceous 113
arthropods vs. sample number for catches at full and new
moons
Figure 4.5 PCA ordination of the invertebrate communities grouped by 115substrate and phase of the moon
Figure 4.6 Summary of invertebrates caught in the Heath trap by order 120
Figure 4.7 Annual cycle of the mean trap catch per calendar month 122
across all sites from two years of trapping, and the mean
minimum air temperature
Figure 4.8 Plot of mean and standard error of the light trap at five sites 123
over 24 sampling periods
Figure 4.9 Differences in log catch at the five sites 124
Figure 4.10 Plot of mean value for the number of insect orders recorded 125
by site
Figure 4.11 Differences in number of coleopteran families caught at the 126
five sites
Figure 4.12 Plot of mean value for the number of coleopteran families 126
recorded by site
Figure 5.1 Mean ratio of recovery rates for fragments from group 6 139
Figure 5.2 Available invertebrate prey and relative effort required to 148
catch them during the winter months in Abu Dhabi

L is t o f P la tes
11
Plate 1.1 The houbara bustard Chlamydotis undulata macqueeni 28
Plate 4.1 The Baynunah study site 91
Plate 4.2 The Medinet Zayed study site 91
Plate 4.3 The Public Hunting Triangle study site 91
Plate 4.4 The Khatam study site 91
Plate 4.5 The Urn Az Zimul study site 91

12
L ist o f A p p e n d ic e s
Appendix 1.1
Appendix 1.2
Appendix 4.1
Appendix 5.1
Appendix 5.2
Appendix 5.3
Appendix 5.4
Appendix 5.5
A preliminary assessment of the arthropods of Abu 205
Dhabi (Tigar 1996a)
Terrestrial Arthropods (excluding insects) (Tigar 1996b) 230
List of species caught in UAE (1992-1996) 243
List of feeding calibration trails 250
A preliminary study of the passage time of digesta of 251
the houbara (Tigar 1995)
Description of invertebrate remains recovered from 259
faecal samples
Photographs of typical fragments from the invertebrate 260
prey groups
List of faecal samples collected in UAE 264

13
Ac k n o w le d g e m e n ts
I thank HH Sheikh Khalifa Bin Zayed and HH Sheikh Mohammed Bin Zayed for
funding research at the National Avian Research Center (NARC), and the
Managing Director, Mr Mohammed AI Bowardi, for his support. I am also grateful
to Paul Goriup, former Chairman of the International Advisory Committee, for
guiding NARC towards bustard ecology and conservation. I also thank him for
sharing his extensive collection of bustard literature.
Particular thanks go to Dr Patrick Osborne, who has juggled the roles of friend
and confidant with those of manager, statistical adviser, proof-reader and
occasional field-assistant, all with equal enthusiasm and ability. I could not have
completed this work without his love and support.
Prof. Charles Godfrey of Imperial College at Silwood Park, acted my adviser and
I thank him for sound technical advice and support. Thanks also go to Prof. John
Lawton CBE, and staff and students at Centre for Population Biology for making
me feel welcome during visits to Silwood. Special thanks go to Dr Stuart McNeill
for his comments and suggestions. Drs Mike Bonsall and Hefin Jones, Anne
Elliot and Emma Croxson are thanked for their friendship and kindness.
At NARC, I was ably assisted in data collection by Andy Griggs, Matt Love, Will
Mitchell and Ollie Wardman. Will’s efforts during the faecal calibration are much
appreciated. Derek Gliddon’s ERDAS skills facilitated the image analysis trials.
John Norton, Dr Mike Oatham and Benno Boer identified plant material, and
John collected and processed the plant epidermes. Dr Fred Launay, Ron
Loughland and Simon Aspinall provided some houbara faeces. Special thanks go
to Jude Hewlett and Maggie Black for their excellent counsel and support.
My thanks also go to the Avian Ecology Group at Stirling University, particularly
Prof. Peter Hudson for his enthusiasm and support. Special thanks go to Dr Tim
Benton for his interest in desert ecology and suggestions for improving draft
chapters. Drs. Dan Thompkins and Ian Stevenson are thanked for their
comments on several chapters, while Mandy Fox, Sue Haysom, Lou Rowe, Zoé
Russell and Laura Sanders provided friendship and advice, along with copious
cups of tea and coffee in moments of despair. The design skills of Bill Jamieson

14
and Dave Aitchison of the Environmental Science Department are much
appreciated (Figures 1.1 & 4.1, Plates 1.1 & 4.1 and Appendix 5.4), and Tracey
Grieve helped to print several of the Figures.
Finally, I want to acknowledge taxonomists around the globe for their knowledge
of and enthusiasm for the Arabian fauna. John Boorman collated numerous
strange specimens that I sent to the Natural History Museum, London, and
ensured that they reached the appropriate specialists (Dr Anne Baker, Dr
Annette Walker, Gary Stonedahl, I. White, John Chainey, Michael Schaffer, M.
Wilson, Nigel Wyatt, Paul Hillyard, Sharon Shute and Richard Thompson), with
particular thanks to Dr George Popov MBE (Orthoptera) and Ted Wiltshire CBE
(Lepidoptera). In addition, Drs Julio Ferrer (Swedish Museum of Natural History,
Tenebrionidae) and Lou Sorkin (American Museum of Natural History, Aranaea)
are thanked for their proficient determinations. Last but not least, I thank Cedric
Collingwood (Leeds City Museum) and Dr Donat Agosti (American Museum of
Natural History) for introducing me to the wonderful world of ants.

15
C h a p t e r 1
G e n e r a l In t r o d u c t io n
Synopsis of Chapter 1
This chapter starts by explaining the rational behind the research, the aims of the
thesis and why there is an urgent need for a better understanding of houbara
bustard ecology. This is followed by a summary of the ecology and biology of the
United Arab Emirates (UAE), especially the terrestrial invertebrates. The Chapter
ends with a discussion of the family Otididae and the houbara bustard.

16
1.1 Identifying the problem; the rational behind the research
Our current understanding of houbara ecology is poor (Collar 1980; Cramp &
Simmons 1980; Johnsgard 1991; Osborne 1996a; Goriup 1997) and is based
mainly on observational data with few scientific studies. However, the species'
range is enormous and threats from habitat loss and hunting are both
widespread and on-going. Accurate data are needed to ensure the success of
international conservation efforts.
In recent years there has been considerable interest in the captive rearing of
houbara, and breeding centres have been established in UAE (Osborne 1996b),
Saudi Arabia (Biquand etal. 1992; Saint Jaime etal. 1996), Kazakhstan (Gubin
& Mukhina in prep.) and Morocco. This is in response to a perceived reduction in
numbers of houbara, which is the favoured prey species of traditional falconry
and of immense cultural importance in the Arabian Gulf (Badr 1976; Remple &
Gross 1993; Osborne 1996a & b). Despite considerable investment and effort,
the success of ex situ breeding and release programmes for rare species is low,
usually because of a lack of understanding of habitat requirements and biology
(Scott & Carpenter 1987). Release programmes can also have undesirable
effects including the overwhelming of unique local populations by non-native
genotypes, and the death of released individuals because the habitat cannot
support them, e.g. the barn owl in UK (Griffiths etal. 1996). Better information on
the ecology and biology of wild houbara is vital if captive-bred birds are to survive
and flourish following release. Research into the role of invertebrates in the diet
could reveal whether the low density of houbara in Abu Dhabi is linked to the
seasonal availability of certain prey species. It could highlight areas of
importance to over-wintering birds and ways to enhance and restore habitats for
houbara. The study may help improve diets for captive birds, e.g. seasonal
changes probably play an important role in bringing birds into breeding condition.
In addition, natural foods may contain micro-nutrients which artificial foods
generally lack (Anderson 1995).
While knowledge of the houbara is poor, even less is known about the
invertebrates of the UAE (see Tigar 1996a & b and Gillett 1996). However, there
is growing recognition of the prominent role invertebrates play in desert
ecosystems (Polis 1991a; Aldryhim, etal. 1992). Their small size belies their

17
importance, and in some desert locations the biomass of ants is ten times that of
mammalian herbivores (MacKay 1991). Invertebrates are also useful indicator
species for assessing habitat quality and biodiversity (Perfecto & Vandermeer
1996; Wilson 1992; Polls 1991a). An understanding of local invertebrates is
essential if we are to protect and maintain fragile habitats from environmental
change. As Abu Dhabi continues to develop, urbanisation, disturbance through
human activity and overgrazing of range lands are becoming increasingly
common (Oatham 1996; Osborne 1996a; Collingwood etal. 1997).
The aims of the thesis are to investigate patterns of seasonal abundance,
distribution and diversity of desert invertebrates in Abu Dhabi and to determine
their importance in the diet of houbara. Secondary objectives are to develop a
reliable method of faecal analysis for the study of houbara diet and to establish
monitoring techniques for the most important invertebrates.
Gizzard contents of wild houbara were also examined, and while emphasis was
placed on the diet of wild birds in their natural environment, captive houbara were
used as models to produce a calibration method for quantifying faecal analysis.
An estimation of the nutritional value of invertebrates was used to test the value
of the technique and to suggest ways of improving diets for captive birds. Where
possible, Abu Dhabi's invertebrates were compared to the invertebrate
communities of other deserts. In addition, a database for invertebrate
observations in UAE and a reference collection of invertebrate specimens and
photographs were established at the National Avian Research Center (NARC).
1.2 Abu Dhabi and the United Arab Emirates
Introduction
The UAE is a federation of seven emirates covering an area of 77,700 km , with
Abu Dhabi emirate occupying over 86% of the land (Figure 1.1). The Tropic of
Cancer crosses the southern tip of UAE which is part of the Great Palaearctic
Desert stretching from North Africa to western Pakistan. Over 90% of the country
is below 250m above sea level (ASL) and consists of flat plains overlain by sand
dunes, interspersed with lagoons and sabkhas (salt flats) at the coast.

18
52° E 53° E 55° E 56° E54° E
26° N - - 26° N
25° N - - 25° N
Public ----- '■ ;■Hunting [
f Triangle j
- 24° N24° N - Baynunah Khatam •
Medinet Zayed
O M A NA B U D H A B I
- 23° N23° N -Dm Az Zimul
km 100S A U D I A R A B I A
53° E 55° E52° E 54° E 56° E
Figure 1.1 Map of Abu Dhabi Emirate showing the five principal study sites;
Baynunah, Medinet Zayed, Public Hunting Triangle, Khatam and Dm Az Zimul

19
The smaller mountainous region of the country reaches about 1500m ASL and
is deeply dissected by several seasonal river valleys or wadis.
Traditionally, society was based on nomadic pasturalism, with herds of camels,
goats and sheep in the desert and limited cultivation at oases such as Liwa
(Anon 1993a). Following the exploitation of oil reserves in the 1960s, economic
development and population growth rose sharply, indirectly causing a lowering
of the water table and an increase in ground water salinity. Now desalination
produces over 80% of the water, much of which is used to irrigate vast forestry
plantations and agricultural developments in Abu Dhabi emirate. Between 1973
and 1990 the area of agricultural land increased from 13,000 to 43,000 ha
(Anon 1993a). Most people live in towns and no longer depend on the desert for
their livelihood.
Clements (1983) has produced a bibliography on the UAE, and there is also a
comprehensive atlas (Anon 1993a).
Geoloqv
The following description of Abu Dhabi’s geology is based on Glennie (1996),
while Anon (1993a) and Feulner (1997) give descriptions for the UAE as whole.
Five land forms occur in Abu Dhabi: sand dunes, interdune areas, coastal
sabkha, inland sabkha and exposed rock. The sand dunes are made of quartz
grains, with a higher carbonate content near the coast. The largest dunes are
static and support scant vegetation, while the smaller dunes (up to 20m high)
are quite mobile. The interdune plains are formed from either fluvial gravels or
low, scrub-covered sand drifts. They often contain gypsum crystals indicating a
history of inland sabkha.
The coastal sabkha occupies a band up to 30km wide along most of Abu
Dhabi’s coastline, at or above high tide level. It is an extremely saline and
evaporative environment, often covered in a crusty layer of halite (salt) and
characterised by a high concentration of carbonates and extensive sheets of
black algal mat. Inland sabkhas lack this algal mat and frequently occupy
interdune areas.

20
Apart from the low mountain at Jebel Hafit, Abu Dhabi emirate is a sandy desert
and three types of exposed rock-based soils occur in the rocky outcrops which
are of Tertiary, Mesozoic and Cambro-Precambrian age.
The two most important abiotic factors affecting the flora and fauna are soil
salinity and compaction. The latter determines whether the substrate is suitable
for burrows and also the ease of root penetration.
Climate
The harsh hyper-arid desert exerts great pressure on life, and many of its
inhabitants are adapted to avoid high temperatures and conserve water.
Bottomley (1996) has described the recent climate of Abu Dhabi. Winter occurs
from December to March, with unsettled periods of rain and strong winds. In
April and May temperatures start to rise and between June and September the
maximum temperature can exceed 40°C. October and November are
characterised by lower temperatures and light winds.
Mean maximum temperatures occur in July (42°C for coastal Abu Dhabi; 44°G
for AI Ain about 100km inland) with mean minimum temperatures in January
(14°C for Abu Dhabi; 12°C for AI Ain). The absolute maximum and minimum dry
bulb temperatures are 47.5°C and 49°C, and 7°C and 3°C for Abu Dhabi and AI
Ain cities respectively. In south-western Abu Dhabi Emirate, near the Rub AI
Khali or Empty Quarter, temperatures are even higher and can exceed 52°C
(Mandaville 1990). The interior may reach close to freezing-point in winter, when
daily temperature fluctuations occur caused by strong, north-westerly winds
known as shamals.
Relative humidity (rh) is highest during the summer along the coastal strip, often
exceeding 90% rh at 30°C, while inland rh below 10% can occur in May
because of a heat low over the Rub AI Khali (Bottomley 1996).
Abu Dhabi receives extremely variable amounts of infrequent rains and a whole
year’s rain can fall in one rain event. The mean annual rainfall is <100mm and
rain mainly falls between December and May. The average number of days on

21
which rainfall is recorded is only ten a year. Rainfall totals are highest in the
north-eastern and lowest in the south-western parts of Abu Dhabi. Summer rain
is rare but storms may occur near AI Ain because of cloud from the Indian
monsoon. The Rub AI Khali is extremely arid and its endemic plant community
is dominated by a few drought adapted shrubs; however complete destruction of
vegetation over great expanses may occur if rains fail repeatedly for several
years (Mandaville 1990).
The cloudless skies, intense solar radiation, high temperatures and low humidity
result in very high evaporation rates, which may exceed the mean annual
rainfall by 35-100 times (Mandaville 1990). This extreme evapo-transpiration
stress is a limiting factor to all forms of life (Holdridge etal. 1971 ; Schulze &
McGee 1978).
Winds are generally light but gusty shamals can reach speeds of 20-25 knots,
occasionally over 40 knots. The strongest winds (>60 knots) are associated with
squall lines or thunder storms. Although mean wind velocity is low, material
carried by winds can damage plant and animal cuticles. Summer shamals can
cause severe moisture stress to already drought stressed plants (Mandaville
1990).
Mist and fog form during the winter and at the end of summer. Dew is an
important form of moisture for desert life, and many shrubs act as centres of
condensation. For example, the sand under Haloxylon persicum Bge. bushes is
often covered with green algae (personal observation). Some desert animals
are able to use dew and although “fog basking” (Hamilton & Seely 1976; Seely
1979) has not been recorded in Arabian tenebrionids, beetles of the genus
Erodius probably extract water from specially constructed tunnels on dune
ridges (Büttiker & Anderson 1988).
Ecoloav
UAE contains elements from three biogeographic zones: the northern
Palaearctic, African and Asian, as well as Arabian endemics, and is potentially a
centre of high biodiversity. In his review of the UAE Satchell (1978) found few
accounts of its ecology, but recent publications include accounts of the natural

22
history of the Emirates (Jongbloed 1987; Vine 1997; Western 1989) and the
ecology of Abu Dhabi (Osborne 1996c). The Emirates Natural History Group
keep reference material and records of flora and fauna.
Introductions to the flora can be found in El Ghomeny (1985), Western (1989)
and Jongbloed (1987, 1997). Mandaville (1990) describes the flora of eastern
Arabia and provides much useful information, as does the Flora of the Arabian
Peninsula and Socotra series (Miller & Cope 1996). Satchell etal. (1981) give a
systematic description of land classes across the UAE, while Roshier et al.
(1996) describe nine characteristic plant species associations of Abu Dhabi. The
Emirates contains plants from both the Saharo-Arabian and Sudanian plant-
geographical regions (Zohary 1973) but their transition is poorly defined and
masked by the hyper-arid conditions in the Rub AI Khali. The Saharo-Arabian
region is a centre of diversity for the genera Suaeda, Fagonia and Zygophyllum.
Much of the fauna of Arabia has only recently been described (see Fauna of
Saudi Arabia series, published since 1980, edited by Wittmer & Büttiker).
However, the birds of the Emirates and Abu Dhabi are fairly well known
(Richardson 1990; Aspinall 1996; Osborne etal. 1996; Richardson 1997) and
there are various publications on land and sea mammals (Gross 1987;
Duckworth 1996; Baldwin 1997; Gross 1997a), fish (Shepley 1997), reptiles and
amphibians (Baha El Din 1996; Gross 1997b) and invertebrates (Gillett 1996;
Tigar 1996a & b).
Invertebrates
Desert arthropods have various mechanisms to conserve water and avoid heat
stress. Their small body size allows them to move into cooler microclimates
during the day and many live in deep burrows or near the roots of plants and
only emerge at night when the temperature has dropped. However, some
invertebrates are able to survive the heat of the day, including ants of the genus
Cataglyphis (Delye 1968) and scorpions which can tolerate higher temperatures
than most other desert arthropods (Polis 1990a). Many predatory arthropods
survive long periods without feeding if prey is scarce (Polis & Yamashita 1991).

23
Arabian invertebrates are not well known but are an abundant and important
component of the desert ecosystem. The majority of Arabian records originate
from scientific expeditions in Saudi Arabia and Oman (see Büttiker & Krupp
1980; Dutton & Bray 1988; Shaw Reade etal. 1980), and records from the UAE
are scant and often come from unrepresentative habitats, such as cultivated or
irrigated areas, where water and exotic vegetation allow non-desert species to
thrive. Taxa by taxa accounts of the invertebrates of Abu Dhabi and the UAE
are given in Tigar (1996a & b; Appendices 1.1 & 1.2) and a summary is
provided here. Information from neighbouring Saudi and Oman was used where
data were lacking.
Arachnids are well represented, with examples from the Scorpiones, Solifugae
and Araneae. Scorpions are perhaps best known, and four families probably
occur in Abu Dhabi: Buthidae, Chactidae, Diplocentridae and Scorpionidae
(Sissom 1990). Buthids are most numerous with 14 Arabian genera, including
the highly venomous Buthacus, Androctonus and Leiurus (Simard & Watt 1990).
Vachoniolus globimanus has been recorded in Abu Dhabi (Levy et al. 1973;
Vachon 1980). Other Arabian arachnids remain poorly described. The solifugids
or camel spiders are common and probably include the families Galeodidae,
Solpugidae and Rhagodidae (Cloudsley Thompson 1987), while desert spiders
may include the Gnaphosidae, Sparassidae, Thomisidae, Hersiliidae, Salticidae,
Lycosidae, Agelenidae, Theridiidae, Tetragnathidae and Argiopidae (Cloudsley
Thompson 1984). Free living mites from arid zones are poorly known but Giant
Velvet Mites Dinothrombium spp. emerge during the rains, and ixoid ticks are
better known because of their importance as vectors of infectious diseases (see
Tigar 1996b).
Many insects live in Abu Dhabi's deserts, including a few entognathans such as
Collembola, which probably survive in humid, underground microclimates (Zak &
Freckman 1991). Thysanura are common and Irish (1991) describes 11 Arabian
species but has no records from UAE, although the Lepismatidae undoubtedly
occur. Other insects in UAE include 12 of the 52 species of Arabian Odonata,
including several long distance migrants (Waterston & Pittaway 1989). Isoptera
are common with 20 Arabian species (Chhotani & Bose 1983, 1991) although
only Heterotermes aethiopicus is recorded from UAE (Boocock 1979). There
are 46 species of mantids in Arabia (Kaltenbach 1984, 1991), and while there

24
are no records for UAE, the curious and cryptically coloured ground mantis,
Eremiaphila bauri Krass, is common on gravel plains and other species live on
plants (Walker & Pittaway 1987).
The Orthoptera contains some important desert species which are occasionally
very numerous, including two locusts Schistocerca gregaria Forskâl and
Locusta migratoria Linnaeus, which are known from Abu Dhabi (see Uvarov
1952, 1966 & 1977). Popov (1980) lists 69 species of Acridoidea from Eastern
Arabia of which 28 occur in UAE. Tettigoniidae, Gryllidae and Gryllotalpidae are
also common across Arabia (Popov 1981; Walker & Pittaway 1987).
Hemiptera are not particularly well adapted for desert life and only occur where
there is abundant vegetation or prey. Matile-Ferrero (1984 & 1988) described 60
species of Aphididae and Coccidae, and Linnavuori (1986) 452 species of
Heteroptera from Arabia but there were no records for UAE. However, the
Arabian cicada Platypleura arabica Myers occurs in stands of trees and a large
shield-bug, Coridius viduatus Fabricusa (Dinidoridae), lives on native or
cultivated cucurbits (Walker & Pittaway 1987).
Two Neuropteran families occur in UAE: Chrysopidae and Myrmeleontidae.
Most larval Myrmeleontidae, or ant lions, construct conical, pit-like traps in the
sand to catch ants, but a few species have active, predatory larvae. Holzel
(1982, 1983 & 1988) lists 156 species from Arabia but UAE species are not
mentioned.
Coleoptera in UAE are abundant and diverse and include Carabidae,
Cicindelidae, Histeridae, Scarabaeidae, Buprestidae, Elateridae, Coccinelidae,
Tenebrionidae, Meloidae and Curculionidae. Large desert carabids, like Anthia
duodecimguttata Bonelli and Scarites spp. are common and tiger beetles, such
as Cicindela lunulata Fabricius and Cicindela immanis (Bates), occur on
intertidal mud flats and sabkhas. The Scarabaeidae include dung beetles, like
Scarabaeus cristatus Fabricius which is very numerous. However, tenebrionids
or darkling beetles are the most abundant and successful desert coleopterans.
They obtain all their water via their metabolism, and the chamber below the
fused elytra of flightless species probably aids water retention. They are
generally detritivores and are usually black or dark-coloured, and crepuscular or

25
nocturnal in habit. Kaszab (1981 & 1982) lists over 300 species for Arabia but
has split some species into several subspecies without providing a key or
description. He lists 18 species from DAE and the most common are Mesostena
puncticollis SoWer, Tentyrina palmeri Crolch, Trachyderma (=Ocnera) philistina
Reiche & Saulcy, Pimelia arabica Klug, Blaps koiiari Seidlitz and Prionotheca
coronata (Olivier). The genus Erodius is also abundant in some areas.
A few Arabian Diptera are well known, particularly disease vectors such as the
Ceratopogonidae and Chironomidae (Boorman 1989; Cranston & Judd 1989).
The Bombyliidae or bee flies have been studied by Greathead (1988) who lists
21 species from UAE. Other families such as Tabanidae, Asilidae and
Calliphoridae also frequent desert areas. Pont (1991) lists 68 species of
Muscidae and the most abundant species are Musca domestica domestica
Linnaeus, Musca domestica calleva Walker, Musca lucidula Loew and Musca
sorbens Wiedermann.
Lepidoptera are fairly well known. Larsen (1983) has written a monograph of
151 butterfly (Rhopalocera) species, and Wiltshire (1983, 1986, 1988 & 1990)
has reviewed the 625 species of Macro-Heterocera from the Arabian Peninsula.
Lepidoptera in UAE include the Cossidae, Pyralidae, Hesperiidae, Papilionidae,
Pieridae, Lycaenidae, Nymphalidae, Geometridae, Lasiocampidae, Sphingidae,
Lymantriidae, Arctiidae, Noctuidae and Psychidae.
Desert Hymenoptera include many ants, wasps and bees, some of which are
well known, for example, the Apidae, Anthophoridae, Megachilidae, Pompilidae,
Chrysididae, Scoliidae, Eumenidae, Vespidae and Sphecidae (see Richards
1984a & b; Guichard 1988a & b,1989a & b; Hamer 1982, 1986a & b; Roche
1981). Ants are an important ecological group and surveys of Saudi Arabia and
Oman revealed 156 and 28 species respectively (Collingwood 1985 & 1988),
although 275 species are now known from Arabia (Collingwood & Agosti 1996).
Tigar & Collingwood (1993) list 14 species from UAE, but these are only a small
proportion of those that occur. They include domestic pests, such as some
Monomorium spp., nocturnal predatory species like Camponotus xerxes Forel
and desert specialists of the genus Cafag/yp/i/s which forage during the day.

26
Other arthropods include a small number of Crustacea such as woodlice
(lsopoda:Porcellionidae), and Chilopoda (centipedes), especially the genus
Scolopendra.
1.3 Introduction to the bustards - family Otididae
This is a brief synopsis of current knowledge. More detailed reviews of the
bustards can be found in Cramp and Simmons (1980), Osborne etaL (1984),
Collar et a i (1986) and Johnsgard (1991), while a comprehensive bibliography
was published by Schulz & Schulz (1991). Specific reviews of houbara biology
have been compiled by Gubin & Mukhina (in prep.), Morris (1991) and Collins
(1984), although the latter mainly covers the Canarian houbara.
Bustards belong to the family Otididae, a homogeneous and ancient lineage
consisting of 22 species representing 6-8 genera (Collar et a i 1986; Osborne
1996a). They are divided into 47 subspecies or races according to differences in
size, plumage and geographic location (Osborne et a i 1984), although their
phylogeny remains in dispute (Cramp & Simmons 1980; Johnsgard 1991).
They are medium to large birds which are characteristic of open plains and
semi-desert, although some African species live in thorn scrub. They are
distinguished by the presence of unusual hexagonal scales on their legs, the
absence of a crop, the absence of a preen or oil gland and by having only three
toes, since a hind toe is lacking. Bustards cannot oil themselves but are covered
in a friable material called “powder down”, which along with dust-bathing is
thought to keep the plumage clean. The young are précédai and nidifugous and
must be bill-fed by the female for their first few weeks. Insects are thought to be
the most important part of the diet for chicks but adult birds are probably
omnivorous, although accurate data are lacking.

27
1.4 The houbara bustard
The houbara is a large bird with a distinctive turkey-like build (Cramp &
Simmons 1980) (Plate 1.1). Males weigh up to 2.36kg and females 1.8kg, but
mean weights are proportionately lower and vary with season; birds are heaviest
in winter (Johnsgard 1991). Houbara are cryptically coloured with a sandy-buff
plumage, and blackish vermiculations or blotches, a black stripe down side of
neck and a relatively long tail. Their white crown-tuft is diagnostic but is not
always visible (Osborne etal. 1984). Juveniles and females are smaller than
males with less well developed crests and ruffs. Birds are sexually mature in
their second year (Johnsgard 1991).
Subspecies and their ranges
Collar (1980) gives an account of the houbara’s general distribution updated by
Goriup (1997). Its taxonomy is described by Goriup & Collar (1980) who
recognise three subspecies, but Gaucher etal. (1996) suggest that Chlamydotis
undulata macqueeni may be a separate species because of differences in its
genetics and behaviour. The nominate North African race, Chlamydotis
undulata undulata, ranges from Mauritania to Egypt, with most of the population
in Morocco. It is sedentary or locally nomadic with a population of about 10 000
birds (Goriup 1997).
The rarest race is the Canarian houbara, Chlamydotis undulata fuertaventurae,
which is sedentary and confined to Fuerteventura, Lanzarote and Graciosa in
the Canary Islands. In the 1980’s there were only 100-300 birds (Collar 1980,
Dominguez 1989) although there are now about 750 (Goriup 1997).
“Macqueen’s” bustard, C.u.macqueeni, ranges from eastern Egypt to Jordan,
Baluchistan, Kazakhstan, Mongolia and eastern China, and Goriup (1997) gives
a population estimate of 39-52,000. The ranges of C.u.undulata and
C.u.macqueeni probably overlap in the Nile valley.
C.u.macqueenI can be migratory, sedentary or locally nomadic. It is paler and
slightly larger than the other races, with a black centre to its white crown tuft, an
olive-green bill and pale creamy-grey feet and legs (Osborne etal. 1984).

28
Plate 1.1 Houbara bustard

29
In Arabia C. u. macqueeni \s primarily a winter visitor, with only small breeding
populations in Saudi, Oman and Yemen (Osborne 1996a). It was also thought
to have bred in Syria, Iraq and Iran but populations are now very low (Collar
1980; Johnsgard 1991; Goriup 1997). The Republics of Kazakhstan and
Uzbekistan, and the Kyzyl-Kum and Turkmenistan deserts are probably the
main breeding areas before the birds move to India or Pakistan. Satellite
tracking has shown that birds over-wintering in Abu Dhabi migrate to
Turkmenistan (Osborne etal. 1997a). Both resident breeding and winter
populations occur in Pakistan, although the latter outnumber breeding birds.
Houbara habitat
The houbara is a bird of harsh, arid, sandy or stony deserts and steppes and is
thought to be among the best adapted birds for desert life. It is associated with
sparse xerophytic and halophytic vegetation such as low shrubs like Artemisia
spp., Tamarixspp., or Haloxylon spp., but may frequent arable land (Osborne et
al. 1984). The climatic and vegetative features of the houbara’s range appear to
correspond with the Irano-Turanian, Saharo-Arabian and Sudanian
phytogeographical regions defined by Zohary (1973) (Osborne 1996a).
EcoIoqv of Chlamydotis undulata macqueeni
Houbara are powerful fliers and can migrate several thousand kilometres
(Osborne etal. 1997a), but they are also cursorial and walk great distances
(Osborne etal. 1984). During migration they form flocks of up to 60 birds
(Cramp & Simmons 1980) but usually occur in loose groups of four to ten birds
which feed and roost together outside of the breeding season (Osborne etal.
1984). Typical population densities are low; between 0.3 (Mian & Surahio 1983)
to 1.7 birds per km in Pakistan (Mirza 1985), and 0.032 to 0.097 birds per km
in Saudi Arabia (Seddon & van Heezik 1996).
Houbara are very wary and are difficult to flush, preferring to crouch behind a
bush or stone, but if forced they will quickly fly out of view. Their capacity for
concealment and cryptic coloration make them surprisingly hard to locate in the
field. Houbara are the favoured prey of Arab falconers, even though such large
birds are difficult quarry. Indeed, houbara may also squirt sticky, caecal faeces

30
over pursuing falcons, causing them to give up the chase unless their feathers
are cleaned by the falconer (Osborne etal. 1984).
In Central Asia breeding occurs from April to June (Gubin & Mukhina, in prep.)
and generally coincides with the spring rains. During the breeding season,
males display at traditional sites with about 500m between rival males. The
unusual “running display" of males is carried out repeatedly through the day
(Osborne etal. 1984). The male initially lowers the tail and spreads and raises
its crown and neck feathers into upward curves. It then retracts its head and
completely puffs out its neck plumage. The mating system is thought to be
polygynous or promiscuous (Collar etal. 1986) but may be monogynous at low
population densities (Osborne 1996a). After copulation the female rears the
young alone. Eggs are olive brown with dark blotches, and are laid in a shallow
scrape with 2 or 3 eggs per clutch. Incubation takes 23 days and the young
fledge in about 35 days but remain with the mother for longer. Details of
Houbara behaviour are described in Cramp & Simmons (1980), Osborne etal.
(1984), Collins (1984), Collar etal. (1986) and Launay & Paillat (1990).
Unusually for a bustard, houbara are virtually silent. Females make a low “quop”
or “quip” like call and chicks a plaintive peeping noise (Cramp & Simmons
1980).
Houbara are generally considered to be opportunistic omnivores. However,
much of the information on their diet is based on historic descriptions of single
samples. In Chapter 2 ,1 review the literature relating to houbara diet and try to
summarise patterns of food consumption by taxa across the range. Some
authors suggest that houbara do not drink and obtain all the moisture they need
from their food or morning dew (Valverde 1957; Mian 1988). However, they also
drink at water holes (Cramp & Simmons 1980) and in captivity.
The conservation and status of the houbara
The houbara is currently on Appendix II of CITES and is subject to strict export
control. It is variously reported as endangered in different parts of the range and
has some degree of protection although is still hunted in many countries. For
example it is listed in the Red Data Book for the USSR and is considered

31
“endangered” in Mongolia (see Johnsgard 1991 for further details). However,
Goriup (1997) has suggested that population estimates for the former USSR
should be at least 20 times higher than previous calculations (Flint etal. 1992),
and that while the Canary Islands subspecies should be listed as “Vulnerable”
according to lUCN criteria, the houbara from other parts of the range are more
numerous and should be listed as “Vulnerable” or “Near threatened” according
to local population levels.
Accurate data on the distribution and status of the houbara are lacking across
much of the range, and population estimates are both difficult and expensive to
obtain because of the patchy distribution of the houbara across desert terrain
(Osborne 1996a). However the population is thought to have declined greatly
during the 20* century (Collar 1980; Goriup 1997). The most probable causes
are excessive hunting (especially from motor vehicles), overgrazing of habitat,
agricultural development, industrial development, egg collecting and the general
pressure of human settlement (Collar 1980). However, hunting records from
Pakistan suggest that the population may have been underestimated because
such high mortality could not have been sustained, and evidence from Central
Asia also suggests that the bird is more numerous (Osborne 1996a). Other
bustard species, such as the great bustard Otis tarda, have also declined over
the last 50 years following agricultural and land-use changes (Hidalgo de
Trucios 1990; Alonso & Alonso 1996), and co-ordinated, international action is
essential to prevent the continued demise of these birds across their ranges
from organisations such as the Bustard Specialist Group (run by the lUCN
Species Survival Commission and BirdLife International) (Goriup 1997).

32
CHAPTER 2
T h e d ie t o f t h e h o u b a r a b u s t a r d - a r e v ie w
Synopsis of Chapter 2
This chapter reviews current literature on the houbara diet. The known food
items of each houbara subspecies are given in turn, followed by detailed
information on animal and plant foods and any examinations of gizzards or
faecal samples. Semi-captive, captive and vagrant birds are mentioned,
especially where natural foods were eaten, and information on gizzard stones is
presented. The discussion compares the methods used to study the diet and
presents an analysis of known items in the diet based on the frequency of
citations for each taxon consumed. This calculated value or citation index
makes it easier to compare between the houbara subspecies. The conclusion
summarises general dietary trends for the species, including seasonal patterns
and specific requirements of birds according to age and reproductive status.

33
2.1 Introduction
Several authors have reviewed the diet of the houbara bustard including Cramp
& Simmons (1980), Collins (1984), Roberts (1991), Morris (1991), Johnsgard
(1991) and Gubin & Mukhina (in prep.). Houbara are often described as
omnivorous, although much of the literature is speculative, observational or
based on single samples, and there are few attempts to quantify the diet. The
only replicated results are some examinations of faeces (see Gubin & Mukhina
in prep.; Collins 1984) and gizzard contents (see Fox 1988; Mirza 1971).
2.2 The review of the diet
All non-speculative data on foods of the houbara were summarised into tables
(Table 2.1 green plants; Table 2.2 fruits, seeds and flowers; Table 2.3
invertebrates and Table 2.4 vertebrates). Some of the literature is unpublished
and has not been peer-reviewed, and may contain inaccuracies. However, it is
often the only source of information on the houbara. Except for the houbara,
scientific nomenclature is not abbreviated to avoid confusion caused by the
large number of genera.
2.2.1 The Canary Island houbara: Chlamydotis undulata fuertaventurae
General descriptions of the diet
Collar (1983) provides a useful translation of early work on C.u. fuertaventurae,
gleaned from the notebooks and letters of bird collectors. General descriptions
of the diet include beetles, wheat and oats (Webb & Berthelot 1836-1844),
snails, lizards and a trefoil (Meade-Waldo 1889a), lizards, plants, seeds and
dung beetles (Volsoe 1951) and invertebrates, snails, lizards {Lacerta atlantica),
geckos {Tarentola delanderei), rabbits, house-mice, rats and hedgehogs
{Erinaceus algirus) (Hooker 1958). However, Hooker (1958) may have implied
that houbara were foraging on necrophilic invertebrates living in carrion, rather
than preying upon live mammals (see Sushkin 1908). Bannerman (1963) states
that houbara eat peas, beans, snails, beetles (dung beetles), caterpillars, a
trefoil and fruits of Lycium intricatum (Solanaceae).

34
Plant name
"Green plants"
Young green shoots
Chlamydotis undulata Chlamydotis undulata Chlamydotis undulatafuertaventurae undulata macqueeni
Vols0e (1951) Valverde (1957), Bédé (1928), Lavauden (1914) Brosset (1961)
Alekseev (1985), Roberts & Savage (1971) Dharmakumarsinhji (1955), Ferguson-Lees (1969), Mian (1988), Roberts(1991), Goriup & Norton(1992 )___________________
MonocotyledonsGramlneaeGrass
Wheat and other crops (see also Table 2 .2)Bromis tectorum Cymbopogon sp. Koeleria phleoides Lasiurus sp.Poa bulbosaOedibasis apiculata
Collins (1984, 1993) Anon (1980)
Lavauden (1914)
Salikhbaev & Ostrapenko (1967), Afanas'ev & Sludskiy (1947), Zhuyuko (1986)Baker (1912), AN & Ripley (1969), Ferguson-Lees (1969), Glutz von Blotzheim etal. (1973) Gubin & Mukhina (in prep.) Mirza (anecdotal) (1985) Mian & Surahio (1983) Mirza (1971)Gubin & Mukhina (in
prep.)Gubin & Mukhina (in
Liliaceae Tulipa Allium sp.(garlic, onion and other Allium sp.)
Gubin & Mukhina (in prep.) Sushkin (1908), Ferguson- Lees (1969), Meinertzhagen (1954), Giutz von Blotzheim etal. (1973)
DicotyledonsAizoaceaeMesembryanthemumspp.
Meade-Waldo (1889a & b), Collins (1984, 1993)
Asclepiadaceae Leptadenia sp. Leptadenia spartinum
Mirza (anecdotal) (1985) Dharmakumarsinhji (1955)
Apocynaceae Rhazya striata Mian & Surahio (1983)
Capparaceae Capparis sp.
Capparis aphylla Capparis decidua Capparis spinosa Dipterygium glaucum
Molchanov (1913), Fox (1988)Surahio (1985)Mian (1988)Seddon, P. (pers. comm.) Launay, F. (pers. comm.)
Chenopodiaceae Anabasis sp. Mian (1988)
Haloxylon sp. Gaucher (1991) Mirza (1971), Surahio (1985), Mian (1988)
Table 2.1
List of plants eaten by the three subspecies of houbara as vegetative material

35
Plant name Chlamydotis undulata Chlamydotis undulata Chlamydotis undulata fuertaventurae undulata macqueeni
ChenopodiaceaeHaloxylon Mian & Surahio (1983)
ammodendronSalsola sp. Mian (1988)
Salsola arbuscula Mian & Surahio (1983),Gubin & Mukhina (in prep.)
Salsola richeri Gubin & Mukhina (in prep.)
Salsola rigida Gubin & Mukhina (in prep.)
Compositae Artemisia sp. (shoots)
Calendula aegyptiaca Lactuca sp.Lactuca serricola Launaea sp.
Collins (1993)
Collins (1993)
Sushkin (1908), Meinertzhagen (1954), Ferguson-Lees (1969), Glutz von Blotzheim e ta l (1973), Gavrin (1962)
Fox (1988)Pavlenko (1962)
Goriup & Taylor (1983)
Cruciferae Collins (1993)
Brassica, mustard, Ali & Ripley (1969),
oilseed and cabbage. Ferguson-Lees (1969), Glutz von Blotzheim e ta l (1973), Surahio (1985)
Brassica campestris Roberts & Savage (1971).
Brassica juncea Mian (1988)
Eruca sativum Mian (1988)
Farsetia sp. Mirza (1971), Gallagher(unpubl.)
Farsetia aegyptia Goriup & Norton (1992)
Farsetia jacquemontii Fox (1988)
Hirschfeldia incana Lack (1983)
Lobularia lybica Collins (1984, 1993)
Malcomia africana Mian & Surahio (1983)
Malcomia sp. Mian (1988)
Notoceras bicorne Collins (1984, 1993)
Gyperaceae Carex pachystilis Carex physodes
Gubin & Mukhina (in prep.) Gubin & Mukhina (in prep.)
Ephedraceae Ephedra sp. Mian & Surahio (1983)
Euphorbiaceae Euphorbia granulata Euphorbia prostrata
Fox (1988)Mirza (anecdotal) (1985)
Geraniaceae Erodium sp. Erodium cicutarium Collins (1984, 1993)
Mian & Surahio (1983)
LeguminosaeTrefoil Meade-Waldo (1889a),
Bannerman (1963)
Table 2.1 (continued)
List of plants eaten by the three subspecies of houbara as vegetative material

36
Plant name Chlamydotis undulata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
Leguminosae Cultivated beans and peas (see Table 1b)
Acacia sp.Alhagi mauraurum Astragalus hamosus Crotalaria sp.
Indigofera sp. Medicago minima Medicago sativa Alfalfa
Polatzek (1909), Bannerman (1963), Lack (1983), Collins (1984)
Collins (1984,1993)
Collins (1984, 1993) Collins (1984) (anecdotal)
Symens (1987) (captive) Dharmakumarsinhji (1955)
Mirza, (1971), Gallagher (unpubl.), Surahio (1985) Symens (1987) (captive)
Symens (1987) (captive), Launay (1989a, b & c) (captive)
Plantaginaceae Plantago ciliata Plantago sp. Collins (1984, 1993)
Mian & Surahio (1983) Fox (1988)
Polygonaceae Calligonum comosum Polygonum afghanicum
Mian & Surahio (1983) Mian & Surahio (1983)
Solanaceae Tomato leaves Lycium shawii
Valverde (1957)Symens (1987) (captive)
Tiliaceae Grewia populifolia Triumfetta rotundifolia
Surahio (1985) Dharmakumarsinhji (1955)
Zygophyllaceae Fagonia sp.
Fagonia indica Tribulus sp.
Tribulus alatus Tribulus terrestrisZygophyllum sp.
Mirza (1971), Gallagher (unpubl.)Symens (1987) (captive) Mirza (1971), Surahio (1985), Mian (1988)Mian & Surahio (1983) Fox (1988), Mian & Surahio (1983)Pavlenko (1962), Meklenburtsev (1990), Mirza (1971)
Unknown Eulaliopsis sp. Mirza (1971)
Table 2.1 (continued)
List of plants eaten by the three subspecies of houbara as vegetative material

37
Plant name Chiamydotis unduiata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
Flowers
Fruits & Seeds
Seeds
Berries
Vols0e (1951)
Whitaker (1905), Anon (1980)Glutz von Blotzheim et al (1973)
Glutz von Blotzheim et a! (1973), Brosset (1961)
Ali & Ripley (1969), Goriup & Norton (1992)Cornwallis (1982)
Dharmakumarsinhji (1955), Ferguson-Lees (1969), Roberts & Savage (1971), Mian (1988)Glutz von Blotzheim e ta l (1973)
MonocotyledonsGramineae Grain and Cereals
Wheat
Oat
BarleySorghum seeds Sorghum bicolor Panicum sp. (seeds) Grass (spikelets)
Palmaceae Dates (fruit)
Polatzek (1909)
Webb & Berthelot (1836- 44)Webb & Berthelot (1836- 44)Cramp & Simmons (1980)
Whitaker (1905), Glutz von Blotzheim et a / (1973)
Jourdain (1915)
Dharmakumarsinhji (1955),
Fox (1988)
Mian & Surahio (1983) Goriup & Norton (1992), Afanas’ev & Sludskiy (1947), Zhuyuko (1986)
Goriup & Taylor (1983)
DicotyledonsAnacardiaceae Pistacia atlantica (nuts)
Glutz von Blotzheim e ta l (1973), Gillett(1988)
Capparaceae Cadaba sp. (fruits) Capparis sp. (fruits)
Fox (1988)Dharmakumarsinhji (1955), Molchanov (1913)
Chenopodiaceae Anabasis articulata (fruits)Haloxyionammodenderon(seeds)
Gillett(1988)
Mian & Surahio (1983)
Cistaceae Heiianthemum ruficomum (buds & flowers)
Gaucher (1991)
CompositaeAchilieafragrantissima(flowers)Argina spinosa (flowers and fruits)
Heim de Balsac & Mayaud (1962), Glutz von Blotzheim et a / (1973)
Goriup & Norton (1992) (casual observation)
Table 2.2
List of flowers, fruits and seeds eaten by the three subspecies of houbara

38
Plant name Chlamydotis undulata Chlamydotis undulata Chlamydotis undulatafuertaventurae undulata macqueeni
Compositae Gagea reticulata (flowers and fruits) Matricaria sp.(flowers)Lactuca serricola (panicles)Launaea spp.(flowers)Launaea arborescens (flowers)____________
Gillett (1988)
Valverde (1957), Goriup & Taylor (1983)
Gubin & Mukhina (in prep.)
Pavlenko (1962)
Hemmingsen (1958), Collins (1984, 1993)
Cruciferae Farsetia aegyptia (fruits)Farsetia hamiltoni (fruits and flowers) Farsetia stylosa (fruits and flowers)
Goriup & Norton (1992)
Goriup & Taylor (1983)
Gaucher (1991)
Geraniaceae Erodium oxyrrhynchum (seeds)______
Gubin & Mukhina (in prep.)
Leguminosae Fruits and seeds Peas
Acacia sp. (seeds) Argyrolobium uniflorum (fruits) Astragalus macroladus (seeds) Astragalus turczaninovii (flowers) Cicer arietinum (Chick pea seeds) Lens culinaris lentil seedsMedicago lactinata (fruits)Reaumuria turkestanica (flowers) Ziila spinosa (flowers and fruits)
Volsoe (1951) Polatzek (1909), von Thanner (1913)
Pavlenko (1962)
Gaucher (1991)
Lack (1983) (anecdotal)
Lack (1983) (anecdotal)
Anegay (1992)
Gubin & Mukhina (in prep.)
Gubin & Mukhina (in prep.)
Fox (1988)
Goriup & Norton (1992)
Gubin & Mukhina (in prep.)
Goriup & Taylor (1983) (observation)
Moraceae Ficus sp. (fruits) Anon (1980)
PolygonaceaePolygonum Mian & Surahio (1983)
afghanicum (seeds)
Table 2.2 (continued)
List of flowers, fruits and seeds eaten by the three subspecies of houbara

39
Plant name Chlamydotis undulata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
Polygonaceae Polygonum sp. Buckwheat (seeds) Calligonum comosum (seeds)
Mian & Surahio (1983)
Mian & Surahio (1983)
Rhamnaceae Zizyphus jujuba (berries)Zizyphus lotus (berries)Zizyphus numularia (berries)Zizyphus sp. (berries)
Gillett (1988)
Glutz von Blotzheim et al (1973), Ali & Ripley (1969)
Surahio (1985)
Cramp & Simmons (1980), Mian (1988), Fox (1988)
Sapotaceae Argina spinosa (fruits and seeds)
Heim de Balsac & Mayaud (1962)
Solanaceae Lycium spp. (fruits) Lycium afrum (fruits) Lycium intricatum (fruits)Heteracia szovitsii (fruits)
Collins (1984; 1993) von Thanner (1912; 1913) Bannerman (1963)
Valverde (1957)
Gubin & Mukhina (in prep.)
Scrophulariaceae Antirrhinum ramosissium (flowers, fruits and seeds)
Goriup & Taylor (1983) (observation)
Tiliaceae Grewia populifolia (fruit)
Glutz von Blotzheim e ta l (1973), Ali & Ripley (1969)
Umbelliferae Bupleurum sp. Lavauden (1914)
Zygophyllaceae Fagonia sp. (seeds) Fagonia sp. (fruits and flowers) Zygophyllum sp. (seeds)
Gillett (1988), Gaucher (1991)
Mirza (1971)
Pavlenko (1962), Meklenburtsev (1990)
Table 2.2 (continued)
List of flowers, fruits and seeds eaten by the three subspecies of houbara

40
Prey item Chlamydotis undulata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
Invertebrates Hooker (1958) Gallagher (unpubl.)
Insecta
Insect larvae Larvae on carrion
Bédé (1928), Brosset (1961)
Roberts & Savage (1971), Dharmakumarsinhji (1955), Mian (1988)Fox (1988)Sushkin (1908)
Orthoptera Acrididae Grasshoppers and locusts
Cramp & Simmons (1980), Collins (1984,1993), Valverde (1957)
Toschi (1969), Anon (1980)
Boehme (1926), Meinertzhagen (1954), Ali & Ripley (1969), Mirza (1971), Glutz von
Sphingonotus caerulans Cedopodinae sp.
Tettigoniidae, Bush crickets Gryllotalpidae, Molecrickets
Heim de Balsac (1926),
Blotzheim et a! (1973), Mian (1988), Gubin & Mukhina (in prep.). Dement’ev & Gladstock (1951), Fox (1988), Pavlenko (1962), Salikhbaev & Ostapenko (1967), Gavrin (1962), Symens (1988a) (captive) Glutz von Blotzheim e ta ! (1973)
Glutz von Blotzheim et at (1973)Glutz von Blotzheim et al (1973)
Mian (1988), Roberts & Savage (1971)
Odonata Gubin & Mukhina (in prep.)
Isoptera, Termites Salikhbaev & Ostapenko (1967), Ali & Ripley (1969), Glutz von Blotzheim e ta l (1973), Gubin & Mukhina (in prep.)________________
ColeopteraBeetles
Webb & Berthelot (1836- 44), von Thanner (1912; 1913), Volsoe (1951).
Timarcha pygidium Pentodon ciliata
Valverde (1957), Anon (1980), Goriup & Taylor (1983)
Lavauden (1914)
Meinertzhagen (1954), Rustamov (1954), Ali & Ripley (1969), Mirza (1971), Glutz von Blotzheim et a / (1973), Mian (1988), Roberts(1991), Goriup & Norton(1992)
Glutz von Blotzheim et al (1973)__________________
Cerambycidae Dorcadion sp. Plocaederus scapularis Acinopus striolatusCarabidae Carabus sp.
Cymindis sp.______
Gavrin (1962)Gubin & Mukhina (in prep.)
Gubin & Mukhina (in prep.)
Glutz von Blotzheim e ta l (1973)
Gubin & Mukhina (in prep.)
Table 2.3
List of invertebrates eaten by the three subspecies of houbara

41
Prey item Chlamydotis undulata Chlamydotis undulata Chlamydotis undulatafuertaventurae undulata macqueeni
CarabidaeHarpalus sp. Glutz von Blotzheim e ta l
(1973)Machozetus lehmanni Gubin & Mukhina (in prep.)
Scarites bucida Gubin & Mukhina (in prep.)
Scarites terricola Gubin & Mukhina (in prep.)
Scarabaeidae Ali & Ripley (1969), Glutz von Blotzheim et al. (1973), Gubin & Mukhina (in prep.)
Dung beetles Webb & Berthelot (1836- Sushkin (1908)1844), Polatzek (1909), Volsoe (1954), Bannerman (1963)
Copris lunaris Glutz von Blotzheim e ta l(1973)
Aethiesa szekessy Gubin & Mukhina (in prep.)
Ootoma bipartita Collins (1984,1993)
Scarabaeus Fox (1988)
gangeticusScarabaeus sp. Gubin & Mukhina (in prep.)
Chafers Heim de Balsac (1926), Valverde (1957)
Gubin & Mukhina (in prep.)
Chineosoma Gubin & Mukhina (in prep.)
kizilcumensePhaeadoretus Gubin & Mukhina (in prep.)
comptusMelanotha sp. Alekseev (1985)
BuprestidaeJulodis variolaris Gubin & Mukhina (in prep.)
Julodis onopordi Toschi (1969)
Chrysomelidae Glutz von Blotzheim etal.(1973)
Barathaea spp. Toschi (1969)
Staphylinidae Glutz von Blotzheim et al.(1973)
Curcuiionidae Collins (1984; 1993) Glutz von Blotzheim et al. (1973), Goriup & Norton (1992), Fox (1988), Anegay (1992), Gavrin (1962)
Ammocleonus sp. Fox (1988)
Cleonus sp. Gavrin (1962)
Chromosomus sp. Fox (1988)
Conorhynchus Collins (1984, 1993)
conirostrisLixus sp. Gubin & Mukhina (in prep.)
Tenebrionidae Gillett (1988), Gaucher (1991)
Sushkin (1908), Kostin (1956), Ali & Ripley (1969), Glutz von Blotzheim etal. (1973), Alekseev (1985), Anegay (1992), Gavrin (1962)
Anatolica sp. Gavrin (1962)
Table 2.3 (continued)
List of invertebrates eaten by the three subspecies of houbara.

42
Prey item
Tenebrionidae Adesmia sp. Adesmia aenescens Adesmia fagergreeni Adesmia panderi Akis sp.Arthrodesis sp.Blaps sp.
Pimelia bengasina Pimelia interpunctata Pimelia indica Pimelia inexpectata Pimelia sp. Prionotheca coronata Mesostena sp. Scorus sp.Thryptera kraatzi Tentyria sp. Trachyderma sp. Trachyderma robusta Zophosis plicataMeloidaeDermestidaeElateridae Mylabris sp.SilphidaeCantharidae
Chlamydotis undulata Chlamydotis undulata Chlamydotis undulatafuertaventurae undulata macqueeni
Symens (1987) (captive) Fox (1988)Fox (1988)Gubin & Mukhina (in prep.) Anegay (1992)Fox (1988)
Lavauden (1914) Glutz von Blotzheim etal.(1973), Fox (1988), Gubin & Mukhina (in prep.), Gavrin (1962)
Toschi (1969)Toschi (1969)
Fox (1988)Fox (1988)Anegay (1992)Anegay (1992)Anegay (1992)Anegay (1992)Fox (1988)Gavrin (1962)Anegay (1992)Fox (1988)
Gavrin (1962)
Gubin & Mukhina (in prep.)
Gubin & Mukhina (in prep.)
Samarin et a i (1986)
Ali & Ripiey (1969), Giutz ____________________________ von Biotzheim et a / (1973)
Coiiins(1984, 1993)
Lepidopteran larvae
Agrotis segetum Trachea atriplicisAdult Lepidoptera
von Thanner (1912,1913) Aiekseev (1985), Gubin & Mukhina (in prep.)Giutz von Biotzheim et al. (1973)Glutz von Blotzheim et al. (1973)
Fox (1988)
Hemiptera (Shield bugs)
Gubin & Mukhina (in prep.) Fox (1988)
Hymenoptera
Apidae, Bees
Vespoidea (wasps)
Ichneumonidae
Gubin & Mukhina (in prep.)Glutz von Blotzheim etal. (1973)
Zhuyuko (1986)
Table 2.3 (continued)
List of invertebrates eaten by the three subspecies of houbara

43
Prey item Chlamydotis undulata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
HymenopteraFormicidae, Ants
Camponotus sp. Messor maurus
Collins (1984,1993)
Collins (1984,1993)
Valverde (1957), Lavauden (1914), Ehrenberg in Neumann, (1834), Goriup & Taylor (1983), Gillett (1988), Gaucher (1991).
All & Ripley (1969), Roberts & Savage (1971), Glutz von Blotzheim etal. (1973), Mian (1988), Roberts (1991), Goriup & Norton (1992), Fox (1988), Anegay (1992), Gubin & Mukhina (in prep.)Anegay (1992)
Diptera, True flies Gubin & Mukhina (in prep.)
Arachnida Cramp & Simmons (1980) Brosset (1961)
Araneae, Spiders
Latrodectustredecimguttata(Cocoons)
Glutz von Blotzheim et al (1973), Anon (1980)
Pavlenko (1962), Meklenburtsev (1990)
Scorpiones ScorpionsCompsobuthusarabicas
Fox (1988)Anegay (1992)
Solifugae Camel spiders Gaieodes sp.
Anegay (1992), Gavrin (1962)
Gubin & Mukhina (in prep.)
Galeodes grantii (?) Anegay (1992)
CrustaceaIsopoda Armadillium sp.
Hemilepistus elegansAlekseev (1985), Rustamov (1954) Gubin & Mukhina (in prep.)
Chilopoda,Centipedes
Ali & Ripley (1969), Gubin & Mukhina (in prep.)
MolluscaGastropoda(Pulmonata)Snails
Theba (Helix) pisana Helix sarcostema
Meade-Waldo (1889a), von Thanner (1912,1913), Cramp & Simmons (1980), Collins (1984, 1993) (anecdotal)Hooker (1958), Lack (1983) (anecdotal)Hooker (1958)
Glutz von Blotzheim at a! (1973), Anon (1980)
Ali & Ripley (1969).
Table 2.3 (continued)
List of invertebrates eaten by the three subspecies of houbara
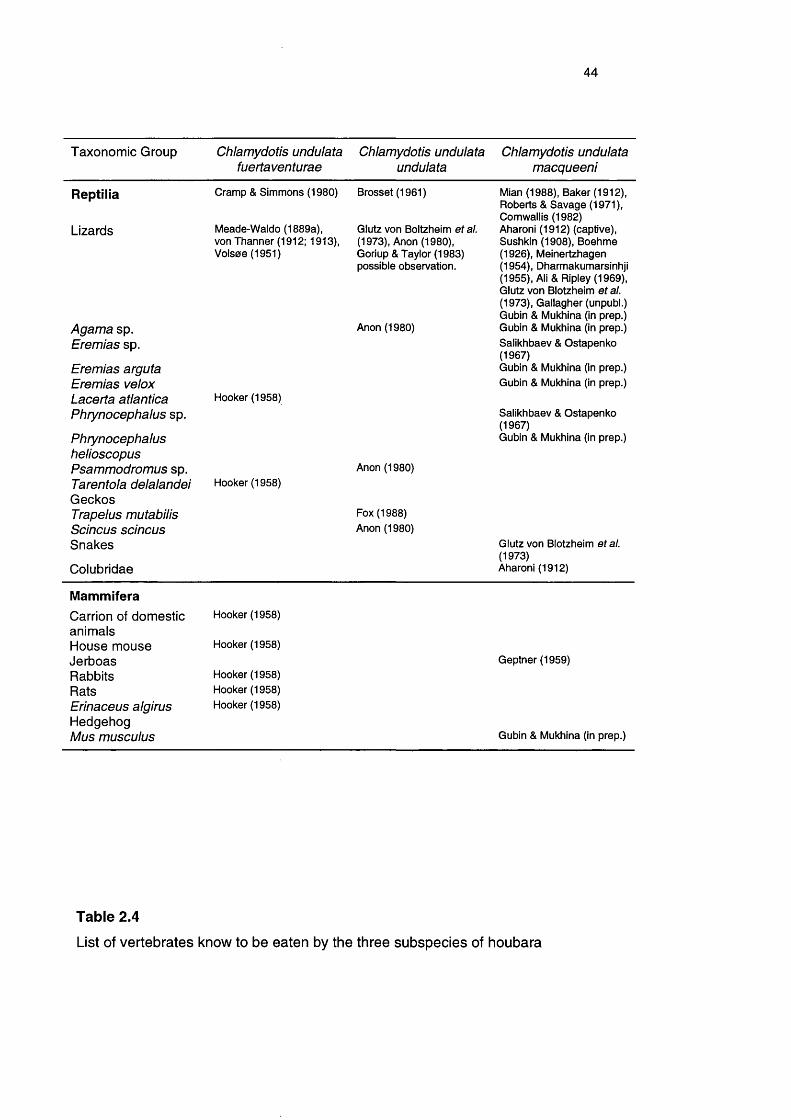
44
Taxonomie Group Chlamydotis undulata fuertaventurae
Chlamydotis undulata undulata
Chlamydotis undulata macqueeni
Reptilia
Lizards
Cramp & Simmons (1980) Brosset (1961)
Meade-Waldo (1889a), von Thanner (1912; 1913), Volsoe (1951)
Agama sp.Eremias sp.
Eremias arguta Eremias velox Lacerta atlantica Phrynocephalus sp.
Phrynocephaius helioscopus Psammodromus sp. Tarentola delalandel GeckosTrapelus mutabilis Scincus scincus Snakes
Colubridae
Glutz von Boitzheim et al. (1973), Anon (1980), Goriup & Taylor (1983) possible observation.
Anon (1980)
Hooker (1958)
Hooker (1958)Anon (1980)
Fox (1988) Anon (1980)
Mian (1988), Baker (1912), Roberts & Savage (1971), Cornwallis (1982)Aharoni (1912) (captive), Sushkin (1908), Boehme (1926), Meinertzhagen(1954), Dharmakumarsinhji(1955), Ali & Ripley (1969), Glutz von Blotzheim et al. (1973), Gallagher (unpubl.) Gubin & Mukhina (in prep.) Gubin & Mukhina (in prep.) Salikhbaev & Ostapenko (1967)Gubin & Mukhina (in prep.) Gubin & Mukhina (in prep.)
Salikhbaev & Ostapenko (1967)Gubin & Mukhina (in prep.)
Glutz von Blotzheim etal. (1973)Aharoni (1912)
MammiferaCarrion of domesticanimalsHouse mouseJerboasRabbitsRatsErinaceus algirus Hedgehog Mus musculus
Hooker (1958)
Hooker (1958)
Hooker (1958) Hooker (1958) Hooker (1958)
Geptner (1959)
Gubin & Mukhina (in prep.)
Table 2.4
List of vertebrates know to be eaten by the three subspecies of houbara

45
Animal foods
Animals are said to be important in the diet, and when pack-animals were
common in the Canaries, houbara visited their tracks to feed on dung-beetles
attracted by excrement (Webb & Berthelot 1836-44; Polatzek 1909). Von
Thanner (1913) states that houbara eat snails, beetles, caterpillars and lizards,
and older literature often mentions snails as a favourite food (Meade-Waldo
1889a; von Thanner 1912), identified by Hooker (1958) as Helix pisana and
Helix sarcostema (both Mollusca:Gastropoda). More recently, local wardens told
Lack (1983) that houbara ate Theba (=Helix) pisana (Müller), but he never saw
this himself. Likewise, local people told Collins (1984) that snails were eaten by
houbara, but he found no trace of snails in 106 houbara faeces.
Plant foods
Several authors list the plants eaten by houbara on Fuerteventura, including a
trefoil (Leguminosae) (either alfalfa Medicago sativa or the native Medicago
minima), a Mesembryanthemum sp. (Aizoaceae) possibly Mesembryanthemum
nodiflorum (Meade-Waldo 1889b; Collins 1984), Lycium a/rum fruits (von
Thanner 1912), Launaea flowers (Valverde 1957), a yellow herb Hirschfeldia
incana (Cruciferae) and perhaps other common shrubs like Launaea
arborescens (Compositae) and Salsola vermiculata (Chenopodiaceae) (Lack
1983). There are also reports of houbara feeding in and damaging crops such
as peas, lentils and cereals (Polatzek 1909; von Thanner 1912, Lack 1983).
Farmers told Collins (1984) that houbara ate chick-peas, lentils and alfalfa
especially in the dry season, but he did not observe this himself during a two
year field-study.
Studies based on faecal analvsis
The most extensive study of the diet of C. u. fuertaventurae is by Collins (1984,
1993) who examined 106 faeces from two field seasons. The majority were
collected at the roost sites of two males (45% and 19% of samples) which
biases his conclusions. He had difficulty in estimating the volume of different
foods because there were no data on their rates of digestion, but estimated the
number of some insects by counting remains of heads, hind tibiae or femora.

46
He estimated the percentage of plant matter and identified plant remains where
possible, but may have under-recorded easily digested foods.
He divided food into four main food types: insects, annual plants, Launaea
arborescens flowers and Lycium sp. fruits. Insects were present in 85% of
faeces and the most frequent were Conorhynchus conicirostris
(ColeopteraiCurculionidae), Zophosis plicata (ColeopteraiTenebrionidae) and
Messor maurus (HymenopteraiFormicidae), with a few Ootoma bipartita
(ColeopteraiScarabaeidae) and grasshoppers. Annual plants were present in
79% of faeces and were identified from their flowers except for Medicago
minima which had distinctive leaves. They included Lobularia lybica (Cruciferae)
(54%), Notoceras bicorne (Cruciferae) (43%) and Medicago minima
(Leguminosae) (43%). Flowers and buds of the spiny shrub Launaea
arborescens (Compositae) were found in 55% of faeces, while fruits of the
thorny shrub Lycium sp. (Solanaceae) occurred in 49% of faeces. Other plant
remains included fragments of Calendula aegyptiaca (Compositae) (20%),
grasses (14%), a Compositae (13%), Erodium cicutarium (Geraniaceae) (10%),
a Cruciferae (4%), Plantago spp. (Plantaginaceae) (3%), Mesembryanthemum
spp. (Aizoaceae) (2%) and Astragalus hamosus (Leguminosae) (1%).
Samples from the two birds that contributed 64% of faeces contained an above
average proportion of weevils (Curcuiionidae), which may reflect the frequency
of weevils in their home ranges or a food preference. Even though many foods,
appeared plentiful some faeces contained only limited items, suggesting
possible prey preferences. For example, there were over 600 Lycium sp. fruits,
150 Zophosis plicata beetles and ten weevils in one faecal sample. During the
rainy season, the estimated amount of plant matter decreased from 82% to 58%
in the bird which contributed 45% of samples, contrasting with Heim de Balsac
& Heim de Balsac’s (1954) suggestion that houbara eat more vegetable than
animal matter during the spring. More Zophosis plicata and fewer ants were
found in the faeces in the wet season, which may indicate a switch from ants to
tenebrionids, since ants were numerous throughout the year while beetles were
only seasonally abundant.

47
Some faeces contained larvae of coprophagous invertebrates (dung-flies and
beetles) which were probably attracted to the faeces rather than being eaten by
the houbara.
Feeding behaviour of C.u.fuertaventurae
The feeding behaviour of C.u.fuertaventurae has been observed by a few
authors. Von Thanner (1913) noted that houbara were inactive during the
hottest part of the day, and Hinz & Heiss (1989) confirmed that they were least
active at noon, and fed mainly in the morning (06.00-8.00 hrs) or afternoon and
early evening (16.00-18.00 hrs). Houbara often stayed in one area, particularly
fields of alfalfa, lentils and chick-peas, where they ate insects and plants. Collins
(1984) described their feeding behaviour, but was unable to get close enough to
see what they were eating because they were easily disturbed.
2.2.2 The North African houbara: Chlamydotis undulata undulata
General descriptions of the diet
C.u.undulata is the least known of the three subspecies and information on its
diet is lacking. General descriptions include plants and insects (Button 1835;
Bédé 1928; Heim de Balsac & Mayaud 1962), and Argania spinosa
(Compositae) fruits in the dry season (Heim de Balsac & Mayaud 1962).
Descriptions of single gizzard contents include; green plants, bush-crickets
(Orthoptera) and large chafers (Scarabaeidae) in the spring (Heim de Balsac
1926); green plants (40%), >100 ants (30%), beetles (20%), and fruits of
Lycium sp. (10%) in June (Valverde 1957); insects, arachnids, reptiles, berries
and young green shoots (Brosset 1961) and ants (Glutz von Blotzheim et ai.
1973).
Other foods include cereals, seeds, pistachio nuts, berries, spiders, snails,
lizards and snakes (Glutz von Blotzheim et ai. 1973) and Launaea sp. flowers
(Valverde 1957), while houbara chicks hunt locust hoppers and eat tomato
leaves (Valverde 1957). Anon (1980) mentions that houbara eat young grass
shoots and the leaves of shrubs in the spring, while in summer and autumn they
consume wild seeds, figs and other windfall fruit in orchards. Small snails.

48
grasshoppers, spiders and beetles, and small lizards such as Psammodromus
sp., Agama sp. and skinks Scincus scincus are also mentioned (Anon 1980).
However, this may be a summary of potential food, since Combreau &
Rambaud (1995) were surprised that C.u.macqueeni gizzards (n=11) did not
contain any of the locally abundant skinks.
Gaucher (1991) tracked an incubating female in the Algerian Sahara and found
feeding damage to buds, fruits and flowers of Farsetia stylosa (Cruciferae),
Argyrolobium unifolium (Leguminosae), Fagonia glutinosa (Zygophyllaceae) and
Heiianthemum ruficomum (Cistaceae), and to fresh shoots of Haloxylon sp.
(Chenopodiaceae). He saw signs of feeding at ants' nests and thought that the
female foraged close to her nest rather than searching for more distant prey.
Observations of houbara in Morocco during April suggested that they often ate
the fruits and flowers of two shrubs, Zilla spinosa (Leguminosae) and Farsetia
hamiltonii (Cruciferae) (Goriup & Taylor 1983). After the rains houbara also
consumed the fruits and flowers of Antirrhinum ramosissium (Scrophulariaceae)
and foraged on the ground, possibly catching lacertid lizards. Gizzards
contained fruits and flowers of Farsetia hamiltonii, flowers and leaves of
Launaea sp. (Compositae), dates, other plant remains, ants and beetles.
Gizzard content analvsis
Lavauden (1914) described two gizzards from houbara in Tunisia. The first
contained two Blaps sp. (Coleoptera:Tenebrionidae), two Timarcha sp.
(Coleoptera:unknown), three “grains”, two umbellifer seeds (probably
Bupleurum sp.) and a green vegetable pulp, possibly young barley shoots. The
other gizzard contained a green pulp, two ants, one Timarcha pygidium and five
“grains”. In March at Bengazi-Agedatia, Toschi (1969) found the following
insects in houbara gizzards: Jolodis onopordi (Coleoptera;Buprestidae), another
buprestid, Pimelia bengasina, Pimelia interpunctata (Coleoptera:
Tenebrionidae), Barathaea sp. (Coleoptera:Chrysomelidae) and grasshoppers
or locusts (Orthoptera:Acrididae).
Gillett (1988) examined 23 gizzards from Algerian houbara in December 1987,
which contained tenebrionids and ants, although he only identified plant

49
remains. There were many fiower-buds of the small shrub Farsetia stylosa
which have high levels of protein (21.1% dry weight) and minerals (20.6% dry
weight). Shoots and fruits of perennial shrubs like Anabasis articulata
(Chenopodiaceae) and Fagonia sp. (Zygophyllaceae), and leaves of Matricaria
sp. (Compositae) were also consumed. He suggested that houbara select
young plant growth such as seedlings, flower-buds and shoots following the
rains, since annuals like Poiycarpaea sp. (Caryopyllaceae) and Trifolium sp.
(Leguminosae) appeared in samples at this time. Gizzards also contained fruits
of Zizyphus lotus (Rhamnaceae) and Pistacia atlantica (Anacardiaceae). Fox
(1988) examined gizzard contents of Algerian houbara which contained lizards
{Trapelus mutabilis) and a scorpion (probably Buthidae). According to his
previous experience. Fox thought that C.u.undulata consumed more reptiles
than C.u.macqueenI.
2.2.3 MacQueen’s bustard: Chlamydotis undulata macqueeni
C.u.macqueenI \s generally migratory, has a wider distribution and is more
common than the other houbara subspecies (Osborne 1996a). Its long
association with Arab falconry has attracted much attention. There are detailed
studies of its diet from several parts of the range particularly the Middle East,
India and Pakistan, and Central Asia, which are considered below. Note
however, that the ranges of some of these populations overlap (see discussion
of houbara distribution in Osborne 1996a).
The Middle Eastern population
General descriptions of the diet include Artemisia sp. shoots, allium bulbs,
beetles (Tenebrionidae), lizards, grasshoppers and locusts (Meinertzhagen
1954), while snakes (colubrids) were a favourite food of houbara in Der-es-Zor
on the Euphrates (Aharoni 1912). In Harrat Al-Harrah, Saudi Arabia, Symens
(1988b) noted that houbara nested in areas with diverse vegetation. Females
and chicks remained close to their nest for over a week after hatching, and ants
appeared to be an important food. Nothing is published on the diet of the
resident houbara of Jiddat AI Harasis in Oman, but potential foods include the
leaves, shoots and corns of Fagonia sp. (Zygophyllaceae), Crotalaria sp.
(Leguminosae), Tribulus sp. (Zygophyllaceae) and Farsetia sp., and

50
invertebrates and small lizards (M. D. Gallagher unpubl. MS). Lavee (1985)
trapped houbara using baby-mice, fly larvae and Blaps sp. (Coleoptera:
Tenebrionidae) but these may not be normal constituents of the diet.
Studies based on faecal analvsis
In Saudi Arabia, Goriup & Norton (1992) noted that houbara foraged in well-
vegetated parts of sabkhas. Ten houbara faeces were examined, of which
seven consisted solely of plant matter, while three contained only insect
fragments. Plant remains included fruits and leaves of Farsetia aegyptia
(Cruciferae), fruits of Medicago lacinata (Leguminosae) and grass spikelets,
while several faeces only contained flower petals (probably Achillea
fragrantissima, Compositae). The faeces containing insects were composed
almost entirely of ants, with parts of a weevil and a large beetle. Goriup &
Norton (1992) suggested that houbara may develop a search image for a
specific food item, such as ants, because of individual preferences or locally
abundant food sources.
The Pakistani and Indian populations
There is much dietary data from this region, including descriptions of gizzard
contents and known foods. However, many studies lack details of methodology
or data analysis, and have not been peer-reviewed. As in the Canary Islands,
houbara feed on crops and may cause considerable damage (Baker 1912), and
crops are used as a lure when catching houbara (P. E. Osborne pers. comm.).
General description of the diet
Indian houbara are said to be omnivorous with a preference towards green
plants, and only eating reptiles when very hungry (Baker 1912). In Saurashtra,
houbara have been recorded feeding upon insects, lizards, Capparis sp.
(Capparaceae) fruits, Leptadenia sparticum (Asclepiadaceae), Triumfetta
rotundifolia (Tiliaceae), mustard, manna {Alhagi mauraurum Leguminosae),
seeds and grains (Dharmakumarsinhji 1955). Ferguson-Lees (1969) states that
houbara over-wintering in India feed on plants, especially Artemisia sp., berries
and seeds of desert plants, wild onions and crops such as mustards, oilseeds

51
and wheat. Ali & Ripley (1969) mention other foods including the fruits of
Zizyphus jujuba (Rhamnaceae) and Grewia populifolia (Tiliaceae), as well as
ants, termites, locusts, grasshoppers, beetles (Tenebrionidae, Scarabaeidae,
Cantharidae and others), and more rarely centipedes, lizards and snails.
Houbara eat seeds and green plants (up to 30% of the diet) including crops,
e.g. Brassica juncea (L.) (Cruciferae) and Cyamopsis tetragonoloba (L.)
(Leguminosae), combined with insects (such as mole-crickets (Orthoptera:
Gryllotalpidae) and ants) and small reptiles (Roberts & Savage 1971). Game-
wardens and hunters thought that over-wintering houbara were omnivorous,
eating seeds, young shoots and slow-moving insects, while in southern and
eastern Baluchistan houbara probably consumed beetles, mole-crickets, ants
and grasshoppers (Mian 1988).
Plant foods
Several authors list plants eaten by the houbara but some may be potential
rather than proven foods. Mian & Surahio (1983) report that in Dak, Zangi
Nower and Kharan, houbara eat young shoots of Calligonum comosum
(Polygonaceae), Koeleria phleoides (Gramineae), Malcolmia africana
(Cruciferae), Haloxylon ammodendron (Chenopodiaceae), Ephedra sp.
(Ephedraceae), Salsola arbuscula (Chenopodiaceae) and Polygonum
afghanicum (Polygonaceae) as well as the dry seeds of Polygonum afghanicum,
Calligonum comosum, Haloxylon ammodendron and Panlcum sp. (Gramineae).
In the Chagai and Yakmuch plains, plants such as Tribulus terrestrls, Tribulus
alatus (Zygophyllaceae), Erodlum sp. (Geraniaceae), Plantago dilata
(Plantaginaceae), Rhazya strlcta (Apocynaceae) and Holosteum umbellatum
(Caryophyllaceae) were probably eaten. Mirza (1985) suggests that food plants
in Pakistan include Leptadenia sp. (Asclepidaceae), Cymbopogon sp.
(Gramineae) and Euphorbia prostrata (Euphorbiaceae). Surahio (1985)
mentions that houbara in Sind (Pakistan) feed on Zizyphus numularla berries
(Rhamnaceae), Capparis aphylla (Capparaceae), Crotolarla sp., Haloxylon sp.,
Grewia populifolia and Tribulus sp., and that mustard crops attract high
numbers of foraging birds. Mian (1988) lists locally available food plants for
several regions of Baluchistan. In Zhob, Pishin, Nushki and Kharan, Salsola sp.,
Haloxylon sp.. Anabasis sp. (Chenopodiaceae), Malcolmia sp., and Tribulus sp.
were said to be important, while in the lowland deserts to the South and East

52
(Gwadar, Dera Bugti, Sibi, Kachi) houbara eat Zizyphus sp. berries, and
vegetative parts of Brassica campestris (Cruciferae), Capparis decidua
(Capparaceae) and Eruca sativum (Cruciferae).
Studies based on gizzard contents
Mirza (1971) examined 100 gizzards of houbara hunted using falcons in
Pakistan. Most contained a mixture of plant and animal matter, such as
grasshoppers and beetles, although about 12.5% consisted entirely of plant
material, including Fagonia sp. (Zygophyllaceae), Haioxylon sp.
(Chenopodiaceae), Farsetia sp. (Cruciferae), Zygophylium sp.
(Zygophyllaceae), Tribulus sp. (Zygophyllaceae), Crotaiaria sp. (Leguminosae)
and Eulaliopsis sp. (unknown). Fagonia sp. seeds occurred in 75% of samples.
Mirza (1985) also mentions a grass, Lasiurus sp., probably from the same
gizzards. Fox (1988) examined 52 gizzards from houbara in Chagai and
Dalbandin in Baluchistan, and Leiah and Rajanpur in the Punjab. They all
contained plants and insects, but vegetation made up 62.9% of the total dry
matter, particularly Farsetia jacquemontii {Cruc\ierae), Capparis sp.
(Capparaceae) and Tribulus terrestrls (Zygophyllaceae), which occurred in 74%,
74% and 84% of samples and contributed 26.5%, 11.9% and 9.9% to the total
dry matter respectively. Other plants included Haloxylon ammodenderon
(Zygophyllaceae), Zizyphus sp. fruits (Rhamnaceae), Euphorbia granulata
(Euphorbiaceae), Cadaba sp. (Capparaceae) and small amounts of chick peas,
water melon and sorghum. Insects contributed 31% of the dry weight with ants
in 84% of the samples. Beetles occurred in many samples, especially the
tenebrionids: Adesmia aenescens (22.6%), Pimelia indica or Pimelia
inexpectata (19.5%), Arthrodosis sp. (9.7%), Biaps sp. (6.5%), Thryptera sp. or
Trachyderma sp. (6.5%) and Adesmia fagergreeni (6.5%). Other beetles and
weevils occurred in 67.7% of gizzards and a few grasshoppers (16.1% of
samples), Hemiptera, adult Lepidoptera and insect larvae were also found.
Houbara gizzards from Pakistan contained leaves, shoots and ground-dwelling
insects such as ants and beetles (Roberts 1991). Seasonal variations in food
availability may influence the diet. In early winter, houbara gizzards contained
17% insects, increasing to 30% in mid-winter and 51% by late winter (although
there is no mention of how these figures were obtained). The amount and types

53
of plants consumed also varied, with a shift from the foliage of perennial shrubs
in early and mid-winter to newly sprouted herbs in the late winter. Houbara may
have a wide diet so that they can survive in arid regions by adapting to different
foods. Observations also suggest that houbara forage during the day
(particularly morning and evening) in undisturbed areas, but feed nocturnally
where human disturbance is high.
The Central Asian population
Central Asia probably contains the largest populations of houbara, including
important breeding grounds where houbara are locally abundant. These areas
facilitate the collection of dietary data since large sample sizes can be obtained
compared with areas frequented by low density migrant birds.
General descriptions of the diet
General descriptions of the diet include green plants, invertebrates (especially
tenebrionids) and lizards (Ponomareva 1982), "ber" fruits, Grewia sp. (Tiliaceae)
berries, lemon-grass and insects (Finn 1915), gerbils (Geptner 1959), lizards,
Tenebrionidae and dung-beetles, as well as carrion-inhabiting Silphidae
(Coleoptera) and insect larvae (Sushkin 1908; Samarin etal. 1986). In the
breeding grounds, houbara eat beetles (especially Scarabaeidae,
Tenebrionidae and Cantharidae) and Orthoptera (locusts, grasshoppers and
bush-crickets), plus a few ants, termites, lizards and small snakes (Glutz von
Blotzheim etal. 1973). In the stony desert of Betpak-Dala, Kazakhstan, houbara
were said to eat beetles, seeds and grasses (Afanas'ev & Sludskiy 1947), while
Boehme (1926) thought that houbara near the Iranian-Kazakhstan border at
Nakhichrev' ate lizards and grasshoppers.

54
Plant foods
Plant foods include Capparis sp. fruits (Capparaceae) in Aybugir, Uzbekistan
(Molchanov 1913), while Artemisia sp. (Compositae), wild onion and garlic (both Ailium
sp. Liliaceae) bulbs are eaten by houbara around the Aral Sea (Sushkin 1908;
Meinertzhagen 1954; Ferguson-Lees 1969; Glutz von Blotzheim etal. 1973). Indeed,
houbara flesh may become tainted by the mixture of pungent plants {Artemisia spp.
and Aliium spp.).
Studies based on gizzard contents
There are many descriptions of houbara gizzard contents from Central Asia, although
most consist of lists of prey from single birds (see summary in Table 2.5). In a few
cases, particular prey types dominated the gizzard contents suggesting a preference
or seasonal glut of food, especially grasshoppers (Acrididae) during the summer
(Bannikov & Skalon 1948; Pavlenko 1962; Piechocki 1968). Most gizzards contained a
mixture of plant and animals foods, and Tenebrionidae were frequently mentioned.
Studies based on faecal analvsis
Gubin & Mukhina's (in prep.) work is based on over 1000 faeces of breeding birds from
four seasons in the Kyzylkum desert, Kazakhstan. Most samples were collected from
breeding males, but there were few from females, chicks or non-breeding birds.
Relative proportions of the constituents of the faeces were estimated by eye and the
frequency of encounter for each prey item was noted. (However, rather optimistically
frequency is quoted to the nearest 0.1%). The species identified from the faeces are
listed in Tables 2.1 to 2.4. Plant material was most abundant in the spring, making-up
about 80% of volume of all the samples, but dropped in June (7%) and September
(20%). Beetles occurred in at least 50% of samples, and were the most frequent
invertebrates in faeces. The highest volumes of beetles were seen between May and
June (56%). Tenebrionids were mainly eaten in May and the most frequently
encountered species was Adesmia panderi (up to 25% of the volume of faeces).
Houbara also consumed weevils (Curculionidae) and scarabs, but rarely ate the large
and abundant Scarabaeus sp. beetles. Buprestidae occurred in samples during May
and June, with a few Carabidae, Cicindelidae and Meloidea, and single records of
Dermestidae and Elateridae.
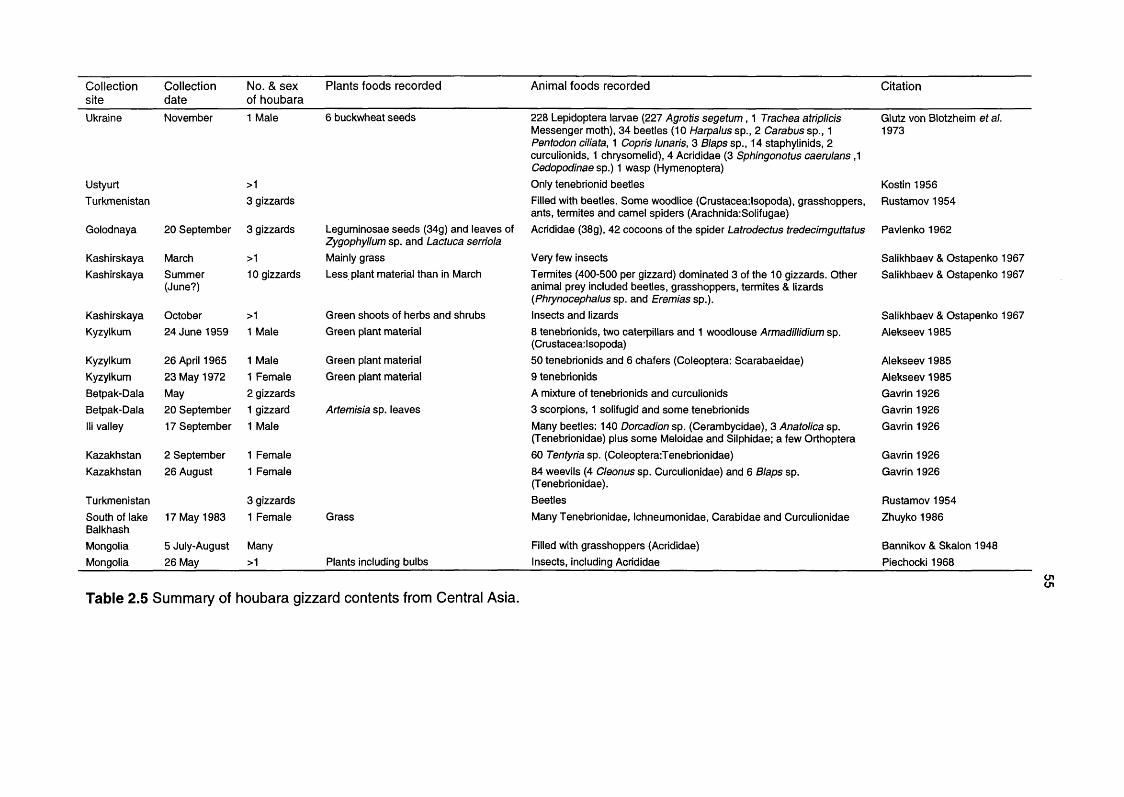
Collection Collection No. & sex Plants foods recorded Animal foods recorded Citationsite date of houbaraUkraine November 1 Male 6 buckwheat seeds 228 Lepidoptera larvae (227 Agrotis segetum, 1 Trachea atriplicis
Messenger moth), 34 beetles (10 Harpalus sp., 2 Carabus sp., 1 Pentodon cillata, 1 Copris lunaris, 3 Blaps sp., 14 staphylinids, 2 curculionids, 1 chrysomelid), 4 Acrididae (3 Sphingonotus caerulans ,1 Cedopodinae sp.) 1 wasp (Hymenoptera)
Glutz von Blotzheim et al. 1973
Ustyurt >1 Only tenebrionid beetles Kostin 1956
T urkmenistan 3 gizzards Filled with beetles. Some woodlice (Crustacea.lsopoda), grasshoppers, ants, termites and camel spiders (Arachnida:Solifugae)
Rustamov 1954
Golodnaya 20 September 3 gizzards Leguminosae seeds (34g) and leaves of Zygophylium sp. and Lactuca serriola
Acrididae (38g), 42 cocoons of the spider Latrodectus tredecimguttatus Pavlenko 1962
Kashirskaya March >1 Mainly grass Very few insects Salikhbaev & Ostapenko 1967
Kashirskaya Summer(June?)
10 gizzards Less plant material than in March Termites (400-500 per gizzard) dominated 3 of the 10 gizzards. Other animal prey included beetles, grasshoppers, termites & lizards (Phrynocephalus sp. and Eremlas sp.).
Salikhbaev & Ostapenko 1967
Kashirskaya October >1 Green shoots of herbs and shrubs Insects and lizards Salikhbaev & Ostapenko 1967
Kyzylkum 24 June 1959 1 Male Green plant material 8 tenebrionids, two caterpillars and 1 woodlouse Armadillidium sp. (CrustaceaJsopoda)
Alekseev 1985
Kyzylkum 26 April 1965 1 Male Green plant material 50 tenebrionids and 6 chafers (Coleoptera: Scarabaeidae) Alekseev 1985
Kyzylkum 23 May 1972 1 Female Green plant material 9 tenebrionids Alekseev 1985
Betpak-Dala May 2 gizzards A mixture of tenebrionids and curculionids Gavrin 1926
Betpak-Dala 20 September 1 gizzard Artemisia sp. leaves 3 scorpions, 1 solifugid and some tenebrionids Gavrin 1926
Hi valley 17 September 1 Male Many beetles: 140 Dorcadlon sp. (Cerambycidae), 3 AnatoHca sp. (Tenebrionidae) plus some Meloidae and Silphidae; a few Orthoptera
Gavrin 1926
Kazakhstan 2 September 1 Female 60 Tentyria sp. (Coleoptera:Tenebrionidae) Gavrin 1926
Kazakhstan 26 August 1 Female 84 weevils (4 Cleonus sp. Curculionidae) and 6 Blaps sp. (Tenebrionidae).
Gavrin 1926
Turkmenistan 3 gizzards Beetles Rustamov 1954
South of lake 17 May 1983 1 Female Grass Many Tenebrionidae, Ichneumonidae, Carabidae and Curculionidae Zhuyko 1986Balkhash
Mongolia 5 July-August Many Filled with grasshoppers (Acrididae) Bannikov & Skalon 1948
Mongolia 26 May >1 Plants including bulbs Insects, including Acrididae Piechocki 1968
Table 2.5 Summary of houbara gizzard contents from Central Asia.cncn

56
Gubin & Mukhina (in prep.) also found that arachnids were an important food,
as evidenced by the hard chelicerate mouthparts of Solifugidae, which occurred
in up to 80% of faeces in mid-June. Ants and termites were as important as
beetles early in the spring, contributing up to 19.7% by volume and occurring in
up to 50% of samples. Orthoptera, especially Acrididae occurred in over 30% of
samples in June (4% by volume). However, they are more easily digested than
beetles and may have been underestimated. In the heat of the summer ants
were one of the few invertebrate groups that did not decline. Woodlice were
eaten at a low frequency and volume, and Myriapoda, Mantodea, Lepidoptera
larvae, Diptera, Odonata, Apidae and Hemiptera were recorded infrequently.
Vertebrate prey were indicated by the remains of bones in faeces. For example,
a few lizard bones (see Table 2.4) were found in faeces from May and June,
and one faecal sample contained the tail of a house-mouse Mus musculus.
2.2.4 Vagrant houbara
Vagrants birds appear in areas outside of their normal range and have often
travelled great distances. When they arrive they are usually starving and out of
condition, and probably eat whatever is available. They are included in Table 2.6
for completeness, but are absent from other Tables.
Placename
List of food Citation
Yorkshire Gizzard contained mainly plant matter, particularly buds and flowers of Senecio aquaticus (Compositae).
Cordeaux 1896
Ukraine Gizzard contained many Lepidopteran larvae in November Glutz von Blotzheim etal. 1973
Suffolk Observed houbara eating winter-wheat, peas, mustard, barley stubble, earthworms and mice.
Axell 1964
UK Insects, including Orthoptera, Lepidoptera (yellow underwing larvae, Noctua pronuba (L.)), carabids and snails.
Roberts 1848; Witherby et al. 1943
Table 2.6 Summary of foods eaten by vagrant houbara

57
2.2.5 Gizzard stones
Houbara ingest small stones to aid the grinding action of their muscular gizzard
in common with other bird orders, such as Galliformes. Gizzard stones are
mentioned by Lavauden (1914) and Surahio (1985) while Fox (1988) found
stones in the 29% of 52 gizzards. They have also been found during post
mortems of captive and wild houbara in the UAE (NARC, unpublished data).
2.2.6 The diet of semi-captive houbara
There are many studies of wild houbara in captivity, and captive-bred birds in
quasi-natural conditions or managed habitats (summarised in Table 2.7). Some
pinioned, wild birds have been kept in areas of natural vegetation planted with
known food plants, and given ad lib supplies of concentrated food pellets,
supplements and irrigated alfalfa. Other free-flying, captive-bred birds have
been released into protected enclosures, with access to supplementary food,
managed habitats and natural foods. Many observational data have been
gathered, including feeding habits, but are likely to be biased towards foods that
are easily identified and to observations taken during daylight hours.
2.2.7 Diets for captive houbara
Various diets for captive houbara have been described (see Table 2.8). They
usually contain a mixture of what is thought to be required (based on studies of
poultry) and foods that can be obtained locally, particularly insects. They may
bear little resemblance to natural diets, for example, yellow mealworms
Tenebrio molitorare easily reared and are often given to houbara but are
unlikely to occur in the wild. Artificial diets are excluded from Tables 2.1 to 2.4
Several authors discuss imbalances in captive diets, such as deficiencies or
excesses of particular nutrients which cause diseases only observed in captivity.
Indeed, the low survival rate of captive chicks is probably diet related (Platt
1985; Nazarov 1992; Haddane 1985). Hermans (1988a) criticised houbara diets
at Taif as being too high in protein with a tendency to turn rancid. He also
suggested that the ratio of energy to protein should be altered according to
stages of growth and reproduction (Hermans 1988b & 1992).
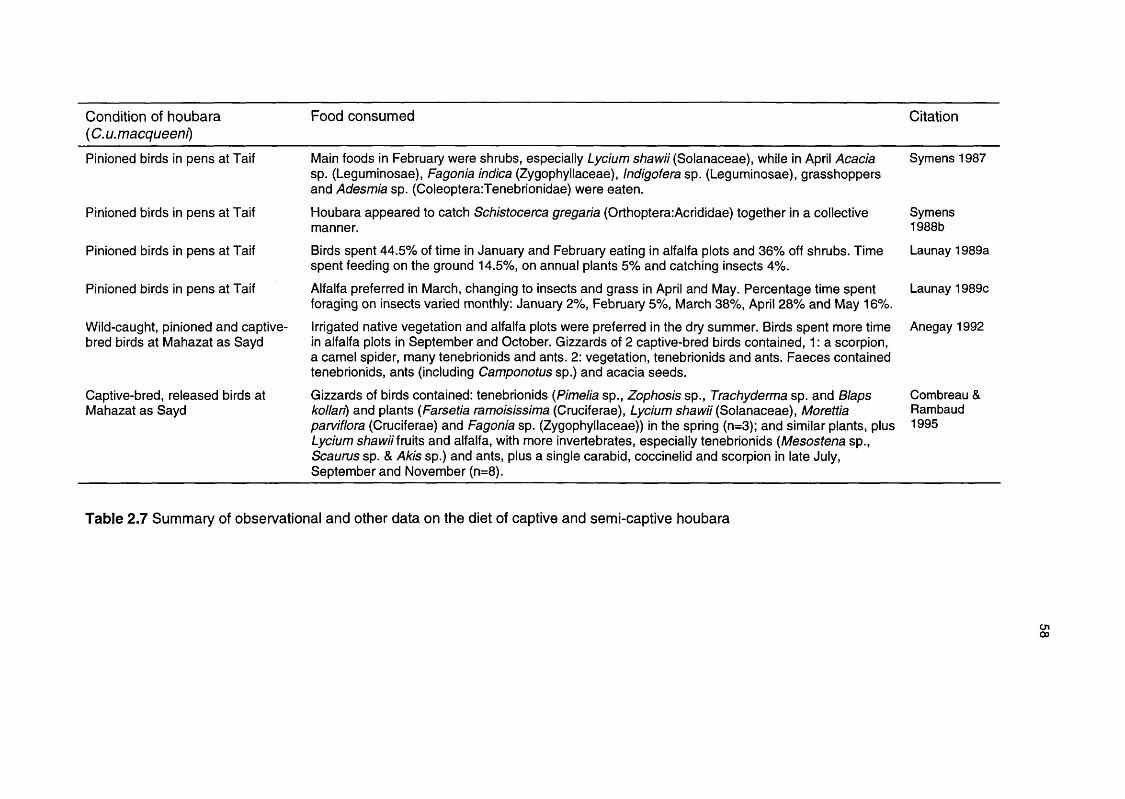
Condition of houbara {C.u.macqueeni)
Food consumed Citation
Pinioned birds in pens at Taif Main foods in February were shrubs, especially Lycium shawii (Solanaceae), while in April Acacia sp. (Leguminosae), Fagonia indica (Zygophyllaceae), Indigofera sp. (Leguminosae), grasshoppers and Adesmia sp. (ColeopteraiTenebrionidae) were eaten.
Symens 1987
Pinioned birds in pens at Taif Houbara appeared to catch Schistocerca gregaria (Orthoptera:Acrididae) together in a collective manner.
Symens1988b
Pinioned birds in pens at Taif Birds spent 44.5% of time in January and February eating in alfalfa plots and 36% off shrubs. Time spent feeding on the ground 14.5%, on annual plants 5% and catching insects 4%.
Launay 1989a
Pinioned birds in pens at Taif Alfalfa preferred in March, changing to insects and grass in April and May. Percentage time spent foraging on insects varied monthly; January 2%, February 5%, March 38%, April 28% and May 16%.
Launay 1989c
Wild-caught, pinioned and captive- bred birds at Mahazat as Sayd
Irrigated native vegetation and alfalfa plots were preferred in the dry summer. Birds spent more time in alfalfa plots in September and October. Gizzards of 2 captive-bred birds contained, 1: a scorpion, a camel spider, many tenebrionids and ants. 2: vegetation, tenebrionids and ants. Faeces contained tenebrionids, ants (including Camponotus sp.) and acacia seeds.
Anegay 1992
Captive-bred, released birds at Mahazat as Sayd
Gizzards of birds contained: tenebrionids {Pimelia sp., Zophosis sp., Trachyderma sp. and Blaps kollari) and plants {Farsetia ramoisissima (Cruciferae), Lycium shaw/V (Solanaceae), Morettia parviflora (Cruciferae) and Fagonia sp. (Zygophyllaceae)) in the spring (n=3); and similar plants, plus Lycium sham/fruits and alfalfa, with more invertebrates, especially tenebrionids {Mesostena sp., Scaurus sp. & Akis sp.) and ants, plus a single carabid, coccinelid and scorpion in late July, September and November (n=8).
Combreau &Rambaud1995
Table 2.7 Summary of observational and other data on the diet of captive and semi-captive houbara
cr«00
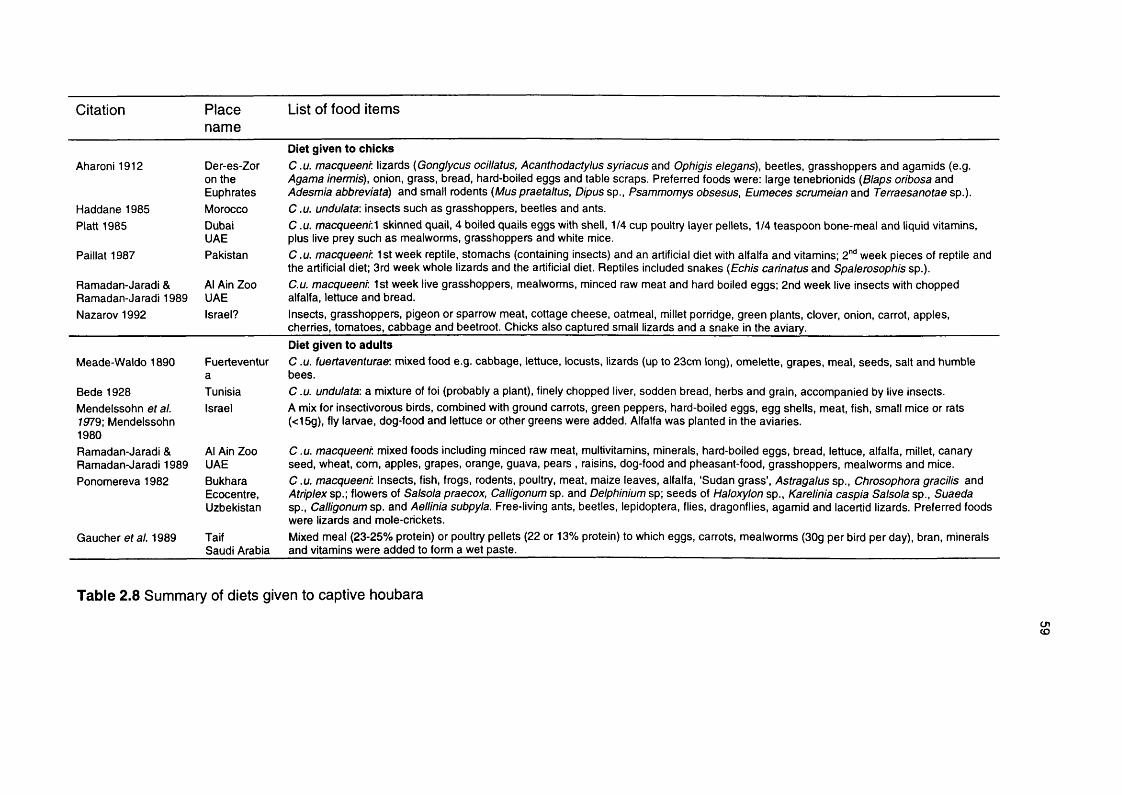
Citation Placename
List of food items
Aharoni 1912 Der-es-Zor on the Euphrates
Haddane 1985 Morocco
Platt 1985 DubaiUAE
Paillat 1987 Pakistan
Ramadan-Jaradi & AI Ain ZooRamadan-Jaradi 1989 UAE
Nazarov 1992 Israel?
Diet given to chicksC .u. macqueenh lizards {Gonglycus ocillatus, Acanthodactylus syriacus and Ophigis elegans), beetles, grasshoppers and agamids (e.g. Agam a inermis), onion, grass, bread, hard-boiled eggs and table scraps. Preferred foods were: large tenebrionids {Blaps oribosa and Adesmia abbrevlata) and small rodents {Mus praetaltus, Dipus sp., Psammomys obsesus, Eumeces scrumeian and Terraesanotae sp.).C .u. unduiata: insects such as grasshoppers, beetles and ants.
C .u. macqueeni-A skinned quail, 4 boiled quails eggs with shell, 1/4 cup poultry layer pellets, 1/4 teaspoon bone-meal and liquid vitamins, plus live prey such as mealworms, grasshoppers and white mice.
C .u. macqueenh 1st week reptile, stomachs (containing insects) and an artificial diet with alfalfa and vitamins: 2"' week pieces of reptile and the artificial diet; 3rd week whole lizards and the artificial diet. Reptiles included snakes {Echis carinatus and Spaierosophis sp.).
C.u. macqueenh 1st week live grasshoppers, mealworms, minced raw meat and hard boiled eggs; 2nd week live insects with choppedalfalfa, lettuce and bread.
Insects, grasshoppers, pigeon or sparrow meat, cottage cheese, oatmeal, millet porridge, green plants, clover, onion, carrot, apples, _________________cherries, tomatoes, cabbage and beetroot. Chicks also captured small lizards and a snake in the aviary._____________________________________
Diet given to adultsFuerteventur C .u. fuertaventurae: mixed food e.g. cabbage, lettuce, locusts, lizards (up to 23cm long), omelette, grapes, meal, seeds, salt and humble a bees.
Tunisia C .u. unduiata: a mixture of foi (probably a plant), finely chopped liver, sodden bread, herbs and grain, accompanied by live insects.
Israel A mix for insectivorous birds, combined with ground carrots, green peppers, hard-boiled eggs, egg shells, meat, fish, small mice or rats(<15g), fly larvae, dog-food and lettuce or other greens were added. Alfalfa was planted in the aviaries.
AI Ain Zoo C .u. macqueenh mixed foods including minced raw meat, multivitamins, minerals, hard-boiled eggs, bread, lettuce, alfalfa, millet, canaryUAE seed, wheat, com, apples, grapes, orange, guava, pears , raisins, dog-food and pheasant-food, grasshoppers, mealworms and mice.
Bukhara C .u . macqueenh Insects, fish, frogs, rodents, poultry, meat, maize leaves, alfalfa, ‘Sudan grass’. Astragalus sp., Chrosophora gracilis andEcocentre, Atripiex sp.; flowers of Salsola praecox, Calligonum sp. and Delphinium sp; seeds of Haloxylon sp., Karelinia caspia Salsola sp., SuaedaUzbekistan sp., Calligonum sp. and Aellinia subpyla. Free-living ants, beetles, lepidoptera, flies, dragonflies, agamid and lacertid lizards. Preferred foods
were lizards and mole-crickets.
Taif Mixed meal (23-25% protein) or poultry pellets (22 or 13% protein) to which eggs, carrots, mealworms (30g per bird per day), bran, mineralsSaudi Arabia and vitamins were added to form a wet paste.
Meade-W aldo 1890
Bede 1928
Mendelssohn et al. 1979] Mendelssohn 1980
Ramadan-Jaradi & Ramadan-Jaradi 1989
Ponomereva 1982
Gaucher e ta l. 1989
Table 2.8 Summary of diets given to captive houbara
( j tCO

60
Bailey (1992) noted that many bustard and stone curlew Burhinus oedicemus
chicks in AI Ain Zoo suffered from deformities associated with too rapid growth
due to excessive protein, possible calcium and phosphorus imbalances and
inadequate exercise. Furley, C.W., Greenwood, A.G. & Solomon, L. (unpubl.)
reported that overfeeding with a high protein diet resulted in obese and
unhealthy birds. They performed post mortems on two houbara which showed
excessive abdominal fat bodies and fatty livers. The intestine of one bird was
completely embedded in semi-necrotic fat, impeding its natural motility.
Anderson (1995) explains the role of micronutrients, such as vitamins, minerals
and amino acids, in the diet of captive bustards. The tendency to simplify
captive diets often results in a limited number of food types compared with wild
diets and therefore specific micronutrients may be lacking.
2.3 Discussion
2.3.1 Relative merits of techniques used to study the diet
A variety of methods have been used to investigate the diet of the houbara, but
all suffer from inherent bias. The simplest way is to interview local people who
are familiar with the houbara, and to catalogue locally available foods (Mian &
Surahio 1983; Mirza 1985; Surahio 1985). However, this only identifies possible
foods and may miss those that are inconspicuous or nocturnal, and gives little
indication of the relative importance of different food items.
Another approach is to observe houbara in the field and calculate a time budget
for different items in the diet (Hinz & Heiss 1989; Launay 1989a, b & c).
However, problems arise when the observer cannot see what the bird is eating
and are most problematic for animal prey. Another disadvantage is that houbara
are very wary and may not behave normally (see Collins 1984). Most
observational data are from semi-captive birds and their behaviour may differ
from wild birds (Table 2.7).
A better approach is to examine gizzard contents, although this necessitates
killing or examining dead birds. However, the diet of the latter may differ from
that of healthy birds if they were weak or sick before they died. Most studies are
based on single houbara (Valverde 1957; Webb & Berthelot 1836-1844;

61
Lavauden 1914; Table 2.6), although a few researchers have obtained gizzards
from hunting expeditions, e.g. Mirza (1971) and Fox (1988).
Examination of faeces is better than other methods because it does not harm
the birds. It also allows repeat sampling from individuals, although
pseudoreplication must be considered in the analysis. Collins (1984,1993) and
Gubin & Mukhina (in prep.) have examined faeces from C.u.fuertaventurae and
C.u.macqueen/respectively. However, partially digested fragments may be
difficult to identify, and easily digested, soft-bodied arthropods and fleshy plant
structures may be underestimated (Collins 1984, 1993; Goriup & Norton 1992;
Gubin & Mukhina, in prep.).
2.3.2 A comparison of the diet of the three houbara subspecies
The taxa recorded as food items for wild houbara are listed in Tables 2.1 to 2.4.
However, I also wanted to use the frequency of citations of foods to indicate the
importance of each taxon and to compare between the subspecies. The
majority of the citations are for C.u.macqueeni, so a system based solely on the
number of citations would underestimate the importance of the foods of
C.u.fuertaventurae and C.u.unduiata. A less biased index was calculated using
the number of taxa recorded for each subspecies, the number of authors who
have recorded the diet for that subspecies and the frequency of citations for
each taxon. This weighted percentage of citations, referred to as the citation
index, was calculated from Tables 2.1 to 2.4 via the following equation:
Citation Index per taxon =
Number of citations for that food taxon x 100(Total citations per subspecies x Total taxa in the table)
Values for the citation index of plant and animal foods are listed in Tables 2.9
and 2.10 respectively.
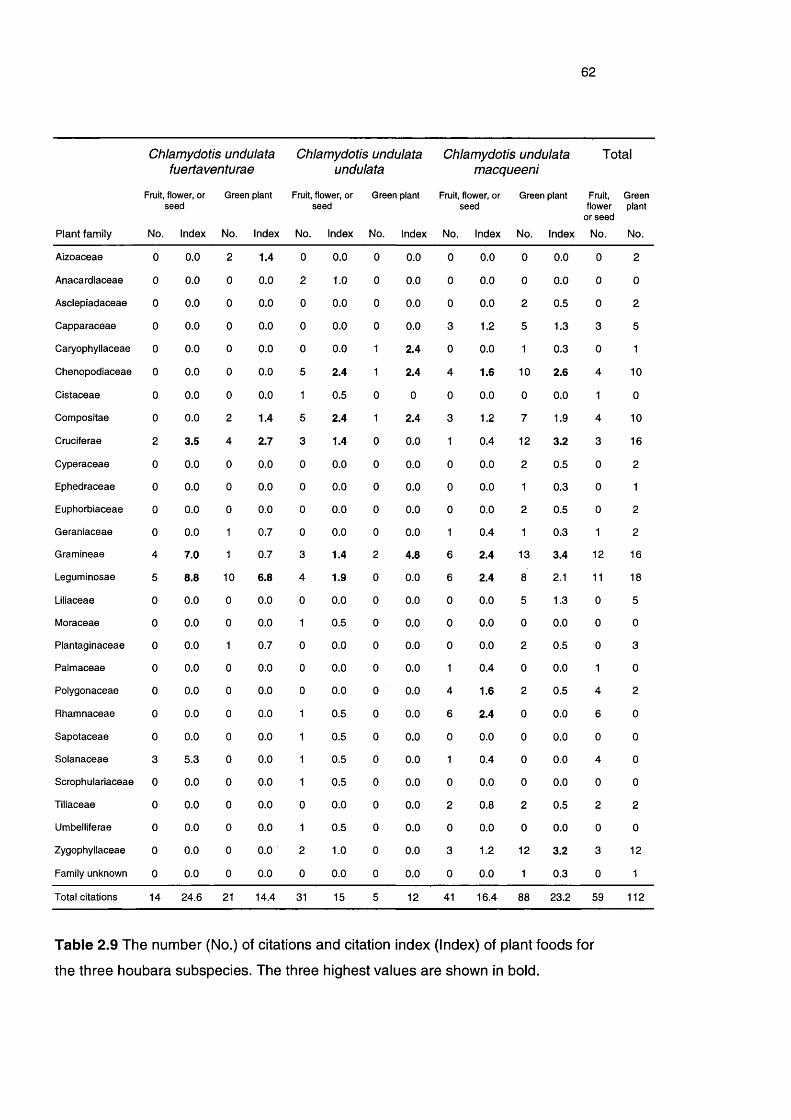
62
Chlamydotis unduiata fuertaventurae
Chlamydotis unduiata unduiata
Chlamydotis unduiata macqueeni
Total
Fruit, flower, or seed
Green plant Fruit, flower, or seed
Green plant Fruit, flower, or seed
Green plant Fruit, flower
or seed
Greenplant
Plant family No. Index No. Index No. Index No. Index No. Index No. Index No. No.
Aizoaceae 0 0.0 2 1.4 0 0.0 0 0.0 0 0.0 0 0.0 0 2
Anacardiaceae 0 0.0 0 0.0 2 1.0 0 0.0 0 0.0 0 0.0 0 0
Asclepiadaceae 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 2 0.5 0 2
Capparaceae 0 0.0 0 0.0 0 0.0 0 0.0 3 1.2 5 1.3 3 5
Caryophyllaceae 0 0.0 0 0.0 0 0.0 1 2.4 0 0.0 1 0.3 0 1
Chenopodiaceae 0 0.0 0 0.0 5 2.4 1 2.4 4 1.6 10 2.6 4 10
Cistaceae 0 0.0 0 0.0 1 0.5 0 0 0 0.0 0 0.0 1 0
Compositae 0 0.0 2 1.4 5 2.4 1 2.4 3 1.2 7 1.9 4 10
Cruciferae 2 3.5 4 2.7 3 1.4 0 0.0 1 0.4 12 3.2 3 16
Cyperaceae 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 2 0.5 0 2
Ephedraceae 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 1 0.3 0 1
Euphorbiaceae 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 2 0.5 0 2
Geraniaceae 0 0.0 1 0.7 0 0.0 0 0.0 1 0.4 1 0.3 1 2
Gramineae 4 7.0 1 0.7 3 1.4 2 4.8 6 2.4 13 3.4 12 16
Leguminosae 5 8.8 10 6.8 4 1.9 0 0.0 6 2.4 8 2.1 11 18
Liliaceae 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 5 1.3 0 5
Moraceae 0 0.0 0 0.0 1 0.5 0 0.0 0 0.0 0 0.0 0 0
Plantaginaceae 0 0.0 1 0.7 0 0.0 0 0.0 0 0.0 2 0.5 0 3
Palmaceae 0 0.0 0 0.0 0 0.0 0 0.0 1 0.4 0 0.0 1 0
Polygonaceae 0 0.0 0 0.0 0 0.0 0 0.0 4 1.6 2 0.5 4 2
Rhamnaceae 0 0.0 0 0.0 1 0.5 0 0.0 6 2.4 0 0.0 6 0
Sapotaceae 0 0.0 0 0.0 1 0.5 0 0.0 0 0.0 0 0.0 0 0
Solanaceae 3 5.3 0 0.0 1 0.5 0 0.0 1 0.4 0 0.0 4 0
Scrophulariaceae 0 0.0 0 0.0 1 0.5 0 0.0 0 0.0 0 0.0 0 0
Tiliaceae 0 0.0 0 0.0 0 0.0 0 0.0 2 0.8 2 0.5 2 2
Umbelliferae 0 0.0 0 0.0 1 0.5 0 0.0 0 0.0 0 0.0 0 0
Zygophyllaceae 0 0.0 0 0.0 2 1.0 0 0.0 3 1.2 12 3.2 3 12
Family unknown 0 0.0 0 0.0 0 0.0 0 0.0 0 0.0 1 0.3 0 1
Total citations 14 24.6 21 14.4 31 15 5 12 41 16.4 88 23.2 59 112
Table 2.9 The number (No.) of citations and citation index (Index) of plant foods for
the three houbara subspecies. The three highest values are shown in bold.

63
Taxonomic Group
Chlamydotis unduiata fuertaventurae
Number of Citation Index citations
Chlamydotis unduiata unduiata
Number of Citation Index citations
Chlamydotis unduiata macqueeni
Number of Citation Index citations
Reptiles 7 4.9 11 6.5 18 2.1
Mammals 5 3.5 0 0.0 2 0.2
Total vertebrates 12 8 .3 11 6.5 20 2.4
Lepidoptera 1 0.7 0 0.0 4 0.5
Hemiptera 0 0.0 0 0.0 2 0.2
Orthoptera 3 2.0 3 1.8 18 2.1
Odonata 0 0.0 0 0.0 1 0.1
Isoptera 0 0.0 0 0.0 4 0.5
Diptera 0 0.0 0 0.0 1 0.1
Total Hymenoptera 2 1.4 6 3.6 13 1.6
Apidae 0 0.0 0 0.0 1 0.1
Vespoldea 0 0.0 0 0.0 1 0.1
Ichneumonidae 0 0.0 0 0.0 1 0.1
Formlcldae 2 1.4 6 3.6 10 1.2
Total Coleoptera 11 7.6 13 7.7 77 9.2
Carabidae 0 0.0 0 0.0 8 1.0
Scarabaeidae 5 3.5 2 1.2 12 1.4
Buprestidae 0 0.0 2 1.2 1 0.1
Chrysomelldae 0 0.0 1 0.6 1 0.1
Staphyllnldae 0 0.0 0 0.0 1 0.1
Curculionidae 2 1.4 0 0.0 9 1.1
Tenebrionidae 1 0.7 5 3.0 28 3.3
Meloidae 0 0.0 0 0.0 1 0.1
Dermestidae 0 0.0 0 0.0 1 0.1
Elateridae 0 0.0 0 0.0 2 0.2
Silphidae 0 0.0 0 0.0 1 0.1
Cantharidae 0 0.0 0 0.0 1 0.1
Total Insects 17 11.8 22 13.0 120 14.3
Araneae 0 0.0 2 1.2 2 0.2
Scorplones 0 0.0 1 0.6 1 0.1
Soilfugae 0 0.0 0 0.0 4 0.5
Total arachnids 1 0.7 3 1.8 8 1.0
Crustacea (Isopoda) 0 0.0 0 0.0 3 0.4
Myriapoda(Chllopoda)
0 0.0 0 0.0 2 0.2
Mollusca 8 5.6 2 1.2 1 0.1
Total Invertebrates 26 18.1 25 14.8 133 15.8
Total animal 38 26.4 36 21.3 153 18.2
Table 2.10 The number of citations and citation index for animal foods of the three
houbara subspecies. The highest values are shown in bold.

64
Citation indices for plant foods
The total citations for green plants (112) were higher than for fruits, flowers and
seeds (59), although this was not the case for C.u.unduiata which had 31 citations
for fruits, flowers and seeds, and only 5 for green plants. C.u.macqueeni showed
the highest diversity of plant foods with 18 families eaten as vegetative structures
and 13 families as fruit and flowers. C.u.unduiata had records for four families as
vegetative structures and 14 as fruit and flowers, while C.u.fuertaventurae had
records for seven families as vegetative structures and four families as fruit and
flowers. This probably reflects the higher number of studies carried out on
C.u.macquee/?/particularly compared with C.u.unduiata which is poorly known.
There may be a tendency for C.u.fuertaventurae to eat a narrower range of fruits
and flowers, perhaps influenced by the diversity of plants in its home range.
The families with the highest citation indices for vegetative foods for each three
subspecies were: C.u.fuertaventurae: Leguminosae (6.8), Cruciferae (2.7),
Aizoaceae (1.4) and Compositae (1.4); C.u.unduiata: Caryophyllaceae (2.4),
Chenopodiaceae (2.4), Compositae (2.4) and Gramineae (4.8); C.u.macqueeni:
Cruciferae (3.2), Gramineae (3.4), Zygophyllaceae (3.2) and Chenopodiaceae (2.6)
(see Table 2.9).
The four families with the highest citation indices as seeds, fruit or flowers eaten by
the houbara subspecies were: C.u.fuertaventurae: Gramineae (7.0), Leguminosae
(8.8) Solanaceae (5.3) and Cruciferae (3.5); C.u.unduiata: Chenopodiaceae (2.4),
Compositae (2.4), Leguminosae (1.9), Gramineae (1.4) and Cruciferae (1.4);
C.u.macqueeni: Chenopodiaceae (1.6), Polygonaceae (1.6) Gramineae (2.4),
Leguminosae (2.4) and Rhamnaceae (2.4).
All three subspecies had records for Gramineae, Leguminosae, Cruciferae,
Compositae and Solanaceae (Table 2.9). The Zygophyllaceae and Chenopodiaceae
were important for C.u.unduiata and C.u.macqueeni. Many plant families were
eaten as both vegetative and fruiting or flowering structures.

65
Citation Indices for animal foods
A wide range of animais were consumed by houbara (Table 2.10). invertebrates
were particularly important, with records from four phyla of the Arthropode, most of
which were insects, for which eight orders were recognised. There were fewer
citations for vertebrates than for invertebrates, although reptiles had citations
indices of 4.9, 6.5 and 2.1 for C.u.fuertaventurae, C.u.unduiata and C.u.macqueeni
respectively. Apart from Fox (1988) and Gubin & Mukhina (in prep.), reptile records
were based on feeding observations. However Symens (1987) noted that houbara
were not always successful when hunting lizards, and observation alone may over
estimate their importance. Mammals were only recorded as foods for
C.u.fuertaventurae and C.u.macqueeni (citation indices of 3.5 and 0.2). It may be
difficult to detect vertebrate remains, for although Gubin & Mukhina (in prep.) found
a mouse tail and reptile bones in faeces, Nazarov (1992) was unable to detect
bones in faeces of houbara fed on small birds.
The citation indices suggested that invertebrates formed a consistently high
proportion of the diet for all subspecies {C.u.fuertaventurae 18.1, C.u.unduiata 14.8
and C.u.macqueeni 15.6), and as with plants, a high diversity of animal prey were
recorded for C.u.macqueeni. Beetles were the most important insect prey in terms
of diversity and number of records and had similar citation indices for
C.u.fuertaventurae, C.u.unduiata and C.u.macqueeni {7.6, 7.7 and 9.2 respectively),
with records from 12 beetle families for C.u.macqueeni. Tenebrionids were
important for C.u.unduiata and C.u.macqueeni, but were rarely recorded for
C.u.fuertaventurae (citation indices of 3.0, 3.3 and 0.7 respectively). However, this
was reversed for Scarabaeidae which had a citation index of 3.5 for
C.u.fuertaventurae, compared with 1.2 for C.u.unduiata and 1.4 for C.u.macqueeni.
Only C.u.fuertaventurae had a high citation index for molluscs (5.6) based on snails,
although more recent work has not confirmed that houbara eat snails (Collins 1984).
There may be differences in the food preferences of the subspecies, influenced by
an impoverished fauna and flora for the island-dwelling C.u.fuertaventurae
compared with mainland dwelling houbara (MacArthur & Wilson 1967). For
example, dung-beetles are more likely to reach the Canaries than apterous
tenebrionids. Ants were also frequently cited for the three subspecies
{C.u.fuertaventurae 1.4, C.u.unduiata 3.6 and C.u.macqueeni 1.2), and the citation

66
indices for Orthoptera were fairly consistent across the range {C.u.fuertaventurae
2.1, C.u.unduiata 1.8 and C.u.macqueeni2.^).
2.3.3 Seasonal trends in the houbara’s diet
Several authors have commented on the seasonal consumption of food types but
their evidence is often based on an interpretation of what they supposed were
abundant foods at the time. VVork by Collins (1984,1993) and Gubin & Mukhina (in
prep.) is more detailed, although their results were biased towards male birds.
Seasonal consumption of plant foods
Collins (1984,1993) states that C.u.fuertaventurae tavouxed Lycium sp. berries in
April. Faeces from one bird consisted almost entirely of Lycium sp. and the bird had
apparently ignored other abundant foods, such as herbaceous plants. Gillett (1988)
suggested that fresh plant growth, seedlings, fruits and flowers were selected by
C.u.unduiata following rain in the winter and spring.
Other work has suggested that the consumption of plants by houbara is
opportunistic. Roberts (1991) mentioned that C.u.macqueeni gizzards contained
perennials in the early winter and herbs in spring. Birds in semi-natural conditions
were also thought to respond to seasonal changes in vegetation (Launay 1989c;
Anegay 1992; Symens 1987) and to consume the vegetative parts of shrubs during
the winter but new growth of annual plants in the spring.
Gubin & Mukhina (in prep.) related the remains of food in faeces to qualitative
descriptions of plant phenology in Kazakhstan. Grasses, Saisoia sp. and tulips
made up much of the spring diet, but by the end of March the amount of plant
material in faeces decreased. At the end of May and the start of summer, seeds
were eaten but the mean proportion of vegetative plant material dropped to about
10%. By June Erodium sp. seeds constituted up to 10% of the volume of some
samples, and by the end of June most ephemeral plants had dried up and there
were more seeds than vegetative plant parts in the faeces. In September, only a
small amount of seeds and vegetation was present in faeces.

67
Seasonal consumption of animal foods
Invertebrates are frequently said to be seasonally important in the diet.
C.u.fuertaventurae appeared to eat tenebrionids in preference to ants, because
while ants were available all year round they were only consumed in the summer
when tenebrionids were rare (Collins 1984,1993). Mirza’s (1971) account of
C.u.macqueenig\zzards suggested an increase in invertebrate consumption
between early and late winter, when houbara may have selected high protein foods
prior to the breeding season. Consumption of insects by houbara in semi-natural
conditions was thought to reflect seasonal abundance (although this was not
measured) with more insects eaten in the winter than in the spring (Launay 1989c,
Symens 1987). Insects were said to be very important for young chicks (Launay &
Paillat 1990) and the hatching of chicks in Kazakhstan may be related to an
increase in invertebrate abundance (F. Launay pers. comm.). Houbara gizzards
from Mahazat as-Sayd suggest a shift towards invertebrate foods in the autumn and
plant foods in the spring (Combreau & Rambaud 1995).
Gubin & Mukhina (in prep.) related a description of the seasonal abundance of
animals to prey remains in faeces. Animals were scarce in early spring but small
amounts of ants, beetles (Tenebrionidae, Curculionidae; Carabidae and
Scarabaeidae), woodlice, Myriapoda and spiders were found in faeces. By the end
of March, the volume of animal remains in faeces and gizzards (n=5) increased to
20% with Curculionidae and Tenebrionidae making-up 12% of the volume of faeces.
Termites, ants, Elateridae and Cicindelidae were also eaten. At the beginning of
May, the volume of animal matter in the faeces continued to increase and 40% with
of the volume of faeces consisted of tenebrionid and buprestid remains.
Curculionidae, Scarabaeidae, Meloidae and Solifugidae were also eaten, but
although Meloidae were very abundant they rarely occurred in faeces probably
because of their high toxicity. By the end of May and the start of June, tenebrionid
and buprestid remains formed up to 70% of the volume of faeces. From mid-June
Solifugidae occurred in 70% of samples and made up 10% of the volume of faeces,
and would have formed a high volume of gizzard contents. Orthoptera were found in
faeces from June onwards and lizards occurred in 9.2% of samples. Woodlice were
said to have been present at all times but were only found faeces in September
when other prey were scarce. Beetles, Solifugidae and a single lizard and mouse
were also eaten at this time.

68
Reports of faeces and gizzards dominated by a single invertebrate species are
common, particularly ants (Goriup & Norton 1992; Glutz von Blotzheim etal. 1973)
and locusts or grasshoppers (see Table 2.5), and Symens’ (1988a) observations in
Saudi Arabia suggest that houbara actively respond to locust swarms. In Central
Asia, Kostin (1956) mentioned gizzards that only contained tenebrionid beetles,
while Salikhbaev & Ostapenko (1967) reported gizzards containing 400-500
termites. Social insects, such as ants and termites, provide an abundant prey for
houbara especially during the dispersal phase of their reproductive castes, and also
if their foraging trails or nests are exploited.
2.3.4 Implications for the management of habitat and captive birds
Evidence to date suggests that the houbara is an omnivore, consuming a wide
variety of prey. The most frequently cited foods were invertebrates, especially
tenebrionid beetles, Orthoptera and ants, and green plants including native species
and cultivated crops. There were few records of vertebrate prey. Differences
between the subspecies probably reflect the local availability of food in their ranges.
Some data suggested preferences for particular food types and seasonal changes
in food consumption with animal foods becoming more important prior to the
breeding season. However, apart from Collins (1984, 1993), no attempts were made
to measure food availability.
There remains much basic information to be learnt about the houbara. However,
current attempts to release captive-bred birds into the wild (Bailey & Hornby 1994;
Biquand etal. 1992) cannot succeed unless suitable habitats are available. It may
be important to try to manage reserves so that sufficient food is available to cover
seasonal trends and stressful times of year, such as extreme dry, hot or cold
periods, which occur in many parts of the houbara's range. Crops such as chick
peas, beans, alfalfa, sorghum and brassicas provide an easily grown source of
cover in open areas with little natural vegetation, and are a source of plant and
animal food. In the Canary Islands, a reward scheme operates for farmers which
ensures that alfalfa is cultivated for the benefit of wild houbara (F. Dominguez
Casanova pers. comm.). Alfalfa is also planted in aviaries and at NARC houbara
often hunt grasshoppers in alfalfa plots (A. Owen pers. comm.). Naturally occurring
foods such as favoured plants should be encouraged, since good quality vegetation

69
will also aid invertebrate and small vertebrate populations. The control of grazing by
domestic stock may be an important factor (Oatham 1996). At Harrat Al-Harrah, the
native Capparis spinosa is thought to be very attractive to houbara (P. Seddon pers.
comm.) and attempts to grow this plant, such as seeding suitable habitat, could
improve the quality of over-wintering areas for wild houbara. It could also be grown
in aviaries and on protected release sites for captive bred birds. It does not occur in
UAE, and an analysis of wild houbara diet in UAE will help to identify other locally
occurring food plants that could be used in similar manner.
Ants are a frequent component of the diet and scavenging species are attracted into
aviaries. Rather than being seen as a threat to captive houbara, they are either
innocuous or a possible alternative food for the birds, and chemical control of ants is
not normally necessary.
2.4 Conclusion
Much of the information on the diet of the houbara is observational and there is a
need for a standardised technique based on faecal analysis to study the diet across
the range (see Chapter 5). The relative importance of foods must be assessed in a
non-subjective method and information is needed on the differential digestion of
prey and passage time in order to interpret samples. In addition, studies of the diet
should incorporate a measure of food availability (see Cooper & Whitmore 1990).

70
C h a p t e r 3
P il o t s t u d ie s f o r m o n it o r in g d e s e r t in v e r t e b r a t e s
Synopsis of Chapter 3
This chapter assesses various sampling methods for examining the distribution,
abundance and diversity of desert invertebrates (to be used in Chapter 4). Both
aerial insects and ground-dwelling arthropods were sampled, although emphasis
was placed on the taxa most likely to occur in Abu Dhabi (see Chapter 1 ; Tigar
1996a; Appendix 1.1) and to be potential food for houbara. Therefore the
Tenebrionidae, Formicidae and Orthoptera were of particular interest as suggested
in Chapter 2. The aim was not to test the absolute sampling powers of the
techniques, but to identify methods that were reliable and practical within a desert
terrain.

71
3.1 General introduction
Invertebrates can be sampled by a variety of methods from casual observation to
more accurate quantified approaches using complex trapping apparatus
(Southwood 1978; Upton 1991). Composition and size of trap catches are
determined by the biology and behaviour of the taxa being sampled, trap design and
local environmental conditions (see Dent & Walton 1997; Muirhead-Thompson
1991). The best techniques achieve a reliable and representative sample which can
be used as a relative measure for comparison between samples obtained in a
similar fashion. Catches are not usually considered absolute measures of
abundance but rather indices of abundance and activity of the population. For
vertebrate predators, like the houbara, such methods can give an idea of prey
availability (Cooper & Whitmore 1990; Wolda 1990). Studies of avian food supply
must be accurate, as well as appropriate to behaviour of the bird (Hutto 1990).
The severe climate and difficult terrain pose a challenge to biological surveys in
UAE (see Chapter 1). In addition, the areas frequented by houbara are used by
herdsmen, livestock, military personnel and for general recreation. Therefore
equipment should be unobtrusive, robust and only left unattended for a short time.
3.2 Pitfall trapping
3.2.1 Introduction to pitfall trials
Knowledge of Abu Dhabi's invertebrates is biased towards relatively mesic
environments (see Tigar 1996a & b; Appendices 1.1 & 1.2) and surface-active,
desert arthropods have largely been ignored. However, in many arid zones they
form a major proportion of the fauna and the bulk of animal biomass (Crawford
1991; Ghabbour & Shakir 1980; Thomas 1979). Pitfall traps catch ground-dwelling
invertebrates and have been used extensively to compare faunal assemblages,
including those of deserts (e.g. Ahearn 1971; Aldryhim etal. 1992; Ayal & MerkI
1994; Faragalla & Adam 1985; Wharton & Seely 1981).
In November 1992, two separate lines of 30 pitfall traps (one pint plastic tumblers)
with ten paces (about 10m) between each trap were set up and left overnight. Upon
collection 75% (± 0.07 S.D.) of traps contained invertebrates, especially ants.

72
silverfish, tenebrionids and spiders (in 70% ± 0.35, 7% ± <0.01, 15% ±0.12 and 5%
± 0.02 of traps respectively). This indicated that pitfall traps were a useful technique
for sampling invertebrates in UAE.
In order to standardise the trapping procedure, several trial transects were set-up.
These examined the effect of the length of the transect and the time of trapping on
the number and diversity of arthropods captured. To reduce the chance of escapees
and to improve pitfall trap efficiency a dry film of Polytetraethylene-fluoride (“fluon”
manufactured by ICI PLC) was painted around the rim of pitfall cups to create a
non-stick barrier.
3.2.2 Methods
The trapping-time trial
Three transects were sited about 1km from each other across uniform sandy desert
in the Public Hunting Triangle to examine how the number of species (a measure of
diversity) and total number of arthropods captured by pitfall traps varied with time.
Each transect consisted of 30 pitfall traps, spaced ten paces apart and left in place
for 14, 24 or 36 hours from the start time (16.30 hrs). Catches were recorded to
taxonomic order.
The transect length trial
To estimate the optimum number of traps to use in a transect, three separate lines
of 60 pitfall traps were set up across uniform sandy desert and left overnight (16 hr
± 0.5 S.D.). Sixty traps represented the maximum number that could practically be
used in field studies. A Monte Carlo method was used to simulate the effect of using
fewer than 60 traps; n traps (where 0<n<60) were selected at random with
replacement on 50 occasions and the average and standard deviation of the catch
were calculated. Catches were examined in terms of the abundance and species
diversity of total arthropods, and of two groups thought to be important foods for the
houbara: Coleoptera and Hymenoptera.

73
3.2.3 Results
The trapping-time trial
The total trap catches at 14 and 24 hours were similar (71 and 69 arthropods
respectively) (see Table 3.1). However, there was a higher catch at 36 hours (216)
because of the increased number of Hymenoptera, especially ants, in the traps.
Taxa Line 1 (14 hrs) Line 2 (24 hrs) Line 3 (32 hrs)Total caught
Coleoptera 2 2 3Hymenoptera 62 60 193Hemiptera 2 0 0Araneae 2 0 1Thysanura 2 3 18Orthoptera 1 4 0Scorplones 0 0 1Total invertebrates 71 69 216
Total caught per hourColeoptera 0.005 0.003 0.003Hymenoptera 0.148 0.083 0.201Hemiptera 0.005 0 0Araneae 0.005 0 0.001Thysanura 0.005 0.004 0.019Orthoptera 0.002 0.006 0Scorplones 0 0 0.001Mean invertebrate capture 5.07 2.88 6
Number of species caughtColeoptera 2 2 3Hymenoptera 23 15 27Hemiptera 2 0 0Araneae 2 0 1Thysanura 2 2 2Orthoptera 1 4 0Scorplones 0 0 1Total invertebrates 32 23 32
Number of species caught per hourColeoptera 0.143 0.083 0.094Hymenoptera 1.643 0.625 0.844Hemiptera 0.143 0.000 0.000Araneae 0.143 0.000 0.031Thysanura 0.143 0.083 0.056Orthoptera 0.071 0.167 0.000Scorplones 0.000 0.000 0.031Total invertebrates 2.286 0.958 0.889
Table 3.1 Numbers of invertebrates and species caught by three pitfall transects in
place for 14, 24 and 32 hours.

74
The mean catch of total arthropods per hour decreased from 5.07 to 2.88 per hour
between 14 and 24 hours, with a slight increase to 6.00 arthropods per hour at 36
hours. Again, the increase in the total invertebrates caught per hour could be
attributed to higher catches of Hymenoptera, although Thysanura also increased.
However, mean catches of Coleoptera, Hemiptera and Orthoptera decreased
between 14 and 32 hours trapping, while scorpions were only caught in traps left in
place for 36 hours.
Results for the number of species caught per hour were mixed. A total of 32 species
were captured following both 14 and 32 hours of trapping, while only 23 species
were caught for the pitfall transect in place for 24 hours (Table 3.1). The mean
number of species caught per hour was higher following 14 hours of trapping (2.3)
than 36 hours (0.9).
The transect length trial
The random simulations of the mean number of total invertebrates, of Coleoptera
and of Hymenoptera caught with increasing number of traps varied between the
three lines and by taxonomic group, although line 1 had a consistently higher catch
(see Figure 3.1). Note that in all cases, the mean catches of fewer than ten traps
were extremely unstable. However, the overall means showed improved stability
between ten and 20 traps, although the estimate for Hymenoptera in line 3
continued to show some instability, even at 60 traps.
The standard deviations calculated during the simulations were not plotted in Figure
3.1 to avoid confusion caused by the considerable overlap between the three lines.
However, a typical example is given in Figure 3.2 for Coleoptera in line 1, showing
the decline in variance with increasing number of traps. Therefore both the means
and standard deviations of total arthropods, Coleoptera and Hymenoptera had a
tendency towards increased stability with at least ten traps.
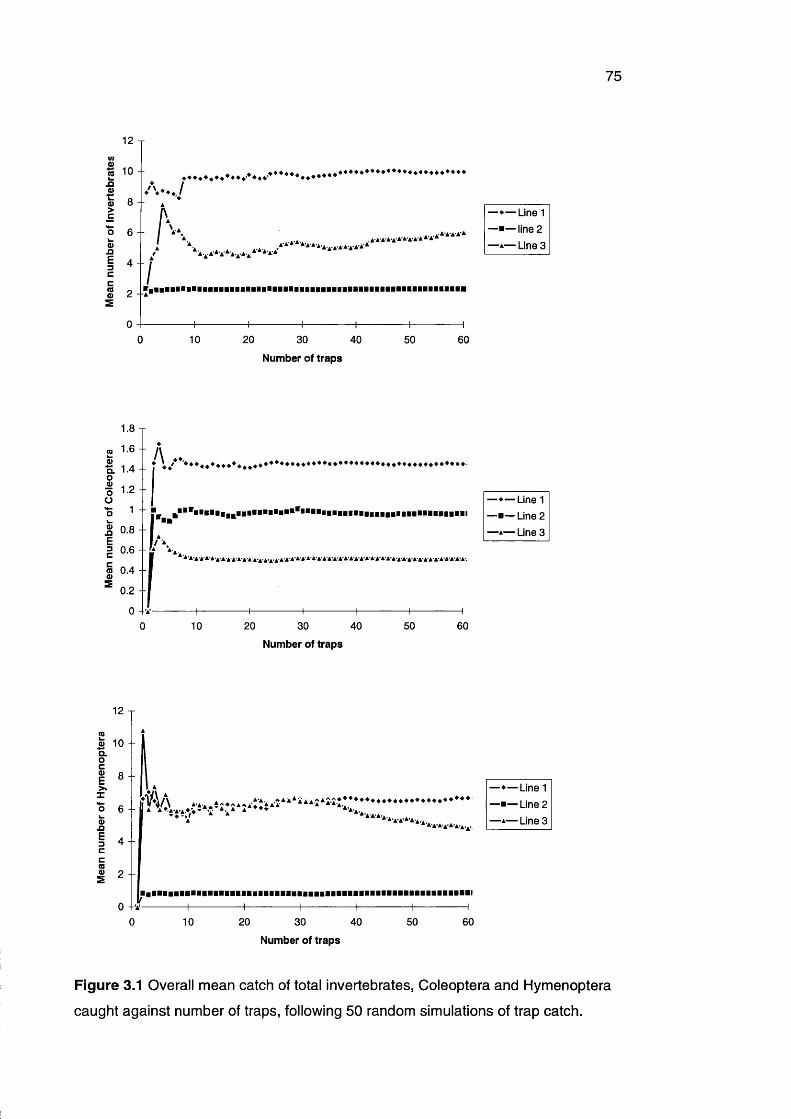
75
12 T
10 -Sn0)CI 8 - -
oa>nE3CC
ss
A-A'A'A 'A.A'A’A .A ..,A *A i
4 -
2 - -■
200 10 30 40 50 60
— ♦— Line 1
— ■— line 2— A— Line 3
Number of traps
10 20 30 40
Number of traps
— I—
50
— ♦— Line 1
— ■— Line 2
— A— Line 3
60
12 T
« 10o
| sX
0 61I 4 +cc(00) pS
0
■A-A.^-A-A-a .^
10 20 30 40
Number of traps
50
— ♦— Line 1
— ■— Line 2— A— Line 3
■■IH60
Figure 3.1 Overall mean catch of total invertebrates, Coleoptera and Hymenoptera
caught against number of traps, following 50 random simulations of trap catch.

76
3.5
I 2.5
0 ^1 1.5
I '5 0.50)S n
-0.50 10 20 30 40 50 60
Number of traps
Figure 3.2 The overall mean catch of beetles for line 1 against number of traps.
Error bars are standard deviation about the mean.
Species-effort curve
The cumulative number of species caught for each of the three lines was plotted
and fitted with a logarithmic trend-line to examine the effect of increasing effort (i.e.
number of pitfall traps) on the number of species captured (Figure 3.3). Line 1
captured the highest number of species (24), while lines 2 and 3 both contained 16
species. However, all three lines showed a similar trend, with a decreasing rate of
return for new species typical of species-effort curves.
To investigate the effect further, the angle of inflection was drawn onto each of the
fitted curves by eye (Figure 3.3). This suggested that the rate of return, or the
chance of capturing new species, declined rapidly with more than nine or 12 pitfall
traps. Although new species were still encountered, the proportion of effort required
to encounter new species was much greater.

77
Line 1
« 20 - -
0 10 20 30 40 50 60
Number of traps
Line 2
« 18 T
w 14 --
° 12 - -
■■■■
0 10 20 30 40 50 60
Number of traps
Line 3
« 16 T
% 14 --
12 - -
20 50 600 10 30 40
Number of traps
Figure 3.3 The cumulative number of species caught along three lines of 60 pitfall
traps. The thin solid lines are fitted logarithmic trends. The dashed lines indicate
the point of inflection of the curve and its intersection with the x axis.

78
3.2.4 Discussion
The total number of arthropods caught in the pitfall transects increased between 14
and 36 hours. However, the taxa of most relevance to the houbara, particularly the
large Coleoptera, did not increase during this time, nor did they increase between
14 and 24 hours. The increased catch was the result of an increased abundance of
ants, which are social insects and communicate via a complex system of
pheromones (Holldobler & Wilson 1990), making it is difficult to interpret their trap
catches. These insects generally find food along communal foraging trails, so if a
foraging trail was intersected by a pitfall transect there would be a high probability of
catching high numbers of ants. In addition, some ants are predators and general
scavengers on other arthropods, and would be attracted to contents of the traps,
particularly dead or dying insects. If a trap is left in place for a long time, especially
during the heat of the day, it would contain a higher proportion of dead to live
insects than a trap in place for a shorter time, and would therefore attract more
scavengers.
In general, an increase in sampling time (effort) would be expected to capture more
individuals and more species, but results suggested that this did not occur in the
short-term (i.e. within a few days) (Table 3.1). Rarer species are less likely to be
captured simply because they are less abundant. However, they are also less likely
to be encountered and eaten by a bird, such as the houbara. Therefore for the
current study, an overnight “snap-shot”, obtained using dusk till dawn trapping
would provide a standardised and practical sampling method, and give a relative
measure of arthropod abundance and diversity for comparison across sites. In
addition, it will sample the time periods when houbara are most likely to be foraging
(Anegay 1994; Combreau & Launay 1996).
Sixty traps were used as an experimental maximum. However, the simulation
suggested that while the use of fewer than ten traps gave an unstable impression of
the catch, the total trap catch became stabilised around 20 traps (Figures 3.1 &
3.2). There was still some instability in the number of Hymenoptera (mainly ants)
caught in line three reflecting the colonial biology and foraging behaviour of ants.
Other factors, such as differences in micro-climate between the sites, as well as ant
colony size and species identity may also have affected the number of ants caught
(see MacKay 1991).

79
The species-effort curve (Figure 3.3) suggested that the rate of return of finding
new species declined markedly after 12 pitfall traps. If 20 traps were used, the most
abundant ground-dwelling arthropods would be sampled, although rarer species
might occasionally be missed. However, the use of temporal and spatial replication
would reduce biases and inconsistencies.
Few scorpions were caught in pitfall traps probably because and some species
detect and avoid traps (Polis 1990b). The scorpions caught in pitfall traps are more
likely to be juveniles or smaller species, which have a higher probability of being
captured and a lower probability of escaping than larger individuals. A short-
wavelength UV light could also be used at night to give an estimate of scorpion
population density (Polis 1990a).
Climate and micro-climate are also likely to influence the size and composition of
pitfall trap catches (see Ahearn 1971). These factors cannot be controlled during
the trapping, but can be monitored using a datalogger.
Other factors to be considered when using live pitfall trapping are losses due to
escapees and predation. However, these would occur with equal probability at all
trap sites. One solution is to use liquid preservatives or killing agents in pitfall traps,
although these have attractant or repellent effects upon certain taxa (see Luff 1968,
1975; Wagge 1985). Preserving fluids are usually used if traps are in situ ior a long
time, while initial tests in UAE suggested that overnight, unbaited traps captured
many ground-dwelling invertebrates. Difficulties in transporting liquids over rough
terrain, combined with evaporation and wind-blown sand filling the traps made the
use of a preservative a less viable option. In addition, it might be difficult to relocate
traps on sandy substrates after a long period of time judging from the poor recovery
of the much larger boards (see Section 3.3). In Saudi Arabia, Combreau &
Rambaud (1995) used dilute picric acid in monthly pitfall traps and their catches
included gerbils and muscarid fly larvae. The latter were presumably breeding on
the decaying trap contents, and are not relevant to the present study.
Another advantage of live trapping is that common and easily identified species can
be released, thereby reducing the effect of removing them from their environment.
Furthermore, the pilot trials revealed the most westerly record for the gecko

80
Teratoscincus scincus (Osborne 1993). Reptiles are also recorded as being preyed
upon by the houbara, and it is useful to have an idea of their distribution.
3.3 The use of refuge boards
3.3.1 Introduction
Some ground-dwelling arthropods, including beetles, scorpions, solifugids and
spiders, seek refuge under pieces of debris and wood in the desert (Muma 1980).
The use of standard-sized, artificial refuges might provide a way of sampling these
invertebrates.
3.3.2 Methods
To test the viability of refuges and the effect of their size on capture, 40 rectangular
wooden boards were made with the following dimensions: 30 x 30 cm (n=20), 30 x
60 cm (n=10) and 60 x 60 cm (n=10). Boards were distributed at random and at
least 250m apart across a sandy habitat, dominated by two grasses: Pennisetum
divisum and Panicum turgidum. The boards were located on 16* May 1993 and
their position was recorded using a Magellan GPS (Global Positioning System) unit
(accurate to 100m) which was used to relocate them 28 days later.
3.3.3 Results
Only 57% of the boards were recovered and 35% of those found harboured a small
number of invertebrates, including termites, spiders and thysanurans (Table 3.2). A
few tracks of larger tenebrionids and reptiles were seen under the boards, but the
individuals responsible for the tracks were not present. The largest boards were
easier to find than the smaller boards, and also acted as better refuges or were
colonised more quickly. However, the effect of size could not be tested statistically
because of the low recovery rate.
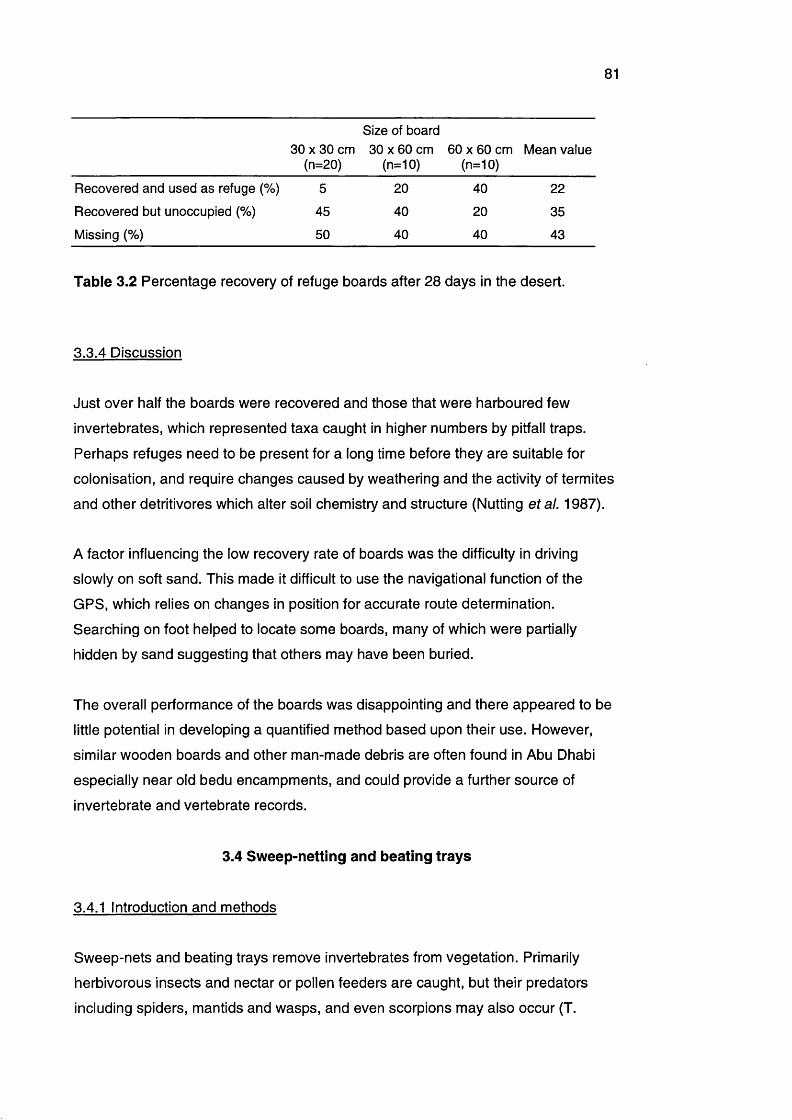
81
30 X 30 cm (n=20)
Size of board30 X 60 cm
(n=10)60 X 60 cm
(n=10)Mean value
Recovered and used as refuge (%) 5 20 40 22
Recovered but unoccupied (%) 45 40 20 35
Missing (%) 50 40 40 43
Table 3.2 Percentage recovery of refuge boards after 28 days in the desert.
3.3.4 Discussion
Just over half the boards were recovered and those that were harboured few
invertebrates, which represented taxa caught in higher numbers by pitfall traps.
Perhaps refuges need to be present for a long time before they are suitable for
colonisation, and require changes caused by weathering and the activity of termites
and other detritivores which alter soil chemistry and structure (Nutting etal. 1987).
A factor influencing the low recovery rate of boards was the difficulty in driving
slowly on soft sand. This made it difficult to use the navigational function of the
GPS, which relies on changes in position for accurate route determination.
Searching on foot helped to locate some boards, many of which were partially
hidden by sand suggesting that others may have been buried.
The overall performance of the boards was disappointing and there appeared to be
little potential in developing a quantified method based upon their use. However,
similar wooden boards and other man-made debris are often found in Abu Dhabi
especially near old bedu encampments, and could provide a further source of
invertebrate and vertebrate records.
3.4 Sweep-netting and beating trays
3.4.1 Introduction and methods
Sweep-nets and beating trays remove invertebrates from vegetation. Primarily
herbivorous insects and nectar or pollen feeders are caught, but their predators
including spiders, mantids and wasps, and even scorpions may also occur (T.

82
Benton, pars. comm.). Sweep-netting is probably the most widely-used method of
measuring invertebrate availability for avian predators, mainly because it is easy
and cheap, although it is biased towards sedentary invertebrates and is sensitive to
changes in vegetation structure, and climatic conditions when sampling (Cooper &
Whitmore 1990). It can be used semi-quantitatively by sweeping or beating for a
specified distance, number of passes of the net, distance or time (e.g. Savory 1974:
Green 1984; Rebel etal. 1995).
In September 1993 sweep-netting was attempted at two locations which had
Zygophyiium sp. - Haloxylon salicornicum plant associations (see definition in
Roshier etal. 1996). The net was passed ten times across the vegetation wherever
it occurred, once every ten paces (approximately 10m), along a 200m line. A total of
12 lines were sampled. All insects captured were removed from the net using a
pooter and recorded. The beating tray was used along 12 transects, which were
parallel to and 20m away from the sweep-net transects. Ten sharp taps to the
vegetation were executed with a thick bamboo cane to loosen any insects.
3.4.2 Results
Very few invertebrates of limited taxa were caught in the sweep-nets (Table 3.3).
Numbers caught were variable (note that standard deviation was twice the mean
value per transect). No insects were captured using the beating trays.
Site Replicate Taxa Number caughtPHT 1 0PHT 2 Hemiptera 3PHT 3 0PHT 4 0PHT 5 0PHT 6 0
B 1 0B 2 0B 3 0B 4 Tentyrina palmeri (Tenebrionidae) 1B 5 0B 6 Crematogaster antaris (Formicidae) 2
Total insects caughtMean insects per transect (±SD)
60.5 (±1.0)
Table 3.3 Number of invertebrates and taxa caught along twelve 200 m Sweep-net
transects. (PHT = Public Hunting Triangle, B = Baynunah).

83
3.4.3 Discussion
Very few insects were caught by sweep-netting and none was caught by the beating
tray. Both methods were difficult to carry out because the vegetation was sparse
(typically <5% cover; Roshier et al. 1996) and consisted of characteristic desert
plants with spiny or twiggy growth. The net frequently became entangled in the
vegetation, and gusty winds made it difficult to control the net and to remove the
catch. Most shrubs were too low and spiny to accommodate the beating tray.
Automated sampling methods, such as D-vacs or suction traps, might be useful.
However, these methods, as well as sweep-netting and beating, are sensitive to the
daily temperature variations that typically occur in Abu Dhabi (see Chapter 1), which
affect insect activity and hence capture rate. In addition, heavy equipment would be
difficult to handle on the sandy, uneven terrain, and suction or vacuum apparatus
would probably fill up and be damaged by dust and sand.
Beating trays and sweep-nets could be used to collect specimens after the rainy
season, but they cannot be recommended as methods for regular sampling.
However, herbivorous insects would be better sampled as winged adults using a
light trap (see Heath trap below), although some apterous insects, like certain
weevil species, may remain difficult to sample.
3.5 Walking transects for Orthoptera
3.5.1 Introduction and methods
In September 1993 Line and band transects are used to assess the abundance of
locusts and other Orthoptera (M. Richie pers. comm.). Twelve transects were
attempted by walking in a straight line for approximately 200m, and recording any
Orthoptera along a 5m wide band.
3.5.2 Results
No Orthoptera were encountered along any of the transects.

84
3.5.3 Discussion
Again, the low vegetation cover meant that this technique is not normally applicable
in Abu Dhabi. However, such transects may be of value when Orthoptera are
seasonally abundant, e.g. following the rains, although identification to species level
may be difficult in the field. Some Orthoptera could be captured using a light trap
(see Section 3.7).
3.6 Flight Interception traps
3.6.1 Introduction and methods
Flight interception traps were considered as a sampling method for flying insects
(see for example Owen 1993). A Malaise trap was positioned on a sandy site at
Sweihan Research Station to look at the types and numbers of flying insects
caught. The reservoir of the trap was filled with 70% ethanol and left in place for 28
days.
3.6.2 Results
The Malaise trap caught a total of 288 insects. However, most were houseflies
(>51%) or small Hymenoptera (38%) with fewer Coleoptera (5%), Lepidoptera
(4.9%) and Neuroptera (<0.2%) (Table 3.4). The average daily catch was 10.3
insects. Some taxa were difficult to identify owing to the lack of taxonomic keys for
the region.
3.6.3 Discussion
The Malaise trap captured flying insects which were not captured in the pitfall traps.
However, daily trap catches were low, about ten to 100 times less than a single
night's catch from a Heath light trap in the same area (see Section 3.7 below).
Houseflies and small Hymenoptera dominated the Malaise trap catch, although
these traps typically capture large Hymenoptera, adult Lepidoptera and only some
Diptera (Cooper & Whitmore 1990). The Abu Dhabi catch contained few likely food
sources for houbara (see Chapter 2).
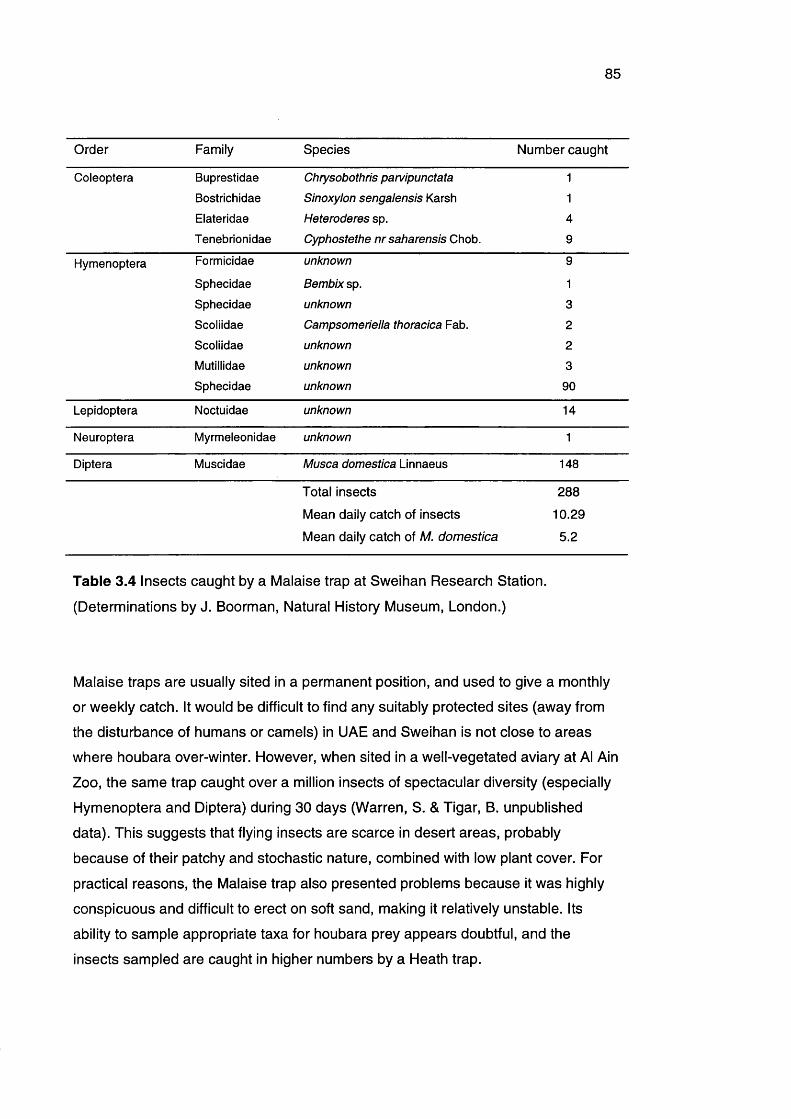
85
Order Family Species Number caught
Coleoptera Buprestldae Chrysobothhs parvipunctata 1
Bostrlchldae Sinoxylon sengalensis Karsh 1
Elaterldae Heteroderes sp. 4
Tenebrionidae Cyphostethe nr saharensis Chob. 9
Hymenoptera Formicidae unknown 9
Sphecldae Bembix sp. 1
Sphecldae unknown 3
Scollldae Campsomeriella thoracica Fab. 2
Scollldae unknown 2
Mutlllldae unknown 3
Sphecldae unknown 90
Lepidoptera Noctuldae unknown 14
Neuroptera Myrmeleonldae unknown 1
Diptera Muscldae Musca domestica Linnaeus 148
Total insects 288
Mean daily catch of insects 10.29
Mean daily catch of M. domestica 5.2
Table 3.4 Insects caught by a Malaise trap at Sweihan Research Station.
(Determinations by J. Boorman, Natural History Museum, London.)
Malaise traps are usually sited in a permanent position, and used to give a monthly
or weekly catch. It would be difficult to find any suitably protected sites (away from
the disturbance of humans or camels) in UAE and Sweihan is not close to areas
where houbara over-winter. However, when sited in a well-vegetated aviary at AI Ain
Zoo, the same trap caught over a million insects of spectacular diversity (especially
Hymenoptera and Diptera) during 30 days (Warren, S. & Tigar, B. unpublished
data). This suggests that flying insects are scarce in desert areas, probably
because of their patchy and stochastic nature, combined with low plant cover. For
practical reasons, the Malaise trap also presented problems because it was highly
conspicuous and difficult to erect on soft sand, making it relatively unstable. Its
ability to sample appropriate taxa for houbara prey appears doubtful, and the
insects sampled are caught in higher numbers by a Heath trap.

86
3.7 The use of light traps for the capture of nocturnal insects.
A Heath trap (supplied by Watkins and Doncaster) was tested during September
1993 at Sweihan. Initially it was filled with egg boxes to act as landing/roosting
places for flying insects attracted into the trap at the night. However, the next
morning the trap quickly heated up and most of the catch, consisting of over several
hundred insects, escaped. Therefore, a jar of ethanol was placed under the funnel,
to kill any insects entering the trap, but it also removed lepidopteran scales making
these insects hard to identify. Instead, a large jar lined with plaster of Paris was
placed under the funnel, and 15 ml of ethylene dichloride was dispensed into the
plaster when setting up the trap. Ethylene dichloride is a killing agent with a low
vapour pressure, and therefore a relatively slow evaporation rate. Upon collection
the next day about 5 ml of ethyl acetate was added to the jar to ensure that all the
material died rapidly. A final refinement was a photoelectric sensor which
automatically switched the trap on or off according to the light level, helping to
conserve battery power.
The light trap caught many flying insects not sampled by pitfall traps, including
Lepidoptera, Coleoptera (especially Scarabaeoidea), Orthoptera, Diptera,
Myrmeleonidae and Hemiptera. Only a single Heath trap was available. Ideally traps
should be replicated to give a measure of variance, but a single trap could be used
to give an indication of the relative abundance of flying insects at each site each
month.
3.8 General discussion and recommendations for trapping methods
No single method can be expected to sample the entire arthropod fauna, since each
trap is designed to exploit particular traits of its intended catch. The pitfall traps
proved an efficient method for sampling ground-dwelling invertebrates in UAE,
particularly the Tenebrionidae which are frequently cited in as a food item for
houbara (see Chapter 2). Herbivorous insects, such as grasshoppers and locusts
(Orthoptera), weevils (ColeopteraiCurculionidae) and lepidopteran larvae are
unlikely to be captured in pitfalls. However, the Heath light trap captured some of
these taxa, including Orthoptera, apterous weevils and adult Lepidoptera.
Therefore, the combination of pitfall traps and a Heath trap can be used to sample
the available ground-dwelling and flying invertebrates (Chapter 4).

87
The other methods tested (refuge boards, sweep-nets, beating trays, grasshopper
transects, and a Malaise trap) either yielded little information or were impracticable.
However, along with hand-searching on vegetation and under rocks, they could be
used to provide additional information on invertebrate diversity and abundance
following seasonal rains.

88
C h a p t e r 4
T h e d iv e r s it y a n d a b u n d a n c e o f in v e r t e b r a t e s in A b u D h a b i's d e s e r t s
Synopsis of Chapter 4
There is scant information on invertebrates in Abu Dhabi (see Tigar 1996a & b;
Collingwood etal. 1997; Chapter 1 & Chapter 3). Therefore base-line data on desert
invertebrates were gathered and examined for general patterns of abundance and
diversity. These aims were achieved through a two-year monitoring programme
using methods evaluated through pilot studies (Chapter 3). Ground-dwelling
invertebrates were sampled with overnight pitfall traps and flying insects with a
Heath light trap. The effect of climate, micro-climate, substrate and the phase of the
moon on trap catch were investigated.
The study established a database and a reference collection of invertebrates for the
UAE, in collaboration with a network of international taxonomists. These resources
facilitated the identification of houbara prey in Chapter 5 and will aid further
entomological and biogeographical research. A list of animals encountered during
the study is given in Appendix 4.1, including 196 new invertebrate records for Abu
Dhabi and ten new species (Collingwood & Agosti 1996).

89
4.1 Patterns of abundance and diversity of ground dwelling arthropods
(Section 4.1 has been published as: Tigar, B. J. & Osborne, P. E. (1997). Patterns
of arthropod abundance and diversity in an Arabian desert. Ecography 20\ 550-
558.)
4.1.1 Introduction
Biodiversity is probably spatially heterogeneous in most ecosystems (Gaston &
Williams 1996). Although desert landscapes appear monotonous and uniform, they
can exhibit patchiness at scales from a few centimetres through to whole continents
due to differences in microclimate, habitat and geography (e.g., Crawford 1988).
Temporal variability may also be marked, occurring on a daily, seasonal, yearly or
historical basis (Crawford 1991) and environmental factors affecting deserts are
also conspicuously stochastic. For example, rainfall is sporadic and variable, and
the total annual precipitation can fall in a single event (e.g. Bottomley 1996 for Abu
Dhabi). Understanding the abiotic and biotic processes that affect species
composition in space and time is an important aspect of community ecology of
deserts (Polis 1991a).
Large-scale studies of biodiversity are scarce, especially outside North America,
Australia and Africa (Gaston & Williams 1996). For deserts, they are even scarcer
and, in Arabia, ecological studies are relatively recent (Osborne 1996c) and basic
knowledge of species' occurrences is poor. Satchell (1978) was among the first to
describe the ecology of the United Arab Emirates and to list some of its
invertebrates (since updated by Tigar 1996a). Faragalla & Adam (1985) and
Aldryhim etal. (1992) have studied the Coleoptera in neighbouring Saudi Arabia.
Ayal & MerkI (1994) studied tenebrionid beetles in the Negev Desert of Israel but
this has a Mediterranean rather than an Arabian influence.
This chapter explores the spatial and temporal patterns in arthropod taxonomic
diversity resulting from a two year monitoring programme. The results show
seasonal (cyclical) changes in numbers and differences between sites related to
climatic factors. In particular, arthropods at near-coastal sites appeared to benefit
from high humidity, allowing a larger number of taxa to occur and to remain active
through the oppressive heat of summer.

90
4.1.2 Methods
Trapping sites
Five sites were established for monitoring invertebrates in separate geographic
areas where houbara or their tracks had been recently recorded (Osborne 1992):
Baynunah, Medinet Zayed, Public Hunting Triangle, Um Az Zimul and Khatam (see
Plates 4.1-4.5, Figure 1.1 & Table 4.1). These areas consisted of mobile sand
dunes intersected by compacted gravel plains with low vegetation cover (<5%)
dominated by low-growing, xerophytic shrubs (Osborne 1992; Oatham 1996;
Roshier etal. 1996). Two sites were coastal, two inland and a fifth intermediate from
the coast. The closest locations were about 100 km apart and the furthest 350 km.
All sites were accessible by 4WD vehicle but were remote from major centres of
human population. A GPS was used to map and locate each sampling point.
Pitfall Trapping
Invertebrates were sampled at the five trapping locations in Abu Dhabi (see Figure
1.1). Table 4.1 shows the site characteristics and summary statistics for the
arthropod communities, with total plant species counts from Osborne (1992) and
cover values from quadrat data in Roshier etal. (1996). Pitfall trapping was used to
assess ground-dwelling invertebrate occurrence and abundance. For many desert
invertebrates, closely spaced pitfall traps yield pseudo-replicates of numbers that
are not spatially independent with respect to the movements of individual animals.
Therefore, to obtain a replicated design for statistical analysis, a line of traps was
used as the unit of replication, pooling the data from individual pitfalls along the line.
At each of the five study sites, three lines of 20 traps were placed on mobile sand
and a further three on compacted inter-dune plains (termed gravel here for brevity),
spacing lines at least 500m apart. Individual pitfalls were 750ml polypropylene
beakers (pint "Rollers"), 83mm diameter and 138mm deep, placed in the ground
with their rims flush with the soil surface.

91
Plate 4.2 Baynunah Plate 4.3 Medinet Zayed
Plate 4.4 Public Hunting Triangle
ui >■ -
Plate 4.5 Khatam Plate 4.6 Um Az Zimul
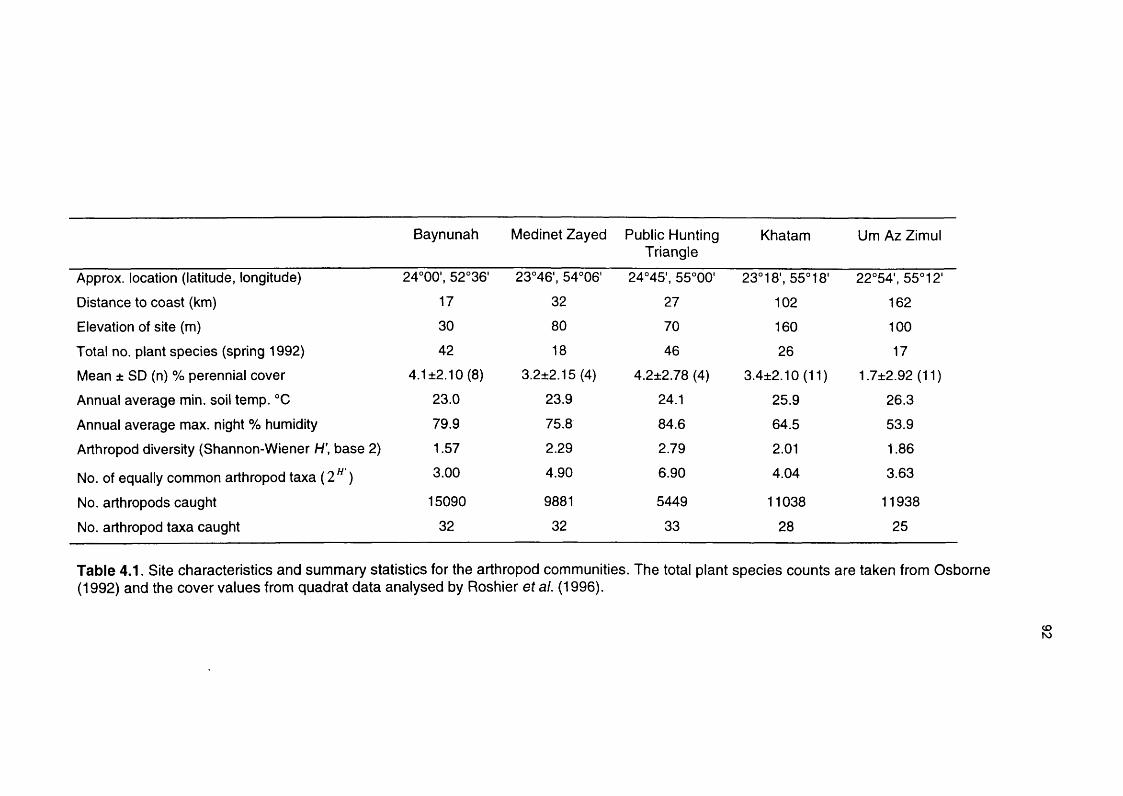
Baynunah Medinet Zayed Public Hunting Triangle
Khatam Um Az Zimul
Approx. location (latitude, longitude) 24°00‘, 52°36' 23°46', 54°06' 24°45', 55°00* 23°18‘, 55°18' 22°54‘, 55°12'Distance to coast (km) 17 32 27 102 162
Elevation of site (m) 30 80 70 160 100
Total no. plant species (spring 1992) 42 18 46 26 17
Mean ± SD (n) % perennial cover 4.1±2.10(8) 3.2±2.15(4) 4.2±2.78 (4) 3.4±2.10(11) 1.7±2.92(11)
Annual average min. soil temp. °C 23.0 23.9 24.1 25.9 26.3
Annual average max. night % humidity 79.9 75.8 84.6 64.5 53.9Arthropod diversity (Shannon-Wiener H\ base 2) 1.57 2.29 2.79 2.01 1.86
No. of equally common arthropod taxa ( 2 " ) 3.00 4.90 6.90 4.04 3.63
No. arthropods caught 15090 9881 5449 11038 11938
No. arthropod taxa caught 32 32 33 28 25
Table 4.1. Site characteristics and summary statistics for the arthropod communities. The total plant species counts are taken from Osborne (1992) and the cover values from quadrat data analysed by Roshier et al. (1996).
CDro

93
Traps were set for one night (approximately two hours before dusk to two hours
after dawn) within a period three days either side of the new moon once every 28
days. Few arthropods are active during daylight hours and the trapping regime
sampled the main periods of arthropod activity.
Trapping was carried out 24 times from 12 October 1993 to 29 August 1995 (i.e. for
about two years) using the same study sites but not the same positions for lines
which were randomly chosen within a block of about lOkm^. The data therefore
consist of three replicated random samples per substrate per site, repeated on 24
occasions, that are independent in both space and time (based on a total of 14,400
individual pitfall traps).
Climatic data
Soil temperature, air temperature and relative humidity were recorded at one
randomly chosen sand pitfall line at each site each month. The severe operating
conditions sometimes caused the data-logger to malfunction and the data set is
unfortunately incomplete. Where data were available, two robust measures from the
logged data (minimum soil temperature and maximum humidity) were used for the
analysis. The mean monthly minimum air temperatures and total rainfall from a
reputable climatic station were also incorporated into the analyses. These data were
averages from the years 1982 to 1992 for Abu Dhabi International Airport
(Bottomley 1996).
Data analvsis
All captured animals were identified to the finest taxonomic level possible, but it was
not always possible to determine to species level. Table 4.2 lists the taxa caught
during the study, excluding those where fewer than 20 individuals were caught over
two years. For reliability and ease of analysis, only the more robust taxonomic
determinations are explored. Differences between sites, substrates and trapping
periods were assessed using ANOVA on log transformed data (to improve
normality). The percentage similarity of the sand and gravel communities at the five
sites was calculated, based on 35 taxa and clustered using an Unweighted Pair-
Group Method using simple Averages (UPGMA) without standardising or
transforming the data (Krebs 1989).

94
Class Order Superfamily/Family
Genus/Species
Arachnida 2.63%
Insecta 97.34%
Crustacea 0.02% Scolopendrida 0.01%
Araneae 1.51% Scorpiones 1.06%
Solifugae 0.06%
Coleoptera 8.35%
Hymenoptera 76.37%
Isoptera 0.07% Diptera 0.16% Orthoptera 0.16% Mantodea 0.06%
Neuroptera 0.01% Hemiptera 0.12% Lepidoptera 0.10% Thysanura 11.95% Isopoda 0.02% Scoiopendromorpha 0.01%
Buthidae 1.06%
Carabidae 0.21%
Curcuiionidae 0.06% Histeridae 0.14% Scarabaeoidea 0.34%
Tenebrionidae 7.54%
Others 0.06% Formicidae 75.36%
Mutiiiidae/Bradybaenidae 0.98% Others 0.03%
Sphaeroceridae 0.16% Tettigoniidae 0.16% Eremiaphiiidae 0.05% Others 0.01% Myrmeieontidae 0.01 %
Lepidoptera 0.10% Lepismatidae 11.95% Porceiiionidae 0.02%
Buthacus yotvatensis nigroaculeatus 0.60% Vachoniolusminipectenibus/globimanus0.26%Compsobuthus arabicus 0.10%Apistobuthus pterygocercus 0.04%nr Galeodes arabs 0.06%
Scarites guineensis 0.08% Anthia duodecimguttata 0.12%
nr Saprinus chalcites 0.14% Scarabaeus cristatus 0.11% Aphodius spp. 0.20% Apentanodes/Arthrodibius 0.70%Akis elevata 0.08%Blaps kollari 0.52%Erodius spp. 0.30%Mesostena puncticollis 3.50% Tentyrina palmeri 0.38% Ocnera/Trachyderma philistlna 0.08%Prionotheca coronata 1.22% Pimelia arabica 0.18% Phaeotribon/Pachycera sp. 0.38%Prochoma sp. 0.05% Sceiosodis besnardi 0.08%
Catagiyphis spp. 0.50% Cardiocondyla gallagheri 0.13%Camponotus spp. 0.36% Crematogaster spp. 8.81% Messor spp. 2.66% Monomorium spp. 62.66% Lepisiota nigrescens 0.17%
Psammotermes spp. 0.07%
Piatycleis sp. 0.16% Eremiaphiia gene 0.05%
Scoiopendra mirabilis 0.01%
Table 4.2. Composition of the 53,396 arthropods caught by pitfall-trapping over two years in Abu Dhabi, nr = near. Genera and species are only listed where more than 20 individuals were captured.

95
Species diversity was calculated using the Shannon-Wiener index, /-/'and its
transposition 2"' (Peet 1974), which is the number of equally common taxa that
would produce the same diversity as /-/’ (Krebs 1989).
4.1.3 Results
The arthropod communitv
Around 53,400 invertebrates were captured over 24 months, of which 97.3% were
insects and 2.6% arachnids (Table 4.2). The dominant insect orders were
Hymenoptera (76.4% of which 75.4% were ants), Thysanura (12%) and Coleoptera
(8.4%). The Araneae comprised 57.4% of the arachnids, the remainder being
scorpions (40.3%) and solifugids (2.3%).
Temporal variation in total catch
Arthropod numbers generally differed significantly over time, among the five sites
and between the two substrates, often with significant interaction (Table 4.3). The
catch of total arthropods closely matched the annual temperature cycle measured
both by the soil probe and the annual average (Figure 4.1). The average catch
doubled with every 6-7°C rise in minimum soil temperature. The total catch also
reached a peak after the average timing of the annual winter rains (Figure 4.1).
The interaction between site and time is well-illustrated by the differences between
the abundance patterns in relation to temperature (Figure 4.1). Simplifying these to
the correlation between log catch and minimum soil temperature showed a
difference in the strength of the relationship between Baynunah, Medinet Zayed and
the Public Hunting Triangle as against Um Az Zimul and Khatam (Table 4.4). Taking
these groups of three and two sites respectively, there was no curvilinear
component in a regression of log catch on temperature for the first group
(temperature: t=2.62, p=0.009; temperature^: t=-0.56, p=0.58; n=396, adj.
r^=35.5%), but a significant quadratic component for the second (temperature:
t=3.23, p=0.001; temperature^: t=-2.57, p=0.011; n=246, adj. r^=12.4%). Plots of the
data (Figure 4.1) showed that arthropod numbers in the second group (inland sites)
rose with increasing temperature until about 28°C but then reached a plateau. For
the other sites, however, numbers rose with temperature to the maximum 37°C
recorded.
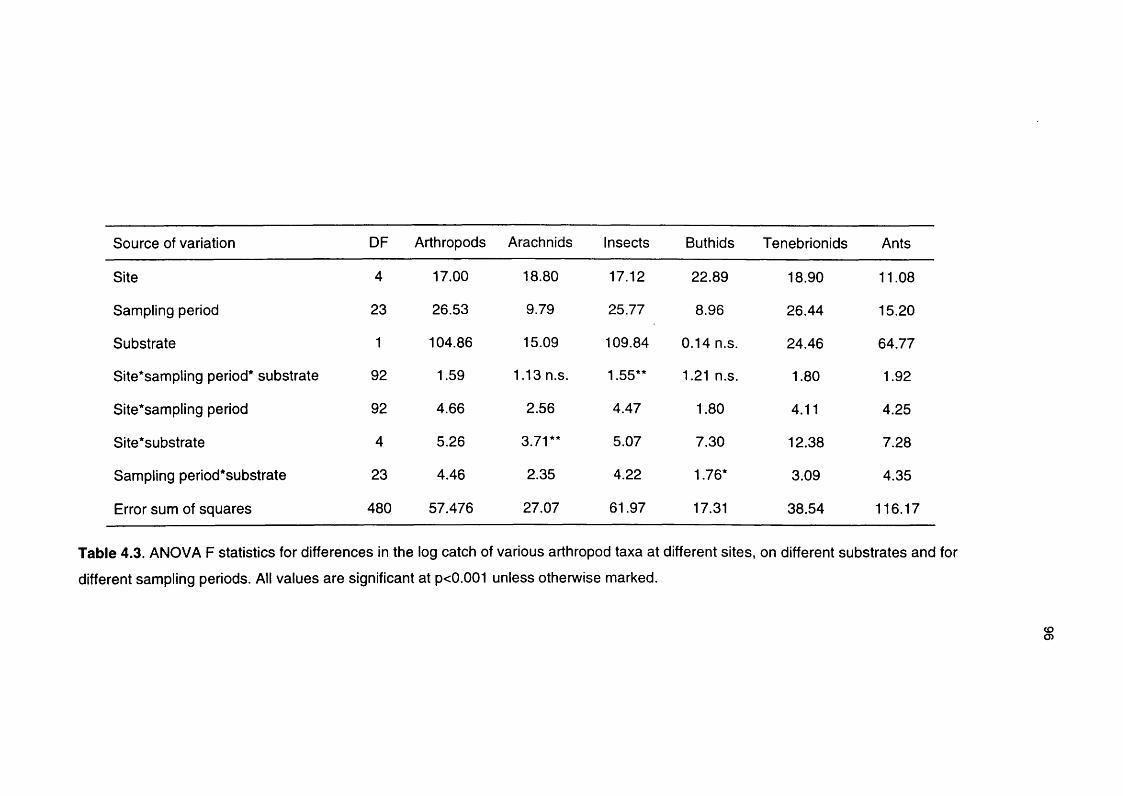
Source of variation DF Arthropods Arachnids Insects Buthids Tenebrionids Ants
Site 4 17.00 18.80 17.12 22.89 18.90 11.08
Sampling period 23 26.53 9.79 25.77 8.96 26.44 15.20
Substrate 1 104.86 15.09 109.84 0.14 n.s. 24.46 64.77
Site*sampling period* substrate 92 1.59 1.13 n.s. 1.55** 1.21 n.s. 1.80 1.92
Site*sampling period 92 4.66 2.56 4.47 1.80 4.11 4.25
Site*substrate 4 5.26 3.71** 5.07 7.30 12.38 7.28
Sampling period*substrate 23 4.46 2.35 4.22 1.76* 3.09 4.35
Error sum of squares 480 57.476 27.07 61.97 17.31 38.54 116.17
Table 4.3. ANOVA F statistics for differences in the log catch of various arthropod taxa at different sites, on different substrates and for
different sampling periods. All values are significant at p<0.001 unless otherwise marked.
(OOÎ
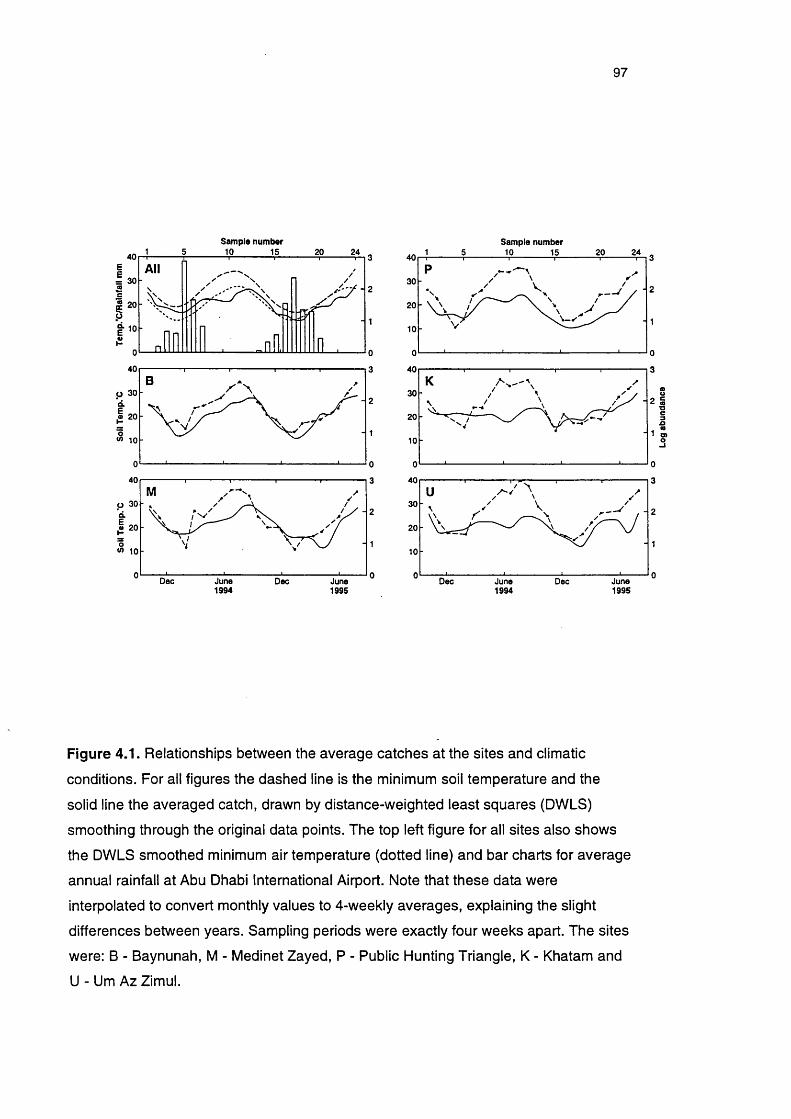
97
Sample number Sample number
40
j 3 0
I 20 P
10
All
I0 0
40 3
p 30
« 20
0
40 3
p 30I
« 20
« 10
Dec June1994
Dec June1995
2040
---/
40 3
20
40 3
30
Dec Dec June1995
June1994
Figure 4.1. Relationships between the average catches at the sites and climatic
conditions. For all figures the dashed line is the minimum soil temperature and the
solid line the averaged catch, drawn by distance-weighted least squares (DWLS)
smoothing through the original data points. The top left figure for all sites also shows
the DWLS smoothed minimum air temperature (dotted line) and bar charts for average
annual rainfall at Abu Dhabi International Airport. Note that these data were
interpolated to convert monthly values to 4-weekly averages, explaining the slight
differences between years. Sampling periods were exactly four weeks apart. The sites
were: B - Baynunah, M - Medinet Zayed, P - Public Hunting Triangle, K - Khatam and
U - Um Az Zimul.
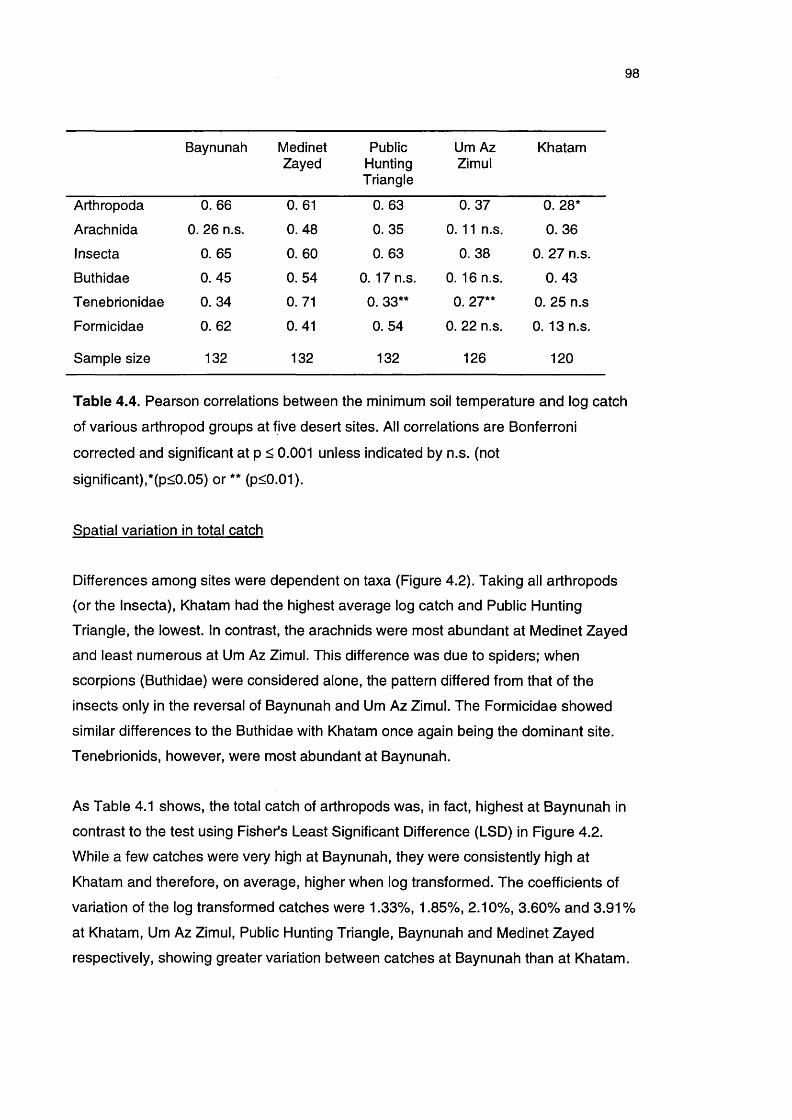
98
Baynunah MedinetZayed
PublicHuntingTriangle
Um Az Zimul
Khatam
Arthropoda 0. 66 0. 61 0. 63 0. 37 0. 28*
Arachnida 0. 26 n.s. 0. 48 0. 35 0. 11 n.s. 0. 36Insecta 0. 65 0. 60 0. 63 0. 38 0. 27 n.s.Buthidae 0. 45 0. 54 0. 17 n.s. 0. 16 n.s. 0. 43Tenebrionidae 0. 34 0. 71 0. 33** 0. 27** 0. 25 n.sFormicidae 0. 62 0. 41 0. 54 0. 22 n.s. 0. 13 n.s.
Sample size 132 132 132 126 120
Table 4.4. Pearson correlations between the minimum soil temperature and log catch
of various arthropod groups at five desert sites. All correlations are Bonferroni
corrected and significant at p < 0.001 unless indicated by n.s. (not
significant),*(p<0.05) or ** (p<0.01).
Spatial variation in total catch
Differences among sites were dependent on taxa (Figure 4.2). Taking all arthropods
(or the Insecta), Khatam had the highest average log catch and Public Hunting
Triangle, the lowest. In contrast, the arachnids were most abundant at Medinet Zayed
and least numerous at Um Az Zimul. This difference was due to spiders; when
scorpions (Buthidae) were considered alone, the pattern differed from that of the
insects only in the reversal of Baynunah and Um Az Zimul. The Formicidae showed
similar differences to the Buthidae with Khatam once again being the dominant site.
Tenebrionids, however, were most abundant at Baynunah.
As Table 4.1 shows, the total catch of arthropods was, in fact, highest at Baynunah in
contrast to the test using Fisher's Least Significant Difference (LSD) in Figure 4.2.
While a few catches were very high at Baynunah, they were consistently high at
Khatam and therefore, on average, higher when log transformed. The coefficients of
variation of the log transformed catches were 1.33%, 1.85%, 2.10%, 3.60% and 3.91%
at Khatam, Um Az Zimul, Public Hunting Triangle, Baynunah and Medinet Zayed
respectively, showing greater variation between catches at Baynunah than at Khatam.

Taxon Sites
Arthropods or Insecta K M U B PI------------- 1h
I--------- 1
Arachnida M B K P U
Formicidae K M B U
Buthidae K M B U P
I 1
Tenebrionidae B M K U PI----------------------------- 1
H
99
Figure 4.2 Differences in log catch at the five sites (Fisher's LSD test). Lines link sites
that were significantly different (p<0.05). Sites are ordered by abundance from left
(highest) to right (lowest). The sites were: B - Baynunah, M - Medinet Zayed, P - Public
Hunting Triangle, K - Khatam and U - Um Az Zimul.

100
Spatial variation in taxonomie diversity
Biodiversity statistics and environmental characteristics for the five sites are
summarised in Table 4.1. Of the possible 35 taxa in the analysis, the best site (Public
Hunting Triangle) had 33 whereas Um Az Zimul had only 25 taxa. Although Baynunah
and Medinet Zayed both had 32 taxa, the diversity as measured by the Shannon-
Wiener index was lower at Baynunah than even at Um Az Zimul. Baynunah showed a
highly skewed distribution of abundance and hence the evenness component of
diversity down-weighted the overall index value.
The number of arthropod taxa at a site was linearly related to relative humidity (r=0.99,
p=0.002) and distance to coast (r=-0.99, p=0.002), a surrogate for humidity amongst
other factors (Table 4.1). The non-significant relationship with number of plant species
appeared good except at Medinet Zayed where the number of arthropod taxa was
higher than expected for the number of plant species.
While all the taxa in Table 4.3 (except Buthidae) differed significantly in abundance
between substrates with higher catches on sand, the gravel communities were more
diverse at four out of five sites (Table 4.5). The exception was Khatam where the
gravel community was particularly poor with a diversity equivalent to only 3.04 equally
common taxa. The richest habitat was the gravel at the Public Hunting Triangle with
9.24 equally common taxa (30 taxa overall). The Public Hunting Triangle also had the
richest sand community with 5.45 equally common taxa.
Using cluster analysis, the least similar communities were separated by site rather than
substrate, suggesting that the greatest differences occurred at the geographic rather
than the local scale (Figure 4.3). Note the separate cluster for the rather impoverished
communities at the Public Hunting Triangle. The strong similarity between the sand
and gravel communities at Baynunah (89.8%) is apparent in Table 4.6, simplified to 14
taxa for ease of interpretation. However, at Medinet Zayed, Um Az Zimul and Khatam,
clustering of substrates and sites was mixed. In particular, the communities at Khatam
resulted in mixed clusters. Khatam's sand community was more similar to that at Um
Az Zimul than the sand and gravel communities were at Um Az Zimul itself. Similarly,
the Khatam gravel community split the adjacent clustering of Medinet Zayed's
communities. Percentage similarities ranged from 43.9% to 89.8%.
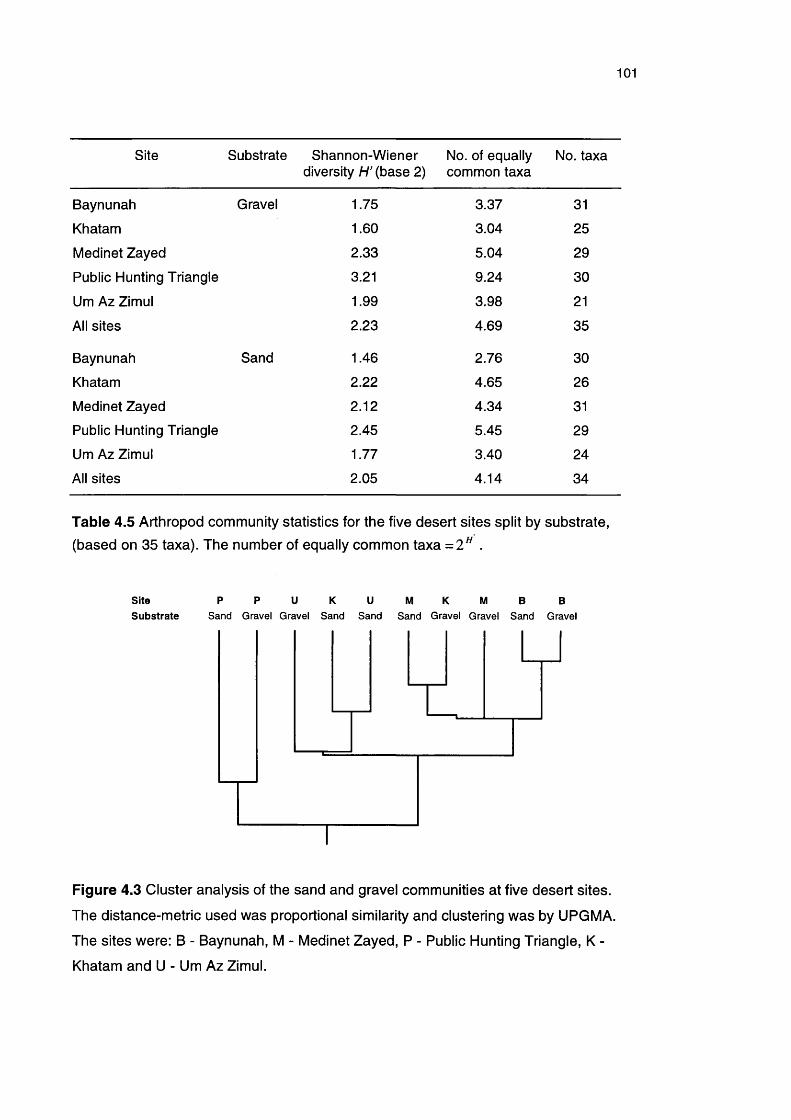
101
Site Substrate Shannon-Wiener diversity H’ (base 2)
No. of equally common taxa
No. taxa
Baynunah Gravel 1.75 3.37 31Khatam 1.60 3.04 25Medinet Zayed 2.33 5.04 29Public Hunting Triangle 3.21 9.24 30
Um Az Zimul 1.99 3.98 21
All sites 2.23 4.69 35
Baynunah Sand 1.46 2.76 30Khatam 2.22 4.65 26Medinet Zayed 2.12 4.34 31Public Hunting Triangle 2.45 5.45 29Um Az Zimul 1.77 3.40 24All sites 2.05 4.14 34
Table 4.5 Arthropod community statistics for the five desert sites split by substrate, (based on 35 taxa). The number of equally common taxa = 2 " .
Site P P U K U M K M B BSubstrate Sand Gravel Gravel Sand Sand Sand Gravel Gravel Sand Gravel
Figure 4.3 Cluster analysis of the sand and gravel communities at five desert sites.
The distance-metric used was proportional similarity and clustering was by UPGMA.
The sites were: B - Baynunah, M - Medinet Zayed, P - Public Hunting Triangle, K -
Khatam and U - Um Az Zimul.

102
Percentage composition of the sand and gravel communities is illustrated in Table 4.6.
Both Baynunah and the Public Hunting Triangle had 13 taxa on sand and 14 on gravel,
and Medinet Zayed had 13 on both sand and gravel. The two inland sites, however,
had fewer taxa, Um Az Zimul having only ten taxa on sand and eight on gravel. The
missing taxa were the Carabidae, Tettigoniidae and Isoptera; the latter two were also
absent from Khatam. In contrast, the Thysanura were both proportionally and
numerically more abundant at these inland sites than elsewhere. Termites and ground
mantids, Eremiaphila gene Febv., were always more common on gravel than sand.
4.1.4 Discussion
Communitv composition
The total catch of around 53,400 arthropods was dominated by ants (Formicidae),
thysanurans, tenebrionids and arachnids. Very few studies have examined entire
desert arthropod assemblages, making comparisons among deserts difficult. Pitfall
traps in Saudi Arabia (Combreau & Rambaud 1995) caught slightly more insects
(98.10%) and fewer arachnids (1.76%) than in this study, and 0.15% crustaceans
compared with only 0.02% from Abu Dhabi. In both areas, by far the most numerous
animals captured were Hymenoptera, particularly ants, with one genus {Monomorium)
making up 62.7% of all invertebrates in UAE. Social insects such as ants and termites
are abundant in all terrestrial ecosystems but, in deserts, may contribute up to an order
of magnitude greater biomass than herbivorous mammals (MacKay 1991). The
diversity of tenebrionids, scorpions and solifugids is greater in deserts than most other
terrestrial ecosystems (Polis 1991b; Polis & Yamashita 1991) and tenebrionids were
well-represented in Abu Dhabi, although solifugids comprised only 0.06% of the catch,
possibly because large adults could escape from the dry pitfall traps.
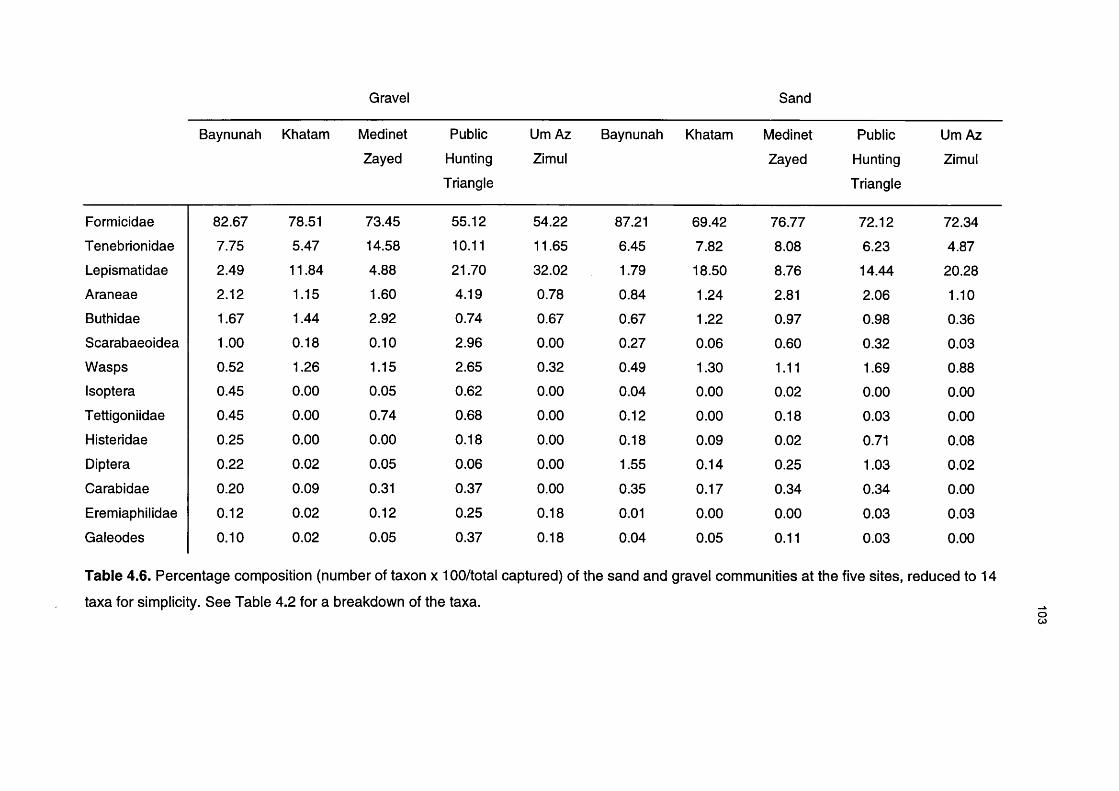
Gravel Sand
Baynunah Khatam MedinetZayed
PublicHunting
Triangle
Um Az
Zimul
Baynunah Khatam Medinet
Zayed
Public
HuntingTriangle
Um Az
Zimul
Formicidae 82.67 78.51 73.45 55.12 54.22 87.21 69.42 76.77 72.12 72.34Tenebrionidae 7.75 5.47 14.58 10.11 11.65 6.45 7.82 8.08 6.23 4.87Lepismatidae 2.49 11.84 4.88 21.70 32.02 1.79 18.50 8.76 14.44 20.28Araneae 2.12 1.15 1.60 4.19 0.78 0.84 1.24 2.81 2.06 1.10Buthidae 1.67 1.44 2.92 0.74 0.67 0.67 1.22 0.97 0.98 0.36Scarabaeoidea 1.00 0.18 0.10 2.96 0.00 0.27 0.06 0.60 0.32 0.03Wasps 0.52 1.26 1.15 2.65 0.32 0.49 1.30 1.11 1.69 0.88Isoptera 0.45 0.00 0.05 0.62 0.00 0.04 0.00 0.02 0.00 0.00Tettigoniidae 0.45 0.00 0.74 0.68 0.00 0.12 0.00 0.18 0.03 0.00Histeridae 0.25 0.00 0.00 0.18 0.00 0.18 0.09 0.02 0.71 0.08Diptera 0.22 0.02 0.05 0.06 0.00 1.55 0.14 0.25 1.03 0.02Carabidae 0.20 0.09 0.31 0.37 0.00 0.35 0.17 0.34 0.34 0.00Eremiaphilidae 0.12 0.02 0.12 0.25 0.18 0.01 0.00 0.00 0.03 0.03Galeodes 0.10 0.02 0.05 0.37 0.18 0.04 0.05 0.11 0.03 0.00
Table 4.6. Percentage composition (number of taxon x 100/total captured) of the sand and gravel communities at the five sites, reduced to 14
taxa for simplicity. See Table 4.2 for a breakdown of the taxa.ow

104
Temporal variation
Arthropod catches varied temporally and trends closely matched the yearly cycle in
minimum air or soil temperature. (The slight decrease in catches around peak
temperatures (Figure 4.1) resulted from combining abundance curves from different
taxa.) As the various measures of temperature (minimum, maximum, mean etc.) show
the same seasonal pattern in Abu Dhabi (Bottomley 1996) it is not wise to be more
specific than to regard catches as correlated with temperature. Ahearn (1971),
however, suggested that minimum temperature was one of two factors (with rainfall)
determining the tenebrionid beetle catch in an Arizona desert. Rainfall in Abu Dhabi
coincides with low winter temperatures and it is not possible in this study to separate
their effects.
Several abiotic and biotic factors could account for this overall pattern, always recalling
that pitfall traps measure abundance and activity rather than population size (e.g. Crist
& Wiens 1995). The simplest explanation is that increasing temperature causes a rise
in arthropod abundance through breeding, increased activity or both (Ahearn 1971). An
explanation involving both abiotic and behavioural factors is that some arthropods alter
their daily cycle of activity depending on temperature (Holm & Edney 1973; Ayal &
Merk11994). In the summer, these arthropods become nocturnal as opposed to diurnal
and so might be caught at night only in this season. In the sandy desert of Abu Dhabi,
there are few day-active arthropods at any time of year, in contrast to the rocky
mountain areas of the country (personal observation). However, the predominantly
diurnal genus Erodius (Tenebrionidae) contributed 0.3% to the overall catch (and was
only caught in the months from December to May) confirming that the sampling regime
could detect diurnal species. A purely biotic explanation is that predation is higher
during the winter when non-resident migrants such the houbara are present. For
example, Ayal & MerkI (1994) believe that white storks {Ciconia ciconia) may
significantly affect tenebrionid abundance in Israel.
Spatial-temporal interaction
The differences in temporal changes among sites may be explained by climate.
Although good climatic data are few, inland sites in Abu Dhabi tend to have higher
summer and lower winter temperatures than coastal sites (Bottomley 1996).
Furthermore, while the maximum relative humidity near the coast rarely drops below

105
85% over the year, inland sites drop to 70% and below from April to September.
Minimum humidity inland is extremely low (10%). Radiation fog is also common,
especially just inland from the coast, although it is less frequent in summer than winter
(Bottomley 1996). At dawn, condensation forms on desert plants with water literally
soaking the sand beneath large shrubs such as Haloxylon persicum. It appears that
desert arthropods can tolerate the high temperatures and remain active near the Abu
Dhabi coast in summer because water is available from the humid air (see also Seely
1979). Even if severely desiccated, tenebrionid beetles exhibit a remarkable ability to
recover if water becomes available (e.g. Naidu & Hattingh 1988). The inland desert
sites were both hotter and drier in summer, resulting in arthropods either avoiding
thermal stress by taking refuge in burrows (Louw & Seely 1982) or dying from
dehydration when prolonged dry periods prevent rehydration.
If this argument is correct, then a general decline in summer catches with distance into
the Empty Quarter it to be expected, along with a general decrease in arthropod
biodiversity as arid intolerant species are eliminated. Winter catches are probably also
lower in the interior as desert temperatures may reach freezing point in December and
January in the Central Region of Saudi Arabia (Faragalla & Adam 1985). The Abu
Dhabi data (Table 4.6) clearly showed fewer Isoptera and tettigoniids at the inland
sites. Overall, inland sites held fewer taxa and fewer individuals than coastal sites. The
exception was at the Public Hunting Triangle. This site held 33 taxa but only 42% of
the individuals found at other near-coast sites (true for both substrates), and did not
appear impoverished in its plant species number, composition or density (Table 4.1).
However, the level of grazing by goats and camels in the Public Hunting Triangle is
higher than at any other site and the lower arthropod numbers may be due to
excessive grazing and disturbance by livestock and animal herders. Ayal & MerkI
(1994) studied non-grazed enclosures and concluded that grazing affected the relative
abundance of tenebrionids species rather than species composition.
Spatial variation
In general, there were greater differences in the arthropod communities between
geographic locations than among habitats. Several studies of community differences
according to substrate show soil type to be important for habitat segregation (Thomas
1983; Sheldon & Rogers 1984; Crawford 1988; Ayal & Merk11994). In Abu Dhabi,
gravel sites always held fewer individuals (from 31% of the sand community at Um Az

106
Zimul to 74% at Medinet Zayed) but the number of taxa and diversity did not differ
consistently by substrate. Crawford (1988) suggests that sands are more suitable
habitats than compacted substrates for desert arthropods, especially when conditions
are extreme. Sand is a poor heat conductor, its porosity allows water to accumulate at
depths below the evaporation zone, and it is easily burrowed. Compacted substrates
resist burrowing activities except by specially adapted animals. However, we know little
of the dispersal abilities of desert arthropods which could be considerable for larger
animals such as the arachnids, and they may simply be caught in pitfalls on gravel
while foraging away from their burrows or retreats elsewhere.
The absence of carabids from the site furthest inland is consistent with their preference
for more mesic habitats and they are generally replaced by the physiologically adapted
nocturnal tenebrionids in extreme hot deserts (Ahearn 1971). Carabids have high
respiratory water loss which must be replaced by feeding on animal prey with a
relatively high water content, while tenebrionids have low rates of water loss and are
predominantly detritivores (Zachariassen etal. 1987). Thysanurans, which were also
more common at inland sites, have very low rates of water loss per unit area and can
survive extremely arid conditions (Edney 1971).
Conclusion
Much of the literature relating to desert invertebrates is based on studies carried out in
regions with less extreme climates and with a higher percentage cover of vegetation.
In Abu Dhabi, the climate is arid or hyper-arid (Bottomley 1996) and perennial
vegetation cover is less than 5% on average (Roshier etal. 1996), only boosted for
short periods by ephemerals following heavy rains. Productivity levels would therefore
be expected to be low in Arabia with fewer arthropods and lower diversity than deserts
with more benign climates. Within Abu Dhabi's deserts (and perhaps Arabian deserts
in general), spatial variation is probably driven by water availability and the extra
humidity near the coast benefits the vegetation and supports a higher taxonomic
diversity of arthropods than further inland.

107
4.2 Does the phase of the moon influence invertebrate trap catch?
4.2.1 Introduction
Moonlight is most apparent in open habitats such as deserts, and the phase of the
moon influences the behaviour of many vertebrates and invertebrates (Beasley &
Adams 1994). In general, predators are thought to be more active at full moon
because they take advantage of the greater light levels when seeking prey (Clarke et
al. 1996; Kirby 1997). Conversely, prey species may be less active, with reduced
foraging effort, because of an increase risk of predation especially from vertebrates
(Clarke etal. 1996; Vasquez 1994; Skutelsky 1996). The effect of moonlight on pitfall
trap catch has not previously been investigated.
The hypothesis that the abundance and diversity of ground-dwelling invertebrates
captured in pitfall traps would differ between new and full moons was tested.
Differences would result from changes in the risk of predation, the ease of
encountering prey and the visual awareness of invertebrates to pitfall traps (although
these hypotheses could not be separated in this study). These factors would tend to
increase predator and decrease prey abundance at the full moon.
4.2.2 Methods
Invertebrate trapping
Ground-dwelling invertebrates were sampled using overnight pitfall traps at the Public
Hunting Triangle, with three replicate lines of 20 trap, pitfall-transects on two
substrates (see Section 4.1.1). Six samples were obtained at the full and new moons,
once every 14 days from 1®* October 1993 until 1S‘ March 1994 (a total of 1440 pitfall
traps). Cloud cover was minimal and it was assumed that full and new moons
represented maximum and minimum moonlight levels, although light levels were not
measured. Temperature and relative humidity were recorded using a datalogger (see
Section 4.1.1).
Data Analvsis
Data were log-transformed to ensure normality and all analyses were carried out using
Systat (Wilkinson 1993). Trends in total capture and taxonomic diversity were
examined, and data were divided into four groups applicable to desert invertebrate

108
communities (Polis 1991c), which were likely to respond differently to moonlight:
predaceous arthropods, macroarthropod detritivores, herbivorous insects and social
insects. Predaceous arthropods exhibit trophic and taxonomic diversity, but are strict
predators, parasites or parasitoids (Polis & Yamashita 1991), Macroarthropod
detritivores are omnivorous, ground-dwelling invertebrates consuming mainly dead and
some living matter, but are not obligate predators (Crawford 1991). Coprophages and
suspected detritivores, such as anthicid beetles which are associated with stones and
debris in sandy areas in North America (Dillon & Dillon 1972), were also included in
this group. Herbivorous insects rely on their host plants for food, which in turn are
dependent upon abiotic factors, such as rainfall that are often limiting in deserts
(Wisdom 1991). Most desert social insects are ground-dwelling ants and termites,
which are characteristically colonial and high in number and biomass (MacKay 1991).
However, they are trophically diverse and include granivores, detritivores, herbivores,
general scavengers and predators.
Recent advances in taxonomy gave greater confidence to the determinations
especially for ants (see Collingwood & Agosti 1996), and it was possible to examine
diversity at a higher taxonomic level than in Section 4.1. However, spiders and
thysanurans remain poorly known and were only recorded by order.
The number of invertebrates captured in pitfall traps is dependent upon date and has
an annual cycle, following a declining curve with ambient temperature during the winter
(see Figure 4.1; Tigar & Osborne 1997). Clearly, invertebrates could not be sampled
from full and new moons on the same date, although the analyses needed to control
for the effect of date. Exploratory regression analysis suggested that sample period
(i.e. date) and its square could adequately describe the general decline in numbers
during the winter. Therefore GLMs (General Linear Models) were used to test whether
the total number of invertebrates or number of taxa caught in pitfall traps were affected
by phase of the moon (new or full), substrate (sand or gravel) and their interaction,
while using sample period (two week increment) and its square as covariates to control
for the effect of sampling at different times. Community composition, according to the
phase of the moon and substrate, was examined using Principal Components Analysis
(RCA) (Gauch 1982).

109
4.2.3 Results
The arthropod communitv
A total of 58 taxa were recorded, and there appeared to be differences in the species
composition between both phases of the moons and substrates, summarised by name
and number of taxa in Tables 4.7 & 4.8 respectively.
A wide variety of predaceous arthropods were caught, with 17 taxa representing nine
orders and at least 11 families, including scorpions, mantids, spiders, solifugids, ticks,
carabids, histerids, mutillids, tiphiids and scolopendrids. There were 16
macroarthropod detritivore taxa, with many thysanurans and tenebrionids, of which 12
species were recorded. Ants (Formicidae) were the only family of social insects
captured showing high diversity (21 species from 5 genera). Herbivorous insects
usually occur on their host plant and pitfall trapping caught only five examples from
three orders: Lepidoptera, Coleoptera and Hemiptera.
TotalCapture
Number of Taxa by GroupPredaceous Macro- Herbivorous Arthropods arthropod Insects
detritivores
SocialInsects
Total taxa 58 17 16 4 21
At both full and new moon 28 7 9 1 11Only at full moon 8 1 2 2 3Only at new moon 22 9 5 1 7
On both sand and gravel 26 9 7 1 9Only on gravel 18 2 6 1 9Only on sand 14 6 3 2 3
Table 4.7 Summary of the number of arthropod taxa captured in pitfall traps, grouped
by phase of the moon (full, new or both) and by substrate (sand, gravel or both).
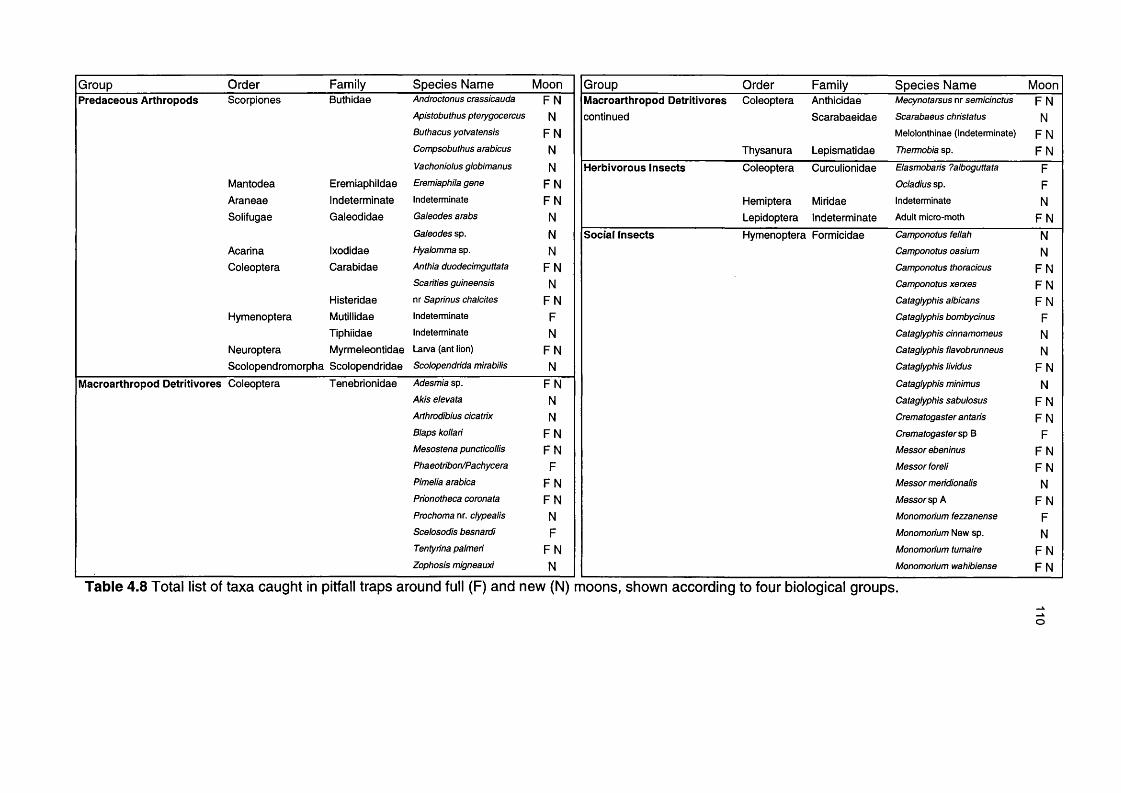
Group Order Family Species Name MoonPredaceous Arthropods Scorpiones Buthidae Androctonus crassicauda FN
Apistobuthus pterygocercus NButhacus yotvatensis FNCompsobuthus arabicus NVachoniolus globimanus N
Mantodea Eremiaphildae Eremiaphila gene FNAraneae Indeterminate indeterminate FNSoiifugae Galeodidae Gaieodes arabs N
Galeodes sp. NAcarina Ixodidae Hyaiomma sp. NColeoptera Carabidae Anthia duodecimguttata FN
Scarities guineensis NHisteridae nr Saprinus chalcites FN
Hymenoptera Mutiiiidae Indeterminate FTiphiidae indeterminate N
Neuroptera Myrmeieontidae Larva (ant lion) FNScolopendromorpha Scolopendridae Scolopendrida mirabilis N
Macroarthropod Detritivores Coleoptera Tenebrionidae Adesmia sp. FNAkis eievata NArthrodibius cicatrix NSlaps koiiari FNMesostena puncticoilis FNPhaeotribon/Pachycera FPimelia arabica FNPrionotheca coronata FNProchoma nr. ciypeaiis NSceiosodis besnardi FTentyrina paimeri FNZophosis migneauxi N
Group Order Family Species Name MoonMacroarthropod Detritivores Coleoptera Anthicidae Mecynotarsus nr semicinctus FNcontinued Scarabaeidae Scarabaeus christatus N
Melolonthinae (Indeterminate) FNThysanura Lepismatidae Thermobia sp. FN
Herbivorous Insects Coleoptera Curcuiionidae Eiasmobaris ?aiboguttata FOcladius sp. F
Hemiptera Miridae indeterminate NLepidoptera indeterminate Adult micro-moth FN
Social Insects Hymenoptera Formicidae Camponotus fellah NCamponotus oasium NCamponotus thoracicus FNCamponotus xerxes FNCataglyphis albicans FNCatagiyphis txombycinus FCataglyphis cinnamomeus NCatagiyphis fiavobrunneus NCataglyphis lividus FNCatagiyphis minimus NCatagiyphis sabuiosus FNCrematogaster antaris FNCrematogaster sp B FMessor ebeninus FNMessor foreli FNMessor meridionaiis NMessor sp A FNMonomorium fezzanense FMonomorium New sp. NMonomorium tumaire FNMonomorium wahibiense FN
Table 4.8 Total list of taxa caught in pitfall traps around full (F) and new (N) moons, shown according to four biological groups.
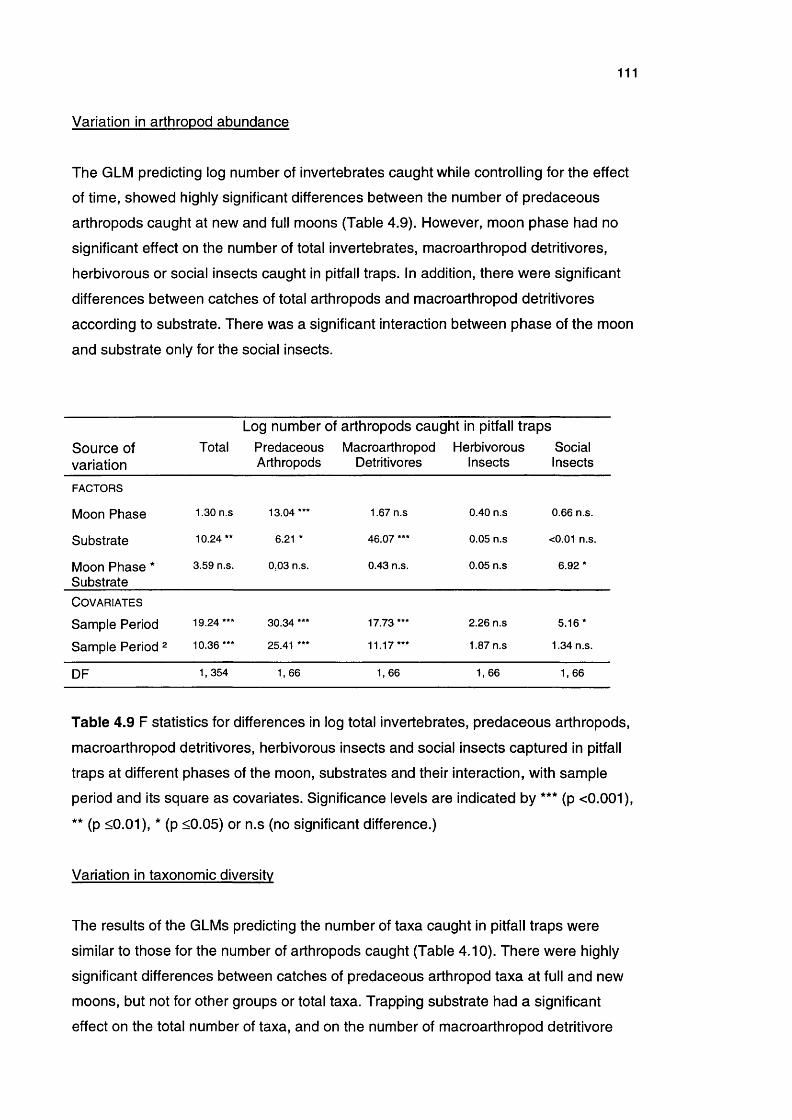
111
Variation in arthropod abundance
The GLM predicting log number of invertebrates caught while controlling for the effect
of time, showed highly significant differences between the number of predaceous
arthropods caught at new and full moons (Table 4.9). However, moon phase had no
significant effect on the number of total invertebrates, macroarthropod detritivores,
herbivorous or social insects caught in pitfall traps. In addition, there were significant
differences between catches of total arthropods and macroarthropod detritivores
according to substrate. There was a significant interaction between phase of the moon
and substrate only for the social insects.
Source of variation
TotalLog number of arthropods caught in pitfall traps
Predaceous Macroarthropod Herbivorous Social Arthropods Detritivores Insects Insects
FACTORS
Moon Phase 1.30 n.s 13.04 1.67 n.s 0.40 n.s 0.66 n.s.
Substrate 10.24 ** 6.21 * 46.07 *** 0.05 n.s <0.01 n.s.
Moon Phase * Substrate
3.59 n.s. 0,03 n.s. 0.43 n.s. 0.05 n.s 6.92 *
C o v a r ia t e s
Sample Period Sample Period ^
19.24
10.36 ***
30.34 ***
25.41 ***
17.73 ***
11.17***
2.26 n.s
1.87 n.s
5 .1 6 *
1.34 n.s.
DF 1,354 1,66 1,66 1 ,66 1 ,6 6
Table 4.9 F statistics for differences in log total invertebrates, predaceous arthropods,
macroarthropod detritivores, herbivorous insects and social insects captured in pitfall
traps at different phases of the moon, substrates and their interaction, with sample
period and its square as covariates. Significance levels are indicated by *** (p <0.001),
** (p <0.01), * (p <0.05) or n.s (no significant difference.)
Variation in taxonomic diversitv
The results of the GLMs predicting the number of taxa caught in pitfall traps were
similar to those for the number of arthropods caught (Table 4.10). There were highly
significant differences between catches of predaceous arthropod taxa at full and new
moons, but not for other groups or total taxa. Trapping substrate had a significant
effect on the total number of taxa, and on the number of macroarthropod detritivore

112
taxa caught. There were also significant differences for the interaction between phase
of the moon and substrate for social insects. There were no other significant effects.
Log number of taxa caught in pitfall trapsSource of variation
Total PredaceousArthropods
MacroarthropodDetritivores
HerbivorousInsects
SocialInsects
FACTORS
Moon Phase 3.64 n.s 12.32 *** 1.37 n.s 0.40 n.s 0.40 n.s.
Substrate 6 .45* 3.70 n.s. 11.70 *** 0.05 n.s <0.01 n.s.
Moon Phase * Substrate
3.16 n.s. <0.01 n.s. 1.66 n.s. 0.05 n.s 6.95 **
C o v a r ia t e s
Sample Period 18.05 *** 17.67 *** 13.40 *** 2.26 n.s 2.21 n.s.
Sample Period ^ 12.88 *** 13.79 *** 9.51 ** 1.87 n.s 0.99 n.s.
DF 1,354 1,66 1, 66 1, 66 1 ,66
Table 4.10 F statistics for differences in log number of taxa for total invertebrates,
predaceous arthropods, macroarthropod detritivores, herbivorous insects and social
insects captured in pitfall traps between phases of the moon, substrates and their
interaction, with sample period and its square as covariates. Significance levels are
indicated by *** (p <0.001), ** (p <0.01), * (p <0.05) or n.s (no significant difference.)
Figure 4.4 illustrates graphically the result for the number of species of predaceous
arthropods and phase of the moon. The curve shows the trend in catch with time,
described by the covariates sample and sample squared. Note that all the full moon
means of the log number of taxa caught lie below the curve, while apart from samples
4 and 8, the new moon means are on or above the curve. Minimum values for the
relationship occurred at the 8th sample (13 January 1994), corresponding with the
minimum soil temperature recorded (10.6°C). Therefore, on average, more
predaceous arthropod taxa were caught around new moons than full moons. The
number of individuals caught showed a similar trend, indicating that both high numbers
and diversity of predaceous arthropods occur around the new moon.

113
<
2ü_
SCO
2.0
1.5
1.0
0.5
0.02 3 4 5 6 7 8 9 10 11 121
Sample no. (2 week intervals)
Figure 4.4 The relationship between log number of taxa of predaceous arthropods vs
sample number (solid line), with means and standard errors for catches at full
(triangles) and new moons (diamonds). Sample number indicates the two week
sampling interval from 1®' October 1993.
Species composition according to the phase of the moon and substrate
PCA identified two factors which together explained 50.2% of variation in species
composition among catches (Table 4.11; Figure 4.5). Factor 1 represents a trend
towards greater numbers of two macroarthropod detritivores: Lepismatidae and
Sceiosodis besnardi, and two social insects: Messor sp. A and Monomorium tumaire
(Table 4.11). Factor 2 represents a trend towards greater numbers of the social insects
Crematogaster antaris and Messor ebeninus. In addition, both factors were positively
influenced by increases in the social insect Monomorium wahibiense.

114
Species or family name Factor 1 Factor 2Adesmia sp. -0.046 -0.031Akis eievata 0.045 0.039Androctonus crassicauda 0.014 0.108Anthia duodecimguttata 0.467 0.047Apistobuthus pterygocercus -0.027 -0.009Arthrodibius cicatrix -0.031 0.015Blaps koiiari -0.093 0.042Buthacus yotvatensis 0.223 0.001Camponotus fellah -0.009 -0.029Camponotus oasium -0.031 0.015Camponotus thoracicus -0.060 -0.001Camponotus xerxes -0.031 -0.003Cataglyphis albicans 0.095 0.113Catagiyphis bombycinus 0.148 0.008Cataglyphis cinnamomeus -0.027 -0.024Cataglyphis fiavobrunneus -0.072 0.073Cataglyphis lividus -0.032 0.008Catagiyphis minimus 0.045 0.039Cataglyphis sabuiosus -0.073 -0.002Compsobuthus arabicus 0.062 0.024Crematogaster antaris 0.467 0.511Crematogaster sp. B 0.053 -0.003Eiasmobaris ?aiboguttata 0.093 0.005Eremiaphila gene 0.056 0.145Gaieodes arabs 0.013 -0.031Galeodes sp. 0.013 -0.031Hemiptera 0.090 0.079Hyaiomma sp. -0.066 0.036Lepidoptera 0.008 -0.014Mecynotarsus nr semicinctus 0.188 -0.029Melolonthinae 0.002 -0.075Mesostena puncticoilis 0.175 0.292Messor ebeninus -0.180 1.631Messor foreii -0.047 0.247Messor meridionaiis -0.108 0.054Messor sp. A 0.572 0.160Monomorium fezzanense 0.004 0.034Monomorium New sp. 0.021 -0.048Monomorium tumaire 1.116 0.163Monomorium wahibiense 1.483 0.504Mutiiiidae 0.391 0.005Myrmeleonidae Ant lion larva -0.053 -0.120nr Saprinus chalcites 0.149 -0.016Ocladius sp. 0.093 0.005Phaeotribon/Pachycera -0.040 -0.021Pimelia arabica 0.165 -0.008Prionotheca coronata 0.219 0.044Prochoma nr. ciypeaiis -0.034 -0.030Scarabaeus christatus 0.001 -0.024Scarites guineensis 0.006 -0.126Sceiosodis besnardi 0.662 0.277Scolopendrida mirabilis 0.001 -0.024Spiders (Araneae) 0.183 -0.152Tentyrina paimeri 0.515 -0.185Thermobia sp. 0.802 0.113Tiphiidae 0.188 -0.029Vachoniolus globimanus 0.052 -0.062Zophosis migneauxi -0.049 0.025
Eigenvalue
Percent of variance expiained
6.126
31.5
3.646
18.8
Table 4.11 Varimax rotated component loadings from the PCA, calculated from the
covariance matrix (n=24). Numbers in bold contributed 5% or more to the explained
variance for that factor.
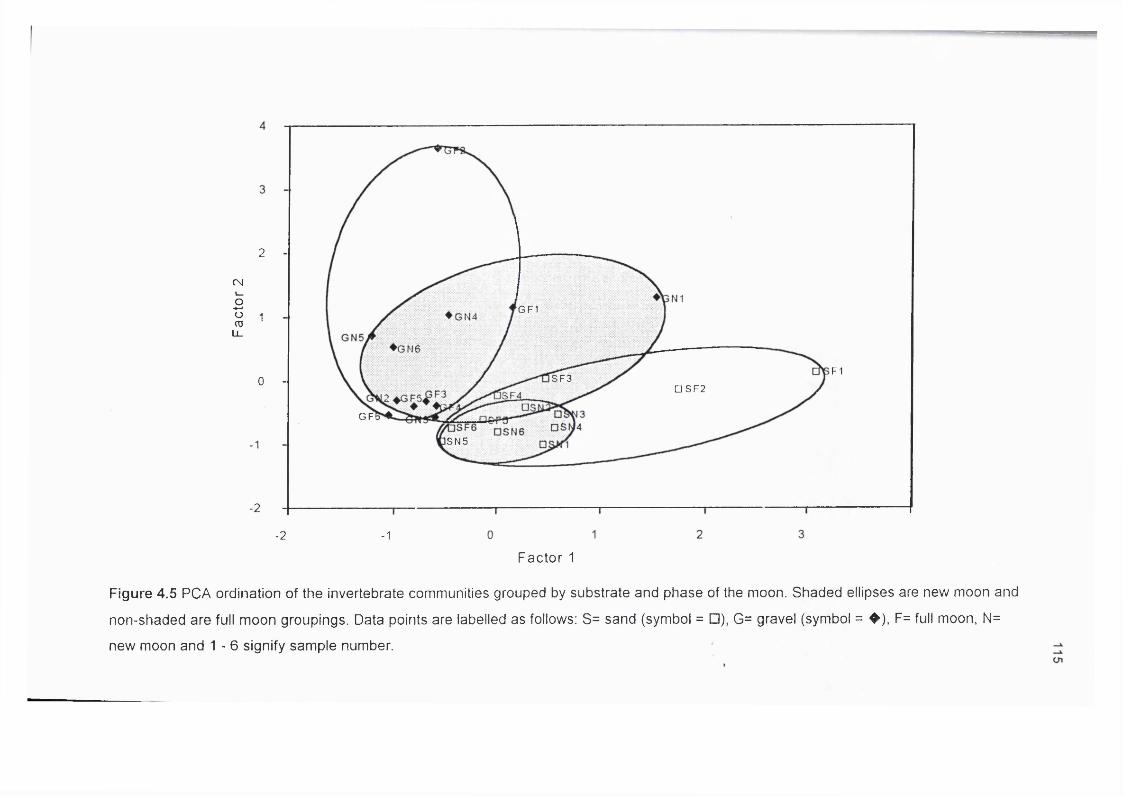
2 -
CNJ
2 % 1 u_
0 -
-1
-2
-2
GF1
SF3□ SF2
GFi
SN5
F1
-1Factor 1
Figure 4.5 PCA ordination of the invertebrate communities grouped by substrate and phase of the moon. Shaded ellipses are new moon and
non-shaded are full moon groupings. Data points are labelled as follows: 8= sand (symbol = □), G= gravel (symbol = ♦), F= full moon, N=
new moon and 1 - 6 signify sample number.

116
Figure 4.5 suggests that the community composition varies according to substrate. In
general catches on gravel had higher values for factor 2, with lower values for factor 1,
apart from GN1. In contrast, most catches on sand had higher values for factor 1, with
lower values for factor 2. Differences in the two factors between phases of the moon
were less distinct. New moon catches on sand were tightly grouped, with low values for
both factors, while full moon catches were more widely spaced. Differences between
full and new moon catches on gravel were both widely spaced with no consistent
pattern.
There also appears to be a trend with sample number (1 to 6), a proxy value for date,
within the groupings by substrate. Earlier samples, 1 and 2, were more widely spaced
with higher values for both factors, while later samples, 5 and 6, were more closely
grouped and had lower values for both factors (Figure 4.5).
4.2.3 Discussion
Higher numbers of more diverse predaceous arthropods were caught at new moons
than full moons, confirming the hypothesis that pitfall trap catches differ according to
the phase of the moon. Predaceous arthropods are intermediate level predators that
must both capture prey and avoid predation (Polis & Yamashita 1991) and a probable
explanation for their reduced activity around full moons is to avoid vertebrate predators
which take advantage of higher light levels when searching for prey. The increased risk
of vertebrate predation to other intermediate predators, including scorpions and
rattlesnakes, has been implicated in reducing their foraging activity at full moons
compared with new moons (Skutelsky 1996; Clarke et a i 1996).
In addition, those predaceous arthropods with well-developed sight may be more
aware of pitfall traps at a full moon and be able to avoid capture. For example the
large-eyed carabid Scarites guineensis was only caught during new moons. However,
scorpions and solifugids generally detect their prey via vibrations rather than visual
cues (Polis 1990b; Cloudsley Thompson 1977). Adult Buthus occitanus Israelis
scorpions showed reduced activity on simulated moonlight nights compared with dark
nights, and those that were active ambushed their prey under bushes rather than in the
open; although juveniles did not alter their activity in relation to light levels (Skutelsky
1996). These behavioural differences were thought to be based on illumination levels,
which were used to evaluate the risk of predation and availability of food. However, an

117
introduced population of Euscorpius sp. in UK also shows reduced activity during full
moons (T. Benton, pers. comm.), even though artificial lighting maintains permanently
high light-levels. Perhaps scorpions perceive moonlight through other means, such as
variations in the gravitational pull of the sun and moon upon the earth. In UAE, nine of
the 16 predaceous arthropods taxa recorded, including three scorpions {Apistobuthus
pterygocercus, Vachoniolus globimanus and Compsobuthus arabicus) were never
captured at a full moon (Table 4.7). As well as vertebrate predation, small species or
juvenile predaceous arthropods will be subjected to intra-guild predation (Polis &
Yamashita 1991). While the size of the arthropods captured in UAE was not recorded,
adult C. arabicus never exceeded 3 cm in body length and are much smaller than other
scorpions. For example, Androctonus crassicauda can measure over 9 cm (Vachon
1979) and was present at both full and new moons.
Other studies from the Negev desert where B. o. Israelis occurs, suggest that
vertebrates such as white storks Ciconia ciconia and stone curlews Burhinus
oedicnemus reduce the activity and abundance of arthropods, especially tenebrionids
(Ayal & Merk11994). In Arabia, houbara often forage at night (Anegay 1994),
particularly on nights with bright moon light, and this is thought to give them a visual
advantage when hunting nocturnal animals (Combreau & Launay 1996).
Preliminary data also suggested that the phase of the moon affected the
activity/abundance of other invertebrates (Tables 4.7 & 4.8), but these effects were not
significant. Some taxa may have been placed in the wrong biological group for the
analysis, e. g. predatory ants could be placed with the predaceous arthropods.
However, the lack of knowledge of the biology of the Arabian fauna makes it difficult to
determine guilds and trophic levels, and is compounded by the complexity of desert
food webs where omnivory and trophic diversity interact, and animals do not always
remain within one trophic level (Polis 1991b). For example, some granivorous ants
deviate from the characteristic trophic level of their genus (MacKay 1991) and will also
feed upon arthropods in order to increase their dietary protein and water levels. In
terms of numbers captured in UAE, ants were dominated by granivores {Messorsp.)
and general foragers associated with aphids and their exudates {Crematogastersp.).
The PCA ordination suggested that changes in the abundance of six species of ants
and three macroarthropod detritivores made a significant contribution to the two factors
which explained about half the variation in the capture of arthropod taxa. PCA also

118
indicated that there was a strong effect of substrate on community composition,
agreeing with the findings of a larger data set (Tigar & Osborne 1997; Section 4.1).
The strong influence of substrate on the numbers and diversity of arthropods captured
in the pitfall trap may have reduced the effect of the phase of the moon, and there was
a significant interaction for these two terms in the social insects.
The experiment was designed around the lunar calendar and although cloud cover is
rare in Abu Dhabi, it is more frequent during winter months (Bottomley 1996). Rather
than using the categorical variables full and new moon, a numerical axis, related to the
number of hours and brightness of moonlight during the trapping period, might explain
more of the variance in the trap catch data. Unfortunately, no suitable equipment to
measure moonlight was available during the experiments. While moonlight was shown
to affect the catch of predators, no effects were found for other arthropods, which may
be more sensitive to absolute levels of moonlight.

119
4.3 Patterns of available biomass and diversity of flying Insects
4.3.1 Introduction
While houbara generally eat ground-dwelling insects (see Chapter 2), some taxa that
are unlikely to be sampled by pitfall traps have also been recorded as prey, for
example Orthoptera, Curcuiionidae and Lepidoptera (see Gavrin 1962; Fox 1988;
Collins 1984). Therefore a light trap was used to sample flying insects and to indicate
their seasonal abundance and biomass in Abu Dhabi.
4.3.2 Methods
The light trap
A Heath light trap (as described in Chapter 3) was placed overnight at a sandy location
once every 28 days for two years at five trapping sites (Section 4.1). Traps were
operated on the same dates as pitfall traps, and air temperature and relative humidity
were recorded (see Section 4.1). The invertebrates captured were sorted to order, and
in a few cases to family, and counted. Most of the catch was not identified further, but
a few representative samples were retained for taxonomic reference and are listed in
Appendix 4.1. In addition, the dung beetle Scarabaeus cristatus was recorded to
species level. The catch was oven-dried to constant weight at 60°C to indicate
available biomass of flying insects. However, if examples had been removed for
taxonomic purposes their dry weights were estimated from the remaining data.
Data Analvsis
Trap catch was log-transformed and was found to conform to a normal distribution
using Lilliefors' test. GLM on Systat (Wilkinson 1993) was used to test for differences
in trap catch between the five sites, between months of the year and for the effect of
minimum air temperature on catch. The catch across the five sites was used to give a
monthly mean catch with standard error. However, analyses are limited because there
is only one trap catch per site per month.
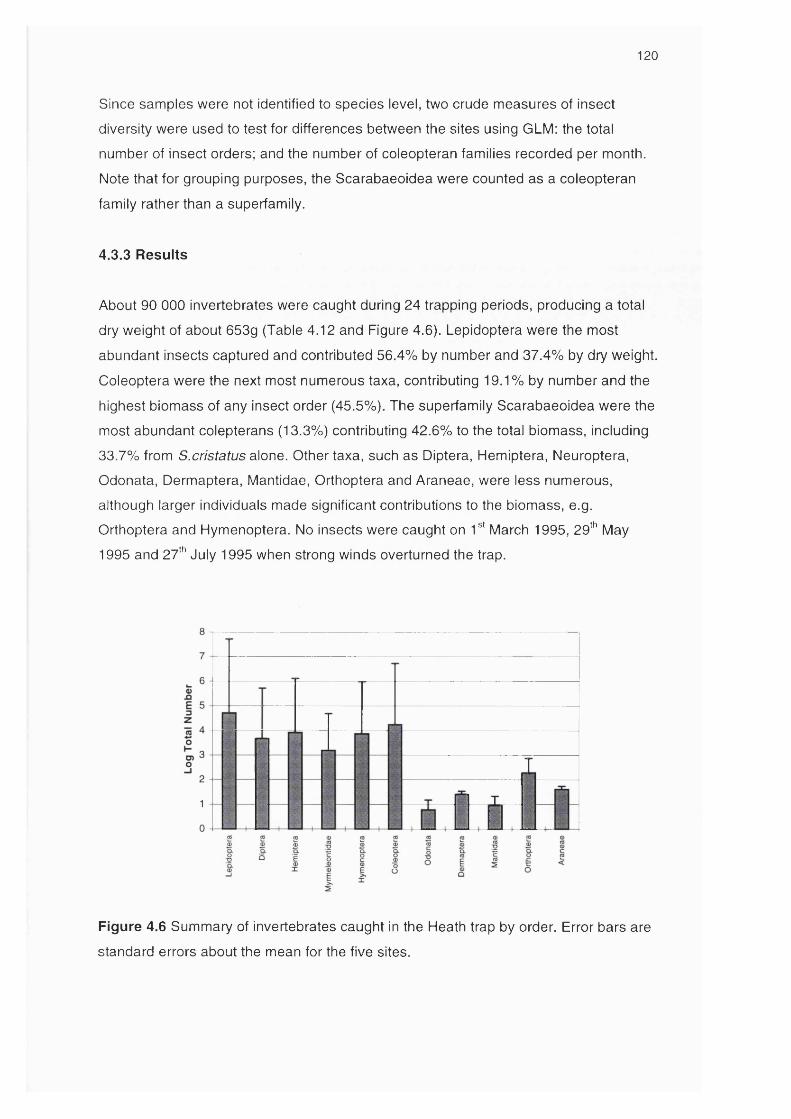
1 2 0
Since samples were not identified to species level, two crude measures of insect
diversity were used to test for differences between the sites using GLM: the total
number of insect orders; and the number of coleopteran families recorded per month.
Note that for grouping purposes, the Scarabaeoidea were counted as a coleopteran
family rather than a superfamily.
4.3.3 Results
About 90 000 invertebrates were caught during 24 trapping periods, producing a total
dry weight of about 653g (Table 4.12 and Figure 4.6). Lepidoptera were the most
abundant insects captured and contributed 56.4% by number and 37.4% by dry weight.
Coleoptera were the next most numerous taxa, contributing 19.1% by number and the
highest biomass of any insect order (45.5%). The superfamily Scarabaeoidea were the
most abundant colepterans (13.3%) contributing 42.6% to the total biomass, including
33.7% from S.cristatus a\or\e. Other taxa, such as Diptera, Hemiptera, Neuroptera,
Odonata, Dermaptera, Mantidae, Orthoptera and Araneae, were less numerous,
although larger individuals made significant contributions to the biomass, e.g.
Orthoptera and Hymenoptera. No insects were caught on 1®* March 1995, 29** May
1995 and 27' July 1995 when strong winds overturned the trap.
Figure 4.6 Summary of invertebrates caught in the Heath trap by order. Error bars are
standard errors about the mean for the five sites.
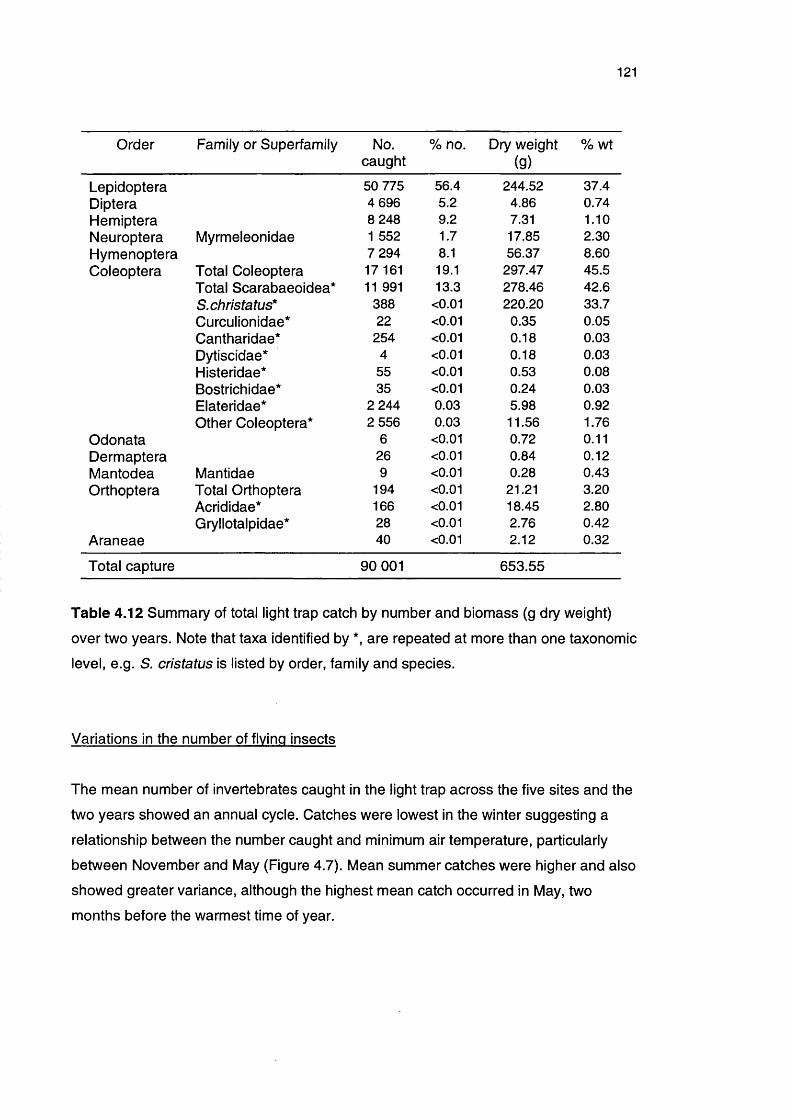
121
Order Family or Superfamily No.caught
% no. Dry weight(g)
% wt
Lepidoptera 50 775 56.4 244.52 37.4Diptera 4 696 5.2 4.86 0.74Hemiptera 8 248 9.2 7.31 1.10Neuroptera Myrmeleonidae 1 552 1.7 17.85 2.30Hymenoptera 7 294 8.1 56.37 8.60Coleoptera Total Coleoptera 17 161 19.1 297.47 45.5
Total Scarabaeoidea* 11 991 13.3 278.46 42.6S. christatus* 388 <0.01 220.20 33.7Curcuiionidae* 22 <0.01 0.35 0.05Cantharidae* 254 <0.01 0.18 0.03Dytiscidae* 4 <0.01 0.18 0.03Histeridae* 55 <0.01 0.53 0.08Bostrichidae* 35 <0.01 0.24 0.03Elateridae* 2 244 0.03 5.98 0.92Other Coleoptera* 2 556 0.03 11.56 1.76
Odonata 6 <0.01 0.72 0.11Dermaptera 26 <0.01 0.84 0.12Mantodea Mantidae 9 <0.01 0.28 0.43Orthoptera Total Orthoptera 194 <0.01 21.21 3.20
Acrididae* 166 <0.01 18.45 2.80Gryllotalpidae* 28 <0.01 2.76 0.42
Araneae 40 <0.01 2.12 0.32
Total capture 90 001 653.55
Table 4.12 Summary of total light trap catch by number and biomass (g dry weight)
over two years. Note that taxa identified by *, are repeated at more than one taxonomic
level, e.g. S. cristatus is listed by order, family and species.
Variations in the number of fivinq insects
The mean number of invertebrates caught in the light trap across the five sites and the
two years showed an annual cycle. Catches were lowest in the winter suggesting a
relationship between the number caught and minimum air temperature, particularly
between November and May (Figure 4.7). Mean summer catches were higher and also
showed greater variance, although the highest mean catch occurred in May, two
months before the warmest time of year.
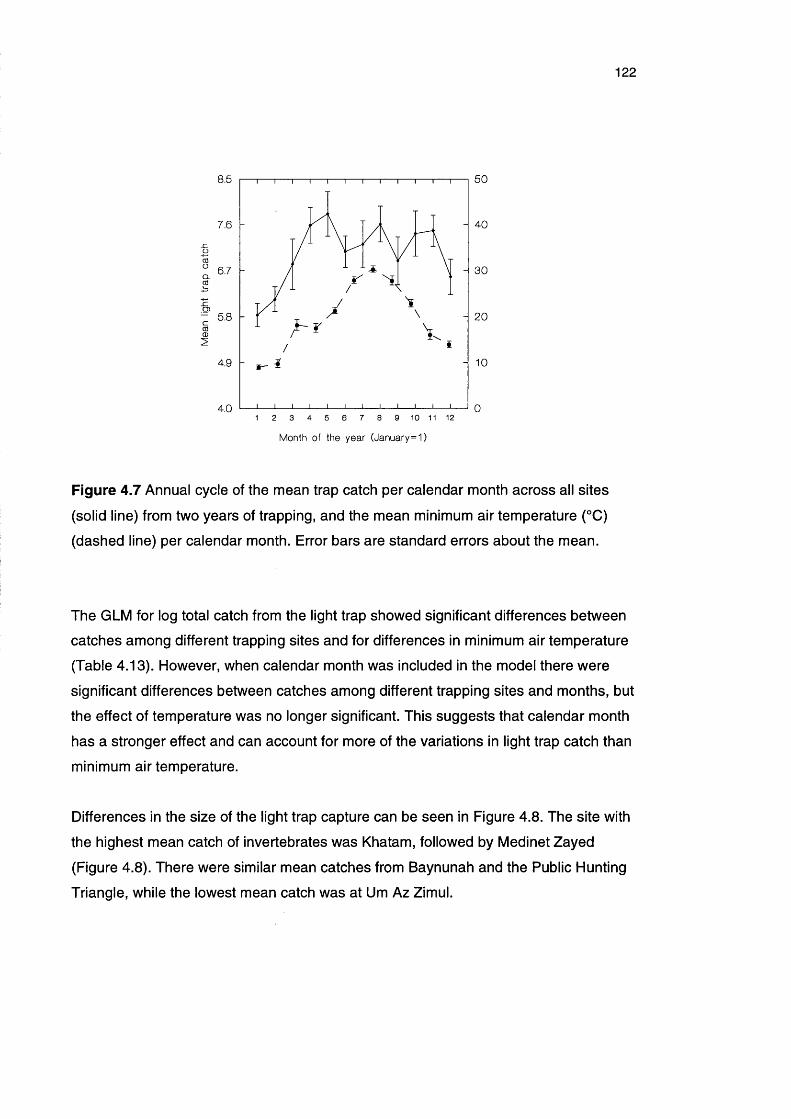
122
8.5
7.6
6.7
5.8
4.9
4.0
50
40
30
20
10
1 2 3 4 5 6 7 8 9 10 11 12
Month of the year (January=1)
Figure 4.7 Annual cycle of the mean trap catch per calendar month across all sites
(solid line) from two years of trapping, and the mean minimum air temperature (°C)
(dashed line) per calendar month. Error bars are standard errors about the mean.
The GLM for log total catch from the light trap showed significant differences between
catches among different trapping sites and for differences in minimum air temperature
(Table 4.13). However, when calendar month was included in the model there were
significant differences between catches among different trapping sites and months, but
the effect of temperature was no longer significant. This suggests that calendar month
has a stronger effect and can account for more of the variations in light trap catch than
minimum air temperature.
Differences in the size of the light trap capture can be seen in Figure 4.8. The site with
the highest mean catch of invertebrates was Khatam, followed by Medinet Zayed
(Figure 4.8). There were similar mean catches from Baynunah and the Public Hunting
Triangle, while the lowest mean catch was at Um Az Zimul.
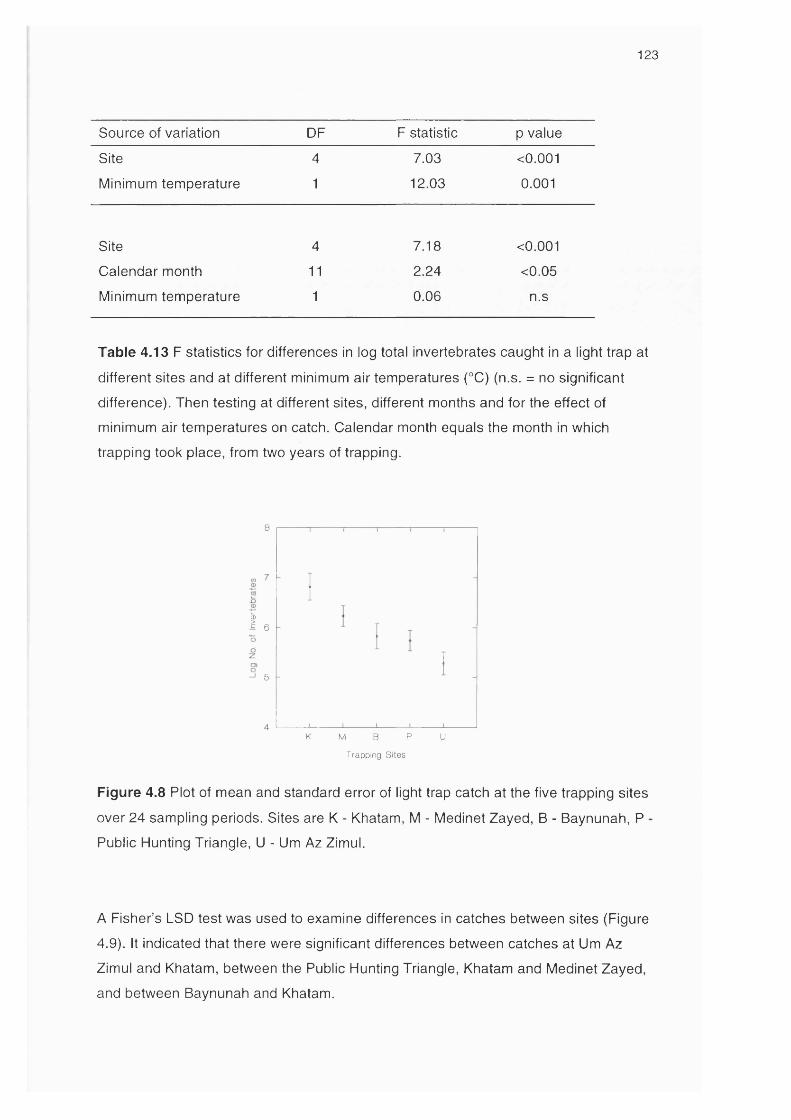
123
Source of variation DF F statistic p value
Site 4 7.03 <0.001
Minimum temperature 1 12.03 0.001
Site 4 7.18 <0.001
Calendar month 11 2.24 <0.05
Minimum temperature 1 0.06 n.s
Table 4.13 F statistics for differences in log total invertebrates caught in a light trap at
different sites and at different minimum air temperatures (°C) (n.s. = no significant
difference). Then testing at different sites, different months and for the effect of
minimum air temperatures on catch. Calendar month equals the month in which
trapping took place, from two years of trapping.
8
I 6
5
B P UK M
Trapping Sites
Figure 4.8 Plot of mean and standard error of light trap catch at the five trapping sites
over 24 sampling periods. Sites are K - Khatam, M - Medinet Zayed, B - Baynunah, P -
Public Hunting Triangle, U - Um Az Zimul.
A Fisher’s LSD test was used to examine differences in catches between sites (Figure
4.9). It indicated that there were significant differences between catches at Um Az
Zimul and Khatam, between the Public Hunting Triangle, Khatam and Medinet Zayed,
and between Baynunah and Khatam.

124
U P B M K
Figure 4.9 Differences in log catch at the five sites (post hoc test by Fisher's LSD
paired comparisons). Sites are ordered by abundance (smallest from left to right) and
the ends of each line joins sites that are significantly different at p<0.05 or better. K -
Khatam, M - Medinet Zayed, B - Baynunah, P - Public Hunting Triangle, U - Um Az
Zimul.
Variations in insect diversitv between sites
There were significant differences between both the number of orders of flying insects
and the number of coleopteran families caught between sites (Table 4.14). Calendar
month and minimum air temperature were used as covariates, since they were likely to
influence the number of invertebrates caught in the light trap (Table 4.13).
A post hoc comparison using Fisher’s LSD paired test showed that the number of
orders of flying insects caught at Khatam was significantly different from all other sites
(p< 0.05). This was due to a consistently higher mean catch of insect orders at Khatam
(Figure 4.10). For the number of coleopteran families caught, Fisher’s LSD showed
that Um Az Zimul was significantly different from Medinet Zayed, the Public Hunting
Triangle and Khatam (p< 0.05) (Figure 4.11). The number of coleopteran families
caught at Baynunah was also significantly different from Khatam and the Public
Hunting Triangle (p<0.01). These differences result from consistently low mean
catches in the number of coleopteran families at Um Az Zimul and Baynunah (Figure 4.12).
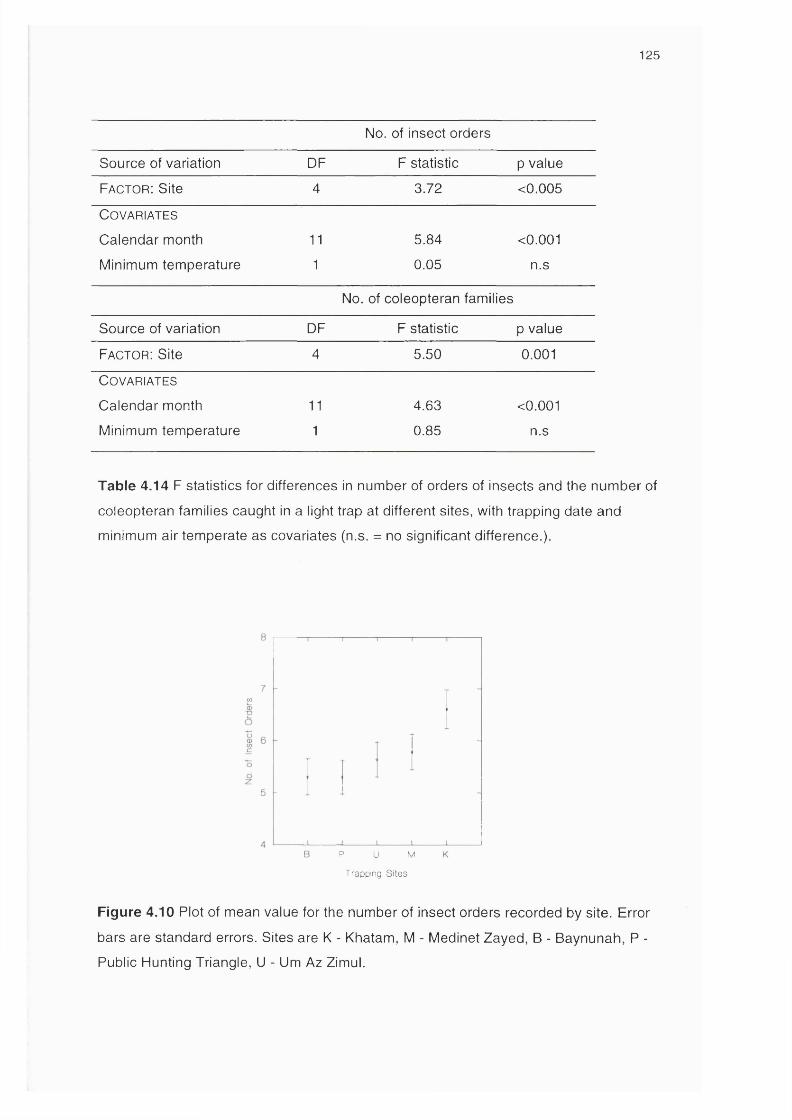
125
No. of insect orders
Source of variation DF F statistic p value
Fa c t o r : Site 4 3.72 <0.005
C o va r ia tes
Calendar month 11 5.84 <0.001
Minimum temperature 1 0.05 n.s
No. of coleopteran families
Source of variation DF F statistic p value
Fa c t o r : Site 4 5.50 0.001
C o va r ia tes
Calendar month 11 4.63 <0.001
Minimum temperature 1 0.85 n.s
Table 4.14 F statistics for differences in number of orders of insects and the number of
coleopteran families caught in a light trap at different sites, with trapping date and
minimum air temperate as covariates (n.s. = no significant difference.).
P U M
Trapping Sites
Figure 4.10 Plot of mean value for the number of insect orders recorded by site. Error
bars are standard errors. Sites are K - Khatam, M - Medinet Zayed, B - Baynunah, P -
Public Hunting Triangle, U - Um Az Zimul.
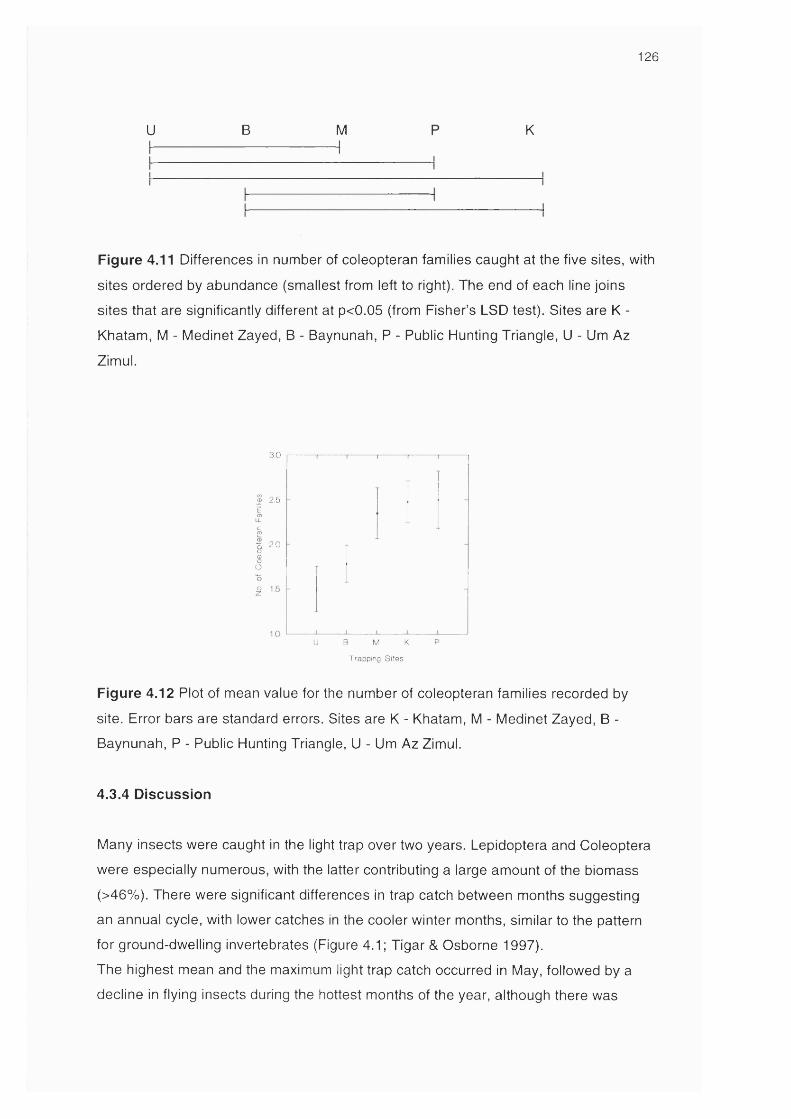
126
U B M K
Figure 4.11 Differences in number of coleopteran families caught at the five sites, with
sites ordered by abundance (smallest from left to right). The end of each line joins
sites that are significantly different at p<0.05 (from Fisher’s LSD test). Sites are K -
Khatam, M - Medinet Zayed, B - Baynunah, P - Public Hunting Triangle, U - Um Az
Zimul.
3.0
I 2,5
1 §K 2,0
f01
u B M K P
Trapping Sites
Figure 4.12 Plot of mean value for the number of coleopteran families recorded by
site. Error bars are standard errors. Sites are K - Khatam, M - Medinet Zayed, B -
Baynunah, P - Public Hunting Triangle, U - Um Az Zimul.
4.3.4 Discussion
Many insects were caught in the light trap over two years. Lepidoptera and Coleoptera
were especially numerous, with the latter contributing a large amount of the biomass
(>46%). There were significant differences in trap catch between months suggesting
an annual cycle, with lower catches in the cooler winter months, similar to the pattern
for ground-dwelling invertebrates (Figure 4.1; Tigar & Osborne 1997).
The highest mean and the maximum light trap catch occurred in May, followed by a
decline in flying insects during the hottest months of the year, although there was

127
considerable variance (Figure 4.7). The catches contained many herbivores, such as
Lepidoptera, Orthoptera and some Hemiptera and Coleoptera, which may have
reduced activity linked with the limited availability of plant foods during the heat of
summer. Insects are poïkilothermie and usually have a body temperature close to that
of their environment (Mordue et al. 1980). However, active metabolic processes,
including flight, generate heat and in warm climates large insects may require rest
periods to avoid overheating (Wigglesworth 1972). The reduction in catch size
between June and October suggests that some insect were trapped less frequently
because they flew for a shorter duration to avoid heat-stress and dehydration. The
highest value for the minimum air temperature recorded during light-trapping in Abu
Dhabi was 34.7°C (August 1995), but maximum daily temperatures for this period
would exceed 50°C (Bottomley 1996). Therefore, during sustained flight large insects
soon reach a lethal body temperature (e.g. 45°C for locusts; Wigglesworth 1972).
The variation in catches between the five sites probably reduced the effect of
temperature on trap catch in the analysis (Table 4.13). The relative abundance and
diversity of invertebrates in light trap catches across sites showed some similarities to
pitfall trap catches (Section 4.1.3). Khatam had the highest mean light trap catch and
the highest measures of diversity of flying insects; while the lowest mean light trap
catch came from Um Az Zimul, the furthest inland and most arid site (Figure 4.7) which
also showed reduced diversity for flying Coleoptera (Figure 4.12). There were
consistently high catches of ground-dwelling invertebrates at Khatam (Section 4.1.3),
although the highest log catch came from Baynunah. The light trap at the Public
Hunting Triangle caught the most coleopteran families, although this result was not
significantly different from Khatam or Medinet Zayed. However, the Public Hunting
Triangle was the most species-rich site for ground-dwelling invertebrates (Section
4.2.3; Tigar & Osborne 1997)
Aerial insects disperse greater distances than ground-dwelling invertebrates, therefore
light traps are likely to sample insects from a greater range than pitfall traps. This may
explain differences between the relative size and diversity of light trap and pitfall
catches at one trapping site. In addition, some of the insects captured in the light trap
are not normally part of the desert community. For example migratory insects, such as
Orthoptera, Lepidoptera, Odonata and Coleoptera, travel thousands of kilometres
across the Arabian Peninsula (Popov 1980; Wiltshire 1990; Walker & Pittaway 1987).

128
Holometabolous insects with aquatic immature stages, such as Odonata and
Dytiscidae, were captured far from any standing water.
A small proportion of the invertebrates in the light trap were predatory (Araneae,
Mantidae and some Hymenoptera), and probably entered the trap in search of prey.
Although ballooning is a well-known method of aerial dispersal in spiders (Wise 1993),
most of the spiders captured were too large to have dispersed in this manner and had
probably climbed into the trap.
One of the aims of this chapter was to look at available prey for houbara. While flying
insects were abundant throughout the year the majority were minute, with a mean dry
weight of <0.01 g, and of little value as food for a bird the size of a houbara. Larger
prey were comparatively rare and only 166 acridid grasshoppers were caught over two
years. Outbreaks of the locusts Schistocera gregaria and Locusta migratoria have
been recorded in the region, but such occurrences are sporadic (Popov 1980).
However, they may be more common in other parts of the houbara’s range such as
Central Asia, where locusts are an important prey (Gavrin 1962). The large dung
beetle S. cr/sfafus frequently occurred in the light trap, but there is evidence to suggest
that even when similar beetles are abundant that they are not eaten by houbara,
although the reason for ignoring such large prey is not clear (Gubin & Mukhina in
prep.). The role of invertebrates in the diet of the houbara is discussed in detail in
Chapter 5.

129
4.4 General Discussion on Abu Dhabi’s Invertebrates
The invertebrate communitv
Previous accounts of invertebrates from Abu Dhabi and UAE have emphasised taxa
from mesic and man-altered habitats (see reviews in Chapter 1; Tigar 1996a & b;
Appendices 1.1 & 1.2). However, desert arthropods are locally abundant and a
frequent component of Abu Dhabi's deserts. Currently, there are 508 records from this
region, of which 303 were found during this study and include 196 new records for the
UAE (Appendix 4.1). Some have yet to be identified to species level, and many more
are undoubtedly awaiting discovery, particularly microfauna (see Zak & Freckman
1991) which were not included in my study.
Large detritivorous tenebrionids formed a major part of the invertebrate community and
49 species or subspecies were recorded (Appendix 4.1). As in other deserts (Holm &
Scholtz 1980; Crawford & Seely 1987) these beetles contributed about one third of the
total desert species in Abu Dhabi (Table 4.7). Tenebrionids, thysanurans and other
omnivores are particularly important in deserts because they recycle energy from
detritus, making it available to organisms higher up the food chain (Seely & Louw
1980). As in other deserts they also contributed much of the biomass (see Crawford
1991).
Predaceous arthropods are another abundant and diverse component of desert
ecosystems, and are more numerous in arid than mesic environments (Polis &
Yamashita 1991). The combination of their physiology, particularly a low metabolic rate
and plasticity of prey choice, allows them to survive conditions of unpredictable and low
food supply typical of deserts (Polls 1991b). In UAE they formed slightly less than a
third of the desert species (Table 4.7), but the taxonomy of some orders, particularly
the Araneae, Mutillidae and other parasitoids, is poorly known and their diversity was
underestimated. The largest arachnids may also have been under-recorded because
they escaped from or avoided capture in the pitfall traps.
Knowledge of Arabian ants continues to improve (Collingwood 1985; Collingwood &
Agosti 1996) and 65 species have been recorded in UAE (Collingwood etal. 1997;
Appendix 4.1). These include 12 species of the genus Cataglyphis\Nh\ch are
structurally adapted for digging in loose sand and are able to survive surface

130
temperatures of 50°C (Delye 1968). However, outside desert locations, Introduced or
tramp species form a high proportion of UAE's ants (about 20%; see Collingwood etal.
1997) and include Solenopsis geminata which is well-known for its deleterious effect
on endemic insects in USA (Williams 1994). Other desert social insects are poorly
known, although there are at least two species of termites (Appendix 4.1). Most ants
and termites live in large colonies and form a high proportion of the desert biomass
(MacKay 1991). They are trophically diverse, although many are detritivores or general
scavengers. In Abu Dhabi ants were the most numerous animals in the pitfall traps.
Aerial insects were plentiful, and included many herbivores especially Lepidoptera, as
well as some predators and omnivores. Studies from North American deserts suggest
that desert-adapted trees and shrubs support a wide variety of invertebrates. For
example, mesquite Prosopis glandulosa has a resident fauna of over 200 arthropod
species which depend either directly or indirectly on the tree for food and shelter
(Simpson 1977; Ward etal. 1977). Each tree acts as a separate habitat patch, and by
moving between different patches herbivorous insects remain persistent at regional
levels in arid environments (Wiens 1976), although at local levels they show
considerable temporal and spatial variation in numbers and community composition
(Wisdom 1991). While native trees are relatively scarce in Abu Dhabi, both trees and
shrubs of Prosopis cinerla and Acacia tortills occur, and are probably used by many of
the aerial insects captured in the light trap for at least part of their life cycle.
Underground herbivory on roots and tubers is another important trophic level in
deserts, where above-ground green plants are scarce and infrequent (Ludwig 1977).
The immature forms of the Cicadidae, Scarabaeoidea, Elateridae and Tenebrionidae
caught in the light trap, are likely to include root and tuber feeders.
Temporal patterns
Both ground-dwelling and aerial insects showed similar patterns of seasonal
abundance (Figures 4.1 and 4.7). Fewer insects were trapped in the colder winter
months, coinciding with the period when rainfall is most likely to occur, although
temperature and rainfall could not be separated in the analysis. Climate-induced
seasonal variations in invertebrate abundance occur in many deserts (Ahearn 1971;
Ayal & Merk11994; Holm & Edney 1973; Seely 1991; Wisdom 1988), and most desert
animals and plants respond rapidly to environmental changes (Polls 1991a).

131
Temporal segregation, whereby different species forage at different times of day, is
thought to allow more species to coexist on the limited food available in deserts. This
has been demonstrated for omnivores, including ants (MacKay 1991) and tenebrionids
(Crawford 1991), and predatory arthropods (Polis & Yamashita 1991).
Spatial patterns
There were differences in the number and diversity of invertebrates at the five trapping
sites. Khatam appeared to have consistently high numbers of invertebrates and the
Public Hunting Triangle was particularly rich in terms of both ground-dwelling and flying
invertebrates, although the reasons for this are not clear. Both biotic and abiotic factors
influence faunal abundance and diversity, and the intensity of grazing is probably an
important factor in Abu Dhabi.
In the Namib Desert, Seely (1991) found that the species richness of ground-dwelling
arthropods on sand depended upon the height of dunes, the ratio of vegetated to non
vegetated surface area, the quantity of detritus and the distance from the coast. The
latter determined the amount of fog, which is the major source of water in this area
with very low rainfall. There were also differences in the faunal composition of
compacted inter-dunes and sand dunes, with similar effects in Abu Dhabi for substrate
(mobile sand or compacted gravel plains). The ever-increasing presence of irrigated
farms and plantations in UAE provides an additional food supply for herbivorous
insects, and may result in many local faunal changes. Invertebrate diversity is also a
consequence of the region's biogeography, with the southern parts of UAE bordering
on Oman containing more elements of the African flora than northern, coastal areas
(Mandaville 1990), which are likely to have their own associated arthropod fauna. Two
sites, Um Az Zimul and Khatam are within 20km of the Omani border, while Baynunah
and Medinet Zayed are coastal and the Public Hunting Triangle is intermediate to both
the coast and the Omani border.
Implications for biodiversitv studies
Biodiversity studies should consider the effect of annual and daily cycles, temperature,
phase of the moon and spatial variation when carrying out surveys. In particular, the
local and regional variation of patchy environments such as deserts demands

132
rigorously designed, randomly-stratified sampling techniques (see Osborne 1991,
1992).
Natural historians often centre their observations in atypical habitats, including wadis,
parks, gardens, irrigated farms, man-made dams and sewage works, where water is
no longer a limiting factor to fauna and flora. While such environments may be useful
for international migrants, particularly birds, the effects of the increasing amount of
mesic areas in UAE on endemic or desert-adapted wildlife are less clear. However, the
ranges of non-desert birds including feral pigeons Columba livia, collared doves
Streptopelia decaocto and house sparrows Passer domesticus have recently expanded
along the tracts of irrigated plantations and by following water transported deep into
the desert for livestock (Osborne etal. 1996). The high proportion of introduced ant
species in UAE (Collingwood etal. 1997), highlights the ease with which small, non
native arthropods can establish themselves. Such minor changes often go unnoticed,
but there are numerous global examples of the devastation to wildlife and agriculture
caused by an accidental or intentional introduction of non-native species (see Mooney
& Drake 1986; Godfrey & Crawley 1998).
UAE is beginning to recognise and value its own fauna and flora (Osborne etal. 1996)
with animals such as the houbara (Bailey & Hornby 1994) and Arabian leopard (Hellyer
1996) acting as flagship species. However, further efforts to understand the natural
history and ecology of the desert should be encouraged. In particular the less
charismatic, arthropod element has largely been ignored and yet forms a greater total
biomass than vertebrates (MacKay 1991 ) and plays a critical role in the cycling of
elements and energy-flow throughout the desert ecosystem.

133
C h a p t e r 5
T h e d ie t o f t h e h o u b a r a b u s t a r d in A b u D h a b i
Synopsis of Chapter 5
This Chapter presents information on the diet of the houbara in Abu Dhabi based on
the examination of faeces and gizzard contents. In addition, access to captive birds
enabled calibration trials to be carried out, so that counts of prey remains in faeces
could be adjusted for differential digestion. While emphasis was on animal foods, plant
remains were identified wherever possible and further refinements for dietary studies
are suggested. The reconstructed diet was used to estimate houbara energy
consumption and compared with the actual consumption of an artificial diet by captive
birds .

134
5.1 General Introduction
Our understanding of the diet and feeding ecology of houbara in Abu Dhabi is poor
(see Chapter 2). More precise information will help habitat management in terms of
food availability for over-wintering and captive-bred, released houbara, and may
explain the seasonal distribution of these nomadic migrants across their range. A full
understanding of the diet of wild birds will indicate how the composition of captive diets
can be improved, including the relative proportions of appropriate food types and
nutrient balance. Better enclosures for houbara could incorporate features that attract
or encourage natural foods, thereby increasing available food and providing a more
stimulating environment.
5.2 Quantifying the diet via faecal analysis
5.2.1 Introduction
Houbara eat a variety of animal and plant food, and invertebrates are frequently cited
as important prey (Cramp & Simmons 1980; Johnsgard 1991). The birds feed
throughout the day but are generally crepuscular (Hinz & Heiss 1989) or nocturnal
(Anegay 1994), which combined with their cryptic coloration and extreme wariness
makes observations of foraging difficult to obtain. Most descriptions of the diet are
reports of gizzard contents or chance observations of feeding (see Chapter 2). Only
Collins (1984, 1993) and Gubin & Mukhina (in prep.) have quantified the diet from food
remains in faeces, but neither assessed the destruction of prey during digestion. As
Green & Tyler (1989) point out, avian dietary studies based on faecal analysis must
account for differential digestion or their conclusions may be invalid.
Houbara faeces contain much finely ground material that is difficult to identify and
quantify. To simplify the examination of faeces prey were categorised into functional
groups, which were more relevant to foraging birds than taxonomy (Cooper etal.
1990), and diagnostic fragments were selected for each group (see Ralph etal. 1985;
Moreby 1988). Feeding trails with captive houbara examined the recovery rates of prey
fragments in faeces, and consistent “key fragments" were identified which enabled the
correction of differential digestion and calculation of initial invertebrate consumption.
Basic accounts of plant foods and calculated estimates of houbara energy
requirements were also included. Finally, estimated invertebrate consumption was

135
compared with prey abundance as measured by pitfall traps (in Chapter 4), to try to
address the question of whether houbara are food limited in UAE.
5.2.2 Methods
Calibration experiments with animal prey
Potential prey were identified from the literature (see Chapter 2) and by qualitative
examination of wild houbara faeces and compared to an invertebrate reference
collection. Prey were classified into 16 groups according to their size, daily activity
rhythm, habit, speed and type of locomotion (Table 5.1). Live prey were available for all
trials apart from adult mice which were used as models for desert rodents.
Wild-caught adult houbara that had been in captivity for about 6 years were used in the
trials. Birds only had access to the experimental diet and were kept separately in
rooms which received natural light and were air-conditioned to about 24°C. The birds
were allowed to settle for two weeks before experiments began, and each received
75g of dry food pellets and free access to water daily. Feeding with pellets stopped 12
hours prior to each experimental meal. On the first day of each trial, birds were given a
known number of a single prey group (Table 5.1) together with pellets to a total weight
of 75g. Any uneaten prey were removed after three hours and counted.
On each subsequent day, birds were returned to a diet of 75g pellets. Faeces were
collected every 12 hours for one week and examined for prey remains. A different prey
type was given at the start of each consecutive experiment so that remains from
previous trials could be distinguished. Where possible trials were replicated for each
prey group on 4-5 birds, although lack of available prey meant that not all prey types
could be tested on every bird (Appendix 5.1).
Preliminary work indicated a rapid passage time through the gut (about 4 days, see
Appendix 5.2) and it was assumed that prey fragments not recovered within 14 days
were digested or crushed beyond recognition. Prey remains were removed from the
faeces and their recovery rate was calculated as the ratio of the number recovered to
the number consumed. Means and standard errors (between trials) were calculated
using the formulae for ratios described by Snedecor & Cochran (1967).

Preygroup
Size (cm) Activity regime Habit ' Speed Type of Movement
Taxonomic groups Examples from UAE
1 <0.5 Nocturnal Terrestrial & arboreal
Slow Walk Hymenoptera: Formicidae. Ants Camponotus xerxes
2 <0.5 Diurnal Terrestrial & arboreal
Slow Walk HymenopteraiFormicidae. Ants Cataglyphis spp., Crematogaster spp., Messor spp.
3 0.5-1.5 Nocturnal Arboreal Slow Walk or crawl Coleoptera:Curculionidae, Lepidoptera (larvae)
eg Bothyrideres anxius Ocladius sp., Ammocleonus sp. (Phytophagous insects)
4 0.5-1.5 Nocturnal Terrestrial Slow Walk ColeopteraiT enebrionidae Small ground-dwelling tenebrionids
Mesostena puncticollis
5 0.5-1.5 Nocturnal Terrestrial and arboreal
Slow Walk ColeopteraiT enebrionidae Small arboreal tenebrionids
Tentyrina palmeri
6 1.5-5 Nocturnal Terrestrial Slow Walk ColeopteraiT enebrionidae Large tenebrionids
Blaps kollari, Prionotheca coronata, Pimelia arabica, Ocnera philistina, Akis elevator.
7 1.5-5 Nocturnal Terrestrial Fast Walk Coleoptera: Carabidae Anthia duodecimguttata, Scarifies guineensis8 1.5-5 Diurnal Terrestrial,
arboreal & aerialFast Walk, jump and
flyOrthoptera:Acridida. Sfiort horned grasshoppers and locusts
Schistocerca gregaria, Pyrgomorpha conica, Truxalis procera, Heteracris littoralis.
9 1.5-5 Nocturnal Terrestrial Fast Run Solifugae. Camel spiders Galeodes sp.10 1.5-5 Nocturnal Terrestrial Slow Walk Scorpionae: Scorpions Buthicus yotvatensis nigroaculeatus,
Vachoniolus spp.11 >5 Diurnal Terrestrial Fast Walk Reptilia. Day active lizards Acanthodactylus spp.12 >5 Nocturnal Terrestrial Fast Walk Reptilia. Gekkos and lizards Bunopus tuberculatus, Stenodactylus spp.13 >5 Nocturnal Terrestrial Fast Walk and jump Rodentia: Small mammals gerbil,
and gerboasGerbillus cheesmani
14 >0.05-4 Nocturnal/diurnal
Terrestrial & arboreal
Fast Walk and run Aranea: Spiders (very diverse) Gnaphosidae, Salticidae, Lycosidae, Sparassidae (indeterminate species)
15 2.0-5 Diurnal & Nocturnal
Aerial and terrestrial
Slow - Fast
Fly or walk Flying insects eg Buprestidae, Scarabaeiodea, Lepidoptera
Julodius spp., Julodella spp. Scarabaeus christatus
16 1.5-4.0 Mainly diurnal Terrestrial Slow - medium
Walk, run or “sand swim”
Surface dwelling, diurnal tenebrionids
Erodius spp., Adesmia spp.
Table 5.1. Functional groups of potential animal prey of houbara bustards in UAE. The term terrestrial implies a predominantly ground-dwelling animal, arboreal indicates one that climbs or lives on plants (usually a herbivore), and aerial indicates an active, free-flying insect.
wO i

137
Faecal sample collection from wild birds
Houbara are the largest of three species of three-toed birds in UAE (the others are
the stone curlew and the cream coloured courser) and have a characteristic
footprint. Wintering houbara occur at low densities in Abu Dhabi (Osborne 1996a)
and unlike resident or breeding birds, they do not have regular roost or display sites
(e.g. Collins 1984; Van Heezik & Seddon, in press). Instead houbara move singly
through the desert and faeces were found by walking along bird tracks, which
remain visible for several days on sandy substrates. Faeces were placed in plastic
bags and kept cool until they could be frozen at -5°C.
Faecal analvsis
Defrosted faecal samples were softened in water and teased apart using a glass
rod over a 200 pm sieve to separate out the prey fragments. Irrigated samples were
transferred to a white ceramic crucible and illuminated under short wave UV light,
which causes scorpions to fluoresce (Polis 1990a). Invertebrate fragments were
removed under a binocular microscope, then dried and counted. They were
identified using a reference collection of whole specimens, body fragments and
photos. Descriptions and photos of fragments are given in Appendices 5.3 and 5.4.
Initially, an attempt was made to measure the volume and dry weight of the animal,
plant and inorganic components per faeces, but manual separation proved
incomplete. Then, a rapid, objective and repeatable index for the relative
proportions of food types in faeces using image analysis was considered (Banks
1990; Chesmore & Monkman 1994). However, poor discrimination between the
components combined with high refraction and reflection of light hampered the
technique. Therefore, a visual estimate of the volume of invertebrate as opposed to
plant material in the faeces was used comprising five categories; <5%, 25%, 50%,
75% and >95% invertebrate material. While limited, such methods are widely used
for birds (see e.g. Savory 1989). The identity of plant remains, including leaves,
flowers or seeds, was noted when possible. In addition, a reference collection of
plant epidermes from UAE was assembled, and as a pilot study slide preparations
were made from captive houbara faeces using the technique of Storr (1961).
Although epidermes and hairs were visible, it was not possible to quantify plant

138
remains because animal fragments in the diet obscured many characteristic plant
microstructures (Norton 1995).
Dietary calculations
When calculating a typical houbara diet from faecal remains, it was desirable to
include a measure of the variation of recovery rates between birds. While standard
errors of ratios provide an indication of variation (Snedecor & Cochran 1967), those
based on small samples are often skewed and may be unreliable (Krebs 1989).
Therefore, a Monte Carlo method was used to estimate a confidence interval for the
overall diet. Firstly, the number of key fragments found per prey group from the total
number of faeces (n=161) was averaged. Then, prey intake per prey group was
calculated by multiplying the average number of key fragments by the reciprocals of
the recovery rates. The recovery rates were chosen at random from the actual
values calculated for each trial bird. This process was repeated 1000 times to yield
1000 estimates of the numbers of each prey group eaten depending on variations in
recovery rates. The number of prey eaten was converted to biomass, using mean
weight per taxon, and then to dry weight, amount of fat and protein from published
data on similar taxa (Redford & Dorea 1984; Robel etal. 1995). Calorific values
were calculated assuming 5.306 kcal/g for insects and 5.480 kcal/g for arachnids in
the absence of more specific data (Robel et al. 1995) and corrected for
metabolizable energy (71.2%; Bell 1990). Results from the 1000 simulations were
ranked to identify the median, and upper and lower 95% confidence limits.
Since there was no information on the houbara’s metabolic rate. Resting Metabolic
Rate (RMR) and Active Metabolic Rate (AMR) were calculated using the following
general equations (Bennett & Harvey 1987):
In (RMR) = 0.68 In (body weight) - 0.28
In (AMR) = 0.61 In (body weight) + 1.18
Energy requirements were also calculated from the weight of food pellets consumed
by captive birds between experiments and the metabolizable energy content of the
pellets (2800 cals/g).

139
A typical diet for the houbara was estimated using the number of each food type
that must be eaten to yield daily energy needs (AMR) assuming an intake
proportional to their relative abundance in the faecal samples. As a crude estimate
of how dietary requirements may determine foraging behaviour, the distance that a
houbara must travel to obtain sufficient invertebrate food was calculated by
assuming that houbara encounter food at similar rate to pitfall traps (Chapter 4).
Pitfall capture from the winter months in Baynunah was used to indicate prey
abundance, since over 92.5% of houbara faeces were collected from this region.
5.2.3 Results
Calibration experiments with animal foods
Key fragments were selected for low variance in the recovery rate between birds
and are shown in bold in Table 5.2. Recovery rates for key fragments ranged from
0.21 ± 0.08 to 0.86 ± 0.11 indicating substantial differences between taxa, although
for some prey groups there was little difference between the recovery rate of
several fragments.
Figure 5.1 and Table 5.3 illustrate how key fragments were chosen, using group 6
as an example. Recovery rates for the 13 recognisable fragments ranged from 0.02
(sternites) to 0.51 (tibia). While tibia had the highest recovery rate they also had a
high variance. The next highest recovery rates were for femora and jaws and both
had lower variance than tibia. In this case the larger and more easily recognisable
femora were chosen in preference to the smaller jaws.
0.90 j 0.80 - 0.70 0.60 + 0.50 0.40 - 0.30 - 0.20 0.10
0.00 - 0.10
Î I - , 11 ITarsus Trochanter Elytra Scutellum Sternites Prostemum Labrum Mentum
Figure 5.1 : Mean ratio of recovery rates (filled squares) for fragments from group 6.
The length of the error bars indicate the upper and lower confidence limits.

Preygroup
Genera used in the trails
No. of trials
Head Thorax Femur Tibia Mandible(mouthparts)
Elytra Pincer (pedipalp) (only in (only in arachnids)
coleoptera)
RecoveryRate
SE RecoveryRate
SE RecoveryRate
SE Recoveryrate
SE RecoveryRate
SE Recovery SE Recovery SE Rate Rate
2 Messor 3 0.66 0.119 0.04 0.031 * * *
3 Ocladius 3 0.45 0.187 0.32 0.192 0.56 0.073 0.45 0.76 * 0.21 0.198
4 Mesotena 6 0.63 0.108 0.38 0.156 0.76 0.043 0.71 0.043 0.72 0.098 0.43 0.125
5 Tentyrina 3 0.47 0.257 0.28 0.255 0.63 0.115 0.45 0.128 0.62 0.187 0.29 0.103
6 Akis,Ocnera,Pimeiia
7 0.35 0.081 0.04 0.026 0.38 0.080 0.51 0.136 0.39 0.088 0.10 0.056
7 Anthia 4 0.33 0.125 0.22 0.052 0.81 0.094 0.80 0.098 0.86 0.109 0.22 0.169
8 Orthoptera 5 * * 0.16 0.043 0.12 0.043 0.67 0.178
10 Vachcnicius 5 * * * • 0.04 0.035 0.21 0.07
16 Erodius 3 0.43 0.147 0.04 0.076 0.76 0.126 0.63 0.005 0.72 0.141 *
Table 5.2. Mean and standard error (SE) of the recovery rate for prey remains in faeces with key fragments in bold. Empty cells indicate that the fragment was not applicable to the prey group and * indicates a recovery rate <0.01%. Fragments occasionally found are excluded, e.g. labrum, mentum, scutellum, trochanter, sternum, sternite and tarsus. Vertebrate prey (groups 12 and 13) are excluded because no fragments could be quantified.
Thorax was represented by the propodeum for group 2, and the thoracic disc for groups 3, 4, 5, 6 and 7
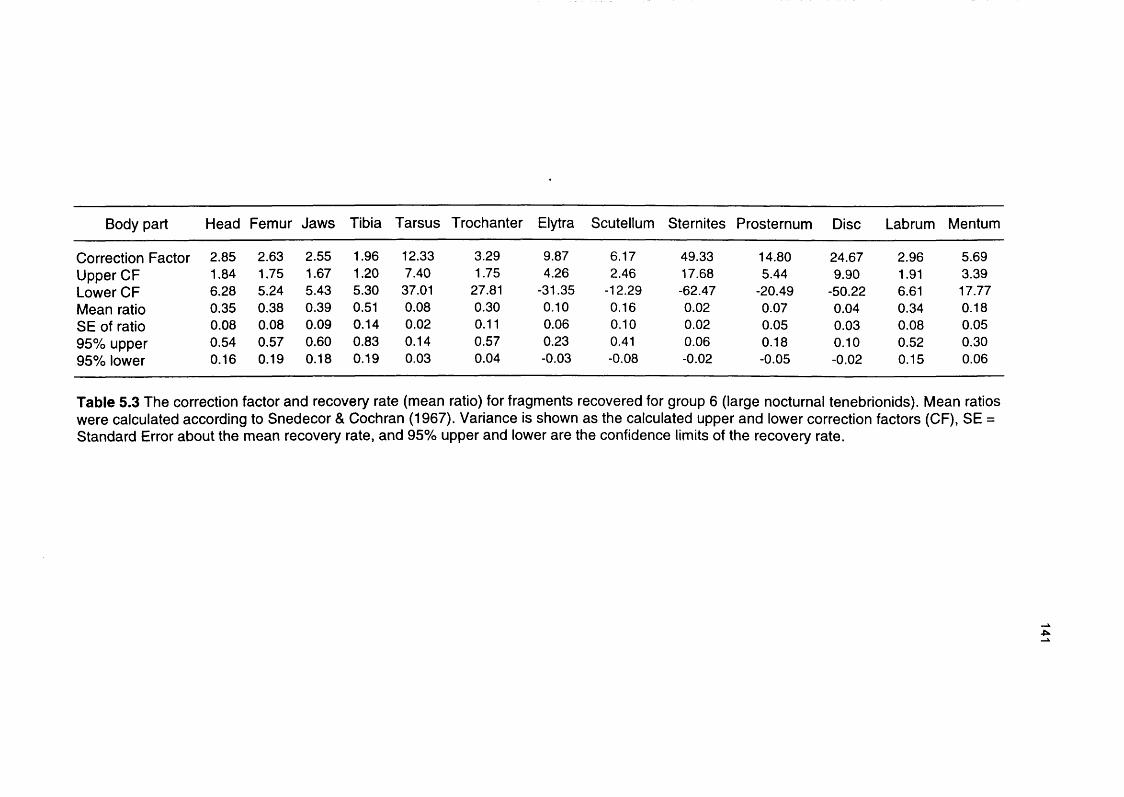
Body part Head Femur Jaws Tibia Tarsus Trochanter Elytra Scutellum Sternites Prosternum Disc Labrum Mentum
Correction Factor 2.85 2.63 2.55 1.96 12.33 3.29 9.87 6.17 49.33 14.80 24.67 2.96 5.69Upper CF 1.84 1.75 1.67 1.20 7.40 1.75 4.26 2.46 17.68 5.44 9.90 1.91 3.39Lower CF 6.28 5.24 5.43 5.30 37.01 27.81 -31.35 -12.29 -62.47 -20.49 -50.22 6.61 17.77Mean ratio 0.35 0.38 0.39 0.51 0.08 0.30 0.10 0.16 0.02 0.07 0.04 0.34 0.18SE of ratio 0.08 0.08 0.09 0.14 0.02 0.11 0.06 0.10 0.02 0.05 0.03 0.08 0.0595% upper 0.54 0.57 0.60 0.83 0.14 0.57 0.23 0.41 0.06 0.18 0.10 0.52 0.3095% lower 0.16 0.19 0.18 0.19 0.03 0.04 -0.03 -0.08 -0.02 -0.05 -0.02 0.15 0.06
Table 5.3 The correction factor and recovery rate (mean ratio) for fragments recovered for group 6 (large nocturnal tenebrionids). Mean ratios were calculated according to Snedecor & Cochran (1967). Variance is shown as the calculated upper and lower correction factors (OF), SE = Standard Error about the mean recovery rate, and 95% upper and lower are the confidence limits of the recovery rate.

142
Faecal analvsis
A total of 161 faeces were collected from 52 separate houbara tracks between 18
March 1993 and 3 September 1995 (Appendix 5.5). The mean number of faeces
per track was 3.23 (± 3.22 SD). It was assumed that the faeces originated from a
maximum of 52 birds, but the minimum is not known since birds may have left more
than one track. For comparative purposes dietary composition was calculated for
both number of faeces and tracks.
All faeces contained invertebrate material, and over 72% of samples contained a
volume of at least 50% or more invertebrate remains following digestion (Table 5.4).
About 12% of faeces contained less than 5% invertebrates, although this figure was
inflated by multiple samples from a single houbara track.
Percentage invertebrate material
<5% 25% 50% 75% >95%
Percentage of faeces 11.8% 16.2% 32.3% 36.7% 3.1%
Table 5.4 Estimated percentage of invertebrate matter of houbara faeces from Abu
Dhabi (n=161).
Dipterygium glaucum and a grass were the most frequently eaten plants, with their
leaves occurring in faeces along 24.5% and 18.9% of tracks respectively (Table
5.5). Other desert plants, including shrubs and forbs, were eaten less frequently.
The percentage frequency of prey per faeces was slightly lower than the
percentage frequency by tracks except for group 16, where a single bird inflated the
totals and reversed the trend (Table 5.6). The most frequent prey from the 52 tracks
were group 6 (94.3%), group 5 (92.5%), group 2 (60.4%) and group 4 (32%) which
are predominantly terrestrial, nocturnal invertebrates representing the
ColeopteraiTenebrionidae and Hymenoptera:Formicidae.
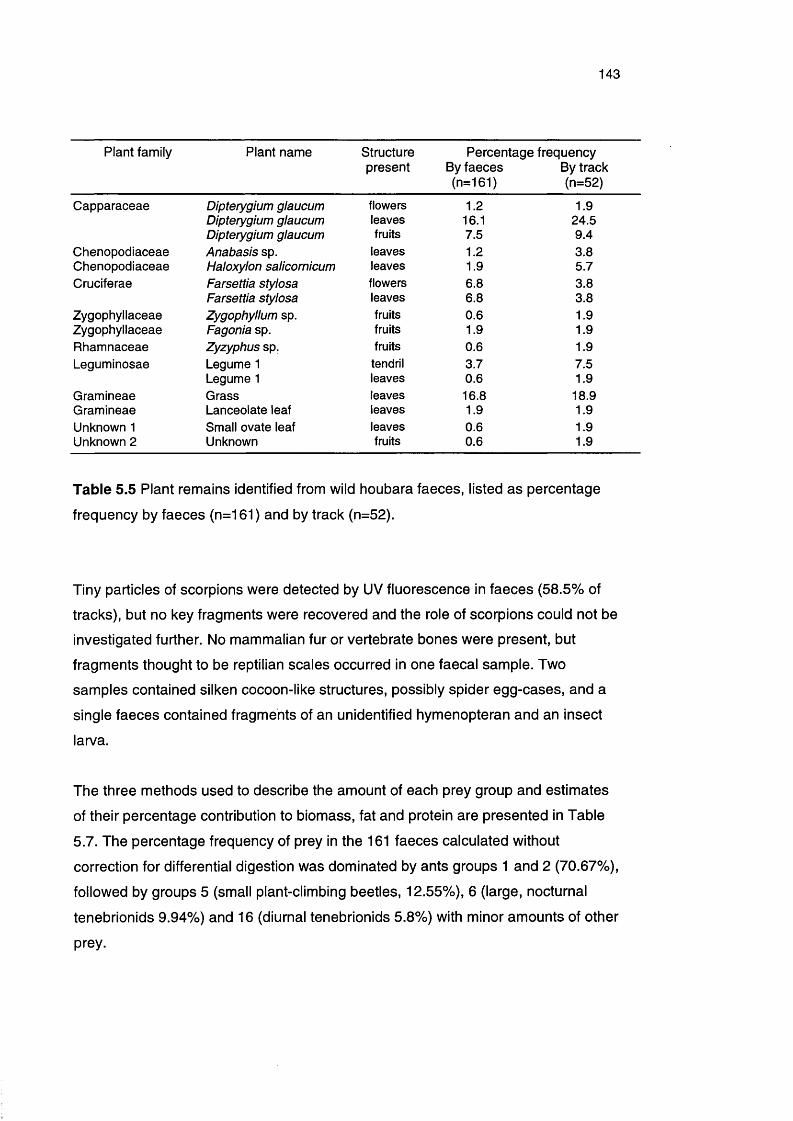
143
Plant family Plant name Structurepresent
Percentage frequency By faeces By track (n=161) (n=52)
Capparaceae Dipterygium glaucum flowers 1.2 1.9Dipterygium glaucum leaves 16.1 24.5Dipterygium glaucum fruits 7.5 9.4
Chenopodiaceae Anabasis sp. leaves 1.2 3.8Chenopodiaceae Haloxylon salicornicum leaves 1.9 5.7Cruciferae Farsettia styiosa flowers 6.8 3.8
Farsettia styiosa leaves 6.8 3.8Zygophyllaceae Zygophyllum sp. fruits 0.6 1.9Zygophyllaceae Fagonia sp. fruits 1.9 1.9Rhamnaceae Zyzyphus sp. fruits 0.6 1.9Leguminosae Legume 1 tendril 3.7 7.5
Legume 1 leaves 0.6 1.9Gramineae Grass leaves 16.8 18.9Gramineae Lanceolate leaf leaves 1.9 1.9Unknown 1 Small ovate leaf leaves 0.6 1.9Unknown 2 Unknown fruits 0.6 1.9
Table 5.5 Plant remains identified from wild houbara faeces, listed as percentage
frequency by faeces (n=161) and by track (n=52).
Tiny particles of scorpions were detected by UV fluorescence in faeces (58.5% of
tracks), but no key fragments were recovered and the role of scorpions could not be
investigated further. No mammalian fur or vertebrate bones were present, but
fragments thought to be reptilian scales occurred in one faecal sample. Two
samples contained silken cocoon-like structures, possibly spider egg-cases, and a
single faeces contained fragments of an unidentified hymenopteran and an insect
larva.
The three methods used to describe the amount of each prey group and estimates
of their percentage contribution to biomass, fat and protein are presented in Table
5.7. The percentage frequency of prey in the 161 faeces calculated without
correction for differential digestion was dominated by ants groups 1 and 2 (70.67%),
followed by groups 5 (small plant-climbing beetles, 12.55%), 6 (large, nocturnal
tenebrionids 9.94%) and 16 (diurnal tenebrionids 5.8%) with minor amounts of other
prey.
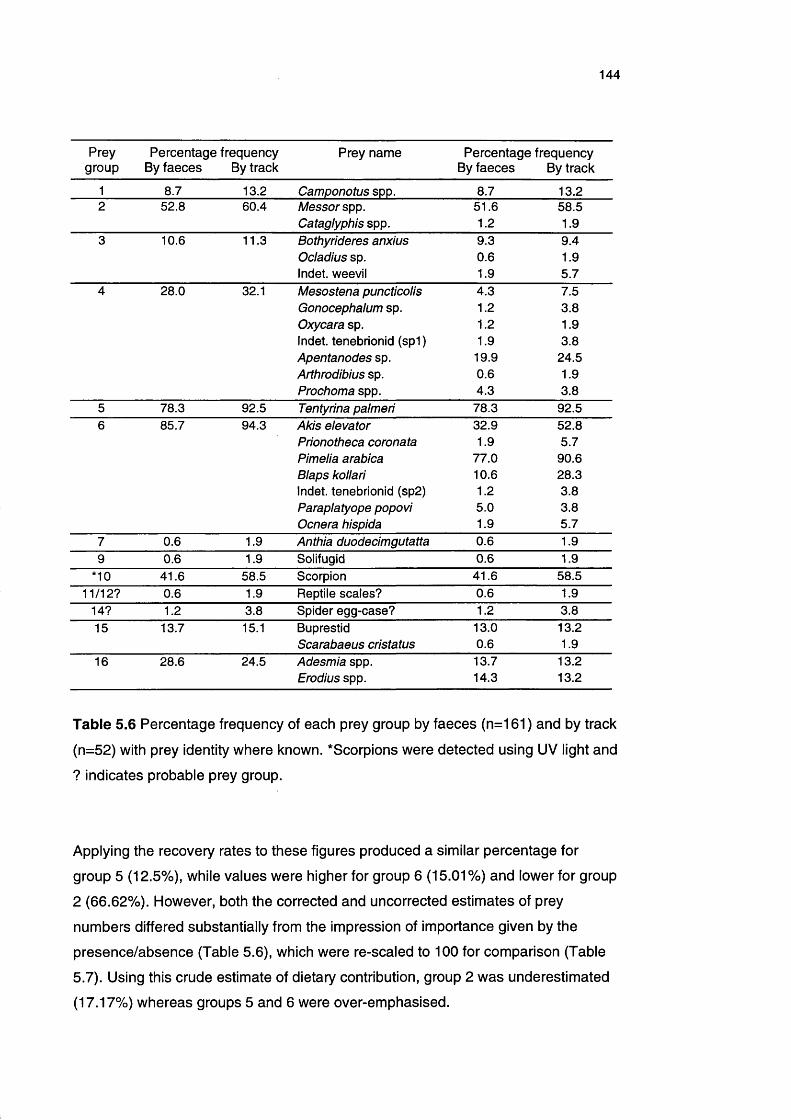
144
Preygroup
Percentage frequency By faeces By track
Prey name Percentage frequency By faeces By track
1 8.7 13.2 Camponotus spp. 8.7 13.22 52.8 60.4 Messor spp. 51.6 58.5
Cataglyphis spp. 1.2 1.93 10.6 11.3 Bothyrideres anxius 9.3 9.4
Ocladius sp. 0.6 1.9Indet. weevil 1.9 5.7
4 28.0 32.1 Mesostena puncticoiis 4.3 7.5Gonocephalum sp. 1.2 3.8Oxycara sp. 1.2 1.9Indet. tenebrionid (sp1) 1.9 3.8Apentanodes sp. 19.9 24.5Arthrodibius sp. 0.6 1.9Prochoma spp. 4.3 3.8
5 78.3 92.5 Tentyrina paimeri 78.3 92.56 85.7 94.3 Akis eievator 32.9 52.8
Prionotheca coronata 1.9 5.7Pimelia arabica 77.0 90.6Blaps kollari 10.6 28.3Indet. tenebrionid (sp2) 1.2 3.8Paraplatyope popovi 5.0 3.8Ocnera hisplda 1.9 5.7
7 0.6 1.9 Anthia duodecimgutatta 0.6 1.99 0.6 1.9 Solifugid 0.6 1.9
*10 41.6 58.5 Scorpion 41.6 58.511/12? 0.6 1.9 Reptile scales? 0.6 1.9
14? 1.2 3.8 Spider egg-case? 1.2 3.815 13.7 15.1 Buprestid 13.0 13.2
Scarabaeus cristatus 0.6 1.916 28.6 24.5 Adesmia spp. 13.7 13.2
Erodius spp. 14.3 13.2
Table 5.6 Percentage frequency of each prey group by faeces (n=161) and by track
(n=52) with prey identity where known. "Scorpions were detected using UV light and
? indicates probable prey group.
Applying the recovery rates to these figures produced a similar percentage for
group 5 (12.5%), while values were higher for group 6 (15.01%) and lower for group
2 (66.62%). However, both the corrected and uncorrected estimates of prey
numbers differed substantially from the impression of importance given by the
presence/absence (Table 5.6), which were re-scaled to 100 for comparison (Table
5.7). Using this crude estimate of dietary contribution, group 2 was underestimated
(17.17%) whereas groups 5 and 6 were over-emphasised.

J• J
Preygroup
Relative importance in diet based on
presence/absence in faeces
Percentage frequency based on key
fragments
Percentage frequency based on key
fragments and their recovery rates.
Percentage contribution to biomass
Percentage contribution to fat
Percentage contribution to protein
1 2.83 0.23 0.22 0.04 0.02 0.04
2 17.17 70.44 66.62 3.23 1.71 3.11
3 3.47 0.13 0.15 0.12 0.03 0.11
4 9.10 0.84 0.69 0.65 0.66 0.65
5 25.46 12.55 12.50 8.91 9.06 8.91
6 27.86 9.94 15.01 73.61 74.88 73.67
7 0.19 0.02 0.01 0.05 0.05 0.05
9 0.19 0.02 0.01 0.01 <0.01 0.01
15 4.45 0.03 0.03 0.07 0.04 0.12
16 9.27 5.80 4.76 13.54 13.54 13.32
Table 5.7 Comparison of three methods of faecal analysis, with estimates of the percentage contribution to biomass, fat and protein using data from Robel etal. (1995). No remains of groups 8 and 10 were recovered from the faeces, although scorpions (group 10) were detected by UV light.
cn

146
By contrast, the contribution of prey groups to the biomass, fat and protein intake
gives an alternative view. Group 2 (ants) contributed only 3.23% of the biomass,
while groups 6 (large tenebrionids) and 16 (diurnal tenebrionids) made up 73.61%
and 13.54% of the invertebrate biomass respectively. Fat and protein gave similar
results, probably because few taxa-specific values were available to alter the
balance between groups.
Using the calibrated figures for number of prey consumed and published energetic
values. Table 5.8 gives Monte Carlo estimates of the calorific value of the
invertebrate food eaten that resulted in an average faecal sample. The median
estimate was 24.6 Kcal but note that the data were bimodal with upper and lower
peaks at about 24.2 Kcal and 15.0 Kcal respectively.
Kcal
Median 24.640
Lower 95% confidence limit 13.119
Upper 95% confidence limit 28.071
Upper peak c. 24.208
Lower peak c. 14.952
Table 5.8. Calorific value of invertebrate diet corrected for metabolizable energy
content per average faecal sample for wild houbara over-wintering in Abu Dhabi.
Lower and upper 95% confidence limits were obtained from 1000 estimates using a
Monte Carlo method. Note that the data are bimodal hence the upper and lower
peaks occurred more frequently than the median value.
Applying Bennett & Harvey's (1987) formula, the daily energy needs for the RMR of
experimental birds was 99.1 Kcal/day, which compares favourably with 104.7 Kcal,
based on their mean daily consumption of food pellets (37.4 ±13.4g). The calculated
AMR for houbara was 258.2 Kcal/day and based on the median energy value of
prey consumed per faeces (Table 5.8), an average houbara would be expected to
produce 10.48 (9.20-19.68) faeces/day to meet its AMR. Assuming prey
composition in proportion to their occurrence in the 161 faeces the figures suggest
that an average houbara bustard can meet its daily AMR by consuming a mixture of

147
about 671 typical desert invertebrates. This excludes the contribution of plant foods
which is more difficult to estimate. However, a calculation based on the mean visual
estimate of the volume of plant and mean weight of faeces suggests that on
average plant foods provide 1.41 to 3.06 Kcals per faeces (using metabolizable
energy values from Savory 1989). This increases the total energy per bird per day
by between 5.4 and 11 %, but does not take into account the metabolic costs of
digestion or differences in the plant structures consumed.
The percentage contribution of each prey group to the total winter pitfall trap catch
(78 lines of 20 traps spaced 20m apart, excluding taxa that do not appear in the
faeces) is shown in Table 5.9 column three. Comparison with the calibrated
percentage frequency in faeces showed good rank agreement (rg =0.93, p=0.001,
n=7) although the typical diet contained more prey from group 2 and fewer from
group 6 than expected by chance encounters.
Preygroup
Typical daily diet (number of prey)
Percentage frequency of prey in pitfall traps
Estimated foraging distance (km)
2 445 51.94 123 1 0.18 84 8 4.14 35 84 6.84 176 101 33.84 47 <1 0.90 <0.116 20 2.16 36
Table 5.9. On the basis of the recovery rates the typical daily diet has been
calculated as the number of each prey group required to meet energetic needs.
Prey availability was assessed by pitfall trapping and the foraging distance is how
far the bird would need to search to find this amount of prey, assuming it
encounters prey at random.
Finally, the average length of pitfall trap line needed to catch the number of each
prey group required by the bird per day was estimated (Table 5.9). The highest
value (36km) provides an upper estimate of the daily foraging distance assuming
random prey encounters, although houbara are likely to encounter other prey along
the same route. Figure 5.2 shows the seasonal variation in prey abundance (group

148
2, group 6 and total prey) and the relative effort needed to catch that amount of
prey. The latter is scaled to one for the average distance values in Table 5.9. Note
that in February, when prey were least abundant, the effort needed to catch
sufficient invertebrates would be expected to rise by up to 11 times the average
value.
12.0
35(AQ.2
10.0
raQ.° 20 6.0
VQ.&
£1E3Z
4.0
2.0
0.0Oct Nov Dec Jan Feb Mar
Group 2— O- - - Group 6- - - A- • • Total — ■— Bfort
Figure 5.2. Available invertebrate prey (groups 2 and 6 and total invertebrates) and
relative effort required to catch them during the winter months in Abu Dhabi.
5.2.4 Discussion
The relative merits of different dietarv calculations and indices
The percentage frequency of faeces containing a particular prey item based on
presence/absence is the simplest way to describe diets, but gives no indication of
the amount of each food consumed. Counting remains of prey in faeces gives a
measure of the relative abundance of prey in the diet, especially when corrected for
differential digestion. Corrected faecal analysis for houbara in UAE indicated that
they mainly consumed group 2, especially ants of the genus Messor, group 6, large,
ground-dwelling insects such as Pimelia spp. (Tenebrionidae); and group 5, small
plant-climbing insects represented by T. pa/mer/(Tenebrionidae).

149
Uncorrected faecal analysis underestimated group 2 and overestimated groups 6 and
16.
However, diet descriptions based on number do not take into account differences in
size of prey. While over 66% of invertebrates eaten were ants, they contributed only
3.2% biomass, 1.7% fat and 3.1% protein consumed (Table 5.7). By contrast, the
large tenebrionids (group 6) contributed about 73.6% invertebrate biomass. Indeed,
the Tenebrionidae as a whole (in groups 4, 5, 6 and 16) made up 96.7% biomass,
98% fat and 96.5% protein of the invertebrate component of the houbara diet.
The calculated value for RMR was close to the energy value of pellets consumed by
experimental birds, giving confidence to the use of the general formula of Bennett &
Harvey (1987). From AMR an average houbara must consume an average of 671
invertebrates and would produce about ten faeces per day. The estimated number of
prey eaten per day assumes a spread across prey groups in proportion to their
occurrence in the faeces which correlated well with invertebrate relative abundance
along lines of pitfall traps. Apart from an estimated foraging distance of 36km for
group 16, the foraging distances in Table 5.9 are within the daily range for houbara
movements in UAE (Launay etal. 1997; Osborne etal. 1997a & b). The assumption
that a foraging houbara captures prey at the same rate as a passive pitfall trap is
probably an underestimate, although the similarity between the prey frequency in the
faeces and the traps suggest that these birds are generally non-selective. The
exceptions are an under-representation of large tenebrionids, possible due to the
chemical defences of certain species such as Blaps kollari (Walker & Pittaway 1987),
and an over-consumption of ants. This generalist foraging strategy is to be expected
of a large predator in a harsh environment with low prey availability.
Hutto (1990) notes that many studies on food availability are inappropriate because
birds do not perceive prey in the same manner as humans. Houbara are strong flyers
but are generally cursorial and walk great distances unless disturbed (Launay et al.
1997). Therefore a pitfall transect is a fair approximation of how they encounter food
since the probability of capturing invertebrates increases with distance for both
houbara and transects. However, while houbara can choose to remain and feed in an
area where there is a locally abundant prey, such as an ant's nest, pitfall traps may
fail to sample clumped or patchily distributed prey.

150
RMR and AMR have not been measured in bustards, and the metabolic rates of
some avian families deviate from the equation based on body weight in Bennett &
Harvey (1987). This is most noticeable in small passerines, whose metabolic rate is
relatively high for their body weight. The calculated metabolic rates for houbara are
of the same order of magnitude as rates measured in birds with similar body
weights (Bennett & Harvey 1987). Avian AMR also rises in response to the
increased energy needs associated with changes in activity and body condition,
such as growing, moulting, breeding and migration, and during particularly harsh
environmental conditions (see Carey 1996). These factors should be considered if
direct measurements of the houbara’s metabolism are undertaken.
Although the calculations suggest that on an average day a houbara can fulfil its
energy requirements by feeding on invertebrates alone, this is unlikely to be true
throughout the winter. Prey abundance and activity, estimated from pitfall capture,
show marked seasonal variation in Abu Dhabi and are at their lowest in January and
February (see Chapter 4). At this time, the estimated foraging distance for a
houbara to meet daily AMR was about 11 times the average winter distance (Figure
5.2). Translating this figure into the number of birds that an area can support
requires data on renewal rates of prey and prey densities which are unknown.
However, the importance of large tenebrionids as a source of biomass suggests
that their abundance could be a key to the presence of houbara in UAE.
The crude estimate of the energy contribution of plants suggests that on average
they provide a further 5.4 to 11% energy during the winter in UAE. However, exact
nutrient and energy content vary according to plant structure. Herbage is least
nutritious, but new shoots, bulbs, tubers, seedlings and seeds contain up to one-
third of the metabolizable energy of animal prey (Karasov 1990). Therefore birds
must consume larger volumes of plants to get the same energy value from animal
prey. This lower nutrient content is compounded by increased costs of digestion,
including the breakdown of cellulose and the detoxification of secondary plant
compounds (Karasov 1990). Foods that are difficult to digest will also contribute a
higher volume to faeces and gizzard contents, and without calibration the initial
amount of plant material consumed cannot be estimated.

151
Identification of key fragments
Various prey fragments were recovered from faeces and the inherently strong,
chitinous and sclerotized structures survived digestion well. The femur was the most
frequently chosen key fragment, and could usually be assigned to a prey group. For
larger beetles it was better to count femoral joints rather than fragments of femur.
Several authors suggest using mandibles for estimating intake of prey especially
beetles (Moreby 1988; Ralph et al. 1985). For the houbara, mandibles were
applicable for large active prey of groups 7 and 8, while for smaller prey (social
insects like ants) head capsules were better because mandibles tended to remain
inside the head, reducing their recovery rate. Collins (1984, 1993) calculated
houbara prey intake on Fuerteventura on the basis of invertebrate fragments in
faeces, but other than ants these differed from my key fragments. Without trials on
captive birds it is difficult to judge which fragments are reliable and as fragment
recovery rates tend to zero, the differences between corrected and uncorrected
faecal analysis become more apparent.
Problems remain in estimating the role of larger more digestible prey, including
arachnids and vertebrates, because few key fragments could be identified.
Solifugae and Araneae (groups 9 and 14) were not available in sufficient numbers
for the trials, but their chelicerae were used by Gubin & Mukhina (in prep.) and
Moreby (1988) to indicate consumption of solifugids and spiders respectively.
Solifugid presence was indicated by single chelicera in single faeces and gizzards
from UAE, and such prey are unlikely to be active during winter months (Cloudsley
Thompson 1987).
During the calibration trials, pedipalps were identified as key fragments for
scorpions but none were recovered in faeces or gizzards from wild birds. A tail
segment and UV fluorescence were detected in faeces, but since no key fragments
were found scorpions were excluded from the energy calculations. The intensity of
the UV fluorescence might indicate how recently scorpions were consumed. In the
five scorpion trials, fluorescence from a single meal lasted for 4 days (± 0.45 SO),
and the intensity decreased with time. The level of UV fluorescence was less
intense in the wild bird faeces.

152
No key fragments were identified for vertebrates (groups 12 and 13), although fur
occurred in faeces from houbara fed on white mice (see Appendix 5.3). A tooth was
recovered from faeces of captive houbara fed on a mixed diet including mice (S. W.
Warren pers. comm.) and Gubin & Mukhina (in prep.) found a mouse tail and
vertebrae in houbara faeces. However, Nazarov (1992) reported that the faeces of
houbara fed on pigeons and sparrows contained no traces of bone or feather.
Perhaps only larger or older vertebrates resist digestion, which combined with low
and inconsistent recovery rates makes them difficult to detect. No mammalian fur
was seen in wild bird faeces or gizzards from UAE, although one faeces contained
two fragments resembling reptile scales. However, there is little evidence for the
frequent consumption of large prey by houbara in UAE during the winter. How to
account for prey which can be detected through means other than key fragments
(e.g. fluorescence, hair or scales) remains a challenge in quantifying the diet.
Weaker fragments, like beetle elytra, were greatly crushed, and their recovery rates
were either too low or too variable for calibration. However, they helped to
distinguish between structurally similar prey which were grouped separately
because of behavioural differences. For example, T. palmeri (group 5) have slightly
longer femora than M.puncticollis (group 4) and their heads, elytra and thoracic
discs are quite distinctive (see Appendices 5.3 and 5.4). Unlike most ground-
dwelling tenebrionids, T. palmeri has a habit of climbing Haloxylon salicornicum
bushes making it easy to find in the field. This may be related to pheromone
production, since beetles rapidly copulate when collected (personal observation). In
the laboratory T. palmeri ate H. salicornicum, and showed signs of being more
phytophagous than other tenebrionids which are generally detritivorous or
omnivorous.

153
5.3 Analysis of houbara gizzards from Abu Dhabi
5.3.1 Introduction
Gizzard contents have been used to describe the diet of birds including the houbara
(see Mirza 1971; Gavrin 1962), but the killing of endangered species for scientific
study is not an acceptable practice. However, the remains of six birds either found
dead in the wild or hunted by Arab falconers were examined. This small sample
may not be totally representative of the normal houbara diet, but is included here for
comparison with gizzard studies from other regions and with the UAE faecal
analysis.
5.3.2 Methods
Fresh or defrosted gizzard contents were placed in a set of analytical sieves (2 mm
and 210 pm), and irrigated with running water. Most invertebrate material floated to
the surface and was removed using forceps, and the rest was filtered off. All food
remains were dried, identified and counted. In most cases digestion was not
complete, but where prey had been crushed by the gizzard the uncalibrated number
of key fragments was used to assess prey intake since the extent of digestion was
unknown. Plant material was identified and the score system for the proportion of
invertebrate material in faeces was applied to gizzard samples.
5.3.3 Results
A list of plants and animals found in the gizzards and faeces is shown in Table 5.10.
In addition, the contribution of animal prey by group was estimated from the number
of prey and their metabolizable energy content (cals/g dry wt) and compared with
faecal estimates (see Table 5.11). Note that the extent of digestion of prey in the
gizzards was unknown.
The most numerous prey in the gizzards were ants (group 2, 48.9%), diurnal
tenebrionids (group 16, 29.3%) and small nocturnal tenebrionids (group 4,17.4%).
The amount of plant material in the gizzards varied (Table 5.11) and seeds, leaves
and shoots of D. glaucum were the most frequent plant remains (Table 5.12).

154
Animal Family Species name (if known) Prey group Gizzards FaecesFormicidae Camponotus spp. 1 X X
Messor spp. 2 X X
Cataglyphis spp. 2 X X
Curculionidae Indeterminate Curculionidae 3 X X
Ocladius sp. 3 X X
Bothyrideres anxius 3 X X
Tenebrionidae Mesostena puncticolis 4 X X
Gonocephalum sp. 4 X
Oxycara sp. 4 X
Indeterminate Tenebrionidae sp1 4 X
Apentanodes sp. 4 X X
Arthrodibius sp. 4 X
Prochoma spp. 4 X
Zophosis sp. 4 X
Tentyrina palmeri 5 X X
Akis elevator 6 X X
Prionotheca coronata 6 X
indeterminate Tenebrionidae sp2 6 X
Slaps kollarl 6 X X
Pimelia arabica 6 X X
Paraplatyope popovi 6 X X
Ocnera hisplda 6 X
Adesmia spp. 16 X X
Erodlus spp. 16 X X
Carabidae Scarites guineensis 7 X
Anthia duodeclmguttata 7 X X
Orthoptera indeterminate Acrididae 8 X
Solifugidae Galeodes sp. 9 X X
Buprestidae Indeterminate Buprestidae sp1 15 X X
Indeterminate Buprestidae sp2 15 X X
Indeterminate Buprestidae sp3 15 X
Hymenoptera (winged) Indeterminate Hymenoptera 15 X X
Scarabaeoidea Scarabaeus christatus 16 X
Plant Family Species name (if known) Plant partCapparaceae Dipterygium glaucum seeds, stems &
leavesX X
Rhamnaceae Zizyphus sp. fruits & seeds X X
Leguminosae Unknown Leguminosae leaves & tendrils X
Chenopodiaceae Haloxylon salicomlcum leaves X
Anabasis sp.? leaves X
Cruciferae Farsetia stylosa flowers & leaves X
Compositae Unknown Compositae flowers X
Zygophyllaceae Zygophyiium sp.
Fagonia spp. fruits & seeds X
Tribulus sp. leaves X
Unknown Unknown dicotyledon seedlings X
Unknown dicotyledon leaves & fruits X
Gramineae Grass 1
Grass 2 leaves X
Table 5.10 List of animal and plant foods from houbara faeces and gizzards. x =
present. Excludes probable prey from Table 5.6.
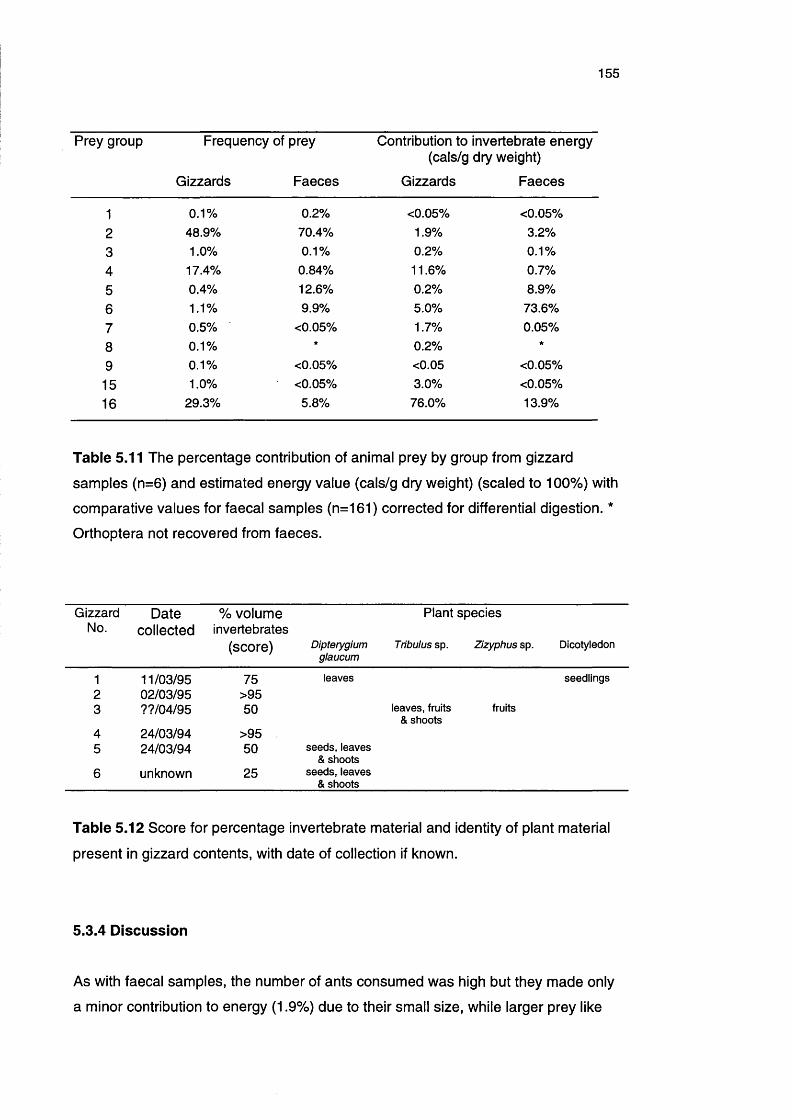
155
Prey group Frequency of prey Contribution to invertebrate energy (cals/g dry weight)
Gizzards Faeces Gizzards Faeces
1 0.1% 0.2% <0.05% <0.05%2 48.9% 70.4% 1.9% 3.2%3 1.0% 0.1% 0.2% 0.1%4 17.4% 0.84% 11.6% 0.7%5 0.4% 12.6% 0.2% 8.9%6 1.1% 9.9% 5.0% 73.6%7 0.5% <0.05% 1.7% 0.05%8 0.1% * 0.2% *
9 0.1% <0.05% <0.05 <0.05%15 1.0% <0.05% 3.0% <0.05%16 29.3% 5.8% 76.0% 13.9%
Table 5.11 The percentage contribution of animal prey by group from gizzard
samples (n=6) and estimated energy value (cals/g dry weight) (scaled to 100%) with
comparative values for faecal samples (n=161) corrected for differential digestion. *
Orthoptera not recovered from faeces.
Gizzard Date % volume Plant speciesNo. collected invertebrates
(score) Dipterygium Tribulus sp. Zizyphus sp. Dicotyledongiaucum
1 11/03/95 75 leaves seedlings
2 02/03/95 >953 ??/04/95 50 leaves, fruits
& shootsfruits
4 24/03/94 >955 24/03/94 50 seeds, leaves
& shoots6 unknown 25 seeds, leaves
& shoots
Table 5.12 Score for percentage invertebrate material and identity of plant material
present in gizzard contents, with date of collection if known.
5.3.4 Discussion
As with faecal samples, the number of ants consumed was high but they made only
a minor contribution to energy (1.9%) due to their small size, while larger prey like

156
tenebrionid beetles contributed most energy (group 16, 76%, group 4, 11.6% and
group 6, 5.0%).
Gizzard contents suggest that there may be differences between the houbara diet in
different parts of the range. In Pakistan, between November and January, 52
gizzards contained a mixture of plant and invertebrate material, but plant remains
made up the highest volume of samples (Fox 1988). The six gizzards from UAE had
smaller volumes of plant material, and only one had a higher volume of plant than
animal material (Table 5.12).
Boobyer (1989) separated out and measured the volume and dry weight of the plant
and animal components in 15 gizzards of the Karoo korhaan {Eupodotis vigorsii:
Family Otididae). On the basis of volume he suggests that plant foods were more
important than animal foods for this bustard, but notes that social insects,
particularly Isoptera, were underestimated because of their rapid digestion
compared with vegetative structures. This highlights the difficulty of studying diet
once digestion has started when information on the extent and speed of digestion of
different foods are unknown.
The gizzards in UAE came from dead or hunted houbara whose diet may differ from
that of healthy birds, and any bias will be amplified by the small number of samples.
One gizzard contained about 230 Erodius sp. and little else, and inflated the mean
for diurnal beetles (group 16). It also had gut parasites; 349 Centrorhynchus lancea
(Acanthocephala) and 2 Harteria rotunda (Nematoda) (Jones etal. 1996).
Comparative data on the effect of parasite burden on houbara are lacking but the
body condition of the bird suggested it was in good health. Erodius sp. also
occurred in 14.3% of faeces, but this contrasts with the trials on captive houbara
where Erodius sp. was not always readily consumed (Appendix 5.1). Five gizzards
were collected in March or April and Erodius spp. were most abundant in pitfall traps
between February and June (38 records, representing 108 beetles, see Chapter 4).
The collection date of the sixth gizzard is unknown, but it did not contain Erodius
spp.

157
5.4 General discussion of the diet
5.4.1 Description of houbara diet in Abu Dhabi
A wide variety of animal prey were identified and quantified from faecal and gizzard
contents suggesting that houbara are probably non-selective, simply consuming
locally available prey. Data on plant consumption were limited and there was little
evidence of vertebrate prey, but neither appeared to be major contributors to
houbara energy needs in UAE. Most prey were nocturnal, confirming behavioural
studies of houbara which indicate nocturnal foraging (Anegay 1994; Combreau &
Launay 1997), although large diurnal tenebrionids, such as Erodius spp. and
Adesmia spp. (group 16), were also consumed. However, classing tenebrionids by
their daily activity pattern may not strictly reflect their field behaviour because some
species show variations in seasonal activity, probably in response to temperature
(Holm & Edney 1973). Ward (1991) thought that some tenebrionids increase their
nocturnal activity in response to vertebrate predation pressure and Ayal & MerkI
(1994) reported smaller pitfall catches of tenebrionids following white stork
predation, although this is probably both a behavioural and a numerical response.
Most of the plant and animal remains found in houbara faeces and gizzards have
previously been noted as prey, but a few new coleopteran prey were recorded:
Ocladius sp. (Curculionidae), T. palmeri, Gonocephalum spp., Apentatonodes spp.,
Arthrodibius spp., Prochoma spp. (all Tenebrionidae) and Anthia duodeclmguttata
(Carabidae).
Several houbara faeces and gizzards were dominated by single prey, such as
T.palmeri, Messor spp. and Eroof/us spp., and other authors have noted similar
results (Collins 1984; Goriup & Norton 1992). In Abu Dhabi, houbara may develop a
search image (Tinbergen 1960) for T. palmeri, since these black beetles are highly
conspicuous when climbing vegetation and despite their resemblance to M.
puncticollis, outnumber them by about 41:1 (n=161) in faecal samples. This
contrasts with results from pitfall trapping where M. puncticollis and T. palmeri
frequently occur, and contributed 3.6% and 0.35% to trap catches respectively (see
Chapter 4). However, M. puncticollis does not climb vegetation and is therefore
more likely to be caught in pitfall traps than T. palmeri.

158
Fewer houbara visited Abu Dhabi between September 1992 and March 1996 in
comparison with previous years (Osborne 1991,1992), restricting the number of
faeces collected. A study of houbara in an area with higher densities and/or resident
birds would facilitate seasonal comparisons and sample collection. However, birds
should be marked so that analytical techniques such as compositional analysis
(Aebisher etal. 1993) can be applied to account for repeated measures from
individuals. It may also be possible to measure hormone levels in faeces of
unknown birds and to investigate differences between sexes.
5.4.2 Comparison with other studies on houbara diet
The frequency of prey in faeces and gizzards can be compared with uncalibrated
studies of houbara diet. Collins (1984,1993) found that weevils, tenebrionids and
ants, including Messor maurus, were the most numerous prey in the faeces of
C.u.fuertaventurae, with ants being eaten at times when tenebrionids were not
available. However, his data for weevils were biased because most faeces
originated from two birds (Collins 1984, 1993). In UAE houbara ate similar ants,
probably Messor ebeninus, but weevils were rarely eaten. The latter contributed
only 0.06% to pitfall trap catches (see Chapter 4), and hand searching for prey in
the calibration trials suggests that weevils have a clumped distribution in UAE.
Most dietary descriptions for houbara emphasise large tenebrionids (see Chapter 2)
but in UAE smaller beetles were also important, especially T. palmeri. This suggests
either a difference in the local availability of prey or that smaller species were
overlooked in the past. In Pakistan, Fox (1988) reported large nocturnal and diurnal
tenebrionids of similar genera to UAE including Adesmia spp. and Pimelia spp. in
20% of houbara gizzards. Gubin & Mukhina (in prep.) found few Scarabaeus spp.
beetles in over 1000 houbara faeces from Kazakhstan, and while the dung beetle
Scarabaeus cristatus is common in UAE (Chapter 1 & 4), only one fragment was
found in 161 faeces. Perhaps these beetles are distasteful or their spines make
them difficult to eat. Other studies indicate that Orthoptera are important prey
(Dement’ev & Gladkov 1951), but only one occurred in a gizzard from UAE, where
they are scarce because of the low density of vegetation.
In Kazakhstan, Gubin & Mukhina (in prep) suggest that seasonal changes in plant
phenology are followed by invertebrate and vertebrate population changes (see

159
Chapter 2), and are reflected in the remains of food in houbara faeces. These
breeding birds appear to be mainly opportunistic, but Gubin & Mukhina’s (in prep)
evidence is based on field observation with little analysis, and the frequency of prey
in faeces was biased by multiple samples collected from the display sites of
individual houbara.
Van Heezik & Seddon, (in press) found that the quantity of vegetation rather than
invertebrate availability influenced houbara habitat use in Harrat al-Harrah, Saudi
Arabia. Vegetation density was higher in this stony desert, with a maximum cover of
25% compared with 10% in Abu Dhabi (Roshier etal. 1996). In Saudi, houbara
presence in the dry season was correlated with the plant Capparis spinosa and the
birds were said to feed on its nutritious fruits. This plant does not occur in the sandy
deserts of Abu Dhabi, suggesting a difference in habitat and/or available food.
Combreau & Smith (1997) mention that released captive-bred houbara prefer well-
vegetated areas outside their normal breeding range, and that the availability of
green shoots and flowers influences their choice of habitat. In addition, the density
of tenebrionids and spiders as measured by pitfall traps, was also higher in
houbara-preferred habitats than areas selected at random. Desert tenebrionids are
generally associated with areas of denser vegetation (Ahearn 1971 ; Ayal & MerkI
1994), and phytophagous invertebrates would also be expected to be associated
with their food plants.
Some foods were hard to detect in faeces and could not be quantified, e.g.
arachnids, vertebrates and plants. Pitfall trapping in Abu Dhabi suggests that
arachnids are rarer than tenebrionids and ants (see Chapter 4). In Kazakhstan,
Gubin & Mukhina (in prep.) reported an average volume of 25.8% solifugid remains
in houbara faeces (n=108) in June. Solifugids are probably eaten only when they
are abundant in the summer, and they show little activity during the cooler winter.
5.4.3 Measurement of the relative proportions of food types
Absolute quantification for comparison between plant and animal foods for
omnivores such as the houbara remains problematic. Future studies should
integrate the quantification of animal and plant material. The dietary composition of
herbivorous birds can be quantified by microscopic examination of plant epidermal
remains in faeces (e.g. Summers etal. 1993). However, there are difficulties

160
associated with calibrating the amount of plant material eaten by birds, such as the
accuracy of measuring the amount of growing vegetation consumed and of
selective feeding on leaves, seeds or fruit, rather than whole plants (Owen 1975).
5.4.4 Nutritional and energetic implications
Many past descriptions of bustard diets concentrated on foods that were easy to
identify (see Chapter 2). The calibration of faecal remains by key fragments gave an
objective estimate of dietary composition, and facilitated calculations of the energy
contribution of foods. The calibrations for differential digestion could be applied
across the range of the houbara to investigate regional and seasonal differences,
although local food groups may need to be tested if they differ from the prey in
Table 5.1. These calibrations may even be used as a model for rarer bustards,
which cannot be studied in captivity. Better faecal analysis provides a non
destructive way to gather data on dietary needs of wild birds which, combined with
energetic studies such as the doubly-labelled water technique (Nagy 1983), would
improve our understanding of factors limiting threatened species.
The accuracy of calculations could be improved by using the nutritional value of the
local invertebrates rather than published data. However, Bedford & Dorea (1984)
found little variation in the nutritional quality of different species of ants and termites,
but stressed that the amount of nitrogen (used as a proxy value for protein) per
sample is misleading if it includes indigestible chitin. Rebel etal. (1995) found that
the energy values for similar taxa from different sites varied, probably because of
differences in species composition and life stages, and concluded that field studies
of diet must be more detailed if energy requirements are to be fully understood.
There may also be differences in the micro-nutrients, such as vitamins and
minerals, between taxa which could be especially important for breeding birds and
young chicks. There is further scope for developing work on captive birds, for
instance, in carrying out choice experiments and looking at differences between
individuals by age, sex or breeding condition. A key question might be to test
whether birds offered both invertebrates and vegetation exhibit preferences.
This study focused on the invertebrate part of diet and the energy contribution of
plant foods appeared to be low. However, while some plant parts are more
nutritious other factors also influence the selection of food by animals, especially the

161
presence of toxins, repellents and digestlblllty-reducing substances (Crawley 1983).
For example, food choice In seven species of sub-arctic ptarmigan, grouse and
capercaillie bore no relation to energy or nutrition but was negatively correlated with
the amount terpenes and phenolic resins (Bryant & Kuropat 1980). These properties
should also be considered when assessing the suitability of particular plants for
houbara. Grazing pressure from livestock In UAE, and In other parts of the range
affect plant community structure, and overgrazing reduces species richness and
Increases the proportion of grazing-adapted plants, which usually contain high
volumes of secondary plant compounds.

162
C h a p te r 6
GENERAL DISCUSSION ON THE ROLE OF INVERTEBRATES IN THE DIET OF THE
HOUBARA BUSTARD
Synopsis of Chapter 6
This chapter reviews the main findings of the thesis and the extent to which they
have addressed the principal objectives:
1. To assess the seasonal abundance, distribution and diversity of invertebrates in
Abu Dhabi.
2. To develop a calibrated method of faecal analysis for examining houbara diet.
3. To use the calibration technique on faeces from wild houbara in UAE and to
assess the role of invertebrates in the diet.
4. To establish monitoring techniques for the most important invertebrates.
The relevance of the results is discussed with reference to: the ecology of arid zone
invertebrates; desert food webs; the study of houbara and other avian diets;
conservation of houbara and wildlife in UAE; and implications for houbara captive
breeding programmes.

163
6.1 The ecology of arid-zone invertebrates
Seasonal abundance of invertebrates
Pitfall and light traps captured 143,397 invertebrates during two years of monitoring
in Abu Dhabi (Sections 4.1 and 4.3). Invertebrate abundance showed significant
spatial and temporal variation, with fewer individuals caught in the winter than
summer months. Invertebrate activity/abundance as measured by the traps was
positively correlated with temperature and there appeared to be an effect of rainfall
(see Section 4.1 ; Tigar & Osborne 1997). Such patterns of abundance have been
noticed in other deserts, reflecting the abiotic and biotic factors which influence their
fauna (Hinds & Rickard 1973; Thomas 1979; Sheldon & Rogers 1984).
Invertebrates survive the extreme climate typical of arid zones through structural
and behavioural adaptations, and the biology of some taxa, such as scorpions and
camel spiders, makes them particularly well-suited to these environments (see
Cloudsley Thompson 1975 & 1983; Crawford 1981; Polis 1991a).
Both light and pitfall traps provided a measure of adult invertebrate abundance, but
apart from antlion larvae, few immature holometabolous insects were captured.
Many surface-active desert insects, including tenebrionids, are long-lived as adults
but their larvae are subterranean feeding on roots (Seely 1991). We know little of
the life history characteristics that govern the susceptibility and response of
invertebrates to favourable and unfavourable conditions, but Crawford (1991)
suggests that desert detritivores show three types of response to changes in
environmental conditions: short-lived with a rapid, opportunistic response (‘r-
selected' species; MacArthur & Wilson 1967) e.g. collembola, thysanurans, mites,
some beetles and Diptera; long-lived with few tight linkages to environment (‘K-
selected' species) e.g. large tenebrionids (and also large predators such as
scorpions); intermediate often with eusociality (some ants and isopods). The role of
root-feeders, such as immature invertebrates, has rarely been assessed in any
natural environment because they generally remain underground, have a long
development time and are difficult to detect (Brown & Gange 1990). Soil moisture is
probably the single most important factor affecting the survival of underground
larvae, which are particularly vulnerable to desiccation and abrasion (Seely 1991).
Attempts to breed tenebrionids in UAE produced copious larvae, fewer pupae but
no adults (Mitchell 1995). The cause of mortality was not known, although

164
insufficient moisture at a critical stage was probably a contributing factor (M. Seely
in litt.).
Distribution of desert invertebrates
In UAE, the spatial differences in diversity suggested a trend towards a more
impoverished entomofauna for the most inland and arid sites. A similar trend in
invertebrate occurrence with distance from coast occurs in the Namib desert, and is
related to the amount of moisture provided by fog, which decreases further inland
(Seely 1991).
Differences in the diversity between sand and gravel substrates were greater than
differences between geographic sites in Abu Dhabi (Section 4.1). Factors
responsible for these differences include microclimate and habitat requirements, as
well as the dispersal abilities of arthropod species. Substrate characteristics are
particularly important to burrowing animals and factors such as the lability of sand
also determine the ease of root penetration by plants. Thomas (1983) noted
differences in the tenebrionid species on sandy or gravelly soil, with six species
occurring exclusively on sand, three on gravel and five on both, which may be
related to their burrowing abilities. Similar trends occur among arachnid species,
and psammophilic (sand-loving) spiders and scorpions can only burrow on sand
(Polls 1990b). Lamoral (1978) suggests that soil hardness governs the distribution
of species of African Opisthophthalmus scorpions, and has led to spéciation with
extant sympatric species separated by their ability and morphology for burrowing in
hard soils. For non-burrowing species, the availability of refuges with suitable
microclimates may be a limiting factor, e.g. pholcid and theridiid spiders rely on
vacant burrows made by other species (Polis & Yamashita 1991).
In terms of dispersal ability, some predators such as large carabids would be
expected to move further than sit-and-wait predators (some scorpions and spiders).
In temperate farmland, Frampton etal. (1995) found that the dispersal of carabids
varied with habitat, and was more rapid through crops than natural grassland.
Causal effects were thought to include prey availability, cover, plant density and
microclimate. Crist & Wiens (1995) studied and modelled tenebrionid movements in
North American arid-grasslands and found that pitfall transects had high probability
of capturing transient individuals from open population structures. Such effects are

165
likely to have influenced the arthropods captured in pitfall traps in Abu Dhabi
(Chapters 2 & 4).
Diversity
The current study increased the records of invertebrates in UAE by over 38% to 508
recognisable species, subspecies or genera (Appendix 4.1; Tigar 1996a). The new
records included many ants and tenebrionids, which were frequently caught in pitfall
traps and had previously been little studied. Base-line data have been published
(see Tigar 1996a & b; Tigar & Osborne 1997; Collingwood etal. 1997) and a
reference collection has been established to facilitate further research.
Deserts are often thought to show poor species diversity (Noy-Meir 1974; Polis
1991a), although certain taxa, particularly ants, tenebrionids and some predatory
arthropods are considered to be more numerous and diverse in deserts than more
mesic environments (see reviews in Cloudsley Thompson 1991 & 1996; MacKay
1991; Crawford 1991; Polls & Yamashita 1991). The comparison of community
composition between deserts is difficult because of the varying amount of effort
(time) and sampling methods used by different authors, and is exacerbated by
confusion over taxonomic resolution. Records for Abu Dhabi suggest that the
Formicidae (Hymenoptera), Sphecoidea (Hymenoptera) and Tenebrionidae
(Coleoptera) are the most diverse taxa, with 65, 115 and 65 species or subspecies
respectively (Appendices 1.1 & 4.1). However, in the case of Hymenoptera in UAE,
this also reflects current enthusiasm among collectors, and much of this information
was collected in surveys for single taxa (see Hamer 1986a & b, 1988; Guichard
1988 a & b, 1989 a & b, 1993; Collingwood 1985,1988; Collingwood & Agosti 1996;
Collingwood etal. 1997).
Nevertheless ants play important and diverse roles, and because of their colonial
nature they contribute a major part of the biomass in deserts (MacKay 1991). This is
also true in Abu Dhabi, where ants were the most numerous taxa caught in the
pitfall traps (Section 4.2) and found in houbara faeces and gizzards. Of the less
well-known Hymenoptera, the Mutillidae were particularly numerous in pitfalls and
light traps, suggesting that these parasitoids exert a heavy mortality on their hosts
(probably other Hymenoptera; Richards & Davies 1980). Other predatory arthropods
are well-represented in deserts compared with more mesic habitats (Polis &

166
Yamashita 1991), and data from other regions suggest that solifugid and spider
diversity were either under-sampled or underestimated in the present study (see
summary in Polis & Yamashita 1991). Many UAE specimens cannot currently be
determined beyond family or genus. However, eight species of scorpion were
identified (Appendix 4.1), which falls within the range for deserts (mean = 7.1 ± 2.5
SD in Polis & Yamashita 1991).
Faunal comparisons and the description of new species provide much of the
impetus for taxonomic studies (see Fauna of Saudi Arabia series). However, a
fauna for which only incomplete taxonomic keys exist, and where within-species
variation and sexual dimorphism are frequent (e.g. Tenebrionidae; Boorman, J. pers
comm.; Hymenoptera and Orthoptera; Richards & Davies 1980), presents a
challenge to the non-specialist. Collaboration between taxonomists and researchers
is vital, and species lists and descriptions must be published. Both Polis (1991c & d)
and MacKay (1991) note that the current unfashionable status of taxonomy
hampers many lines of research on ecological theory in arid zones.
By understanding the characteristics of a species, particularly life history and
autecology, and the heterogeneity of environment it is possible to predict broad
scale patterns of species abundance. Wiens (1976) even suggests that the spatial
and temporal heterogeneity of deserts can result in a higher species abundance
and diversity than in more uniform environments because competing species co
exist in different habitat patches. In addition, differences in adaptation to host-plant
chemistry and plant architecture influence the number of herbivorous species
occupying a plant (Lawton 1983; Strong etal. 1984; Wisdom 1988). Grazing by
livestock has a considerable effect on both the architecture of woody plants and
plant community composition (Thalen 1979; Floret 1981; Oatham 1996).
In North America, there is good knowledge of the invertebrates associated with
particular plants, e.g. Prosop/s trees (Wisdom 1991) but most Arabian studies have
considered either insects or plants in isolation (Oatham 1996; Roshier et ai. 1996;
see also Fauna of Saudi Arabia series). Broader ecological studies linking animal
communities with the underlying distributions of plants will lead to a greater
understanding of general underlying ecological processes. Geographical
Information System (GIS) in combination with robust analytical techniques such
GLM, would provide powerful tools for hypothesis testing. Deserts also provide good

167
places to study and test metapopulation and patch dynamics (see Hanski 1991 ;
Wiens 1976 & 1985), with the relatively stable microhabitats provided by perennial
plants and burrows acting as refuges from physical conditions and poor food
availability, connected to similar habitat patches via dispersal.
Permanent study sites, like the Coachella Valley in California (Polis 1991b & d),
emphasise the value of long data series, where each researcher has added to the
layers of ecological information. Indeed, short-term studies in stochastic
environments may be highly misleading since annual variations in biological and
climatic data are a characteristic of these ecosystems. In Abu Dhabi, Baynunah is
emerging as a potentially useful study site, and baseline data on invertebrates,
houbara and other fauna and flora have been published (Osborne 1996a; Osborne
et al. 1996; Oatham 1996).
6.2 The desert food web
Polis (1991b & d) provides an example of a desert food web (see definition in
Lawton 1989) from Coachella Valley based on a 20-year data set and about 820
publications. This web is complex because of two factors: the diversity of
interactions, and omnivory. Indeed, Polis suggests that many published food webs
are incomplete because of poor taxonomic discrimination and an inadequate
knowledge of trophic interactions, features also recognised by Pimm etal. (1991)
and Cohen etal. (1993).
Apart from the houbara (Chapter 5), the Abu Dhabi study was not designed to
investigate trophic relationships, and taxonomic discrimination was a problem for
some groups. However, a few observations of feeding were noted for arthropods
(Table 6.1), and most are typical of desert interactions although the laboratory and
pitfall observations should be treated with caution. In the absence of better data, it
seems likely that a web for Abu Dhabi would show much of the complexity of the
Coachella web, although the species would be different. Indeed, it is hard to
compete with Polis’s (1991b) observations of 2,000 man-hours in the field for a
single scorpion species.

168
Trophic relationship Species (Order or Family)
Food Source Type of observation
Coprophage Scarabaeus cristatus (Scarabaeidae)
Camel dung (provisioning for nest)
Field
Detritivore/Omnivore Mesostena puncticolis (Tenebrionidae)
Lab diet Lab
Pimelia arabica (Tenebrionidae)
Lab diet Lab
Ocnera philistina (Tenebrionidae)
Lab diet Lab
Blaps koiiari (Tenebrionidae)
Lab diet Lab
Herbivore-host plant Tentyrina palmeri (Tenebrionidae)
Haloxylon salicornicum (Chenopodiaceae)
Field & lab
Granivore Messor sjpjp. (Formicidae)
Seeds {Tribulus sp.?) Field
Predator-prey Apistobuthuspterygocercus(Scorpiones)
Scarabaeus cristatus (Scarabaeidae)
Field
Galeodes sp. (Solifuges)
Various Acrididae Field & lab
Mesostena puncticolis (Tenebrionidae)
Isoptera Field (under wood)
Scarites guineensis (Carabidae)
Bunopus tuberculosis (Reptilia)
Pitfall trap
Scarites guineensis (Carabidae)
Erodius spp. (Tenebrionidae)
Pitfall trap
Cannibal Mesostena puncticolis (Tenebrionidae)
Mesostena puncticolis (larva) (Tenebrionidae)
Lab
Cannibal/detritivore Mesostena puncticolis (Tenebrionidae)
Mesostena puncticolis (Tenebrionidae)
Lab
Ocnera philistina (Tenebrionidae)
Ocnera philistina (Tenebrionidae)
Lab
Blaps koiiari (Tenebrionidae)
Blaps koiiari (Tenebrionidae)
Lab
Pimelia arabica (Tenebrionidae)
Pimelia arabica (Tenebrionidae)
Lab
Intra-guild predation Scarites guineensis (Carabidae)
Parabuthus liosoma (Scorpiones)
Pitfall trap
Galeodes sp. Mantidae LabNecrophage Anthia
duodeclmguttata(Carabidae)
Chicken (Aves) Field
Scarabaeus cristatus (Scarabaeidae)
Gerbilus cheesmanii (Rodentia)(provisioning for nest)
Field
Table 6.1 Trophic relationships in Abu Dhabi (excluding houbara). Cannibal/
detritivores may have been scavenging on dead or dying insects rather than
depredating them. Laboratory diet: rolled oats, dates, almonds and carrots.

169
Polis’s suggestion that omnivory is a common feature of desert animals is
supported for many vertebrates (Wiens 1991), including the houbara, and
invertebrates (Seely 1991; MacKay 1991; Cloudsley Thompson 1996). Desert
arthropods may be more generalised and opportunistic in prey choice compared
with those of mesic environments, increasing the likelihood of interactions between
species. For example, there are many records of intra-guild predation in deserts
(Polis & Yamashita 1991). Spiders, solifugids, scorpions, mantids and reduviids
conform to this pattern; they are resource generalists and are frequently intolerant
of each other. Parasitism and hyper parasitism also occur, and predatory
arthropods are hosts for many parasitoids, e.g. during winter in the Chihuahuan
desert pompilid wasps consumed up to 65% of the population of the spider
Geolycosa rafaelana, although this dropped to <5% in summer (Conley 1985).
Temporal variations in desert food webs are also apparent and age structure is
particularly important (Polis 1991b & d). Age-dependent mutual predation is a
common occurrence, and while immature scorpions eat different and smaller prey
from adults, they are themselves subjected to intra-guild predation or cannibalism
by larger scorpions. Some authors suggest that loops in food webs, where species
A eats B, B eats C, and C eats either B or A, are unlikely events in the real world
(Pimm 1982). Again this is refuted by numerous examples in Polis (1991c). In Abu
Dhabi, laboratory observations of Galeodes sp. suggested intra-guild predation on a
mantid, and cannibalism of larvae by adult M. puncticolis. Intra-guild predation of a
small scorpion by the large carabid Scarites guineensis was observed in a pitfall
trap, while two tenebrionids, M. puncticolis and T.palmeri, which were assumed to
be detritivorous, showed predatory and herbivorous tendencies (respectively) in the
field.
Other generalisations about food webs, such as short linkages between trophic
levels, do not apply to the Coachella web which generally has long linkages
(between 6 and 11), often lengthened by soil biota interactions. Polis (1991d)
includes all possible linkages, even rare observations, because he argues that
without a thorough understanding of the diet of a species we cannot recognise
unrepresentative foods. Such information is usually lacking and some food webs
may simply be incomplete.

170
Certain features of deserts, including relatively high proportions of arachnids and
other predators, significant temporal and spatial stochasticity with pulses of food
and incoming migrant or nomadic animal populations, may result in more flexible
species interactions, with food webs that are more complicated than those of mesic
environments. The question of whether desert webs are unique can only be
resolved by further research, using standardised methods as advocated by Cohen
etal. (1993).
6.3 Studying houbara and other avian diets
The method for faecal analysis developed using captive houbara was successfully
applied to food remains in faeces of wild birds, and provided a calibration for the
differential digestion of prey (Chapter 5). The use of a standard technique based on
key fragments enables faecal analysis to be carried out more quickly and
objectively. However, if novel foods are suspected in different parts of the houbara’s
range further calibration trials should take place to identify appropriate fragments.
When studying the diet or foraging tactics of animals it is important to understand
how they choose prey. The prey groupings based on body size, speed and mode of
locomotion (Table 5.1) worked well on the faecal samples. Some groups were more
taxonomically diverse than others, but such groupings are justified because birds
are unlikely to discriminate on taxonomic features alone (Cooper etal. 1990).
Indeed, prey location and size are more likely to be used by birds to judge the time
needed to capture prey and prey energy content (Maurer 1990). Avian vision is
different from our own and in addition to the human range of vision, birds can see
well in the UV spectrum (Bennett & Cuthill 1994), which they use during mate
choice (Bennett etal. 1996). Birds may also find prey using UV (see raptor foraging;
Viitala etal. 1995), and this may affect food choice in the houbara, although the
most likely candidate prey, scorpions which show UV fluorescence, are active
nocturnally when natural UV light is low.
The role of plant material in the houbara diet should also be investigated further.
The recommended method is the microscopic examination of epidermal layers
across a slide transect (see Summers etal. 1993; Section 5.2), which is less
subjective than examining whole plant remains.

171
Faecal analysis is not without its problems (see Section 5.2), but providing it is
calibrated, it is the best technique for studying the diet of wary and endangered
birds including the houbara and other bustards. Gastro-intestinal flushes (Major
1990), emetics and ring-collars have been used to study avian diets (see review by
Rosenberg & Cooper 1990) but are only suitable for smaller and more common
species. The effort and risk in catching bustards is great, and may result in their
death or a non-fatal injury, including capture myopathy (Windingstad etal. 1983).
This is a temporary paralysis affecting long-legged birds, which prevents them from
standing or walking. Any injury caused during catching would render a wild bird
more vulnerable to predators.
The estimate of energy consumption provides a guide to houbara metabolic needs,
but is not a direct measure. Bryant & Tatner (1991) examined variations in energy
expenditure of birds with body masses at least an order of magnitude less than the
houbara (from 9.5 to 143.4g) and suggest that it is unwise to assume the costs of
one species apply to another. They found that different factors were correlated with
energy expenditure in different species, although flight and brood-provisioning were
common causes of increased energy expenditure. However, other authors have
proposed general equations for energy consumption in birds (Kendeigh etal. 1977),
and provide examples for different bird taxa including passerine or non-passerines;
and during different periods or conditions which influence energy expenditure:
migration, reproduction, moulting, extremes of temperature, changes in
photoperiod, wind velocity, insolation, radiation and other environmental factors (see
reviews in Carey 1996). Adaptation to local climate is important, with desert and
tropical species generally having a lower metabolic rate than related birds from
mesic environments (Ricklefs 1996). Ideally, the estimate for the houbara should be
tested using the doubly labelled water technique (Nagy 1983).
Much of the work on energy expenditure centres on critical values for the
reproductive cycle of birds (e.g. Martin 1987; Carey 1996). Nesting and brood-
provisioning are very energy-intensive processes, and altricial birds may desert the
nest if they lack sufficient energy reserves. The breeding strategy of precocial birds,
such as the houbara, avoids this risk since their young feed themselves from a very
early age. However, precocial birds may require more energy prior to the breeding
season, because their eggs are larger and their yolks have a higher calorific value
than those of altricial birds. Therefore, the over-wintering period in Abu Dhabi may

172
be important in preparing houbara for the following breeding season in Central Asia.
Houbara are long-lived birds and may delay breeding or wait until the following year
if their energy reserves are too low. Any work on energy expenditure or food
limitation would do well to examine resident houbara first, before considering the
costs of more complex migratory birds.
Models of foraging theory hâve been applied to many birds, with varying success
(see reviews in Stephens & Krebs 1986; Stephens 1990). The theory assumes that
animals will act economically and choose feeding strategies that yield more food, or
another currency such as energy or protein, in less time. The functional response of
predators to their prey changes as prey abundance increases, and smaller less
profitable prey may only be eaten when sufficiently common, e.g. in a clumped
distribution (Caraco etal. 1980). Such a strategy could occur in the houbara in
response to social insects or locust swarms, which may only be included in the diet
when sufficiently numerous (note the distribution of Messor spp. ant remains in
houbara faecal samples; Chapter 5). Prey selection in omnivores has been largely
ignored or considered as intermediate between carnivores and herbivores, since
omnivores consume both high-quality animal prey and lower-quality plant foods
(Westoby 1978). Carnivores are thought to be constrained by the search and
handling time needed to obtain prey, while herbivores are constrained by their need
for a nutritionally balanced diet (Pyke 1984). Ball (1994) provides an excellent
account of foraging theory for an omnivorous, diving duck (the canvasback Aythya
valisineria) which did not conform to an energy-maximisation model when tested
with four foods. He suggested that size, texture and perhaps taste were more
important in prey choice than energy (some ducks even rejected prey they had
caught by spitting it out!). However, this situation is more complex than foraging in
the houbara, because of the high energetic costs of diving.
6.4 Houbara diet in UAE
Houbara are omnivorous, with invertebrates making up much of the bulk and most
of the energy of UAE birds. The most important prey were ants in terms of number,
and tenebrionid beetles in terms of total biomass and energy contribution. Bustards
are directly exposed to the same environmental stresses of desert life as their
foods, and by eating insects which have a high water content compared with seeds

173
or dried plant-parts, they avoid the need for supplementary water, although they
drink when in captivity on an artificial diet.
The use of different methodologies, each with their own inherent biases, make it
difficult to compare between studies from other parts of the houbara’s geographic
range (see Chapters 2 & 5), although Fox’s (1988) work suggests that more reptiles
are eaten by C.u.undulata in Morocco. Future work should look at the pattern of
foods eaten by houbara, to see whether they respond to differences in locally
abundant foods or consistently choose particular food types across the range.
Why do houbara occur in UAE when their principal food, invertebrates, is least
abundant, indeed most foods are at their least abundant? The answer may lie in
their breeding grounds in Central Asia rather than Abu Dhabi. The former frequently
experience sub-zero temperatures with deep snow-cover (Osborne etal. 1997a & b)
and it may be more profitable in terms of energy gain to migrate to Arabia where the
winters are less extreme and food is available. In addition, the high summer
temperatures experienced in Abu Dhabi may be too stressful for houbara.
An alternative hypothesis is based on the “source-sink” model of den Boer (1968).
Perhaps the resident, breeding, Arabian houbara in Oman and Saudi are source
populations which quickly fill vacant habitat patches where food availability is not
limited. Any excess birds might then become a sink population, occupying sub-
optimal habitat patches, and perhaps becoming nomadic or migratory across
Arabia. Indeed, Seddon & van Heezik (1995) suggest that the houbara population in
the Harrat al-Harrah reserve in Saudi Arabia is boosted by seasonal migrants,
although their origin is not known. The migration route of houbara over-wintering in
UAE to their breeding grounds in Central Asia has only recently been confirmed,
and is based on a few individuals (Osborne etal. 1997a). We know nothing of other
migration routes or whether there is any interchange of genetic material between
houbara populations in the Middle East. Such work would involve satellite-tagging
wild birds and genetic analyses to examine the relatedness of populations, which
would require capturing houbara and therefore carry an inherent risk of mortality
(see Section 6.3).

174
6.5 C onservation o f H oubara and w ild life in UAE
The houbara is a useful flagship species for the desert. Planning for its dietary
requirements and need for undisturbed areas with reduced human activity and
grazing pressure would also benefit other wildlife, although long-term habitat
management may benefit from controlled grazing (Oatham 1996). Most of Abu
Dhabi's desert is open-access, apart from urban areas, private estates, oil-fields,
forestry plantations and a few military zones. The need for protected areas has
been recognised but to date, none have been designated (Osborne & Hornby
1995). The successful functioning of protected areas requires strong political will
and financial commitment, and lessons from other areas suggest that grazing and
hunting rights may be a cause for particular concern in the Middle East (Child &
Grainger 1990; Seddon etal. 1995). The potential harmful effects of development
and environmental change have attracted growing recognition in UAE and in 1993
the Federal Environmental Agency was established (Anon 1993b).
Nomadism is often considered a characteristic of desert birds, which move in
response to changes in food availability (Wiens 1991), and the houbara’s long and
short-distance movements are probably responses to local environmental
conditions. Historical records of resident-breeding houbara in UAE may reflect
previous eras when rainfall was more frequent (Thesiger 1959). The mobility of
birds allows them to exploit differences in habitat and micro-habitat patches in all
deserts such as potential food supply, nest sites etc.
In view of the increasing pace of development in UAE (Oatham 1996; Collingwood
etal. 1997), it would be wise to monitor environmental change. Much of the recent
interest in wildlife has centred on houbara and other large vertebrates (Hellyer
1996; Aspinall 1996). However, within the UK, Lawton (1996) noted that while birds
were useful flagship species, they were not good indicators of species-rich sites of
high conservation value for other taxa. Elsewhere, ants have been used as
indicators of habitat quality and biodiversity (Perfecto & Snelling 1995; Bestelmeyer
& Wiens 1996; Majer & Beeston 1996) and would be a useful Arabian group
because they are well known, easy to collect and are speciose with considerable
variations in habitat requirements. Majer & Beeston (1996) noted that ant diversity
in Australia was reduced by road construction, and to a lesser extent by agricultural

175
development, urbanisation and rangeland grazing. In UAE, Collingwood etal.
(1997) noted that 14 of the 65 species of ants collected were tramp species,
including several cosmopolitan pests and a fire ant Solenopsis geminata. The latter
is of particular concern because of its deleterious effect on native fauna and flora
following an accidental introduction into North America (Williams 1994). Similarly,
the distribution of other typical desert arthropods, such as scorpions, camel spiders
and tenebrionids, might provide useful indications of habitat quality once their
biology and taxonomy are better known.
The only note of caution is that the perception of biodiversity varies according to the
taxa being considered (Gaston 1998). Indeed Gaston found little evidence of either
congruence in geographical hot spots among different taxa, or strong correlations in
biodiversity between different groups across multiple areas. While measures of
biodiversity are highly variable, it is often the less popular taxa, such as
invertebrates, that are ignored in conservation plans (Sutherland 1998).
6.6 Implications for captive breeding programmes
Houbara have been successfully reared at Taif, Saudi Arabia, and released into a
protected reserve, Mazhat az-Sayd (Seddon et al. 1995). However, this was not
without high initial mortality, thought to be due to predator-naive birds being taken
by foxes. Other factors may also have influenced predation, including food
availability away from supplementary feed sites and irrigated plots. Naïve houbara
in poor condition would, according to state dependent foraging, feed in areas and at
times with a high risk of predation (see McNamara 1990), and the foxes may have
responded to this. Foxes may also have scavenged from the carcasses of houbara
that had died of other causes, including starvation.
Evidence from this study (Chapter 5) and other authors (Anegay 1994; Combreau &
Launay 1997) suggests that houbara forage nocturnally, particularly around a full
moon. However, captive birds are generally fed in the morning, when bird keepers
in UAE start work. A change in feeding-times to late afternoon would allow houbara
to forage around dusk, in tune with their innate behavioural rhythms. Indeed,
Hermans (1988a) remarked that much of the food given to houbara at Taif was not
eaten, and soon became rancid and unpalatable to the birds. Fresh foods, including
mince, apples and lettuce, are mixed with moistened food pellets and given to

176
houbara in UAE where unless they are eaten Immediately they soon become
spoiled (personal observation). Artificial lighting might also be beneficial to houbara,
particularly during the summer when the birds could feed nocturnally and avoid heat
stress.
The planting of alfalfa in houbara aviaries is already general practice, and is doubly
effective since it provides a source of fresh green material and herbivorous insects,
such as grasshoppers and weevils, which houbara readily hunt (A. Owen pers.
comm.). In addition, a native plant of the UAE Dipterygium giaucum occurred in
both faecal samples and gizzard contents. Attempts to cultivate and encourage this
plant would be beneficial in both aviaries and areas where houbara over-winter, or
for houbara release sites.
6.7 Conclusions
This study has answered its primary objectives as far as possible. It has provided
information on invertebrate diversity, as well as data on general trends in the
abundance of desert invertebrates. The diet of the houbara was determined using a
calibrated faecal analysis and the examination of uncalibrated gizzard contents.
Houbara take a wide variety of food and invertebrates, especially tenebrionid
beetles, make up much of the bulk in Abu Dhabi.

177
Epilogue
Two main themes run through the thesis; variations in the abundance and diversity
of desert arthropods, and the diet of the houbara. However, abiotic factors of the
desert explain much of this variation. In his classic work “Arabian Sands”, the great
explorer Wilfred Thesiger tells of his travels with the bedu of Abu Dhabi and his
respect for their nomadic way of life. On many occasions, the lives of Thesiger and
his companions were dependent upon the bedu's ability to find the scarce grazing
for their camels. I end with a quote in which Thesiger ponders the question of how
life arrives at isolated habitat patches (Thesiger 1959).
“Four hours later we came to large red dunes set close together. There were green
plants growing there as a result of heavy rain which had fallen two years
before \Ne camped In a hollow and loosed our camels to revel among the Juicy
shrubs.
Larks were singing round our camping place. Butterflies flitted from plant to plant.
Lizards scuttled about, and small black beetles walked laboriously across the
sand. still marked where jerboas and other small rodents had scampered about
during the night. I wondered how they got here, how they had located this small
green island. In the enormous emptiness which surrounded it.”

178
References
Aebisher, N. J., Robertson, P. A. & Kenward, R. E. (1993). Compositional analysis
of habitat use from animal radio-tracking data. Ecology!^: 1313-1325.
Afanas'ev, A. V. & Sludskiy, A. A. (1947). Report on the mammals and birds of
Central Kazakhstan. Izv. Akad. NaukKaz. SSR Ser. Zoo/. 6: 48-64. Alma-Ata.
Aharoni, J. (1912). Houbara macqueenii Gray. Orn. Jahrb. 23: 1-15.
Ahearn, G. A. (1971). Ecological factors affecting population sampling of desert
tenebrionid beetles. The American Midland Naturalist 86: 385-406
Aldryhim, Y. N ., Mills, C. W. & Aldawood, A. S. (1992). Ecological distribution and
seasonality of darkling beetles (ColeopteraiTenebrionidae) in the Central
Region of Saudi Arabia. Journal of Arid Environments 23: 415-422.
Alekseev, A. F. (1985). The houbara bustard in the north-west Kyzylkum. Bustard
Studies 3: 87-93. (First published in Russian in 1980 as Zoo/. Zhurn. 59:
1263-1266.)
Ali, S. & Ripley, 8. D. (1969). Birds of India and Pakistan. Oxford University Press,
Delhi.
Alonso, J. C. & Alonso, J. A. (1996). The great bustard Otis tarda in Spain: Present
status, recent rends and evaluation of earlier censuses. Bioiogicai
Conservation 77: 79-86.
Anderson, S. (1995). Bustard micronutrient review. Unpublished report. National
Avian Research Center External Report 4. National Avian Research Center,
Abu Dhabi.
Anegay, K. (1992). Observations on the feeding behaviour of a group of houbara
bustards {Chiamydotis unduiata macqueenii) in semi-captivity in the Mahazat
As Said Reserve (April 1991-March 1992). Unpublished report. National
Wildlife Research Center, National Commission for Wildlife Conservation and
Development, Kingdom of Saudi Arabia.
Anegay, K. (1994). Diurnal and nocturnal activity patterns of semi-captive houbara
bustard Chiamydotis unduiata. Sandgrouse 16: 41-46.
Anon. (1980). Tunisian delegation: report on the houbara bustard. In: Coles, C. L. &
Collar, N. J. (eds.) Symposium papers of the small game commission of the
Conseil International de la chase. The great bustard, Sofia 1978. The houbara
bustard, Athens 1979. The Game Conservancy, Fordingbridge.
Anon. (1993a). National Atlas of the United Arab Emirates, United Arab Emirates
1993. Published by GEOprojects, Reading.

179
Anon. (1993b). The Federal Environmental Agency. Federal Law Number 7
Government of the United Arab Emirates, Abu Dhabi.
Aspinall, S. (1996). Status and conservation of the breeding birds of the United Arab
Emirates. Hobby Publications, Dubai and Warrington.
Axell, H. E. (1964). Houbara bustard in Suffolk. British Birds 57: 247-249.
Ayal, Y. & MerkI, O. (1994). Spatial and temporal distribution of tenebrionid species
(Coleoptera) in the Negev Highlands, Israel. Journai of Arid Environments 27:
347-361.
Badr, Y. (1976). Falconry as a sport: our Arab heritage. Ministry of Information and
Culture, Abu Dhabi.
Baha El Din, S. (1996). Terrestrial reptiles of Abu Dhabi. In: Osborne, P. E. (ed.)
Desert ecology of Abu Dhabi- a review and recent studies. Pp. 124-147.
Pisces Publications, Newbury.
Bailey, T. (1992). Veterinary aspects of bustards at AI Ain Zoo and HH Sheikh
Khalifa's farm at AI Ain, UAE. Unpublished report. Nationai Avian Research
Center Internal Report Series 11. National Avian Research Center, Abu
Dhabi.Bailey, T. & Hornby, R. (1994). National Avian Research Center annual report 1993.
Motivate Publishing, Dubai.
Baker, S. E. C. (1912). The gamebirds of India, Burma and Ceylon. Part 6. Journal
of the Bombay Natural History Society 21: 303-337.
Baldwin, R. (1997). Marine mammals. In: Vine, P. J. (ed.) Natural Emirates. Pp.
199-212. Trident Press, London.
Ball, J. P. (1994). Prey choice of omnivorous canvasbacks: imperfectly optimal
ducks? Oikos 70: 233-244.
Banks, S. (1990). Signal processing, image processing and pattern recognition.
Prentice Hall International, UK.
Bannerman, D. (1963). Birds of the Atlantic Islands. 1. History of birds of the Canary
Islands and Salvages. Oliver & Boyd, Edinburgh.
Bannicov, A. G. & Skalon, W. N. (1948). Ornithologische Notizen über die Mongolei.
Ochrana Prirody (Naturschutz) 5:17-31.
Beasley, C. A. & Adams, C. J. (1994). Relationship between environmental factors
and capture time of the pink-bollworm (Lepidoptera, Gelechiidae) moths in
traps baited with sex-pheromone. Journai of Economic Entomology Q7: 986-
992.

180
Bédé, P. (1928). Notes d'ornithologie. Jardin Zoologique de Sfax Fascicule 1. Sfax
Tunisia.
Bell, G. P. (1990). Birds and mammals on an insect diet: a primer on diet
composition analysis in relation to ecological energetics. In: M. L. Morrison,
Ralph, C. J., Verner, J. & Jehl, J. R. (eds.) Avian foraging: theory,
methodology and applications. Studies in Avian Biology'IZ: 104-109. Cooper
Ornithological Society, California.
Bennett, A. T. D. & Cuthill, I. C. (1994). Ultraviolet vision in birds: what is its
function? Vision Research 3^: 1471-1478.
Bennett, A. T. D. & Cuthill, I. C., Partridge, J. V. & Maier, E. J. (1996). Ultraviolet
vision and mate choice in zebra finches. Nature 380: 433-435.
Bennett, P. M. & Harvey, P. H. (1987). Active and resting metabolism in birds:
allometry, phylogeny and ecology. Journal of the Zoological Society of London
213: 327-363.
Bestelmeyer, B. T. & Wiens, J. A. (1996). The effects of land-use on the structure of
ground-foraging ant communities in the Argentine Chaco. Ecological
Applications 6: 1225-1240.
Biquand, S., Gaucher, P., Paillat, P., Saint-Jalme, M. & Seddon, P. (eds.) (1992).
Houbara propagation. Unpublished report. National Wildlife Research Center,
National Commission for Wildlife Conservation and Development, Kingdom of
Saudi Arabia.
Boehme, L. B. (1926). A list of birds collected by P. I. Zhukov in northern Turkey in
1915-1917. Izv. Gorsk. s.-khoz. Inst 3: 97-102.
Boobyer, M. G. (1989). The eco-ethology of the Karoo korhaan Eupodotis vigorsii.
Unpublished M.Sc. thesis. University of Cape Town.
Boocock, D. (1979). Termites. Emirates Natural History Group Bulletin (Abu Dhabi)
8: 27-28.
Boorman, J. (1989). Culicoides (Diptera:Ceratopogonidae) of the Arabian Peninsula
with notes on their medical and veterinary importance. Fauna of Saudi Arabia
10: 160-224.
Bottomley, N. (1996). Recent climate of Abu Dhabi. In: Osborne P. E. (ed.) Desert
ecology of Abu Dhabi- a review and recent studies. Pp. 36-49. Pisces
Publications, Newbury.
Brosset, A. (1961). Ecologie des oiseaux du Maroc Oriental. Trav. Inst. Soi chérif
sér. Zoo/. No.22.

181
Brown, V. K. & Gange, A. C. (1990). Insect herbivory below ground. Advances in
Ecological Research 20: 1 -44.
Bryant, D. & Tatner, P. (1991). Intraspecies variation in avian energy expenditure:
correlates and constraints. Ibis 133: 236-245.
Bryant, J. P. & Kuropat, P. J. (1980). Selection of winter forage by subarctic
browsing vertebrates: the role of plant chemistry. Annual Review of Ecology
and Systematlcs 11: 261 -285.
Buffon (1835). Oeuvres completes. Tome VII Oiseau 1. Dumenil, Paris.
Büttiker, W. & Anderson, E. (1988). Water-extracting tenebrionid beetles from the
Wahiba sands, Oman. Journal of Oman Studies, Special Report 3: 321-324.
Büttiker, W. & Krupp, F. (eds.) (1980). Fauna of Saudi Arabia 1. Hurbert & Co.,
Gottingen.
Caraco, T., Martindale, 8. Whitham, T. S. (1980). An empirical demonstration of
risk-sensitive foraging preferences. Animal Behaviour 23: 820-830.
Carey, C. (ed.) (1996). Avian energetics and nutritional ecology. Chapman & Hall,
New York.
Chesmore, D. & Monkman, G. (1994). Automated analysis of variation in
Lepidoptera. The Entomologist'\^3: 171-182.
Chhotani, O. B. & Bose, G. (1983). Distribution of Isoptera in Arabia and their
affinities with the termites from the Indian Desert and neighbouring countries.
Fauna of Saudi Arabia 5:121 -128.
Chhotani, O. B. & Bose, G. (1991). Isoptera from Saudi Arabia and Kuwait, with a
key to Arabian species. Fauna of Saudi Arabia 12: 256-265.
Child, G. & Grainger, J. (1990). A system plan for protected areas for wildlife
conservation and sustainable development in Saudi Arabia. Unpublished
report. National Commission for Wildlife Conservation and Development,
Kingdom of Saudi Arabia.
Clarke, J. A., Chopko, J. T. & Mackessy, S. P. (1996). The effect of moonlight on
activity patterns of adult and juvenile prairie rattlesnakes {Crotalus virldus
virldus). Journal of Herpetology 30:192-197.
Clements, F. A. (1983). United Arab Emirates. World bibliography series, volume
43. Clio Press, Oxford.
Cloudsley Thompson, J. L. (1975). Adaptations of Arthropoda to arid environments.
Annual Review of Entomology 20: 261-283.
Cloudsley Thompson, J. L. (1977). Adaptational biology of Solifugae. Bulletin of
British Arachnology Society 4: 61-71.

182
Cloudsley Thompson, J. L. (1983). Desert adaptations in spiders. Journal of Arid
Environments 6: 307-317.
Cloudsley Thompson, J. L. (1984). Arachnids. In: Cloudsley Thompson, J. L. (ed.)
Sahara Desert (Key Environments). Pp. 175-204. Pergamon Press, Oxford.
Cloudsley Thompson, J. L. (1987). Solifugidae and keeping them in captivity. In:
(eds.) Cooper, J. E., Pearce-Kelly P. & Williams, D. L. Arachnida -
proceedings of a symposium on spiders and their allies. Pp. 52-56. Chiron
Publications Ltd., London.
Cloudsley Thompson, J. L. (1991). Ecophysiology of desert arthropods.
(Adaptations of desert organisms). Springer, Berlin.
Cloudsley Thompson J. L. (1996). Biotic interactions in arid lands. (Adaptations of
desert organisms). Springer, Berlin.
Cohen, J. E., Beaver, R. A., Cousins, S. H., DeAngelis, D. L., Goldwasser, L.,
Heong, K. L., Holt, R. D., Kohn, A. J., Lawton, J. H., Martinez, N., O’Malley,
R., Page, L. M., Pattern, B. C., Pimm, S. L., Polis, G. A., Rejânek, M.,
Schooner, T. W., Schoenly, K., Sprule, W. G., Teal, J. M., Ulanowicz, Warren,
P. H., Wilbur, H. M. & Yodzis, P. (1993). Improving food webs. Ecology 74\
252-258.
Collar, N. J. (1980). The world status of the houbara: a preliminary review. In:
Coles, C. L. & Collar, N. J. (eds.) Symposium papers of the small game
commission of the Conseil International de la chase. The great bustard, Sofia
1978. The houbara bustard, Athens 1979. The Game Conservancy,
Fordingbridge.
Collar, N. J. (1983). A history of the houbara in the Canaries (Report of the ICBP
Fuerteventura houbara expedition 1979). Bustard Studies 9-29
Collar, N. J., Goriup, P. D. & Osborne, P. E. (1986). Otididae, bustards. In: Urban,
E. K., Fry, C. H. & Keith, S. (eds.) The birds of Africa. Volume 2. Pp. 148-179.
Academic Press, London.
Collingwood, C. A. (1985). Hymenoptera: Fam. Formicidae of Saudi Arabia. Fauna
of Saudi Arabia 7: 230-302.
Collingwood, C. A. (1988). The ants (Hymenoptera:Formicidae) of the Wahiba
Sands, Oman. Journai of Oman Studies Special Reports: 361-362.
Collingwood, C. A. & Agosti, D. (1996). Formicidae (lnsecta:Hymenoptera) of Saudi
Arabia (Part 2). Fauna of Saudi Arabia 15: 300-385.
Collingwood, C. A., Tigar, B. J. & Agosti, D. (1997). Introduced ants in the United
Arab Emirates. Journai of Arid Environments S7: 505-512.

183
Collins, D. R. (1984). A study of the Canarian houbara bustard {Chiamydotis
unduiata fuertaventurae) with special reference to its behaviour and ecology.
Unpublished M.Phil. thesis. University of London.
Collins, D. R. (1993). The diet of the houbara bustard Chiamydotis unduiata
fuertaventurae in the Canary Islands. Boletim do Museo Municipal do Funchal
Supplement 2: 57-67.
Combreau, O. & Launay, F. (1996). Activity rhythms of houbara bustards
{Chiamydotis unduiata macqueenii) in relation to some abiotic factors. Journal
of Arid Environments 33: 463-472.
Combreau, O. & Rambaud, F. (1995). The houbara bustard programme in Mahazat
As-Sayd, April 1992-November 1994. Final report (unpublished). National
Wildlife Research Center, National Commission for Wildlife Conservation and
Development. Kingdom of Saudi Arabia.
Combreau, O. & Smith, T. R. (1997). Summer habitat selection by houbara
bustards introduced in Central Saudi Arabia. Journal of Arid Environments 35:
140-160.
Conley, M. R. (1985). Predation versus resource limitation in survival of adult
burrowing wolf spiders (AraneaeiLycosidae). Oecologia 67: 71-75.
Cooper, R. J. & Whitmore, R. C. (1990). Arthropod sampling methods in
ornithology. In: M. L. Morrison, Ralph, C. J., Verner, J. & Jehl, J. R. (eds.)
Avian foraging: theory, methodology and applications. Studies in Avian
Biology ^3: 29-37. Cooper Ornithological Society, California.
Cooper, R. J., Martinet, P. J. & Whitmore, R. C. (1990). Dietary similarity among
insectivorous birds: influence of taxonomic versus ecological catagorization of
prey. In: M. L. Morrison, Ralph, C. J., Verner, J. & Jehl, J. R. (eds.) Avian
foraging: theory, methodology and applications. Studies in Avian Biology\3:
104-109. Cooper Ornithological Society, California.
Cordeaux, J. (1896). Zoologist20: 433-444.
Cornwallis, L. (1982). Notes on a post-symposium tour to survey the position of
bustards (Otididae) in India. In: Goriup, P. D. & Vardhan, H. (eds.) Bustards in
decline. Pp. 353-364. Jaipur.
Cramp, S. & Simmons, K. E. L. (1980). Handbook of the birds of Europe, the Middle
East and North Africa. Hawks to bustards. Volume 2. Oxford University Press,
Oxford.
Cranston, P. S. & Judd, D. D. (1989). Diptera:Fam. Chironomidae of the Arabian
Peninsula. Fauna of Saudi Arabia 10: 236-289.

184
Crawford, 0. S. (1981). Biology of desert invertebrates. Springer-Verlag, New York.
Crawford, C. S. (1988). Surface-active arthropods in a desert landscape: influences
of microclimate, vegetation and soil texture on assemblage structure.
Pedobiologia 32: 373-385.
Crawford, C. S. (1991). The community ecology of macroarthropod detritivores. In:
Polis, G. A. (éd.). The ecology of desert communities. Pp. 89-112. The
University of Arizona Press, Tucson.
Crawford, C. 8. & Seely, M. K. (1987). Assemblages of surface-active arthropods in
the Namib Dunefield and associated habitats. Revue de Zoologie Africaine
101:397-421.
Crawley, M. J. (1983) Herbivory, the dynamics of animal-plant interactions. Studies
in ecology Blackwell Scientific Publications, Oxford.
Crist, T. O. & Wiens, J. A. (1995). Individual movements and estimation of
population size in darkling beetles (Coleoptera:Tenebrionidae). Journai of
Animal Ecology 6A: 733-746.
Délye, G. (1968). Recherche sur l’écologie, la physiologie et l’éthologie des fourmis
du Sahara. Unpublished thesis. Faculty of Science, Aix-Marseille, France.
Dement’ev, G. P. & Gladkov, N. A. (1951). (eds.) Ptitsy Sovietskogo Soyuza, 2. (in
Russian), Moscow. (Translated as: Birds of the Soviet Union, Jerusalem,
Israel.)
den Boer, P. J. (1968). On the survival of populations in a heterogeneous and
variable environment. Oecologia 50: 39-53.
Dent, D. R. & Walton, M. P. (eds.) (1997). Methods in ecology and agricultural
entomology. CAB International, London.
Dharmakumarsinhji, K. S. (1955). Birds of Saurashtra, India with additional notes on
the birds of Kutch and Gujerat. Pp. 561. Times of India Press, Bombay.
Dillon, L. S. & Dillon, E. S. (1972). A manual of the common beetles of eastern
North America. Second edition. Dover Publications, New York.
Dominguez, F. (1989). Censo de la pobulacion de hubara canaria {Chiamydotis
unduiata fuertaventurae) en la isla de Fuerteventura (diciembre de 1988).
Unpublished manuscript in Spanish.
Duckworth, W. (1996). Land mammals of Abu Dhabi. In: Osborne P. E. (ed.) Desert
ecology of Abu Dhabi- a review and recent studies. Pp. 147-171. Pisces
Publications, Newbury.

185
Dutton, R. W. & Bray, D. A. (eds.) (1988). The scientific results of the Royal
Geographical Society's Oman Wahiba Sands project 1985-1987. The Journal
of Oman Studies Special Report 3. Hindson Print Ltd, Newcastle upon Tyne.
Edney, E. B. (1971). Some aspects of water balance in tenebrionid beetles and a
thysanuran from the Namib Desert of southern Africa. Physiological Zoology
44: 61-76.
El Ghomeny, A. A. (1985). Ecology and flora of AI Ain region. Volume 1 ecology and
monocotyledons. United Arab Emirates University, AI Ain.
Faragalla, A. A. & Adam, E. E. (1985). Pitfall trapping of tenebrionid and carabid
beetles (Coleoptera) in different habitats in the Central Region of Saudi
Arabia. Zeitschrift fdr Angewandte Entomologie 99: 466-471.
Ferguson-Lees, I. J. (1969). Birds of the world volume 3. UK.
Feulner, G. (1997). Geology of the United Arab Emirates In: Vine, P. J. (ed.) Natural
Emirates. Pp. 21-40. Trident Press, London.
Finn, F. (1915). Indian sporting birds. Edwards, London.
Flint, V. E., Gabuzov, O. S. & Khrustov, A. V. (1992). Strategy for the conservation
of bustards. Bustard Studies 5: 2-7.
Floret, C. (1981). The effects of protection on steppic vegetation in the
Mediterranean arid zone of southern Tunisia. Vegetation 46: 117-129.
Fox, N. (1988). Notes on the analyses of houbara bustard stomach contents from
Baluchistan and Punjab. Unpublished manuscript.
Frampton, G. K., Çilgi, T, Fry, G. L. A. & Wratten, S. D. (1995). Effects of grassy
banks on the dispersal of some carabid beetles (Coleoptera:Carabidae) on
farmland. Bioiogicai Conservation 71: 347-355.
Furley, C. W., Greenwood, A. G. & Solomon, G. (unpubl.). Disease of bustards.
Unpublished manuscript.
Gallagher, M. D. (unpubl.). The bustards (Otididae) in Oman. 2nd draft.
Unpublished manuscript.
Gaston, K. J. (1998). Biodiversity. In: Sutherland, W. J. (ed.) Conservation science
and action. Pp. 1-19. Blackwell Science Ltd., Oxford.
Gaston, K. J. & Williams, P. H. (1996). Spatial patterns in taxonomic diversity. In:
Gaston, K. J. (ed.) Biodiversity - a biology of numbers and difference. Pp.
202-229. Blackwell Science, Oxford.
Gauch, H. G. (1982). Multivariate analysis in community ecology. Cambridge
University Press, Cambridge.

186
Gaucher, P. (1991). On the feeding ecology of the houbara Chiamydotis unduiata
unduiata. Alauda 59:120-121.
Gaucher, P., Paillat, P. & Eichacker, X. (1989). Houbara bustard {Chiamydotis
unduiata) report on NWRC breeding season. Unpublished report. National
Wildlife Research Center, National Commission for Wildlife Conservation and
Development, Taif, Kingdom of Saudi Arabia.
Gaucher, P., Paillat, P., Chappuis, C., Saint Jaime, M., Lotfikhan, F. & Wink, M.
(1996). Taxonomy of the houbara bustard Chiamydotis unduiata subspecies
considered on the basis of sexual display and genetic divergence. Ibis 138:
273-282.
Gavrin, V. F. (1962). Order bustards - Otides. In: Dolgushin, I. A. (ed.) The birds of
Kazakhstan 2, pp 5-39. Alma-Ata.
Geptner, V. G. (1959). Report on the birds of Badkhyz (southern Turkmenistan).
Trudy Inst. Zoo/. Parasltol. (Ashkhabad) A\ 125-44.
Ghabbour, S. I. & Shakir, S. H. (1980). Ecology of soil fauna in Mediterranean
desert ecosystems in Egypt. III. Analysis of Thymelaeae mesofauna
populations at the Mariut frontal plain. Revue Ecologie et Biologie du so/14:
429-459.
Gillett, H. (1988) Analysis of stomach contents of Algerian houbara. Unpublished
report. National Wildlife Research Center, National Commission for Wildlife
Conservation and Development, Taif, Kingdom of Saudi Arabia.
Gillett, M. (1996). Insects. In: Vine, P. J. (ed.) Natural Emirates. Pp. 95-106. Trident
Press, London.
Glennie, K, W. (1996). Geology of Abu Dhabi. In: Osborne, P. E. (ed.) Desert
ecology of Abu Dhabi- a review and recent studies. Pp. 16-35. Pisces
Publications, Newbury.
G lutz von Blotzheim, U. N., Bauer, K. M. & Bezzel, E. (1973). Handbuch der Vogel
Mitteleuropas, 5. Akademische Verlagsgesellschaft, Frankfurt.
Godfray, H. C. J. & Crawley, M. J. (1998). Introductions. In: Sutherland, W. J. (ed.)
Conservation science and action. Pp. 39-65. Blackwell Science Ltd., Oxford.
Goriup, P. D. (1997). The world status of the houbara bustard Chiamydotis
unduiata. Bird Conservation International (1997) 7: 373-397.
Goriup, P. D. & Collar, N. J. (1980). The ICBP Fuerteventura houbara expedition.
Spring 1979 - a preliminary account. In: Coles, C. L. & Collar, N. J. (eds.)
Symposium papers of the small game commission of the Conseil International

187
de la chasse. The great bustard, Sofia 1978. The houbara bustard, Athens
1979. The Game Conservancy, Fordingbridge.
Goriup, P. D. & Norton, J. A. (1992). Houbara bustard field research project. Report
for 1990. Private report to National Commission for Wildlife Conservation and
Development, Saudi Arabia.
Goriup, P. & Taylor, N. (1983). The houbara bustard in Morocco, report on a pilot
study. Unpublished report. International Council for Bird Preservation,
Cambridge.
Greathead, D. J. (1988). DipteraiFamily Bombyliidae of Saudi Arabia (Part 2).
Fauna of Saudi Arabia 9: 90-113.
Green, R. E. (1984). The feeding ecology and survival of partridge chicks {Alectoris
rufa and Perdix perdix) on arable farmland in East Anglia. Journai of Applied
Ecology 21: 817-830.
Green, R. E. & Tyler, G. A. (1989). Determination of the diet of the stone curlew
{Burhinus oedicnemus) by faecal analysis. Journai of the Zoological Society of
London 217: 311 -320.
Griffiths, H. I., Davison, A. & Birks, J. (1996). Species réintroductions. Conservation
Biology ^0: 923.
Gross, C. (1987). Mammals of the Southern Gulf. Motivate Publishing, Dubai.
Gross, C. (1997a). Terrestrial mammals. In: Vine, P. J. (ed.) Natural Emirates. Pp.
213-226. Trident Press, London.
Gross, C. (1997b). Terrestrial reptiles and amphibians. In: Vine, P. J. (ed.) Natural
Emirates. Pp. 121-134. Trident Press, London.
Gubin, B. & Mukhina , E. (in prep.). The houbara bustard. Manuscript in Russian,
English translation provided by M. Wilson, Nature Conservation Bureau Ltd.,
Newbury.
Guichard, K. M. (1988a). Hymenoptera:Sphecidae subfamily Sphecinae in Arabia.
Fauna of Saudi Arabia 9:114-131.
Guichard, K. M. (1988b). The genus Pa/arus (Hymenoptera:Sphecidae) in Arabia.
Fauna of Saudi Arabia 9: 132-137.
Guichard, K. M. (1989a). The genus Bemb/x (Hymenoptera: Sphecidae) in Arabia.
Fauna of Saudi Arabia 10:134-151.
Guichard, K. M. (1989b). The genera Stizus and Stizoides (Hymenoptera:
Sphecidae) in Arabia. Fauna of Saudi Arabia 10:152-159.
Haddane, B. (1985). The houbara bustard in Morocco: a brief review. Bustard
Studies 3: 109-111.

188
Hamer, I. (1982). Hymenoptera Highlights 1. Emirates Natural History Group
Bulietin (Abu Dhabi) 18: 16-18.
Hamer, I. (1986a). Hymenoptera Highlights. Emirates Natural History Group Bulletin
(Abu Dhabi) 29: 17-22.
Hamer, I. L. (1986b). Hymenoptera Highlights III Emirates Natural History Group
Buiietin (Abu Dhabi) 30: 12-17.
Hamer, I. L. (1988). Hymenoptera Highlights V. Emirates Naturai History Group
Bulletin (Abu Dhabi) 35: 6-12.
Hamilton, W. J. & Seely, M. K. (1976). Fog basking by the Namib Desert beetle,
Onymacris unguicuiaris. Nature 262: 284-285.
Hanski, I. (1991). Single-species metapopulation dynamics: concepts, models and
observations. Bioiogicai Journai of the Linnaean Society A2: 17-38.
Heim de Balsac, H. (1924). Contributions à l'ornithologie du Sahara central et du
Sud-Algérien. Mem. Soc. d'Hist. nat. Afrique du Nord, Nr.1, Alger.
Heim de Balsac, H. (1926). Contributions à l'ornithologie dans la Sahara
septentrional en Algérie et en Tunisie. Paris.
Heim de Balsac, H. & Heim de Balsac, T. (1954). De l'oued Sous au fleuve
Sénégal. Oiseaux nicheur. Alauda 22: 145-205.
Heim de Balsac, H. & Mayaud, N. (1962). Les oiseaux du Nord-Ouest de l'Afrique.
Paris.
Hellyer, P. (1996). The natural history movement. In: Vine, P. J. (ed.) Natural
Emirates. Pp. 12-20. Trident Press, London, UK.
Hemmingsen, A. M. (1958). Field observations of birds in the Canary Islands.
Vidensk. Medd. Dansk naturh. Foren. 120:189-206.
Hermans, F. (1988a). The Houbara project at the NWRC, Taif, Kingdom of Saudi
Arabia in March. Unpublished report. National Wildlife Research Center, Taif,
Kingdom of Saudi Arabia.
Hermans, F. (1988b). Captive breeding of Bustards at AI Ain Zoo, Abu Dhabi, UAE.
Unpublished report. National Avian Research Center, Abu Dhabi.
Hermans, F. (1992). Captive breeding of Bustards at AI Ain Zoo, Abu Dhabi, UAE.
First report May 1992. Unpublished report. National Avian Research Center,
Abu Dhabi.
Hidalgo de Trucios, S. J. (1990). World status of the great bustard {Otis tarda) with
special attention to the Iberian Peninsula. Miscelianeous Zooiogy\^: 167-180.

189
Hinds, W. T. & Rickard, W. H. (1973). Correlations between climatic fluctuations
and a population of Philolithus densicollis {Horn) (ColeopteraiTenebrionidae).
Journal of Animal Ecology 42: 341 -351.
Hinz, C. and Heiss, E. M. (1989). The activity patterns of houbara bustards: aspects
of a field study in the Canary Islands. Bustard Studies 4: 68-79.
Holdridge, L. A., Grenke, W. C., Hathaway, W. H., Liang, T. & Zosi, J. A. (1971).
Forest environments in tropical life zones. Pergamon Press, Oxford.
Holldobler, B. & Wilson, E. O. (1990). The ants. Springer, Berlin, Heidelberg and
New York.
Holm, E. &. Edney, E. B. (1973). Daily activity of Namib Desert arthropods in
relation to climate. Ecology 54\ 45-56.
Holm E. & Scholtz, C. (1980). Structure and pattern of the Namib Desert ecosystem
at Gobabed. Madoqua 12: 5-12.
Holzel, H. (1982). Insects of Saudi Arabia - NeuropteraiFam. Myrmeleonidae.
Fauna of Saudi Arabia 4: 244-270.
Holzel, H. (1983). Insects of Saudi Arabia - NeuropteraiFam. Myrmeleonidae.
Fauna of Saudi Arabia 5: 210-234.
Holzel, H. (1988). Neuroptera of Arabia - Fam. Sisyridae, Hemerobiidae and
Myrmeleonidae (part 3). Fauna of Saudi Arabia 9: 52-67.
Hooker, T. (1958). Birds seen on the eastern Canary Island of Fuertevertura. ibis
100: 446-449.
Hutto, R. L. (1990). Measuring the availability of food resources. In: M. L. Morrison,
Ralph, C. J., Verner, J. & Jehl, J. R. (eds.) Avian foraging: theory,
methodology and applications. Studies in Avian Biology'll: 20-28. Cooper
Ornithological Society, California.
Irish, J. (1991). Lepismatidae (Thysanura:lnsecta) of the Arabian Peninsula. Fauna
of Saudi Arabia 12: 225-241.
Johnsgard, P. A. (1991). Bustards, hemipodes and sandgrouse. Birds of dry places.
Oxford University Press, Oxford.
Jones, A., Bailey, T. A., Nothelfer, H. B., Gibbons, L. M., Samour, J. H., AI Bowardi
M. & Osborne, P. (1996). Parasites of wild houbara bustards in the United
Arab Emirates. Journai of Helminthology 70: 21-25.
Jongbloed, M. (1987). The living desert. Motivate Publishing, Dubai.
Jongbloed, M. (1997). Plant Life. In: Vine (ed.) Natural Emirates. Pp. 83-94. Trident
Press, London.
Jourdain, F. C. R. (1915). Notes on the bird-life of eastern Algeria, ibis 10:133-169.

190
Kaltenbach, A. (1984) Insects of Saudi Arabia - New species and further records of
Mantodea from Saudi Arabia and Oman. Fauna of Saudi Arabia 6; 203-209.
Kaltenbach, A. (1991). A further contribution to the knowledge of the Mantodea of
the Arabian Peninsula. Fauna of Saudi Arabia 12: 246-256.
Karasov, W. H. (1990). Digestion in birds: chemical and physiological determinants
and ecological implications In: M. L. Morrison, Ralph, C. J., Verner, J. & Jehl,
J. R. (eds.) Avian foraging: theory, methodology and applications. Studies in
Avian Biology 3: 391-415. Cooper Ornithological Society, California.
Kaszab, Z. (1981). Insects of Saudi Arabia - Coleoptera:Fam.Tenebrionidae (part
1 ). Fauna of Saudi Arabia 3: 276-416.
Kaszab, Z. (1982). Insects of Saudi Arabia - Coleoptera:Fam.Tenebrionidae (part
2). Fauna of Saudi Arabia A\ 124-243.
Kendeigh, S. C., Dol'nik, V. R. & Gavrilov, V. M. (1977). Avian energetics. In:
Pinowski, J. & Kendeigh, S. C (eds.) Granivorous birds in ecosystems, their
evolution, populations, energetics, adaptations, impact and control. Pp. 129-
204. Cambridge University Press, Cambridge.
Kirby, J. S. (1997). Influence of environmental factors on the numbers and activity
of wintering lapwings and golden plovers. Bird Study 4: 97-110.
Kostin, V. P. (1956). Report on the mammal fauna of the left bank of the lower
Amu-dar'ya and the Ustyurt together with an account of the distribution of
vertebrate animal species. Trudy Inst. Zoo/. Parasitol. (Tashkent) 8: 7-80.
Krebs, C. J. (1989). Ecological methodology. Harper and Row, New York.
Lack, P. C. (1983). The Canarian houbara: survey results. (Report of the ICBP
Fuerteventura expedition 1979.) Bustard Studies 1: 45-50.
Lamoral, B. H. (1978). Soil hardness, an important and limiting factor in burrowing
scorpions of the genus Opisthophthalmus C. L. Kock, 1837 (Scorpionidae,
Scorpionida). In: Merrett, P. (ed.) Arachnology: Seventh International
Congress. Symposia of the Zoological Society of London No.42. Pp. 171-181.
Larsen, T. B. (1983). Insects of Saudi Arabia - Lepidoptera:Rhopalocera. A
monograph of the butterflies of the Arabian Peninsula. Fauna of Saudi Arabia
5: 333-478.
Launay, F. (1989a). First results of the behaviour of a semi-captive group of
houbara Bustard {Chiamydotis unduiata macqueenii). Quarterly report Spring
1989 (unpublished). National Wildlife Research Center, National Commission
for Wildlife Conservation and Development, Taif, Kingdom of Saudi Arabia.

191
Launay, F. (1989b). Feeding of a semi-captive group of Houbara Bustard. Quarterly
report Spring 1989 (unpublished). National Wildlife Research Center, National
Commission for Wildlife Conservation and Development, Taif, Kingdom of
Saudi Arabia.
Launay, F. (1989c). Behavioural ecology of houbara bustard preliminary report.
Internal report, January to June 1989. (unpublished). National Wildlife
Research Center, National Commission for Wildlife Conservation and
Development, Taif, Kingdom of Saudi Arabia.
Launay, F. & Paillat, P. (1990). A behavioural repertoire of the adult Houbara
bustard {Chiamydotis unduiata macqueenii). Revue d’ Ecologie {Terre Vie) 45:
65-86.
Launay, F., Roshier, D., Loughland, R. & Aspinall, S. J. (1997). Habitat use by
houbara bustard {Chiamydotis unduiata macqueenii) in arid shrubland in the
United Arab Emirates. Journai of Arid Environments 35:111 -121.
Lavauden, L. (1914). Notes sur Houbara unduiata et quelques points relatifs a la
faune ornithologique de Tunisie. L'Oiseau et Rev. France d'Orn. 3: 308-311.
Lavee, D. (1985). The influence of grazing and intensive cultivation on the
population size of the houbara bustard in the northern Negev, Israel. Bustard
Studies 3: 103-108.
Lawton, J. H. (1983). Plant architecture and the diversity of phytophagous insects.
Annual Review of Entomology 28: 23-29.
Lawton, J. H. (1989). Food webs . In: Cherrett, J. M. (ed.) Ecological concepts. Pp.
43-78. Blackwell Scientific Publications, Oxford.
Lawton, J. H. (1996). Population abundances, geographic ranges and conservation:
1994 Witherby Lecture. Bird Study 3\ 3-19.
Levy, G., Amitai, P. & Shulov, A. (1973). New scorpions from Israel, Jordan and
Arabia. Journai of the Linnaean Society {Zoology) 52:113-140.
Linnavuori, R. E. (1986). Heteroptera of Saudi Arabia. Fauna of Saudi Arabia 8: SI-
197.
Louw, G. & Seely, M. (1982). Ecology of desert organisms. Longman Scientific and
Technical, Harlow.
Ludwig, J. A. (1977). Distribution adaptations of root-systems in desert
environments. In: Marshall, E. (ed.) The belowground symposium - a
synthesis of plant-associated processes. Range Science Department Science
Series 26: 85-91. Colorado State University.

192
Luff, M. L. (1968). Some effects of formalin on the numbers of Coleoptera caught in
pitfall traps. Entomologist’s Monthly Magazine 104:115-116.
Luff, M. L. (1975). Some features influencing the efficiency of pitfall trap. Oecologia
19: 345-357.
MacArthur, R. H. & Wilson, E. O. (1967). The theory of island biogeography.
Princeton University Press, New Jersey.
MacKay, W. P. (1991). The role of ants and termites in desert communities. In:
Polis G. A. (ed.) The ecology of desert communities. Pp. 113-150. The
University of Arizona Press, Tucson.
Major, J. D. & Beeston, G. (1996). The biodiversity integrity index; an illustration
using ants in western Australia. Consen/ation Biology W: 65-73.
Major, R. E. (1990). Stomach flushing of an insectivorous birds: an assessment of
differential digestion of prey and risk to birds. Australian Wildlife Research 17:
647-657.
Mandaville, J. P. (1990). Flora of Eastern Arabia. Kegan Paul International and
National Commission for Wildlife Conservation and Development, London and
Riyadh.
Martin, T. E. (1987). Food as a limit on breeding birds: a life-history perspective.
Annual Review of Ecology and Systematlcs 18: 453-487.
Matile-Ferrero, D. (1984). Insects of Saudi Arabia - Homoptera:Suborder
Coccoidea. Fauna of Saudi Arabia 6: 219-229.
Matile-Ferrero, D. (1988). Sternorrhyncha suborder Coccoidea of Saudi Arabia (Part
2). Fauna of Saudi Arabia 9: 23-38.
Maurer, B. A. (1990). Extensions of optimal foraging theory for insectivorous birds:
implications for community structure. In: M. L. Morrison, Ralph, C. J., Verner,
J. & Jehl, J. R. (eds.) Avian foraging: theory, methodology and applications.
Studies in Avian Biology A3: 455-461. Cooper Ornithological Society,
California.
McNamara, J. M. (1990). The policy which maximises long-term survival of an
animal faced with the risks of starvation and predation. Adv. Appi. Prob. 22:
295-308.
Meade-Waldo, E. G. (1889a). Notes on some birds of the Canary Islands. /b /s6 :1-
13.
Meade-Waldo, E. G. (1889b). Further notes on the birds of the Canary Islands, ibis
6: 503-520.

193
Meade-Waldo, E. G. (1890). Further notes on the birds of the Canary Islands. Ibis
7: 429-438.
Meinertzhagen, R. (1954). Birds of Arabia. Henry Sotheran, London.
Meklenburtsev, R. N. (1990). Family Otididae. In: Kashkarov, D. Yu. & Ostapenko,
M. M. (eds.) The birds of Uzbekistan Volume 2. Pp. 7-16. Tashkent.
Mendelssohn, H. (1980). Development of houbara {Chiamydotis unduiata)
populations in Israel and captive breeding. In: Coles, C L & Collar, N.J. (eds.)
The great bustard, Sofia 1978: The houbara bustard, Athens 1979. The Game
Conservancy, Fordingbridge.
Mendelssohn, H., Marder, U. & Stavy, M. (1979). Captive breeding of the houbara
bustard {Chiamydotis unduiata macqueenii) and a description of its display.
internationai Council for Bird Preservation X lil Buiietin 1979. Cambridge.
Mian, A. (1988). A contribution to the biology of the Houbara {Chiamydotis unduiata
macqueenii)’, some observations on the 1983-84 wintering population in
Baluchistan. Journai of the Bombay Natural History Society 85: 9-25.
Mian, A. & Surahio, M. I. (1983). Biology of the houbara bustard {Chiamydotis
unduiata macqueenii) with reference to western Baluchistan. Journai of the
Bombay Naturai History Society 80: 11-118.
Miller, A. G. & Cope, T. A. (1996). The flora of the Arabian Peninsula and Socotra.
Volume 1. Edinburgh University Press, Edinburgh.
Mirza, Z. B. (1971). Preliminary report on the study of the population, mortality, sex
ratio, age structure and food habits of the houbara bustard, in Cholistan
desert, 14th January-24th January 1971. Unpublished report.
Mirza, Z. B. (1985). A note on the Houbara Bustards in Cholistan, Punjab. Bustard
Studies 3: 43-44.
Mitchell, W. (1995). Invertebrate culturing and feeding trials with captive houbara
bustards. Unpublished student placement report 1994-95. Nationai Avian
Research Center Internai Report Series 26. National Avian Research Center,
Abu Dhabi
Molchanov, L. A. (1913). On the birds of Aybugir (Über die Vogel von Ajbugir: von L.
A. Moltschanow). Pp.151-7.
Mooney, H. A. & Drake, J. A. (eds.) (1986). Ecology of biological invasions of North
America and Hawaii. Springer Publications, New York.
Mordue, W. Goldsworthy, G. J., Brady, J. & Blaney, W. M. (1980). Insect
physiology. Blackwell Scientific Publications, Oxford.

194
Moreby, S. J. (1988). An aid to the identification of arthropod fragments in the
faeces of gamebird chicks (Galliformes). Ibis 130: 519-526.
Morris, R. P. (1991). A literature review of the houbara bustard Chiamydotis
unduiata. Unpublished report. National Avian Research Center Internal Report
Series 2, Abu Dhabi.
Muirhead-Thompson, R. C. (1991). Trap responses of flying insects. Academic
Press, London.
Muma, M. (1980). Comparison of three methods for estimating solpugid (Arachnida)
populations. Journai of Arachnoiogy Q\ 267-270.
Nagy, K. A. (1983). The doubly-labelled water method - a guide to its use.
University of California, California.
Naidu, S. G. & Hattingh, J. (1988). Water balance and osmoregulation in
Physadesmia giobosa, a diurnal tenebrionid beetle from the Namib Desert.
Journai of Insect Physiology 34: 911 -917.
Nazarov, A. P. (1992). Experience of keeping houbara bustards in captivity. Bustard
Studies 5:196-197.
Norton, J. (1995). Methods for analysis of plant remains in houbara faeces and
preparation of reference collection materials. Unpublished report. National
Avian Research Center Internai Report Series No. 23, Abu Dhabi.
Noy-Meir I. (1974). Desert ecosystems: higher trophic levels. Annual Review of
Ecoiogy and Systematics 5:195-213.
Nutting, W. L., Haverty, M. I. & La Page, J. P. (1987). Physical and chemical
alteration of soil by two subterranean termite species in Sonoran Desert
grassland. Journal of Arid Environments 12: 233-239.
Oatham, M. (1996). Restoring habitats for the houbara bustard in Abu Dhabi. In:
Osborne, P. E. (ed.) Desert ecology of Abu Dhabi-a review and recent
studies. Pp. 66-85. Pisces Publications, Newbury.
Osborne, P. E. (1991). Preliminary field survey of the houbara bustard in Abu
Dhabi, United Arab Emirates. Phase 1 - survey of wintering houbara bustards
in Abu Dhabi October-December 1991. Unpublished report. Nationai Avian
Research Center Internal Report Series 4. National Avian Research Center,
Abu Dhabi.
Osborne, P. E. (1992). Preliminary field survey of the houbara bustard in Abu
Dhabi, United Arab Emirates. Phase 2 - survey of potential breeding habitat in
Abu Dhabi, February-March 1992. Unpublished report. National Avian

195
Research Center Internal Report Series 5. National Avian Research Center,
Abu Dhabi.
Osborne, P. E. (1993). Teratoscincus scincus - the gecko with large reflective eyes.
Tribulus 4: 23.
Osborne, P. E. (1996a). The houbara bustard in Arabia with special reference to
Abu Dhabi. In: Osborne, P. E. (ed.) Desert ecology of Abu Dhabi-a review and
recent studies. Pp. 86-97. Pisces Publications, Newbury.
Osborne, P. E. (1996b). Introduction. In: Osborne, P. E. (ed.) Desert ecology of Abu
Dhabi-a review and recent studies. Pp. 2-15. Pisces Publications, Newbury.
Osborne, P. E. (ed.) (1996c). Desert ecology of Abu Dhabi-a review and recent
studies. Pisces Publications, Newbury.
Osborne, P. E. & Hornby, R. J. (1995). A proposal to create managed Resource
Protected Areas for houbara bustard in Abu Dhabi. Unpublished report.
National Avian Research Center, Abu Dhabi.
Osborne, P. E., AI Bowardi, M. & Bailey, T. A. (1997a). Migration of the Houbara
Bustard, Chiamydotis unduiata from Abu Dhabi to Turkmenistan; the first
results from satellite tracking studies. Ibis 139: 192-196.
Osborne, P. E., Collar, N. & Goriup, P. (1984). Bustards. Dubai Wildlife Research
Centre and the Cotswold Printing Company, UK.
Osborne, P. E., Launay, F. & Gliddon, G. (1997b). Wintering habitat use by houbara
bustards Chiamydotis unduiata in Abu Dhabi and implications for
management. Biological Consen/ation 8^: 51-56.
Osborne, P. E., Norton, J. A. & Aspinall, S. (1996). Desert birds of Abu Dhabi. In:
Osborne P. E. (ed.) Desert ecology of Abu Dhabi- a review and recent
studies. Pisces Publications, Newbury.
Owen, J. A. (1993). Use of a flight interception trap in studying the beetle fauna of a
Surrey woodland over a three year period. The Entomologist'\A2:141-155.
Owen, M. (1975). An assessment in faecal analysis technique in waterfowl feeding
studies. Journal of wildlife Management 39: 271-279.
Paillat, P. (1987). Houbara and wildlife investigation in Pakistan, winter 1987.
Unpublished report. National Wildlife Research Center, Taif, Kingdom of
Saudi Arabia.
Pavlenko, T. A. (1962). Vertebrate animals of the Golodnaya steppe. In: The animal
fauna of the Golodnaya steppe. Pp. 127-75. Tashkent.
Peet, R. K. (1974). The measurement of species diversity. Annual Review of
Ecology and Systematics 5: 285-307.

196
Perfecto, I. & Snelling, R. (1995). Biodiversity and the transformation of a tropical
agroecosystem- ants in coffee plantations. Ecological Applications 5 :1084-
1097.
Perfecto, I. & Vandermeer, J. (1996). Microclimatic changes and the indirect loss of
ant diversity in a tropical agroecosystem. Oecologia 108: 577-582.
Piechocki, R. (1968). Beitrâge zur Avifauna der Mongolei. Tell 1. Non-
Passeriformes. Mitt. Zoo/. Mus. Berlin 44:149-292.
Pimm, S. L. (1982). Food webs. Chapman & Hall, New York.
Pimm, S. L, Lawton, J. H. & Cohen, J. E. (1991). Food web patterns and their
consequences. Nature 350: 669-674.
Platt, J. B. (1985). Houbara bustard research in Dubai, United Arab Emirates.
Bustard Studies 3:101-102.
Polatzek, J. (1909). Die Vogel der Canaren. Orn. Jahrb. 20:1-24.
Polls, G. A. (ed.) (1990a). Biology of scorpions. Stanford University Press,
California.
Polls, G. A. (1990b). Ecology. In: Polls, G. A. (ed.) Biology of scorpions. Stanford
University Press, California.
Polls, G. A. (1991a). Desert communities: an overview of patterns and processes.
In: Polls, G. A. (ed.) The ecology of desert communities. Pp. 1-26. The
University of Arizona Press, Tucson.
Polls, G. A. (1991b). Food webs in desert communities: complexity via diversity and
omnivory. In: Polls, G. A. (ed.) The ecology of desert communities. Pp. 383-
439. The University of Arizona Press, Tucson.
Polls, G. A. (ed.) (1991c). The ecology of desert communities. The University of
Arizona Press, Tucson.
Polls, G. A. (1991d). Complex trophic interactions in deserts - an empirical critique.
American Naturalist A38\ 123 -155.
Polls, G. A. & Yamashita, T. (1991). The ecology and importance of predaceous
arthropods in desert communities. In: Polls, G. A. (ed.). The ecology of desert
communities. Pp. 180-222. The University of Arizona Press, Tucson.
Ponomareva, T. S. (1982). Distribution, nesting biology and prospects for captive
breeding of the Houbara bustard in USSR. Acta XVIII Congr. Int. Orn. 2:
1159-60.
Pont, A. C. (1991). A Review of the Fanniidae and Muscidae (Diptera) of the
Arabian Peninsula. Fauna of Saudi Arabia 12: 312-365.

197
Popov, G. B. (1980). Acridoidea of Eastern Arabia. The scientific results of the
Oman flora and fauna survey 1977 (Dhofar). Journal of Oman Studies.
Special Report 2: 113-149.
Popov, G. B. (1981). Insects of Saudi Arabia - Orthoptera Fam:Tettigoniidae
(bushcrickets) Fauna of Saudi Arabia 3\ 114-148.
Pyke, G. H. (1984). Optimal foraging theory: a critical review. Annual Review of
Ecoiogy and Systematics 15: 523-572.
Ralph, 0. P., Nagata, 8. E. & Ralph, 0. J. (1985). Analysis of droppings to describe
diets of small birds, Journai of Field Ornithology 56 :165-174.
Ramadan-Jaradi, G. & Ramadan-Jaradi, M. (1989). Breeding the houbara bustard
at the AI Ain Zoo and Aquarium Abu Dhabi, UAE. Zoo/. Garten N.F. 59: 229-
240.
Redford, K. H. & Dorea, J. G. (1984). The nutritional value of invertebrate with
emphasis on and ants and termites as food for mammals. Journai of the
Zoological Society of London 203: 385-395.
Remple, D. & Gross, 0. (1993). Falconry and birds of prey in the Gulf. Motivate
publications, Dubai.
Richards, O. W. (1984a). Insects of Saudi Arabia - Hymenoptera:Fam. Masaridae
(The Arabian species). Fauna of Saudi Arabia 6: 413-422.
Richards, O. W. (1984b). Insects of Saudi Arabia - Hymenoptera:Fam. Vespidae
(social wasps of the Arabian Peninsula). Fauna of Saudi Arabia 6: 423-440.
Richards, O. W. & Davies, R. G. (1980). Imms' general textbook of entomology.
Volume 2 - classification and biology. Tenth edition. Chapman & Hall, London.
Richardson, 0. (1990). The birds of the United Arab Emirates. Hobby Publications,
Dubai and Warrington.
Richardson, 0. (1997). Birds and birdwatching. In: Vine, P. J. (ed.) Natural
Emirates. Pp. 149-166. Trident Press, London.
Ricklefs, R. E. (1996). Avian energetics, ecology and evolution. In: Carey, C. (ed.)
avian energetics and nutritional ecology. Pp. 1-30. Chapman & Hall, New
York.
Robel, J. R., Press, B. M., Henning, B. L., Johnson, K. W., Blocker, H. D. & Kemp,
K. E. (1995). Nutrient and energetic characteristics of sweepnet-collected
invertebrates. Journai of Field Ornithology 66: 44-53.
Roberts, A. (1848). First record of houbara bustard in Britain. Zoologist 1969-2065.
Roberts, T. J. (1991). The birds of Pakistan. Oxford University Press, Oxford.

198
Roberts, T. J. & Savage, C. D. W. (1971). Houbara bustard {Chiamydotis unduiata
macqueenii) and its management. The Outdoorman - April 1971: 37-39.
Roche, C. G. (1981). Notes on the bees and wasps of UAE. Emirates Natural
History Group Buiietin (Abu Dhabi) 15: 12-20.
Rosenberg, K. V. & Cooper, R. J. (1990). Approaches to avian diet analysis. In: M.
L. Morrison, Ralph, 0. J., Verner, J. & Jehl, J. R. (eds.) Avian foraging: theory,
methodology and applications. Studies in Avian Biology 80-90. Cooper
Ornithological Society, California.
Roshier, D. A., Boer, B. B. & Osborne, P. E. (1996). Vegetation of Abu Dhabi and a
preliminary classification of its plant associations. In: Osborne, P. E. (ed.)
Desert ecology of Abu Dhabi - a review and recent studies. Pp. 50-65. Pisces
Publications, Newbury.
Rustamov, A. K. (1954). The birds of the Kara-Kum desert. Ashkhabad.
Saint Jaime, M., Combreau, O., Seddon, P., Paillat, P., Gaucher, P. & van Heezik,
Y. (1996). Restoration of Chiamydotis unduiata macqueenii populations in
Saudi Arabia- a progress report. Restoration Ecology^: 81-87.
Salikhbaev, K. S. & Ostapenko, M. N. (1967). Birds. In: Ecology, conservation and
wise use of the vertebrates of the Karshirskaya steppe. Pp. 76-128. Tashkent.
Samarin, E. G., Saraev, F. A., & Bidashko, F. G. (1986). Short communications on
the houbara bustard. In: Rare animals of Kazakhstan. Pp.79-86. Alma-Ata.
Satchell, J. E. (1978), Ecology and environment in the United Arab Emirates.
Journai of Arid Environments 1: 201 -226.
Satchell, J. E., Mountford, M. D. & Brown, W. M. (1981). A land classification of the
United Arab Emirates. Journai of Arid Environments 4: 275-285.
Savory, C. J. (1974). The food of red grouse chicks Lagopus L scoticus. /b/s 119:
1-9.
Savory, C. J. (1989). The importance of invertebrate food to chicks of gallinaceous
species. Proceedings of the Nutrition Society 113-133.
Schulz, M. & Schulz, H. (1991). Working bibliography of the bustards (Otididae).
National Wildlife Research Center, Taif, Saudi Arabia.
Schulze, R. E. & McGee, O. S. (1978). Climatic indices and classifications in
relation to the biogeography of Southern Africa. In: Werger, M. J. A. (ed.)
Biogeography and ecology of Southern Africa. Pp. 19-52. Junk, The Hague.
Scott, J. M. & Carpenter, J. W. (1987). Release of captive reared or translocated
birds. What do we need to know? Au/c 104: 544-545.

199
Seddon, P & van Heezik, Y. (1996). Seasonal changes in houbara bustard numbers
in Harrat al-Harrah Saudi Arabia- implications for managing a remnant
population. Biological Conservation 75 :139-146.
Seddon, P. J., Saint Jaime, M., van Heezik, Y., Paillat, P., Gaucher, P. &
Combreau, O. (1995). Restoration of houbara bustard populations in Saudi
Arabia: developments and future directions. Oryx 29 :136-142.
Seely, M. K. (1979). Irregular fog as a water source for desert dune beetles.
Oecologia 42: 213-227.
Seely, M .K. (1991). Sand dune communities. In: Polis, G. A. (ed.) The ecology of
desert communities. Pp. 348 -382. The University of Arizona Press, Tucson.
Seely, M. K. & Louw, G. N. (1980). First approximation of the effects of rainfall on
the ecology and energetics of a Namib Desert dune ecosystem. Journal of
Arid Environments 3: 25-54.
Shaw Reade, S. N., Sale, J. B., Gallagher, M. D. & Daly, R. H. (eds.) (1980). The
scientific results of the Oman flora and fauna survey 1977 (Dhofar). Journal of
Oman Studies Special Report 2.The Cray Press Ltd, Sidcup.
Sheldon, J. J. & Rogers, L. E. (1984). Seasonal and habitat distribution of
tenebrionid beetles in shrub-steppe communities of the Hanford Site in
eastern Washington. Environmental Entomology 3: 214-220.
Shepley, M. (1997). In: Vine, P. J. (ed.) Natural Emirates. Pp. 21-40. Trident Press,
London.
Simard, J. M. & Watt, D. D. (1990). Venoms and toxicity. In: Polls, G. A (ed.) The
biology of scorpions. Pp. 414-444. Stanford University Press, California.
Simpson, B. B. (ed.) (1977). Mesquite: its biology in two desert scrub ecosystems.
Dowden, Hutchinson & Ross, Stroudsburg, USA.
Sissom, W. D. (1990). Systematics, biogeography and paleontology. In: Polls, G. A
(ed.) The biology of scorpions. Pp. 64-160. Stanford University Press,
California.
Skutelsky, O. (1996). Predation risk and state-dependent foraging in scorpions -
effects of moonlight on foraging in the scorpion Buthus occitanus. Animal
Behaviour 52: 49-57.
Snedecor, G. W. & Cochran, W. G. (1967). Statistical methods. Iowa State
University Press, Iowa.
Southwood, T. R. E. (1978). Ecological methods. Second edition. Chapman & Hall,
London.

200
Stephens, D. W. (1990). Foraging theory: up, down and sideways. In: M. L.
Morrison, Ralph, C. J., Verner, J. & Jehl, J. R. (eds.) Avian foraging: theory,
methodology and applications. Studies in Avian Biology “\3: 444-454. Cooper
Ornithological Society, California.
Stephens, D. W. & Krebs, J. R. (1986). Foraging theory. Princeton University Press,
New Jersey.
Storr, G. M. (1961). Microscopic analysis of faeces, a technique for ascertaining the
diet of herbivorous mammals. Australian Journal of Biological Science 14:
157-164.
Strong, D. R., Lawton, J. H. & South wood, T. R. E. (1984). Insects on plants -
community patterns and mechanisms. Harvard University Press, Cambridge,
USA.
Summers, R. W., Stansfield, J., Perry, S. Atkins, C & Bishop, J. (1993). Utilization,
diet and diet selection by brent geese Branta bernlcia bernicia on salt-
marshes in Norfolk. Journal of the Zoological Society of London 231: 249-273.
Surahio, M. I. (1985). Ecology and distributions of houbara bustards in Sind.
Bustard Studies 3: 55-58.
Sushkin, P. P. (1908). The birds of the central Kirgiz steppe (Turgay region and the
eastern part of Ural'sk region). Pp. 240-5. Moscow.
Sutherland, W. J. (1998). Managing habitats and species. In: Sutherland, W. J.
(ed.) Conservation science and action. Pp. 202- 219. Blackwell Science Ltd.,
Oxford.
Symens, P. (1987). Houbara bustards in semi-captivity April-June. Quarterly Report,
Spring 1987 (unpublished). National Wildlife Research Center, Taif, Kingdom
of Saudi Arabia.
Symens, P. (1988a). Influence of the mass migration of migratory locusts
{Schistocerca gregaria) on birds in the Taif area. Quarterly Report, Winter
1988 (unpublished). National Wildlife Research Center, Taif, Kingdom of
Saudi Arabia.
Symens, P. (1988b). Houbara Bustard survey in Harrat al-Harrah, March 1988
report (unpublished). National Wildlife Research Center, Taif, Kingdom of
Saudi Arabia.
Thalen, D. C. P. (1979). Ecology and utilisation of desert shrub rangelands in Iraq.
Dr. W. Junk, B. V. The Hague
Thesiger, W. (1959). Arabian Sands. Harper Collins, London.

201
Thomas, D. B. (1979). Patterns in the abundance of some tenebrionid beetles in the
Mojave desert. Environmental Entomology S: 568-574.
Thomas, D. B. (1983). Tenebrionid beetle diversity and habitat complexity in the
eastern Mojave Desert. The Coleopterlst's Bulletin 37 :135-147.
Tigar, B. J. (1996a). A preliminary assessment of the arthropods of Abu Dhabi. In:
Osborne, P. E. (ed.) Desert ecology of Abu Dhabi- a review and recent
studies. Pp. 172-195. Pisces Publications, Newbury, UK.
Tigar, B. J. (1996b). Terrestrial Arthropods (excluding insects). In: Vine, P. J. (ed.)
Natural Emirates. Pp. 107-120. Trident Press, London, UK.
Tigar, B. J. & Collingwood, C. A. (1993). Some new ant records for Abu Dhabi
emirate. TribulusZ: 13.
Tigar, B. J. & Osborne, P. E. (1997). Patterns of arthropod abundance and diversity
in an Arabian desert. Ecography 20: 550-558.
Tinbergen, L. (1960). The natural control of insects in pinewoods. I factors
influencing the intensity of predation by songbirds. Arch. NeeriZool. 13: 265-
343.
Toschi, A. (1969). Introduzione alia ornithologia della Libia. Lab. Zoo/. Appl. Caccia
y Bologna VI: 191 -192.
Upton, M. S. (1991). Methods for collecting, preserving and studying insects and
allied forms. Fourth edition. Australian Entomological Society Miscellaneous
Publication 3. Brisbane.
Uvarov, B. P. (1952). Studies in the Arabian Orthoptera III - new genera, species
and subspecies, collected by the anti-locust missions. Journal of the LInnaean
Society {Zoology) 42: 176-194.
Uvarov, B. P. (1966). Grasshoppers and Locusts: A Handbook of General
Acridology Volume I. Anti-Locust Research Centre, Cambridge
Uvarov, B. P. (1977). Grasshoppers and locusts - a handbook of general acridology.
Volume II - Behaviour, ecology, biogeography, population dynamics. Anti-
Locust Research Centre, Cambridge.
Vachon, M. (1979). Arachnids of Saudi Arabia - Scorpiones. Fauna of Saudi Arabia
1: 30-66.
Vachon, M. (1980). Arachnids of Saudi Arabia. Fauna of Saudi Arabia 1: 30-66.
Valverde, J. A. (1957). Aves del Sahara Espahol. Institute de Estudios Africanos.
Consejo Superior de Investigaciones Cientificas, Madrid.
van Heezik, Y. M. & Seddon, P. J. (in press) Seasonal changes in habitat use by
houbara bustards in Northern Saudi Arabia. Condor.

202
Vasquez, R. A. (1994). Assessment of predation risk via illumination level -
facultative central place foraging in the cricetid rodent Phyllotis darwini.
Behavioural Ecology and Socloblology 34: 375-381.
Viitala, J., Korpimali, E. Palokangas, P. & Koivula, M. (1995). Attraction of kestrels
to vole scent marks visible in ultraviolet light. Nature 323: 425-427.
Vine P. J. (ed.) (1997). Natural Emirates. Trident Press, London.
Volsoe, H. (1951). The breeding birds of the Canary islands. 1 Introduction and
synopsis of the species. VIdensk. Medd. fra Dansk naturh. Foren. 113:1-153.
von Thanner, R. (1912). Von den Kanaren. Orn. Jahrb. 22: 221-228.
von Thanner, R. (1913). Wild und Jagd auf den Kanaren. Deutsche Jager-Zeitung,
61: 569-572, 586-589, 600-604, 616-618 & 631-633.
Wagge, B. E. (1985). Trapping efficiency of carabid beetles in glass and plastic
pitfall traps containing different solutions. Fauna Norvegica Series B 32: 33-
36.
Walker, D. H. & Pittaway, A. R. (1987). Insects of Eastern Arabia. Macmillan
Publishers Ltd, London and Basingstoke.
Ward, 0. R., O’Brien, 0. W., O’Brien, L. B., Foster, D. E. & Huddleston, E. W.
(1977). Annotated checklist of new world insects associated with Prosopis
(Mesquite). U.S. Department of Agriculture Technical Bulletin 1557.
Ward, D. (1991). A test of the ‘maxithermy’ hypothesis with three species of
tenebrionid beetles. Journal of Arid Environments 21: 727-738.
Waterston, A. R. & Pittaway, A. R. (1989). The Odonata or dragonflies of Oman and
neighbouring territories. Journal of Oman Studies 10:131-168.
Webb, P. B. & Berthelot, S. (1836-44). Histoire naturelle des Iles Canaries, 2, pt.ll.
Zoologie. Paris.
Western, R. A. (1989). The flora of the United Arab Emirates - an introduction.
United Arab Emirtes University, AI Ain.
Westoby, M. (1978). What are the biological bases of varied diets? American
Naturalists'll: 627-631.
Wharton, R. A. & Seely, M. (1981). Species composition of and biological notes on
Tenebrionidae of the lower Kuiseb River and adjacent plains. Madoqua 13: 5-
25.
Whitaker, J. I. S. (1905). The Birds of Tunisia 2. London.
Wiens, J. A. (1976). Population responses to patchy environments. Annual Review
of Ecology and Systematlcs 7:81-120.

203
Wiens, J. A. (1985). Vertebrate responses to environmental patchiness in arid and
semiarid ecosystems. In: Pickett, S. T. A. & White, P. S. (eds.). The ecology
of natural disturbance and patch dynamics. Pp. 169-193. Academic Press,
New York.
Wiens, J. A. (1991). The ecology of desert birds. In: Polis, G. A. (ed.) The ecology
of desert communities. Pp. 278-310. The University of Arizona Press, Tucson.
Wigglesworth, V. B. (1972). The principles of insect physiology. Seventh edition.
Chapman & Hall, London.
Wilkinson. L. (1993). SYSTAT 5.02 for Microsoft Windows. SYSTAT Inc., Illinois,
USA.
Williams, D. F. (ed.) (1994). Exotic ants - biology, impact and control of introduced
species. Westview Press, Boulder Colorado.
Wilson, E. O. (1992). The diversity of life. Penguin Books, London
Wiltshire E. B. (1983). Insects of Saudi Arabia - Lepidoptera:Fam. Cossidae,
Sphingidae, Thyretidae, Geometridae, Lymantriidae, Artiidae, agaristidae,
Noctuidae, Ctenuchidae (Part 3). Fauna of Saudi Arabia 5: 293-332.
Wiltshire E. B. (1986). Insects of Saudi Arabia - Lepidoptera:Fam. Cossidae,
Sesiidae, Metabelidae, Lasiocampidae, Sphingidae, Geometridae,
Lymantriidae, Arctiidae, Nolidae, Noctuidae (Heterocera); Fam. Satyridae
(Rhopalocera) (Part 4). Fauna of Saudi Arabia 8: 262-323.
Wiltshire E. B. (1988). Lepidoptera of Saudi Arabia - Fam. Metabelidae,
Geometridae, Lymantriidae, Arctiidae, Argastidae, Noctuidae (Part 6). Fauna of Saudi Arabia 9: 68-82.
Wiltshire, E. B. (1990). An illustrated, annotated catalogue of the Macro-Heterocera
of Saudi Arabia. Fauna of Saudi Arabia 11: 91-250.
Windingstad, R. M., Hurley, S. S. & Sileo, L. (1983). Capture myopathy in a free-
living greater sandhill crane from Wisconsin. Journal of Wildlife Diseases 19:
289-290.
Wisdom, C. (1988). Comparisons of insect use and chemical defence patterns of
two Sonoran desert shrubs. In: R. G. Zohary (ed.) Desert ecology 1986 - a
research symposium. Pp. 39-49. Southern California Academy of Sciences
and the Southern Desert Studies Consortium.
Wisdom, C. (1991). Herbivorous Insects. In: Polls, G. A. (ed.) The ecology of desert
communities. Pp. 151-179. The University of Arizona Press, Tucson.
Wise, D. H. (1993). Spiders in ecological webs. Cambridge University Press,
Cambridge.

204
Witherby, H. F., Jourdain, F. C. R., Ticehurst, N. F. & Tucker, B. W. (1943).
Handbook of British birds, Volume 5. H. F. & G. Witherby, London.
Wolda, H. (1990). Food availability to an insectivore and how to measure it. In: M. L.
Morrison, Ralph, C. J., Verner, J. & Jehl, J. R. (eds.) Avian foraging: theory,
methodology and applications. Studies in Avian Biology 3: 38-43. Cooper
Ornithological Society, California.
Zachariassen, K. E., Andersen, J., Maloiy, M. O. & Kamau, J. M. Z. (1987).
Transpiratory water loss and metabolism of beetles from arid areas in East
Africa. Comparative Biochemistry and Physiology S6A: 403-408.
Zak, J. C. & Freckman, D. W. (1991). Soil communities in deserts - microarthropods
and nematodes. In: Polis, G. A. (ed.) The ecology of desert communities. Pp.
55-88. The University of Arizona Press, Tucson.
Zhuyuko, B. P. (1986). Short communications on the houbara bustard. In: Rare
animals of Kazakhstan. Pp. 79-86. Alma-Ata.
Zohary, M. (1973). Geobotanical foundations of the Middle East. Volumes 1 & 2.
Gustav Fischer Verlag, Stuttgart.

205
Appendix 1.1
A preliminary assessment of the arthropods of Abu Dhabi
by Tigar, B. J. (1996a).
In: Osborne, P. E. (ed) Desert ecology of Abu Dhabi- a review and recent studies,
pp 172-195. Pisces Publications, Newbury, UK.

RECORDS OF ARTHROPODS occurring in the UAE, particularly Abu Dhabi, are very few, rcHecting the general lack of knowledge of invertebrates in the Middle East. This makes it impossible to compile an authoritative account of the arthopods of Abu Dhabi at the current time. The majority of records that have been published within the last IS years are based on data collected during scientific expeditions to the neighbouring countries of the Kingdom of Saudi Arabia and the Sultanate of Oman. Where no records exist for UAE, these data have been used to suggest which arthropods may occur in Abu Dhabi based on their occurrence in similar habitats elsewhere.
The taxa that occur most frequently in the desert ecosystem and form the highest biomass are from the classes Arachnida and Insecta. The arachnids are mainly represented by the orders Scorpiones, Solifugae and Araneae, and the insects by the orders Colcoptera (especially the Tenebrionidae and Scarabaeidae), Hymenoptcra (family Formicidae) and the Orthoptera. The ability of desert arthropods to survive the harsh climatic conditions prevailing in Abu Dhabi is discussed in general terms but there remain many arthropods that play an important role in the desert ecosystem about which little is known.
206
C H A P T E R 10
A preliminary assessment of
the arthropods of Abu Dhabi
b y B a rb a ra J T ig a r
^ J. C.Ijuyi yjj ysjiuNij»
Vita uu to Ji«'i 11 jM ÿ xic. . i j f i jUc. tikL, J vjdl ÂSLmB (JSJ Âj<Jc j
y d ij ^ .-iiji.a .» j c jKiL-su i i* ca vu '.a v j v jo i ^ > V '
.s jjV a JXÏ Ih ^1 am ^ u v i> ^ .u, ^ u j ^ j
U iü ijjS; J ji J (Jij tàJfJi* ijj*i j j L i J v - j j v - l iV l ^ UjItM cL u J mS . Insecta j A rachn ida
kUjiiJi J l û j .A raneae j Solifugae Vho* 'Scorp iones *— etc) *— Â jl— i t '(S carabaeidae j T e n e b rio n id a e lÿJ ï—c. ..a lt) C o leo p tera
V j l * SÜ jL<U«> • j x i t-.lc . uilLü O rth o p tera j (F o rm ic id aeI j j j ^ 0- i j £ . J-Ijd ^ j s i j ^ J i M
• lWîÏ t»je- ' j j s ijjS j q jija—aï Âjjjï î-jL ia uik
. I t irti.s l/.H- / i i i i i i c / / s / i i i lc iy / i \ - t i l l \ locilhccl. t m d i h t y irc rc ' ci i i i i i in >ii in t i l l h u t th e
m ost e ir id / ) / t ic i ’s. 7J.H'\' i r o n ’ as m u c h as th re e in c h e s tien iss . i r i t h h n ir y . rec id is h /e,iis. e in d
/H 'n d n lo n s h o i/ie s ... / s n i r o n e ... n n d t r ie d to h i l l it h n t it esen /H 'd . .1 l i t t le I t i t e r h n y
e o in /x in io n l b in K d h in n t ie h le i l th e h n c h o f m y neeh . t t i i i l th tn h in a it it'its th is .s f id e r /
in m { x ‘d e o n r n ls i r e ly n m l zr/wc/ m y ten . " W'li.i Ki II I'l IIMl ,i-1< I I 04V ).
1 7 2

■\ I ’ KI-'I I M I \ M n W",! SS\ l | : \ r | ()l- I HI A K I I I K U H O D S ( ) r \ lU DHABI
- 207 -
E n t o m o l o g y in A rab ia
IU irtiker( IVSO) has rc\ icw c il rhe work ol I he pioneering explorers w ho made mention o f the fanna and flora o f the Arabian Peninsula in their aeeoiints. The first o f these was Ihn Battuta, a merchant, scientist and Islamic pilgrim from Tangier, w ho tra\ elled extensi\e ly in the Middle and Far East as well as Russia laetween 1525 and 135.5. H ow e\ er. the most notable from the point o f \ iew o f collecting and recording art hropods were St John Philby. Bertram 'I'homas and W ilfred Thesiger. P iiilby made many expeditions to the Arabian Peninsula from 1917 until 1953 and collected a considerable qnantity o f material that is hon.sed in the Natural H isto iy Museum in London. The material has been poorly w orked. howe\ er. and no com plete lists or articles ha\ e e\ er been ])tiblished from his specimens. Similarly. Thomas collected in,sect material fo r the Natural History Museum mainly during 1931 but this also remains little worked. The famous explorer W ilfred Thesiger tra\ elled extensi\ ely in the reg ion between 19a6 and 1948 and made many o b se rva tions on the flo ra and fauna, in c lu d in g in \ ertebrates (Thesiger 1959). Indeed, his first cro.ssingof the Rufi al Khali was made possible by a request from O.B. Lean, de.sert locust specialist o f the Foocl and A gricu ltu re O rganisation, that he docum ent locust breeding sites in Arabia. Since the 194Os. most o f the entom ological w ork in the M iddle Fast has been o f an applied nature dealing w ith specific pest problems, in particular mosquitoes and related malaria and locust outbreaks, la ir exam ple. Ta lhouk ( 1969) lists the agricultural ; rests o f the G u lf region.
In the late 1970s. \ arious .scientific expeditions were carried out to some relativ ely unknow n regions o f both the K ingdom o f Saudi Arabia and the Sultanate o f Oman (see B iittike r 1980. Shaw Reader’/r//. 1980. O ullon 1988). These have encouraged further .scientific research and have highlighted the interesting fauna and flora that occur in the Arabian I’eninsula and the lack o f know ledge concerning the liio logy o f the region. They also show the potential for uncovering new records and species as yet unknow n to .science. In [xirticular. Prof. W. B iittike r and Michael Gallagher have endeavoured to ensure not only that valuable biological material is collected and recorded but also that it reaches experts who are a file to study and identify it or describe new species in the scientific literature.
Despite the interest in ne ighbouring countries, ver\ little entom ological w ork has been carried out in the UAE. In consequence, this chapter is not an authoritative account o f the arthropods o f Abu Dhabi but rather an in troduction to their occurrence and abundance, ba.sed on the relatively few records that exist. The Emirates Natural H isto iy G roup have provided some o f the more recent and specific records for UAE. although much of
their material has come from the less repre.sentative habitais, such as w adis and gardens, where the availability o f water and exotic vegetation allows many non-de.sert specialists to become numerous. The true de.serts o f Abu Dhabi have, largely, been ignored. The potential fo r the d iscoveiy o f unrecorded species is high, since not only is there a paucity o f records but also LLA.E lies at the meeting point o f three biogeographical zones, i.e. noithern Palaearctic. African and Asian. The National Av ian Research Center ( NARC) in A fiu Dhabi is collecting and identify ing desert arthropods as part o f its research on the feeding ecology o f the houbara bustard Cblaiiiyciotis iin d u la ia {.see Chapters I, 5 and 6). These records w ill make a valuable contribu tion to the entom ological know ledge o f the Emirate. Records in this review that are not a.scribed to a published source are the first results from the w ork o f NARC.
Review of the arthropods occurring in Abu Dhabi
The phylum Arthropoda denotes a major taxonomic d ivision that consists o f invertebrates characterised by having jointed liml.xs and an exoskeleton. In particular, the SLibphyla Chelicerata and Mandibulata are well represented in desert environs such as Abu Dhabi Emirate (see Barnes 1974 for a fu ll cla.ssification of invertebrates). The form er includes many arid-zone .specialists o f the class Arachnida such as the orders Scorpiones (scorpions), Solifugae (solifugids or camel spiders). Araneae (spiders) and Acarina (mites and ticks). The M andibtila ia includes the classes Insecta (insects), Crustacea (crustaceans) and the m yriapodous or m anylegged Chilopoda (centipedes). By far the most diverse and abundant o f these groups is the in.sects which contains more members than any other single animal group w orldw ide. In contrast, the Crustacea are only repre.sented in the deserts o f Abu Dhal.ii by a single order. Isopoda ( (he isopods ). since the majority o f their members have an aquatic lifestyle
S c ih p h y lu m C h e licc fn taCla.s.-s A r a c h n id aO r d e r .S c o r p io n e s ( s c o r p io n s )
Scorpions have existed on the earth for over 4ÜÜ m illion years and present-day scorpions show- both prim itive and advanced features. This com bination gives them an overall plasticity in response to their environm ent that allows them to survive in the most arduous conditions, in c lud in g desert eco.systems. For exam ple, desert scorpions are able to w ithstand tempeiatures .sex'eral degrees higher than most other de.sert arthropods (see Polis I99(.) for a full description o f .scorpion bio logy).
173

\ I ’ K I . l . I M I I N A K ' i A.S.SL.S.SMhIN I < )l - I I 11. A 1< I I I K( ) l ’ < ) D.S O I' A l U I D H A H I
208
Mosl scorpions prey on other arlliropotls, p ;irlie iil;irly insects and arachnids, but some ha\ e been known to k ill and consume \ eilebia les such as li/ards (see account lo r SfeiKxIciclyliis s lc i'i iii in ( ihaptcrb), snakes and rodents. They k ill by means ot neurotoxins injeeled From their sringinti tails. Some ot the.se toxins are potent enough to k ill a human but most are not deadly, although the sting may be painful for several days.
The seorpions are divided into nine families (Sissom 1990) and four o f these (the buthidae, Chactidae, lOiploeentridae and Seorpionidae) are known to exist in the M iddle East and are therefore like ly to occur in Abu Dhabi, although at present only Buthidae have been recorded.
From the family Buthidae, the genera A i u I i x k I o h i i s ,
Apitboh iitb iis . B irulatus. D iitbaciis, D iitbcoliis. Butbotus, C a iiipso bu tb iis , L c iiin is , L io h iilb iis , M tc ro h iilb tis ,
Ckloiiiobiiibiis. Ortbocbmis. Parahiitb iisAnd I acboiiio liis are known to exist in the Arabian Feninsula. There arc- tw o records o f V 'acbo iiio liisg lob inu iiiiis Levy et al. from Bada Haza in Abu Dhabi (Levy c/rt/. 1973. Vachon 1980). This species has also lieen recorded recently by NAKC from Abu Dhabi Emirate, as have B tilbac iis yoti’ateiisis ii liin x ic iilc a tiis le v y et al. ( Plate 10.1 ). Parahutbasliasoim i ( I lem prich and Ehrenberg), and Coinpsohatbasarabicas Levy- et al. (all specimens verified by |. Boorman in litt.). The genera Anclroctoaas, Bntbacns and Leinrns are considered the most medically im portant .scorpions in Arabia according to Simard and Watt (1990), i.e. tho.se whose venom shows a high toxic ity to humans.
The family Chactidae is repre.sented by the single genus Hnscoipins in the M iddle East, w h ile for the family Diplocentridae, there are records o f tw o genera, Nebo and Heteronebo. In the family Seorpionidae, the three
P h itc lo . I . n iill.u ii. ii.'iy i'li'iiU -ii.'ii.'i i i i i in x K i ik 't i l i i. ' i l .v \ y cV ti l . i.s p n ih . ih ly i Ik - iim .st o i i n i n o n .s c o rp u m I'o iin U o n s:ini.l in Ahii Dluhl.
lUHbttcusymtili'iisis C».jlv j l i J JiSi U-' V m til. iilf;nitii iilt‘tilii.<
P h ili- 1 0 .2 . A i : in u - l .spiUci ( It l i f t it It •.'t .sp. m :ik i.s ;i h i l l II i \ \ b y p ii.s h in x .Siinil \ \ l l l i it.s p ii l ip . i lp .s , n n l i i . I lik i. .1 h i i l l i l i ix v i
Gtlktxies .sp (J*»J • ^ >1J-, US JUjI
/ - /

A IM O ' I I M I N A I O A S S K S S M l ' N r O K T I U-: A K i l l K O I’ O I )S O l A I H i D M A M I
209
gcnciLi Hcniiscoipiiis. F a iic liiius and Scoipio ha\ e liecn recorck'tl in die M iddle Hast.
O ix lcr SoliFei^nc ( s o l i f u g id s o i c a m e l sp id e r s )
'I'here are ren know n fan iilies o f these fast m oving and extraonlinary-looking arachnids, three o f which probably occur in Abu Dhabi: the G aleodidae, Solpugidae and Khagodidae (C loudsley-Thom pson 1987). A lthough armed w ith powerfu l jaws that are capable o f penetrating the skin, they do not readily attack humans and, despite much controversy in the past, there is no evidence that they are venom ous (C loudsley-Thom pson 1984). They are predatory, m ainly feeding on o ther invertebrates and, in captivity, their preferred prey consists o f members o f the fam ily O rthoptera (C loudsley-Thom pson 1987)
The fam ilies G aleodidae and Solpugidae are s im ilar in appearance w ith very long legs and sandy colon red, ha iry bodies w h ile the Rhagodidae are sm aller and generally black. Several as yet un id en tifie d species o f Galeodidae ( Plate 10.2 ) and Solpugidae occur in isolated desert areas o f Abu Dhab i and Rhagodidae are often seen around Al Ain where they are attracted to the lights at n ight
O r d e r A r a n e a e ( s p id e r s )
There is very little know n about the spiders occurring in Abu Dhabi, the UAE or Arabia as a whole. Desert species are probably represented by families sim ilar to those- found in the Sahara wh ich are better known. These include ground-hunting spiders (Gnaphosidae), giant crab spiders and crab spiders (Sparassidae andThomisidae
K h ilc I O .S. A n I > rh - \\< .- ; i \fr Cl ul n ix n x l l i i ik ic m .Ai t> io p iU ;n .') lN ini> in w a ll In ils w I. h s ir i in i ' aem s.s a
sp. s liru b
Tcirajin.ithitlac ) gL.: r - i .^ J e I J ü u Jijj (ArRlopidac
■/A’gophyUum sp
IM atc 1 0 .4 . ( ’l ia n t \ e l \ e l n ilu - ! i i iK t lh n in ih i i i i i i s p n m s r^ iinx iVi im w c l s a n il a n x in t l a I lu is i w a tia p i p u
Dhiiilh ivnib iiin i sp. U3Ue <jUe. s i t - \ .
m m s m
1 7 5

\ I ’ K K l . l i M I N A K ' l A. S. sL . sSi M IZN' I t J T T1 I P. ' M Î T H R O P Ü D S O F A B L ' D F I A B I
210
iespeL'ti\ ely ). b;u k spiders ( H ersiliiilue ), juni]Dingspiders (Sukicidiie ), w o li spiders ( Lyeosiikie ), sheet-web spiders ( Agelenidue ), e o iiib ib o ii il spiders (Theridiichie ) und orb- weuvers (Tetragnarhidue and Argiop idae) (Cloudslcy- Thonipson 108 i ). i'w ek e species of Sakicidae have been recorded from Saudi Arabia (Proszynski 1993). They are all predatory and feed m ainly on insects and other in \ ertebrates that they catch liy various means according to the ir b io logy and adaptation. For example, Lycosidae and Sakicidae have excellent sight and actively seek out and hunt the ir prey. On the other hand, web-spinners such as Tetragnathidae, Therid iidae and Argiop idae lay their webs as traps to entangle fly ing insects that the spider can then de\ our ( Plate 10.3). Many desert spiders survive the harsh conditions by staying in burrows during the day and on ly emerge at night. Sonic species can also survive long periods w ithou t feeding when prey is scarce.
O r d e r A cn rin ti ( m i ces :ind t ic k s )
The free-li\ ing mites from arid regions have not been studied nor collected to any extent. However, they probably survive in damp microclimates such as the soil at the base o f plants or on some more specialised habitat such as the bodies of larger insects, e.g. beetles. One interesting obser\ ation from Abu Dhabi is a record o f a giant vek et mite, D in o th ro n ih iiin i sp.. from near the Dubai border fo llow ing a burst water pipe that apparently triggered the emergence o f the mite (Plate 10.4). These are ve iy large mites, about 12 mm in length, and as their name suggests they are covered in a thick layer o f dense, scarlet hair. They spend most o f their lives in the soil in . 1 state of diapause and only emerge fo llow ing hea\y rains when they come to the surface to feed on termites ( CloLKlsley-Thompson 1962).
0 \ \ ing to their blood sucking habit and their potential l( ) transmit rlisease and rerluce the condition o f livestock, ixodid ticks ha\e been studied in the Arabian region though not from Abu Dhabi itself. Hoogstraal elcil. ( 1981) ha\'c re\ iewed the argasid and ixodid ticks o f Saudi .Aiabia and Papadoulos el al. (1991) those occurring in Oman, but neither mentions any records for the UAE. Hoogstraal et al. (1981) also give taxonom ic keys and in form ation on the medical and veterinary importance o f these ticks. They identified 37 species and subspecies, many o f w h ich com m only feed on livestock such as camels, goats and sheep. As we ll as causing blood loss, the ticks may induce anaemia, toxic reactions and paralysis and afso transmit agents pathogenic to humans and domestic stock. The most common tick in desert areas is H ya h m m a (hya lo n in ia ) c lrnn iedarii Kock, the camel hyalom mine, a h igh ly adapted desert species that, as ils name suggests, chiefly infests camels. This species undi lubted ly occurs in the deserts o f Abu Dhabi where ii'' m.iin host is abundant. A few rarer species have also
been identified from Saudi Arabia by Hoogstraal (1982), Hoogstraal et al. (1983a), Hoogstraal et al. (1984) and Pegram et al. (1989).
A few species have also been found infesting marine birds and pigeons (see above references and Hoogstraal and Rafort (1982). Hoogstraal et al. (1983b)). A lthough the ir distributions and range o f hosts are probably w ide, there are no records for UAE. Many lizards caught in Abu Dhabi also bear one or more small red miles or ticks but the identity o f these has not been established.
S u b p h y lu m M a n d ib u la ta C la ss E n to g n a th a O r d e r C o llc m b o la
The Collembola or springtails are m inute, wingless arthropods. They derive the ir com m on name from their ab ility to leap in to the air w h ich they do w ith the aid o f a special springing organ know n as the furca A lthough generally thought to be dependent on high humidityy Collem bola have been found in sandy areas o f Abu Dhabi where they probably feed on the roots o f plants or other decaying matter. To date the ir identity is unknow n.
C la ss I n s e c ta ( tr u e in s e c t s )
Insects are the most numerous members o f the animal kingdom . They exhib it a w ide variety o f form and function, enabling them successfully to exp lo it even the most extreme o f habitats. Desert insects use various mechanisms to con.sen e water and sun ive heat stress and the ir small body size allows them to move into com paratively cooler microclimates during the day. For example, many retreat to deep burrows or the ba.se and roots o f plants where moisture le\ els and hum id ity arc h igher and temperatures low er than in exposed areas. They then emerge at night to feed once the tetiip e rature has dropped. However, there are .some in.sects that are active even during the intense heat o f the day. For example, ants o f the genus Cataglyphis can tolerate temperatures up to 30°C for at least one hour (Delye 1968), Conversely, there are also many insects o f tropical orig in in UAE that inhabit agricultural o r urban areas where they can on ly survive on irrigated plants w h ich are often non-native. This chapter m ainly considers the true d e s e rt-d w e llin g and desert-adap ted species. The classification system used is according to CSIRO (1991 ).
O r d e r T h y sa iiu r a
The Thy .sa n Lira are com m only know n as s ike rfish or bristle tails and are small wingle.ss arthropods usually covered in metallic .scales. They are often found in houses but some species also inhabit the desert, includ ing a few that arc commensal w ith ants. A small num ber o f
/ ~i,

A H K Ü U M I N A K ' i A S S r . S S M E N T O T T H E A R T I I R O P O D S O H A B U D H A B I
TFT
cleserr examples have been found and although their identity is uncertain they are probably from the family Lepismatidae, Irish (1991) revised the Thysanura o f the Arabian Peninsula and found 11 species but had no actual records from UAE.
O rclei' O d o n a to
The o rder O donata includes the suborders Zygoptera (dam selflies) and Anisoptera (dragonflies). Both have a fu lly aquatic larval pe riod and are genera lly associated w ith perm anent freshw ater w h ich , in the Emirates, is la rge ly restricted to some w adis and irriga tio n ditches. There fore they cannot no rm a lly com ple te the ir life cycle in the desert. Dam selflies are sm aller than dragonflies and are least lik e ly to be found away from water, but some o f the dragonflies are m igratory and fly across the open desert. F ifty -tw o species ha \e been recorded fo r the A rabian Peninsula and tw e lve o f these were collected in northern UAE (W aterston and Pitta way 1989). AluixpaiibeiiopeSe\yf=,-^\'\d C rocotbeniiseiythm ea (B ru lle ) have been found in desert areas (W alke r and Pitta way 1987) and are recorded from the UAE in Schneider and K rupp (1993). A s im ila r species that inhabits salt marshes is HeiuianaxeJ)hippi}jerBurme\MeT. This may be more w idespread in Abu Dhabi, but to date has on ly been recorded from Q arnein Island (B row n 1989b). Heath ( 1989) has records o f Selysiothemis n ig ra ( V a nder Linden ), A . partbenopc and Pan ta la J la i 'escens (Fabric ius) from Das Island. Schneider and K rupp ( 1993) also list the fo llo w in g as occurring in the UAE but g ive no specific localities: Iscbn n rn e ra ns i M orton. A rah ic in an isca e rn la W aterston, O rtbutrun i cb iys iig ina (B urm e ister). O rlb o lr iin i sab ina (D ru ry ), Diplacodes le fe b rr ii ( Rambur), Tritbem is a n n n ia ta ( Béarnais), T ritbenusa rte riosa (B urm e ister) and P. flaeescens.
O r d e r Rlattc^dea
Cockroaches generally li\'e in association w ith man and the ir o m n i\o ro u s habit and ab ility to transm it disease th rough con tam ina tion o f foodstuffs means that they can be serious pu b lic health pests. Members o f the genus Heterogamodes are true desert species and can to le rate extrem e arid cond itions (Abusham a 1984) but it i.s uncerta in w hethe r any occur in Abu Dhabi. Urban cockroaches, such nsP erip laneta a m e rican a Linnaeus and BJateiia ge rm a n ica Linnaeus are w idespread, cosm opo litan species (W a lke r and P ittaway 1987).
O r d e r T,sc:>ptera
The Isoptera or termites are soft-bodied, social in.sects that live in colonies made up o f many sterile, wingle.ss soldiers and ^'orkers, and a few reproductive forms. The
latter start life w ith the ab ility lo fly but shed their w ings fo llow ing a mass dispersal phase from w hich just a fev.' sun ive to set up new colonies. Dispersal generally coincides w ith rainy periods and the w inged termites pro \ ide an abundant and rich food source for many animals. Desert species know n from th e ,Sahara construct deep subterranean nests and workers are capable of gathering damp sand from the water table at depths o f up to 40 m (Popov.el al. 1984). T w o foraging strategies are adopted in the desert; species are either nocturnal, or they construct protective soil sheeting or tunnels where they are shielded from the heat and desiccating effects o f the sun. Termites feed on plant material that may be liv ing, dead or decaying. Some species are capable of reducing w ooden structures to dust and may cause serious damage to buildings. Twenty species are thought to occur in the Arabian Peninsula (Chhotani and Bose 1983, 1991 ) but the only record for UAE is o f a colony of Heierolennes ae lb iop ic iis (^o s ie d i) infesting a house in A l A in (B oocock 1979). Judg ing from its know n distribution, Anacantbotennes ochracens (Burm eisier) probably also occurs in UAE although it has not been recorded so far. Se\'eral species o f termites ha\ e been collected in the desert environs o f Abu Dhabi but theii identity' is unknow n (Plate 10.3).
O r d e r M a n to d eu
F am ily M an tidaePorly-.six species o f mantids are known from the Arabian Peninsula, but there are no published records from UAE (Kaltenbach 1984, 1991). However, a desert specialist, the common ground mantis Erem iapbila ba iie ri Krass, has recently been collected from the Baynunah area. Like other members o f this family, it is extremely well- camouflaged and is hard to differentiate from the rocky substrate upon w hich it lives. O ther cryptically-coloured species exist in the desert, liv ing hidden among grass ot shrubs w h ile they lie in wait for the ir prey w hich includes other insects and occasionally small lizards (Plate 10.6).
O r d e r O r th o p te r n
The order Orthoptera includes the grasshoppers (Plate 10.7), locusts, bush-crickets and crickets, and contains some im portant desert specialists. Indeed, tw o species that can cause great econom ic loss to crops, the desert locust Scbislocerca gregaris Forskal and the migratory locust Lociisla m ig ra to ria Linnaeus, have been recorded breeding in the deserts o f Abu Dhabi. Studies o f locusts and related Orthoptera have been carried out by the Anti- Locust Research G roup (now Natural Resources Institute, Chatham, UK) because o f the ir importance as agricultural pests (see Uvarov 1952, 1966 and 1977). They lay their eggs follow 'ing the rains in remote sandy areas. The eggs
1 77

\ I * K I - I . 1 M I N A K > A S S K S S M H N I' O l I I 11 A K T H i t O I ' O I J S O H A M U O I I A M I
212
Plalc 1().‘>. I'L r i n i l c s a r c c o m n i o n in I In
lIo S L T C n i lL lL T |- ) lo i'L -S o l W O O L I I I II a l
a h a n L l o i i L - i l h L ' i l i i L a m p s . I ' I i c .s l - iL iu t i l e s
( p o .s .s ih ly I lolcni lcni ics s p > w l - i l ' I n i n n
l I l ' V o l i i l ' l I l>y 11IL- l i - - i > t - l> r io n i i l Mfsiislcuii />iii ic/icti//is .<«t l iL 'i I P l a t e I I I . I I ) . n o r m a l K
I o n s l i l L T L - i l l o I x - a L lL - l r l t i \ I >iv.
^ jUlA/l Sa-J ) (,_>:Ajj')|l JaJI) i-JojS/' 3-\ ..i* ^ ^ USjlj Jâ 4 5 JL.I
Oils { l l i ‘i i ’n)U‘>vu's sp. jjSS j l Ja Ia J ja ) jV i sJiUll SAjUl M csi)sieiiti p iiH C IIco llis Snller .l-jiiJi
.y ia j ysi liU . TenL-hrionid-.iL
P l a t e l ( ) . ( > . . A l l h o i i L - l l l I l m i K l i s l l i I l - a . u a insl
I he haiki>roimLl ol'.sailLl. this LmiLk-nl il'iisl maitllLl 1s almost in\ isihlL- on l I l - s l - i I
VL’.HL’tation u Iilmv it amlxisliLs its ptL \
I j* *c -jj O jA jV j i l l .J^ill j i jA f i-J t ^
J ‘ J ‘ J i* ’ CV S-i ‘ J - j H l3 >• C J f k i.<-11 j« » lij <*jI>»-aJ1 OljLill
Plate lO."'. An aL t ieloiel in sin itl-lnntiL<1 ,i>tasshoppL-r
. j jjU Acridoitl v-t . cajI
/ -.V

A P I Î F . I . I M I N A i n A S . s C S . S M E N T ( ) ! ■ T H E A K ‘ l ' M K O EC) O S O P A B U D H A B I
hatch into nigiirless hoppers w hich in certain years may become gregarious and join to form large marching i)ands. The hoppers develop into w inged adults that congregate and migrate in huge swarms. The gregarious phases not on ly behave diffe rently from the ir non- gregarious forms, but also d iffe r in their appearance. Most true desert-dwelling grasshoppers do not fly long distances but prefer to hop using their powerfu l, well- developed hind legs. Many afso have the ab ility to produce .sound using special s tridu la to iy organs or scrapers on the wings.
S u p erfa m ily T e tt ig o n io id e a F am ily T e tt ig o n iid a eTettigonids or bush-eriekets are generally associated w ith vegetation such as herbs, bushes or trees and their occurrence in deserts is therefore lim ited to times or places w ith abundant vegetation. O nly tw o species have f)een recorded in the UAE: Trigouocoiypha a iig iis tn ta Uvaro\' from Sharjah and a m inor pest species Platyciois a lh ifrn tis (Fabricius) at Ras Al Khaimah (Popo\- 198J). There are no know n records for Abu Dhabi,
S u p erfam ily G ry llo id ea F am ily Gry llid aeThese are the true crickets o f which two species commonly occur in Aaabia. Firstly, the house cricket/it7)e/r/ domes!ica Linnaeus w hich is associated w ith human habitation, and secondly, the tropical field cricket G)yIIus bim aeula lus de Greer wdiich is found in vegetateLl areas. A minute, nocturnal species has also been found in desert areas but has not been identified
F am ily G r y llo ta lp id a eMole crickets pos.scss pow erfu l front legs which are fossorial, that is, adapted for digging. They spend much o f the ir life underground w'herc they feed on plant roots and small insects. The adults are w inged and are often attracted to lights. There are tw o species, Gry/lotnlpn gry lln ta lpa Linnaeus and the smaller G aj'ricatia . that are common throughout Arabia and probably occur in L?AE. Gorochov ( 1993) has records o f a third species, GiyJlolalpa dehilis Gersaeker. from Al Madam. L'AE.
S u p erfam ily A cr id o id eaThis superfam ily includes the more numerous and familiar grasshoppers and locusts, some o f w h ich are well- studied pest species. Popo\ (1980) gives 69 species as occurring in Eastern Arabia along w ith a \e iy u.seful taxonom ic key and comments on their biology, ecology and biogeography. There are records for 28 species in the UAE and although most o f these specimens were collected from Shtirjah, Ras Al Khaimah and Khor Fakkan ( in a part o f Sharjah on the east coast), it is like ly that they afso occur in Abu Dhabi
A u.seful w ay o f cla.ssifylng Acridoidea is to split them on the basis o f the habitat and terrain in w liich they arc found. Thus terricoles live on the ground and feed on plants w ithou t actually c lim b ing them (Uvarov 1977) and those w liich live in open deserts can be separated as deseriicoles. In turn, these can be either rr/'e/z/co/ccs', liv ing on sand oy saxicoles. liv ing on rocky substrates. In the deserts o f Abu Dhabi, we w ou ld expect to find mainly te r r i-desert I coles, te rr i-a ren i coles a n d te r r i-.sa.xicoles. Therefore, according to Popov (1980), the fo llow ing species are most like ly to be occur:
Tcrri-dcserticolesA liosc irtiis leagneri (K itta ry ). S ph iiigono tns fe m o ra lis Uvarov. S phiiigonotns octo/ascicitiis (Aud'inet Serville), Sphi ngonot I ispredtetshenskyM\s[ehenko. Sphiiigonotns rnbescens (W a lk e r ) , S p h iiig o n o tn s la e a n d n ln s Popov, P sendospb ingonotnsparadoxus ( B e i-B ienko ), Psendosphingonotns dentatns ( Predtetshensky ).
Terri-arenicolesTeiinitaisnsaiignstnsCQlMnch-MxM.Hyalorrhipiscanescens (Saussure), H ya to rrh ip is a ra b ica Uvarov. Acrotylns insnbricns longipes (Charpentier).
Terri-saxicolesScin iharis ia notablis b lanchard iana (Saussure).
O r d e r P h a sm a to c le a
Fam ily P hasm aticlaeThe phasmids or s lick insects are m edium -sized, phytophagous insects that are very d is tinc tive because o f the ir rem arkable resem blance to sticks or the stems o f plants upon w h ich they li\ e. They are not like ly to be \ ery num erous in the desert ecosystem, be ing confined lo the more vegetated areas. Several s lick-like specimens ha\ e been found liv in g on the desert giwssPennisetnm d ir is n n i ( Gm el ) a lthough the ir id en tity is unkno w n at present
O rd er H e m ip te r a
This order covers the true bugs as w ell as lea llioppe rs , cicadas, aphids and scale insects. True bugs e xh ib it d ive rs ity in both form and lifesty le , bu t they are all chtiracterised by having suck ing m outh parts. Most Hem iptera feed on the sap o f plants bu t some are predatory and feed on anim al tissue or b lood . There are three suborders in the H em iptera : the S lernorrhyncha, the A uchenorrhyncha and the Heteroptera. They arc not pa rticu la rly w e ll adapted fo r desert life and are genera lly on ly num erous w here there is abundant vegetation or, in the case o f p redatory species, o ther insects or prey
7 79

A I" l (K L I 11 A l ( \ A S . S K . s S . M t N ' l O h I l ib: A K I H K O P O L)S O l A 13 L 1)1 I A 131
^ 4
S u b o r d e r S tcrn o rrh y n ch aThis incliiclcs m o him ilies rhnt o in hccomc serions pests o f crops. They ure the families Aphiclidae and Coccoidae, know n as aphids or plant lice and mealy-bugs or .scale insects respectively. Both tend to Ise veiy small and li\ e by sucking plant sap. In many species, females can reproduce w ithout mating ( pathenogenesis) a llow ing populations to bu ild up \ ery rapidly. As well as causing damage to the plant by rem oving sap, they can also transm it plant \iruses. Sixty species of coccidids are kno w n from Saudi Arabia (M atile -l’errero 19<Si, 19BS). These ha\ e the potential to cause serious economic damage because o f aridity o f the countiy (Matile-Ferrero 1984 ). A lthough they undoubtedly occur in both cultivated areas and the deserts o f Abu Dhabi, there are no records to confirm this.
S u b o r d e r A u c h e n o r r y n c h a F am ily C ica d id a eCicadas ha\ e a \ ery long-1 i\ ed nymphal stage that learls a subterranean existence and is seldom seen. Adults are com parati\ ely short-1 i\ ed and spend their rime on trees. The males are conspicuous because they can produce a \ ery loud noise w ith w hich they attract females. They do this by means o f a paii o f special organs, the tymbals. w h ich arc situated on the afidomcn. The Arabian cicaila PU ilyp/c iirn (in ih ica Myers i.s common among stund.s ot trees in L'AH.
S u b o rd er H e te ro p te ra S u p er fa m ily P e n ta to m o id e aIn Arabia the most frequently oeeuiring families are the D in idoridae anil the Pentatomidae both o f w hich feeil on p lant sap. The D inidoridae includes the melon bug C orid i/is f if l iK if i is Fabricusa. a large black bug that feeds on native Cucurbitac but which may becotne a pest il numbers inerease anil it in\ ailes cu lti\ ateil erops. The Pentatomidae are com m only known as shielil bugs because o f their shape, or stink bugs because o f theii ab ility to exude ;t foul-sm elling flu iil when handleil. L innaxuori (1986) lists 4s2 species ol Heteroptera as occurring iti Saudi .Arabia, many o f w hich are new taxa. It is like ly that Abu Dhabi shares many o f these species an il they w o u lil be expecteil to lie w iilespreail w here\ er the ir plant hosts occur.
O r d e r N eu rop L era
In the desert ecosystem tw o families of N euioptera com m only occur: the Chrysopiilae or lacewing.s and the M yrm eleontidae or ant-lions. Both ha\ e predatory lar\ ae, the C hrysop iilae specia lis ing on aphids an il the M yrm eleontidae on ants, but o f the adults on ly the Chrysopiilae are predatory. The M yrm eleontitlae are by lar the most numerous and include many species whose
laiAae are adapted to desert conditions. They gel i Ik ii comtnon natne o f ant-lion from the Ian ae w hich kx d ferociously on ants. Most species con.strucl conical p itlike traps in the sa ml and flick sand grains ;it any passing ant. Ants that fall into the trap are im m ediate ly attacked anil de\ oured by the lari ae using the ir sharp, sickleshaped mouth-]iarts. A few ha\ e free-li\ ing larvae that do not produce traps but actively seek the ir prey instead. Adults superfic ially re.semble dragonflies but can be ilifferentiated by their clubbed antennae and net-\ eined wings, llo lz e l (1982, 1983 and 1988) lists 136 species know n to occur in the Arabian l^eninsula but there are no published records o f the species that occur in UAE.
O fc le f C o le o p c e i a
More than 300.000 species o f Coleoptera or beetles ha\ e been describeil w o rlil w iile an il about 40 "n of all insects and 30" I, o f a II animal species are beetles. Not surprisingly, they are one o f the largest groups in the de.sert and pi(>bably make up the largest biomass o f in \ertelirates that occur there ow ing both to the size anil abutidatice o f some ilesert spei ies. The Coleoptera m entioned here :ire those that are most numerous in .3bu Dhabi and w h ii h show .some ilegiee ol ilesert ailaptLilion
S u b o rd er A d cp h a g a Famil>’ C arab ldaeOne o f the most fam iliar ilesert beetles is A iith ia cliioc/cciiuii’ iiltcilci Bonelli. com m only know n as the ilom ino beetle (Plate 10.8). Like other t. arabiils it is an acti \ e predator and is often seen scurrying around the ilesert both by ilay and night. It prei son other in\ ertebrates but w ill also seax enge on earcasses o f \ ertebrates. Another common carabiil i.s .S'(.Y//V//e,s spp. ;m aggre.ssix e nocturnal pre ila tor w ith \ cry large, pow erful mouthparts.I he l.i biinae and Brachininae carabids o f Arabia ha\ e been re\ iew eil by Mateu ( 198()) but there are no records for Abu Dhabi or the UAE.
Fam ily C ic in d c llid a cTiger beetles are fast-mo\ ing, actixe preilators xx ith large sickle-shapeil jaxx s xx hich feeil on small inx ertebrates. There is a species o f tiger beetle that often frequents the seashore, muilllat.s anil sabkha in Abu Dhabi anil this is \m )\yA\')\y liiin ihi/iir-dbÙLUs. tilthough its iilentityhas not been confirmed. Txxo species o f tiger fseetle, p iobably Cicindclci ii in iu ii i is (Bates) an il C. I it in ila la . haxe recently been fourni on Meraxxah IsLinil.
S u b o rd er P o ly p h a g a F am ily H is ter id a cH iste rills are genera lly sn ia ll shiny, b lack, tough and very rounded beetles that are found on carrion o r dung. It is though t that they are actually p redatory an il lix e o ff
/ , S ( I

A MUHLIMIN Xs.sLSMMliNI' Oh I H I: ARTHROPODS OK A B l' D IIA B I
"21^
the laiATie o f 11 y and o ther beetle species that inhabit these places. Several d iffe re n t species o f histerids ha\ e been found in Abu Dhabi but the ir iden tity and b io logy are unknow n .
F am ily S carab aeid aeThe fam ily o f scarab beetles includes the dung beetles and chafers both o f w h ich can be quite numerous.
The most cotnm only occurring dung beetle in UAE is Scarabaeus crislatus Fabricius. It may frequently be seen fly ing over the desert and is often attracted to lights at night. It fo llow s herds o f camels and other livestock and can be seen ro lling balls o f precious dung across the sand searching for a suitable place to b u iy them. Having buried the dung the beetle lays a single egg and seals the entrance to the tunnel. The lanTt then hatches into a secure environm ent w ith the dung acting as a food store fo r its development. It is not on ly dung that is u.scd, however, and dung beetles may dissect and ro ll-o ff dead animals such as gerbils (Plate 10.9k This behaviour is also know n from some African species o'f Scarabaeus ( Scholtz and H o lm 1985). G ille ll ( 1994) has collected 32 species o f Scaraliaeidae m ainly from the Al A in area. Table 10.1 lists all the species o f dung beetles he has identified.
Chafers arc plant-feeders and their Ian ae generally Ih'e on roots o f plants and in .some cases can cause economic dtimage to crops and ornamental plants. Many adult beetles are attracted to artificial light at night. Several species have been collected in Abu Dhabi but their identity is unknown. G illett (1994) has records for 14 chafers found in the Al A in region.
F am ily B u p restic laeBuprestid or jewel beetle lan'ae feed under the bark of trees w h ile the large, m etallic-coloured and bullet-shaped adults are free flying, feeding on pollen and flowers. They are generally restricted to plantations or forested
areas but w ill also sun ive on i.solated Prusopis and Acacia trees. Several have been collected at Al A in and Um Az Zim ul but their exact identity is unknow n. Their elytra form a conspicuous com ponent o f little ow l (Athene noctua) pellets found near the Abu D hab i/ Dubai border.
F am ily E laterid aeElaterids or click beetles have larvae that feed on the roots o f plants. They get their com m on name from the ab ility o f adults to turn themselves over by Hexing their body and producing a c lick ing noise in the process. The body shape o f the adults is typ ica lly elongated and flattened Chassa in (1983) has records fo r 12 species o f Elateridae from Arabia and. although none are specifically from UAE, this beetle fam ily certainly occurs in Abu Dhabi as they are often attracted to artificia l light at night.
F am ily D e r m c st id a eDermestids are com m only know n as carpet or hide beetles and are scavengers, feeding on carrion and animal carcasses. In the desert they and other carrion feeders are responsible for the .swift remox al o f dead animals. There is currently nothing know n about those that occur in Abu Dhabi
F am ily C o c c in c llid a eMost cocc ine llids feed on aphids but a few are phytophagous. The commonest ladybird in Arabia is the seven-spot ladybird Coccine/lasepfenipiincfata Linnaeus, a prechitory species that may be found on desert plants in Abu Dhabi
F am ily T e n e b r io n id a eIn terms o f desert survixal, tenebrionids or darkling beetles are the most numerous and succe.ssful coleopteran family. They are highly adapted to water con.seiAxition
Tabic 10.1. .Spccic.s o f .ScLirabaciclac iV o m I he HA f
c o llc c tc U h\ (.'lilk-ll (190-1)
Âlilill pl\ 1 -( ■
LûljU y i jA Scanihiicidae .G illett 0 9 9 4 ) US
Species n a m e
S c a ra b a e u s c ris ta l us F ahric lo .s
G v n iiio p le u ru s s p l
G y n iiio p /e u ru s s p 2
C a tb a rs iu s iu eru U s (C a -s tc in n o )
C h iro n it is osiricUs R c ic h e
O u th o p b a g u s n itic iu lu s King
O u th o p h a g u s u a rie g a liis C Fahnc iu .s )
O n th o p h a g iis tra n s c a p ic u s ( K o e n ig )
O n tic e /h is .sp]
O n U ce llu s ,sp2
ApbocUus s p l
AphocUus .sp2
Aphuctius .sp3
A p b o d iu s .sp i
A p haciins s p 5
L o c a lity
A l A in , A l A in A l F a y c la h , A l A i n - D i i h a i r o a d , A l O ita .
M a h t la h a n d F(.>ssil \ a lle y
M a h d a h a n d A l A in A l F a y d a h
M a h d a h
A l A in —D i ib a i r o a d , A l A in —A b u D h a b i lo a d ,
A l A in —A l W ig a n r o a d a n d A d h D h a id
M a h d t ih a n d A l A in —A l W ig a n ro a d
Al A in —A l W ig a n ro a d . A t A in A l F a y d a h , M a h d a h
a n d Fo.ssil v a l le y
A l A in —A l W ig a n r o a d a n d M a h d a h
A l A in —i3 u b a i ro a d a n d M a h d a h
M a h d a h
A l A in
A l A in —A l W ig a n ro a d a n d M a h d a h
A l A in —D u b a i r o a d a n d M a h d a h
A l A in . A l A in —A l W ig a n ro a d , M a h d a h a n d Fossil v a l le y
A l O h a a n d A l A in —A l W ig a n r o a d
A l A in —D u b a i r o a d a n d M a h d a h
181

A l > K l £ L l M I N A i n A S . S K S . S M h N I O T T l l l v A U T H R O P O D S O F A l U I D l l A U I
ZTB
P la tt: 1 0 .8 . T h e d o m in o b e e t le . \ i i l h i t i r l i io c le c in in n lU lU l b o n e l l i is a n a e t i\ e p r e t la lo ty e a ia b id . o f te n s e e n a ro u n ti s u n se t.
A tith ia c liu x lec ln if’ i i l ta la Uonclli «L-iüJ a - \ . i» . jl J Carabidae iU J l ^
P la te lO .y , O u n j i b e e t le s .V<.Y/;v'/b/'«'/f.s' c r fs l i i / i is F a b iie iu s w i l l i l r a x a n t i ro ll e are as s es l ik e th is t ie a t l > -e ib il iC Jerh iH ns t7 > e e .v /; /< /; / /T h o m a s ) s e e e ra l m e tre s b e fo r e te a rin i> ih e n i a p a r t a n t i b u r y in j; th e m in p ie c e s
Scamlxicn.s chsUiltis Fabricius i j j H ^Thomas) làt J l. .lùdl j aw f Â
r  J ! W*j>“ Jfi (CerhiUns chcvsmaui■ U».V 1+Ü1J
P la te 10. I t ) . I ‘r it> in > lh cc ti c iin n u i lc i ( O l i \ ie r ) is th e la rg e s t o f th e de.sert te n e b r io n id s in U A F .
jjSI Piionotbeca cnronata (O liv ier) i . - 1 .> 7 ' tfi Tenebrionidae Âüdl W l l j jL iiJ
/.S’J

A I ’ K K I . I M I N A K ' » A S S R S S M I N r ( } | I I I I : A K I 11 K l ) I ' i ) I )S ( ) | \ I U I D H A H I
2tr
Plate I 0.1 I. M c s i i s l i ’i u i iH i i u l i C d l I i s Soller is pn ihahly ihe nu isi numerous Il nr I «I il m ill in Ahii |)liahi.
^ M esosteiia fw iiL lic n tlis S o h e r M - l . ê.^1 ■ lï^ j^ ' l-ut T fn i'h r io n id a e é tU il i» jlill
Plate 10.12. / ' i i i i f / i i i < i i t i h i c i i Kluj; i.s
anoiliei eommon ile.se-ri lenehrioniil
> j t-»* J* arabica KIuh i'f-> • i^ jlTenebhnnidue s ü J l >— J ,>L üJi

A I ’ K U L I M I N \ R > - A S S U S S M H N T ( ) H T H T . A R I H H O R O D S C.5F A H l ' D l l A B I
218
L in d can obtLiin all the water they need through their own metabolic processes. Most are scavengers or plant feeders. They are generally black or dark colou ied and show d i\ ersity in size ranging from the m inute to the ve iy large. The largest species found in UAE IsP rionotboa i coro)iata (O liv ie r), com m only know n as the urchin beetle, w hich is about 40 mm from the head to the tip o f the abdomen (Plate 10.10). Many o f the larger tenebrionids have their elytra fused and although they have lost the ir ab ility to fly they are thought to have an increased ab ility to retain water. A lthough large and obvious, many are avoided by vertebrate predators because they are distasteful. Most are crepuscular or nocturnal in habit.
Kaszab (.1981, 1982) lists over 300 species o f tenebrionids as occurring in the Arabian Peninsula, but veiy few examples have actually been collected from UAE and even few e r from Abu D hab i Em irate. Tenebrionids are an abundant and im portant com ponent o f the desert ecosystem and many species exist in Abu Dhabi. Table 10.2 lists the species presently know n to occur in UAE but it is like ly that many more w ill l)e found in the future. Typical examples include Alesostena p iin c lic o llis Sol ier (Plate 10.11) and the hvgev P in ie lia arab ica K ing (Plate 10.12). Sand swimmers, w hich are members o f the genus Froclius are also abundant in some desert areas.
Fam ily M elo ic laeA du lt Meloiclae or o il beetles are often very co lou rfu l and may be seen feeding on flow ers o f various plants. T h e ir close association w ith f low e rin g plants means that they are generally on ly seen in desert areas fo llo w in g the rains, but some species can cause serious econom ic damage to crops. Many have an aposematic o r w a rn in g co lo ra tion to advise predators that they are distasteful o r even toxic w hen eaten. Lan ae are thought
to be parasites or parasito ids o f o ther a rth ropods but there is litt le kno w n about the ir b io lo gy in Arabia. E igh ty-e igh t species have been listed fo r the Arabian Peninsula (Kaszab 1983, Schneider 1991) bu t records fo r IIA E and Abu D habi are scarce, a lthough this p ro ba b ly reflects a lack o f know ledge rather than the occurrence o f desert species. G ille tt ( 1992 ) htTs found a potentia l pesi species E p ica iita erytbrncepba la (Pallas) feeding on w ild plants in Oman, fa irly close to the Al Ain area o f Abu Dhabi.
F am ily C er a m b y c id a eThese are large longhorn ' beetles whose larval stages live and feed under the bark o f trees such as acacias. One species A cantbopboriis a rab icas (Thom son) has been found at Svvcihan in Abu Dhabi Emirate. Holzschuh (1993) lists 54 species or subspecies o f cerambycids occurring in Arabia but has no records for the UAE.
Fam ily C u r cu lio n id a cThe CurcLilionidae are com m only know n as w eevils. They are all phytophagous and their lanae usually li\e and feed in a protected en\ ironm eni such as the root oi shoot o f a plant, wh ile adults feed on the external parts o f plants. Adult w ee \ils are easy to distinguish from olher Coleoptei'a because they have prom inent snouts or rostrums and elbowed or angled antennae. They are not w ry common in the de.sert but a few smaller species such as Anuin)cleo)ias ascbahaiicnsis Faust have been found on ZyiidpbyHiim spp. plants in Abu Dhabi.
O r d e r D ip te r a
W orldw ide this is one o f the largest orders o f in.sects. although in the deserts o f Abu Dhabi there are many individuals from just a lim ited number o f species. The\
Species n a m e ,
Znpbosis n i ig e a n x i s im p le x K a .s zn li (1 9 S 1 )
P a c h y c e ra p y g m a e a a m b ic a (C K o c l i )A ie s o s lf iu ip n iic r ic o l/is S o lie r
M ic m d e ra in c iig iiin ta d es e itic o la (B la i r )T e n ty n n a d e s e n a sp a isep u iic ta tc i K a.s /.a lj ( 1 9 8 t )
O xy c c im h u e tt ife r i K a s z a ti ( 1 9 8 1 )A desm ic i c o rh iirn a /o o n ia n e n s is K a.sza lr (1 9 8 1 .)
A d e s m ia c n tb n rn a ta ra th je n s i S c lii is te r
Adesm ict s io e c k le ir ii ra s a lk h a m n t ia K n s z a li ( 1 9 8 1 )
P a ra p la ly o p e a ra b ic a a r a b ic a (B ta ir )
T ra c h y d e rn ta h is p id a (F o rs l< )T rn c b y d e rm a p b ih s t in a R e ic l ie a n d Sa n ic y
T b r ip ira b r a a iz i H a a g -R .P im e U a a r a b ic a o m a n ic a K a s z a b (1 9 8 2 )P irn e iia a r a b ic a a ra b ic a Kln^P irn e tia a r a b ic a e m ir i K a s z a b (1 9 8 2 )
C o n o c e p b a in m b e s n a rd i K a s z a b ( 1 9 8 2 )
O p a ira id e s pu n c ia la tiL S B r u lte
O p a tro id e s v ic in n s (F a ir m a r ie )P b a le r ia p ro H x a F a im ia r ie
L o c a lity
Ra.s A l IC Iia iin a li Ra.s A t K l ia lm a l i . D ilil^ a
Ras A l K h a im a h , D u b a i D i l ib aRas A t K l ia im a l i , lOitrlna
Ra.s A l K h a im a h
Ras A l K l ia im a t i D ib b a
R as A l K l ia im a h
Ras A l K h a im a h A b u D l ia b i
Ras A t K l ia im a h . D u b a iM a sat'iDulxiiD u t? a iRas A l K l ia lm a l i
R as A l K l ia im a h Ras A l K h a im a h
R as A l K h a im a h
“T r u c ia l O m a n "
Tabic 10.2. I.isi 11| tcncbi ii m ill bc fllcs kiK iwn tn im OAK.
Alter Kaszab t 1081 1V82)
Hjldl sajUJI is il ^1^1 CÛÜ T —t < d jd* Tenebrionidae
.Kaszab (1981, 1982) j c .s.w iall
is /

A PRm.I.MI \ \K 'i ASSrsSMTNT OT TIIP: ARTHROPODS O P A B l^ P ^ lA B l
CLin be cliscingLiishecl from other flighted insects liy tiie ii single p iiir o l wings. The second pair is reckiced to a small club-shaped structure know n as a haltere, which is im portant for balance and stabilirs' during flight. The hcause fly M iisca clonieslic{i Linnaeus is ub iquitous and capable o f spreading di.sea.se as well as being a general nuisance and irritation. In the M iddle East, some w ork has laeen done on those flies that are a potential hazard to the health o f humans and the ir livestock, but little interest has been shown in those families that contain m ainly non-pest species.
The D iptera are split in to tw'o suborders, Nematocera and Brachycera, on the basis o f their m orphology. The B rachycera are fu rth e r sp lit in to tw o d iv is io ns , O rthorrhapha and Cyclorrhapha.
S u b o rd er N e m a to c e r a F am ily C u lic id a eA num fier o f species o f mosquito has been recorded from the Arabian Peninsula, and studies o f these insects have concentrated on their im portance as medical or \ eterinaiy pests since some species are capable o f transmitting diseases. One such disea.se i.s malaria, although in recent years there have been few cases reported from the Arabian Peninsula.
Fam ily C e r a to p o g o n id a cBoorm an (19S9) has w ritte n an account o f the genus CuHcoides or b iting midges o f Arabia and lists 2-f species a long w ith notes on th e ir b io lo g y , d is tr ibu tion and ab ility to transm it disease. Five species were found in UAE: Cti/icoicles irc ird i Boorman. C. rcii'iis de M e illon . C. o.xystonia K ieffer. C. in nco la K ie ffe r and C. cizerbcijdzhciiucits'Dzhw'i-Moy. Apart from C. oxystonia and C. inucoki. there is little know n about the pest status o f these species. C oxystonia is kno w n to be a serious
b itin g pest o f cattle and it may also be able to transm it cattle filariae. C. inuco la is a very serious veterinary pest and is a kn o w n vecto r o f A frican horse sickness, b lue tonguc v irus and A kabanc virus.
C. inesgbalii Navii has recently been added to the list for UAE, identified from the Baynunah area (J. Boorman in litt.). Noth ing is know n about disease transmi.ssion in this species w h ich was caught b iting the author!
F am ily C h ir o n o m id a eFifty-three species o f Chironom idae or non-b iting midges have been recorded fo r the Arabian Peninsula (Cranston and Judd 1989). C hirononnis cciliptenis is the only species collected from Abu Dhabi where it was reported to be a nuisance pest around the A l Ain sewTige works, occurring in veiy large numbers.
S u b o rd er B ra ch y cera D iv is io n O ith o r r h a p h a F am ily T a b a n id a eThe ta ban ids are com m only know n as horse-flies or clegs. Males are harmless but females require a blood meal for the developm ent o f their eggs and some species are pests o f livestock and an irrita tion to humans Tabanids probably occur in Abu Dhabi but have not been recorded.
F am ily A silid a eThe asilids are know n as robber flies, and are active predators that capture other insects on the w ing. Members o f the genus Apodea are com m on in desert areas o f Abu Dhabi.
F am ily B o m b y liid a eThese are com m only know n as bee flies because o f their superficial resemblance to bum ble bees and the ir sim ilar
Tahle I (I..S. UcinhyliiiliUs hLC lliu.s iVoiii I'
.M 't f l- C ' . I f ; i t h f ; K l I UI.S.Si
ji Bdnibyliidaf CUr. j-ijii r - \ .
•Circarhcatl (19HK) j .
Species n a m e L o c a lity
B a n ib y liiis iiieÿciccpheiliis Poivllin.skii Harm—At Madam RuadA iu is lo e c h iis niL’cus Hermann HarmSystoccbns h o rr it ln s Greathead HartaB<ii>ibylisrima SPiiegn/ense ( Macc|ii:irO Harra, .Sweihan roadPcirorossei tropiccilis liJezzi HartaPclixtirissci (tlb ifd c ies (MaccjLJart) LIAJZD e sD ia lo iie iirc i b rec ip en n is (.Bezzi) Sliweila—At Madam RoadDesinciU nieiirei .sp. Sweihan ruadA n th r a x in fd sc la tc i Meigen Wadi Al Fay, near DibbaSpogostylnni cdn clic liin i I Sack I As Sa adSpogostyluni tnippoytd (Wiedemann) ■Shweib roadSp ugvslylnni ucyd le (Wiedemann) Hartan y r ic ld ) i lb r d x c lec ip iild (Ansren) Shwcil-»—Al Madam Road7 b 1 Tictd n th r a x p e r i c i Id n's p e r ic ila ris (Loew) Wadi Al Fay. near DibbaFxt.tyata m b r a x b ec h erian n s (BezzO Wadi Al Fay, near DibtiaP d c h y n tt ih ra x n n n u u td n n (Greathead) Shweils—Al Madam RoadV e rib n ijd d iig d s te o c n ld liis (Becker) Ras Al CharV erib n b o a im s ( Wiedemann^ Al Bahrani I.siandH e te ro to n id itieg er/c i (Meigen ) FlartaH e le ro /a n id n n ic n re a (King) As SaadJ ie te ro h m ia o tiv ie r i (MacquarO As Saad

A l ’ R l i L I M I N A K ' ï A S S F S S M h N f O F T H F A R T H U O P O H S O F A H H D H A B I
“ 220
liab it o f feeding on nectar-pmciueing Howers. They are im portan t as po llina to rs and several species occur in Abu D hab i. G reathead ( 198<S) lists the records in Table 10.3 fo r UAE.
D iv is io n C y clo rrh a p h a F am ily C a llip h o r id a eThese are the fam ilia r b luebottles and greenbottles w hose la rvae feed on ca rrion and o th e r dead organ ic matter. Some, such as the w in te r greenbottle L u c ilia seh ca la M eigen, w ill also lay the ir eggs on the w ounds o f livestock, causing them considerable pa in and d iscom fo rt as w e ll as preven ting norm al healing . N ot recorded from Abu D habi a lthough they p ro ba b ly occur.
F am ily M u sc id a ePont ( 1991 ) lists 68 species o f the fam ily Muscidae from the A rab ian Peninsula but o n ly fou r o f these have been recorded from UAE. Many such flies are po tentia l p u b lic health pests because o f the ir ab ility to transm it disease by the con tam ination o f foodstuffs. The most im portan t o f these is M iiscci c/oiiieslicci cloiueslica Linnaeus, the com m on house fly, w h ich can be found even in isolated desert areas. S im ilarly, the subspecies M usca doniesUca ca lle ra W alker has also been recorded fo r L^AE as have M u s c y i l i ic id i ik i Loew and AIiiscci .so / 7;e/ / s W i e d e r m a n n .
O r d e r L e p id o p te r a
M oths and bu tte rflies tend to be the best-studied insects because o f the spectacular beauty o f some m em bers o f th is o rder. They h a \e a com p le te m etam orphosis and the ir larvae spend most o f the time eating to b u ild -u p the food resources required to
ach ieve th is tra n s fo rm a tio n . The m a jo rity have phytophagous larvae that feed on one o r a few specific food plants l.iut some larvae are insectivorous. In general, the adults have a co iled proboscis w ith w h ich they feed on the nectar 6 f plants or o ther flu ids , bu t some have a troph ied m ou th parts and are incapable o f feeding. They spend most o f the ir life as a ca te rp illa r and on ly em erge a.s adults fo r a very short tim e to mate and die. Some species m igrate as adults and may occur in rem ote reg ions a long way from w here the ir larval food plants are found.
Larsen (1983) has wndtten a c o m p re h e n s ive m onograph o f the 151 species o f bu tte rfly ( Rhopalocera ) wdiich occur in the Arabian Peninsula. W iltsh ire ( 1983, 1986, 1988 and 1990) has rev iew ed the d is tr ib u tio n and abundance o f the 625 species o f M acro Heterocera (la rger m oths) w h ich are found in Arabia, Those that have been recorded in LIAE o r are though t to occur there are listed be low w ith comments.
S u p erfu m ily C o sso id e a F am ily C o ss id a eThe cossids are com m on ly kno w n as goat and leopard moths. / lo lce rce riis j^ lurios iis Erscho\- is com m on in grassy sand dunes and ho llo w s in eastern Arabia (W alker and Pitta way 1987). It has not been recorded in UAH although it p robab ly occurs there.
S u p crfa m ily I^ ra lo id e a F am ily P yralidaeThere are no actual records o f pyra lid moths for UAE a lthough they undoub ted ly occur there and it is su rp ris ing that e\ en the M editerranean Hour moth EphcsHa kticd in ie lln Zeller, a serious pest o f stored products, has not been recorded. O ther non-no.\ious species such dsLaiuorici ciiiollci Denis and Schifferm üller
Species: n a m e
G o in a lia e h n a e ln ia Trimfn
S p ia lia d o n s d n ris Wiiltcer
S p ia lia cololes se triico n jliie iis De Jong
S p ia lia z e b ra b if id a Higgin.s
P e lo p id a s m a lb ia s m a th ia s (Fabricus)
G e g e n e s p n n U lio ssp
C o m m o n n a m e
African mallow .slvipper
Deserc grizzled skipper
Grizzled skipper
Zebra grizzled skipper
Lesser millet skipper
Pygmy skipper
L o ca lity
UAF. (lîrown 1992)
UAE (brown 1992)
UAE (Brown 1992)
UAE (Brown 1992)
Khor Fakkan (Larsen 1983), UAE (Brown 1992)
UAE (Brown 1992)
Table Id.-I. The skippei btit lerll ies c >1 I A F
al>^1 J i ilkill jiljill t- H Jja» .1.13,0*11 4 *11
Sp ecies ':riam e. . - .: ' f C o m m o n ■name: .
P a p il io d em o leu s d en to le iis Linne Citrus swallowtail
P a p it io m a c h a o n m u e lin g i Seyer Swallowtail
'.Locality
Felij, Al Ain and Sharjah Clairsen 1983),UAE fBrown 1992),Al Ain Zoo (Wingate 1992), Das Island (Reaney 1987)
UAE (Drown 1992),Al Ain Zoo (Wingate 1992)
Table lO.S. The s\\ allow tail
biittertlles o f I I.AE.
CjIjUyi >. 11,1.11 jü jill Ô-1 .
!S ( )

A P R F . IJ .M IN A i n A S .sE .S S .M F N ’l O F T H F A R T H R O P O D S O F A H LI D H A H I _ 221-----
T a b le K I 6 T in - pii. r id In iltr i Ilie> ol I 'AH
^ Pifriclae ^U l j li ji ll . Jja»
; Comwiore,:K«me.“De Niccvillc Small while
P o n tia i ila n c o n a n ie (.K\up.) Desert white
E u ch lo e b e le tn ia belem ici CEsperl Green striped white
E lp h in s to n ia c h a r lo n ia a m s e li Desen black tip(Gross and Ebert)
A n a p b a e is a i im ta (Fabricius) Caper white
Cn/otis c a la is a m a m s Fabricius Small salmon Arab
ColoHs p h is a d ia p h is a d ia (Godurt) Blue spotted Arab
Colotis H agore Klug
M a d a is fa iis ta fa u s ta (Olivier)
C a to p s ilia J lo re lla (Fabricius)
Desert orange tip
Salmon Arab
African emigrant
A p harU isacc irnash ypaigym s(\iux\t::\) Leopard butterfly
E u re m a h ccah c senegaleusis Grass yellow
i j^ i Çjjdi CjijUyi A rto g c ia ra p a e Linnaeus Small cabbage while
UAE (Brown 1992)
Al Ain Zoo, Fujairah (Larsen 1983),UAE (Brown 1992),Das Island (Reaney 1987), Merawah Island (Brown 1990)
Sharjah (Larsen 1983)
Wadi Shaab (Larsen 1983), UAE (Brown 1992)
Felij, Jebel Flafit, Dubai (Larsen 1983),UAE (Brown 1992),Das Island (Reaney 1987)
UAE (Brown 1992),Das Island (Reaney 1987)
Sharjah, Al Ain (Larsen 1983), UAE (Brown 1992)
UAE (Brown 1992)
Al Ain (Larsen 1983),UAE (Brown 1992)
Khor Fakkan, UAE (Brown 1992),Das Lsland (Reaney 1987)
Al Ain (Ltirsen 1983)
UAE (Brown 1992)
Das Island (Reaney 1987)
did likd l) to b t‘ tound in odses and s tone-s lie \\ n plain.s (W a lke r and Pittaway I )
S L ip c r la n iil> H e s p e io i t lc a F a m ily H c s p c riic la c -rhese aid co inn ion ly know n as sk ippd i h iittd illids and can hd d is tinguished from other b iitte rn ies liy the way they lay the ir hind w ings flat lin t keep the ir forew ings in a vertica l pos ition w hen at rest. Table KJ.t lists the species recorded as occu rring in UAE.
S u p e r fa m i ly P a p ilio n o ic le a F a m ily P a p ilio n ic la eThese are the fam ilia r sw allow tail bu tte rflies that ha\ e a gracelu l flig h t and generally possess a p ro jection or ta il' on the back edge o f the ir h ind w ings. Table lO.S
gives the species recorded in l^AE
F a m ily P ie r ic la eThe p ierids are com m only know n as w h ite or su lphur bu tte rflies since most are p redom inan tly w h ite , cream o r y e llo w in co lour, a lthough a few species are light p ink o r orange. The most com m on lar\ al food plants o f the pierids are members o f the Brassica fam ily, includ ing crops such as cabbages and radishes. Sometimes the laiATie can occur in sufficient numbers to cause economic damage. Many species are strong m igrants and occur w ide ly across the whole Arabian Peninsula. For example,
m igrant species such Aiu ipbcie is ctnraUi (Fabric ius) M dcldis/c iiis lc i fd iis id (O li\ ie r) a iirl C citops ilid J lo iv lla (Fabric ius) ha \e an unpred ic tab le annual d is tribu tion and abundance. H o w e \e r, some pierids have on ly a lim ited d is tribu tion such as F Jph iiis lo iiia cbctrlon ia (in ise li (Gross and Ebert) w hose larvae feed on locally occu rring w ild cruc ife rous plants. The species know n from the UAF are listed in Fable 1 ().().
F a m ily Ly c a c n ic la cMost lycaenids are predom inantly blue in colour, hence the com m on name o f blue butte rflies In some species there is a m arked sexual d im orph ism w ith showy, blue males and brow n, c ryp tica lly co loured females. Mans s])ecies are m igrants w ith a w idespread occurrence, such :\s Ldiiipicles hoeticiis ( Linnaeus) w hich uses a w ide range o f larval food plants. Others are con fined to areas where the ir lar\ al food plants occur. For example. T c in ia is loscicens ( Austraut) and more rare ly T ariic iis bcilkcin iciis (Freyer) are found where Z izypb iis sp grows. A lthough Zizeeha karsancbri karscuuira (Moore) is a weak flie r, it is qu ite wdciespread p robab ly because cu ltivated alfalfa is its most im portan t larval food plant in Arabia. As a whole , the fam ily has a w ide \'ariety o f larval food plants in c lud in g tree species such as Prosopis and A cac ia w h ich are used by Cbilades p a rrb a s iiis (Fabric ius) Table 10 7 gives the species know n from the UAH.
7,S':

A I ’ R l i L I M I N A K ^ A S S F S S M |-:N 1 O l ' T H l i A K T I I R O PC) O S <.)1- A B I j D I I A B I
^ 22-
F am ily N ym p h a lic laeThe n y n ip h iilid h im ily incliiile .s tlie to rto iseshe ll, m ilkw eed , b ro w n und fr il i lh iry bu tte rfiies w h ich make up a c o lo u rfu l and \ aried ^ ro u p 'I'hey are ve iy capable llie rs , but the ir fron t legs are small and they w a lk using on ly the ir m idd le and h ind legs. D a n a iis chrys ipp iis cbrys ipp iis (L innaeus) is abundant and has a very- w idespread d is tr ib u tio n th roughou t Arabia.
Vanessa ca rd u i ca rc iiii (Linnaeus) is a true migrant and it is veiy com m on at times across the wdiole Arabian Peninsula. Hypolinn ias misippus (Linnaeus) is also a
m igrant but shows considerable fluctuation in numbers, w h ile 1 p th in u i bo lanica Marshall is thought to be confined to the more m ountainous regions. Its laiA'ae probably feed on grass as do those o i'H ippa rcb iapa risa tis (K o \h r). Table 10.8 lists the species recorded in UAE.
Sui:>erfamily G e o m e tr o id e a F am ily G e o m e tr id a eThe geometrids are com m only know n as the looper m oths or inch w orm s because o f the way the ir larvae b ring the ir rear pro legs up to t lie ir fore pro legs w ith the
Species n a in e . . C om m on, n a m e
A p h a r it is n iyn iiec o p h ilc i Dumont De.sert leopard
A n tb e n e a n u n 'a b a m a r a h (Guerin-) Leaden ciliate blue
L am p id es hoen'ciis (Linnaeus) Pea blue
T a n ic iis i-osaceiis (Ati.straut ) Mediterranean pierrol
T a riic u s b c ilh au ic iis (Fre>er) Balkan pierrot
Zi=L‘e n n b a rs n m h n b a is i i i id n i Asian rass blue( Moore )
A 2:n n u s u h a lc liis Cramer
A zcm u s Jesons (Guerin)
A<^rudkicli(s lo e u 'ii ssp. Zeller
C b dtides p a r rb a s in s (Fabricius)
Vr'het .spotted blut
.A.l'iiean babul blue
Loew s blue
Small cupid
h'rcyerici tm cl.iyh is Iro ch yh is Kreyci Grass jewel
L o c a li ty
LIAH (Brown 1992)
Shargiyin (Larsen 1983)
Al Ain, Jebel HaFit, Dubai (Larsen 1983),UAE (Brown 1992),Das Islanil (Reaney 1987, Western 1988)
'Trucial Coasts", Fujairah,Al Ain (Larsen 198.S), l.IAE (Brown 1992)
Wadi Al Jizzi (T.ar.scn 1983), UAE ( Brown 1992),Das Island. At Khazna (Reaney 1987)
Al Ain (Larsen 1983).IIAE (Brown 1992),Das Island (Western 1988, Reaney 1987)
LIAF ( Brow n I V92)
Khor Fakkan (Larsen 1983). UAH ( Brown 1992)
UAE (Brown 1992)
Al Ain, Fujairah. Dubai (Larsen 198.3),UAF. (Brovin 1992),Das Island ( Reane\ 1987)
Meravi ah Island ( Bn >wn 1990). IIAF (Brown 1992)
Table 10.7. The blue butterllies ot I I.AE
^ j j j y i J i j W V - ) . J jo , .iWi.ll
Species n a m e
U c in a iis c b iys ip p iis cbrys ipp iis ( Linnaeus)
C o m m o n n a m e
Plain tiger or milkweed
\ 'anessa c a r d u i c a rd m ( Linnaeus) Painted lady
J u u o u ia o r itb y a berc Lang Blue pansy
H y p o lin iiia s m is ippus (Linnaeus) Diadem or egglly
y p ib im a h n ia n ic a Marshall
U ip p a rc b ia p a r is a t is ( Kollar)
Baluchi ringlet
White-edged rockbio^\ n
L o c a lity
Felij. Abu Dhabi Das Island (Reaney 198' ).Meraw ah Island ( Brown 1990) Al Ain, .AI Ain Al Faydah, LIAE (Brown 1992)
Das Island (Reaney 1987), Meitiwah Island (Brown 1990), Al Ain Zoo. Baynunah, UAE (Brown 1992)
Das Island ( Reaney 1987),Al Ain, UAE (Brown 1992)
Das Island (Reaney 1987). UAE (Brown 1992)
Al Ain Zoo (Brown 1992)
Das Island (Reaney 1987), Dibba—Masafi road (Brov ’n 1992)
Table I 0.8. Ths nympi lalid iTUtlerllies 11| 1 C\E
jjllll jStjilt A - t .
ÂHjdi C jijUyi ^ Nym phalidae

A I ' K H I . I M I N A K ' . A .SS CSS M K N I ' O F T H E A E T 1 I K < ) l->< ) D s ( ) I- A H U D H A B I
22T
body fo rm ing u loop w lic n moving. The on ly records Foi the UAE me both fo r E np ithecia n ie k ra iu i c in iim /is W iltsh ire from "T rnciu l O m nn” and Masafi (W iltsh ire1986). In general, geom etrids w o u ld be expected to have a w idespread occurrence w ith m any species feed ing on cu ltiva ted and oas is-dw e lling plants. For example, the jasmine emerald Cblorissa c/iscessa W alker is o ften found in oases where its larvae feed on C lorode iic lro ii /'/?e/v;/e(W iltshire 1990).
S u p crfa m ily B o m b y c o id e a F am ily L a s io ca m p id a eLasiocampids are com m only know n as eggars and their lanxie are usually covered in dense hair. In some species the hairs contain a poi.son that can cause an irrita tion if touched. Chilenn lansta>ia Daniel is found in sandy desert areas o f UAE where its laiwae feed on Ccilligom ini com osiim (W iltshire 1990). Strebloie s ira Lefebvre, the Jujube lappet, is one o f the commonest species in Eastern Arabia and has been noted at three coastal locations: Ras Ghanada (ow n ob.seivation). Das Island (Reaney 1987) and Merawah Island (B row n 1990). Lasincampci s'em ila palaestinensLs Staudinger has been recorded in Masafi and w ou ld be expected to occur elsewhere in E^AE (W iltshire 1990). 'J'rnniinda m im dissin ici W alker is a w ide ly occurring species across Arabia and has also been found in UAE (W iltshire 1990)
S u p crfa m ily S p h iiig o id e a F am ily S p h in g id a eS phingids o r haw km oths are some o f the largest moths and the ir bodies are covered in th ick scales g i\ ing them a fu rry appearance. The da y-fly in g M cicroglossiin i stelU itc irnn i Linnaeus o r hu m m ingb ird haw km o th has been seen at Ras Ghanada (ow n obser\ a tion ), Das Island (Reaney 1987), Hatta and D ibba (B row n 198-t) and Al A in ( G ille tt 1993) and is probab ly a m igrant from no rthern O m an w here it breeds (W alke r and Pitta way1987). d c6 erm 7 / /r/ .\V t tv West w o o d . the Eastern death's- head haw km oth (Plate 10 13). frequents oases and towns, and is know n from Al A in (W ingate 1992, G ille tt 1993), Das Island (Reaney 1987) and Abu D habi (B row n 198-j). It is \'e ry large and makes a curious squeaking noise by passing air through its spiracles when disturbed. D a p h n is i ie r i i Linnaeus, the o leander haw km oth . is fou nd w h e re \’er its la r\'a l food p lant, N e riiin i sp. or oleander, is abundant (fo r exam ple. Das Island (Reaney 1987)). O leander is a com m on ornam enta l shrub in tow ns and cities but also occurs natura lly in some wadis (J. B row n 1991 ). The silver-striped hawkmoth/7//?/?o//o77 ce le iio Linnaeus has been recorded at Al A in A l Faydah (W ingate 1992) and the striped hawkm oth Z;/)7e.s'//7'0/v?/C77 Esper at Abu Dhab i (B row n 1979) and Al A in (G ille tt 1993). The convo lvu lus haw km oth A g riiis c o n ro lv ii/ i i Linnaeus is kno w n from Abu D hab i (B row n 1979).
S u p crfa m ily N o c tu o id c a F am ily L ym an triid aeLym antrids are know n as tussock m oths and Casanici L'ilis (W a lke r) is recorded as com m on in UAE and no rthern O m an (W iltsh ire 1990).
F am ily A rc tiid a eThe arctiids or tige r m oths are b r ig h tly co lou red w h ich serves as a w a rn in g to vertebrate predators since many are po isonous w hen ingested. The d a y -fly in g crim son speckled foo tm an Utelbeisa p tilc b e lla Linnaeus has been recorded on Das Is land (Reaney 1987), Q arnein Island (B row n 1989b) and at A l A in Zoo (W ingate 1992).
F am ily N o c tu id a eThe noctu ids o r ow le t m oths fo rm the largest fam ily in the o rder Lepidoptera. H ow ever, the o n ly record for UAE is o f a s ingle exam ple o f H ete ropa lp ia ve liis la (W a lke r) from Sharjah (W iltsh ire 1988). The lack of records is u n lik e ly to be due to the ir ra rity but rather because noctu ids are noctu rna l and genera lly a drab b ro w n or grey co lou r and so o f litt le interest to most co llecto rs The larval food plants are o ften grasses and some noctuids are agricu ltura l pests, such ^f^Spoduptcru /7V/07Y7/7S B oisduval, the co tton leafworm .
Fam ily P sy c h id a eO f the p sych id sA iid c ta D iiir in a K lug has been found in UAE. Its larva makes a four-s ided sack o f tw igs for p ro tec tion and these are com m on ly seen in desert areas around Baynunah and M edinat Zayed in Abu D habi on Jlrdo.xy/on s c ilic o n /ic iin i and H. p c rs ic in ii (Plate 10.14).
O r d e r H y m e n o p te r a
The o rder H ym enoptera includes all the bees, wasps, ants and sawfJies. The sawfJies are members o f the suborder Symphyta whose adults d iffe r from those o f the suborder Apocrita (bees, wasps and ants) by the lack o f a m arked cons tric tion bef^'een abdom ina l segments one and two. They are phytophagous rather than preda to ry or ]ia ras itic and there are no know n records o f the ir occurrence in UAE p robab ly because they ha\ e been ove rlooked .
A lthough some o f the Apocrita are so lita ry, many bees, wasps and ants characteris tica lly live in h igh ly organised, social colonies where most o f the ind iv iduals are sterile, fem ale w o rkers o r sold iers w ith on ly one or a few specia lised females and males capable o f rep roduc ing . The H ym enoptera contains some o f the most h igh ly advanced an im al societies. In most cases, the larvae are apodus ( legless) w ith small heads. They have lost the a b ility to m ove since the adults p rov ide them w ith all the necessaiw food or food stores fo r the ir

A l'IŒLIMINAin- ASSliSSMLNT l)F THF, ARTHROPODS OF AHU DHABI
-224
i
i
Plate 10.13. Ku.slern tleath's-head hawkmoth A chen>tiiU i siy.x We.stwoocl at rest on a PiXK-tcfpis tree. Its cryptic coloration allows it to blend in with the bark of trees and so avoid predation.
AcberoiUia siyK I j j j sjjj ji tr - t . i»jl l«ji jll j! .Pmsopis kJtt i ^ fin Westwood
j U J jb-Ail .U Jljï 4- ^
Plate 10.14. Uirval case of the psychid A inictc i i i i i i r h u i King on Hcilo.xylou fx.-is iciim .
Amicta muiiiui klug ÂVU Ujj 11- \ . ■//cilwcylon persiatm ij*»-i Jc- Psychidae kliUli
Plate 10. IS. PcircipsciiiiinophiUi t iira iiic c i Morawitz is one of the solitary sand wasps found in Abu Dhabi's de.serts
Parapsammopbila turanica Morawitz 'o-l •^ ^ , ^ > 7 1 d.ji _*uj A.I
/VU

\ I ’ K I - L I M I N ■\K' i ‘\ S M - S S M 1 - N 1 O I - I I I I-: A I M ' I I K O l'< ) l ) S ( ) I- M i l l I J i l A H I
225-
(.lc \e lopm ent In the deserts ol Abti D h iih i, ants are particu larly numerous anil form an im portant part o f the d iet ol many p re ila to ry in \ ertehrates and vertebrates Some ants are sca\ engers erf carrion and w ill soon strip any carcass that they find , w hile others are specialist seed-eaters and the ir e ffic ien t forag ing leaves litt le on the surface o f the sand. Many o f the sm aller wasps are parasitoids, laying the ir eggs on a liv ing host The wasp laiA'ae then develop inside the liv in g host, usually resulting in its death. Some species are parthenogenetic atid femtiles are able to lay v iable eggs w ith o u t the need to mate, Parasitoids have not been studied in UAE in any detail and are re la tive ly unknow n in Ara liia as a w hole
S u b o r d e r A p o c r i t a F a m ily A n th o p h o r id a cThe anthophorids gather wood pu lp to construct their nests and are com m only know n as carpenter bees. I'hey are large and \ ery noisy fliers m aking them quite conspicuous. The fo llow ing ha\ e been recorded in Abu Dhabi: ( Friese ), AniegiUa hyssiiui( K lug). I ’i lh it is tcirscita ( Mor. ) and Xylocupnpiihesccns Spitio la (Ham er 19(S6). Xylocopci/eiicstrci Pabricius was recorded from lla tta ( I lam er !9<Sh)
F a m ily M e g a c h i l id a eThis family includes the leal-cutting bees and se\eral species have been collected from \arious localities in HAP (Hamer 1986 and Table 10 9)
S u p c ifa m i ly C ih iy s ic lo id c a F a m ily C h iy s id id a cChrysids are commonly known as cuckoo wasps and tend to be bright, metallic green or blue in colour The\ are fast llie rs and are stnall to m edium in size. Koche (1981) collected a po.ssible 11 species from the fo llow ing genera in HAP Cicptiiicie. Chiysis Purnopes. H o ilych rid inn i. Heclychnmi and Onutliis
S 11 p c r fa m i h V c s p o ic lc a F a m ily P o m p i l id a cThe pom piliils are known as the spider-hunting wasps. They are m ainly confined to stony locations such as wadis and gra \e l plains The females provision their nests w ith spiders that they paralyse. The spiders do not
immediately die but act as a liv ing food store for the de\ eloping wasp Ian a The males do not hunt spiders but feed on flowers and po llen and are very fast flying. Se\ eral un identified pom pilids were collected by Roche(1981).
F a m ily S c o liid a cThe scoliids are know n as beetle wasps. Roche (1981) collected between two and four species from two genera Scolia and E/is. It is often d ifficu lt to tell which males and females belong to the same species because o f the marked sexual dimorphism.
F a m ily E u m e n id a eEumenids are semi-social o rso lita iy w asps. The fo llow ing species have been recorded in Abu Dhabi by Hamer(1982): Cl.doroclyiienis c/.T/oriticus (Spino la), Della canipcin iforn ieg ra c ile iS-Aussuve), Della c lin iinua tipen iie (Saussure). D e lla b u lle iito ta n i elegans (Saussure). Etioclyneriis e.xcelleiis (Perez) and Eiiocly)ienis iiilo lic iis (Saussure)
F a m ily V e s p ic la crhe.se are the true wasps that li\ e in social colonies Richaids ( 198-tb) has records for three species: Ves/)a orie iila lis arieiila li.'i Linnaeus and Fulislcs iiu lic iis Stolfa at AI ,Ain. and Folisles ir a l l i Cameron at Hatta.
SVIbfamil\ Ma.sarinaeThese are wasps that feed on pollen and Mowers The fo llow ing have been recorded in HAE by Richards (168 ta): (J iia il i i/ ia m ih ia iia Richards at Hatta and As Saad. Celonilesjotisseam iiei R du Huys.son at As Saad.
J i i g i i i l i a Jeniei/e iisi .s Kostylei at Hatta
Family^ F o r m ic it ia cAnts are an im portant eco logica l group, some o f w h ich are w ell adapted to desert cond itions w h ile others can on ly survive in irrigated areas and oases In spite o f the c osm opo litan occurrence o f ants, few records exist for the M idd le East and e\ en few er fo r ll.AE Surveys ol Saudi Arabia and Oman have found 156 and 28 species respectively (C o lling w oo d 1988, 1988) and recent material has boosted the total num ber o f ant species in Arabia to around 278 (C. C o llingw ood in l i l l . ) . Table 10.10 is based on T igar and C o llingw ood ( 1993) atid
Taille Ida; r t ie leal-o.unn.t> tivcs
Mexai liiliLlai n f rom I 'A t ' tVisi il on
I l.iinei ( I VN(,)
M fi'aO iiiidac VLUU j p l ^ U li <l-\. ; a ^ l V u jJ I O IjL -V I j . . .
Species n a m e
A lc g a c h ile p c ite llin ic iiu i Spinota Me,L(ac/.}ile u 'c ilkeri lA. Tor. M e p a c h ile in iilis s iiiu i Karl C h alicoctonu i n ih rip e s Mor C re ig b lo n e lk i an ic ih ilis (Civil. ) M e s a u lh ic lin m a l/c r iu is (Klug) .4 ii/hcicapa h e /o iifu .'ia (I riese)
Lo ca lity
Abu Dtnatri MaltaAlsu It)ha hi Hatta Malta I tall a AI Ain
191

A i >K i i . i M i N A i n a s s i -:s s m i ; n i l i t - I i n - a k t i i k ( •)!><.) n s o i - m î t i a i i m ü
226
Species n a m e
Subfam ily M yrm tcm ae
M essor e b e n in n s I Orel Aiessor rn /u les lac e iis Foer.stt-r C'rei)ia/Of>cister a i ita r is Fore!A io iio in d r iii I I I abe iU ei Anil re A k m o n io h in n tu a y r i Fore I A lo n a n to riu n t l i in u i ir i Collingwoocl ( in b it ) M o iio n io n ii in iir ib ib ie n s is Collingwood ( in /ill.')
Subfamily Formicinae(b in ip tin n lN s .w r.w .i Fore I
Ckilci,iilypbis b r id a André C a ia g irp h is n ig e r (André)
C a la g /ypb is J /a ro b n m n e n s Collingwootl ( in b n .)C a ia g /)p b is m in im a Collingwood ( in bn ) C a ia g ly p b is sa /m /n sa Kugler A c a n lb o /e p is nigrescens K:ir:iw:ijew
Locality a n d recorder
Ras Glr.inada Island (Tigar)2--l°46.R'N Sh°SS.3’E (Tigar)2-)°a6.S'N Ai°5T.3'E (Tigar)Ras Ghanada Island (Tigar)24°4b.H'N 3 °53.3'E (Tigar)Jebel Halu (Heatwole). Ras Ghanada Island (Tigar) lebel Halit ( HealAvole), Ras Ghanada Island (Tigar)
Ras Ghanada Island (Tigar), .AI Ain Al Faydah Hotel and Fark (Wingate)lebel Halit ( Heatwole). Ras Ghanada Island (Tigar) Al Ain Zoo. Al Ain Al Faydah Hotel anil Park.AI Wigan (Wingate)lebel Halit ( Heatwole), Ras Ghanaila Islanil (Tigar )
2 t° ib.S'N 5+"3t.3'E and Ras Ghanada Islanil (Tiiaar) lebel Halit (Heatwole). 24°h(-.,.S'N ôa°Â5.,VE (Tigar) Ras Ghanada Island and 2 t°-t(i.H'N 3'E (Tigar)
I'alale I () I () List i it atit spi-i
rent in lei I h i un ,\ln i I )h.ibi
Finit ate
ijt^i j*iii j-ty ■ J jA -
Species'^ ”I -xii-wtJH ■ ' . •- I 1- i-S ■ ■ ■>>-■«- ' L. - . .C b /o rio n sem en o ivt o c c u /e n ia le Beaumont C b /o rio n b ir tn n i (Kohl)C /ylorion J in io ro u m CjiibodoC b a l) •bionJle/jite ( Lepeletie de Sa int-Fargea it )S cebpbnm m a d ra s p a ia n n m p ic ln m (S m \(\) )S cebpbron p iiicb ic lb im re c ln m KholS p b e x fn m ic a tn s ChristSpbe.x p rn in o s iis GermarS pbex / la r ip e n n is FabrieiiisP r io n y x c n id e /is SmithP r io n y x n ire a ln s (Duloitr)/^ r io n rx r i i /n a ln s (Christ )/k ira jjs a m m il/lb i/a I l ir a n i r a P re im icb ares sp.
/^ iid a /im ia iy d e i (Le Guilloit)A m m u /> h i/a /ja o c i/iic iie m is Mot iee A m m ii/ ib i /a e i in in e a Kohl A m m o /)b i/a g r a d / / i m a Tasi henbetg A m im i/ i / i i /a r iib i/ies Spinola
Locality a n d recorder
Al Ain ( Brown )Near Dibba (Roehe)Wadi Mor. Jebel Ali, Shweib (Hamer)Dubai, Abu Dhabi ( I lamer)Hatct. Khor Fakkan. Waili Asitnah ( \ lamer. Roehe) Hatta (Hamer). Hatta ( Roche), Al Hyayntth ( Hamer) Widespread ( G nicha til )Abu Dhabi (Roche)Al llyaynah ( I lamei )Widespteail ( Giiicharil )Widespi ead ( Guii hat il )Abu Dhabi. Hatta ( I lamer 19B6)As Saad (Roche)Abu Dhabi ( Roehe). Futaisi Island ( I Lmii i ) Widespread (Guichard)Asimah. Khoboos-Kawan (Llaniei )As Saail (Roche)Abu Dhabi. As .Saatl. Hatta (llami-i I9S(i )Abit Dhabi. As Saail. I lalta (Hatner 19H(i)
Table lO II Recot ils ol the subI.imiK Sphecinae Irom the HAE. taken Irom Guii haril I iu.SiSa) unless otherwise
siitc. ■ - i-v'll .*,*-) 1- ./i ) ) ~ ) , d jA i
; .iAilJl CiijlaV' ^ S|ilieiinac jji. J,I jdtl tji VI Guichard ( l9KKa)
.allj
Species
D ry iu /e //a tr ic o /o r (V„\r\ Linilen) n iy i id e / /a i ie p b e r li li (Pulawski) P )iy iid e //a n s ir ic a (Pulawski /:> iynd e//a h ifa s c ia la (Pulawski)
Locality
Abit Dhabt. Hatta. Al Markhaniyalt Al Markhaniyah Al M a rkhaniyah AI Markhaniyalt
I aisle 10,12 Ki-i oi'ils < >1 the
sublamily Astatinai- Irom the
ILAF. bas iil on 1 Ltmei i lO.S.S)
dilt aurt.i./, D - i . Jjaa jc .iAguli iajdi ejtjt-yi y Aslalinac
.l-laiiiiT ( 19SS)
incorpoTdtcs the icco ids ot Heatwole (1991) aiiU ohseiA arions by \X ingaie ( 1992) O nly l-i .species are listed and they are probab ly on ly a small p ropo rtion o l those that oecnr They range in habits from com m on honseho ld pests, such as members o f the genus M o iio n io r iiin i. night foraging ants such as Ciin ipo i/o /ns .wr.xos Forel. to those that are h igh ly adapted to the descri environm ent, such as the genus Ci/taglyphis w h ich can forage even du ring the heat o f the day. The species list a lm ijst certainly re 11 eels the areas where the ants were collected u hieh included a residential fie ld l eseareh station as w e ll as several desert areas around As Samhah, A l A in and Ras Ghanada Island, all in Abu Dhabi Tmirate. Doubtless many more species wait to be recorded in the Emirate
S iip c i fa m ily S p iic c o ic le aThe family Sphecidae comprises small to medium sized solitary wasps that make their nests by digging tunnels in io soft .sandy banks (Plate lU. Is). They capture and paraly.se other insects w ith their stings to proxasion theii nests. Some families and e\ en genera ha\ e been fairly w ell w orked for L'AE by Guichard ( 19HSa. 19HSb, 1989a. 1989b, 199.4). Twenty-seven species o f Zlc/)//;/.vare listed for the Arabian Peninsula o f w hich 16 ha\ e been found in UAE. O f the three species o f S'/fro/V/es. tw o are found in UAE and fix e o f the 12 known Arabian S/iziis have also been recorded. UAE records are particularly p lentifu l from the Sweihan road and from Abu Dhabi, although this probably reflects intensity o f collection rather than abundance o f the wasps themsehes. Tables 10.11 to
/o j

A l - K U l I M I N A I O A S . s l „ S . s M i ; \ I O T1 I i : A K l I I K( > l ' ( ) O S ( ) I \ H I D H A l i l
227
I ih ic lO lA. Kun xils wl tliL
suhhiniily L.iiriti.ii' IVoiii ihc I lAI:. Ii;isn l 1111 I liiniui ( I'J.s.s)
un ie » o iIk tw isi.- suili il
üilxi ' V- \ . J_,i,
j t eji jU"!(l Uirriniie
J ! ij) V) llum er (1988)
Species
I . i r i s (i/L>i/is (f-'. sin)/Jris a ln ia la (Spinohi)U ris n i^ ra Kohl/. i / is iti^ i ic a us (Walker)L iris prcieleniiissct ( Kichartls )U ris s iib fa s c ia tti (Walker) G cislniscricus elcclus Nurse G astrosericus m o r ic t i (E. Saunrl ) C astrosericns s u u c /n s Pulawski Gcis/roseiicus u n / f / i t Spinola Tachytes com f ie ri Turner Tachytes cH i'e ts ico ru iau u s Turner Tctcbytes u i/o tic iis Turner Tachytes p ig m a e u s Kohl 7’cicbyles s iu t ia liis Pulawski Tachytes tricbo pygus Pulawski Tacbyspex a lh o c iu c tu s (Lucas) Tacbyspex cheaps de Beaumoni Tachyspex costae (De Stef.) Tacbyspex e ry th ro p b a n is D. Tor T acbyspex e iy tb ix jp iis (Spinola) T a c b y s p e x fu /t ic o ru is Turner Tacbyspex g ra n d is s im n s Guss Tachyspex in certu s (.Rad.) Tacbyspex m aicH i de Beaumont Tachyspex m y c e rin u s tie Beaumont Tacbyspex n itid n s (.Spinola) Tacbyspex osiris de Beaumont Tachy spex p a lo p te n is ( Da hi ) T a c b y s p e x p u lc h e r Pulawski T a c b y s p e x p a n z e r i (Van Linden) Tacbyspex rttgosus Cuss P a la rn s d an g a le n s is Klu.q P a ia ru s sp iu o la e sp in o ia e Saussure PaU trus iaetus Klug P a la rn s p a re u h is de Beaumont M iscopbns ctenopns Kohl .Miscopbns h e lre tic n s Kohl M iscopbns im ita n s Gin Mar M iscopbns in im e tic n s Hon T iy a x y /a n a e g y p tin m Kohl T iyo xy /o n a ra b ic n n i Guss O xybe/ns la m e /la /n s Oli\ iei (Xxybe/ns c o /ia ris Kohl Oasy/>n>ctiis a ra b s Kohl
Locality a n d recorder
Abu Dhabi, Hatta Abu Dhabi. Liwa Abu Dhabi. Hatla Abu Dhabi Abu Dhalii HattaAbu Dhabi, As Saad. Hatta. Sweihan Abu Dhabi. Sweihan Abu Dhabi, Hatta Abu DhabiAs Saad. Al Markhaniyah, Shweil->-Al Madam roadAs Saad. HattaAbu DhabiHattaHatlaHattaAbu DhabiAbu Dhabi, Hatta, Shwell>-Al Madam road Abu Dhabi Abu DhabiAl Ain. Abu Diiabi. As Saad, HattaAs SaadSweiiianHatlaI lattaAbu Diiabi, Al Markhaniyali, Sweihan As Saad. Hatta As Saad HatlaAs Saad, SweiiianAbu Diiabi. As Saad. SweihanHatlaAbu Dhabi (Hamer). Abu Dhabi. As Saatl (Roche) Hatta. Shweib Al Madam road (Roche) Widespread (Guichard)As Saad. Shweils—Al Madam road ( Rot he )Abu Dhabi, Sweihan, As SaatlSweihan. As SaadSweihanSweiiianI lattaHattaAbu Dhabi. As Saad. I latta. SweihanAl Ma rkhaniyahHatta
r.lblc it) i t. Recortls ol the subfamily Nyssoninae Irom the ( lAE. taken Irom GuichartI ( lOSOa) unless otherw ise
i lilc Cis'.ll . l i i L ^ t fc - 1 .
^ j d i .jijL .y t Ny,s.sonin:te jd .\ G V) Gtiich.-trd ( 1989a) >
Species
A m m a to m n s mesnstenns ( Hantlerlirshe) A m m a to m n s rn jo n o d is (Radowski) l ie m b ix tra n c jn e b a ric a (Gmelin)
B e m b ix g a z e lla G ttichard B e m b ix saadensis GuichartI B e m b ix c h a p a rd i BerlantI B e m b ix fre y g e s s n e ri Mtirice B e m b ix d a b ib o m i Handlirst11 B e m b ix c b lo ro tic a Sptnola B e m b ix o c n la ta La tie II le B e m b ix n ig r is /iin a Guithaitl B e m b ix ra d a s zk o n s k y i I lantllirsch B e m b ix rn f ire n tr is I’riesner B e m b ix p r ie s n e ri Beaumont
B e m b ix p a l/e s c e n s Priesner B e m b ix hbo lt Monte B e m b ix n H o tica Priesner
B e m b ix m c b i GuichartI
B e m b ix b a m e t i GuichardStizo id espoec i/op terc is HanderlirschStizns c itr in n s KlugStizns fn s c a tn s Mot iceS tizns b iz o n a tn s SpinolaS tizns n a d ig i Roth in NailigStizns rn fic a rn is (Etirster)Stizns s a i'ig n i Spinola Stizns a ra e ic n s Guichard
Localiiy a n d recorder
Hatta (Hamer 1988)Hatta (Hamer 1988)Shweil>-Al Matlam roail (Rociie).Sweihan road (I lamer)Shweib-AI Matlam roatl. Sweihan road (Roehe)As Saatl (Roche)lebel Ali and Ril'aa (Hamer)Witlespretitl (Guichard)Witlespread (Guichard)As Saatl (Roche). Sweihan rtiatl (I lamer) Witlespread ( Guichai tl )Sweihan Roatl (I lamer)As Saad (Roche)Hatta ( Rot lie)As Saad. Shweil3-Ai Madam road. Al Marklianiyah ( Roche). Jebel Diianna, Sweihan (Hamer)As Saatl. Abu Dhabi. Sweihan. I latta (Hamer 1988) As .Saad. Al Marklianiyah (Roche)Abu Dhtibi. Sayh Huwayyah. Shweili—Al Matlam roatl (Roclie)Siiwtib—Al Madam rotitl. Sweihan road. As Saatl. Bitia Al Ajam, Al Marklianiyah (Roche)Sweiiian (Hamer)Aiitt Dhabi (Roche), Hatta (Hamer 1988)Hatta (Hamer)Abu Dhabi (Hamer)Abu Dhabi (Hamer and Roche)Shw'eili-Al Madam roatl (Roclie)Sweiiian road (Hamer)Shweib-Ai Madam road (Roche)Abu Dhabi, Lullat'yah, Jebel Ali, Khor Pakltan (Hamer). Hatta (Rot he )
19 A

A I ’ K l i L l M I X A I O AS.M;sSM1:N I O f I I I f AK I I I K O l ’ O n s Of- AUf | - ) t f \ l^ l228
Species
F b ila n th n s coe irctanis Spinnln F h ila u tb u s ^ e ita lis Kolil P b ilc n itb u s p c illid u s Klug I 'b ila n tb u s Ir ia itj^ t ilu in (Faf-)riciu.s) C ercens a lb ic b ic ic i Klug C erceris c b ra m a tic d Sell I
Ce> -cens f is b e r i Spinola C erceris Iciterip ixiclucrn Moc hi C e rc e ris J 'itx g e ra k li Empey C erceris bcin io ri Guichard C erceris In rkestc iiiicn Rndoszkowski Cerceris a lb o a trc i Walker C erceris s t ra n iii ie a Dufour C erceris uittalci eitrypgct Kohl C erceris iiugcj.x Arnold
C erceris e n g e n ia Schlellerer C erceris d iff ic il is Guichard C ercens v a g a ta Kohl Cerceris s o lH a ria Dahl C erceris tricu lo irilc i Spinola
Lvccdity a n d recorder
Al Ain, Abu Dhabi, A.s Saad, Haifa (Hamer)
Abu Dhabi (Hamer.)
Abu Dhabi, As Saad ( I lamer)Abu Dhabi (Hamer)Abu Dhabi, As Saad, Sweihan (Hamer)Abu Dhabi, As Saad (Hamer), Sweihan. Sh\veil>-Al Madam road (Roche)Abu Dhabi, Hatta (Hamer)Abu Dhabi. As Saad (Hamer)Abu Dhabi, Remah, As Saad, Jebel Ali (Guichard ) Sweihan, Remah (Hamer)Ilayl (Hamer)Wadi Asimah (Hamer)Al Babha(Roche)Sweihan road, Dubai—flatta road (Hamer) Shweili—Al Madam road (Hamer), (Roche) Sweihan road. Al Mtidam road. Al Awir—Mileiha road (Hamer)Sweihan road ( Hamer)Hatta (Hamer)Wadi Asimah. Kalba, 1 lallti (Hamer) flatta (Hamer)As .Saad. Hatta, Abu Dhabi (I lamer)
fable lO IS Recoiils ol the
subl'amily I’ hilanihinae From
the LIAH. based on f lamer ( Ib.S.S) and ( jn il hard ( 1003 )
djU. ) 0- ) .^ jd l Ciljt-yi I’hilantliinae
• Guichard ( 1993) j I lamer ( 198K)
10. Is list by subbim ilic.s all iccorcls o f sphccicl species reeoixled in the Emirates. T h irty three species ol Ccrcdris ha\ e been foun(.l in .Arabia o f w hich 16 occur in V.\E
S L ip c r fa i i i i i> A p o i t l c a
F a m i ly A p ic la c
Members o f the apid fam ily are the fam iliar and economically im poiian l honey bees, such ;is, l/)b iiic llifen i lannaeus that is kept in commercial hi\es. The small •Asiatic honey bt'c ApisJlo ivd Fabricius has also become established in Abu Dhabi ant.1 is reporlcxl to be the most com m only occurring honey bee (1 lamer 19.S2)
Cda.s.s C r iis ta c :e a ( c fc is ta c e a n s )
The Crustacea are characterised by ha\ iiig tw o pairs o f antennae, three pairs o f m outhparts and many leus. Most Crustacea are aquatic, fo r exani|)le crabs, praw ns. lobsters and shrim ps However, a lthough the o rder Isopoda generally contains many marine or Iresh-w ater dw e llin g species, it also has some that are terrestria l and surpi is ingb ' do occur in desert en\ ironm ents
C la s s C h ik ) | ) ( ) c la ( c e n l ip e c ie s )
The cla.ss Chilopoda are m yriapodous arthropods. i,e the) ha\ e many le.gsand were form erly grouped together w ith other similar arthropods. They are prcdaton and reh ili\e l\ sofl-bodied Their bodies are dor,so-\cntra lh flattened and the head possesses a pair o f simple antennae, a group o f simple eyes, three pairs o f mouihparls and one pair o f poison glan(.ls. The rest ol the bod\ consists o f numerous segments each ol W h ich bears one pair o f legs .Some species can give a painful bite though it is not likely to be very serious. To date, onh the order Scolopendromorpha has been foun(.l in Abu Dhabi, bui a few Ceophilom orpha may also occur. Lewis and Gallagher (1993) list four locations for S a ih ifd iid ra iiiirc ih i/is (Porat) in CAF anil Scolopcmlm I'alidci Lucas once in Sharjah. Nine species ha\e been recorded in Saudi .Arabia and there is no reason to suppose that those occurring in Abu Dhabi are different species. They are not thought to be especially well adapted to the desert, are nocturnal and are not usually seen (.luring ifie day unless disturbed un(_lei stones and other refuges (Lewis 19,S I)
O r d e r I s o p o d a ( i s o p o d s ) P o s t s c r ip t
Tw enty species o f terrestrial isopods have been recorded fo r the Arabian Peninsula (Taiti and Ferrara 1991). Dcsert-dw e lling isopods from the fa mi I \ Porcellionidae. com m only know n as wood lice . have been found in Abu Dhabi, although the ir identity is unknow n They sun ive by constructing special burrow s that a llow them to m aintain a m icroclim ate w ith su ffic ien tly high hum id ity du rin g the day. They emerge to forage on the surface o f the sand at n igh t (Lewis 198j ).
ln \ ertebrates form one ol the most numerically important groups in arid regions and show a irem endous \ ariety of structure and lifestyle, besides their direct contribution to desert b iod ive rs ity , they are undoub ted ly im portant in the food chains o f \ ertebrates such as reptiles, birds and mammals. Yet invertebrates remain poo rly studied and the even the most basic co llecting can help im p iove ou r know ledge o f them in Arabia. The in fo rm ation in this chapter is based on the data a\ a ilab le up to

A PRELIMINARY' A.S.SES.SMENT OF THE ARTHROPODS OF ABU----------------------------------------------------------------------- 22y-----
February 1994, bu t the s itua tion is rap id ly changing. NARC’s studies on the feed ing eco logy o f the houbara bustard inc lude a m o n th ly program m e o f p itfa ll trapp in g at five sites across the Emirate. D is tr ibu tio na l records o f the invertebrates caught are stored on a database fo r easy re trieva l and sorting. By Decem ber 1 9 9 4 , the database con ta ined 20,368 observations o f invertebrates from ove r 200 species, m ostly ground- d w e llin g species and desert specialists. W herever possib le, id en tifica tion s are be ing carried ou t w ith the he lp o f in ternationa l experts, fo r exam ple, at the Natural
H is to ry Museum , London, bu t th is is becom ing the lim it in g factor. Inve rtebra te taxonom y has become unfashionable, the rem aining experts often being retired pro fessionals w h o con tinue the ir w o rk as a labour o f love. Abu D h ab i’s geograph ica l pos ition at the m eeting z o n e o f th re e b io g e o g ra p h ic re g io n s m akes id en tifica tion s especia lly d if f ic u lt and the reference m ateria l itse lf is p o o rly w o rked . A ll too soon, it w ill no longe r be possib le to f in d experts w h o kn o w A rab ia ’s invertebrates. New species and form s that are today und iscovered may be lost fo reve r in the sands o f time.

230
Appendix 1.2
Terrestrial Arthropods (excluding insects)
by Tigar, B. J. (1996b).
In: Vine, P.J. (ed.) Natural Emirates. Pp 107-120. Trident Press, London.

Terrestrial Arthropods (excluding insects) Bmimrn Ti^nr
Argiope, the orb or
signature spider,
packaging its prey.
The zigzag anchoring
chords may serve to
draw the attention of
a potential victim
away fiv iii the web
which captures it.
T he f l o r a a n d fa u n a o f th e UAE contain a mixture of elements from three
major biogeograpluc zones: the Northern Palaearctic, Africa and Asia. This,
in combination with an extremely dry climate, results in a characteristic
invertebrate fauna with typical arid-zone species, such as camel spiders and
scorpions, being relatively abimdant. Legends and tales of the terrible scourge
that these creatures cause aboimd. Eadeed, on first sight they look like products
of an araclmaphobe's nightmare, ha this chapter I hope to dispel such myths and
show the important roles these animals play in their environment. 1 also explain
how, tlarough their biology and ecology, mvertebrates are well adapted to one of
the most inhospitable parts of the world: while most humans venture only
briefly from the conafort and safety of their air-conditioned, four-wheel drive
vehicles, the ai thi opods take the desert m their stride, albeit usuag many legs!
H is t o r ic a l r e c o r d s o f t e r r e s t r ia l a r t h r o p o d s
Invertebrates of the Arabian region as a whole remained poorly studied until
the late 1970's, and the UAE still has far fewer records than neighbouring
Saudi Arabia and Oman (see the Fauna of Saudi Arabia series and the lournal
of Oman studies for further details). This is mainly due to the lack of collectors
and records rather than a paucity of fauna, although the other two countries
are larger with greater climatic and biological diversity. In this chapter I have
gathered together all published records and some of my own unpublished
findings of terrestrial arthropods in UAE. I have also added information on
arthropod groups which have not been found in the Emirates but which
probably do occur on the basis of records from neighbouring countries and
sim ilar climatic regions elsewhere. Wherever possible I have included
information on first aid for bites and stings.
A n IN TR O D U C TIO N TO THE ARTHROPODS
The arthropods form a major taxonomic division, also known as a phylum
which consists of invertebrates (animals without backbones) that possess
jointed bodies and limbs, and an exoskeleton. For more detailed information
on arthropod biology and phylogeny see Barnes (1987), Manton, (1977) or
Meglitsch (1967). The exoskeleton or cuticle can either be thin and flexible
forming the joints, or deposited in thick, stiff plates called sclerites which
function like armour. Crack open the claws of crabs or lobsters and you will
know just how strong the sclerites can be. The cuticle also acts as a barrier
against water loss and therefore helps arthropods to survive in arid
environments. Arthropods are further classified according to the similarity of
their body structure, in particular the shape of their mouthparts which reflects
the range of diets and feeding habits.
Five sub-phvla are recognised of which three, the Crustacea, the Insecta-
Mvriapoda and the Chelicerata, occur in UAE. The vast majority are insects,
which differ from other arthropods by having specialised antennae, two pms
of wings. SIX pairs of legs and compound eyes. The insects are described in a

T e r r e s t r ia l A r t h r o p o d s
separate chapter of this book. Here I shall consider the remaining terrestrial
arthropods which are more diverse although they tend to be either predatory
or parasitic animals.There are few species of terrestrial Crustacea (crustaceans) in UAE although
they are more common than might be expected of these generally mesic (ie
moderate moisture) creatures. Besides the vast array of insects, several other groups belonging to the Insecta-Myriapoda frequently occur in arid-zones like
the Emirates, particularly the Scolopendrida or centipedes. By contrast, the
Chelicerata (chelicerates) are very well-represented by various members of the
class Arachnida which again can be divided into 11 sub classes. Of these sub
classes, nine are likely to occur in UAE. They are the scorpions (Scorpiones), pseudoscorpions (Pseudoscorpiones), solifugids or camel spiders (Solifugidae), whip scorpions (Schizopeltida and Thelyphonida), tail-less whip scorpions
(Amblypygi), spiders (Araneae), harvestmen (Opiliones) and mites and ticks
(Acari). In terms of biodiversity and biomass, or the amount of biological
material determined by body size and relative numbers, the scorpions and
solifugids are the dominant groups in UAE. Some of these animals are
predatory and a few are venomous, but they are not generally aggressive
towards humans and only use their poison to kill and capture their prey or as
defensive weapons. Most stings or bites orüy occur when these animals come
into accidental contact witih man.All invertebrates are poikilothermic, or cold-blooded, so their body
temperature is dependant on that of their environment. Activity therefore is
often directly related to temperature and there are noticeable differences in
activity levels between day and night, and the summer and winter months.
Indeed, during the day, deserts appear devoid of animal life and it is not until
dusk that their many arthropodian inhabitants become apparent. Conversely,
during the winter, if the ambient temperature is too low for their basal metabolic
rate or that of their prey, mvertebrates remain inactive and will rarely be seen.
Although most arthropods have a relatively small body-size, their biology is
quite varied and many species show a high degree of specialisation.
Everything from parental care through to complex behaviour and adaptation
to environment are exhibited. In some arthropods, the juvenile stages are exact
miniatures of the adults while in others they not only look very different, but
also have totally different habitats and diets. In order to grow most arthropods
have to moult, known as ecdysis, before they can increase in size. During this
time they are very vulnerable until the new cuticle has hardened.
In terms of classification, I have tried to follow Sheals (1973) and Sheals &
Rice (1973), but there is considerable disagreement in the scientific literature
about how closely related some of the arthropods are to one another. Indeed, it is not always clear at what level they should be split and whether they are
separate phyla, classes or orders. By keeping to one system of classification I
hope to simplify matters and avoid confusion. I shall therefore start by
introducing the various groups, then deal with each in turn by describing their
general appearance, biology, ecology and natural history. Finally, for each taxa, I shall list those species known to occur in UAE. In most cases this consists of a
scientific name because they have no known common name.
Sub-phylum Crustacea
The crustaceans are a very diverse group dominated by marine and aquatic
species. In many deserts, numerous tiny brine shrimps are found in temporary
pools following the rains. These creatures are able to withstand immense
periods of desiccation and reproduce very rapidly to ensure the continual
survival of their kind. Such pools have not been studied in UAE but it is
possible that ephemeral crustaceans do occur. The only terrestrial Crustacea are
die sub-class or class isopoda.
Crustacea Isopoda
Since most isopods are either aquatic or associated with very humid
environments, it is perhaps surprising that the family Porcellionidae or woodlice is common in desert areas. These creatures have small, ovoid bodies
up to about 6mm in length, with two long antennae and a shorter pair of
antennules at the front of the head, and a thorax divided into seven segments. fs>
N

T e r r e s t r ia l A r th r o p o d s
separate chapter of this book. Here I shall conMOcr the remaining terrestrial
arthropods which are more diverse although :ne\ tend to be either predatory
or parasitic animals.
There are few species of terrestrial Crust.rce.r t crustaceans) in UAE although
they are more common than might be e\pected of these generally mesic (ie
moderate moisture) creatures. Beside,- the \ a-t array of insects, several other
groups belonging to the Insecta-M\ riapoda trequently occur in arid-zones like
the Emirates, particularly the Scolopendrida or centipedes. By contrast, the
Chelicerata (chelicerates) are \ er\ well-represented by various members of the
class Arachnida which again can be divided into 11 sub-classes. Of these sub
classes, nine are likely to occur in U.AE. The\ are the scorpions (Scorpiones),
pseudoscorpions (Pseudoscorpiones), solifugids or camel spiders (Solifugidae),
whip scorpions (Schizopeltida and Thelvphonida), tail-less whip scorpions
(Amblypygi), spiders (Araneae), harvestmen (Opiliones) and mites and ticks
(Acari). In terms of biodiversity and biomass, or the amount of biological
material determined by body size and relative numbers, the scorpions and
solifugids are the dominant groups in UAE. Some of these animals are
predatory and a few are venomous, but they are not generally aggressive
towards humans and only use their poison to kill and capture their prey or as
defensive weapons. Most stings or bites onlv occur when these animals come
into accidental contact w ith man.
A ll invertebrates are po ikilotherm ic, or cold-blooded, so their body
temperature is dependant on that of their em ironment. Activity therefore is
often directly related to temperature and there are noticeable differences in
activity levels between day and night, and the summer and winter months.
Indeed, during the day, deserts appear devoid of animal life and it is not until
dusk that their many arthropodian inhabitants become apparent. Conversely,
during the winter, if the ambient temperature is too low for their basal metabolic
rate or that of their prey, invertebrates remain inacti\ e and w ill rarely be seen.
Although most arthropods have a relativeh small body-size, their biology is
quite varied and many species show a high degree of specialisation.
Everything from parental care through to complex behaviour and adaptation
to environment are exhibited. In some arthropods, the juvenile stages are exact
miniatures of the adults while in others they not only look very different, but
also have totally different habitats and diets. In order to grow most arthropods
have to moult, known as ecdysis, before they can increase in size. During this
time they are very \ ulnerable until the new cuticle has hardened.
In terms of classification, 1 have tried to follow Sheals (1973) and Sheals &
Rice (1973), but there is considerable disagreement in the scientific literature
about how closely related some of the artliropods are to one another. Indeed, it
is not always clear at what level they should be split and whether they are
separate phyla, classes or orders. By keeping to one system of classification I
hope to s im plify matters and avoid confusion. I shall therefore start by
introducing the various groups, then deal w ith each in turn by describing their
general appearance, biology, ecology and natural history. Finally, for each taxa,
1 shall list those species kiTown to occur in UAE. In most cases this consists of a
scientific name because they have no known common name.
S u b - p h y l u m C r u s t a c e a
The crustaceans are a very diverse group dominated by marine and aquatic
species. In many deserts, numerous tiny brine shrimps are foimd in temporary
pools following the rains. These creatures are able to withstand immense
periods of desiccation and reproduce very rapidly to ensure the continual
survival of their kind. Such pools have not been studied in UAE but it is
possible that ephemeral crustaceans do occur. The only terrestrial Crustacea are
the sub-class or class isopoda.
C r u s t a c e a
I s o p o d a
Since most isopods are either aquatic or associated w ith very hum id
environments, it is perhaps surprising that the fam ily Porcellionidae or
woodlice is conunon in desert areas. These creatures have small, ovoid bodies
up to about 6mm in length, w ith two long antennae and a shorter pair of
antennules at the front of the head, and a thorax divided into seven segments. 109

N atural Emiiumes
Of the various species
of scorpions, that
occur in the UAE, the
small i/ellow
Buthacus
yotvatensis
iiigroaculeatus
is the most poisonous.
Tliey are detritivores and survive by remaining in the humid micro-climate of
burrows at the base of desert shrubs and only emerge to feed at night. They are
probably relics of a more moist environment.
T e r r e s t r ia l I s o p o d s i n U A EFerrara & Taiti (1985) and Taiti & Ferrara (1989, 1991) list 20 species of
woodlice which occur on the Arabian peninsula. Of these there are two species
introduced by man (PorcellioniAes prunniosus and Protracheneonisus inexpectatus)
and three cosmopolitan species {Porcellio assimilis, Porcellio evans/s and
Koweitoniscus tamei). All of these are likely to be found in the Emirates. Taiti &
Ferrara (1991) also describe a new species Littrorophiloscia stronhali which was
first discovered in Sharjah along with another isopod Somalodillo paeninsulae.
Isopods remain a relatively poorly known group in UAE
S u b - p h y l u m C h e l ic e r a t a
C l a s s A r a c h n id a
Most chelicerate arthropods belong to the class Araclmida and include some of
the most typical arid-zone animals. All araclmids have a pair of feeding organs
called chelicerae, a pair of pedipalps and four pairs of walking legs. The
chelicerae and pedipalps are variously modified according to biology, usuallv
reflecting diet. In structure the chelicerae resemble a pair of pincers with teeth
along their inner edge. The arachnid body is divided into two regions: the
anterior prosoma or céphalothorax and the posterior opithosoma or abdomen.
However, in the Acari and Opliones, the prosoma and opithosoma are not
clearly differentiated. Arachnids are nearly all terrestrial predators of other
arthropods. For further information on Arachnids see Savory (1964 & 1977)
and Cloudsley Thompson (1958),
C h e l ic e r a t a
A r a c h n i d a
S c o r p io n e s ( s c o r p io n s )Almost everybody knows what a scorpion looks like e\en if they have never
actually seen one. The image of their body-shape is ingrained in our minds
from earliest times, and myth and superstition surround them, Manv people
fear and loath scorpions but in common with other venomous animals they
normally avoid humans and w ill only sting when threatened. Most stings
occur when people inadvertently come into contact with scorpions, partic
ularly during camping trips, when they are found hiding under rocks, stones,
tents, clothing, debris and rubbish,
Tlie scorpions are stiucturally a very homologous group and have been highly
successful for over 450 million years. Tliey can be relatively large although some
are only 9mm long. Their bodies are heavily sclerotized with a thick cuticle
forming a compact sliield or carapace over the prosoma, Tliey have one pair of
median eyes and from two to five smaller simple eyes or ocelli, Tlieir chelate-
pedipalps or pincers are massive and powerful and are used to grasp and
manipulate prey. Tliey have four pairs of walking legs, and the basal segments of
the first two pairs are modified for chewing. Tlieir opithosoma is broad and made
up of a seven-segmented pre-abdomen and a five-segmented tail or post-roabdomen with a sting at the tip. On their ventral side is a pair of debate
comb-like organs known as pectines which are thought to be sensory in function.

T l KKIM KI \ i A i\I I IK ’( ir o n s
Scorpions prey on other arthropods,
particularly insects and arachnids but ma\
also feed on small \ertebrates. Scorpions
that ha\ e verv lar^e pincers do not al\va\>
use their sting to kill prey, but may simpK
relv on the crushing power of their claw s
The chelicerae are used to tear-up the pre\
and digestion actually starts outside the
mouth. They ha\e a long gestation period
and surprisingly are viviparous, meaning
that they give birth to li\e voung Toung
scorpions are smaller versions of their
parents. Maternal care of the \ oung ranyes
trom a few da vs to several months and lan
include complex beha\ iour such a- i n
operative feeding and burrow building.
The lifespan of most scorpions is trom tw o
to fi\e vears but in at least one speaes it
exceeds 23 years! To find out more about
scorpions I thoroughly recommend an exiellent hook edited by Cary P o l i s
(IWO).An interesting phenomenon concerning scorpions is the fact that the\
fluoresce when exposed to ultra violet iL \ i light. \'o-one knows wh\ this
happens; it is |ust a natural propeilv ot the aiticle However, it prov ides an
easy wav ot detecting scorpions and w u -ed Iw biologists to estimate their
numbers w ithout having to capture them It i- a lso an effective wav ot
avoiding scorpions when camping, althougli m -ome desert locations it is
alarming to see iust how many the LA light re' e.,1-'
SCORPIONS IN U A E\'achon (I4SM) li^ts 14 species or s u b - s p e i i e s ol ormon- tiom the Arabian
peninsula. They belong to two families: the huthul.s- .md the Vorpionidae
There are eight genera of buthids: Aiidroilonii^i,
OrlluKluni^, Panil'ullm^ and Vihiioiiioliis, but only two
genera ol scoioionids: Hriin>cor/>iiis and Scorpio. To date only buthids have
Lx'en round m the hmirates. They are .^iidrocioiiio^ cni^^iciiiido, Ap/t/i(>l’iitliih
/ 1 ,'t'rirgeccrci/-. Hiilliiicih i/eCwtcus/s iuyroiiciiU’iitii>, Coiiio>ohiillnc onilucih^,
Porol'iiiiiii- Tesmi.'.n \\ichomohi> iiiniipcctibiniiC' and possiblv Vihhoiiiolu>
‘pol’inuiiiti^. although it is not clear w hether the latter are two separate species,
or male and female of a single species. There mav well be further scorpions
aw aiting discoverv m live l laiar mountains.
and /\.pli'nipOn'rcii> are the two largest scorpions in UAf:.
\diilts of both species can measure up to l3Hmm from their head to the tip o'
I heir sting and thev can occur near human habitation. They are i]uite different
ill appearame . r.os/,//la/a is black w ith verv cliunkv claws and a highlv
A. crassicauda /s
one ot llic /ifrgi'sf
LIAP ■'lorpioih.
ixa

N a t u r a l E m ir a t e s
1 1 2
sculptured tail, while A.pten/goccrais is pale-yellow with very long, slender
pincers and a swollen disc-shaped second segment on the tail.
The most common species in sandy areas is B.y.iiigwnciilcntiis which is
mostly yellow, but is black on the last segment of tail and sting. Adults can
reach up to about 75mm in length. Tlie Vnclmiolus spp. are slightly smaller and
also yellow with a darker-coloured tip to their sting. The first Vnchouioliis spp
ever collected were from Bada Haza, Abu Dhabi. Both C.ivnbiciis and P.liosoiun
are smaller and only reach about 25mm in length. Tlie former is all vellow,
while the latter is mainly pink, except for the last t%'o segments of the tail and
sting which are brown. I have also found a fragment from the tail of another
scorpion, possibly Ortliocliiriis iniiesi, which is awaiting confirmation.
S c o r p io n v e n o m a n d s t in g s
In recent years there has been much interest in the biology of scorpions, partic
u larly their venom which is a m ixture ot some of the most potent and
biologically active compounds in the animal kingdom. The venom of all buthid
species contains powerful and dangerous neurotoxins. Of the buthids,
Audroctouus nnstralis is considered to be very venomous and symptoms of its
sting resemble strychnine poisoning. Its close relative, A.crnssicnudn. occurs in
the Emirates and should be treated with caution. Victims of stings feel a sharp
pain followed by numbness, drowsiness and an itching of the throat. This can
be accompanied by excessive saliva and the tongue becomes sluggish with the
jaw muscles contracted. If large amounts of venom have entered the blood
system, difficulties in co-ordination arise and body temperature increases
while the production of saliva and urine are reduced. Touch and sight can be
affected, with sensitivity to strong light. Tliere may also be haemorrhages and
convulsions with increasing severity. Most victims are normally out of danger
w ith in three hours but they should receive medical supervision for at least
eight hours.
Primary first aid for scorpion stings is to reassure the victim who w ill be
su ffe ring from shock. Clean the wound and then try to isolate it by
immobilising the sight of the sting. Use a firm supporting bandage but not a
tourniquet, and hold the limb up to avoid the venom going directly to the
heart. If possible keep the site of the sting cold by placing it in iced water.
Although fatalities are \'ery rare, do seek medical help, particularly in the case
of small children and invalids who are most at risk. In some cases, an anti
venom can be administered and these work well if given early enough.
However, in other cases a pain killer is all that can be given but recovery
should be rapid.
C h e l ic e r a t a
A r a c h n i d a
P s e u d o s c o r p io n e s (p s e u d o s c o r p io n s )Pseudoscorpions are similar in structure to scorpions although they lack the
post abdomen and sting. They are distinguished by their minute size and only
measure between 1 and 7mm long. Their bodies are flattened in appearance
and some species lack eyes. Their palpal chelae or claws are large, like those of
scorpions, with a swollen "hand" and a moveable finger or digit. They use
their chelae to climb up hairs of other animals. Although they normally walk
forwards, they are equally good at going backwards. Sometimes they are
gregarious and found in large groups.
Pseudoscorpions are predators and possess a poison gland at the base of
their pincers which they use to anaesthetise prey. They feed on other tiny
arthropods such as spring-tails (Collembola), book-lice (Psocidae), mites
(Acari) and silverfish (Thysanurana). Food is digested externally by a fluid
poured over the prey and the liquefied remains are ingested by the chelicerae.
Pseudoscorpions regularly clean their palps to remove remains of food so that
they can easily suck up their next meal through special grooves.
Some pseudoscorpions are phoretic which means that they use other
animals for transport and dispersal over larger distances. They do so by
attaching themselves to the legs of insects (such as flies) using their pincers.
Others live under the elytra of large beetles where they prey upon parasitic or
phoretic mites also living on the beetles. They have even been seen h e ck lin g
to beetles which then let them mount their bodies!

T (‘r k f s t r ia i A r t h r o p o d s
Despite their minute size pseudoscorpions demonstrate a tremendous
variety and complexity of lifestyle. They have silk glands and construct nests
of silk for moulting, brooding and hibernation. Tlieir courtship dances may be
very complicated. Females carry their eggs in a brood sac attached to their
genitalia and actually provide nourishment in the form of a nutritive fluid
which passes to the embryos in the brood sac. Some species exhibit parental
care with the young riding on the back of females, but thev generallv disperse
very quickly. The young are identical to adults in all but size and undergo
three moults before they are fully grown. For further information on these
minute but fascinating animals see Weygoldt (1969) and Legg & |ones (1988).
P s e u d o s c o r p io n s in UAEAlthough pseudoscorpions are generally associated with moist habitats such as
leaf litter and crevices, the families Olipiidae and Cheliceridae prefer dry
habitats and may well occur in UAE. Manhert (1980) identified se\ en species of
pseudoscorpions from the Arabian peninsula, tour of them new to science and
Manhert (1991) found 12 species from Oman alone. So tar only two uniden
tified specimens have been found in UAE. Thev were both inside a light trap
and had probablv been using a fly or large beetle tor transport
C h e l ic e r a t a
A r a c h n i d a
S o L iF U G iD A (S o l if u g id s o r c a m e l s p id e r s )
Solifugids have four pairs of long, hairy legs and enormous, well-de\eloped
jaws. They appear to ha\ e a large "head" which can be as long as the rest of
the prosoma and houses the powerful muscles needed to operate the
chelicerae. The teeth on the chelicerae consist of solid chitin (an exfremeb
strong protein) and a solifugid bite can be severe. The chelicerae are co\ ered
with numerous spines and setae which help to remo\ e solid pre\ so that the\
can ingest their liquid diet. Males are usuallv smaller than females and in some
species, their teeth are reduced to a ridge pre\ enting efficient feeding and the\
do not live long
Camel spiders are one of the fastest running arthropods. Although thev
have four pairs of legs, they run using only three pairs. The first pair of legs or
pedipalpi are held up in front of them and used in a similar manner to the
antennae of insects. Thev have very long, silky setae and are constantly
moving in order to locate and pick-up prey. On the underside of the last pair of
legs are five malleoli or racquet organs which are thought to be sensory in
nature and function like the pectines of scorpions. They use their second pair
of limbs as rakes to push loose soil when constructing burrows and move the
soil in rapid mox ements, looking like little bulldozers'
Despite their fearsome appearance and their strong bite, solifugids are
unlikely to harm humans. In the past they were considered venomous and
extremely dangerous but it is now thought that the only risk of injury resulting
from them is caused by shock or infection follow ing a bite. There is no
evidence of venom m any part of their body.
Camel spiders are nocturnal predators of other arthropods including
scorpions and are \ oracious teeders. Some species kill and feed on lizards and
it is speculated that others kill mice and birds. They rely solely on their speed
and stealth to catch their prey. In desert areas they are often attracted to lights
Civiicl spiders iirc
ciillcd solifiiyids.
bcciiiisc tlici/ nvoid
('ciiiy out III Ilk' >1111.
It onciiL'cidciitnlli!
UIICOVCIS II Lllllll'l
>pidcr, it will
friiiiticnlhi look for
■blinde, u>iinlhi found nt
oiicd foot Tins clinriK-
tcri>iic lins y i i ’iTi it on
tiiidc>i'ivcd ropiitnlioiiK>
eUiygressieii w

Camel spider,
Solifugid galeodes.
1 1 4
at night in search of food and their appearance can cause alarm if they enter
tents. It IS rare to see them during the winter months in UAE and they are
thought to hide or hibernate during cold periods.
Females dig a burrow in which they lay over 200 eggs. They guard then-
young for two to three weeks until the first moult. Although they are quite
large, solifugids are thought to live for less than one year.
C a m e l s p id e rs in UAEDespite their conspicuous appearance and size, little is known about these
creatures and we are not e\ en sure of their exact identity. There are probably
three families of solifugids in UAE. These are the long-legged and sandy-
coloured Galeodidae and Solpugidae, and the black-coloured Rhagodidae
which has shorter legs and is better adapted for digging. The Galeodidae are
commonly seen in sandy areas and the largest species is probablv Galeodes
arabs which is particularly hairy and bulky with limbs spanning up to l^Omm
Other species probablv occur in the mountains and Rhagodidae are thought to
occur around .Al Ain
C h e l ic e r a t e s
A r a c h n i d a
S c h i z o p e l t i d a , T h e l y p h o n i d a a n d A m b l y p y g i
( w h ip s c o r p io n s a n d t a il - le s s w h ip s c o r p io n s )These obscure arachnids are very poorly-known and superticiallv resemble
scorpions but lack a sting. However, the whip scorpions, Schizopeltida and
Thelyphonida, have slender, whip-like appendages or tails and exhibit a
defensive reaction, by discharging a caustic, dust-like cloud trom an anal
gland, which smells strongly of acetic acid or chlorine gas. Schizopeltida are
similar in habits to scorpions but are only 5-7mm long. They are nocturnal and
hide under stones or in burrows during the day. Thelyphonida are relatively
large, nocturnal predators feeding on a wide variety of arthropods. They are
quite ferocious in appearance and although their chelicerae arc not claw-like
their pedipalps are very powerful. The Amblypygi can be relatively large (8-45
mm in length) and are flattened in appearance. They lack a tail and are
sometimes called tail-less whip scorpions. They are also nocturnal predators
and have raptorial pedipalps armed with strong spines and a moveable hook
with which they grab their prey.
There are no known records of these arachnids in UAE but Schizopeltida are
common m other arid regions and may have been over-looked. Thelyphonida
and Amblypygi are generally found in the tropics and could occur in moister
en\'ironments such as wadis.
C h e l ic e r a t a
A r a c h n i d a
O p il io n e s ( h a r v e s t m e n )
Opiliones or harvestmen are cosmopolitan predators inhabiting a wide range
of habitats. They superficially resemble spiders but their legs are very long and
spindlv and the prosoma is not divided from the opithosoma. They teed on
dead nr recently dead tissues and probably require a humid environment.
Ehey have not been recorded from UAE, but probably occur in t l^ more
humid areas.

C h e l ic e r a t a
A r a c h n i d a
A r a n a e a
Aranaea are the true spiders and arid-zone species tend to be cryptically
coloured, often with brushes of hair on their undersides to help them move
through sand. Spider chelicerae are hook-like with moveable digits that carry
poison glands, but most are harmless because their chelicerae are too weak to
pierce human skin. They are predators and are further classified into separate
families according to their anatomy and biology which reflect the way in
which they capture their prey. For example, some spm webs and sit and wait
for their prey while others are active hunters. Spiders are found world-wide
and occur is all types of habitats.
S p id e rs in UAEAt present little is known about spiders m the Emirates but current investi
gations by the American Museum of Natural History w ill soon tell us more.
However, the spider fauna of the Sahara region is known, and includes a wide
range of spider families such as ground hunting spiders (Giwtliosidni’), crab
spiders (Thomisiiiae), giant crab spiders (Spnrnssiiine), bark spiders {Hcrsiliulne),
jumping spiders (Salticidae), wolf spiders {Uicosidae), sheet-web spiders
{A^ekuidae), comb-footed spiders (Tlieridiidiu'} and orb-weavers {Tetrn^uatliuini
and Ar^wpidiie). All these families probablv also occur in the Emirates
Spiders of the Arabian peninsula as a whole are not well studied. Twenty-
nine species of Salticidae have been found, many of which were undescribed
but resembled African or Asian spiders (PrÛszynski,1993). Eight species from
S I X genera of crab spiders or Thomisidae also occur (Dippenaar-
Schoeman,1989). One of these, Thouiisiis dtnnelliis, has a wide distribution and
probably inhabits UAE. In Saudi Arabia, two species of Linyphiidae have been
recorded although they are not normally considered arid-zone species
(]ocquE,1981).
B l a c k w id o w s ( T h e r i d i i d a e ; L a c t r o d e c t u s s p p )
Two species of black widow are common in the Middle East, Lactrodectus
pallidiis, occurring from Libya to Azerbaidjan, and Lactrodectus Injsterix, known
from Aden and Yemen. In the Emirates, black widows occur along the coastal
strip and hide in rubbish or objects left outside overnight. Care should
therefore be taken when picking up debris etc. Their bite is rarely felt but is
noticed later when the wound starts to swell and two tiny spots become visible
where the fangs have penetrated the skin. The bite is very painful and the
whole body can ache especially the legs. Other symptoms include shock, fever,
nausea, headache, raised blood pressure, difficulty in breathing and heavy
sweating. However most bite victim s
recover completely w ith in two
days. If bitten seek medical
attention and always reassure
the v ictim . Treatment
includes intravenous
salts and pain
kille rs which
alleviate
symptoms
The crab spider,
camouflaged by its
colour, waits at the
heart of a floioer, to
catch Its prey.

Natural Emirates
The velvet mite oiihf
nppeni'S nbove pinmmi
after rams.
1 1 6
C h e l ic e r a t a
A r a c h n i d a
A c a r in a o r A c a r i ( M it e s a n d t ic k s )The Acari are the smallest chelicerates hut are also the most numerous world
wide in terms of the number ot species. Thev are commonly known as mites
and ticks. Adults have four pairs of legs while the voung have three pairs
Thev include both parasites and predators which generallv live on other
animals or inside plants, although some inhabit the soil. Seven orders of Acari
are recognised: Notostigmata, Tetrastigmata, .Mesostigmata, Metastigmata.
Cryptostigmata, Astigmata and Prostigmata.
The Notostigmata and Tetrastigmata are predatory mites frequenting
warmer regions ot the world, and probablv live in the Emirates. The other
orders are cosmopolitan and probably occur, but are relatively unknown
because of their small size. However, the Metastigmata nr ticks arc better
known because thev are ectoparasites of terrestrial vertebrates and some are
important pests of livestock and vectors of disease. Crvptostigmata are verv
small and dark in colour and live in soil and leaf litter where thev feed on dead
and decaying plant material. Astigmata are small and generallv feed on fungiw
or detritus, but thev also include some parasites of vertebrates known as fur
and feather mites. Prostigmata are more heterogeneous and include tree-living
predators, phytophagous mites, parasites and aquatic mites.
Ticks are divided into two types: the hard ticks or Ixodidae and the soft ticks
or Argasidae. Ixodids have a hardened dorsal scutelum which is absent in
argasids. Argasids are most abundant in dry regions. The family Argasidae
contains two genera: Ar^as which are associated with birds, bats or their
nesting/resting places, and Oniitliodorus which frequent burrows, corrals or
houses. The larger Ixodidae fam ily comprises six genera: Hualomma.
Amblpomma, Rliicicephalus, Boopliihis, Apoiwmma and Haemapbpsalh
Tick bites can be painful and irritating, and severe cases ot multiple tick
infestation in a single animal, can result in anaemia, toxic reaction and
paralysis. However, it is the tick's potential as a vector ot pathogenic disea-^c"
that is more .seriou.- . Although man is onlv an incidental host and their natuia'
hosts are wild animals,
some of the tick species
present in Arabia can
t r a n s m 1 1 h u m a n
d i s e a s e s . M a n \
po ten tia lly infected
ticks are accidentally
im ported along w ith
foreign livestock but
are equally able to feed
on local livestock and
c o u l d e s t a b l i s h
permanent populations.
If thev are infected with agents pathogenic to humans or domestic animaH
these may also be spread by local ticks. Although cases ot tick-borne human
disease are rare there is an element of risk.
T ic k s f r o m U A E
There are no published records of ticks for the Emirates but the following
species are native to Arabia (Hoogstraal et al, 1984) and therefore likelv to be
part of the local tick fauna. Hi/alomma In/alomma aiiatolicitm aiiatoliciim is verv
common and its hosts include domestic stock, lizards, rodents, hedgehogs,
hares and humans. It is the ma)or vector of Crimean-Congo Haemorrhagic
Eever (CCHF) in the Southern Soviet republics, Pakistan and Nigeria. Othei
common Hvalomma ticks include H.li.ilromedani which is both a vector tor
CCHE and a natural reservoir of Q-tever; and H.li.impeltatiim and
H.li.mar<^iiiatiim turcanicum which have also been implicated as vectors tor
CCHF The kennel or brown dog tick, Rliipicephalus rliipicephaius siutytimciis,
generallv feeds on dogs as its name suggests, but can carry CCHF as can the
closely related R.r.tiiranicus. Another vector of CCHF is Roophilus aimiilatus
although its preferred hosts are goats. Human cases of CCHF are verv^re in
the Emirates, but it has been reported from Dubai.

O t h e r m ite s in UAEProbablv the largest mite in UAE is the giant velvet mite,
Diiwntlironibiiiiti sp. Adult mites measure up to 12mm and are covered in a
thick and dense scarlet tur which warns predators that they are distasteful.
Adults onlv emerge after heavy rain when thev wander around sandy areas m
search of termites. I have found them on se\ eral occasions near Sweihan and
Al Samha. Their larvae are parasitic on grasshoppers.
Oribatid mites are one of the richest soil arthropod groups both in terms of
numbers and species diversity. Bavoumi & Al-Khalifa (1985) report 48 species
from Saudi Arabia but thought that the number of species was limited bv soil
humidity.
I n s e c t a - M y r ia p o d a
The Insecta-mvnapoda are the most niimericallv important group of animals
in the UAE. As well as the insects this group includes millipedes (Diplopoda),
centipedes (Chilopoda or Scolopendrida), and the lesser known Pauropoda
and Symphyla. The name Myriapoda reflects the fact that these animals have
manv legs and, as w ill be seen, they are a di\ erse group.
I n s e c t a - M y r ia p o d a
D ip l o p o d a
The diplopoda are commonly known as millipedes. Their body is made up ot a
large number of double-sided segments each bearing two pairs of legs. Thev
are slow moving detriti\ ores and are usuallv found in leaf litter and rotting
vegetation. Millipedes are most abundant in the tropics and have not been
recorded from UAE but probabh' occur m relativeh' humid areas such a
wadis, parks and gardens.
I n s e c t a - M y r ia p o d a
C h il o p o d a o r S c o l o p e n d r id a
( C e n t ip e d e s )
There are three fam ilies of centipedes; Ceophilom orpha,
Scutigeromorpha and Scolopendromorpha. However, only the latter occur in
the Emirate^, Scolopendrid centipedes have an elongated, dorso-ventrallv
flattened hod\ w ith a distinct head. The head bears a single pair of antennae
and three pairs ot feeding appendages (a mandible and two pairs of maxillae).
The trunk can ha\ e trom 15 to over 10Ü segments each bearing a single pair of
legs which results in their characteristic locomotion, and centipedes can measure
up to 12ilmm m length The first pair ot trunk appendages is known as the
maxillipedes or toxognatha and has powerful terminal claws at the tips, which
have ducts leading to poison glands. Centipedes cannot usuallv pierce human
skin and are unlikely to be poisonous but it is best to treat them with respect
The\ are generallv nocturnal predators and feed on other smaller
arthropods. During the day thev hide under rocks, stones and debris. Female
Scolopendrids care tor their voung bv making a protective basket between the
body and legs, formed bv curling their legs around the voung.
C e n t ip e d e s in UAELewis & Callagher (1993) mention \ arious records of scolopendrids tor U.AE
although the\ probablv occur in most areas. Scolopciidiidn iiiinibilis has been
found in Masah, Sharjah and jebel Faivah while Scolopciidiidn volidn was collected
near Sharjah. Both species are thought to be very resistant to desiccation.
C e n t ip e d e b it e s
The medical importance ot centipedes was probabh oxerestimated in the past
since onh \er\ old records mention human fatalities (Lewis, 1986). However,
the bite nt S lunnhili^ is though to be like that ot a s im ila r genuso
17

N atu r a l Em irates
Many interesting Tmchycormocephahts which causes pain, swelling and
spiders remain as yet subcutaneous bleeding. The area around the bite is
unidentified. particularly tender but most symptoms disappear
within 24 hours. First aid treatment for a centipede bite
is similar to that for scorpion stings although it is less
likely to be life threatening
I n s e c t a - M y r ia p o d a
P a u r o p o d a a n d S y m p h y l a
These two groups of myriapods are less common fhan
the others. They are both multiple-limbed, small and
soft-bodied. They live m soil and leaf litter where they
are thought to feed on fungi and detritus, although the
Symphyla also feed on living plant material. Little is
known of their biology and distribution and they have
not been reported from UAE, but they could occur in
the soil fauna of moderafely moist environments.
118
I n s e c t a - M y r ia p o d a
S u b - p h y l u m o r c l a s s T a r d ig r a d a
I could not finish this chapter without mentioning the tardigrades. These
minute animals only measure 50-500pm in size and cannot be seen without the
aid of magnification. They have six legs ending in claws and are commonly
known as water bears because of their rotund appearance. They live in the
siuface film of water covering mosses and in fresh and salt water. Since they
can withstand prolonged periods of desiccation they could easily occur in
temporal rain pools or beaches in UAE. To my knowledge, there is no
published research work on the desert tardigrades but they would be an
interesting subject for study.
Arthropods are very successful and many are able to flourish m the dry
climate of the UAE either through adaptation to their environment or by
behavioural avoidance of extreme conditions. They include a few defritivores
and herbivores, but are predominantly predators. However they are all
potential prey for vertebrates such as birds, mammals and reptiles. The larger
species, such as scorpions and camel spiders, are particularly good sources of
food for fhose able to catch them. However, there sHll remains much that we
do not know about them, including which species actually occur in the
Emirates. There are probably numerous new records and species waiting to be
discovered all over Arabia. Next time you come across an arthropod be content
to watch and admire it for its powers of survival.
A c k n o w l e d g e m e n t s
I thank Dr john Balfour for information on public health pests. Dr Vojin Sjlivic
for discussions about scorpions stings and Dr Patrick Osborne for commuting
on an earlier draff of the text.

R e f e r e n c e s
Hoogstraal, H., Wassef, H. Y. & Buftiker, W. (1984). Ticks (Acarina) of Saudi Arabia.
Family Argasidae. Ixodidae' F n iii in o fS n iid i A rn h in , 3. 23-110
Barnes, R.D. (1987). I in ' t r h ' l ’n i t i ' Z o o h j^ /. Fifth edition. Saunders College,/Holt, Rhinehart
and Wilson, USA.
Bayoumi, B. M. & Al-Klialifa, M. S. (1985).' Oribatid mites (.Acari) of Saudi Arabia'. F n iiiw
o f S aud i A rn b in , 7,66-92.
Cloudsley Thompson, J. L. (1958). Spiders, scorp ions, ic n fip e d rs an d m iles. Pergamon Press,
London.
D ippenaar-Schoeman, A. S. (1989). 'A n annotated check lis t ot Crab Spiders
(Araneae:Thomisidae) of Saudi Arabia'. Fauna of Saudi A ra b ia ,1 0 ,20-30
Ferrara, F. & Taiti, S. (1985). 'The terrestrial isopods (Oniscoidea) of the Arabian
Peninsula'. Fauna o f S aud i A ra b ia , 7 , 93-121
Polls, G. A. (ed.) (1990). The b io logy o f scorp ions. Stanford University' Press, USA.
JocquE, R. (1981) 'Araneae:Fam. Linyphiidae'. Fauna o f S aud i A ra b ia . 3,111-113.
Legg, G. & Jones, R. E. (1988). 'Pseudoscorpions'. S pnops is o f the B r it is h F auna (New
Series) Kemack, D. M. & Barnes, R. S. K. (eds.) No. 40. W BackJnivs, Leiden,
Lewis, j. G, E.(1986). 'Chilopoda ot Saudi .Arabia: centipedes of Saudi Arabia'. Fauna ot
S aud i A ra b ia , 8, 20-30
Lewis, j. G. E. & Gallagher, ,V1. D. (1993). 'Scolopendromrph and Geophilomorph
centipedes trom Oman and UAE'. Fauna o f S aud i A ra b ia , 13, 55-62.
Manhert, V. (1980). 'Arachnids of Saudi Arabia: Pseudoscorpiones'. Fauna o f Saudi A rab ia ,
1,32-48.
Manhert, V. (1991). 'Arachnids of Saudi Arabia. Pseudoscorpions (Arachnida) from the
Arabian Peninsula'. Fauna o f S aud i A ra b ia , 12,171-199
Manton, S. M. (1977). The A r th ro p o d a - h a b its , fu n c t io n a l m o rp h o lo g y a n d e v o lu t io n
Clarenden Press, Oxford.
Meglitsch, P. A. (1967). In ve rte b ra te zoology. Oxford University Press, Oxford.
PrÛszynski,]. (1993). 'Arachnids of Saudi Arabia: Salticidae (Araneae) of Saudi Arabia II'
Fauna o f S aud i A ra b ia , 13 ,27-54.
Savory, T. H. (1964). A ra ch n id a . Academic press, London.
Savory, T. H. (1977). S p id e rs a n d o th e r a ra ch n id s The English Universities Press Ltd,
London.
Sheals, J. G. (1973). 'Arachnida'. In: Smith, K. G. V. (Edi Insects an d o th e r a rth ro pods ot
m ed ica l im p o rta n c e , pp 417-472. Trustees of the British Museum (Natural History),
London
• %
- - ' J '
%
- V
■ . , 4. ^■* ■ ( A
Sheals, j. G. (1973). 'Other Arthropods' In: Smith, K. G. V. (Ed) Insects a n d o th e r a rth ro pods
o f m e d ica l im p o rta n c e , 473-482. Trustees of the British Museum (Natural History),
London
Taiti, S. & Ferrara, F, (1989). Terrestrial isopods of Saudi Arabia (Part 2)'. Fauna o f S aud i
A rab ia , 10, 78-86.
Taiti, S. & Ferrara, F (1991). '.New species and records of terrestrial isopods (Crustacea)
trom the Arabian Peninsula' F auna o f S aud i A ra b ia , 12,209-224.
\achon, VI. ( 1989) Arachnids of Saudi Arabia. Scorpions'. Fauna o f S aud i A ra b ia , 1, 30-65
IVevgoldt, P (1969). The biolosiu ot iise iidosco rp io iis . Harvard University Press, Harvard.119

243
Class Order (Division) Family(subfamily)
Genus & Species (if known) Methodof
capture
Verifiedby
Arachnida Scorpiones Buthidae *Androctonus crassicauda (Olivier) p JB
Arachnida *Aplstobuthus pterygocercus Finnegan p JB
Buthacus yotvatensis nigroaculeatus p JB
Levy et al.Compsobuthus arabicus Levy et al. p JB
Parabuthus liosoma (H. & E.) p JB
*Vachonioius minipectinibus (Levy et p JB
al.)Vachonlolus globlmanus Vachon p JB
*Nr Odhochlrus InnesI Simon F BJT
Solifugae *Ga!eodes arabs Koch P, H PH
*Galeodes? spA. P PH
indet. (not Galeodes) P PH
Araneae Gnaphosidae *Pterotrlcha sp A. P LS
Araneae Gnaphosidae *Pterotrlcha sp B. P LS
*Lachesana sp P LS
Lycosidae indet. P PH.LS
Salticidae indet. P PH.LS
Theridiidae indet. P PH, LS
Philodromidae indet. P PH.LS
Sparassidae indet. H PH
Eresidae *Stegodyptus llneatus P PH
Acarina Ixodidae Hyalomma sp. P JB
Caeculidae indet. P AB
Trombidiidae DInothromblum sp. H AB. BJT
Entognatha Colembolla Arthropleona indet. P BJT
Insecta Thysanura Lepismatidae Nr Thermobla domestica Packard P JB
indet. P JB
Odonata Aeshnidae Hemlanax ephlpplger Burmeister L JB
Orthoptera Gryllotalpidae Gryllotalpa africana Pal. L JB
Gryllidae Acheta domestica Linnaeus H JB
Indet. P JB
Pyrogomorphidae Pyrgomorpha indet. L GP
Pyrgomorpha conica- bisplnosa gp. L GP
Chrotogonus homalodemus H GP
(Blanchard)Tenultarsus angustus (Blanchard) H GP
Acrididae Derlcorys cyrtostema Uvarov L GP
Acorypha glaucopsis (Walker) L GP
Heteraciis annulosa (Walker) L GP
Heteraciis llttoralls Rambur L GP
*Hyalorrhlpls canescens (Saussure) H GP
Sphlngonotus rubescens (Walker) H GP
Acrotylus longlpes (Charpentier) H GP
Truxalls procera Klug H GP
*Ochrllldla tibialis (Fieber) H GP
Ochrllldia genlculata (Bolivar) L GP
*OchrlHdla persica Salfi L GP
*Leptoptemls gracilis (Eversmann) H GP
"Tridactylus fasclatus Guerin L GP
Tettigoniidae Nr Platyclels sp. nymph P GP
*Conocephalus concolor (Burmeister) L GP
*Ruspolla nitldula (Scopoli) H GP
*Dlogena fausta Burm L GP
Dermaptera Labiduridae *Labldura ripaiia (Pallas) L JB
Mantodea Eremiaphilidae *Eremlaphlla gene Febv. P JB
Mantidae *Blepharopsls mendica Fab. H JB
*Rlvetlna Inermis Uvar. H JB
*Mlcrothespls dmltiiewl VJern. L JB
Ins pitched Kalt H JB
*Hypslcorypha gracilis Burm. H JB
Appendix 4.1 List of species caught in UAE (1992-1996).

244
Class Order (Division) Family(subfamily)
Genus & Species (if known) Methodof
capture
Verifiedby
Insecta Mantodea Mantidae *Sinaiella sabulosa Uvarov L JB
Mantodea Mantidae *Sinaiella nebulosa Uvarov L JB
Isoptera Rhinotermitidae Psammotermes hybostoma Desneaux P JB
Indet. Indet. P JB
Phasmatodea Phasmatidae *Ramulus sp. H JB
Hemiptera(Homoptera) Fulgoridae Indet. P MW
Dictyopharidae Indet. P MW
Cicadidae Platypleura arabica Myers H, L BJT
Aphididae Indet. H JB
Hemiptera(Heteroptera) Tingidae *Phaenotropis Cleopatra (Horvith) L GS
Reduviidae *Reduvius sp. P GS
*Putoniola sp. P JB
Miridae *Eurysty!us bellevoyei (Reuter) H GS
Indet. H GS
Lygaeidae Indet. L GS
Coreidae *Homoeocerus varlabills (Dallas) P GS
Cydnidae *Amaurocoris aspericollis Puton P GS
Dinidoridae Cordius viduatus Fabricius H, L BJT
Pentatomidae *Chorantha ornatula (H-S) L GS
*Bagrada sp. P GS
Neuroptera Myrmelionidae *Tomatarella markii Kimmins L JB
*Paipares cephalotes Klug L JB
*Neuroleon parvus Kimmins L JB
*Neuroleon nr nemausiensis Bork. L JB
*Myrmeceiaurus nr caudatus Navas L JB
*Gepus invisus Navas L JB
*Nr Cueta sp. L JB
Lepidoptera Sphingidae Agrius convolvuli Linnaeus H BJT
Acherontia styx Westwood H BJT
Macroglossum stellarum Linnaeus H BJT
Daphnis netil Linnaeus H BJT
*Hyles llvomica Esper L BJT
HIppotlon celeiio Linnaeus L BJT
Noctuidae *Agrotls biconica Koilar L JB, EBW
*Hellcoverpa nublgera (H.-S) L JB
*Amlcta munna Kiug H, L EBW
*Anumeta eberti zaza Wiitshire L EBW
*Anumeta spllota Wiltshire L EBW
*Cerocala sana Staudinger & Rebei L JB
Lasiocampidae *Chrondrostega fasclana felsall L EBW
WiitshireStreblote sIva Lefèbvre BJT
*Lambessa decolorata L EBW
Cossidae "Lamellocossus arias Pungeler L JB
*Lameiiocossus cheesman/Tams L JB
Pyralidae *Hercynodes afflnis Rothschiid L JB
*Epizonara confusalls Hampson L JB
"Epizonara sinaica Rebei H MS
Microlepidoptera Indet. P JB
Diptera Therevidae indet. L JO
Tephritidae *Gonlurellla tridens (Hendei) H IW
Asilidae *Apociea femoralls Weidemann H NW
Bombyliidae Anthrax sp. H NW
*Heteropus sp. H JO
Ephyridae *Ephydra sp. H NW
Sarcophagidae *Blaesoxlpha sp. H NW
Galliphoridae *Vlllenuevlella sp. H NW
Sphaerocideridae Indet. L JB
Appendix 4.1 Continued List of species caught in UAE (1992-1996).

245
Class Order (Division) Family(subfamily)
Genus & Species (if known) Methodof
capture
Verifiedby
Insecta Diptera Sphaerocideridae nr Telomerina sp. L NW
Muscidae Musca domestica Linnaeus M. L BJT
Heliomyzidae *Trichoscei!s sp H NW
Ceratoponogidae Cullcoides mesghalil Naval H JB
Hymenoptera Bethylidae Indet. L AW
Braconidae, Indet L JB
RogadinaeScoliidae *Campsomeneiia thoracica Fabricius H
Tiphiidae Indet. L JB
Hymenoptera Mutillidae Indet. sp1 P AW
Indet. sp2 P JB
Indet. sp3 P JB
Braenidae Indet. P
Bradynobaenidae *Pterogyna savigni Klug P JB
Indet. P JB
Formicidae(Ponerinae) *Pachycondy!a sennaarensis (Mayr) H.B CAC
(Myrmicinae) *Cardiocondyia emery! Forel H,B CAC. DA
*Cardlocondyla gallagheri Collingwood H.B CAC
& Agosti*Cardiocondyla stiuckardi Forel H .P CAC. DA
*Crematogaster aegyptiaca Mayr P CAC
Crematogaster antaris Forel P CAC
*Crematogaster mosis Emery P CAC
*Leptothorax //V/ae Collingwood & P.H CAC. DA
AgostiMessor ebeninus Santschi P CAC
*Messor forel! Santschi P CAC
*Messor h/sma/Collingwood & Agosti P CAC
*Messor mendionaHs (André) P CAC
*Messor muscatus Collingwood & P CAC
AgostiMessor rufotestaceus (Foerester) P CAC
Monomorium abeille! André P CAC
*Monomorium arenlphllum Santschi P CAC
*Monomorlum barbatulum Mayr P CAC
*Monomorlum buxtoni Crawley H,B CAC
*Monomorlum chobauti Emery P CAC
*Monomorium destructor (Jerdon) H CAC
*Monomoiium fezzanense Co\\\ri^ood P CAC
& Agosti*Monomorlum gallagheri Collingwood & P CAC
Agosti*Monomorlum Indlcum Forel H CAC
*Monomorlum mayr! Forel P CAC
*Monomorlum m/ntmbe Collingwood & P CAC
Agosti*Monomor!um nllotlcum Emery H CAC
*Monomorium tumaire CoWmgwood &. P CAC
Agosti*Monomorlum wahlblense CoW'mgwood P CAC
& Agosti* Pheldole megacephala (Fabricius) H.B CAC
*Pheldole sculpturata Mayr H.B CAC
*Pheldole tenerlffana Forel H.B CAC
*Solenopsls gemlnata (Fabricius) H CAC
*Tertramoiium blcaiinatum (Nylander) H CAC.DA
*Tertramorium calldum Forel H.B CAC
*Tetramoiium juba Collingwood H.P CAC
(Dolichoderinae) *lrldomrmex anceps (Roger) H.B CAC
*Leneplthema humlle (Mayr) H.B CAC.DA
*Taplnoma simrothi Krausse H.B CAC.DA
*Taplnoma megacephalum (Fabricius) H .B CAC.DA
Appendix 4.1 Continued List of species in UAE (1992-1996).

246
Class Order (Division) Family(subfamily)
Genus & Species (if known) Method Verified of by
capture
Insecta Hymenoptera
Coleoptera
PompilidaeVespidaeSphecidaeSphecidaeApidaeCarabidae
StaphilinidaeCicindelidae
DytiscidaeNitulidaeMeloidaeCoccinelidae
Curculionidae
(Formicinae) 'Camponotus compressus (Fabricius)*Camponotus fellah Dalla Torre
*Camponotus oasium Forel 'Camponotus sehceus (Fabricius) Camponotus thoraclcus Fabricius
Camponotus xerxes Forel *Cataglyphis acutinodis Collingwood & Agosti*Cataglyphls adenensis (Forel) 'Cataglyphis albicans (Roger) "Cataglyphls arenarius Finzi *Cataglyphls cinnamomeus Karawaiew
*Cataglyphis flavobrunneus Collingwood & Agosti Cataglyphis lividus (André)Cataglyphis minimus Collingwood Cataglyphis niger (André)Cataglyphis sabulosus Kugler
"Cataglyphis urens Collingwood
"Cataglyphis viaticus (Fabricius) "Lepisiota gracillicornis (Forel) "Lepisiota nigrescens Karawaiew
"Lepisiota opaciventris (Finzi) "Lepisiota spinisquama (Kunetsov- Ugamsky)"Paratrechina flavipes (Smith) "Paratrechina jaegerskioeldi (M ayr) "Paratrechina longicornis (Latreille) "Polyrhachis lactipennis F. Smith
Indet."Vespa orientalis Linnaeus
Bembix sp Indet.."Pseudapsis n Hot ica Smith Thermophilum (=Anthia) duodecimguttata Bonelli "Scarites guneensis Dejean
"nr Harpalus spl "nr Harpalus sp2
Indet."Salpingophora nr ruppelli (G -M ) "Lophyridia aulica Dej."Eretes sticticus Linnaeus
Indet.Indet.Coccinella septempunctata Linnaeus "Hyperaspis vinciguerrae Capra
"Elasmobarls nr alboguttata (Brisoutj "Lixus (Phillixus) sp."Ocladlus sp."Bothynoderes anxius (Gyllembal) "Ammocleonus aschabadensis Faust "Gronops nr pallidulus "Bagous cyperorum Bagoussp.*Nr Xylinophorus sp.*Nr Pseudostytyphlus sp.
(Entiminae) indet. Tanymeclnl
Chrysomelidae "Pseudocolaspls sp."Macrocoma leprieuri Lefevr.
p
p
p
p
p
p
p
p
p
P, H
p
P, H
p
p
H
P, H
P. H
HH.B
P, H
H.B
H,B
H.B
H,B
H.B
H,B
L
H
ML
L
P
P
P
P
P
H
H
L
P
P
H
H
P
L
P
H
H
P
P
P
P
P
P
H
H
CAC.DA
CAC
CAC
CAC
CAC
CAC
CAC, DA
CAC
CAC, DA
CAC
CAC, DA
CAC, DA
CAC, DA
CAC, DA
CAC, DA
CAC, DA
CAC, DA
CAC, DA
CAC
CAC
CAC
CAC
CAC
CAC
CAC
CAC
JB
BJT
AW
JB
JB
JB
JB
JB
JB
JB
JB
JB
JB
JB
JB
BJT
JB
RT
AT
AT
AT
AT
AT
AT
AT
AT
AT
AT
SS
ss
Appendix 4.1 Continued List of species caught in UAE (1992-1996).

247
Class Order (Division) Family(subfamily)
Genus & Species (if known) M ethodof
cap ture
Verifiedby
Insecta Coleoptera Elateridae *nr Heteroderes sp. L JB
Histeridae *nr Saprinus chalcites ill. P JB
Tenebrionididae *Prochoma nrclypealis Blair P JB
*Prochoma bucculenta Koch P JF
*Microtelus nejdanus Kaszab P JF
*Ammogiton sonyae Kaszab P JF
*Nr Falsocatomulus sp. P JF
*Foleya brevicomis Peyerimhoff P JF
Pimelia arabica Klug P JB.JF
*Pimella arabica s.sp.? P JB
*Pimelia arabica edomita Kaszab P JF
*Pimeiia ionguia Kweiton P JF
*Parapiatyope popovi Koch P JB, JF
*Biaps koiiari Seidlitz P JB. JF
*Adesmia khaiiensis Blair P JB
Adesmia stoekieini Koch P JB
*Adesmia arabica Reitt P JB
* Adesmia persiana Reitt P JB
*Tentyrina paimeri (Crotch) P JB, JF
*Sceiosodis besnardi P JB
*Arthrodibius cicatrix P JB
*Zophosis migneauxi Deyr. H JB
*Cyphostethe nr saharensis Chob. M JB
*Phaeotribon/Pachycera P JB
*Prochoma sp. P JB
*Phaeotribon puicheiium P JB
*Gonacephaium rusticum (Olivier) P JF
*Gonacephaium setuiosum P JF
(Falderm ann)*Anemia sardona (G ene) P JB, 88
*Apentanodes sp1 P JB
Mesostena puncticoiiis Sol. P JB, JF
*Oxycara breviuscuium Fairm. P JB
*Trichosphaena arabica Kaszab H JB
*Nr Phaeotribon P JB
Pachycera pygmaea arabica Kaszab P JF
Pachycera sp P JB
*Erodius reichei Allard P JB.JF
*Erodius nr octocostatus Peyerimhoff P JB
*Erodius sauditus Kaszab P JB, JF
*Arthrodibius cicatrix (Fairm ) P JB
Trachyderma(=Ocnera) phiiistina P JB
Reiche*Trachyderma striatogranosa P JF
(Fairm aire)*Apentanodes arabicus (Kirchberg) P JB, JF
*Prionotheca coronata ovaiis Ancey P JF
*Prionotheca coronata Olivier P JB
*Akis eievata Solier P JB, BJT
*Akis eievata f. scuiptior Koch P JF
*Eurycauius granuiatus Reitt. P JB
*Eurycauius buettikeri Schawaller P JF
*Leichenum mueiieri Gridelli P JF
Zoptiosis migneauxi Deyr P JB
Nr Stenosis ?sp P JB
Cerambycidae Acanthophorus arabicus Thom s H BJT, JB
Buprestidae *Juiodis whithiiii Gray H JB
Juiodis sp. H JB
Acmaeodera? Sp. P JB
*Chrysobothris parvipunctata Obenb. M JB
A ppend ix 4.1 Continued List of species caught in UAE (1992-1996).
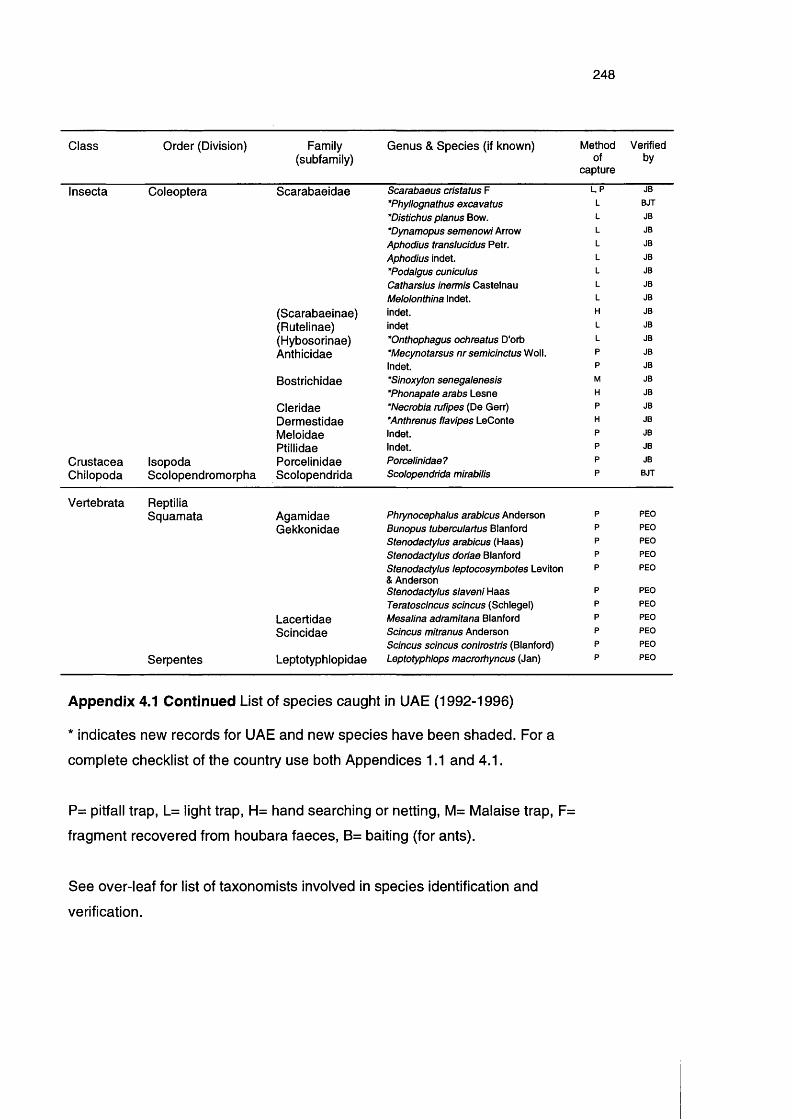
248
Class Order (Division) Family Genus & Species (if known) M ethod Verified
(subfamily) of bycap ture
Insecta Coleoptera Scarabaeidae Scarabaeus cristatus F L. P JB
*Phyllognathus excavatus L BJT
*Distichus planus Bow. L JB
*Dynamopus semenowl Arrow L JB
Aphodius translucidus Petr. L JB
Aphodius indet. L JB
*Poda!gus cuniculus L JB
Catharslus inermis Castelnau L JB
Melolonthina indet. L JB
(Scarabaeinae) indet. H JB
(Rutelinae) indet L JB
(Hybosorinae) *Onthophagus ochreatus D'orb L JB
Anthicidae *Mecynotarsus nr semicinctus W oil. P JB
Indet. P JB
Bostrichidae *Sinoxylon senegalenesis M JB
"Phonapate arabs Lesne H JB
Cleridae *Necrobia rufipes (D e Gerr) P JB
Dermestidae ”Anthrenus flavipes LeConte H JB
Meloidae indet. P JB
Ptillidae Indet. P JB
Crustacea Isopoda Porcelinidae Porcelinidae? P JB
Chilopoda Scolopendromorpha Scolopendrida Scolopendrida mirabilis P BJT
Vertebrata ReptiliaSquamata Agamidae Phrynocephalus arabicus Anderson P PEO
Gekkonidae Bunopus tuberculartus Bianford P PEO
Stenodactylus arabicus (H aas) P PEO
Stenodactylus dorlae Bianford P PEO
Stenodactylus leptocosymbotes Leviton P PEO
& AndersonStenodactylus slaveni Haas P PEO
Teratosclncus sclncus (Schlegei) P PEO
Lacertidae Mesallna adramitana Bianford P PEO
Scincidae Sclncus mitranus Anderson P PEO
Sclncus sclncus conlrostris (Bianford) P PEO
Serpentes Leptotyphlopidae Leptotyphlops macrorhyncus (Jan) P PEO
Appendix 4.1 Continued List of species caught in UAE (1992-1996)
* indicates new records for UAE and new species have been shaded. For a
complete checklist of the country use both Appendices 1.1 and 4.1.
P= pitfall trap, L= light trap, H= hand searching or netting, M= Malaise trap, F=
fragment recovered from houbara faeces, B= baiting (for ants).
See over-leaf for list of taxonomists involved in species identification and
verification.
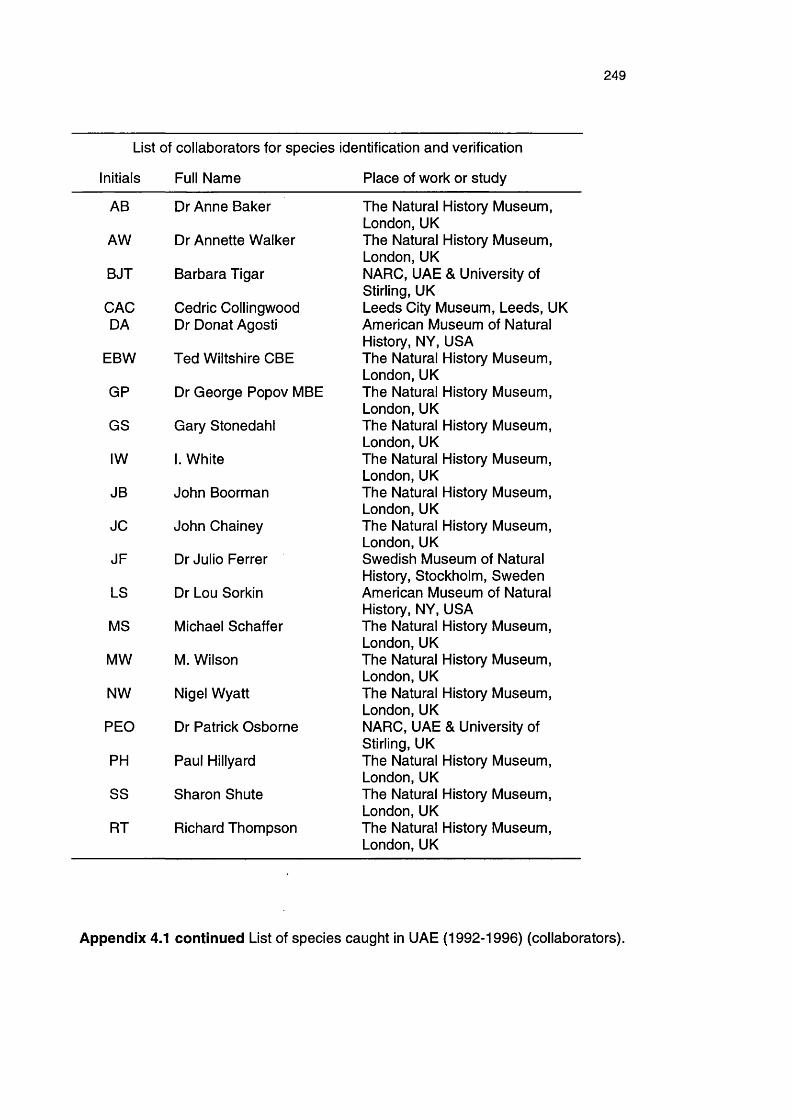
249
List of collaborators for species identification and verification
Initials Full Name Place of work or study
AB Dr Anne Baker The Natural History Museum, London, UK
AW Dr Annette Walker The Natural History Museum, London, UK
BUT Barbara Tigar NARC, UAE & University of Stirling, UK
CAC Cedric Collingwood Leeds City Museum, Leeds, UKDA Dr Donat Agosti American Museum of Natural
History, NY, USAEBW Ted Wiltshire QBE The Natural History Museum,
London, UKGP Dr George Popov MBE The Natural History Museum,
London, UKGS Gary Stonedahl The Natural History Museum,
London, UKIW 1. White The Natural History Museum,
London, UKJB John Boorman The Natural History Museum,
London, UKJC John Chainey The Natural History Museum,
London, UKJF Dr Julio Ferrer Swedish Museum of Natural
History, Stockholm, SwedenLS Dr Lou Sorkin American Museum of Natural
History, NY, USAMS Michael Schaffer The Natural History Museum,
London, UKMW M. Wilson The Natural History Museum,
London, UKNW Nigel Wyatt The Natural History Museum,
London, UKPEG Dr Patrick Osborne NARC, UAE & University of
Stirling, UKPH Paul Hillyard The Natural History Museum,
London, UK88 Sharon Shute The Natural History Museum,
London, UKRT Richard Thompson The Natural History Museum,
London, UK
Appendix 4.1 continued List of species caught in UAE (1992-1996) (collaborators).
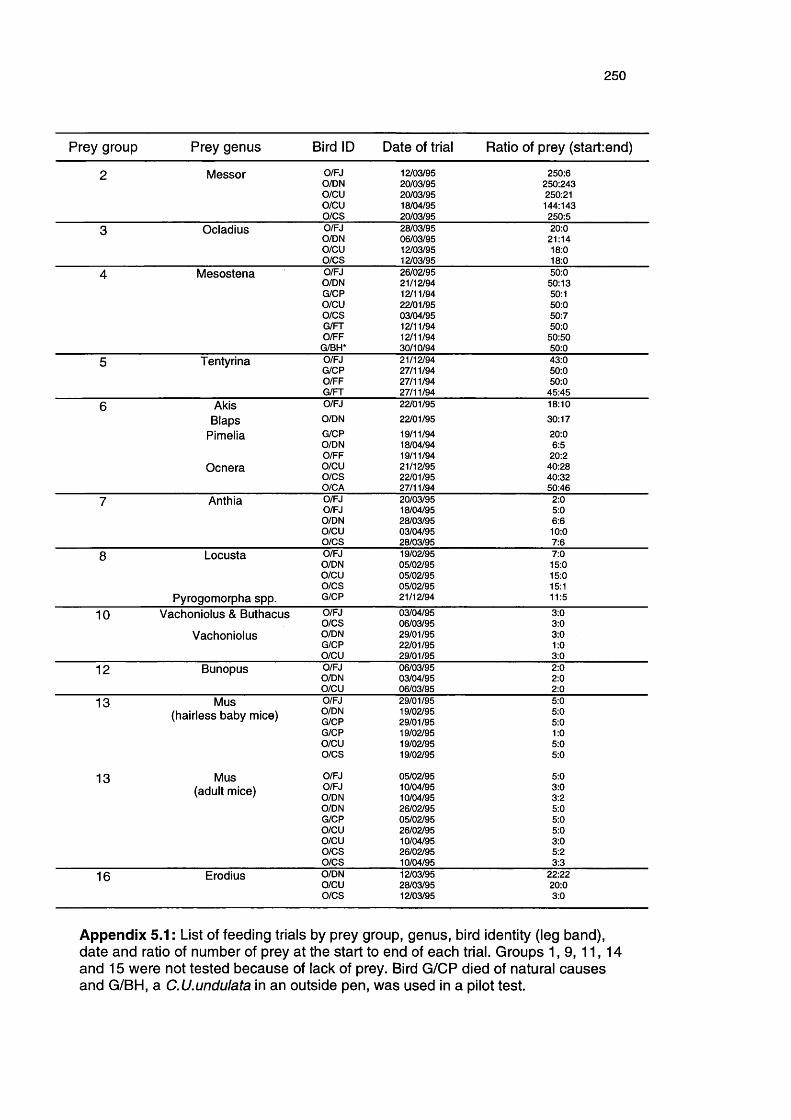
250
Prey group Prey genus Bird ID Date of trial Ratio of prey (start:end)
2 Messor O/FJ0 /D N0/C U0/C U0/C S
12/03/9520/03/9520/03/9518/04/9520/03/95
250:6250:243250:21144:143250:5
3 Ocladius O/FJ0 /D N0/C U0 /0 8
28/03/9506/03/9512/03/9512/03/95
20:021:1418:018:0
4 Mesostena O/FJ0 /D NG/CPO/CU0/C SG/FT0 /F F
G/BH*
26/02/9521/12/9412/11/9422/01/9503/04/9512/11/9412/11/9430/10/94
50:050:1350:150:050:750:0
50:5050:0
5 Tentyrina O/FJG/CP0 /F FG/FT
21/12/9427/11/9427/11/9427/11/94
43:050:050:0
45:45
6 Akis O/FJ 22/01/95 18:10
BlapsPimelia
Ocnera
0 /D N
G/CP0 /D N0 /F F0 /C U0/C S0 /C A
22/01/95
19/11/9418/04/9419/11/9421/12/9522/01/9527/11/94
30:17
20:06:5
20:240:2840:3250:46
7 Anthia O/FJO/FJ0 /D N0/C U0/C S
20/03/9518/04/9528/03/9503/04/9528/03/95
2:05:06:610:07:6
8 Locusta
Pyrogomorpha spp.
O/FJ0 /D N0/C U0/C SG/CP
19/02/9505/02/9505/02/9505/02/9521/12/94
7:015:015:015:111:5
10 Vachoniolus & Buthacus
Vachoniolus
O/FJ0 /C S0/D NG/CP0/C U
03/04/9506/03/9529/01/9522/01/9529/01/95
3:03:03:01:03:0
12 Bunopus O/FJ0/D N0/C U
06/03/9503/04/9506/03/95
2:02:02:0
13 Mus(hairless baby mice)
O/FJ0/D NG/CPG/CP0/C U0/C S
29/01/9519/02/9529/01/9519/02/9519/02/9519/02/95
5:05:05:01:05:05:0
13 Mus (adult mice)
O/FJO/FJ0/D N0/D NG/CP0/C U0/C U0/C SO/CS
05/02/9510/04/9510/04/9526/02/9505/02/9526/02/9510/04/9526/02/9510/04/95
5:03:03:25:05:05:03:05:23:3
16 Erodius 0 /D N0 /C U0/C S
12/03/9528/03/9512/03/95
22:2220:03:0
Appendix 5.1 : List of feeding trials by prey group, genus, bird identity (leg band), date and ratio of number of prey at the start to end of each trial. Groups 1,9, 11, 14 and 15 were not tested because of lack of prey. Bird G/CP died of natural causes and G/BH, a C.U.undulata in an outside pen, was used in a pilot test.

251
Appendix 5.2
Preliminary Study of the Rate of Passage of Digesta in Houbara bustard
by Tigar, B. J. (1995).
National Avian Research Center Internal Report Number 5 1995

252
Preliminary Study of the Rate of Passage of Digesta in Houbara bustard
Barbara Jane Tigar
Rate of passage through the gut of the houbara was investigated in two separate trials on four birds in Sweihan Quarantine Centre. Plastic pellets were used as a marker and were given to the birds in a mash of houbara feed pellets. Different coloured markers were given on three consecutive days and their consumption was highest on the first day and lowest on the third. Overall consumption of plastic pellets was low and highly variable both between experiments and individual birds. The first pellets to be recovered appeared within three hours and the majority recovered were obtained within 24 hours. One sample appeared after 37 days and may have been either a contaminant or may have been retained by the gizzard. Fewer plastic pellets were consumed in the second trail than the first. Reasons for the variation are discussed and ways to improve such work in the future are suggested. This includes a summary of a review of experiments on passage time which has recently been added to our library. It is highly relevant and contains much useful information on the limitations, design and statistical analysis of such trials.
Introduction
The Houbara Feeding Ecology Program is primarily concerned with the diet of wild houbara in Abu Dhabi. However, a series of experiments was carried out using captive birds to enable us to identify and quantify invertebrate remains in faeces. The tests were carried out in Sweihan Quarantine Centre (SQC) and upon completion the experimental birds were kept a further six weeks to investigate the rate of passage through the gut. This is defined by Warner (1981) as the time taken for an individual portion of digesta to be mechanically mixed, digested, absorbed and excreted.
Warner's (1981) review of the rate of passage in birds and mammals was not available at the start of the experiments but may have influenced the experimental design and choice of marker. He points out that three important factors influence all experiments on the rate of passage:
1. There is usually a large coefficient of variation and many replicates are needed to demonstrate small effects.
2. The amount of food consumed during the experiments affects the retention time and ad lib intakes are difficult to interpret.
3. Different components of the diet may have independent passage rates and markers must behave in a similar manner to the experimental diet.
Points two and three are relevant to our work because the marked food was left with the birds for 12 hours and plastic beads were used as the marker and it is unlikely that they behaved in a similar way to the houbara diet.
The speed of the passage through the gut has implications for several areas in which NARC has an interest, particularly the rate at which drugs are absorbed and the time taken for non- digestible fragments to appear in faeces. For example, a single meal may appear in numerous faeces which are voided at different times. Differential digestion of various items may also affect the rate of passage time. Data from the feeding trials with invertebrate prey will also increase our understanding of these factors. However, the aim of this study was to establish a standard determination of passage time which could be used as a reference for the interpretation of both faecal analysis and pharmokinetic studies in houbara.
Methods
Experimental Conditions
The four Chlamydotis undulata macqueenii used in the trails were fed a diet consisting solely of dry houbara maintenance pellets from November 1994 until June 1995. Each bird was given 75 g of pellets per day (i.e. excess food) and food intake was monitored and is
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard
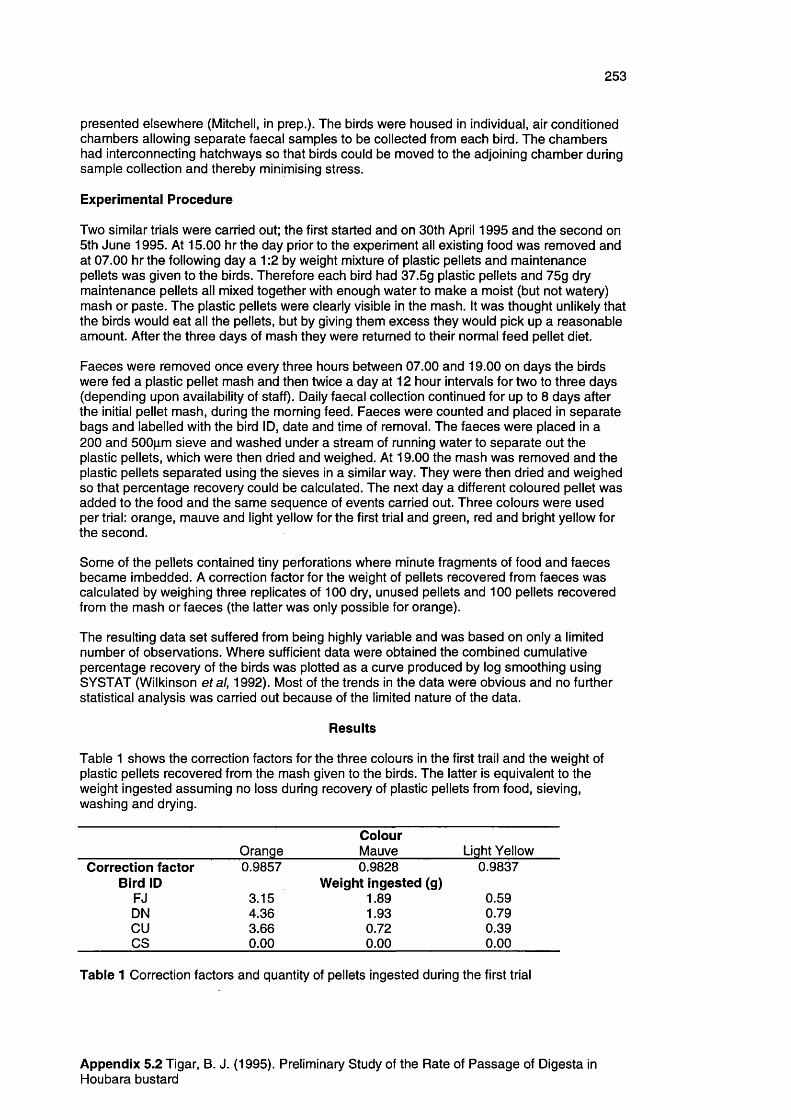
253
presented elsewhere (Mitchell, in prep.). The birds were housed in individual, air conditioned chambers allowing separate faecal samples to be collected from each bird. The chambers had interconnecting hatchways so that birds could be moved to the adjoining chamber during sample collection and thereby minimising stress.
Experimental Procedure
Two similar trials were carried out; the first started and on 30th April 1995 and the second on 5th June 1995. At 15.00 hr the day prior to the experiment all existing food was removed and at 07.00 hr the following day a 1:2 by weight mixture of plastic pellets and maintenance pellets was given to the birds. Therefore each bird had 37.5g plastic pellets and 75g dry maintenance pellets all mixed together with enough water to make a moist (but not watery) mash or paste. The plastic pellets were clearly visible in the mash. It was thought unlikely that the birds would eat all the pellets, but by giving them excess they would pick up a reasonable amount. After the three days of mash they were returned to their normal feed pellet diet.
Faeces were removed once every three hours between 07.00 and 19.00 on days the birds were fed a plastic pellet mash and then twice a day at 12 hour intervals for two to three days (depending upon availability of staff). Daily faecal collection continued for up to 8 days after the initial pellet mash, during the morning feed. Faeces were counted and placed in separate bags and labelled with the bird ID, date and time of removal. The faeces were placed in a 200 and 500pm sieve and washed under a stream of running water to separate out the plastic pellets, which were then dried and weighed. At 19.00 the mash was removed and the plastic pellets separated using the sieves in a similar way. They were then dried and weighed so that percentage recovery could be calculated. The next day a different coloured pellet was added to the food and the same sequence of events carried out. Three colours were used per trial: orange, mauve and light yellow for the first trial and green, red and bright yellow for the second.
Some of the pellets contained tiny perforations where minute fragments of food and faeces became imbedded. A correction factor for the weight of pellets recovered from faeces was calculated by weighing three replicates of 100 dry, unused pellets and 100 pellets recovered from the mash or faeces (the latter was only possible for orange).
The resulting data set suffered from being highly variable and was based on only a limited number of observations. Where sufficient data were obtained the combined cumulative percentage recovery of the birds was plotted as a curve produced by log smoothing using SYSTAT (Wilkinson etal, 1992). Most of the trends in the data were obvious and no further statistical analysis was carried out because of the limited nature of the data.
Results
Table 1 shows the correction factors for the three colours in the first trail and the weight of plastic pellets recovered from the mash given to the birds. The latter is equivalent to the weight ingested assuming no loss during recovery of plastic pellets from food, sieving, washing and drying.
OrangeColourMauve Light Yellow
Correction factor 0.9857 0.9828 0.9837Bird ID Weight ingested (g)
FJ 3.15 1.89 0.59DN 4.36 1.93 0.79CU 3.66 0.72 0.39OS 0.00 0.00 0.00
Table 1 Correction factors and quantity of pellets ingested during the first trial
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

254
Table 1 also shows that there was a decline in the amount of plastic pellets consumed with most taken on the first day (orange), fewer on the second (mauve) and virtually none on the third (light yellow). One of the birds, CS, did not eat any plastic pellets.
Cumulative percentage recovery of the three colours from faeces during the first trial is plotted against time lapsed from the start of the trial (Figures 1 to 3). Data points from individual birds are represented by the same symbols in all the figures i.e. a circle for FJ, a small diamond for DN, and a square for CU. The curve was produced by log smoothing of the combined data. The time axis indicates the number of days since the start of the experiment. Therefore mauve pellets do not appear until after day two and light yellow pellets after day three.
60
50
40
30MI 20 Ü
0 2 3 5 6 7 81
Time (days)
Figure 1 Plot of cumulative percentage recovery for orange pellets
60
50
%40
X 30
20O
80 2 3 5 6 71
Tim e (days)
Figure 2 Plot of cumulative percentage recovery for mauve pellets
X 30
0 1 2 3 4 5
Time (days)
Figure 3 Plot of cumulative percentage recovery for light yellow pellets
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

255
The highest cumulative percentage recovery of pellets from faeces was 58% of mauve from DN samples and occurred between three and four days after the start of the trial, i.e two to three days after ingestion. However, most of the recovered pellets were obtained within 24 hours of ingestion, seen as a rapid increase followed by a levelling out of the curves in Figures 1 to 3. A higher percentage of orange pellets was recovered from FJ and very few light yellow pellets were either ingested or recovered from any of the birds.
RedColourGreen Yellow
Correction factor 0.9457 0.9900 0.9855Bird ID Weight ingested (g)
FJ 0.30 0.12 0.03DN 0.01 0.03 0.03CU 0.16 0.02 0.06CS 0.04 0.03 0.00
Table 2 Correction factors and quantity of pellets ingested during the second trail
Table 2 shows the correction factors and amount of pellets ingested in the second trial. Fewer plastic pellets were ingested than in the first trail and apart from red and green for FJ and red for CU pellet intake was negligible (less than 0.1 g). Again there was a decrease in the amount of pellets taken, with most being consumed on day one, in this case red pellets.
The cumulative percentage recovery of red and green pellets is shown in Figure 4. The top curve represents red and is based on 5 data points from FJ and one from CU, the bottom curve is for green and is based on only two data points from FJ.
80
70
60
50
I30
Ü
0 2 3
Time (days)
Figure 4Plot of cumulative percentage recovery for red and green pellets
One unusual result was that a single mauve pellet was recovered from CU on 6th June 1995. Assuming that this was from the first trial and not a contamination then it took 37 days to be voided by the bird.
Discussion
Time for recovery of the first sample was fairly rapid, within three hours for FJ. This is similar for other birds species, for example Duke e ta /(1968) recovered the first marked sample from the ring-necked pheasant in 1.5 hours and the last in 8.5 hours. The latter authors also measured the through-put time of the caecae which was slower, with the first sample appearing at 6.6 hours and the last at 39 hours. However, the last sample we recovered appeared after 36 days. This may suggest that plastic (or other) beads are not suitable for this type of study because they act like grit and are retained to aid the crushing action of the gizzard. This retention may also apply to invertebrate and other hard remains in faecal samples. Analysis of the captive bird feeding calibration trails should help illuminate any such
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

256
affects. The only way to be sure that no further pellets remain would be to euthanase the birds and examine their gizzard contents which is not considered justifiable for this trial.
The reason for the small intake of plastic pellets by the birds is unclear. It may indicate that they were actively avoiding them or had a preference for particular colours e.g. orange. It is also possible that they did not find the mash mixture as palatable as their usual diet of dry pellets, although such a mash with various additional ingredients is given to most of NARC's birds. However, it can go rancid and quickly spoil, and mould growth was noted on the top of the mash when separating the plastic pellets in the lab the next day. Whatever the reason, the small quantity of pellets ingested meant that any inaccuracy in recovering and weighing the pellets had a large effect on the percentage of pellets recovered.
There was some variation in the consumption of plastic pellets by the different birds, with FJ and DN taking the most. CS took none at all and fewer were taken by the other birds in the second trial which may suggest avoidance (although since different colours were used it is hard to say). Intake of plastic pellets may be related to the normal food intake of the birds. Records of daily food consumption may show that some birds always eat more than others and so automatically ingested more plastic pellets (Mitchell, in prep). 0 8 and DN were females and when weighed on 22nd March 1995 registered 1090g and 1009g respectively.FJ and CU were males and therefore heavier than the females, weighing 1304g and 1647g respectively. This is a very small sample size but there was no obvious effect of weight or sex on the intake of plastic pellets. However, daily consumption of food pellets by captive houbara is highly variable (Jacquet, in prep; Mitchell, in prep) and it may just have been a natural low in food intake rather than an avoidance of the pellets by individuals.
It is possible that some plastic pellets were not recovered from either the food dish or faeces. The chamber floor was covered with a layer of sand, and plastic pellets could easily have been coated with sand especially when wet. If the birds had taken more of the plastic pellets such losses would have had less effect. Warner (1981) also concedes that it is often difficult to get 100% recovery of many types of markers due to unknown causes.
Problems during the trials
During sample collection the birds were moved to the adjoining room to avoid stress caused by the researcher entering in the room. However, all these birds were wild- caught adults and often showed signs of distress including defaecation on being moved. This was probably a defensive action and there is evidence that houbara defaecate onto attacking falcons to deter them (Cramp & Simmonds, 1980) and as a preparation for flight when alarmed (personal observation). In the wild, houbara appear to defaecate fairly frequently as indicated by the high rate of encounter of faeces along their tracks (pers. observation), so through-put time is probably always fairly rapid. However, too frequent a visit to the study birds may have stressed them and affected their behaviour increasing the speed of the gut and number of samples.
The pellets and food pellets were given as a mash but it was possible for the birds to avoid the colours. A more viscous mixture of foods would make it easier to conceal the plastic pellets and harder for the birds to avoid or remove them. Peanut butter is used to conceal the plastic pellets from badgers (C. Cheeseman, in lift.) and this might be suitable for houbara although it is not a natural food item and may not be palatable to the birds. A mixture of mince or even placing the pellets inside mice might work although any change in the diet might also affect the through-put time.
Gut length is known to be affected by diet for other birds and switching from a captive based diet of concentrated food such as pellets, to a natural based, higher fibre content diet has been shown to increase the gut length in red grouse (Moss & Parkinson, 1972). However, the quantity of fibre may also affect retention time, and Mattocks (1971) found that in geese, grass started to appear in faeces 1.7 hours after feeding while meal (lower fibre but higher nutritional content) took only 1.3 hours. Originally a trial using a mixed diet like that given to most captive birds at NARC had been planned, i.e. switching to a mixture of soaked houbara pellets, minced meat, cabbage and apple. This could be used to examine any differences in through-put time and the affect of changing the diet over say a four week period. However, it
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

257
was decided not to attempt this because of poor consumption and recovery of the plastic pellets from the maintenance pellet trail.
Future work.
This work was carried out after the trials with invertebrate prey because it was thought to be relatively simple to execute and resources were available at the time. Manpower is not currently available to develop these techniques and preliminary results and the review of Warner (1981 ; see summary below) show that such studies are more complicated than anticipated. If in the future such work becomes a higher priority the following comments on choice of marker and experimental design, based on Warner (1981), should be taken into account and a full literature search carried out.
Choice of marker
Markers can be divided into four categories, but no practical marker is ideal although all can give an approximate rate of passage. Choice of marker will depend on the species being studied and resources available (some can only be detected by sophisticated analytical techniques or x-rays). The list below explains the biases that affect recovery of different types of markers from faeces.
1. Normal dietary constituents such as lignin.The recovery of all dietary markers is difficult to measure accurately and they may also suffer some digestion.
2. Solute markers which probably stay in solution in the gut, for example polyethylene glycol (PEG) and the chromium complex of ethylenediamine tetra-acetic acid (EDTA).It is not known how solute markers are distributed between the free gut fluid and the fluid within the digesta, which affects the time of detection in faeces. Some adsorption into the bloodstream may also occur.
3. Particulate markers such as chromic oxide and radio-opaque plastic pellets. The plastic pellets we used come into this category.All particulate markers probably differ in size, specific gravity or surface properties from the food and so may behave differently from it. Size has been shown to effect the rate of passage of particulate markers in mammals with a trend for smaller sizes to pass more rapidly. Substances of a high specific gravity are retained for longer, particularly in the stomach and caecum. Plastic pellets of specific gravity 1.12 had a mean retention time of about 60 hours in cattle fed on hay, but this increased to 110 hours when fed on a concentrate diet (Campling & Freer, 1962).
4. Particle markers which are originally in solution but become embedded in food e.g. stains and the ruthenium-phenathroline complex.Particle markers have been shown to transfer between particles and may be adsorbed into the bloodstream.
Experimental design
The simplest experimental design is to use a pulse dose, i.e. the marker is given as single pulse (discrete meal) and then the faeces are collected at regular intervals. Passage rate may vary according to the frequency of feeding and the time between each feed. In geese, dietary markers travelled more slowly when given mid-meal than when given on an empty gut (Mattocks, 1971). The recovery pattern for the marker should give a sigmoid curve. Various calculations and indices are discussed by Warner (1981), particularly the time to excrete a certain percentage of the marker which is a common theme in many papers. For example, useful reference points are the time for first recovery of marker, which is often termed transit time or t , and the time for half the marker to be recovered also called half time or t o- Warner recommends calculating the mean recovery time, which is the average time of retention of all the digesta being studied and presents mathematical formulae for calculating the defined parameters. If the passage time for different segments of the gut is required, radio-opaque markers and x-rays must be used, or the experimental animals killed and dissected.
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

258
Warner (1981) highlights the complexity of this topic and factors which must either be controlled or accepted as biases. Sufficient replication is very important as there may be tremendous variation between individuals of the same species. Future studies of houbara should avoid the use of particulate markers. Actual choice of marker will depend on resources and equipment available, but a preliminary trial to investigate the best markers should be carried out. We originally tried using colour dyes in our trials (Mitchell, in prep.) but we could not detect their presence in the faeces probably because they were either digested or absorbed by the bloodstream. It maybe worthwhile looking at both solid faecal and caecal faecal through-put times. We did not differentiate between the them in this trail and it is sometimes hard to do so because they are present within a single sample (personal observations). Mean retention time as suggested by Warner (1981) would be a good reference value to aim for. We were unable to calculate it because of the poor recovery of markers.
Acknowledgements
The coloured plastic pellets were kindly provided by Dr Chris Chessman of Central Science Laboratory, UK and his work on badger territorial ranges acted as an inspiration for using them in these trials. William Mitchell is duly thanked for many hours of technical assistance, in particular for collecting and processing of samples. Dr Patrick Osborne is thanked for his advice on data handling and analysis. Judith Howlett, Mohammed Nafeez and Dr Jaime Samour of Veterinary Science Department are also thanked for their help at SQC.
References
Campling, B.C. & Freer, M (1962). The effect of specific gravity and size on the mean time of retention of inert particles in the alimentary canal of the cow. British Journal of Nutrition 16, 507-518.
Cramp, S. & Simmons, K.E.L. (1980). Handbook of the Birds of Europe, the Middle East and North Africa. Vol.2. Hawks to Bustards. Oxford University Press.
Duke, G.E., Petrides, G.A. & Ringer, R.K. (1968). Chromium-51 in food metabolizablity and passage rate studies with the ring-necked pheasant. Poultry Science 47,1356-1364.
Jacquet, J. M. (in prep). Food intake in the houbara bustard, relation with body weight and ambient temperature. NARC Internal Report Series
Mattocks, J.G. (1971). Goose feeding and cellulose digestion. Wildfowl 22,107-113.Mitchell, W. L. (in prep). Sandwich student placement report 1994-95. NARC Internal Report
SeriesMoss, R. & Parkinson, J.A. (1972). The digestion of heather {Cailuna vulgaris) by red grouse
{Lagopus lagopus scoticus). Br. J. Nutr., 27, 285-298 Warner, A.C.I. (1981). Rate of passage of digesta through the gut of mammals and birds.
Commonwealth Bureau of Nutrition, Nutrition abstracts and review series ”B", 51, 789-819.
Wilkinson, L., Hill, M, Howe, P & Miceli, S. (1992). SYSTAT for Windows, Version 5 Edition. Evanston, IL: SYSTAT, Inc., 1992.
Appendix 5.2 Tigar, B. J. (1995). Preliminary Study of the Rate of Passage of Digesta inHoubara bustard

Prey group Description Remains likely to be present in faecal samples
Tough, square-shaped head capsule is usually intact. Jaws typical, “hand-like” shape, occurring loose and hidden inside head capsule. Pieces of cuticle, propodeums and typical legs seen floating on the surface tension of samples.
1 & 2 Nocturnal and diurnal ants.HymenopteraiFormicidae
3 Phytophagous, plant dwelling insects e g weevils
4 Nocturnal, small tenebrionids e.g Mesostena puncticollis
5 Diurnal, small, plant climbing tenebrionids Coleoptera:T enebrionida e Tentyrina palmen
6 Large, noctunral tenbrionids
7. Fast-moving, predatory carabid beetles
8 Fast-moving, plant dwelling insects (can fly or jump)Orthoptera:Acridida
9 Very fast, large predatory arachnids. Solifugae
10 Slow, large predatory arachnids. Scorpionae
11 Diurnal Reptilia12 Nocturnal Reptilia13 Rodentia14 Fast moving, small
predatory arachnids Aranea
15 Diurnal/nocturnal aerial insects
16 Large diurnal tenebrionids
Weevils are highly sclerotised and usually covered in scales which are worn-off during digestion. When threatened they withdrawn their appendages close into the body, and a few almost complete bodies were recovered. They have characteristically shaped limbs and their femora are distinctive and easy to identify for each species. They are often attached to tibia. Head and rostrum, tarsi, disk, prosternum (often fused to disk), elytra (with evidence of scales), sternites (usually fused together) also recovered less frequently.A variety of fragments, including jaws, heads, tibiae and femora were recovered during the trials M.puncticollis has a row of tiny serrations at front of head capsule, parallel rows of dots along the elytra, and a very shiny, smooth thoracic disc. Other species identified on the shape of their femoraT.palmeri has no serration at front of head capsule but a small peg-like, off-centre projection is present. The elytra are plain and smooth. Two dimples are present on the thoracic disk. When large numbers are eaten some beetles remain almost complete following digestion.
Much finely-ground, black material present following digestion, e.g. fragmented elytra, thorax and head, but the jaws often remain intact and femora are recovered in large numbers, although they maybe broken. If broken count leg joint only. Sometimes tibia are a similar size to femora, but they are usually more flattened.Most remains are finely ground but the distinctive, large sickle-shaped mandibles and notched tibia are conspicuous.
Generally very well digested. However, the mandible are usually intact, although they may be broken into two halves. The femur joint is often present.
Very characteristic long, orange hairs can be seen in the sample. The highly sclerotized jaws or chelicerae may be present.
Body highly fragmented, but fragments fluoresce under UV light. Occasionally a chelicera may be recovered.
Possibly a few scales. No bones recovered from wild or captive birds Possibly a few scales. No bones recovered from wild or captive birdsFur seen as glossy film on surface tension of sample or compact and matted fur-balls. Bones and teeth not recovered in trials. Chelicera and hairs might remain. Silken cocoons also possible.
Femur and metallic cuticle of Buprestids very distinctive. Characteristic, spiny fore-limb and front of head indicate dung beetles.
Generally rotund beetles with distinctive, half-moon shaped thorax. Adesmia spp. fragments have a blue metallic sheen. All have species specific sculpturing on first femur.
Appendix 5.3 Description of invertebrate prey remains recovered from faecal samples, with prey groups according to Table 4.1 rocnCD

260
: î
9
Group 1 Head capsule of Camponotus xerxes Group 2 Head capsule of Messor sp
Group 3 Femora and tibiae of Ammocleonus sp Group 3 Lateral view of Ammocleonus sp headcapsule
«WKNI
_____________Group 4 Head of Mesostena puncticollis showing small, angular eyes
Group 4 Close-up of M. puncticollis head showing small eyes and serrated front edge
Group 4 Elytrum of M. puncticollis showing parallel rows of dots.
Group 4 Fore-, mid- and hind-femora of M. puncticollis
Appendix 5.4 Photographs of typical fragments from the invertebrate prey groups.

261
Group 5 Head of Tentyrina palmeri showing large round eyes
Group 5 Close-up of T. palmeri head with large eyes and projection at front of head
Group 5 Fore-, mid- and hind-femora of T. palmeri
Group 5 Elytrum of T. palmeri
ft I t 3 4 ,: :5 S ? ' $ « |
Group 6 Wax-covered femora of Ocnera philistina
Group 6 Fore- and mid-femora of Blaps koliari
0M______________
'
'*
Group 6 Fore-, mid- and hind-femora of Akis elevator
Group 6 Fore-limb of Paraplatyope popovi showing distinctive tibia
Appendix 5.4 Photographs of typical fragments from the invertebrate prey groups.

262
.. .
Group 7 Sickle shaped-jaws of Scarities guineensis
Group 7 Sickle-shaped jaws of Anthia duodecimguttata
_______ _Group 9 Chelicerate mouthparts of camel spider, Galeodes sp.
Group 15 Fore-tibia of Sacer christatus
Group 15 Head capsule of Sacer christatus, showing spines at front edge
Group 15 Buprestid femora, showing pitted surface
4.%% y ^Group 16 Erodius sp, fore-limb, with fringes of Group 16 yApenfanoc/es sp limbs, note the largehair on femur and spine on tibia fore-limb
Appendix 5.4 Photographs of typical fragments from the invertebrate prey groups.

263
Group 16 Hind-limb of Adesmia sp. Group 16 Fore-, mid- and hind-limbs of Adesmia sp.
Appendix 5.4 Photographs of typical fragments from the invertebrate prey groups.
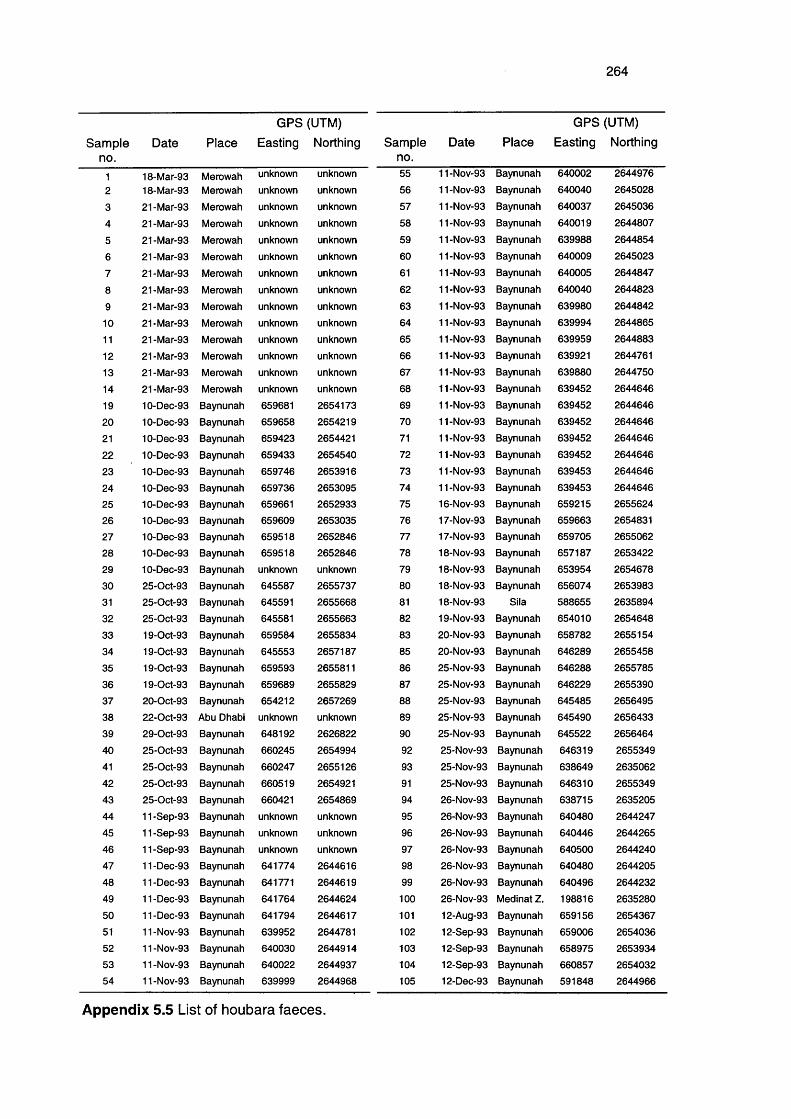
264
Sampleno.
Date PlaceGPS (UTM)
Easting Northing Sampleno.
Date PlaceGPS (UTM)
Easting Northing
1 18-Mar-93 Merowah unknown unknown 55 11 -Nov-93 Baynunah 640002 2644976
2 18-Mar-93 Merowah unknown unknown 56 11 -Nov-93 Baynunah 640040 2645028
3 21-Mar-93 Merowah unknown unknown 57 11 -Nov-93 Baynunah 640037 2645036
4 21-Mar-93 Merowah unknown unknown 58 11-Nov-93 Baynunah 640019 2644807
5 21-Mar-93 Merowah unknown unknown 59 11-Nov-93 Baynunah 639988 2644854
6 21-Mar-93 Merowah unknown unknown 60 11-Nov-93 Baynunah 640009 2645023
7 21-Mar-93 Merowah unknown unknown 61 11-Nov-93 Baynunah 640005 2644847
8 21 -Mar-93 Merowah unknown unknown 62 11-Nov-93 Baynunah 640040 2644823
9 21-Mar-93 Merowah unknown unknown 63 11-Nov-93 Baynunah 639980 2644842
10 21-Mar-93 Merowah unknown unknown 64 11 -Nov-93 Baynunah 639994 2644865
11 21-Mar-93 Merowah unknown unknown 65 11 -Nov-93 Baynunah 639959 2644883
12 21-Mar-93 Merowah unknown unknown 66 11 -Nov-93 Baynunah 639921 2644761
13 21-Mar-93 Merowah unknown unknown 67 11-Nov-93 Baynunah 639880 2644750
14 21-Mar-93 Merowah unknown unknown 68 11-Nov-93 Baynunah 639452 2644646
19 10-Dec-93 Baynunah 659681 2654173 69 11-Nov-93 Baynunah 639452 2644646
20 IO-Dec-93 Baynunah 659658 2654219 70 11-Nov-93 Baynunah 639452 2644646
21 10-Dec-93 Baynunah 659423 2654421 71 11-Nov-93 Baynunah 639452 2644646
22 IO-Dec-93 Baynunah 659433 2654540 72 11-Nov-93 Baynunah 639452 2644646
23 IO-Dec-93 Baynunah 659746 2653916 73 11 -Nov-93 Baynunah 639453 2644646
24 IO-Dec-93 Baynunah 659736 2653095 74 11 -Nov-93 Baynunah 639453 2644646
25 IO-Dec-93 Baynunah 659661 2652933 75 16-NOV-93 Baynunah 659215 2655624
26 IO-Dec-93 Baynunah 659609 2653035 76 17-NOV-93 Baynunah 659663 2654831
27 10-Dec-93 Baynunah 659518 2652846 77 17-NOV-93 Baynunah 659705 2655062
28 IO-Dec-93 Baynunah 659518 2652846 78 18-NOV-93 Baynunah 657187 2653422
29 IO-Dec-93 Baynunah unknown unknown 79 18-NOV-93 Baynunah 653954 2654678
30 25-Oct-93 Baynunah 645587 2655737 80 18-NOV-93 Baynunah 656074 2653983
31 25-Oct-93 Baynunah 645591 2655668 81 18-NOV-93 Sila 588655 2635894
32 25-Oct-93 Baynunah 645581 2655663 82 19-NOV-93 Baynunah 654010 2654648
33 19-Oct-93 Baynunah 659584 2655834 83 20-NOV-93 Baynunah 658782 2655154
34 19-Oct-93 Baynunah 645553 2657187 85 20-NOV-93 Baynunah 646289 2655458
35 19-Oct-93 Baynunah 659593 2655811 86 25-NOV-93 Baynunah 646288 2655785
36 19-Oct-93 Baynunah 659689 2655829 87 25-NOV-93 Baynunah 646229 2655390
37 20-Oct-93 Baynunah 654212 2657269 88 25-NOV-93 Baynunah 645485 2656495
38 22-Oct-93 Abu Dhabi unknown unknown 89 25-NOV-93 Baynunah 645490 2656433
39 29-Oct-93 Baynunah 648192 2626822 90 25-NOV-93 Baynunah 645522 2656464
40 25-Oct-93 Baynunah 660245 2654994 92 25-NOV-93 Baynunah 646319 2655349
41 25-Oct-93 Baynunah 660247 2655126 93 25-NOV-93 Baynunah 638649 2635062
42 25-Oct-93 Baynunah 660519 2654921 91 25-NOV-93 Baynunah 646310 2655349
43 25-Oct-93 Baynunah 660421 2654869 94 26-NOV-93 Baynunah 638715 2635205
44 11-Sep-93 Baynunah unknown unknown 95 26-NOV-93 Baynunah 640480 2644247
45 11-Sep-93 Baynunah unknown unknown 96 26-NOV-93 Baynunah 640446 2644265
46 11-Sep-93 Baynunah unknown unknown 97 26-NOV-93 Baynunah 640500 2644240
47 11-Dec-93 Baynunah 641774 2644616 98 26-NOV-93 Baynunah 640480 2644205
48 11-Dec-93 Baynunah 641771 2644619 99 26-NOV-93 Baynunah 640496 2644232
49 11-Dec-93 Baynunah 641764 2644624 100 26-NOV-93 Medinat Z. 198816 2635280
50 11-Dec-93 Baynunah 641794 2644617 101 12-Aug-93 Baynunah 659156 2654367
51 11-Nov-93 Baynunah 639952 2644781 102 12-Sep-93 Baynunah 659006 2654036
52 11 -Nov-93 Baynunah 640030 2644914 103 12-Sep-93 Baynunah 658975 265393453 11 -Nov-93 Baynunah 640022 2644937 104 12-Sep-93 Baynunah 660857 265403254 11-Nov-93 Baynunah 639999 2644968 105 12-Dec-93 Baynunah 591848 2644966
Appendix 5.5 List of houbara faeces.

265
Sampleno.
Date PlaceGPS (UTM)
Easting Northing Sampleno.
Date PlaceGPS (UTM)
Easting Northing
106 13-Dec-93 Baynunah 661349 2654402 137 02-Sep-94 Baynunah 642270 2551500
107 14-Dec-93 Baynunah 661411 2653969 138 02-Sep-94 Baynunah 642082 2651622
108 19-Dec-93 Baynunah 660597 2635290 139 02-Sep-94 Baynunah 641723 2651810
109 20-Dec-93 Baynunah 640596 2635274 140 02-Sep-94 Baynunah 641525 2651690
110 20-Dec-93 Baynunah 640512 2635901 141 30-Oct-94 Baynunah 642732 2602759
111 20-Dec-93 Baynunah 640592 2635747 142 30-Oct-94 Baynunah 642633 2602715
112 21-Dec-93 Baynunah 640851 2635937 143 30-Oct-94 Baynunah 642713 2602681
113 21-Dec-93 Baynunah 640959 2636000 144 30-Oct-94 Baynunah 642648 2642722
114 21-Dec-93 Baynunah 641147 2635846 145 30-Oct-94 Baynunah 642710 2602668
115 21-Dec-93 Baynunah 640650 2635360 146 30-Oct-94 Baynunah 642868 2602809
116 21-Dec-93 Baynunah 660654 2657109 147 25-Oct-94 Baynunah 629685 2612246
117 22-Dec-93 Oishtan 641879 2649302 148 27-Oct-94 Baynunah 678701 2650850
116 01-Oct-94 Baynunah 660802 2653781 149 19-May-94 South 248529 2568646
119 01-Mar-94 Baynunah 660848 2653003 150 20-May-94 South 248607 2568347
120 01-Mar-94 Baynunah 660786 2654802 151 20-May-94 South 248607 2568347
121 01-Mar-94 Baynunah 660824 2653040 152 20-May-94 South 248529 2568347
122 01-Mar-94 Baynunah 660790 2653795 153 20-May-94 South 248607 2568347
123 01-Apr-94 Baynunah 662393 2654150 154 20-May-94 South 248607 2568347124 01-Apr-94 Baynunah 662292 2654088 155 19-May-94 South 248529 2568646125 01-Apr-94 Baynunah 657590 2653021 156 19-May-94 South 248529 2568646126 01-Apr-94 Baynunah 660953 2654149 157 20-May-94 South 248607 2568347127 23-Jan-94 Baynunah 660890 2654053 158 20-May-94 South 248607 2568347128 23-Jan-94 Baynunah 660953 2654149 159 19-May-94 Baynunah 248529 2568646129 23-Jan-94 Baynunah 660752 2654172 160 20-May-94 Baynunah 248529 2568646130 23-Jan-94 Baynunah 660698 2654152 161 20-May-94 Baynunah 248529 2568646131 15-Feb-94 Baynunah 638882 2647694 162 19-May-94 Baynunah 248529 2568646132 02-Apr-94 Baynunah 660922 2654159 163 03-Sep-95 Baynunah 24.009 52.738133 02-Apr-94 Baynunah 660881 2654040 164 03-Sep-95 Baynunah 24.009 52.738134 02-Sep-94 Baynunah 642258 2651510 165 22-Apr-94 unknown unknown unknown135 02-Sep-94 Baynunah 642267 2651507 166 11-Apr-95 Baynunah unknown unknown136 02-Sep-94 Baynunah 642270 2551500 167 20-May-94 Baynunah 248607 2568347
Appendix 5.5 continued List of houbara faeces collected In UAE GPS= Global Positioning System and UTM= Universal Transverse Mercator Captive birds (15-18) and caecal faeces (128 & 165) were excluded from calculations In Chapter 5




















