
Identification_of_downstream_t.pdf - UCL Discovery
242
identification of downstream targets of Mirror, a transcription factor in Drosophila meianogaster. Mike Mohns Developmental Patterning Laboratory Cancer Research UK, London A thesis submitted for the degree of Doctor of Philosophy at the University of London August 2003
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Identification_of_downstream_t.pdf - UCL Discovery

identification of downstream targets of Mirror, a
transcription factor in Drosophila meianogaster.
Mike Mohns
Developmental Patterning Laboratory
Cancer Research UK, London
A thesis submitted for the degree of
Doctor of Philosophy
at the University of London
August 2003

ProQuest Number: U642545
All rights reserved
INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest U642545
Published by ProQuest LLC(2015). Copyright of the Dissertation is held by the Author.
All rights reserved.This work is protected against unauthorized copying under Title 17, United States Code.
Microform Edition © ProQuest LLC.
ProQuest LLC 789 East Eisenhower Parkway
P.O. Box 1346 Ann Arbor, Ml 48106-1346

Abstract
M irror, a member of the Iroquois family in Drosophila, is involved in
embryonic development and in planar polarity in the eye. m irror encodes a
homeodomain containing protein, which is believed to act as a transcription factor.
Mirror is expressed in the dorsal half of the Drosophila eye, where it plays a key
role in forming the equator. This dorsal-ventral midline is established by an interplay of
different signalling molecules and complex pathways. Mirror restricts expression of the
protein Fringe to the ventral half of the eye and mediates Notch activation at the fringe-
expressing boundary, which leads to formation of the equator.
In the developing embryo Mirror is expressed most strikingly in stripes at the
anterior side of the segmental border and in the central nervous system (CNS), however
its function there is unknown. Furthermore it is not known if Mirror acts as an activator
or repressor in the embryo. The absence of Mirror leads to segmentation and dorsal
closure defects in the embryo and results in embryonic lethality. The formation of the
CNS is also disrupted.
The aim of this work was to identify genes that are controlled by Mirror during
embryonic development. To detect downstream targets of Mirror I have over-expressed
Mirror in fly embryos and compared their modified gene expression with control
embryos. Mirror loss-of-function embryos have also been analysed. Changes in gene
expression were assessed using Drosophila Genome Arrays. To evaluate the microarray
data I analysed the expression of potential target genes in Mirror over-expressing and
mirror loss-of-function embryos with in situ hybridization and antibody staining
techniques. The analysis of potential Mirror targets is presented in my thesis.

Acknowledgements
I would like to thank my supervisor Dr. Helen McNeill for all the support, never ending
ideas, encouragement and advice she has given me during this project.
I would like to thank all past and present members of the Developmental Patterning
Laboratory for being such great friends.
I would like to thank the members of the Ish-Horowicz and Lewis Lab for their help and
advice.
I owe a lot of thanks to Simon Tomlinson and Gavin Kelly for their help and
enthusiasm to analyse all my microarray data.
I would like to thank Sophie for her patience with me and for the interest she developed
in the last four years for Mirror, microarrays and fruitflies to make our life easier. I
wouldn't have started doing all this without her.

to Kirsten

TABLE OF CONTENTSTitle page page 1Abstract page 2Acknowlegements page 3To Kirsten page 4Table of contents page 5List of figures page 9List of tables page 12
CHAPTER 1 : Introduction
1.1 Homeodomain transcription factors page 151.1.1. Homeotic genes and the homeodomain page 151.1.2. Classification of homeodomain proteins page 161.1.3. Iroquois belong to the TALE class of homeodomains page 17
1.2. The Iroquois family page 181.2.1. The Iroquois Complex page 181.2.2. Genomic organisation page 22
1.3. The role of the Iro/Irx genes in vertebrates page 231.3.1. Xenopus laevis page 251.3.2. Zebrafish page 271.3.3. Chick page 291.3.4. Mouse page 301.3.5. Human page 331.3.6. Irx genes share similar function in different organisms page 33
1.4. The Iroquois complex in Drosophila page 341.4.1. The role of Mirror and the Iro-C in the eye page 341.4.2. The role of Mirror and the Iro-C in the wing and the notum page 411.4.3. The role of Mirror during oogenesis page 471.4.4. Mirror in the embryo page 501.4.5. Formation of the CNS in Drosophila and mirror expression page 54
1.5. Aim of the project: page 581.5.1. The working hypothesis and the techniques page 581.5.2. The model page 581.5.3. Does Mirror act as an activator or a repressor? page 591.5.4. What are possible downstream targets of Mirror? page 60
CHAPTER 2 : Materials and Methods
2.1. Drosophila meianogaster techniques page 632.1.1. Flies page 632.1.2. Collection and fixation of embryos page 642.1.3. Embryo collections used in microarray experiments page 64

2.2. Molecular biology techniques page 652.2.1. RNA extraction and purification from Drosophila embryos page 652.2.2. Double stranded cDNA synthesis page 652.2.3. Drosophila Genome Arrays page 662.2.4. Polymerase chain reaction (PCR) page 702.2.5. Cloning page 742.2.6. DNA preparation, purification and analysis page 752.2.7. Sequencing page 75
2.3. in situ hybridisation page 762.3.1. Generation of anti-sense RNA probes page 762.3.2. Pre-hybridisation and hybridisation page 762.3.3. Signal detection for single in situ hybridisation page 772.3.4. Double in situ hbridisation with FAST RED and NBT/BCIP page 77
2.4. Bio- and Immunohistochemistry techniques page 782.4.1. Staining of Embryos page 782.4.2. Protein Extraction page 802.4.3. SDS PAGE page 802.4.4. Western Blots page 81
CHAPTER 3 : Mirror
3.1. Mirror expression in the embryo page 843.1.1. Mirror expression from stage 10-14 page 843.1.2. Mirror expression in various other stages of development page 86
3.2. Analysis of Mirror over-expressing embryos page 903.2.1. P-tubulin-WŸ 16xlJAS-mirror embryos page 903.2.2. Cuticle phenotype of mirror over-expressing embryos and its page 90 molecular origin3.2.3. Detection of Mirror protein by western blotting page 92
Discussion page 96
CHAPTER 4 : heat shock m/rror versus heat shock “control”
4.1. The heat shock Mirror over-expression model page 994.1.1. Direct comparison of expression profiles from heat shocked page 99 pKB12-mz>ror m dyw embryos4.1.2. CGI2199 seemed to be repressed by Mirror page 1014.1.3 Strain variation in gene expression page 1034.1.4. “pKB/yw-mi>ror” a new genetic background page 105
Discussion page 108

CHAPTER 5 : heat shock versus non heat shock
5.1. The heat shock subtraction model page 1125.1.1. Indirect comparison of expression profiles from mirror over- page 112expressing and yw embryos5.1.2. Over-expression of Mirror downregulates atonal expression in page 116the embryo5.1.3. Over-expression of Mirrro represses gem expression page 118
5.2. Over-expression of Mirror inhibits the expression of segment page 123polarity genes
5.2.1. gooseberry page 1235.2.2. hedgehog page 1255.2.3. wingless and sloppy paired page 125
Discussion page 128
CHAPTER 6 : Short heat shock
6.1. 30 minutes heat shock page 1356.1.1. The modified heat shock subtraction model page 13 56.1.2. “Slow and fast” heat shock response genes page 135
6.2. The “fast Mirror response” genes page 1406.2.1. alpha tubulin at 84B is positively regulated by Mirror page 1406.2.2. dalmatian, CG8066 and CG8013 are upregulated by Mirror page 1466.2.3. Fringe, CG2467 and CGI0650 are downregulated by Mirror page 150
6.3. The “slow Mirror response” genes page 1526.3.1. carnation, plutonium and CG4586 are positively regulated by page 152Mirror6.3.2. CGI5922, CG6900 and gleeful are negatively regulated by page 154Mirror
Discussion page 156
CHAPTER 7 : Mirror confers neuroblast identity
7.1. Mirror regulates Kriippel expression in the Drosophila embryo page 1627.1.1. Introduction to Kriippel page 1627.1.2. Kriippel expression in the developing CNS page 1647.1.3. Krüppel and Mirror are expressed in mutually exclusive domains page 1667.1.4. Over-expression of Mirror represses Kriippel in the embryo page 1677.1.5. Kriippel is ectopically expressed in Mirror loss-of-function page 172embryos

7.2. The CNS phenotype in mirror mutant embryos page 1797.2.1. Krüppel controls cell fate decisions in the CNS page 1797.2.2. Loss of Mirror leads to aberrant Engrailed expression in the CNS page 1807.2.3. Eagle expression is lost in Iroquois mutant embryos page 1827.2.4. Loss of Mirror function leads to defects in the CNS page 184
Discussion page 188
Final remarks
Bibliography P g
Appendix page 212

LIST OF FIGURES
CHAPTER 1
Figure 1.1. Schematic diagram of proteins of the Iroquois family. page 19Figure 1.2. The Iroquois phenotype. page 21Figure 1.3. Similar genomic organization of the Iro/Irx genes in page 24
Drosophila and vertebrates.Figure 1.4. Mirror is involved in forming the equator. page 36Figure 1.5. Development of the eye-antennal imaginai disc. page 37Figure 1.6. Mirror expression pattern in the third instar eye disc. page 37 Figure 1.7. A model for equator specification in the developing eye. page 39Figure 1.8. Effects of mirror loss-of-function in the adult eye. page 39Figure 1.9. Effects of Iro-C loss-of-function in the eye imaginai page 42
disc.Figure 1.10. Overview of gene expression in the dorsal (D) and page 42
ventral (V) compartments of the wing and the eye disc.Figure 1.11. Cartoon of a mature wing disc with indication of the page 44
fates of the relevant regions.Figure 1.12. Lack of alula formation in mirror mutant flies. page 44Figure 1.13. The Iro-C confer lateral identity to sensory neurons in page 46
the notum of the fly.Figure 1.14. Loss of Mirror results in ventralization of the egg page 49
chamber.Figure 1.15. The amnioserosa at germ band extension in a mirror page 51
mutant embryo.Figure 1.16. in situ hybridisation pattern of mirror in the embryo. page 53Figure 1.17. Mirror is expressed in delaminating neuroblasts. page 56
CHAPTER 3
Figure 3.1. Mirror antibody staining in wild type embryos from page 85stage 10-14.
Figure 3.2. in situ hybridisation pattern of araucan in the embryo page 87Figure 3.3. Further characterisation of Mirror expression in the page 89
embryo.Figure 3.4. Ectopic Mirror expression in page 91
Gal4xUAS-wz>ror embryos.Figure 3.5. Western blot to detect Mirror protein in P-tubulin VP 16- page 91
Gal-4xUAS-m/rror embryos.Figure 3.6 Cuticle phenotype in Iroquois over-expressing embryos page 93Figure 3.7. Molecular origin of the “wingless class” cuticle page 93
phenotype in ubiquitously mirror over-expressing embryos.
Figure 3.8. Western blot to detect Mirror protein in heat shocked page 94Mirror over-expressing embryos.

CHAPTER 4
Figure 4.1. The Mirror over-expressing model. page 100Figure 4.2. CGI2199, repressed by Mirror? page 102Figure 4.3. in situ hybridisation of CGI2199 in wild type embryo page 102Figure 4.4. Scatter Graphs visualises the differences in the genetic page 104
background.Figure 4.5. CG727PP a false candidate. page 106
CHAPTER 5
Figure 5.1. Heat shock subtraction model. page 113Figure 5.2. Over-expression of Mirror reduces the level of atonal page 117
transcript in the embryo.Figure 5.3. atonal in situ hybridisation in stage 11 embryos. page 117Figure 5.4. Histogram of gem expression. page 120Figure 5.5. Ectopic Mirror expression represses glial cells missing. page 120Figure 5.6. gem in situ hybridisation and Mirror antibody staining. page 122Figure 5.7. Mirror loss-of-function embryos lack Repo expressing page 122
cells in the CNS.Figure 5.8. Histogram of gooseberry expression. page 124Figure 5.9. Over-expression of Mirror repressed gooseberry (gsb) page 124
expression.Figure 5.10. Ectopic expression of Mirror represses page 126
expression in the embryo.Figure 5.11. Mirror antibody staining combined with hh in situ page 126
hybridisation.Figure 5.12 Ectopic expression of m/rror represses page 128
expression in the embryo.Figure 5.13. Ectopic Mirror or ectopic Araucan can repress sloppy page 128
pairedl (slpl)
CHAPTER 6
Figure 6.1. Mirror protein is detected after a short heat shock of 30 page 136minutes.
Figure 6.2. The fast heat shock response genes. page 138Figure 6.3. The slow heat shock response genes. page 139Figure 6.4. alphaTub 84B, a positive regulated fast Mirror response page 143
gene.Figure 6.5. Anti-alpha tubulin staining in germ band extended page 143
embryos.Figure 6.6. Anti-alpha Tubulin staining is altered in mirror loss-of- page 145
function embryos.Figure 6.7. The upregulated fast Mirror response genes. page 147Figure 6.8. /ro^wow mutant embryos exhibit a phenotype, page 149Figure 6.9. Genes with a fast response to Mirror over-expression. page 151Figure 6.10. The upregulated slow Mirror response genes. page 153
10

Figure 6.11. Downregulated slow Mirror response genes. page 155
CHAPTER 7
Figure 7.1. Krüppel expression pattern in the forming CNS. page 165Figure 7.2. Krüppel and Mirror are expressed in mutually exclusive page 168
domains.Figure 7.3. Over-expression of Mirror reduces the level of Kriippel page 169
transcripts in the embryo.Figure 7.4. Over-expression of Mirror downregulates Krüppel page 169
expression in the embryo.Figure 7.5. Over-expression of Mirror alters Krüppel expression and page 171
leads to morphological defects in the CNS.Figure 7.6. Kriippel is ectopically expressed in mirror loss-of- page 173
function embryos.Figure 7.7. Characterisation of ectopic Krüppel expression in page 175
Iroquois loss-of-function embryos.Figure 7.8. Anti-Krüppel and anti-Bgal staining in a mirror-lacZ page 175
mutant embryo.Figure 7.9. Double in situ hybridisation for mirror an Krüppel. Page 177Figure 7.10. Loss of Mirror results in aberrant Krüppel expression in page 178
the forming CNS at stage 14.Figure 7.11. Aberrant Engrailed expression in wzrror loss-of-function page 181
embryos.Figure 7.12. Eagle expression is lost in Mirror loss-of-function page 183
embryos.Figure 7.13. CNS phenotype of Iroquois mutants. page 186
11

LIST OF TABLES
CHAPTER 2
Table 1. Fly list page 63Table 2. Primer list page 70Table 3. Sequencing Primer page 75Table 4. Primary Antibody list page 79Table 5. Secondary Antibody list page 80Table 6. Primary and secondary antibodies used for detection of Western page 82
blots
CHAPTER 4
Table 7. Tested genes from the direct comparison approach
CHAPTER 5
page 103
Table 8. Tested genes from the direct comparison approach page 114
CHAPTER 7
Table 9: Penetrance of the two cell phenotype in mirror LOT embryos at page 173 stage 11
12

APPENDIX
Analysis of variance between replicates page 212Figure A; Long heat shock scatter plots page 213Figure B: Non heat shock scatter plots page 214Figure C: Clustering Based Correlation Matrix page 226
Table I: Initial sorting of upregulated and downregulated potential Mirror page 215target genes using fold change from the yw long hs versus pKB long hs comparison.
Table II: Initial sorting of consistently up- and dowregulated potential page 216 Mirror target genes of the combined pKB and pKBy w data
ANOVA: Analysis Of Variance between groups page 217Table III: Differentially expressed genes of yw long hs versus pKB long hs page 218
comparisonTable IV: Differentially expressed genes of yw long hs versus yw non hs page 219
comparisonTable V: Differentially expressed genes of pKB long hs versus pKB non hs page 220
comparisonTable VI: Differentially expressed genes of pKByw long hs versus pKByw page 221
non hs comparisonTable VII: Differentially expressed genes of the combined pKB and page 222
pKByw-mirror data.Table VIII: Differentially expressed genes of yw short hs versus yw non hs page 223
comparisonTable IX: Differentially expressed genes of pKB short hs versus pKB non page 224
hs comparisonTable X: Number of significant genes in each of the Anova categories page 225Table XI: Correlation Matrix page 226Table XII: Raw data of 800 randomly selected genes. page 227
13

14

Chapter 1: Introduction
CHAPTER 1
Introduction
1.1. Homeodomain transcription factors
1.1.1. Homeotic genes and the homeodomain
Homeotic genes or Hex genes are master control genes that specify the body and
regulate development of higher organisms. The Homeotic genes, which specify
segmental identity and positional information along the antero-posterior axis, are
organised into clusters on the chromosome.
Hex genes share a common sequence element of 180 bp, the homeobox, which
was first discovered in Drosophila as a region of homology common to several
homeotic genes and to the segmentation gene fushi tarazu (McGinnis et al., 1984; Scott
and Weiner, 1984). Subsequently it was shown to occur in all metazoa that have been
analysed, from sponges to humans, plants and fungi (Levine and Hoey, 1988).
Importantly, homeobox containing genes have also been found outside the Hox gene
clusters. The homeobox encodes a 60-amino acid homeodomain, which represents the
DNA-binding helix-tum-helix motif of the much larger homeodomain protein. The
homeodomain allows the sequence-specific recognition of sets of target genes by these
proteins. The recognition sequence generally contains a common ATT A core (Gehring
et a l, 1994). A glutamine at position 50 of the homeodomain makes direct contacts with
the DNA sequence just 5’ of the ATT A motif (Carroll, 1990). The structure of several
homeobox genes has been analysed by NMR spectroscopy and X-ray crystallography
15

Chapter 1: Introduction
and the prototype of a homeodomain has been determined. It consists of three alpha-
helices, which pack around a hydrophobic core. The homeobox binds specific DNA
sequences, which indicates that these proteins would regulate other genes on the level of
transcription. In other words they act as transcription factors (Desplan et al., 1985;
Gehring et al., 1994; Levine and Hoey, 1988).
1.1.2. Classification of homeodomain proteins
The classical homeodomain sequences have been classified into two
superclasses, the Complex Superclass and the Dispersed Superclass (Gehring et al.,
1994). This classification is based mainly on sequence similarity and on the fact that the
original homeodomain proteins, the Hox family, exist in chromosomal clusters or
complexes. The well conserved Drosophila and vertebrate Hox genes therefore form the
Complex Superclass which is divided into six classes named after the corresponding
Drosophila genes such as labial, deformed or sex combs reduced. Almost all homeobox
genes within the Complex Superclass share a hexapeptide motif of the consensus
sequence lYPWMK located N-terminal to the homeodomain. All the rest of the
homeodomain proteins are part of the Dispersed Superclass. There is a lot of divergence
within the latter class of homeodomains, both within the homeodomain and with respect
to additional domains found in these proteins. The Dispersed Superclass can be
subdivided into more than 16 classes or families (Gehring et al., 1994). Some of these
families are characterised by their similarity within the homeodomain, such as the even
skipped, caudal and distalless classes, whereas other families are distinguished by
additional domains they contain. For example, the p a ire d class proteins are
characterized by a paired domain, the PO U class have the POU domain and the
16

Chapter 1: Introduction
engrailed class contains four highly conserved protein segments, named EH-domains,
in addition to the homeodomain.
Besides the two superclasses of classical homeodomains, there are also some
atypical homeodomains which have more or less than 60 amino acids when the
sequences are aligned (Bürglin, 1997; Gehring et al., 1994). Nevertheless, the overall
three-dimensional conformation of these domains is still the same as for the typical
homeodomains.
1.1.3. Iroquois belong to the TALE class of homeodomains
A particular subset of atypical homeodomain transcription factors has emerged that
has three extra residues in the loop between helix 1 and helix 2 of the homeodomain and
has been given the name TALE (three amino acid loop extension) (Bertolino et al.,
1995). The TALE superclass of homeobox genes can be organized into four classes in
animals: PBC, MEIS, TGIF and IRO; two types in fungi: the mating type genes NI
AT YP and the CUP genes, and two different classes in plants: KNOX and BEL. The
IRQ class has a unique conserved motif downstream of the homeodomain. For the
KNOX class, a conserved domain, the KNOX domain, was defined upstream of the
homeodomain. TALE homeobox genes were already present in the common ancestor of
plants, fungi and animals and represent a branch distinct from the typical homeobox
genes (Bürglin, 1997). The loop between helix 1 and helix 2 at position 24-26 of the
homeodomain consists always of a proline-tyrosine-proline motif, except for the TGIF
group, which has an alanine instead of the proline at position 24. The polar residue (i.e.
glutamine) at position 50 of the typical homeodomains is replaced by a small, non polar
residue in the DNA-binding helix of the TALE homeodomain. In the case of the IRO
17

Chapter 1: Introduction
class it is an alanine, in the PBC class it is a glycine and in most of the other genes it is
an isoleucine. Position 50 is very critical for the DNA binding specificity of the
homeodomain. The fact that in TALE homeodomains a small non polar residue is at this
position suggests that the DNA-protein interaction of TALE proteins could be of a
different nature than that of typical homeodomains. In addition, DNA binding
specificity might be conferred by other parts of the protein (Bürglin, 1997). TALE class
homeodomains are thought to form homodimers or heterodimers with other
homeodomain proteins (Mann and Chan, 1996).
1.2. The Iroquois family
1.2.1. The Iroquois Complex
The Iroquois (Iro) family is a highly conserved subclass of TALE genes, which is
present in Drosophila, C.elegans, Xenopus, zebrafish, chick, mouse and humans. A
comparison of IRO sequences from different organisms revealed that the sequence
similarity is mainly restricted to the homeodomain region. In addition to their highly
conserved homeodomain, IRO proteins share a novel region of unknown function in the
C-terminal half of the molecule, known as the IRO-box (McNeill et a l, 1997).
Noteworthy is also an acidic patch downstream of the homeodomain, which has been
proposed to serve as a transcriptional activation domain (Bürglin, 1997). A motif
reminiscent of the central part of the EOF repeats of the Notch receptor protein is
located N-terminal of the homeodomain (fig. 1.1.), (Dahlsveen, 2002).
Members of the Iroquois family usually occur in genomic clusters of three genes,
similar to clusters formed by the original homeotic genes. In Drosophila, three such
closely related genes are located on the third chromosome at 69D (Netter et a l, 1998).
18

Chapter I: Introduction
Mirror
Ara
Caup
N
N
HD 640a
716a
E E 693a
clrx4
hlRX2
mlrx3
Xiro3
Ziro3
485a
HD
HD
417aa
507aa
448aa
420aa
Figure 1.1. Schematic diagram o f proteins o f the Iroquois family. The figure shows the different protein
domains o f Drosophila Iro proteins (Mirror, Ara and Caup) as well as some vertebrate representatives
(c=chick, h=human, m=mouse, X=Xenopus, Z= zebrafish). l l ie Iro/Irx proteins are highly homologous
within the homeodomain (HD) and the IRO Box (1). In addition, all Iro/Irx proteins contain an acidic
domain (A). The Drosophila proteins contain an EGF-like domain (N), proline rich (P) and glutamine
rich (Q) motifs. The latter is a eommon feature o f transcription factors in Drosophila. The fly proteins are
more similar to each other than to the vertebrate proteins (from Ina Dahlsveen, 2002).
19

Chapter 1: Introduction
They are called araucan and caupolican, which are the founder members of the
Iroquois family, and mirror (Gomez-Skarmeta et al., 1996; Kehl et al., 1998; McNeill
et al., 1997). Together they form the Iroquois Complex (Iro-C) that spans approximately
130 kb of DNA (Netter et al., 1998). The Iroquois Complex was initially identified
through fly mutations lacking the lateral bristles of the notum, leaving a median stripe
of hairs unaffected (fig. 1.2.), (Leyns et al., 1996). This pattern resembles the hairstyle
of the Iroquois American Indians (also known as Mohawk) - hence the name of the
locus.
The distance between the araucan (ara) and caupolican (caup) genes is less than
25kb whereas mirror (mirr) is found over 70kb downstream of caupolican. This
organisation reflects the degree of similarity between the gene products as well as the
similarity in expression patterns, araucan and caupolican encode highly similar proteins
which are identically expressed throughout development. The close proximity of the
genes and the similarity in their expression patterns suggests that araucan and
caupolican are controlled by the same enhancer elements and that they may functionally
replace each other (Cavodeassi et al., 2001; Gomez-Skarmeta et al., 1996). In contrast
mirror has an independent expression pattern that only occasionally overlaps with the
other Iro genes, and is not functionally redundant (Gomez-Skarmeta et al., 1996
McNeill et al., 1997). The expression pattern of mirror overlaps with araucan and
caupolican in the lateral epidermis, the proventriculus, the brain, the eye discs and in
small regions of the wing disc (Gomez-Skarmeta et ah, 1996; McNeill, 1997). mirror is
uniquely expressed in some areas during embryonic development, especially during
early stages, notably in the ventral nerve cord of the embryo. As mirror expression
overlaps with ara and caup in some areas, some ara/caup enhancers may also act on
mirror.
20

( 'huptcr ! : huroctucPi);}
Figure 1.2. The Iroquois phenotype. Lateral view o f the thorax in wild-type A) and in the Iroquois
mutant B). A) The macrochaetes are, from anterior (left) to posterior (right): Hu, humerais, aNP, anterior
notopleural, PS, presutural, pNP, posterior notopleural, aSA and pSA, anterior and posterior supraalars,
aPA and pPA, anterior and posterior postalars, aDC and pDC, anterior and posterior dorsocentrals, aSC
and pSC, anterior and posterior scutellars. (B) irol/iro2. The lateral region o f the notum lacks both
macrochaetes and microchaetes. Note that the humeral bristles (white arrow) are not all affected by the
mutations (from Grillenzoni el a i , 1998).
Zl

Chapter 1: Introduction
1.2.2. Genomic organisation
The genomic organisation of the Iroquois genes seems to be conserved among
species (Gomez-Skarmeta and Modolell, 2002). In mouse and humans six Iro genes
have been found on two different chromosomes, in two clusters of three genes each, just
like in Drosophila (Ogura et a i, 2001; Peters et al., 2000). The mammalian clusters
consist of Irx I, Irx 2 and Irx4 in one complex (cluster A) and Irx3, Irx5 and Irx6 in the
other complex (cluster B). The members of each cluster are most similar to the Irx gene
in the same position in the other cluster (i.e. IrxI~Irx3, Irx2~Irx5 and Irx4~Irx6). In
general the pattern of expression of the first two genes of each cluster is highly similar
and in some tissues, the paralogs Irxl/Irx3 and Irx2/Irx5 are also identically expressed
(Houweling et a l, 2001). This suggests the duplication of regulatory elements for the
Irx genes within the different clusters. It is interesting to note that the expression of the
third gene of a cluster, Irx4 or Irx6 is in general more divergent. This is reminiscent of
the situation with mirror and the other Iro members in Drosophila.
In Zebrafish, four genes have been identified to date. Z/ro7(putative Irx A
cluster) and Ziro3 and 5 (putative Irx B cluster). Intriguingly Ziro l is found at a third,
genomic location, raising the possibility that more than two Iro/Irx cluster exist in
Zebrafish (Lecaudey et a l, 2001). This would be consistent with the extra gene
duplication that took place during teleost evolution (Postlethwait et al., 1998; Robinson-
Rechavi et al., 2001).
Five genes have been identified in the chick and three in Xenopus. Mapping of
these genes indicates that they are also found in two clusters (Ogura et al., 2001; Wang
et al., 2001b). Caenorhabditis elegans (C.elegans) is, with one Iro/Irx gene, the only
exception to this rule. The expression of some orthologs in similar domains in different
vertebrate lineages also suggests that regulatory regions may be conserved during
22

Chapter 1: Introduction
evolution. For example, Xenopus Iro3, chick Irx3, mouse Irx3 and zebrafish Iro3 are all
expressed in an equivalent region of the dorso-ventral axis of the neural tube and in the
lateral mesoderm.
Vertebrate and invertebrate Iro/Irx genes are similarly regulated by Wnt and
TGF-B signalling molecules, which further suggests that controlling elements regulating
the ancestral Iro/Irx gene have been evolutionary conserved. Despite these similarities
in gene regulation and the relationship between the vertebrate Irx cluster and
Drosophila Iro cluster it is not clear if they originated from a common ancestral IroHrx
cluster. It could well be the result of an independent duplication event, giving rise to
three-gene clusters in ancestors of the insect and vertebrate lineages (Peters et a i,
2000). The main argument for the latter is the observation that the orientation of
transcription of genes within a cluster varies between Drosophila and the vertebrates
(fig. 1.3.) (Gomez-Skarmeta and Modolell, 2002). However, the best characterised
example of a conserved gene cluster is represented by the original Hox genes, which is
believed to have been duplicated along the evolution of vertebrates from a single
ancestral Hox gene cluster (reviewed in Finnerty and Martindale, 1998).
1.3. The role of the Iro/Irx genes In vertebrates
Iro genes have diverse functions in vertebrate development. Intially they are
expressed in large domains, where they apprear to specify the identity of large
territories, such as the neural plate in frogs and mice. Later in development Iro genes
are expressed in subdomains, where they are thought to function again to subdivide
those territories into smaller domains. The following paragraphs will summarise briefly
what is known to date about the role of the Iro/Irx genes in different vertebrate
organisms.
23

Chapter I : Introduction
IrxA cluster
IrxB cluster
Iro-C
frx3
>
frx5
caup
S p e d e s
Hu mon Mouse
HumanMouse
Chr
613
168
D fosoph ,lJ 31
' uiieni ‘ ipnlcn in & Cw^opnxnl
Figure 1.3. : Similar genomic organization o f the Iro/Irx genes in Drosophila and vertebrates. In human
and mouse, two clusters o f three Iro/Irx genes are located in the indicated chromosomes. Paralogous
genes in IrxA and IrxB clusters are labeled with similar tone colors. The Drosophila Iro genes, araucan
{ara), caupolican {caup) and mirror {mirr), are located in a unique cluster. Orientation o f arrows
represents direction o f transcription. Chr, chromosome (from: Gomez-Skarmeta and Modolell, 2002).
24

Chapter 1: Introduction
1,3.1. Xenopus laevis
Injection of mRNAs that encode wild type or modified versions of the Xiro
proteins has shown that the Xiro genes are essential for development of the neural plate
(Bellefroid et al., 1998; Gomez-Skarmeta et al., 2001; Gomez-Skarmeta et al., 1998).
Xiro genes have complex roles in neural development, as they appear to control both
proneural and neurogenic genes. Proneural genes promote neuronal differentiation
while neurogenic genes such as Notch and Delta, have a role in limiting the number of
cells that differentiate into neurons within the proneural domain (Chitnis et al., 1995;
Coffman et al., 1993).. The expression of neurogenic genes is often activated by bHLH
proneural transcription factors. Xiro genes positively regulate the expression of the
bHLH proneural genes ATH-3, Xash3 andXngnrl, which are implicated in neurulation.
The Notch ligand Xdeltal (XDll) is repressed by Xiro gene products. This repression is
probably direct as expressing a chimeric form of Xiro 1 that converts the Xirol repressor
into an activator, expands the X D ll expression domain (de la Calle-Mustienes et al.,
2002a). Like the Iro/Irx genes, bHLH proneural genes which function early in
development are expressed in wider domains that encompass the expression domain of
late expressed proneural genes. In order to emphasize the distinct roles of early and late
acting bHLH genes, they have been termed, “neuronal determination” and “neuronal
differentiation” genes, respectively (Bellefroid et al., 1998).
In Xenopus laevis, the formation of the presumptive neural plate is initiated by
the signalling activity of a small region in the mesoderm, which functions as an
organiser center, called the “Spemann organiser” (Spemann and Mangold, 1924). Bone
morphogenetic protein (BMP), a growth factor of the TGPP superfamily, is initially
expressed in the whole embryo. During gastrulation and formation of the Spemann
organiser, expression disappears from the dorsal side of the embryo, including the
25

Chapter 1: Introduction
dorsal mesoderm and the prospective neural plate (Fainsod et al., 1994). Xirol (and
probably Xiro2 and 3) regulate the size of the neural plate through repression of Bmp-4
signalling (Bellefroid et al., 1998; de la Calle-Mustienes et al., 2002b; Glavic et al.,
2001; Gomez-Skarmeta et al., 2001). Formation of the neural plate requires suppression
of the function of Bmp-4, as Bmp-4 represses neural fate and promotes epidermal
differentiation. Expression of Xirol is activated by Wnt signalling and expression of
Xirol is found in the Spemann organiser from the beginning of gastrulation (Glavic et
al., 2001). Xirol and Bmp-4 are initially expressed in an overlapping pattern in the
region of the prospective neural territory. Later this expression becomes
complementary. It has subsequently been shown that Xirol acts as a transcriptional
repressor of Bmp-4 and vice versa. Moreover, Xirol binds in vitro to the Bmp-4
promoter region, indicating that this suppression might be direct (Gomez-Skarmeta et
al., 2001). In addition, another gene XCoREST which is expressed at the borders of the
Xirol expressing territories has also been proposed to be repressed by Xirol (de la
Calle-Mustienes et al., 2002b).
Overexpression of Xirol induces a secondary axis as well as the ectopic
expression of several organiser genes like chordin, goosecoid and X lim -1. For that
reason, it is believed that Xiro genes have at least two different functions: control of the
neural plate and development of the Spemann organiser. Consistent with this model is
that Xiro3 expression is found in pluripotent precursors of both primary and secondary
neurons and glia cells, where it is positively regulated by the combined activity of the
gene products of the fibroblast growth factor and noggin (Bellefroid et al., 1998). This
suggests that Xiro3 is an earlier acting gene than all proneural genes identified in
vertebrates. Proneural genes act later in descendant cells and regulate subsequent steps
of neurogenesis (Ma et al., 1996).
26

Chapter 1: Introduction
1.3.2. Zebrafish
Ziro3 is highly homologous to Xenopus and mouse iroquoisS. Expression of
Ziro3 starts during gastrulation in the dorsal axial mesoderm that develops into the
notochord. Later, the expression is limited to the chordo-neural hinge in the tailbud.
Ziro3 expression also occurs in the central nervous system (CNS), excluding the
telencephalon (Tan et a l, 1999). In zebrafish, the organiser is thought to consist of two
regions, the yolk syncytial layer (YSL) and the shield. It appears to send signals that
affect formation of the shield in the overlying mesendoderm. The shield mainly gives
rise to the prechordal plate and notochord and is the zebrafish equivalent of the
Spemann organiser. A domain of dorsal deep cells located between the YSL and the
shield is marked by expression of the Ziro3 gene. This domain might be an additional
organiser subdomain in zebrafish residing at the vegetal side of the shield in the dorsal
mesendoderm (Kudoh and Dawid, 2001). As gastrulation proceeds, the Ziro3 positive
domain involutes and migrates to the animal pole. Ziro3 expression is regulated by
Nodal and Bmp antagonists. Overexpression of Ziro3 induces ectopic expression of
shield-specific genes. This effect is mimicked by a Ziro3-Engrailed transcriptional
repressor domain fusion, whereas an Ziro3-VP16 activator domain fusion behaved as a
dominant negative or antimorphic form (Kudoh and Dawid, 2001), These results
suggest that Ziro3 acts as a transcriptional repressor, and is another example of an
Iro/Irx gene acting in organiser formation.
Expression of Zirol is complementary to that of Ziro3 in the notochord and later
in the gut. Ziro5 expression mostly overlaps with that of Ziro3 (Wang et a i, 2001b).
Later, Ziro3 transcripts are present in the CNS from the mid-brain to the posterior end
of spinal cord and in the midline axial structures (notochord, fioorplate and hypochord).
27

Chapter 1: Introduction
It is also interesting to note that ZiroS expression is stronger in the even rhombomeres
(r2 and r4), weaker in the odd rhombomers (rl,r3 and r5) and absent in rhombomere 6
(Tan et a l, 1999).
Ziro7 is a novel, divergent member of the Ira family. It is expressed at early
epiboly stages in the dorsal half of the zebrafish embryo, with a higher level in the
dorso-lateral margin. From mid-gastrulation stages onward, Zirol is expressed in a large
transversal stripe in the future neural plate (Lecaudey et a l, 2001). At the tailbud stage
Zirol and Zirol are expressed in partially overlapping pattern with the proneural gene,
neurogeninl (ngnl) in the neuroectoderm This raises the question if the expression of
ngnl is a consequence of Ziro function. Indeed ectopic expression of both Zirol and
Zirol is able to promote expression of this proneuronal gene. In addition, both the use
of a chimeric protein, En-Zz>o7HD, which should act as a suppressor, and the equivalent
form of an activator, VP-Z/ro7HD, induce ectopic expression of ngnl (Itoh et ah,
2002). In this context, induction of ngnl using the repressor form has only been seen in
the ventral ectoderm where high levels of Bmp are present. Consequently, this effect
could be indirect by antagonising Bmp and neuralizing the ectoderm. This possibility is
supported by the ability of Zirol, Z iro l and En-ZirolHD to suppress expression of
gatal, a BMP target. The activator form only induces ngnl expression dorsally in the
neurectoderm, a tissue which is already neuralized. Here Ziro genes might function
directly to induce expression of ng n l (Itoh et al., 2002). However, endogenous
expression of ngnl is only observed in a restricted subdomain of the normal Ziro
expression domain, suggesting that additional factors regulate ngnl expression.
In addition, a knockdown experiment using morpholino oligos of Zirol and
Zirol revealed an essential function of Ziro l for the determination of neurons in the
trigeminal ganglia and formation of the neural crest. They are also essential for the
28

Chapter 1: Introduction
establishment of the isthmic organiser, a specialised tissue with secondary organiser
properties. Expression of Zirol and Ziro7 is also positively affected by Wnt signalling.
(Itoh et al., 2002).
1.3.3. Chick
Members of the Iroquois family have also been found in the chick, where they
are involved in many different developmental processes such as heart formation and in
the development of the CNS. The vertebrate heart consists of two types of chambers,
the atria and the ventricles. The chicken Iroquois homologue clrx4 is expressed only in
the ventricle at all stages of heart development. Moreover, chick, mice and zebrafish all
show ventricle-specific expression of Irx4, suggesting that Irx4 has an evolutionary
conserved role in heart development (Bao et al., 1999; Bruneau et al., 2000). In
contrast, the ventricle myosin heavy chain-1 (VMHCl) and the atrial myosin heavy
chain-1 (AMHCl) genes are expressed in the entire heart tube at the initial stages of
development. Later VMHCl is restricted to the ventricle and AMHCl to the atria. The
determination of the chambers is most likely mediated by clrx4 as misexpression of
clrx4 in the atria upregulates VMHCl and downregulates AMHCl expression (Bao et
ah, 1999; Wang et al., 2001a). Furthermore, a chimeric RCAS-H+en^ Irx4 protein,
principally consisting of the Irx4 homeodomain fused to the engrailed repressor domain,
has no effect if injected in the atria but leads to AMHCl expression if injected in the
ventricle. Thus, it seems that Irx4 is also required to maintain the ventricular profile of
myosin heavy chain gene expression during normal development. clrx4 is also
expressed in the eye, in the hindbrain and in the developing feather buds (Bao et al.,
1999).
clrx2 is initially expressed in subdomains of the neural plate where its
29

Chapter 1: Introduction
expression domain is largely complementary to that of Bmp4. For this reason clrx2
might also play a role in neurulation. Later in development clrx2 is found in the
developing hindbrain. Unlike its homologue clrx4, clrx2 is not expressed in the
developing heart, nor in the eye. Furthermore, clrx2 is not expressed in the limbs,
opposite to its murine ortholog mlrx2, (Goriely et al., 1999).
clrx3 was found to have a role in the régionalisation of the forebrain, where it
might act to promote midbrain identity. It is expressed posterior to a boundary of
subdivision within the forebrain called the zona limit ans intratalamica (ZLI). This
border is determined by the mutual repression of clrx3 and the homeodomain encoding
gene Six3. Also, Fgf8 and Sonic Hedgehog (SHH) signalling are modulated in different
ways in the different domains of clrx3 and Six3 expression. As a result, only in the
clrx3 domain Enl and Nkx6.1 are activated in response to Fgf8 and SHH, respectively.
Furthermore, ectopic expression of clrx3 in the forebrain induces the formation of
abnormal bulges (Kobayashi et al., 2002). Within the developing neural tube, clrx3 acts
in combination with other transcription factors to create dorso-ventral (D/V) territories,
from which different types of neurons arise (Briscoe et al., 2000; Gomez-Skarmeta and
Modolell, 2002). In this context, OUg2, a bHLH transcriptional repressor has been
identified to downregulate clrx3 in the ventral domain of the dorsal spinal cord. On the
other hand, 0Ug2 is repressed by clrx3 in the dorsal domain (Mizuguchi et ah, 2001;
Novitch et ah, 2001).
1.3.4. Mouse
As in the chick, members of the Iroquois family seem to play a role in various
developmental processes in the mouse. In general, murine Irx (mirx) genes of both
30

Chapter I: Introduction
cluster A (mirxl, -2 and -4) and cluster B (mlrx3, -5 and -6) are expressed in the brain,
the neural tube, the lung, the teeth anlagen, the limbs, the heart and in the primordial
cells of the vibrissae which give rise to the straight and stiff sensory hairs in the face of
the animal (Becker et al., 2001; Cohen et al., 2000; Houweling et al., 2001;
Mummenhoff et al., 2001). Moreover, in situ hybridisation analysis revealed cluster
specific expression in some tissues. For example, only members of the mIrxA cluster are
found in the pancreas, the digits or in the rib primordia. In contrast, specific expression
of genes from the mIrxB cluster has been reported in the gonads, the mammary gland
primordia and the interdigital mesenchyme. These results indicate that the mIrx genes
are coordinately regulated at the level of the cluster. On the other hand, some tissues are
void of expression of individual members of a cluster. For example, mlrx4 is not
expressed in the dorsal spinal cord nor in the limbs (Houweling et ah, 2001). However,
mlrx4 expression is found early in the linear heart tube in the ventricular segment and is
later restricted to the ventricular myocardium similar to clrx4 in the chick (Bao et al.,
1999; Bruneau et al., 2000). Four other mirx genes are also expressed in the mouse
heart in more or less overlapping patterns (Christoffels et al., 2000). mlrx4 expression
in the ventricular myocardium is initiated and maintained by the gene products of the
transcription factors Nkx2-5 and dHand, respectively (Bruneau et al., 2000). Mice
deficient for the mlrx4 gene exhibit reduced ventricular expression of the bHLH
transcription factor eHand placing it downstream of mlrx4. Furthermore, derepression
of the atrial natriuretic factor (ANF), an atrial chamber specific gene, plus increased
levels of a-skeletin actin has been observed in the ventricle shortly after birth. Normally
mlrx4 expression is maintained in the postnatal ventricular myocardium, supporting a
role in repressing these genes after birth as well as in the embryo, mirx4 deficient mice
are viable but develop cardiomyopathy as adults. These mice have normal embryonic
31

Chapter I: Introduction
ventricular expression of m irx l but exhibit increased levels of mirx2 expression.
Therefore, it has been suggested that other murine Irx genes may substitute for loss of
mlrx4 function during early heart development. In conclusion, mlrx4 is not essential for
ventricular chamber formation but is required for normal ventricular function (Bruneau
et al., 2000).
Similar to their Irx orthologs in Xenopus, mirx are believed to positively regulate
expression of the proneural gene MashI during early neurogenesis. Mashl promotes
neuronal differentiation at the expense of the glial fate in the CNS (Cohen et al., 2000).
Besides neurons and glial cells, oligodendrocytes and astrocytes are also found in the
CNS. Two genes Oligl and 0Ug2 are specifically expressed in oligodendrocyte
precursors and are required for motoneuron and oligodendrocyte differentiation. These
precursors emerge from a highly restricted, ventral domain in the dorsal spinal cord,
demarcated by Olig gene expression and restricted dorsally by mlrx3. Analogous to the
observed mutual repression of clrx3 and OHg2 in chick, Oligl and 0Hg2 double mutant
mice display ventral expansion of Irx3 expression. This expansion is coupled with cell
fate changes of oligodendroytes into astrocytes (Zhou and Anderson, 2002). Therefore it
is likely that Oligl and 0Ug2 are negatively regulated by mlrx3. In addition, mirx3 is
the only murine Irx gene active in the notochord, which stands in contrast to the three
zebrafish members found here (Wang et al., 2001b).
Finally, the molecular analysis of the fused toes (Ft) mutation in mouse has been
reported recently (Peters et al., 2002). This mutation is deficient for the entire mirx B
cluster as well as for three other genes. Heterozyotes mice are viable, but have thymic
hyperplasia and fusion of the forelimb digits. Homozygous Ft mice die during
embryonic development, probably due to malformation of the heart and defects in left-
right axis formation. In addition, they exhibit polycactyly, deformation of cranial
32

Chapter 1: Introduction
structures and malformation of the ventral domain of the spinal cord (Van der Hoeven et
al., 1994). Many of the observed defects correspond to areas in which the mirx genes
would have been expressed normally, suggesting that the Irx genes are responsible for
at least some of the patterning defects .
1.3.5. Human
The analysis of human Irx genes is so far just at its onset. Nevertheless, it is known
that human Irx2 is expressed in different tissues such as the lung, uterus, the salivary
gland and kidney. Unlike its mouse ortholog mlrx2, hlrx2 transcript has been detected
in the mammary gland. Furthermore, hlrx2 has been reported to be maintained in human
breast cancer cells while many mouse homeobox genes lose their expression during
neoplasia (Lewis et al., 1999).
1.3.6. Irx genes share similar functions in different organisms
The vertebrate Irx genes play a role in many developmental processes, however,
in all species analysed to date, functional studies and the expression patterns suggest
that they are involved in neural development and formation of the CNS. Initially, Irx
genes are implicated in the specification of the neuroectoderm as prepattern factors that
regulate proneural genes, whereas later they subdivide the neural plate into smaller
regions. This observation raises the possibility that members of the Iroquois Complex in
Drosophila might also be involved in the formation of the CNS.
Like the Drosophila Iroquois genes, the vertebrate Irx genes usually occur in
clusters of three genes and their close proximity to each other might allow them to be
controlled by shared regulatory elements leading to overlapping expression in several
vertebrate species (Cavodeassi et al. 2001). These shared enhancers might be the reason
33

Chapter 1: Introduction
why the Iro/Irx gene complexes have persisted in both Drosophila and vertebrates. It is
also interesting to note that both Drosophila (see chapter 1.4.1) and vertebrate Iro/Irx
genes can be positively regulated by Wnt signalling and negatively regulated by TGFp
signalling, suggesting that parts of the regulatory mechanisms could be conserved. So
in addition to the homology within the homeodomain, and the conserved genomic
organisation, it seems that the Iro/Irx proteins are controlled by similar regulatory
mechanisms in Drosophila and vertebrates.
Finally, it is well documented that araucan and caupolican are
functionally redundant in the Drosophila. The observation that the mouse Irx2 gene
might be able to substitute for Irx4 during heart development, supports the possibility of
functional redundancy of members of the Iroquois family in vertebrates. Another
example is found in zebrafish, where the use of morpholinos for Zirol and Ziro?
revealed redundant function in the determination of specific neurons and in the
formation of the neural crest. Therefore a limited amount of functional redundancy is
characteristic of the Iro family.
1.4. The Iroquois complex in Drosophila
1.4.1. The role of Mirror and the Iro-C in the eye
mirror (mirr) was first discovered due to a P element insertion, in the 69D region of
the third chromosome, which carries a miniwhite gene and lacZ reporter gene. Flies
carrying such a transgene show strong expression of the miniwhite gene exclusively in
the dorsal half of the eye (fig. 1.4.), (McNeill et al., 1997). The fly eye is composed of
dorsal and ventral fields of photoreceptor clusters, called ommatidia. Ommatidia in the
dorsal half of the eye are the mirror-image of those in the ventral half. They are
34

Chapter 1: Introduction
characterised by opposed orientation and chirality, thus demonstrating enantiomorphic
polarity. The boundary where the dorsal and the ventral fields meet is known as the
equator (fig. 1.4.).
mirror expression in the dorsal half of the eye is dependent on Wingless and
Hedgehog signalling and it is believed that Mirror plays a key role in the formation of
the equator (Cavodeassi et al., 1999; Cho and Choi, 1998; Heberlein et a l, 1998; Kehl
et a l, 1998; McNeill et a l, 1997). This midline is a highly regular structure that bisects
the eye into a dorsal and a ventral half. It rarely deviates more than one ommatidial
width as it crosses the eye. In a wild type eye, ommatidia of dorsal polarity are never
found in the ventral field and vice versa (Wolff and Ready, 1991). Moreover, clones of
cells lacking mirror function in the eye tend not to mix with surrounding mirror-
expressing cells. Therefore Mirror might also act to sharpen the equator by reducing the
mixing of dorsal and ventral cells through control of cell affinities (Yang et a l, 1999).
Formation of the equator begins in the eye imaginai disc - the larval precursor of the
adult eye. The undifferentiated eye imaginai disc consists of a single layer of
epithelium. Differentiation starts at the posterior margin of the disc during the third
instar larval stage. A wave of differentiation, which is marked by an indentation in the
epithelium, known as the morphogenetic furrow, sweeps across the eye from posterior
to anterior. The ommatidial preclusters which form behind the furrow are initially
identical in both halves of the eye. The first cell to differentiate in the cluster is the R8
photoreceptor precursor in the posterior-most position, followed by the R2/R5 pair and
the R3/R4 pair. The R1/R6 pair and R7 join the cluster later. The more polar cells of
each pair become R4-R6 and the more equatorial R1-R3 (flgl.5.). Finally, the clusters
rotate 90 degrees in opposite direction in the two halves of the eye in a process thought
to be regulated by signals emanating from the dorsal/ventral boundary formed earlier
35

( 'haplcr I : Introduction
At ,
Figure 1.4. Mirror is involved in forming the equator The left panel shows the enhancer trap line (mirr^')
expressing the miniwhite gene (which results in red pigmentation) controlled by Mirror. The expression
pattern is restricted to the dorsal half o f the eye The right panel shows a section through the equatorial
region o f an adult eye. Ommatidia located dorsal to the equator (red line) are reflected like a mirror image
in the ventral half, hence the name for the gene mirror. The polarity o f the ommatidium is visualised by
the positions o f the rhabdomers from each photoreceptor arranged in a stereotypical trapezoid shape.
Usually, the equator moves up an down one ommatidial width as it traverses the eye (pictures by Helen
McNeill).
Vo

( 'haplcr 1 : Iiilrotluclioii
M orphogenetic furrow
H quftfor - -
Dorsal pole
^ ^§ %
I© §
% A
%> i§3 Qb I©£&■- #)%j ©
<> A
Anterior TÉL Po^enor
Ventral pole
Figure 1.5. Development o f the eye-antennal imaginai disc. The photoreceptor clusters differentiate
behind the morphogenetic furrow and start rotating in opposite directions in the dorsal and ventral half
only once the cell fate is determined. Photoreceptor differentiation starts with the specification o f the R8
cell (yellow) followed by the R3/4 pair (green) and the R2/5 pair (blue). The R l/6 (brown) and R7 (red)
photoreceptors are recruited later to the cluster (from Blair, 1999, and Ina Dahlsveen, 2002).
Mirror
Figure 1.6. Mirror expression pattern in the third instar eye disc. A) Mirror protein (green) is found in the
dorsal half o f the eye disc and expression ends at the midline (arrow-head) The morphogenetic furrow is
marked with an arrow. B) Mirror and Fringe (red) are complementary expressed and meet at the equator
(arrow head) (pictures by Helen McNeill).
37

Chapter 1: Introduction
(Blair, 1999). As a consequence, the eye does not become simply divided by the
midline, but polarized in the plane of the epithelium. This organisation of cells or
clusters of cells is also known as planar polarity.
Mirror is highly expressed in dorsal cells in front of the furrow and maintained
in the R3/R4 and R2/R5 pair after the furrow (Dahlsveen, 2002). It is believed that
Mirror restricts expression of Fringe to the ventral half of the eye and mediates Notch
activation at the ^rrngg-expressing boundary, which leads to formation of the equator
(fig. 1.6). In addition, mirror is not the only Iroquois member expressed in the dorsal
half of the eye. araucan and caupolican also exhibit a dorsal-specific expression
pattern. In fact, the expression boundary of all three genes of the Iroquois complex lies
at the equator and it is believed that they can functionally substitute each other
(Dominguez and de Celis, 1998; Yang et ah, 1999).
The dorsal expression of the Iro-C results in ventral expression of the
glycosyltransferase Fringe which activates the Notch ligand Serrate. Another Notch
ligand. Delta is expressed dorsally (fig.1.7.). Serrate and Delta are differentially
modulated by Fringe (Papayannopoulos et ah, 1998) and it is this interplay that leads to
high levels of Notch activity at the dorsal/ventral boundary in a process almost identical
to the formation of dorsoventral boundary cells in the wing imaginai disc (Cho and
Choi, 1998). Ectopic expression of Caup or Mirr can repress fringe expression in the
eye, indicating that fringe might be restricted to the ventral half because it is normally
repressed dorsally by the proteins of the Iroquois complex (Dominguez and de Celis,
1998). Misexpression of the Iro-C throughout the entire eye generally results in a small
eye phenotype. Also, removing mirror function from the eye and antennal disc, results
in ectopic fringe expression and development of new mini-equators (fig. 1.8A) at the
equatorial side of the border (Yang et ah, 1999).
38

( 'haplcr 1 : Introduction
Figure 1.7. A model for equator specification in the developing eye. A number o f signals - Unpaired
(Upd) from the posterior equator. Wingless (Wg) from the poles and unknown factors (?) from the ventral
pole - combine to determine the extent o f dorsal Iro-C expression. Iro-C (only Mirr is shown) controls the
dorsal production o f Delta (Dl) and the ventral production o f Serrate (Ser) and Fringe (Png). Fringe
blocks Serrate reception ventrally, but potentiates Delta reception at the dorsal/ventral midline. Ventral
Serrate signals to adjacent dorsal cells and dorsal Delta signals to adjacent ventral cells. This interchange
situates and reinforces Notch signalling at the equator and induce the production o f a unknown factor X,
responsible to set up planar polarity in the eye. Wingless suppress Decapentaplegic (Dpp) to prevent
formation o f ectopic furrows from the lateral margins and is important in the regulation of Mirror
expression. Anterior is to the right, posterior to the left and doral is up (modified after Cho and Choi,
1998).
•'A
: ^A ' A '■ * : A
Figure 1.8. Effects o f mirror loss-of-function in the adult eye. A) A section through the eye showing a
clone o f mirror (marked by the absence of pigmentation) which leads to the generation of ectopic mini
equators and reversal o f ommatidial polarity at the equatorial border B) Dramatic protrusion in the dorsal
posterior region of the eye (from Yang et a i , 1999).
VI

Chapter 1: Introduction
Protrusions form occasionally in the dorsal posterior region of the eye (fig.l.SB),
(Cavodeassi et a l, 2000; Yang et al., 1999).
In addition, the removal of Iro-C occasionally results in a duplication of the
antenna disc, which differentiates an ectopic antenna and maxillary palpus and
transforms the dorsal head capsule into ventral structures (Cavodeassi et al., 2000; Yang
et al., 1999). These findings support a dorsal selector role for the genes of the Iro-C in
the eye and head capsule. It is noteworthy that the removal of the entire Iro-C produces
the same phenotypes as removal of ara and caup together. Thus, mirror might be
dispensable to suppress the ventral identity in the dorsal head. In contrast, Iro-C genes
ectopically expressed in the presumptive ventral head territory do not cause the reverse
transformation (Diez del Corral et al., 1999). A reciprocal transformation of for
example a haltere into a wing and vice versa is characteristic for homeotic genes like
Ubx in loss-of-function and gain-of-function experiments. For that reason it has been
proposed that the proteins of the Iro-C act as transcription factors in multimeric
complexes, maybe as cofactors for Hox proteins. Their absence would impair the
function of these complexes, thereby causing the dorsal-to-ventral transformations, but
their ectopic expression in ventral cells would not necessarily result in the opposite
effect if other members of such a complex were unavailable in these cells.
mirror/Iro-C clones in the dorsal half of the eye give rise to very round clones
with smooth borders as if mutant cells minimise their contact with surrounding cells. In
contrast, when Iro-C clones are induced in the ventral part of the eye they form smooth
borders with boundary cells at the midline but they have wiggly borders where they are
in contact with cells of ventral origin (Cavodeassi et al., 1999; Yang et al., 1999)
(fig. 1.9.). Thus the Iro-C has a dual role in dorsal/ventral patterning of the eye.
It restricts the activation of Notch signalling to the border where dorsal and
40

Chapter 1: Introduction
ventral cells meet by repressing Fringe in the dorsal cells. In addition, it acts to sort the
dorsal cells from ventral cells by reducing cell-cell mixing at the boundary. As for
Apterous in the wing, it is not known which genes are directly regulated by the Iro-C in
the eye to confer the affinity difference between dorsal and ventral cells.
Analogous to the above, induction of fringe clones in the ventral half of the eye
leads to non-autonomous repolarisation of wild type ommatidia at the equatorial border
of the clone but at long distances. Additionally, ventral fringe clones ectopically express
the Notch ligand Delta and dorsal fringe clones have wiggly borders. Early ubiquitous
expression of Fringe causes reduction in the size of the eye disc, which is due to a
failure to activate Notch at a fringe expressing - non expressing border. In contrast,
Mirror expression is not significantly changed in these eye discs (Cho and Choi, 1998;
Dominguez and de Celis, 1998). Thus Mirror represses fringe but not vice versa. In
conclusion of the above, it has been proposed that the Iro-C genes specify the identity
of dorsal cells in the eye and that the juxtaposition of Iro-C expressing and non
expressing cells serves to establish a pattern-organising centre (Cavodeassi et al., 1999).
1.4.2. The role of Mirror and the Iro-C in the wing and the notum
Similar to the eye/antenna disc, the Iro-C is expressed at least from the early
second instar developmental stage, in the entire dorsalmost part of the wing disc, which
will give rise to the dorsal mesothorax (mostly the notum) of the fly. The transcription
factor Apterous has been described to act as a dorsal selector, subdividing the wing disc
into a dorsal and a ventral compartment. The dorsal and ventral domains of Serrate,
Delta and Fringe expression are inverted in the wing (Milan et a l, 2001a; Milan et al.,
2001b). Interestingly, in the wing the Iro-C do not appear to have a direct role in
41

C'fiaptcr I: Inlrocliiclion
Figure 1.9. Effects o f Iro-(^ loss-of-function in the eye imaginai disc. A) Iro-C mutant celis in the dorsal
half o f the eye form round clones with smooth borders B) Iro-C mutant cells in the ventral half have
wiggly borders (from Cavodeassi et a i, 1999).
D
Wing
Apt, Iro-C
Fng, Ser,
Caps, Trn, Msh,
— ff .
Dl
Eve
Iro-CD
V Ser, Fng
Figure 1.10. Overview o f gene expression in the dorsal (D) and ventral (V) compartments o f the wing
and the eye disc Notch becomes activated at the midline. Apterous (Apt), Fringe (Fng), Serrate (Ser),
Capricious (Caps), Tartan (Tm), Muscle segment homeobox (Msh), Notch (N), Delta (Dl), Iroquois
complex (Iro-C).
4 2

Chapter 1: Introduction
oonXxoWmg fringe expression, instead in this tissue Apterous functions to regulate fringe
expression. In fact, Iro-C proteins and fringe are co-expressed in some regions of the
wing disc, suggesting that the enhancer elements that drive fringe expression in the
wing do not contain Iro binding sites. Significantly, apterous is not expressed in the eye
imaginai disc and in its place the Iro-C may be acting as dorsal selector genes to
establish the dorsal and ventral compartments (fig. 1.10.), (Blair, 1999).
It is believed that the Iro-C contributes to confer dorsal identity and set up an
organiser centre that patterns the tissue at both sides of the boundary of Iro-C
expression in the wing. This boundary corresponds to the border between the
developing notum and the wing hinge (Fig. 1.11.). Removal of Iro-C function causes the
prospective notum cells to adopt a wing hinge fate. Thus, expression of the Iro-C is
necessary for notum specification, in contrast Apterous is largely dispensable for the
development of the notum (Cohen et a l, 1992; Diez del Corral et ah, 1999).
Mirror expression overlaps with that of Ara and Caup in the wing, but seems to
be missing in areas of wing vein formation. Correspondingly, mirror mutants do not
show any wing vein defects. Instead, mirror is essential for formation of the alula, a
basal structure of the wing located in the hinge region between the notum and the base
of the wing veins (fig. 1.12.). Moreover disruption of mirror function give rise to adult
flies with held-out wings and crumpled halters (Kehl et al., 1998; McNeill et al., 1997).
The Drosophila notum, the dorsal body wall of the thorax, bears 11 pairs of
precisely located large bristles, the macrochaetes and about 200 smaller bristles
arranged in rows, the microchaetes (see fig. 1.2.). Macrochaetes are mechanosensory
organs composed of four cells: two cells form the bristle shaft and its socket and the
other two give rise to a sensory neuron and its associated sheath cell. These four cells
43

( 'haplcr I : huroduclion
Iro-C clone
Iro-CLNdomain ;
Hinge
Wing
Figure 1.11. Cartoon of a mature wing disc with indication o f the fates o f the relevant regions. Dark and
light blue lines indicate compartment borders (D, dorsal, V, ventral; A, anterior; P, posterior
compartments). Iro-C domain o f expression is coextensive with the presumptive notum in young wing
discs. This expression is necessary for the specification o f the notum, as Iro-C- clones transform notum
into wing hinge (indicated by a green H) The border o f notum Iro-C expressing and hinge Iro-C-non-
expressing cells is a pattern organizing center (arrows), as revealed by the effects o f Iro-C- clones on the
surrounding wild-type tissue which acquires properties o f the lateral notum (LN) (from Cavodeassi ei a i,
2001 ).
Figure 1.12. Lack o f alula formation in mirror mutant flies. A) Wild-type alula (arrow). B) The
mirrSail/mirrBI-12 mutation is semi-lethal but escapers show outheld wings and the complete lack of
alula (arrowhead) (from Kehl et a i , 1998).

Chapter 1: Introduction
originate from a single progenitor cell, the sensory organ mother cell (SMC). The
formation of SMCs at the right position depends on the function of the proneural genes
acheate (ac) and scute (sc) (reviewed in Campuzano and Modolell, 1992). The acheate-
scute complex (AS-C), consists of achaete, scute, asense and lethal o f scute which
encodes four related proteins, each containing a basic helix loop helix (bHLH) domain.
lethal o f scute (I ’sc) together with another bHLH protein, daughterless, has been shown
to act as a transcriptional activator in a yeast assay (Cabrera and Alonso, 1991). Ectopic
expression and rescue experiments have shown that the four bHLH genes of the AS-C
can substitute for each other to produce morphologically indistinguishable bristles
(Brand et ah, 1993; Dominguez and Campuzano, 1993; Giebel et al., 1997; Hinz et a l,
1994; Martin-Bermudo et al., 1993; Rodriguez et al., 1990). The genes of the Iroquois
Complex are important for the development of 8 of the 11 macrochaetae in each hemi-
notum as combinations of some Iro alleles leads to loss or reduced levels of these
bristles (Leyns et al. 1996). In addition, it has been shown that a hypomorphic allele of
mirror leads to the loss of a specific subset of bristles, namely the PS and PA bristles
(Kehl et al., 1998; Dahlsveen, 2002), (see also figure 1.2.A).
The Iroquois phenotype of bristle loss is due to loss of expression of the
proneural genes of the AS-C complex in the imaginai disc epithelium that gives rise to
the lateral notum (Gomez-Skarmeta et al., 1996). Even though the AS-C defines the
exact position of the SMCs they are not involved in specifying the type of sensory
projection of the notai bristles. It has been shown that the genes of the Iro-C have an
additional role in conferring a lateral identity (ipsilateral projection) to sensory neurons
(fig. 1.13.), (Grillenzoni et al., 1998). Each bristle in the notum is innervated by a single
bipolar neuron, which extends its dendrite towards the base of the bristle shaft, and its
axon towards and into the CNS. The morphology of the axon projection varies
45

( haplcr I : fniriHhirln >n
V
Figure 1.13. The Iro-C confer lateral identity to sensory neurons in the notum o f the fly. A) Contra
lateral projection o f a wild type medial macrochaete (aDC). B) Ipsilateral projection o f a WT lateral
macrochaete (pSA). C) Sensory projection with contralateral branches o f a rarely formed lateral
macrochaete in an Iro-C mutant background carrying a gain o f function allele {H airy-w ing^) o f the AS-C
(HlV*‘ ;iro'/iro^). The dashed line indicates the midline (from Grillenzoni el a i, 1998).
44)

Chapter 1: Introduction
depending on the position of the bristle. Contralateral branches are characteristic for
neurons innervating the medially located macrochaetes (DCs, SCs and pPA, see
fig. 1.2.) and the vast majority of the microchaetes. In contrast, the neurons innervating
the lateral macrochaetes (NPs, aSA, pSA and aPA) and the most lateral microchaetes
have a projection confined to the ipsilateral half of the CNS. The Iro-C confers a lateral
identity on these neurons, resulting in appropriate lateral projections.
1.4.3. The role of Mirror during oogenesis
Events during oogenesis are very important for the regional specification of the
Drosophila embryo, and the egg is laid with considerable pattern already specified. At
the start of oogenesis, one cell divides four times to produce 16 cells, one of which
becomes the oocyte while the other 15 become nurse cells. The whole cluster of oocyte
and nurse cells is surrounded by follicle cells to form the egg chamber. The follicle cells
are derived from the gonads and are of somatic rather than germ-line origin. The signal
for dorsal/ventral asymmetry originates from the follicle cells outside the embryo and a
signal transduction pathway carries the information across the plasma membrane of the
embryo. The formation of the dorsal/ventral axis in the embryo is another example of
the role of the Iro-C to set up an organiser centre. Once more, mirror and fringe are
expressed in a complementary pattern. In fact, the ligand Gurken activates the BGFR
pathway which induces mirror expression. Mirror in turn spatially restricts fringe
expression, leading to a Notch activation border between the anterior-dorsal and the
posterior-ventral follicle cells (Zhao et al., 2000). From this border diffuses a long range
signal, possibly encoded by the Drosophila Bmp-4 homologue dpp, which can repress
expression of pipe, a potential glycosaminoglycan modifying enzyme (Jordan et al.,
47

Chapter 1: Introduction
2000; Peri et al., 2002). Pipe regulates the activity of a protease cascade that activates
the transmembrane receptor Toll and determines the dorsal/ventral pattern of the
Drosophila embryo. The dorsal/ventral polarity of the eggshell covering the embryo is
characterized by the positions of the dorsal appendages, which form at the dorsal
anterior side of the egg. Disruption of Mirror function leads to several defects during
oogenesis. Early defects in the encapsulation of the 16-cell cyst and its separation from
the germarium have been reported (Jordan et al., 2000; Zhao et al., 2000). However,
most impressive is the ventralisation of the eggshell, characterised by loss of the dorsal
appendages (fig. 1.14.), (McNeill et al., 1997). In contrast, overexpression of mirror
induces a stripe of rhomboid expression in the posterior follicle cells and leads to
dorsalisation of the eggshell, which can be seen by the formation of dorsal appendages
rudiments from anterior-ventral follicle cells (Jordan et al., 2000; Zhao et al., 2000). In
addition, over-expression of Mirror in ventral follicle cells leads to the repression of
twist, a gene required for the ventral cell fate, in the embryo. Such embryos lack the
ventral denticle belts completely or partially. Therefore, it has been suggested that
Mirror plays a role in the dor sal/ventral patterning of the embryo (Zhao et al., 2000).
However, the mirror loss-of-function analysis with respect to twist regulation in the
early embryo has not yet been conducted. Interestingly, removal of mirror leads to
expansion of pipe into dorsal follicle cells (Jordan and Ruohola-Baker, 1999) which is
activated by Windbeutel. Pipe is normally required in the ventral region of the follicular
epithelium to activate tw ist expression via Dorsal in the embryo (Nilson and
Schuepbach, 1998). For that reason it might be that over-expression of Mirror in the
ventral follicle cells affect twist expression in the embryo by repressing the expression
of pipe or even windbeutel in the follicle cells. Taken together, these results suggest that
Mirror is not only involved in setting up an organiser center in the oocyte but also
48

'Ii‘.ipirr ! : Ih im J ih lion
'4
%
m
Figure 1.14. Loss o f Mirror results in ventralisation o f the egg chamber. A) Dorsal appendages o f a wild
type embryo. B,C) Shows two examples o f the loss o f the dorsal appendages in mirror mutant embryos
(from Jordan et a i, 2000).
49

Chapter 1: Introduction
indirectly represses the ventral fate of embryonic cells (Zhao et al., 2000).
1,4,4. Mirror in the embryo
Loss-of-w/rror function or ectopic expression of mirror results in embryonic
lethality (McNeill et al., 1997; Zhao et al., 2000). mirror is expressed in a highly
dynamic pattern during embryogenesis. It is not maternally loaded into the embryo and
zygotic expression starts at the cellular blastoderm, at stage 6 as an anterior patch at the
site of the presumptive anterior midgut invagination (fig.l.l6A ). At the same time
mirror transcript accumulates dorsally prior to and while the dorsal folds form during
stage 7-8 (fig.l.l6B). The dorsal folds will give rise to part of the amnioserosa. During
germ band extension (stage 8-9), a morphological event similar to the vertebrate
convergent extension process, mirror continues to be expressed in these tissues
(fig.l.l6C). The role of mirror in these early stages is not very clear. Phenotypes that
may be associated with the early expression pattern of mirror are delays in germ band
retraction and dorsal closure (Helen McNeill, unpublished). Interestingly mirror mutant
embryos are often twisted along the anterior/posterior axis and show overgrowth of the
amnioserosa (Helen McNeill unpublished, fig. 1.15.).
From the beginning of stage 10 mirror is expressed in a one-cell-wide pattern in
segmentally repeated bands of ectodermal cells posterior and adjacent to the engrailed
expression domain (fig.l.l6D). Consequently mirror is expressed at the anterior side of
each segment. As the border between engra iled and wingless expressing cells
determines the parasegmental boundary (Lawrence and Struhl, 1996), so might be the
juxtaposition of m/rror-expressing and non-expressing cells be important for defining
the segmental border. In fact, mirror mutant embryos have cuticles which are smaller
50

Chupler I: Iniroducium
o p ra llc
cephalic lurro»
Figure 1.15. The amnioserosa at germ band extension in a mirror mutant embryo. A) The left panel
shows a dorsal view of a wild type embryo at germ band extension stage The germ band compressed the
flattening amnioserosa and pushed it between the posterior end of the extended germ band and the
cephalic furrow. B) The right hand panel shows an EM photograph o f a mirror mutant embryo at a
comparable stage with an overgrown amnioserosa. (A: from http://flybase.bio.indiana.edu; B: Courtesy
from Helen McNeill, unpublished).
SI

Chapter 1: Introduction
than wild type and with sparse denticles (McNeill et al., 1997). Most embryos fail to
hatch and the very few escapers die shortly after hatching. The most anterior expression
at this stage marks the region of the stomodeal invagination, mirror is also found in
dorsal lateral cells of the ectoderm (fïg.l.lôD). Shortly after that, mirror is expressed in
neuroepithelial cells and in delaminating neuroblasts. As the embryo undergoes germ-
band retraction, mirror expression appears transiently in the proventriculus as it
undergoes folding and is retained in the brain and the ventral nerve cord (fig.l.l6E and
F), (McNeill et al., 1997).
The expression pattern of araucan and caupolican in the embryo overlap with
mirror in the lateral epidermis, the proventriculus and in parts of the procephalon
(Gomez-Skarmeta et al., 1996; McNeill et al., 1997). Intriguingly, only mirror is
expressed in the ventral nerve cord, mirror mutant embryos have severe CNS and head
involution defects (Helen McNeill, unpublished), suggesting that mirror plays an
important role in the developing CNS. This would be consistent with the observation
that members of the Iro/Irx family are involved in the formation of the CNS in
vertebrates. The mirror mutant embryos {mir/"^^) used in this study die during late
embryonic development. However, mirr^^^ embryos still express low levels of mirror
transcript and is therefore not a true null mutation. Although no Mirror protein is
detected using an anti-Mirror antibody, it is still possible that some residual Mirror
activity persist in these mirr "* embryos, which might be sufficient to provide some
function.
Iroquois mutant embryos lack expression o f all three members of the Iroquois
Complex and therefore represent a true null mutation of Mirror activity. We initially
assessed changes in gene expression of potential target genes using Iroquois mutant
embryos to analyse the effect of complete loss of Mirror activity.
52

( 'haplcr I : Inlroduclion
D
Stage 6
B
Stage 7-8
i f
Stage 8-9
Stage 11
f fi-
V J.Stage 13
pre-hatching
Figure 1.16. in situ hybridisation pattern o f mirror in the embryo. A) mirror is expressed in dorsal and
anterior patches at stage 6. B and C) During gastrulation and germband extension mirror is expressed in
the dorsal folds and amnioserosa. D) At stage 11, mirror is expressed in a segmentally repeated pattern
E) mirror is expressed in the brain and the ventral nerve cord as well as the proventriculus at stage 13. F)
mirror expression continues in the CNS till late in embryonic development (from McNeill et a i, 1997).
S3

Chapter 1: Introduction
Whenever an Iro-C phenotype is observed a more detailed analysis using
specifically mirror mutant embryos is necessary to determine if the observed defects are
also observed by removing exclusively Mirror activity. This is in particular necessary
when Mirror function is analysed in the peripheral nevous system, where its expression
overlaps with Araucan and Caupolican. Nevertheless, the Iroquois mutant embryos can
be used to analyse Mirror function in the CNS, as mirror is the sole Iroquois Complex
member expressed in this tissue in the Drosophila embryo (Gomez-Skarmeta et al.,
1996; McNeill et a i, 1997).
1.4.5. Formation of the CNS in D r o s o p h i l a and mirror
expression in the CNS
Following gastrulation, a stripe of ventral ectoderm, the neurogenic region, gives
rise to neuromeres, which represent the segmented CNS. Unlike vertebrate neurulation,
in which the entire neural anlage invaginates as the neural tube. D rosophila
neurogenesis begins as single stem-cell-like precursors within the neurogenic region
enlarge and delaminate into the embryo. The cells remaining at the external surface give
rise to the epidermis. About 30 neuroblasts (NB) are formed in five spatiotemporally
different waves (S1-S5) in a stereotyped, roughly orthogonal array within a
hemisegment. A hemisegment is a bilateral half of a neuromere and is the
developmental unit of the segmented portion of the developing CNS. The hemisegments
are arrayed end to end down the length of the embryo (Skeath, 1999). The first 10 (SI)
neuroblasts partially fill in a 3x4 array, with three colunms running the length of the
embryo and four rows across each segment. Initially, there are four neuroblasts in the
medial and the lateral column and two neuroblasts in the intermediate colunrn with
54

Chapter 1: Introduction
respect to the midline (Goodman and Doe, 1993). Following this stage, the pattern
becomes more and more complex. However, each neuroblast has a unique, invariant cell
lineage to produce a diverse population of neurons and glia, and can be identified due to
its position, time of formation, or pattern of gene expression (Doe, 1992; Isshiki et aL,
2001). NBs that develop in the same relative position and at the same time but in
different hemisegments acquire the same fate (reviewed in Skeath, 1999). For that
reason, Drosphila neurogenesis may provide another example of planar polarity. NBs
divide asymmetrically to “bud o ff’ a series of smaller daughter cells, the ganglion
mother cells (GMC s). The GMC typically will give rise to two postmitotic neurons
and/or glia. For example the first GMC from NBl-1 always divides to produce the aCC
and pCC neurons. The neuroblasts in which mirror is expressed have been identified
using the mirror-lacZ enhancer trap (fig.1.17.), (Broadus et al., 1995; Doe, 2003).
Almost all NBs that express mirror delaminate at the anterior border of the neuromere.
At stage 9 the SI NBs 1-1, 3-2, 2-5 and at stage 10, the S2 NBs 1-2 and 2-2 begin to
express mirror. During stages 10-11 mirror is found in the NBs 6-1, 3-4, the glial
precursor and in all row 2 NBs. In addition, mirror is also expressed in the median
neuroblast. At stage 16 mirror-lacZ is found in a subset of neurons and glia in the
ventral nerve cord including the aCC and pCC neuron and the longitudinal glia. The
expression in these and other cells suggests that mirror might be lineally maintained
(Doe, 2003).
The proneural genes of the AS-C promote NB formation, whereas the
neurogenic genes antagonize proneural gene function and NB formation (reviewed in
Skeath, 1999). achaete {ac), scute (^c) and lethal o f scute (I’sc) are expressed in clusters
of cells in the ventral neuroectoderm from which the neuroblasts delaminate. Analysis
of the ventral neuroectoderm before and during delamination of the first two
55

( ihtpirr I : hiir.'.!h:rii(in
eS1 (18)o * oo ooooo oS3 (e10)
81 (e9)
o • o oooO : O
84 (11)
0 - 0 ^ ^
8 2 ( 9 )
i8 5 ( I 1 1 )
• =ear*efmnmgo iwrnhg* = *aDtiea pna
Figure 1.17. Mirror is expressed in delaminating neuroblasts. Schematic representation o f the
development o f neuroblasts (NB) in one hemi-segment from stage 9-11. NBs that express the mirror-lacZ
enhancer trap are marked in black, (from the Doe Lab website).
56,

Chapter 1: Introducrion
populations of neuroblasts shows, that cells in all regions of proneural gene activity
change their form prior to delamination (Martin-Bermudo et al., 1991; Skeath et al.,
1992).
Before delamination of the first population of neuroblasts, the coincident
expression pattern of ac and sc overlaps partially with that of Vsc. Even though the
ability to promote bristle formation is shared by all members of the AS-C, some of them
have their own specificity at the level of neuronal differentiation. In the CNS lethal o f
scute can not fully substitute for either achaete or scute. (Parras et ah, 1996; Skeath and
Doe, 1996). Altogether the three proneural genes are expressed in ten adjoining clusters
per hemisegment that determine where NBs develop.
This number corresponds to the final number of Nbs in the first wave (SI)
(Martin-Bermudo et al., 1991; Skeath et al., 1992). All cells with high levels of
proneural proteins have the potential to adopt the neural fate (Campos-Ortega, 1993).
Proneural gene expression, however, is progressively restricted to a single cell in each
cluster, i.e., the future neuroblast. This restriction results from inhibition of proneural
gene expression in remaining cells by the delaminating neuroblast and is mediated by
the neurogenic genes. In embryos lacking any one of the neurogenic genes this
restriction does not occur; instead all cells of each cluster continue to express proneural
genes and eventually become neuroblasts (Campos-Ortega, 1993). The expression
pattern of lethal o f scute (Romani et al., 1987) overlaps with mirror in the developing
CNS at stage 11.
Interestingly, it seems that the role of mirror as a prepattern gene and the
proneural genes is reversed - so far it is believed that the members of Iro-C act initially
in large territories which they subdivide later into smaller domains. This is clearly not a
function of Mirror during neurogenesis. Mirror is detected only in the forming NB
57

Chapter 1: Introduction
suggesting that its role must be more specific than simply acting as a prepattem gene.
Each of the four known AS-C genes is expressed in overlapping subsets of neuroblasts
at the right time and place to confer neuroblast identity. However their role in NB
specification remains unclear (Martin-Bermudo et aL, 1991). In this context, it might be
possible that Mirror assigns neuroblast identity.
1.5. Aim of the project:
The aim of the project was to identify genes that are regulated by the Drosophila
transcription factor Mirror and to study its role during the embryonic development of
the fly. The main goal was to identify direct Mirror targets.
1.5.1. The working hypothesis and the techniques
The working model is based on the understanding that increased levels of Mirror
protein will change the levels of transcription of target genes. Therefore, I analysed the
effects of ectopic overexpression of Mirror on gene expression in the embryo and
compared it to control embryos. To assess the changes in gene expression, I used
Drosophila Genome Arrays to identify potential Mirror targets.
1.5.2. The model
I used fly embryos which carry a mirror transgene under the control of the heat
shock promoter HSP70. Over-expression of Mirror can be achieved by exposing the
animal to a heat shock inducing temperature, normally around 37°C. The major
58

Chapter I: Introduction
advantage of this system is clearly the possibility to choose the moment and the
duration of ectopic Mirror expression. However, many genes respond with
transcriptional regulation to the heat shock and need either to be identified or otherwise
subtracted from the analysis. By examining gene expression shortly after induction of
ectopic Mirror expression, I hoped to identify primary Mirror targets, rather than genes
which change the level of their expression further down in the hierarchy. Nevertheless,
the microarray approach is only a tool to identify potential target genes. It should be
stressed that the microarray analysis does not automatically provide evidence that
candidate genes are indeed directly regulated by Mirror. To establish that a gene is a
real direct target of Mirror, additional experiments are required and in particular the
analysis of mirror mutant embryos is necessary to validate that expression of a putative
target is controlled by mirror in vivo.
To sum up, a positively regulated gene should be present in the same cell at the
same time as Mirror. Transcription of such a gene should be increased in Mirror over
expressing cells and decreased in corresponding cells of Mirror loss-of-function
mutants. For a negative regulated target gene the reverse case applies.
1.5.3. Does Mirror act as an activator or a repressor?
The variety of examples given previously demonstrates that members of the
Iroquois family may act as transcriptional activators or repressors or both. This makes it
difficult to predict a possible function for Mirror. It might even be possible that Mirror
acts differently in the different tissues during the various stages of the embryonic
development dependent on the availability of cofactors. Alternatively, the amount of
Mirror protein might be crucial for its activity. Expression of Mirror at low levels could
59

Chapter 1: Introduction
have an activating function, whereas the accumulation of Mirror in a cell could lead to
binding to low affinity DNA sites and turn it into a repressor.
A DNA binding site has been identified for another member of the Iro-C,
Araucan, located on the ac-sc enhancer element which drives expression of the
proneural genes achaete and scute at the presumptive vein L3 and the twin sensilla of
the wing margin (TSM). In vivo studies using a lacZ reporter construct carrying this
L3/TSM enhancer element suggested that Ara and Caup directly bound to it to act as
transcriptional activators (Gomez-Skarmeta et aL, 1996). However, over-expressing a
chimeric protein containing the Ara homeodomain fused to the Engrailed repressor
domain, which should function as a repressor, also expanded the proneural cluster. This
indicates that the regulation of the L3/TSM enhancer is not direct and that Ara acts as a
transcriptional repressor to control this enhancer (Cavodeassi et aL, 2001).
1.5.4. What are possible downstream targets of Mirror?
Fringe is expressed in complementary domains to Mirror in different tissues of
Drosophila. It has been shown that Mirror restricts the fringe expression domain in the
oocyte and in the eye disc (Cavodeassi et aL, 2001; Yang et aL, 1999; Zhao et aL,
2000). Furthermore, removal of Mirror expands the fringe expression domain.
Therefore, I would expect that fringe expression is repressed in Mirror overexpressing
embryos and that this downregulation can be recognized in the microarray analysis.
This would be extremely helpful to validate the specificity and relevance of this
approach. Nevertheless, it is not sure that fringe is directly regulated by Mirror. Other
expected candidates would be the members of the AS-C. It is believed that the proteins
of the Iroquois complex control these genes in the wing disc (Gomez-Skarmeta et aL,
60

Chapter 1: Introduction
1996).
Aside from the “expected” downstream targets of Mirror, the microarray
analysis should reveal unexpected genes. These could be genes of unknown function or
known genes which are involved in the different processes of embryonic development
which are marked by Mirror expression. Genes which are controlled by Mirror could
functionally be involved in the formation of the amnioserosa, dorsal closure, germ band
retraction, head involution, segmentation and/or in the formation of the CNS. Those
genes could play a role in cell-cell communication, cell adhesion, - proliferation, -
growth or could be other transcription factors. The role of Mirror in the various stages
of embryonic development is not known, nor have transcriptional targets of Mirror in
the embryo been unambiguously identified.
61

Chapter 1: Introducrion
62

Chapter 2: Materials and Methods
CHAPTER 2
Materials and Methods
Usually chemicals were obtained from BDH, Fluka or Sigma. Commonly used
buffers and media were made by the Cancer Research UK media services according to
(Sambrook et a i, 1989). Other materials are described within each section.
2.1. Drosophila me/anogaster techniques
2.1.1. Flies
The following flies listed below were used in this work:
Table 1. FIv list
Stock name: Genotype: Source
pKB12-m/>ror hs-sev-mirr Helen McNeillyw Ish-Horowicz labpY2Qlyw-mirror y , w ; hs-sev-mirr Mike Mohnsmirr Helen McNeillIro2 Df(3L) lR02/TM3-LacZ Juan Modolell\]A^-araucan ^7775; Juan ModolellUAS-mz>ror ^7775. uAS-mirr Helen McNeill^-Gal4 w; kr-Gal4/TM3 Ish-Horowicz labVP16-Gal4 w; P-tubulin-VP16-Gal4 Amo Müllersca-GdXA w; scaGal4/TM3 Heinrich Reichertprd-GdXA w; prd-Gal4/TM3 Bloomington, 1947
63

Chapter 2: Materials and Methods
2.1.2. Collection and fixation of embryos
Embryos were collected on apple juice plates for either 2-5 hours and aged to
reach the desired stage(s), or overnight. The embryos were dechorionated in 100%
bleach for 2-3min, rinsed thoroughly in 0.1% Triton X-100 and water, and transferred
into a vial containing equal amounts of fixing solution (4% paraformaldehyde in PBS)
and heptane. After 20 min. incubation (on rocker), the lower phase (fix) was removed
and replaced by 100% methanol and the vial vortexed for 20 sec. The de-vitellinised
embryos settle to the bottom of the vial and phases containing the heptane and the
vitilline membrane were discarded. The embryos were washed twice with 100%
methanol and stored at -20°C in methanol. These embryos were used for in situ
hybridisation studies or antibody stainings.
2.1.3. Embryo collections used In microarray experiments
Fly cages were set up one night before the egg collections were performed. The
next day two 30 minutes pre-lays were collected on apple juice plates and discarded.
Subsequently embryos were collected for exactly 30 minutes and aged for 4 h at RT.
The embryos were rinsed off the plates with water and transferred into a 1.5ml
Eppendorf tube and placed in a water bath at 36°C for Ih. After the heat shock
treatment the embryos were kept at RT for 30 min. and then shock frozen in liquid N2
and stored at -80°C till further treatment. Alternatively embryos were aged for 5 h and
heat shocked for 30 minutes and immediately shock frozen and stored at -80°C. Non
heat shocked control embryos were collected for 30 minutes and aged for 5^^^h. The
effective staging of the embryos was confirmed by examining a subset of the collected
population under the light microscope (Nikon eclipse E800 microscope). Embryos
staged in the above manner were between developmental stage 10 and 11. Embryos of
64

Chapter 2: Materials and Methods
these stages are characterised by their fully extended germ band (stage 10), and the
formation of tracheal pits, and furrows that subdivide the germ band into metameric
units (stage 11).
2.2. Molecular biology techniques
2.2.1. RNA extraction and purification from Drosophila embryos
Embryos were ground with a pestle on dry ice in the Eppendorf tubes. Total
RNA extraction was performed using the RNeasy Mini® kit (Qiagen) following the
manufacturer’s instructions. RNA was precipitated adding O.Spl (5 mg/ml) glycogen
(GIBCO BRL), 1/10 volume 3M NaOAc (pH 5.2) and 2.5x volume 100% cold ethanol.
The mixture was left on ice for 30 minutes, collected by centrifugation (13000 rpm for
30 min) and washed with 80% ethanol. The supernatant was discarded and the air dried
pellet re-suspended in RNase free water. The integrity of the extracted RNA was
analysed on a 1% agarose gel (GIBCO BRL). The concentration of the RNA was
determined by measuring the absorbance at 260nm in a spectrophotometer
(BioPhotometer, Eppendorf). Approximately 200 embryos are needed to obtain > 5pg
of total RNA. The ratio of the readings at 260nm and 280nm (A260/A280) provides an
estimate of the purity of the RNA preparation. Pure RNA has an A260/A280 ratio of 1.9 -
2.1 in lOmM Tris-Cl, pH 7.5.
2.2.2. Double stranded cDNA synthesis
5 pg of high quality total RNA were used for the synthesis of double stranded
cDNA following the instruction manual of GIBCO BRL Superscript Choice® system.
Double stranded cDNA prepared for microarray experiments were synthesized
following the instructions of the Affymetrix GeneChip® Expression Analysis Manual
65

Chapter 2: Materials and Methods
using a T7-(dT)24 primer to prime first strand cDNA synthesis in place of the oligo (dT)
or random primers provided with the Superscript kit.
T7-(dT)24 primer:
5'- GGCCAGTGAATTGTAATACGACTCACTATAGGGAGGCGG-(dT)24- 3'
2.2.3. Drosophila Genome Arrays
Drosophila Genome Arrays were purchased from Affymetrix. More than 13500
predicted transcripts from the annotation of the Drosophila genome (version 1, August
2000) are represented on the array. This includes over 8000 genes for which
confirming EST or full-length cDNA evidence is available. About 3000 genes were
identified using prediction algorithms . The remaining -2500 have not only a gene
predicted algorithmically, but also show some homology to genes from other organisms.
In addition, several hundred ESTs close to predicted gene sequences, but lying outside
predetermined gene inclusion boundaries, are also included on the array. The final
microarray analysis presented in this thesis was carried out using updated annotation of
the Drosophila genome (March 2003). The genes are represented in form of single
stranded DNA oligonucleotide probes. Each probe type is located in a specific area on
the probe array (or microarray) referred to as a probe cell and consists of 25 nucleotides
originating from the 3' region of each gene. Every gene is represented in a probe set,
containing 14 different probe pairs. A probe pair is made of two probe cells, a Perfect
Match (PM) probe cell and its corresponding Mismatch (MM) probe cell. The MM
probe cell is designed to be complementary to the reference sequence except for a
homomeric base mismatch at the central (13*) position and serves as a control for cross
hybridisation.
All microarray experiments were performed following the instructions of the
Affymetrix GeneChip® Expression Analysis Manual (Affymetrix 2000). The
experiments were replicated three times, starting with a new embryo collection and
66

Chapter 2: Materials and Methods
were therefore biological replicates. The major steps of the Genechip expression
analysis are outlined below. Steps 2-5 were done in the Institute of Child Health in
Mike Hubanks laboratory in London. The bioinformatic analysis was done in
collaboration with Simon Tomlinson and Gavin Kelly at Cancer Research UK.
1) Target preparation
Biotin-labeled RNA fragments referred to as the cRNA target were generated
following the instructions of the Bio Array High Yield RNA Transcript Labeling Kit
(Enzo). The anti-sense cRNA is purified using the RNeasy kit (Qiagen), quantified and
qualified using a spectrophotmeter (Eppendorf). 20pg of cRNA are fragmented by heat
treatment at 94°C for 35 minutes in a 5x RNA Fragmentation Buffer (200mM Tris-
acetate, pH 8,1, 500mM KG Ac, 150mM MgOAc). This procedure produce a
distribution of cRNA fragments sizes from approximately 35 to 200 bases.
2) Target hybridisation
The fragmented biotin-labeled cRNA is added to a hybridisation cocktail
(GeneChip® Eukaryotic Hybridization Control Kit, Affymetrix), which contains
different probe array controls as well as BSA (Gibco BRL) and herring sperm DNA
(Promega). This is then hybridised to the oligonucleotide probes on the microarray
during 16-hour incubation at 45°C under rotation at 60rpm (GeneChip Hybridization
Oven, Affymetrix). Each target cRNA is hybridised to one microarray.
3) Probe Array Washing and Staining
The hybridised probe array is stained with a streptavidin phycoerythrin
conjugate (SAPE) and undergoes an automated washing and staining protocol (EukGE-
WS2v4) using the GeneChip Fluidics Station 400 from Affymetrix.
67

Chapter 2: Materials and Methods
4) Probe Array Scan
Each probe array is scanned 2 times by the Hewlett-Packard Gene Array^^
Scanner at the excitation wavelength of 488 nm. The amount of light emitted at 570nm
is proportional to the bound target cRNA at each location on the probe array. The
scanner acquires an image of each of the probe cell. The Affymetrix® Microarray Suite
5 software automatically overlaps the 2 scanned images and averages the intensity of
each probe cell.
5) Data Analysis
Initially an absolute analysis is carried out using the probe array's hybridisation
intensities measured by the scanner. The Signal Intensity is calculated by taking the
difference between every Perfect Match probe cell and its control Mismatch probe cell
of every probe pair and averaging the differences over the entire probe set. This Signal
Intensity serves a relative indicator of the level of expression of the transcript. To
calculate the background, which is a measurement of the Signal Intensity caused by
autofluorescence of the array surface and nonspecific binding of target or stain
molecules (SAPE), the array is divided into 16 sectors. The software ranks probe cells
by fluorescence intensity and identifies the lowest 2% for each sector and calculates
their avarage. The resulting value is the sector's background, which is than subtracted
from the average intensities of all probe cells within that sector. The output of any
experiment is multiplied by a Scaling Factor to make its Average Intensity equal to an
arbitrary Target Intensity (TOT = 300), set by the user. The average intensity of an array
is calculated by averaging all the Signal Intensity values of every probe set on the array,
excluding the highest 2% and lowest 2% of the values. Scaling allows a number of
experiments to become normalised to one Target Intensity, allowing comparison
between multiple experiments.
68

Chapter 2: Materials and Methods
6) Statistical analysis
For analysis within Genespring (Silicon Genetics®) the data was normalised so
that each chip's data was centred around the median expression value. This approach
makes the assumption that the expression measurements are symmetrically scattered
about the median. This assumption could be inappropriate if a severe treatment would
lead to an assymetric shift of the expression measurements, for example due to general
repression of gene expression. However this was not the case in our experiments. An
alternative approach to normalisation would be to scale the data to a set of non
differential expressed genes (housekeeping genes). The normalisation then critically
depends upon the selection of the housekeeping genes. This can be a difficult task
compounded by the non-independence of the expression patterns of these genes.
Differentially expressed genes were identified using a Welch t-test and applying
multiple testing correction to adjust the individual p-value. Correction for multiple
testing should be considered when significance tests are applied multiple times. When
testing the statistical significance of group comparisons for many genes, relying on the
nominal p-value, many genes will pass the filter by chance alone. The expected number
of genes that are positive by chance (p<=0.05) is the product of the a level for a single
test multiplied by the number of times the test is applied. For instance, when 10,000
genes are tested for reliable changes between groups at significance a level 0.05, then
assuming the tests are independent, the expected number of genes that are significant is
a multiplied by the number of genes 0.05 *10000=500. Thus, about 500 genes will be
misidentified as significant, even when there is no real difference in gene expression.
This means that from 1,000 genes showing significant behavior by this approach, half of
the genes on the list will have appeared by chance. The Benjamini and Hochberg test
was used to control False Discovery Rate (FDR), which is defined as the expected
69

Chapter 2: Materials and Methods
proportion of truly non-differential genes in the differential gene list. This allows lists of
genes to be selected based on a given false discovery rate. For example, if a false
discovery rate of 0.5 is used to select differential genes then this indicates that 50% of
the genes selected from a given experiment are expected to occur erroneously. The
Genespring Global Error Model was used when selecting differentially expressed genes.
The Global Error Model performs a variance components analysis, using measurement
variation and sample-to-sample variation to accurately estimate standard errors and
compare mean expression levels between experimental conditions.
2.2.4. Polymerase chain reaction (PCR)
Genes which were identified from the microarray analysis were isolated by RT-
PCR and cloned in order to generate an anti-sense RNA probe for in situ hybridisation
studies. All primers were synthesized by the Cancer Research UK oligo services and
were either pre-purified or ethanol precipitated as suggested. Stock solutions of 1 mg/ml
were prepared in TE Buffer (lOmM Tris-HCl, ImM EDTA, pH 8.0) and stored at -
80°C. PCR was performed using a Primus 96plus PCR machine from MWG-
BIOTECH. Taq polymerase (in house) and buffers from Stratagene or the Tag PCR
Master Mix kit from Qiagen were used to amplify DNA 2fragments. Approximately 50-
lOOng of double stranded cDNA was generally used as a template, primers at 125 ng
and dNTPs (Roche) at a final concentration of 2.5mM each per reaction. Reaction
components were mixed on ice before dénaturation at 94°C for 3 min followed by 30
cycles of dénaturation at 94°C for 30 sec, annealing at 55-60°C (depending on the
primer melting temperature) for 30 sec and elongation at 72°C for 1-3 min (depending
on the length of the resulting DNA fragment). The PCR was finished with a final
extension step at 72°C for 10 min.
70

Chapter 2: Materials and Methods
The following primers were used to amplify DNA fragments to generate probes
for in situ hybridisation studies. All primers were designed to be to the predicted cDNA
using Primer 3 and Amplify-2.5|3r (Software for PCR, University of Wisconsin) to
eliminate primer-dimer formation and to optimize annealing temperature.
Table 2. Primer list
GTACTTCAATCCCATGGCAGC atonalFCCGTGCCGTGTAATTATCGTC atonal RCGTCGCTCCAGAGAAAAGTT gcm_FATTGGTGAGATGCGGAAAAG gcm_RCTGTTTCGGCTCTCAACATGAC gsb_FCAAAATCCAAAGTATGGATAGGTTGG gsb_RGCCACAGTTCCAGTTCCAGT hh FTGGGTTTTTGATTTGTGGTG hh_RCAGCAGTCTCAGCCAAGTCGAGG wg_FCTCCTCGCCGTAGACCAGACCAG wg_RACGTGACCCCAGTCAAGAAC sry-alpha_FTCAGCAAGTGAGTCCTGTGG sry-alpha_RATGTCCAAAATGAAAATGCTCC yan_FTTAGCTCAGGTAATGTTGTGTGTGT yan_RGGTGCTGTACCTCACCAACA yin_FCAATGCCGTTTGTTTTCCTT yin_RGCTCCAGCGCAATATATCATTG l(3)mbt_FCGACCTTCTCTGGAGATGTAACAAC l(3)mbt_RCGCGGCGAGGTCAATATCTC lola FCGGCTGTCGAGGATGTTAGTTTAG lola_RCTGGATGATAGTGATCCAAACTGG StamFGCTGCTGCTGGTAGATAGGAATG Stam RGAAACTGTTCGTATTCCTGGCCTTG Ser99Db FGTTGTCGCGGATCCAGTCCAG Ser99Db_RACGCAGTTCCTGCTGCAGAAC pdm2_FACTGATGGAACTGCCGCAGAAG pdm2_RCTCCAGAACGGAATAGCGCC u s h FGTTCTTCTGCTGGCCGTGTC ush_RGTGCTCGCTGTGCTATGCTA a s pFAGCTTGCACAAAATCGTATCG a s p RCATTGACTTCCGCGAGAACC uzipFGAGACGCTTCTCGTAGCTGAAGAG u z i pRATGAAGTGCCAACTGCTTTTCG bnb_FCTCCTTCGATTCGCTGCTCTC bnb_RCATGGAGCGACGTGGTCTCTAC mdr49_FCGACACCTGTTGCATCAGATAGAG mdr49 R
71

Chapter 2: Materials and Methods
CTGACGGCTCGACTCGATTAGAC MLC2 FGGATGTGATCGGATGTGCCTTAG MLC2_RCAGCCAACTATATGCAGGGCAG neur FCGCGACCTTTCTACGTGGTG neur RTGTTTTTGTGACATAACAGCTGTTC Z600 FTTAGAAGTTCAAAGATTTGCGAGC Z600_RGAATTGCATTTAGCTAACAATTGG myotubularin_F
GCATCCGAGTGCTTGTTGTC myotubularin_RCGATATGGATATCGGACCCTTAG nAcRalpha-8 OB_FGTGTGCATAGTGCAGTTTATGTGC nAcRalpha- 8 OB_RCTGGGACGCAGGTTCTTATTG echinoid_FCTAACGCCCTGAGTATGCCG echinoid RGCATTAGTCTTAACGCTGCTGC fc96Cb FGTGCTACAGCTAGACTTCACAGACC fc96Cb_RCCTGACCATCTCCATCCTAGGAAAG beaten p a t h RCAAATTGTGCGCAGCGAGAG beaten p a t h FGCGTTATTAAGGACGCGAAAATG huntingtin_FCCTGCTTGTCCTCATCCGAG huntingtin_RCTCGACCAGACAGAAATCAGC Lcp2_FGAACAGTCCGTGGAGTGGTC Lcp2_RTGGTTTGAGCTATCCATCGTT Lcp65AG2q FGTTCTTATCGGCGATGTAGTTG Lcp65AG2q RGAGATCACAGGCCGGATTTA mew_FCCCTTATTGCAGTCCAGCTC mew RTATCCGGGATGTAGCAAAGC ci_FCGTTGATGAGCTGACGGTAA ci_RGAGCGATCGATCTCCAAGTC bap_FAGACATGGTCACTCCCAACC b a p RGCATTCGCCAAATTCGATAC dve_FTTTCGTTGTCCTCCTCATCC dve_RTGGCCTACACACAGAAGACG patchedFCTGCTCCCAGAAGATGAAGG patchedRTGTGTCTGTGTTTCCCAAGC dropFTTCTGCTTGCTTGTTTGCTC d ro pRGTCGAGCGTGAAAGAACCTC dorsocrossFCTACCCGTTGAGTCCTGCTC dorsocrossRTCCTTCATTTCGAACCCATC m-spondin_FGCAGGACGAATGACTCCAAT m-spondin_RTGCCATCTCTAACAGCAACG arm adilloFGGAAGAGGGCTATTGTGCAG armadillo_RTCTAATTTGCTGGCCGAAAG bloated tubulesFCAGGCACTCGAATCCTTGAT bloated tubules_RCAGCATTACCAGCTGACCA lethal of scute_FATAGCCACCTCCGGAAATCT lethal of scute_RGCCAAAAGCAAAACCAAAAA knockout_FACCTTGCCGCTGTCAATACT knockout RCACCATCAACTGTCACTTCGAC CGI 1659 FGCCATTCGATTGGTCTTTCC CGI 1659 RTGGCTACATACGCAACCAGA CG6900 FAAAGGAGCAGTTTGGTAAGCA CG6900 R
72

Chapter 2: Materials and Methods
GGTTCTAATGGCCCTACCGCC CG12199_FCGC ATCGAGTACTGCTGACC AG CG 12199_RCATGGAAGATCCTCGCATTTGC CG5704_FCACAACTTGCTGCGCTTTTTG CG5704 _RAGAAATCCAGTCGCAAGTCG CG14112_FGTCTTCTTGCCGTTGAGGAT CG14112_RGCAAAGCTTGCTCATCTCTG CG3045_FCTGATATTGACGCGGATTGA CG3045_RCAAAAAGCAGACACCAACCA CG6770_FCAAACTTCATTATTTGAAACGCTCT CG6770_RATCCCATCGACAACAAGGAC CGI 1212_FGTAGGCGTAGCTGGTCAAGG CGI 1212_RGGGAAACCATTCTCGTTTGT CG6709_FCGCCAGCATCGATCATTATT CG6709_RTTCCACCACCAGTGGTTTTC CG7271_FATGCTGTCCACGGAGAAGTC CG7271_RCGCAAAGTAAATCAGATCACAGAG CG5784 FCCTCTGACTTGCGACTCGTC CG5784_RGAGTC AAGCTC AGAATTC AACC AC CG 16844_FGTTTTGTTTGGAATATATTTAATATGGGC CGI 6844_R CAATTAGGCCAACACTGCTTGG CG14014_FCCGTGTAGACGAACTCCTCGTG CG 14014_RGAACCTGAGCGACCTACAGG CG8349_FGTGCACGAGGAAGTCACTCC CG8349_RCTGAGGCAAACTAGTCGGCTG CG13532_FGGAACTGTCGCAATTGGAGC CG13532_RCCGCTTGAAACCTC AC ACTTC CG 13 000_FGAAAGCGCATCTGTGAGTGG CGI 3000_RCAGCGACGAATGGCAAACAG CG6256_FCTCGATTGATGAGCATGGAGC CG6256_RCAAGTGAAGAGCGAGTAGAGTGATG CGI 3213_FGAGTACACCGTGTGATGTGGC CGI 3213_RGCCCAGGAAGTCCATCAACC CG7290_FCGTCGAAGTTCCATCGCTTTG CG7290_RGAAGTTAGCTGCGCTGCAATG CG3775_FCTTGAGGCGATAGTATGCATTCAC CG3775_RGCCCAGGAAGTCCATCAACC CG7290_FCGTCGAAGTTCCATCGCTTTG CG7290_RGAAGTTAGCTGCGCTGCAATG CG3775_FCTTGAGGCGATAGTATGCATTCAC CG3775_RC AAGCTTATGCTGCGGTAGTAAC CG9109_FGTTTGCTGCAGC AATCCAAC CG9109_RCCCAGCGATCGCACAGATAG CG7294_FCTAGATAATCCGCTCTCTGGCAGC CG7294_RGCGCACTCTGACATATGTGC CGI 1152_FGAGGCGGCGTAATAGAGGAAG CGI 1152_RGGCGCACAAGTTCATCATAA CGI 5444_FACGAACAGGACCACGATTTC CGI 5444_RGGGAAACCATTCTCGTTTGT CG6709_FCGCCAGCATCGATCATTATT CG6709 R
73

Chapter 2: Materials and Methods
TTCCACCACCAGTGGTTTTCATGCTGTCCACGGAGAAGTCGGCCAGTGTTTTCGATGAGTGATTTGCAAGGACCTTCGAGAGCGCTACAATCGCGTCTATTGCTTGGCCAGTTGGATAATCCTCCGATCAGACCGTTTTACAGACTCAGCCTCATCCACAAAAAATGGCCATCCATTCAGGCTCCCAGTCCAAGATGAAGACACCACGGAGGAGTACCTGGGCCCAGACTTCAGGATGTACGACTTCTATGCCTGCCAGTGTGCAGTCGTAAGACGCTGAGCCGGCCATTCTTCTCATTAAGGAAGGAGCAGCAAAGGATCAGACTGGAGTGGGTTGCTTCCATTTTTGCTTGTGCCATCAAGCTACCAGGACATCAGCGCTCCATTGATTATTCATGAATACCAACCCATCGCCAGCATGGAGATACGTTGCGAATAATTCCAGGAATAATGCAACAAGCGTGGATCAGTTAGCGTGATGTCACTGTCGGCTTGTAG
CG7271_FCG7271_RCG7204_FCG7204_RCG5272_FCG5272_RCG14521_FCG14521_RCG9555_FCG9555_RCG2191_FCG2191_RCG7876_FCG7876_RCG7920_FCG7920_RCG8089_FCG8089_RCG17778_FCG17778_RCG10362_FCG10362_RCG17084_FCGI 7084 R
2.2.5. Cloning
PCR products were cloned using the vector pBluescript II SK(+/-) or TOPO TA cloning
kits from Invitrogen according to the instruction manuals. Ligations were either
transformed into Invitrogen One Shot® Chemically Competent TOPIC cells according
to the manufacturers instructions or TOPIC cells that had been made competent using
the Calcium Chloride method described in Short Protocols in Molecular Biology
(Ausubel et aL, 1989). Transformed bacteria were plated on LB agar plates containing
the correct antibiotics and incubated at 37°C for 16-24 hours. All clones were analysed
by preparation of DNA and restriction enzyme digests or verified by sequencing. The
cDNA s for the genes race, zen, dpp, u-shaped and pannier were obtained from Amo
Müller. The full length araucan cDNA was obtained from Juan Modolell.
74

Chapter 2: Materials and Methods
2.2.6. DNA preparation, purification and analysis
DNA was prepared from bacterial cultures grown overnight in LB medium using
the Qiagen Mini prep kit according to the instruction manuals. Digested DNA were
purified either by using the QIAquick Gel Purification or PCR Purification kits
(Qiagen) according to manufacturer’s instructions. Alternatively, DNA was purified by
phenol : chlorofrom extraction and ethanol precipitation. Briefly, an equal volume of
phenol : chlorofrom was added to the DNA sample, mixed and centrifuged to separate
the phases. To the aqueous phase was added 1/10 volume 3M NaOAc (pH 5.2) and 2.5x
volume 100% cold ethanol. DNA was precipitated at -20°C for 30 min, collected by
centrifugation (13000 rpm, 20-30 min), washed with 70% ethanol and re-suspended in
TE buffer. Gel electrophoresis was carried out using 1-1.5% agarose gels in TAB.
2.2.7. Sequencing
DNA sequencing was carried out by the Cancer Research UK in house facilities
using the ABI PRISM Dye terminator cycle sequencing kit. Each reaction contained 8pl
of the Dye Terminator Ready Reaction Mix (Perkin Elmer), 500ng-lpg of plasmid
DNA template and lOpmol of primer. PCR cycle sequencing and purification of the
products was performed as described in the ABI Cycle Sequencing Kit instructions. The
reactions were loaded on an automated sequencing machine (ABI PRISM 377, Perkin
Elmer) and analysed using ABI PRISM software (Sequencing Analysis V 3.0).
The sequencing primers used were:
Table 3. Sequencing Primer
Primer: Sequence: Vectors:
M13Fwd GTAAAACGACGGCCAGT pCR*II-TOPO, pBluescript II SK(+/-)
M13Rev CAGGAAACAGCTATGAC pCR®II-TOPO, pBluescript II SK(+/-)
75

Chapter 2: Materials and Methods
2.3. in situ hybridisation
2.3.1. Generation of anti-sense RNA probes
Digoxigenin-labeled or fluorescein-labeled anti-sense probes were made from
plasmid DNA, linearized with the appropriate restriction enzymes. The template was
purified using the PCR purification kit (Qiagen) or by phenol:chloroform extraction.
The in vitro transcription reaction was carried out as described by the company with
SP6, T3 or T7 RNA polymerase (DIG RNA Labeling Kit, Roche). The probe was than
purified with the RNeasy kit (Qiagen) in combination with an on-column DNasel
digestion using the RNase-free DNase Set (Qiagen) to remove the DNA template. An
aliquot of the RNA probe was analysed by agarose gel electrophoresis. The probes were
stored at -20°C. The full length RNA antisense probe used in this study for KrUppel and
sloppy paired 1 were obtained from Simon Bullock and Kirsten Hardiman, respectively.
The mirror probe does not contain the homeodomain and was generated by Helen
McNeill.
2.3.2. Pre-hybridisation and hybridisation
Embryos were re-hydrated through serial washings in 95%, 70%, 50% and 30%
methanol in PBS, followed by 20 min post-fixation in 4% paraformaldhyde in PBS.
Next, the embryos were washed five times in PTW (PBS + 0.1% Tween), then rinsed
for 5 min in equal volumes of PTW and hybridization buffer (50% deionised
formamide, 20x SSC, 100 pg/ml tRNA, 50 pg/ml Heparin, 0.1% Tween in ddHiO).
After that the embryos were pre-hybridised at 70°C for at least one hour in the
hybridization buffer. The anti-sense RNA probe is denatured through heating at 80°C
for 2 min and immediately put on ice. The pre-hybridisation buffer was replaced with
pre-warmed hybridisation buffer and 0.5 ng/pl of the digoxigenin-labeled probe (usually
Ipl) were added. Embryos and probe were mixed and incubated at 70°C overnight.
76

Chapter 2: Materials and Methods
2.3.3. Signal detection for single in situ hybridisation
After removal of the probe, the embryos were washed in pre-warmed
hybridization buffer at 70°C for 20 min, followed by a 20 min washing step in pre-
warmed PTW and hybridization buffer (1:1). Subsequently five washes in PTW at RT
were carried out, each for 20 minutes. Once the embryos were treated for 30 min at RT
in PTW + 1% heat inactivated goat serum (Vector) they were incubated for 1 hour in
PTW +1% heat inactivated goat serum with a 1:2000 dilution of alkaline phosphatase-
conjugated anti-digoxigenin Fab fragments (anti-DIG, Roche). After washing 4 x 20
min. at RT or overnight at 4°C in PTW, the embryos were rinsed 2 x 3 min. in AP
buffer (100 mM Tris, pH 9.5, 100 mM NaCl, 50mM MgCli, 0.1% Tween). The alkaline
phosphatase was detected using NBT/BCIP solution (Roche, 4pl in 1ml AP buffer).
Development of the dark blue diformazan/indigo precipitate was monitored under a
dissecting microscope and the reactions stopped by washing the embryos several times
in PTW. Embryos were incubated in 50% glycerol, allowed to settle down and mounted
on slides in 80% glycerol. Pictures of mounted embryos were taken using a Nikon
DXM1200 digital camera attached to a Nikon eclipse B800 microscope.
2.3.4. Double in situ hbridisation with FAST RED and NBT/BCIP
In a double in situ hybridization experiment the dig-labeled and fluorescein-
labeled anti-sense RNA probes were hybridised simultaneously at 70°C overnight.
Washing and blocking were performed as described for the single in situ hybridisation.
Following the block, the embryos were incubated with a 1:2000 dilution of anti-
fluorescein Fab fragments for 1 hour at RT, washed 4 x 20 min or overnight in PTW
and rinsed twice in lOOmM Tris pH 8.0. A FAST RED tablet (Roche) was dissolved in
2ml of lOOmM Tris pH 8.0 and filtered through a syringe driven filter unit (45pm,
Millipore). The embryos were washed once and then incubated in this solution in the
dark. Staining was allowed to develop for 1-3 hours before the embryos were washed
77

Chapter 2: Materials and Methods
once in lOOmM Tris pH 8.0 followed by three washes 5 min each in PTW. The first
antibody was then heat inactivated by incubating the embryos at 70°C for 15 min.
Afterwards the embryos were rinsed once in PTW and blocked in PTW + 1% heat
inactivated goat serum followed by one hour incubation with the anti-DIG antibody.
Washing, staining and mounting of the embryos were done like described for the single
in situ hybridisation. In my experience it does not seem to make a difference which of
the two antibody staining is carried out first. However, FAST RED staining was only
visible for highly expressed transcripts.
2.4. Bio- and Immunohistochemistry techniques
2.4.1. Staining of Embryos
Collection and fixation of embryos was as described above and stored at -20°C
methanol. To re-hydrate the embryos, the entire methanol was removed and replaced
directly with PBTX (PBS, 0.1% BSA, 0.1% Triton X-100) for 1-2 hours. The embryos
were then blocked in PBTX + 2% normal goat serum (Vector). Primary antibodies were
diluted in blocking solution and embryos incubated overnight at 4°C. Following three
washes of 20 min each in PBTX, the embryos were blocked again for 30 min.
Secondary antibodies were diluted in blocking solution and embryos incubated for 2
hours at room temperature. The embryos were then washed 3 x 20 min in PBTX and if a
biotinylated secondary antibodies had been used, incubated with labeled streptavidin for
1 hour at room temperature. Embryos stained with fluorescent antibodies were mounted
in Slow Fade reagents (Slow Fade® Antifade kit. Molecular Probes). DAB staining was
performed following the instructions of the ABC kit (Vectorshield) together with the
3.3-diaminobenzidine tablet set from Sigma. Embryos were analysed and images
78

Chapter 2: Materials and Methods
captured using a Nikon DXM1200 digital camera attached to a Nikon eclipse E800
microscope.
Primary Antibodies used for immunohistochemistry:
Table 4. Primary Antibody listAntibody Dilution used Species Sourcea-PGal 1:2000 Rabbit ICN/Cappel
a-PGal 1:1000 Mouse Promega
a-Castor 1:2000 Rabbit Ward Odenwald
a-engrailed 1.20 Mouse DHSB
a-gooseberry 1:4 Rat Yu Zhang
a-Mirror 1:500 Rat Helen McNeill
a-Mirror 1:2000 Rabbit Helen McNeill
a-Krüppel 1:500 Guinea pig Yasushi Hiromi
a-Repo 1:500 Rabbit DHSB
a-tubulin 1:500 Mouse Sigma
a-Eagle 1:500 Rabbit Joachim Urban
BP102 1:100 Mouse DHSB
22C10 1:100 Mouse DHSB
79

Chapter 2: Materials and Methods
Secondary antibodies used for fluorescent labelling
Table 5. Secondarv Antibodv list
and DAB staining:
Antibody Dilution Source
Alexa Fluor 594 conjugated a-Gp IgG 1:500 Molecular probes
Alexa Fluor 488 conjugated a-Gp IgG 1:500 Molecular probes
Alexa Fluor 488 conjugated oc-Ms IgG 1:500 Molecular probes
Alexa Fluor 546 conjugated a-Rb IgG 1:500 Molecular probes
Alexa Fluor 488 conjugated a-Rb IgG 1:500 Molecular probes
Alexa Fluor 488 Streptavidin conjugate 1:500 Molecular probes
Biotinylated a-Ms 1:300 Vector
Cy3 conjugated a-Ms Ig 1:500 Jackson ImmunoResearch Labs
Abbreviations: Ms: mouse, Rb: rabbit, Gp, Guinea pig
2.4.2. Protein Extraction
Staged embryos were ground with a pestle in an Eppendorf tube in lOOpl of
protein grinding buffer (8M Urea, 0.2% Triton, 0.2% Nonldet P40, 1 Protease inhibitor
cocktail tablet (Complete, Mini EDTA free) from Roche (in 10ml PBS). Samples were
centrifuged for 1 min at 14000g and the supernatant transferred into a new tube and
stored at -80°C.
2.4.3. SDS PAGE
Polyacrylamide gel electrophoresis was performed using the Bio-Rad Protean
systems II and III. Resolving gels were made to contain 7.5-12% polyacrylamide
(37.5:1 acrylamide/Bis, Bio-Rad) in 375mM Tris pH 8.8, 0.1% SDS and stacking gels
80

Chapter 2: Materials and Methods
contained 4% polyacrylamide in 125mM Tris pH6.8, 0.1% SDS. Quantified protein
samples (Bio-Rad protein Assay) were mixed with 4X sample buffer (125mM Tris pH
6.8, 20% glycerol, 4% SDS, 10% P-mercaptoethanol, 0.008% Bromophenol Blue) and
boiled for ~ 5min. before loading with appropriate molecular weight markers. Gels were
run in SDS-PAGE running buffer (25mM Tris pH8.3, 192mM Glycine, 0.1% SDS).
Electrophoresis conditions were 200V constant voltage for small gels and 50mA
(stacking gel) and 20-70mA (resolving gels) constant current for large gels.
2.4.4. Western Blots
Western blots were performed using the Semi Dry Electrophoretic Transfer Cell,
from Bio-Rad, according to the instruction manual. Polyacrylamide gels were
equilibrated in transfer buffer (48mM Tris pH8.3, 192mM Glycine, 20% methanol,
0.05% SDS) and proteins transferred onto ECL Hybond nitrocellulose membranes
(Amersham Pharmacia) at 1OV for 60-90 min. using the Semi Dry system. Membranes
were blocked in PBST (PBS, 0.1% Tween 20) with 5% milk for 1 hour at room
temperature or overnight at 8°C. Primary antibodies were diluted in PBST, 0.2% milk
and incubated with the membrane for 1-2 hours at room temperature. Following two
rinses and 3x 10 min washes in PBST, the membrane was incubated with a horseradish
peroxidase conjugated secondary antibody (diluted in PBST, 0.2% milk) for 1 hour. The
membrane was washed as before and the HRP detected using the ECL Western Blotting
chemiluminescent detection agent (Amersham Pharmacia) according to the
manufactures instructions.
81

Chapter 2: Materials and Methods
Table 6. Antibodies used for detection of Western blots
Antibody: Dilutions: Source:
a-Mirror, 1:1000-1:2000 Helen McNeill,
rabbit polyclonal antigen = GST-Mirror C-term fusion
a-rabbit Ig horseradish 1:5000 Amersham Lifescience
peroxidase conjugate
82

Chapter 2: Materials and Methods
83

Chapter 3: Mirror
CHAPTER 3
Mirror
3.1. Mirror expression in the embryo
3.1.1. Mirror expression from stage 10 -14
Before the presentation of candidate genes obtained from the microarray
analysis, a more detailed description of mirror expression in the embryo during
development is relevant to this study. Even though the major characteristics of the
expression pattern have been reported (McNeill et al., 1997) and are summarised in the
introduction (see page 50), some previously unnoticed aspects were revealed by
performing Mirror antibody stainings in the embryo.
At the beginning of stage 10 Mirror expression is localised in a segmental
pattern at the anterior border of the segment in a transient one cell wide stripe per
hemisegment. These stripes do not surround the entire embryo, as do most segment
polarity genes. The expression pattern quickly expands in the neuroectoderm (fig.
3.1. A) and the epidermal stripes seem to grow fainter (fig.B.l.B). At the onset of germ-
hand retraction at stage 11, Mirror expression becomes further restricted in the
neuroectoderm (fig.S.l.C). During germ-hand retraction at stage 12, Mirror expression
is found in the delaminating neuroblasts, the ventral midline and in the head. (fig.B.l.D)
At this stage the lateral Mirror expressing cells form segmentally repeated more or less
round clusters (fig.S.l.E).
84

Chapter 3: Mirror
Figure 3.1. Mirror antibody staining in wild type embryos from stage 10-14. A) Mirror protein is
localised in a segmental pattern at the anterior border o f the segment at stage 10. B) Same embryo as
in A but slightly turned to the side to demonstrate that the Mirror expressing stripes do not surround
the entire embryo. Mirror is also expressed in lateral cells in the epidermis and at the site o f the
stomodeal invagination (arrow). C) At the onset o f germ-band retraction at stage 11, the striped
expression is lost and Mirror expression becomes restricted ventrally. D) At stage 12, Mirror
expression is found in the delaminating neuroblasts, the ventral midline and in the head. E) Same
embryo as in D turned to the side. Mirror expression persist in the lateral epidermis in more or less
round patches o f cells (see dotted line). F) Mirror expression continues in several cells o f the forming
CNS and the ventral midline at stage 14. G) Lateral view o f the embryo shown in E. Mirror protein is
found in a cluster o f lateral cells giving rise to the PNS. Panels are ventral view o f the embryo, except
B+E+G and anterior is to the right.
85

Chapter 3: Mirror
At stage 14 Mirror expression continues in several neuroblasts of the forming
CNS and the ventral midline (fig.S.l.F), while Mirror protein persist in cluster of lateral
cells giving rise to the PNS (fig.3.1.G). In the lateral epidermis the expression pattern of
Mirror overlaps with that of Araucan and Caupolican (Gomez-Skarmeta et al., 1996;
McNeill et al., 1997). The expression pattern of the latter two has been reported to be
identical (Gomez-Skarmeta et al., 1996). Figure 3.2. shows the in situ hybridisation
expression pattern for araucan. Similar to mirror, araucan is expressed in clusters of
cells in the lateral epidermis from stage 11 to 14 (fig.3.2.A+C) and is strongly expressed
in the proventriculus (frg.3.2. C+D). araucan is not detected in the ventral nerve cord
(flg.3.2.B+D). Mirror is the only Drosophila Iroquois Complex member which is
expressed in the developing ventral nerve cord (McNeill et al., 1997: Juan Modollel,
personal communication).
3.1.2. Mirror expression in various other stages of development
Less well documented is Mirror expression in various other stages of
development. In addition to its expression in anterior and dorsal cells at the cellular
blastoderm stage, mirror is also weakly expressed in three patches of cells at the ventral
side of the embryo (fig.3.3.A). At the onset of germ band retraction mirror is expressed
in a narrow mid-dorsal partition of the blastoderm, which will give rise to the
amnioserosa and in an anterior dorsal region of the procephalon (fig.3.3.B). Anti-p-Gal
staining of a mirror-lacZ embryo demonstrates the expression in the lateral cells of the
epidermis at stage 10 (fig.3.3.C). Shortly after, at stage 11, Mirror expression is
restricted to clusters of cells originating from the earlier broader, potentially
mesodermal, expression domain (fig.3.3.D).
86

4,
B D ^ JP; V'Vfr
Figure 3.2. /w .sv/m hybridisation pattern of araucan in the embryo. A) araucan is expressed in cells o f the
lateral epidermis at stage 11 B) The same embryo as in A but shown from the ventral side araucan is not
expressed in the developing CNS. C) At stage \ A araucan is expressed in cells o f the lateral epidermis. D)
Ventral view o f the embryo shown in C. araucan transcript is not detected in the ventral nerve cord o f the
embryo. Strong expression is detected in the proventriculus (arrow). Anterior is to right in all panels.
87

Chapter 3: Mirror
This periodically arranged expression resembles the expression pattern of the
gene bagpipe in the trunk visceral mesoderm promordia (Azpiazu and Frasch, 1993) or
the gene gleeful in the mesoderm (Furlong et ah, 2001). bagpipe is required for visceral
mesoderm formation and the promotion of somatic muscle and heart formation (Lee and
Frasch, 2000). gleeful is believed to play a role in somatic muscle development (Furlong
et ah, 2001). Mirror protein is found in foregut structures such as the esophagus (es), the
proventriculus (pv) and the pharynx (ph) (fig.3.3.E+F). Moreover, at stage 15, Mirror is
expressed in four cells in the posterior spiracles in a non-overlapping pattern with
Engrailed expressing cells (fig.3.3.G+H).
88

( 'haplcr 3: Mirror
B
Figure 3.3. Further characterisation of Mirror expression in the embryo. A+B) in situ hybridisation using
a mirror riboprobe. A) At the cellular blastoderm (stage 5), mirror is expressed in dorsal and anterior
patches but also in three patches on the ventral side o f the embryo B) At stage 8, mirror is expressed in
the amnioserosa and in cells o f the procephalon (arrow). C) The lateral view o f a mirror-lacZ embryo at
stage 10 stained with (3-gal antibody (green) reflects Mirror expression in cells o f the lateral epidermis in
an initially connected stripe. D) Mirror antibody staining (green) in a wild type embryo o f stage 11.
Mirror expression becomes restricted to clusters o f cells within each segment in the lateral epidermis. E)
Lateral view and F) ventral view o f the head region o f the embryo shown in (C). Mirror is expressed in
foregut structures such as the esophagus (es), the proventriculus (pv) and the pharynx (ph). G) Costaining
with anti-Mirror (green) and anti-Engrailed (red) in a stage 15 embryo showing a dorsal view on the
posterior spiracles. Mirror and Engrailed are expressed in mutually exclusive cells G) Higher
magnification o f the Mirror expressing cells in the posterior spiracles. Anterior is to the right in all panels.
89

Chapter 3: Mirror
3.2. Analysis of Mirror over-expressing embryos
3.2.1. P-tubulin-\/PASx\JAS-mirror embryos
P-tubulin-VP 16-Ga\A females crossed to UAS-Mirror males give rise to a
progeny which strongly over-expresses Mirror. Misexpression of Mirror in these
embryos is ubiquitous and starts during the cellularisation of the blastoderm at stage 5
and remains active till stage 13-14 of development (fig.3.4, see also fig.3.6.B). These
embryos die during development. The onset of Mirror expression in these embryos was
also analysed in western blots. After two 30 minutes pre-lays, embryos were collected
for 30 minutes and staged to different stages of development (see figure 3.5). Mirror
protein becomes first visible 2.5h AEL and appears as a clean double band, possibly due
to phosphorylation. Dephosphorylation experiments performed in the lab using various
Mirror constructs expressed in Drosophila Schneider 2 cells suggested that Mirror is
phosphorylated (Trevor Littlewood and Helen McNeill, unpublished). Indeed, the C-
terminal part of Mirror contains potential phosphorylation sites for the mitogen-
activated protein kinase (MAPK).
3.2.2. Cuticle phenotype of m/rror over-expressing embryos
and its molecular origin
The epidermis of the mature embryo is a segmentally arranged pattern of bands
of naked cuticle and belts with hair-like protrusions called denticles (fig.3.6.A)
(Martinez Arias, 1993). To analyse the effects of misexpression of mirror using the Gal-
4/UAS system (Brand et al., 1994) in the embryo, a P-tubulin-VP 16-Gd\4 enhancer trap
line was crossed to a GAS-mirror responder line. 100% of these embryos exhibit severe
90

Figure 3.4. Ectopic Mirror expression in fi-iubulin-VP16-G2^Ax\]A.^-mirror embryos. A) Embryos o f
this genotype ectopically express Mirror in the entire embryo from cellular blastoderm stage onwards.
Characteristic for this stage o f development are the outstanding pole cells (white arrow) at the posteriez
end of the embryo. B) Ubiquitous Mirror expression is shown in an embryo around stage 10-11. C) This
expression persist in embryos during germ band retraction (stage 12). D) Ectopic Mirror expression is
fading in fi-tubulin -V P]6-G ai4\\]A H -m irror embryos at 14. Still visible is a small part o f the amnioserosa
(dotted line). Dorsal is up and anterior is to the right in all panels.
Ih A E E 2 h A fiE 2 .5 h AEE 3h AEL 3.5h AEL 5h AEL Kh AEL
. . 89 kDa
..6 4 kDa
Figure 3.5. Western blot to detect Mirror protein in V/*76-Gal-4xUAS-m/rro/- embryos. Embryos were
collected for 30min and aged for different time points and total protein were extracted. Mirror protein
becom es first visible 2.5h AEL. A lower second band of Mirror jwotein is also detected. This might be an
unposphorylated form o f Mirror. 300pg o f total protein were loaded into each lane
91

Chapter 3: Mirror
fusions of adjacent segments characterised by the loss of naked cuticle and the joining
together of denticle belts (fig.3.6.B). This phenotype is generally associated with defects
in the establishment of the segmental border and could be due to alterations in the
expression of segment polarity genes. The cuticle phenotype is that of the segment
polarity mutants of the “wingless class” in which the posterior region of every segment,
often including the segment boundary, is deleted and the remaining anterior regions
display mirror image duplications of denticles. Double in situ hybridisation of mirror
and wingless in ^tubulin-VP16-GdXAy3JAS-mirror embryos demonstrates the loss of
wingless expression and illustrates the ubiquitous expression of mirror (compare
fig.3.7.A + B). In contrast, over-expression of araucan using the same Gal-4 driver line
results only to 60% (18/30) fusion of adjacent segments (fig.3.6.D). However, this
might be due to lower levels of produced protein rather than different transcriptional
activity.
3.2.3. Detection of Mirror protein from heat shocked embryos
by western blotting
To optimise the time point, the temperature and the duration of the heat shock
treatment of pKB12-mz>ror and yw embryos, different conditions were tested. Total
proteins were extracted from non heat shocked and heat shocked embryos at different
stages of development and analysed by western blot to determine when Mirror can be
expressed. Mirror has an apparent molecular weight of 83kDa and usually has a
“smeary” appearance on Western blots from total proteins extracted from heat shocked
embryos. Mirror protein first becomes inducible Ih AEL + Ih heat shock at 37°C.
Higher amounts of protein were detected in embryos aged for 2h AEL + Ih hs (fig.3.8.).
The amount of detected protein does not seem to reach higher levels in older embryos.
92

( 'haplcr 3: M irror
f $
Figure 3.6. Cuticle phenotype in Iroquois over-expressing embryos. A) Shows the evenly spaced denticle
belts which are separated by naked cuticle o f two wild type embryos. B) Ubiquitous overexpression of
Mirror (f7^y6-Gal4xUAS-/w/rror) is lethal and results in a wingless class cuticle phenotype with severe
fusions o f denticle belts. The denticle rows seemed to be compressed and shifted towards the posterior
half o f the embryo. C) Embryos ubiquitously over-expressing araucan (VP 16-Gal4xUAS-arawcaw)
exhibit a less severe denticle fusion phenotype
Figure 3.7. Molecular origin o f the “wingless class” cuticle phenotype in ubiquitously mirror over
expressing embryos. A) wingless in situ hybridisation in a stage 11 wild type embryo. Clearly visible is
its segmented expression in 14 stripes. B) Double in situ for mirror (red) and wingless (black) in a j8-
tu b lilin EP/6-Gal4 x \JA^-mirror embryo. The segmented expression o f wingless is almost completely
abolished. Lateral view with anterior to the right.
93

( 'haplcr 3: .\ hrror
3h AEL
non-hs
Oh AEL
+ 1h hs
1h AEL
+ 1h hs
2hAEL
+ 1h hs
3h AEL
+ 1h hs4h AEL
+ 1h hs
•
pKB yw pKB yw pKB yw pKB yw pKB yw pKB yw
. . 89 kDa
■ ■ 64 kDa
Figure 3.8. Western blot to detect Mirror protein in heat shocked Mirror over-expressing embryos.
pKB12-/w/>ror and yw embryos have been collected for Ih and aged from 0-4h prior to a one hour heat
shock at 37C. Mirror protein first becomes inducible Ih AEL + Ih hs at 37°C. 300pg o f total protein
extract were loaded in each lane Note that endogenous Mirror protein is below the detection level in yw
samples. Mirror has an approximately molecular weight o f about 83kDa.
94

Chapter 3: Mirror
However, this could be due to saturation of the detection signal,
90% of heat shocked pKB 12-mirror embryos during development, and about 5%
show a weak cuticle phenotype with fused denticle belts. In contrast more than 50% of
yw embryos subjected to heat shock treatment at 37°C develop normally. Lowering the
temperature of the heat-shock to 36°C resulted in greater than 90% survival for yw
embryos, with only 10% of pKB 12-mirror embryos surviving. Therefore a 36°C heat-
shock protocol was used for the microarray experiments.
95

Chapter 3: Mirror
Discussion
The Drosophila embryo, like the adult fly is segmentally organised.
Segmentation is a morphological process, that is established by the interplay of various
genes, commonly referred to as “segmentation genes”, which act in a strictly
hierarchically controlled manner. Initially the gap genes define broad contiguous sets of
segments, followed by the primary and secondary pair-rule genes which act in
alternating segments, and last but not least the segment polarity genes which have single
segment periodicity and cause the subdivision of the anterior-posterior axis into
parasgements (Pankratz and Jackie, 1993). It is a common feature of most, if not all,
genes involved in this process that their initial broader expression domain becomes
more and more restricted and defined during embryogenesis.
The fused denticle phenotype in mirror loss-of-function embryos (McNeill et
al., 1997) and the observed fusion of segments in mirror over-expressing embryos,
combined with mirror expression in a transient stripe at the anterior border of each
segment at stage 10, point to a possible role of Mirror in finishing the segmentation
process.
The segmentation defects of araucan over-expressing embryos appear less
severe and less frequent than the observed defects in Mirror overexpressing embryos.
An explanation could be that the amounts of produced protein might be lower for
Araucan than for Mirror due to different insertion sites into the genome of the
transgenes.
Cuticle defects are rarer in heat shocked pYCQll-mirror embryos than in
Gal4xUAS-wz>ror embryos. This is most likely due to the short time of Mirror activity
in these embryos. Mirror protein is still detected in western blots of total protein extracts
30 min after heat shock but is significantly reduced compared to protein extracts from
96

Chapter 3: Mirror
embryos processed directly after heat shock. No Mirror protein is detected Ih after heat
shock (not shown). The rapid degradation of Mirror protein after heat shock stands in
contrast to the extended time of Mirror activity in Gal4xUAS-m/rror embryos. Over
expressing Mirror using the P-tubulin-VP 16-Gsi\4 driver line leads to large amounts of
Mirror protein which accumulates for several hours and therefore during different stages
of development in the embryo. This in turn results in severe cuticle defects like shown
in figure 3.6.B.
Encouraged by the observed cuticle defects in mirror mutant and overexpressing
embryos, and having shown that high levels of Mirror protein were detected in heat
shocked pKB12-mz>ror embryos, we decided to stage the embryos for 4h at RT prior to
heat shock at 36°C. This allowed us to obtain a staged collection in which the majority
of embryos would normally express Mirror in a striped pattern, potentially allowing us
to detect Mirror targets that normally function at stage 11. Several more reasons led to
the choice of this developmental stage. Firstly, mirror expression is negligible at earlier
stages and so might be the availability of cofactors, which could be indispensable for
proper Mirror function during development. And secondly, if Mirror acts to suppress
genes which are not expressed in early developmental stages the analysis would be in
vain. Finally, embryos younger than embryonic stage 10 do not survive a heat shock
treatment. It has been reported that embryos which were heat shocked earlier than stage
10 are irreversibly arrested in development and that the intermediate filament
cytoskeleton collapses around the nuclei as a result of the treatment (Walter et al.,
1990). To avoide complications due to these effects, earlier heat-shock treament was
avoided. Thus, it was decided to carry out the microarray analysis at a time where the
majority of embryos reached embryonic stage 11 (including staging + heat shock +
recovery).
97

Chapter 3: Mirror
98

Chapter 4: Heat shock mirror versus heat shock ^control”
CHAPTER 4
Heat shock m/rror versus heat shock “control”
4.1. The heat shock Mirror over-expression model
4.1.1. Direct comparison of expression profiles from heat
shocked pKBI 2-m/rror and yw embryos
The development of the Drosophila embryo, in particular at early stages is a
rapid process, with dramatic changes in gene expression. In order to obtain a tightly
staged collection of embryos in which the majority of embryos are at embryonic stage
11, two initial 30 min pre-lays were discarded to aid in a tight staging of embryos.
pKB\2-mirror and yw control embryos were collected for 30 min on apple juice plates
and kept for 4h at RT. Both types of embryos were subjected to heat shock at 36°C for
Ih followed by a recovery period of 30 min at RT, referred to as the “long hs”. A
similar approach has been used to identify heat shock response genes in the Drosophila
embryo (Leemans et al., 2000). The purpose of the recovery period was to allow down
regulation of heat shock induced genes and to permit Mirror protein to be produced and
to act on target genes, mirror must be transcribed, translated, bind to a regulatory
element of a downstream targets and induce changes in the level of its relative
expression. A positively regulated gene needs to be transcribed and the existing
transcripts of a negatively regulated gene need to be degraded to exhibit differences in
the amount of mRNA in the embryo at the moment of processing for the microarray
analysis (fig.4.1).
99

Chapter 4: heat shock mirror versus heat shock “control”
Heat shock over-expression model
pKBl2- mirror embryos 4h AEL
yw control embryos 4h AEL
36°C water bath for Ih +30 minutes at RT
iii
RNA extraction
cDNA synthesis
Target preparation
iii
Microarrayanalysis
PotentialMirror
targets
GeneA with similar expresaon profile in both pm ulations are sobtracted from the
analysis
Figure 4.1. The Mirror over-expressing model. Both, pKB 12-m/rrar and _yiv control embryos were
collected for 30 minutes, aged and subjected to heat shock. After a 30 minutes recovering period at RT,
RNA was isolated from stage 11 embryos, processed and differences in gene expression were assessed
using Drosophila Genome Arrays. The expression profile of pKBI2-m/rr6>r embryos was compared to
that of yw embryos. This comparison should directly eliminate the heat shock response genes from the
analysis and only genes with altered levels of expression between both populations were considered as
potential Mirror targets.
100

Chapter 4: Heat shock mirror versus heat shock ^control”
4.1.2. CG12199 seemed to be repressed by Mirror
The direct comparison of the gene expression profile from heat shocked pKB12-
mirror and heat shocked yw embryos revealed many genes with altered levels of
expression (see appendix figure A) which were ranked according to their average fold
change. A list showing upregulated and downregulated genes like it was generated
during the course of this work is shown in table I in the appendix. Flags were not taken
into account and an arbitrary ± 2 fold cut-off was used. The availability of tools (cDNA,
antibodies, P-elements) were also considered to shortlist genes for an in vivo analysis.
Therefore selected candidate genes do not necessarily appear on the very top of the list.
A more recent analysis of the data uses a ranking system dependent on a false discovery
rate and is presented in table III in the appendix. The previously uncharacterised gene
CGI2199 will be presented as a representative of the initial comparison (fold change
ranking). The bioinformatic analysis showed almost 3 times more transcript of this gene
in the yw control embryos than in the mirror over-expressing embryos. The difference
suggested that this gene is downregulated by Mirror in this experiment (fig.4.2).
Subsequently, the expression pattern of this gene was analysed by in situ hybridisation.
At stage 11, CGI 2199 is expressed at low levels in the entire embryo and enriched in a
segmented pattern in the epidermis and the developing hind gut where it remains till
later stages (fig.4.3.A). At stage 14, transcript is present in the salivary glands, the CNS,
the proventriculus and the hindgut but not in the amnioserosa (fig.4.3.B). This pattern of
expression made CGI 2199 an attractive candidate. However, embryos over-expressing
or deficient for mirror showed the normal wild type expression pattern (not shown),
demonstrating that CGI2199 is not regulated by Mirror in the embryo. This was also the
case for all other 20 potential candidate genes which were selected for the in vivo
analysis. Table 7 summarises which genes were tested.
101

Chapter 4: heat shock m irror versus heat shock "control"
800
700
600
500
400
300
200
100
0
F igure 4.2. C G12199, repressed by Mirror? The direct comparison o f amounts o f biotinylated cRNA
from heat shocked pKBI2-m /rr«r embryos versus heat shocked yw embryos led to the identification of
the uncharacterised gene CG12199. The histogram suggests that over-expression o f Mirror inhibits
C G I2199 expression. Here and in all following histograms is shown the average signal intensity o f the
normalised replicates. I’lie error bar represents the standard deviation, which is a measure o f how widely
values are dispersed from the average value (the mean). n=3 for each experimental condition.
F igure 4.3. in situ hybridisation of CG 12J99 in wild type embryo. A) A stage 11 embryo shows
ubiquitous expression o f unknown gene C G I2199 with enrichment in the hindgut and in a segmented
pattern. B) At the embryonic stage 14, CG 12199 is expressed in many tissues including the salivary
glands, the proventriculus, the hindgut and the CNS. Anterior is to the right and dorsal is up.
102

Chapter 4: Heat shock mirror versus heat shock ^control”
Table.7. Tested genes from direct comparison approach.
Gene name Observed expression pattern/commentrace dorsalzen dorsalU-shaped dorsalpannier dorsaldpp dorsalpdm2 segmentally repeated stripeslola didn't workSerine protease 2 ubiquitousMyosin light chain 2 ubiquitousabnormal spindle ubiquitousStam initially ventral, later ubiquitousyan ubiquitousLcp2 ubiquitousLcp65AG2q ubiquitousMdr49 ventral expressionl(3)mbt ubiquitousnAcRalpha-80B ubiquitousCG12199 ubiquitous, highly expressed in hind gutCG7271 proventriculus/CN SCG13213 ubiquitousCG14014 ubiquitousCG13532 ubiquitously low expression levels
4.1.3 Strain variation in gene expression
These data led us to examine if the observed differences in gene expression for
CGI2199 and other candidate genes are indeed due to Mirror activity. The expression
profile of non heat shocked ipKBXl-mirror and non heat shocked yw embryos were
compared in an additional microarray analysis. To do so, embryos were again collected
for 30 minutes and aged for 5^^^h AEL at RT to obtain comparable stages of
development (around stage 11). If CGI2199 was a true Mirror target the comparison of
normal developing embryos should reveal that the levels of detected transcript are
similar in the untreated embryos from both pKB12-mz>ror flies and yw controls. This
was not the case. The analysis showed that many genes are expressed in different levels
in Drosphila embryos from different strains at the same stage of development (fig.4.4.).
103

Chapter 4: heat shock m irror verstts heat shock 'h otttroV'
A) Signal log2100000 - 100000
stage 11 pKB non hs embryos10000, 10000
1000, 1000
stage 11 yw non hs embryos
2x tOx
010.1 10 100 1000 10000 1000001
Signal log2
B) Signal log2100000
stage 11 pKB hsi embryos10000
1000
lOx2x stage 11
pKB hs2 embryos10
2XlOx
~ - l a i100000
0.11 10 100 1000 10000
Signal log2
F igure 4.4. Scatter Graphs visualises the differences in the genetic background. A) Expression fffofde of
non hs pK B12-m /m ;r embryos compared to non hs yw embryos. Note the broad scatter graph with many
genes exhibiting a more than two fold difference, which points towards the endogenous differences in
gene expression o f the two fly strains. B) A repUcate o f the same experimental condition using pKB12-
m irror embryos exhibit a tight expression profile around the Ix line (no difference). The other lines
indicate a 2x and lOx difference o f the measured Signal intensity. Take note o f the differences in the
shape o f scatter graph in A) and B).
101-

Chapter 4: Heat shock mirror versus heat shock „control”
(See also figure B in the appendix)
To rule out the possibility that leakiness of the heat shock promoter at RT drives
Mirror expression at low levels and might be responsible for the observed differences,
the expression profiles of stage 11 embryos raised at 18°C were also compared. The
differences in gene expression were the same as observed for the embryos kept at RT.
To put it briefly, the flies are different even in the absence of m irror
overexpression. The signal intensity, representing the level of gene expression for
CG12199 in non heat shocked ipKRXl-mirror is significantly lower compared to that
of non heat shocked jiv embryos (fig.4.5.). It turned out, that this comparison was
extremely important to identify what is the “baseline” of expression of individual genes
prior to heat shock and in embryos from different genetic backgrounds.
4.1.4. "pKB/yw-m/rror" a new genetic background
The fly stock used to generate the original transgenic pKB 12-mirror flies does
not exist anymore and it was therefore not possible to repeat the experiment using
control flies with the same genetic background. For that reason I attempted to cross the
mirror transgene of the pKB 12-mirror flies into the yw background. A male pKB 12-
mirror fly was crossed to a yw female fly and red eyed males were crossed for 8
generations to yw females. Subsequently red eyed yw flies, carrying the mirror
transgene were crossed for several generations to each other in order to obtain a nearly
homozygous stock named pKB/yw-m irror. Embryos from these flies have been
collected and treated (heat shocked and non heat shocked) in the same way as described
above and their expression profile was compared with that from the other samples
(fig.4.5).
105

Chapter 4: heal shock m irror versus heat shock "coutoV
i/
Figure 4.5. C G 12J99 a false candidate. The expression profiles o f D rosophila embryos with diffeent
genetic background can not be compared directly to identify Mirror targets. I'he observed differcnc of
more than two fold for the gene C G 12I99 between pK B-m im ;r hs and yw hs embryos is due to diffeent
expression levels in embryos with different genetic background. The comparison o f the expresion
profiles obtained from pKB-mirror non hs and yw non hs embryos also displays an almost 2 old
difference. Ihe ''pseudo downregulation” is not due to Mirror activity. C G I2199 expression in a lird
genetic background is also presented in the histogram (yellow ) (see text). n=3 for each experimeital
condition.
106

Chapter 4: Heat shock mirror versus heat shock ^control”
It became evident that these embryos still display dramatic differences in gene
expression compared to yw (and also pKB12-w/rror) embryos. pKB/yw-mirror flies
represent therefore another, third genetic background and can not be used for a direct
comparison of expression profiles to identify genes which alter their level of expression
due to Mirror activity.
107

Chapter 4: Heat shock mirror versus heat shock ^control”
Discussion
To conclude, the expression profile of heat shocked pKBl2-m irror and heat
shocked yw embryos is not directly comparable to identify potential target genes which
are regulated by Mirror. The assumption that genes would be expressed at similar levels
in Drosophila embryos of different genetic backgrounds proved to be false. 13
differentially expressed genes were analysed but not one candidate gene could be
related in vivo to Mirror function.
Using this direct comparison approach, 96 potential downstream targets of the
homeodomain transcription factor labial were identified in a genome wide analysis
using microarrays (Leemans et al., 2001). Labial is the most proximal gene within the
Antennapedia complex and is required for proper head development and specification of
cellular identity in the brain and the midgut (Hirth et a l, 1998; Hoppler and Bienz,
1994). It would be interesting to see, which of the proposed candidate genes prove to be
dependent in vivo on labial function.
Another study identified 682 transcripts which showed highly significant
changes in expression levels in response to Otx2 over-expression in flies (Montalta-He
et al., 2002). Otx2 is the human orthologue of the Drosophila homeobox gene
orthodenticle (old). Members of this family have a conserved role in the embryogenesis
of the brain and the head (Leuzinger et al., 1998). Gene replacement experiments have
shown that otd and Otx2 are functionally equivalent, in that over-expression of either
gene in a null mutant background of Drosophila or in mouse can rescue brain and
cephalic defects (Acampora et al., 1996; Sharman and Brand, 1998). Changes in gene
expression were compared between heat shock induced over-expression of otd and Otx2
and it was postulated that about 90 genes are common downstream targets of the fly otd
gene and the human Otx2 gene in Drosophila (Montalta-He et al., 2002). The in vivo
108

Chapter 4: Heat shock mirror versus heat shock „controï’'
validation of this microarray approach is still ongoing.
Targeted misexpression of the transcription factor glial cells missing in the
neuroectoderm was used to identify potential downstream targets by means of
microarray analysis. A sca-Gd\-A enhancer trap line was crossed to a UAS-gcm
responder line and the expression profile was compared to that of wild type embryos
(Oregon R) at stage 11 and at stage 14. From about 14000 genes represented on the
microarray, over 1200 genes were reported to be differentially expressed as a result of
gem function. The subsequent in vivo investigation confirmed two genes with coherent
alteration in the level of expression in gem over-expressing and gem loss-of-function
embryos, namely the genes bangles and beads and wrapper (Egger et al., 2002).
Thus it seems that the over-expression approach can identify many genes with
different levels of expression, however since few candidate genes were reportedly
altered in mutant embryos there may be a relative high percentage of false candidates in
the analysis. A recent microarray study which also investigated downstream targets of
Gem using a similar approach to this study, reported a 88% false positive rate in their
analysis (Freeman et al, 2003). This study also predicts that a similar rate of false
positives among the 1200 putative Gem regulated genes proposed by Egger and
collaborators. The complexity of the whole-embryo tissue has been mentioned to be
responsible for the high numbers false positives. However, it is also possible that genes
are expressed at different levels in flies from different genetic background used in these
comparisons which contributes to the high number of false positives.
At a first glance, this hypothesis seems to stand in contrast to a cDNA
microarray study, which analysed the contribution of sex, genotype and age to
transcriptional variance of two Drosophila fly strains (Oregon R and Samarkand) (Jin et
al, 2001). Combinations of differently labelled cDNA probes from adult flies of
109

Chapter 4: Heat shock mirror versus heat shock „control”
different sex and age (1 week or 6 weeks), were hybridised on the same array. The
microarray analysis revealed that gene expression in adult flies is affected most strongly
by sex, less so by genotype, and only weakly by age (Jin et al., 2001). However, in none
of the experiments were samples from Oregon R and Samarkand flies directly compared
on the same array. The authors stated that: “If we had contrasted Oregon R and
Samarkand on each array instead of the two ages, it is likely that the genotype effect
would have been much more significant” (Jin et al, 2001).
Thus, the observation that gene expression in Drosophila embryos of different
genetic background varies considerably stands by no means in contrast to the above
mentioned study. The major advantage of the Affymetrix Genome Arrays is that each
microarray is hybridised with cRNA from only one sample as opposed to other
techniques, in which the array is hybridised with differently labelled probes from two
samples. This allowed us to reprocess the generated data by simply varying the
bioinformatic analysis to circumvent the issue of the different levels of gene expression
in different genetic backgrounds. The new approach I used to identify downstream
targets of Mirror will be presented in the next chapter.
110

Chapter 4: Heat shock mirror versus heat shock ^control”
111

Chapter 5: Heat, shock versus non heat shock
CHAPTER 5
Heat shock versus non heat shock
5.1. The heat shock subtraction model
5.1.1. Indirect comparison of expression profiles from mirror
over-expressing and yw embryos
The direct comparison of transcript levels in heat shocked p¥iB\2-mirror and
heat shock yw embryos was misleading due to differences in gene expression of the
different genetic backgrounds. Therefore the expression profiles of heat shocked versus
non-heat shocked embryos of the same genetic background were compared separately.
Three lists of genes with altered levels of expression were generated and compared
(fig.5.1). Genes which altered their level of expression in heat shocked versus non heat
shocked yw embryos were considered as heat shock response genes (see appendix table
IV). These genes were subtracted from the two lists containing heat shock and the
Mirror response genes of the pKBl2-mirror and pKB/yvz-mirror background (see
appendix table V + VI). Only genes which consistently altered their levels of expression
in embryos of the two different mirror over-expressing backgrounds, and which
remained unchanged in the yw background, were considered as potential Mirror target
genes. Table II and VII in the appendix were generated using a ± 2 fold cut off and a
FDR of 0.5 respectively. This approach will be referred to as the “heat shock subtraction
model”. The identified genes were analysed further in mirror over-expressing embryos
using various Gal4 driver lines and in mirror loss of function embryos.
112

Chapter 5: heat shock versus non heat shock
Mirrortargets
pKB-iffirr long hs versus
pKB-m/rr non hs
pK R tyyv-m irr long hs versus
pK B^^-m irr non hs
Hsresponse
genes
yw long hs versus
yw non hs
Figure 5.1. Heat shock subtraction model. Embryos from the three different genetic backgrounds were
collected for 30 min, aged and subjected to heat shock at 36°C. A sample o f each collection was kept at
RT and aged equally to about stage 11. Two fly strains, pKB-w/rror and pKB/yw-/w/>ror contain a mirror
transgene under the control o f a heat shock promoter. Yw embryos were used as a wild type control. RNA
from heat shocked and non heat shocked embryos was processed and used for microarray analysis. A list
o f genes with altered expression in hs and non hs embryos was generated independently for each genetic
background. These lists were than compared and the heat shock response genes, identified in the yw
background were subtracted. Genes which didn’t change their level o f expression in the control yw
embryos but in the two Mirror over-expressing embryos were considered as potential Mirror targets.
113

Chapter 5: Heat shock versus non heat shock
As the aim of the project was to identify genes which are directly regulated by
Mirror, I will not present clusters of genes which are organised according to their
putative function. Even though this technique summarised nicely a study which aimed
to analyse circadian gene expression in Drosophila (McDonald and Rosbash, 2001), the
meaning of these gene clusters in order to identify genes that are regulated by a specific
transcription factor is not always very clear (Egger et ah, 2002; Leemans et aL, 2000;
Leemans et aL, 2001; Montalta-He et a l, 2002). On the contrary, I will present the
analysis of some candidate genes and focus on results obtained in the course of the
project. The table 8 summarises which genes were analysed by in situ hybridsation or
antibody staining to validate if their expression is regulated in vivo by Mirror.
Table 8: Tested genes from heat shock subtraction approach.
Gene name Observed expression pattern Commentatonal segmentally repeated lateral
patchesrepressed by Mirror
wingless segmentally repeated stripes repressed by Mirrorhedgehog segmentally repeated stripes repressed by Mirrorsloppy paired 1 segmentally repeated stripes repressed by Mirrorsloppy paired 2 segmentally repeated stripes repressed by Mirrorgooseberry segmentally repeated stripes repressed by Mirrorengrailed segmentally repeated stripes repressed by Mirrorci broad segmentally repeated
stripesnot tested
huckebein CNS repressed by Mirrorbeaten path la CNS repressed by Mirrorfc96Cb CNS repressed by MirrorKrüppel CNS repressed by Mirror +
ectopic expression in LOT mutants
Sry-alpha ubiquitous not testedbloated tubules Segmentally repeated
stripes/CNSno changes
Z600 dorsal no changesm-spondin didn't work not testedmyotubularin ubiquitous no changesechinoid ubiquitous not testedhuntingtin ubiquitous no changesLcp65AG2q ubiquitous not tested
114

Chapter 5: Heat shock versus non heat shock
Gene name Observed expression pattern CommentLcp2 ubiquitous not testedmew broad segmentally repeated
stripesno changes
ko CNS no changesine gut/CNS no changesdve didn't work not testedgem CNS repressed by Mirror
upregulated in LOFarmadillo ubiquitous no changesbagpipe segmentally repeated patches in
mesodermno changes
yin ubiquitous no changesdorsocross didn't work not testedpatched didn't work not testedCG6900 ubiquitous no changesCG14112 forgut no changesCG2191 amnioserosa not testedCGI 5068 ubiquitous no changesCG7920 amnioserosa and gut not testedCG6770 ubiquitous not testedCG7204 ubiquitous not testedCG6709 ubiquitous not testedCG1300 ubiquitous not testedCG 13532 ubiquitous not testedCG8349 ubiquitous not testedCG3775 ubiquitous not testedCG13213 ubiquitous not testedCG7290 ubiquitous not testedCG8089 ubiquitous not testedCGI 1659 few cells in the gut not testedCG5704 ubiquitous not testedCGI 7778 ubiquitous not testedCGI 0362 ubiquitous not testedCG7294 ubiquitous not testedCG6256 ubiquitous not testedCG9109 ubiquitous not testedCGI 1152 ubiquitous not testedCG7876 didn't work not testedCG5272 ubiquitous not testedCG3045 ubiquitous not tested
115

Chapter 5: Heat shock versus non heat shock
5.1.2. Over-expression of Mirror downregulates a to n a l
expression in the embryo
One of the genes identified using the heat shock subtraction model is the
proneural gene atonal, which encodes a bHLH protein and has transcription factor
function. It behaves in many ways just like the proneural genes of the achaete-scute
complex (AS-C) and is required for the differentiation of sensory organs such as
photoreceptors and chordotonal organs. The latter are mechano-sensory organs and
function as stretch or proprio-receptors (Jarman et aL, 1993a; Jarman et al., 1993b). In
atonal mutants, all chordotonal organs are absent except for one scolopidium of Ich5.
The scolopidium is the basic unit of the chordotonal organ (Jarman et aL, 1995). Early
in embryonic development, atonal is expressed in all chordotonal organ progenitor
cells, called founder precursors and gets restricted later to only a particular set of
precursors in a Notch independent process of lateral inhibition which is not well
understood. Flies with mutated atonal function have disrupted eye development.
However, expression of atonal is not restricted to the ventral half of the eye but is
produced in all cells just anterior to the morphogenic furrow (Jarman et aL, 1995). This
suggests that atonal is not regulated by mirror in the eye. Nevertheless, atonal is an
interesting candidate because it functions as a neurogenic gene in the place of the bHLH
proneural genes of the AC-S in the chordotonal organs. Expression of atonal is
downregulated in mirror over-expressing (heat shocked) embryos and is unchanged in
yw embryos (fig.5.2). The in situ hybridisation using an antisense RNA probe for atonal
demonstrates the almost complete abolishment of atonal expression in ubiquitously
Mirror over-expressing embryos (19/20) with the genotype P-tubulin-VP 1 6-Gsl\4 x
\JAS-mirror (fig.5.3.). However, preliminary analysis of mirror loss-of-function
embryos showed normal expression of atonal (not shown).
116

Chapter 5: heat shock versus non heat shock
700
500c2c 400
1 300O)« 200
^ 100
Figure 5.2. Over-expression o f Mirror reduces the level o f atonal transcript in the embryo. I he histogram
displays the average signal intensity measured for biotinylated cRNA after normalisation o f the gene
atonal prepared from embryos o f three different genetic backgrounds. In this and all the follow ing
histograms cRNA from yw embryos is represented in blue, pK B-m /m ;r cRNA is labeled in red and
cRNA from pK B /yw -m irror is colored in yellow. Stage 11 embryos were subjected to heat shock or in
parallel kept at RT and processed for the microarray analysis. Heat shocked and non heat shocked yw
embryos were used to identify genes which alter their level o f expression due to the treatment. The heat
shock procedure does not vary the level o f atonal transcript significantly in yw control embryos. pKB-
and pKBY W -m/rror embryos are two different fly strains containing a m irror transgene under the control
o f heat shock promoter. The Mirror over-expressing (heat shocked) embryos have reduced levels of
atonal transcript compared to non heat sh œ k ed embryos o f the same strain. Embryos were collected for
30 min on apple juice plates, aged for 4h, heat shocked for Ih at 36°C and kept another 30 min at RT
(named long hs) with the intention that their developmental stage is about stage 11. Non heat shocked
embryos were kept at R1 and prtxx'ssed 5'^ h AEL. n=3 for each microarray condition.
B
» #
Figure 5.3. atonal in situ hybridisation in stage 11 embryos. A) The wild type embryo shows evidence of
atonal transcript in clusters o f cells in the lateral epidermis within the forming PNS. B) Ubiquitous over
expression o f Mirror using a J^-tubulin-VPlCGaiA driver line crossed to a V A S -m irror responder line
nearly com pletely abolished atonal expression in the embryo. Unaffected are clusters o f cells in the head
region. Anterior is to the right and dorsal is up.
117

Chapter 5: Heat shock versus non heat shock
This still needs to be confirmed using carefully staged embryos and antibody staining
techniques.
Thus, ubiquitous expression of mirror is able to repress atonal expression in the
embryo. On the contrary, it seems that the absence of mirror alone is not sufficient to
expand atonal expression in cells of the proneural clusters which give rise to the
chordotonal organs.
5.1.3. Over-expression of Mirror represses gem expression
The transcription factor glial cells missing (gem) encodes a transcription factor
with a new type of DNA-binding domain with no overall homology to other known
proteins (Schreiber et aL, 1997). Gem is the primary regulator of glial cell fate in
Drosophila. In many cases neuroblasts are the common precursors of glial cells and
neurons. The role of gem is to direct the cell fate switch between neurons and glial cells
by activating the glial developmental program in the neuroblast. Glial cells express gem
as soon as they are born and if gem is lacking, presumptive glial cells are transformed
into neurons. Conversely, when gem is ectopically expressed neurons are transformed
into glial cells. By stage 11, gem expression becomes evident in the glial cells of the
peripheral nervous systems in the lateral epidermis and in the central nervous systems.
In the CNS, gem is expressed in progeny of specific neuroblasts and in the brain. These
cells include glioblast precursors of longitudinal glia, exit glia, glia asssociated with
peripheral axons, and glia associated with sensory organs (Jones et aL, 1995). gem is
not expressed in midline glial cells where mirror is present and gem mutants have
abnormal longitudinal CNS tracts (Jones et aL, 1995).
Ectopic expression of Mirror throughout the embryo downregulates the level of
118

Chapter 5: Heat shock versus non heat shock
gem expression using the experimental conditions described above (fig.5.4). Repression
of gem can also be seen in embryos over-expressing mirror under UAS control by a
prd- Gal4 driver. In a stage 11 embryo the prd-GdXA enhancer trap drives expression in
every other segment providing an internal control for gene expression in alternate
segments. Double in situ hybridisation using a mirror antisense RNA probe (red) and a
gem probe (black) demonstrates the repression of gem by mirror (fig.5.5.A+B).
As it is possible that members of the Iroquois Complex in Drosophila can
functionally replace each other I used embryos which are deficient for the entire
Iroquois locus for in vivo studies. This should eliminate the possibility of functional
replacement by members of the Iroquois Complex in PNS in the absence of Mirror
function. The analysis of Mirror function in the CNS is unambigious as it is the only
Iro-C member expressed in this tissue in the Drosophila embryo. If mirror normally
acts in the embryo to inhibit expression of gem, the analysis of Iroquois mutant embryos
should show an increase in gem expression in corresponding cells which have lost
mirror expression in the CNS. Quite the opposite seems to be the case: gem expression
seems to be lost in embryos which lack mirror function as revealed by in situ
hybridisation (fig.5.6.). In addition, an antibody directed against the protein of the gene
reversed polarity (repo), can be used to assess if Mirror regulates gem expression. The
gene repo is known to be positively regulated by gem (Akiyama et aL, 1996). Repo is
essential for the differentiation and maintenance of glia function (Halter et aL, 1995).
Repo expression starts at stage 11 in one cell per hemisegment, the longitudinal
glioblast, which divides symmetrically and both daughter cells, named longitudinal glial
(LG) cells start migrating towards the midline. After further division, the group of
longitudinal glia stretches in anterior-posterior direction and additional Repo expressing
cells intermingle with them.
119

Chapter 5: heat shock versus non heat shock
inc2c
«c
in
<
900800700600500400300200100
0
\0
I
< f\0
/ /Figure 5.4. Histogram o f gem expression. Heal shocked pK B -m irror embryos have reduced levels o f gem
transcript compared to their non hs control (red). I he slightly lower levels o f gem expression in long hs
pKBYW embryos (yellow ) or long hs yw control embryos (blue) are statistically not significant. n=3 for
each microarray condition.
» * *
Figure 5.5. Ectopic Mirror expression represses glial eells missing (gem). A) In situ hybridisation o f a
wild type embryo at stage 11 reveals expression o f gem in small clusters of cells (arrow) in the
neuroectoderm. B) Over-expression o f m irror (red) in a prd-G al4 x V A S -m irror embryo abolishes
expression o f gem (black) in these cells in every other segment (arrows). lateral views, anterior to the
right and dorsal is up.
120

Chapter 5: Heat shock versus non heat shock
At stage 14 about 28 Repo positive cells are found per hemineuromere (see
fig.5.7.A+C). Peripheral and exit glia begin to migrate out of the ventral nerve cord at
stage 15 to line up along the segmental nerve (fig.5.7.E; (Halter et a l, 1995).
Consistent with the reduced levels of gem expression in vivo, but in contrast to
the microarray read-out is the loss of Repo positive cells in the developing CNS of
Iroquois mutant embryos (fig.5.7. B+D). In addition the organisation of Repo positive
cells within the ventral nerve cord at stage 16 appears severely disrupted (fig.5.7.F).
121

Chapter 5: heat shock versus non heat shock
Figure 5.6. gem in situ hybridisation and Mirror antibody staining. A) wild type embryo at stage 14
shows expression o f gem (black) and Mirror (green) in the CNS. B) The Iroquois mutant embryo,
identified by the lack o f Mirror staining seems to have reduced levels o f gem expression in the CNS.
Ventral view with anterior to the right.
Figure 5.7. Mirror loss-of-function embryos lack Repo expressing cells in the CNS. Wild type embryo
(A, C, E) and Iroquois mutant embryos (B, D, F) at stage 14, 15 and 16 respectively. A,C,E) From stage
14 onwards about 28 cells per hemineuromere are labelled by the anti-Repo antibody. This number
remains about the same in embryos o f later stages but cell migration takes place till stage 16. B,D,F) The
pattern o f the Repo expressing cells in Iroquois mutant embryos is severely disrupted and some cells
might be missing. The disorganisation o f glia cells within the CNS is most obvious at stage 16. The arrow
in E+F, point towards the exit and peripheral glia which seem to have migrated normally in the absence
o f Mirror function. All panels are ventral views with anterior to the right.
122

Chapter 5: Heat shock versus non heat shock
5.2. Over-expression of Mirror inhibits the expression
of segment poiarity genes
5.2.1. gooseberry
The segment polarity gene gooseberry (gsb) encodes a homeodomain
transcription factor of the paired class (Baumgartner et a l, 1987). Gooseberry protein is
required for the specification of naked cuticle in the epidermis and specific neuroblasts
in the central nervous system (Zhang et aL, 1994). gooseberry is first detected during
gastrulation of the embryo in 7 stripes. By convention these stripes are called the odd
numbered stripes and 7 additional, even numbered stripes will appear later in
development (fig.5.9.A). All 14 stripes are finally established at embryonic stage 9-10.
Reduced levels of gooseberry transcript were detected in the course of the microarray
analysis in mirror over-expressing embryos (fig.5.8.). The segmental expression pattern
of gooseberry overlaps with the expression of engrailed in the epidermis of the embryo.
Therefore gooseberry like engrailed, is expressed at the posterior border of the segment,
adjacent to the mirror expression domain (fig.5.9.C). Ubiquitous over-expression of
m irror in the embryo severely disrupts gsb expression (fig.5.9.B). Similarly,
misexpressing mirror using a prd-GdXA driver inhibits gsb expression shown in figure
5.9.D. Nevertheless, mirror mutant analysis did not reveal aberrant gooseberry
expression.
123

Chapter 5: heal shock versus non heal shock
1800
1600
1400
12001000
800
600
400
2000
(A
# /Figure 5.8. Histogram o f gooseberry expression. I he Mirror over-expressing (heat shocked) embryos
have reduced levels o f gsb transcript but not the yw control embryos. All embryos processed for the
microarray analysis were about stage 11 (5'^ h AEL). n=3 for each microarray condition.
&B
D
Figure 5.9. Over-expression o f Mirror repressed gooseberry (gsb) expression. A) gsb in situ hybridisation
in a wild type embryo. At stage 11 gsb is expressed in 14 stripes. B) The segmental expression pattern o f
gsb is severely disrupted in Mirror overexpressing embryos o f the genotype VP16-Gal4 x VA S-m irror.
C) Mirror antibody staining (brown) and gsb in situ hybridisation in a wild type embryo. Mirror and gsb
are expressed in cells adjacent to each other at the segmental border. D) The Gooseberry antibody
staining in a prrf-Gal4xUAS-/7urrt>r emtffyo shows repression o f Gsb in every other segm ent due to
ectopic expression o f Mirror
m

Chapter 5: Heat shock versus non heat shock
5.2.2. hedgehog
Another segment polarity gene, hedgehog (Hh) also exhibits reduced levels of
expression in mirror over-expressing embryos, hedgehog encodes a protein which is
secreted by the engrailed expressing cells in the posterior (P) compartment (Kopp et al.,
1997; Struhl et al., 1997). Hedgehog protein spreads into the mirror expression domain
across the anterior and the posterior boundaries to form opposing concentration
gradients that organize cell pattern and polarity. On the other side Hedgehog protein is
required to stabilise wingless expression. Misexpression of mirror in the paired pattern
results in repression of hedgehog even before the normal onset of the segmentally
repeated pattern of mirror expression in the embryo (fig.5.10.). The mirror loss-of-
function analysis using Iroquois mutant embryos of stage 10 and 11 was indistinct.
Early at stage 10, mirror is expressed in a thin one cell wide stripe next to hedgehog
(fig.5.1 l.A,+B). In Iro' embryos of similar stages, hedgehog expression is generally
normal (fig.5.1 l.D+E), although very rarely (< 10% of mutant embryos) hedgehog
seems to expanded into the neuroectoderm at stage 11 (fig.5.11.compare C+F).
5.2.3. wingless and sloppy paired
Wingless (wg) and sloppy paired (sip) are segment polarity genes which are
expressed in overlapping domains adjacent to hedgehog, engrailed and gooseberry. The
interplay between these different segment polarity genes marks the parasegmental
border. Wingless encodes a secreted molecule which spreads between the anterior and
posterior compartments of segments where it has a role in establishing different cell
fates. Wingless secretion is dependent on hedgehog, produced in adjacent
compartments. The Wg protein is distributed more widely than its mRNA.
125

Chapter 5: heat shoek versus non heat shock
Figure 5.10. Ectopic expression o f Mirror represses hedgehog expression in the embryo. A) hh in situ
hybridisation in a wild type embryo at stage 8. hh is expressed in a segmented pattern interrupted by cells
in the ventral midline. B) Double in situ hybridisation for mirror (red) and hh (black) in a prd-Gd\A x
UAS-m/>ror embryo o f stage 8. Expression o f hh is significantly disrupted by ectopic expression o f
Mirror. Ventral view with anterior to the right.
Figure 5.11. Mirror antibody staining combined with hh in situ hybridisation. A+B) Ventral and dorsal
view o f a wild type embryo at stage 10, respectively. Mirror (green) is expressed in a stripe o f 4-5 cells
per hemisegment adjacent and posterior to the hh (black) stripe. C) At stage 11 Mirror is expressed in
delaminating neuroblasts and in the ventral midline. D-F) The absence o f Mirror staining marks the
Iroquois mutant embryos o f comparable stages to the wild type A-C, respectively. Hedgehog expression
seems normal in the absence o f Mirror. F) Sporadically Iro' embryos show an expansion o f hh into the
Mirror expression domain (dotted line). Anterior is to the right.
l2(o

Chapter 5: Heat shock versus non heat shock
Transcription of wingless is in single-cell-wide stripes, whereas the protein is
detected over 3-4 cell diameters (Lawrence et al., 1996).
The genes sloppy paired 1 and 2 encode transcription factors with a fork head
domain. During the patterning of the epidermis, sloppy paired genes functions upstream
of wingless to maintain wg expression (Cadigan et al., 1994). Conversely, it has been
demonstrated that the sip genes function downstream of Wingless signalling to generate
specific neuroblasts in the ventral nerve cord of the Drosophila embryo. Expression of
sip is lost from the neuroectoderm in wingless mutant embryos and ectopic expression
of wingless induces ectopic expression of sip. In addition gooseberry interacts with
sloppy paired to prevent wingless from conferring identity to specific neuroblasts (Bhat
et al., 2000). Interestingly, none of these genes is coexpressed with mirror in the wild
type embryo, and over-expression of mirror in the embryo is able to repress all of the
segment polarity genes mentioned above. As representatives for this observation,
wingless (fig.5.12.A+B) and slpl (fig.5.13.A+B) expression in Mirror over-expressing
embryos using a Krüppel-GdXA (fig.5.12.C) and/or a paired-GdXA (fig.5.12D) enhancer
trap line are presented. In addition, ectopic expression of araucan, another member of
the Iroquois complex is also able to repress sloppy paired 1 (fig.5.13.C). However, all
the analysed segment polarity genes seemed normally expressed in Iroquois or mirror
mutant embryos (not shown).
127

Chapter 5: heat shock versus non heat shock
/
Figure 5.12. Ectopic expression o f mirror represses wingless expression in the embryo, wingless in situ
hybridisation in wild type embryos o f stage 9 (A) and 11 (B). Double in situ hybridisation for mirror
(red) and wingless (black) in Mirror misexpressing embryos. C) Over-expression o f Mirror in a A>-Gal4 x
UAS-m/rror embryo interrupts the segmental expression pattern o f wingless. D) Repression o f wingless is
also seen in an embryo at stage 11 o f the genotype prd-QdAA x UAS-w/rror (compare with B). Lateral
views, anterior to the right.
*%
n i t * * ’
Figure 5.13. Ectopic Mirror or ectopic Araucan can repress sloppy paired! (slpl). A) In situ
hybridisation o f a wild type embryo at stage 11 shows the segmented pattern o f the slplgene. B) Ectopic
expression o f mirror (red) in a prd-Ga\A x UAS-/w/rror embryo represses slp l (black) in every other
segment (double in situ hybridisation). C) The same driver was used to over-express Araucan, another
member o f the Iroquois complex which also repressed slp l expression. Lateral views, anterior to the
right.

Chapter 5: Heat shock versus non heat shock
Discussion
The expression pattern of 57 candidate genes, short listed by applying the heat
shock subtraction model, was analysed using in situ hybridisation or antibody staining
techniques in wild type embryos. 29, mostly uncharacterised genes, were ubiquitously
expressed. 7 of these ubiquitously expressed genes were analysed in vivo but were
uneffected by Mirror. Therefore genes with ubiquitous expresion in wild type embryos
were later discarded from further investigations. 23 showed a distinct expression
pattern, 20 were further analysed in mirror over-expressing embryos. 12 of these genes
showed repression of expression in domains where ectopic Mirror was present. These
were the genes mentioned above and in addition the genes: sloppy paired 2, engrailed,
huckebein, fd96Cb and beaten path la and Kriippel. The analysis of 5 genes were
unsuccessfull.
Initially, ubiquitous mirror over-expression using the Gal4/UAS system was
used in the in situ hybridisation screen to validate the microarray data. The P-tubulin-
FP7d-Gal4 enhancer trap line drives expression of a UAS response gene early in
development and at high levels. This leads to severe morphological defects when mirror
is driven. Moreover, the onset of expression is so early in development, about 2 ^ h AEL
(see fig.3.6. in chapter 3), that many candidate genes haven’t initiated expression yet.
The analysis of potential target genes hours after the onset of ectopic mirror expression
might not be relevant for the identification of direct Mirror target genes. Therefore other
Gal4 drivers, such as 7Cr-Gal4 or prd-GdXA, which become active later in development
have also been used. In particular the prd-Gd\A line provides a nice tool to misexpress
mirror under UAS control in every second segment in the Drosophila embryo, leaving
the intervening segment to serve as a control.
The in vivo examination of selected candidate genes, which were identified
129

Chapter 5: Heat shock versus non heat shock
using the heat shock subtraction model, confirmed that ectopic Mirror expression
indeed inhibits the expression of most of the analysed genes. Thus, the bioinformatic
analysis seems to reflect the situation in the embryo. This demonstrates that the
microarray technique is sensitive enough to detect the different levels of gene
expression under the experimental conditions. However, the effect of ectopic Mirror
could only be confirmed for negatively regulated candidate genes in the embryo. Genes
which were positively regulated according to the microarray analysis showed the
normal wild type expression pattern in Mirror over-expressing embryos. It might be that
Mirror acts primarily as a repressor in the embryo and that the upregulated genes are a
result of the variability in the microarray analysis. Nevertheless, it cannot be ignored
that the majority of analysed potential Mirror downstream targets seemed completely
normally expressed in mirror loss-of-function embryos. The only exception from this
observation was the gene glial cells missing, which encodes a transcription factor.
However, it was expected that expression of gem is enhanced in an embryo which lacks
Mirror function, which is not the case. The in situ hybridisation using a gem riboprobe
and the antibody staining for its known positively regulated target Repo, showed
reduced levels of expression. These results are inconsistent and might suggest that either
the observed decrease of gem expression is an indirect effect of a negative regulatory
feedback loop, activated in the absence of Mirror function, or that the way the
microarray experiment was designed didn’t lead to the identification of direct Mirror
targets.
The fact that most analysed target genes failed to show an aberrant expression in
a mirror mutant background argues that the microarray experiments were not revealing
genuine mirror targets. One possible artifact is that too high amounts of Mirror protein
in the embryo could lead to Mirror binding to DNA sequences for which it normally has
130

Chapter 5: Heat shock versus non heat shock
only low affinity. This in turn would inhibit or disrupt the transcription of a gene
situated downstream of this site. In fact, this could even inhibit the transcription of an
otherwise positively regulated target gene. A similar search for downstream targets of
the transcription factor glial cells missing has been performed by another group. The
microarray analysis was carried out on embryos (at stage 11 and at stage 15), which
over-expressed gem under UAS control using a sca-Gd\A enhancer trap line. This driver
becomes active at embryonic stage 9. The embryos were compared to “control”
embryos and a dramatic increase of the transcript for the known direct target repo was
detected at stage 11. Interestingly, no repo transcript at all has been detected in embryos
of the same cross at stage 15 (Egger et a l, 2002). This may support the theory that there
is an inhibitory effect caused by large amounts of a transcription factor on a normally
positively regulated gene.
Alternatively, it could be that the choice of the time point to analyse the effect of
ectopic expression of a transcription factor is crucial. If its activity proceeds for an
excessively long period of time in the embryo, genes which act further downstream in
the hierarchy will also change their level of expression. These changes of primary or
secondary targets cannot be easily distinguished based on the microarray analysis.
Mirror binding to DNA cannot be completely random and non-specific, as the
majority of genes detected in the microarray analysis didn’t change their level of
expression. For example, the amounts of transcript detected for the two genes fringe or
rhomboid are not significantly different in long hs mirror over-expressing embryos
compared to normal developing embryos. It has been shown that the gene fringe is
negatively regulated by Mirror in the eye imaginai disc (Yang et al., 1999) and ectopic
rhomboid expression has been observed in mirror over-expressing follicle cells in the
Drosophila egg (Jordan et a l, 2000). Therefore, we expected significant changes in
131

Chapter 5: Heat shock versus non heat shock
their expression levels in our analysis, which is not the case. However, it is not clear if
these genes are directly regulated by Mirror in the embryo. In addition, the results
described above showed that Mirror and Araucan can repress the transcription of sloppy
paired 1 in the embryo. This observation vs not unexpected as the homeodomain is
highly similar in members of the Iroquois complex.
Taken together, the obtained results suggest that the microarray analysis using
the proposed heat shock subtraction model is representative for the changes in gene
expression in the embryo but does not necessarily reflect real dovmstream targets of
Mirror activity. Only if a gene alters its pattern or level of expression consistently in
mirror over-expressing and in mirror mutant embryos, would it have been considered as
a potential direct target of Mirror activity and analysed further. So far, this wasn’t the
case for any of the analysed potential candidate genes. Possible explanations for this
might be that Mirror expression proceeded either for too long in the analysed embryos
and introduced changes of secondary or tertiary target genes, or that its level of
expression was to high and resulted in the binding of the protein to low affinity DNA
sites, inhibiting the transcription of genes in a non specific manner. Both possibilities
could be addressed by optimising the heat shock and staging protocol for the embryos in
a way to reduce the time of Mirror activity and the amount of produced protein in the
embryo.
It was also unclear what happened during the 30 minutes recovery period after
the heat shock in terms of transcriptional regulation of Mirror target genes. The aim of
the recovery period was on one hand to allow enough time for transcriptional regulation
by Mirror and on the other hand to reduce the levels of highly abundant transcripts of
heat shock inducible genes. However, if the changes in gene expression of Mirror target
genes “recover” with the same pace as the heat shock response genes, the induced
132

Chapter 5: Heat shock versus non heal shock
differences in expression levels of Mirror targets might also diminish. To test this, we
decided to omit the recovery period and to analyse differentially expressed genes
directly after a short heat shock treatment.
133

Chapter 5: Heat shock versus non heat shock
134

Chapter 6: Short heat shock
CHAPTER 6
Short heat shock
6.1. 30 minutes heat shock
6.1.1. The modified heat shock subtraction model
The procedure for ectopic misexpression of Mirror in the embryo using the heat
shock approach was modified in order to achieve lower amounts of protein and a shorter
period of Mirror activity in the embryo (fig.6.1.). To do so, pKB12-m/rror and>^w
embryos were collected for 30 minutes, aged for 5h AEL at RT and heat shocked for 30
minutes at 36°C. The recovery period was omitted and embryos were processed
immediately after heat shock, referred to as the “short hs”. The embryos were staged as
before to allow comparison with the non heat shocked embryos of stage 11 from the
previous analysis. The heat shock subtraction model was applied for pKB12-m/rror and
yw embryos only. The microarray analysis revealed differences of many genes with
altered levels of expression in response to heat shock and provided new data for the
analysis of potential Mirror target genes (see appendix, tables VIII and IX).
6.1.2. “Slow and fast” heat shock response genes
A side product of the search for downstream targets of Mirror was the
identification of genes which were not known to change their level of expression in
135

Chapter 6: Short heal shock
pKB pKB yw
89 k l)a —
0 0 0 M irror protein (83k Da)
64 kDa —
52 kDa —
30 m in 45m in 45m in
heat shocked
Figure 6.1. Mirror jyotein is detected after a short heat shock o f 30 minutes, yw and pK B 12-m irror
embryos at stage 11 were heat shocked for 30 and 45 minutes and total protein extracts analysed by
western blot. Heat shock induced Mirror protein is detected after 30 minutes in significantly lower
amounts than after 45 minutes o f heat shock. I he yw control embryos were heat shocked for 45 minutes
and no Mirror protein is detected by the antibody.
iVo

Chapter 6: Short, heat shock
response to heat shock. Even though it was not the aim of the analysis to identify such
genes in particular, it is worth having a closer look on their expression profile according
to the different conditions used in this study as it might mimic what happened to
transcripts of potential Mirror target genes (mRNA turnover). Given that the entire
study is based on Mirror over-expression due to heat shock, a better understanding of
the general transcriptional response of the Drosophila embryos in the model, might be
useful for the identification of Mirror target genes.
There are both slow and fast heat shock response genes. The fast heat shock
response genes increased their level of expression rapidly within 30 minutes of
exposure to an inducing temperature. Strikingly, these genes have restored their level of
expression to almost normal levels after 1 h heat shock followed by a 30 minutes
recovery period. Such a behaviour is observed for examples for the genes hsp22, alpha
tubulin at 84D, posterior sex combs, frizzled but also for twist and many uncharacterised
genes such as CGI8282. Their relative levels of detected transcript in response to heat
shock decreases quickly within 30 minutes (fig.6.2). In contrast, the slow heat shock
response genes, such as Hsc70Cb or P h a s f the latter encodes a eukaryotic-initiation-
factor-4E binding protein or the unknown genes CG4484, CG2676, CG2708 (and many
others) seemed to reach significantly increased levels of expression only during the
recovery period (fig.6.3.). This observation is consistent with a recent study, which
investigated the tissue and development specific turnover of hsp70 transcripts after heat
shock and during recovery in Drosophila. A remarkable two hours delay in the
transcriptional activation of the hsp70 gene was observed in cyst cells and others
unidentified cells of the adult testes (Lakhotia and Prasanth, 2002). The larval
Malpighian tubules are also known to synthesize hsp70 only during recovery but not
immediately after heat shock (Krebs and Feder, 1997). It is interesting to note that not
137

Chapter 6: Short heat shock
alpha Tub 84D
6 0 0
^ 500 1® 400
= 300
f 200
< 100
0
y
1600
>. 1400
posterior |sex combs 1
2 800 600 400 200
0
/ / / /
1O)in<
y yy yf ' /V V
frizzled
600
^ 500 «I 400 ci 300
200
100
O)'c5
<0
Figure 6.2. The fast heat shock response genes. The histograms shows the relative expression levels o f 3
representative genes for this group, namely alpha tubulin at 84D, posterior sex com bs and frizzled . Fast
heat shock response genes increase their level o f expression rapidly during a 30 minutes heat shock and
they restored their baseline levels o f expression after a 30 minutes recovery period. Short hs = 30 min
heat shock, long hs = Ih heat shock + 30 minutes recovery at RT, n=3 for each microarray condition.
138

Chapter 6: Short heat shock
hsc70Cb
2o>«<
1400
1200
1000
800
600
4 00
200
0
1
HH’ Vv)
/ / / / / /CG2676
CG2708
1600
>. 14001 12002 1000
2g>«<
800
600
4 0 0
2000
±
/
gO)0)<
1000 9 00 - 8 0 0 - 7 00 - 6 0 0 - 5 0 0 - 4 0 0 L 3 0 0 L 200 100 k
0 r-
y #
Figure 6.3. The slow heat shock response genes. The relative expression levels for the gene HscJOCb and
the unknown genes CG 2676 and CG2708 are shown according to the different heat shock conditions used
in this study. All three genes show highest levels o f expression after Ih heat shock follow ed by a 30
minutes recovery period at RT. All e m k y o s used for the microarray analysis were about embryonic stage
11. Short hs = 30 min heat shock, long hs = Ih heat shock + 30 minutes recovery at RT, n=3 for each
microarray condition.

Chapter 6: Short heat shock
all tissues increase their level of hsp70 transcripts in response to heat shock. For
example, no induction of any of the hsp70 genes of Drosophlia was found in the
proventriculus of the late third-instar larval gut (Lakhotia and Prasanth, 2002).
A decrease in expression in response to heat shock for a subset of genes has been
noted previously (Leemans et al., 2000) and our analysis confirms this observation.
Genes which are negatively regulated in our model could also be separated into the two
classes of fast and slow heat shock response genes. However, our analysis was not
designed to properly distinguish if a slow heat shock response gene reaches its lowest
level of expression only during the recovery period or if this is a result of the extended
heat shock time of 1 hour (as opposed to 30 minutes) used in the experiments. On the
other hand, it became obvious that genes with dramatically decreased levels of
expression after the short heat shock had already restored their baseline level of
expression (as defined by the non heat shock control) after recovery in the initial
experiment (Ih hs + 30 min RT). Examples would be the genes wunen, the eukaryotic-
initiation-factor-4G or the uncharacterised genes CG3271 and CGI5168.
These observation are purely based on the analysis of the microarray data and no
in vivo validation has been done to confirm these theory. However, this concept raises
the question if genes that are controlled by Mirror exhibit a similar transcriptional
behaviour (mRNA turnover). To answer this question and in analogy to the above it
appears useful to distinguish between “slow and fast Mirror response” genes.
6.2. The “fast Mirror response” genes
6.2.1. alpha tubulin at84B is positively regulated by Mirror
The most impressive example so far of an upregulated “fast Mirror response”
140

Chapter 6: Short heat shock
gene is alpha tubulin at 84B {a-Tuh84E). A massive increase of a-Tub84B transcript
level is observed in the short heat shocked pKBl2-mirror embryos compared to the
control. The histogram demonstrates nicely that the induced differences in gene
expression for a-Tub84B are almost restored to normal levels after recovery in pKB12-
mirror embryos. However, levels of expression are still high in pKB/yw-mirror embryos
even after the long heat shock treatment. Many genes appear to show a longer time of
elevated expression levels in the pKB/yw-mirror background compared to pKB 12-
m irror embryos. We do not understand why this is so, however one possible
explanation is that by outcrossing to yw embryos we have rid the chromosomes of
accumulated mutations that protect the fly agains the deleterious effects of ectopic
mirror expression, yw embryos do not exhibit increased levels of a-Tub84B transcript
(fig.6.4.) There are four alpha tubulin genes in Drosphila, each encoding a distinct
isoform. The major isoforms are a-Tub84B and a-Tub84D, which are expressed at all
developmental stages and encode proteins that differ at only 2 of 450 residues. The
constitutive isotype encoded by these genes shares 96% sequence identity with
abundant mammalian tubulins. Expression of the a-Tub85e gene is restricted to the
testes and a subset of cells in the peripheral nervous system (Bo and Wensink, 1989)
and a-Tub67c is present only in ovaries and very early embryos (Kalfayan and
Wensink, 1982). Alpha tubulins are found dimerised with beta tubulins for which there
are also four isoforms in flies. The alpha-beta tubulin heterodimer is the functional form
of tubulin in the cell. The heterodimer is the structural subunit of microtubules, which
compose the cytoskeleton. The microtubules are essential for intracellular transport and
cell division in all eukaryotes (Nogales et al., 1998). Alpha tubulin is found in two
isotypes in the embryo, a maternally supplied form, which is synthesised in the ovary
and deposited in the oocyte and a highly conserved constitutive isotype that is both
141

Chapter 6: Short heat shock
maternally loaded and zygotically synthesised (Theurkauf, 1992). Both isotypes are
abundant and assemble into all classes of microtubules from the syncytial blastoderm
stage until completion of germ band retraction. During later stages of development the
maternal isotype is retained in the ventral midline and in the developing CNS while the
expression throughout the embryo fades. Expression of the constitutive isotype remains
high in all tissues during development. However, it is in particular enriched in the
esophagus, the epidermis and most strikingly at the segment boundary. The constitutive
isotype appears to be more stable than the maternal isotype.
It has been shown that mRNA encoding the maternal isotype and the maternally
supplied constitutive isotype are degraded before blastoderm cellularisation (Matthews
et al., 1990; Theurkauf, 1992). The perdurance of the proteins they encode was analysed
using isotype specific antibodies. Both maternally provided isotypes are still detected
between 15-18h AEL. Expression of the zygotically expressed constitutive isotype
peaks between 9-15h AEL. Therefore, the alpha tubulin isotypes present in the embryo
change during embryogenesis from a mixture of maternal and constitutive isotypes to
primarily the zygotically expressed constitutive isotype (Theurkauf, 1992).
To investigate if over-expression of Mirror indeed induces high levels of a-
Tub84B in vivo, antibody stainings using a mammalian anti-tubulin antibody (DM1 A,
Sigma) were carried out. This antibody cross reacts with both tubulin isoforms, a-
Tub84B and a-Tub84D. Nevertheless, the accumulation of the a-Tub84B is easily
recognisable in the ventral midline and at the segmental border (fig.6.5.A). Ectopic
expression of a-Tub84B is observed in embryos misexpressing Mirror under UAS
control using various Gal4 enhancer trap lines (fig.6.5.B-D). Remarkable is the strong
staining around the midline (fig.6.5.B+C) and in the entire neuroectoderm (fig.6.5.D) in
embryos around stage 11-12. In addition, the effect of over-expressing another member
142

Chapter 6: Short heat shock
8000
4000
y/
\0
Figure 6.4. alphaTuh S4B, a positive regulated fast Mirror response gene. Note the dramatic increase in
the signal intensity for alpha Tub 84B after the short hs in pK B I2-m irror embryos. This difference in
expression is almost restored to normal levels after the recovery period in the long hs experiment.
pK B /yw -m irror embryos still exhibit high levels o f transcript after recovery. Short hs = 30 min heat
shock, long hs = Ih heat shock + 30 minutes recovery at RT, n=3 for each microarray condition.
p rd - . \ r a
p rd - . \ r akr-niirr
Figure 6.5. Anti-alpha tubulin staining in germ band extended embryos. A ) W ild type embryo showing
ubiquitous staining with enrichment in the ventral midline and at the segmental border (arrow). B+C) Kr-
Gal4xUAS-m/rror embryos show abberrant staining around the ventral midline. Note the broader midline
in C than in wild type embryos. D) sca-G a\4xl]A S-m irror embryo prove enrichment o f alpha tubulin
specifically in the ventral neuroectoderm. E+F) Ectopic expression o f Araucan under UAS control using a
prd-G ai4 driver also results in misexpression o f alpha tubulin around the midline. All embryos are ventral
view s with anterior to the right.
i t s

Chapter 6: Short heat shock
of the Iroquois complex was assessed driving \]h^-araucan with a prd-GPAA enhancer
trap line. Similar to Mirror, Araucan can also induce abnormal high levels of alpha
tubulin in the embryo (fig.6.5.E+F).
These results are remarkable as they proved for the first time that Mirror can act
as a transcriptional activator. It should also be mentioned that alpha tubulin at 84B is
the first gene identified and analysed after using the 30 minutes heat shock subtraction
model. This could suggest that this new approach to identify downstream targets of
Mirror might be more sensitive and biologically relevant than the previous attempts.
However, to validate a-Tub84B for consideration as a potential direct Mirror target
gene, mutant analysis should reveal its misexpression in the absence of Mirror. To
assess changes in expression of a ubiquitously expressed gene with a strong maternal
contribution, which in addition encodes an extremely stable protein is a priori difficult.
Nevertheless, the analysis of mirror mutant embryos revealed the most outstanding
results to this point. As mentioned above, both isotypes of a-Tub84B are ubiquitously
expressed in the embryo. In addition, the zygotically synthesised isotype is found
enriched at the segmental border whereas the maternally supplied form is strongly
expressed in the ventral midline. Interestingly Alpha tubulin staining appears to be lost
at the segmental border (fig.6.6. compare A+B with C+D) and strong staining is
observed in a single cell within each segment (arrows in fig.6.6.B+D). Both isotypes are
expressed in the developing CNS of the wild type embryo (fig.6.6.E+G). Severe defects
in the ventral nerve cord were observed in Iroquois mutant embryos (fig.6.6.F) and the
CNS of mirror mutant embryos often give the impression of being stained less intensly
and clearly exhibits the loss of longitudinal connections (fig.6.6.H).
144

Chapter 6: Short heat shock
Figure 6.6. Anti-alpha tubulin staining is altered in m irror loss-of-function embryos. A+C) W ild type
embryos showing ubiquitous alpha tubulin staining with enrichment at the segmental border at stage 14.
B+D) This staining is not detected in m irror mutant embryos. In addition, each segment seem to
accumulate alpha tubulin strongly in one or two cells per segment (arrows in B+D) which are not visible
in wild type embryos. E+G) Alpha tubulin is detected strongly in the CNS o f a wild type embryo at stage
16. II Iroquois mutant embryo exhibit dramatic defects in the ventral nerve cord. H) CNS staining in
m irror mutant embryos at stage 16 seems less intense and exhibit gaps in the longitudinal connections
(arrow). E-H are ventral view s o f stage 16 em lxyos. A -D are lateral view s o f stage 14 embryos with
anterior to the right and dorsal up.
145

Chapter 6: Short heat shock
Altogether, these results are particular intriguing as it has been hypothesised that
Mirror might play a role in refining the segmental border (McNeill et al., 1997).
Costaining with Engrailed or DAPI should be carried out to assure that the comparison
is truly made within the same focal plane in wild type and mirror mutant embryos. To
further investigate the relationship between Mirror and a-Tub84B, in situ hybridisation
studies using a a-Tub84B riboprobe which matches specifically the 3’UTR of the
constitutive isotype could be used. This should generate results with less background
staining to investigate the lack of a~Tub84B at the segmental border and help to
determine if Mirror regulates a-Tub84B in vivo.
6.2.2. dalmatian, CG8066 and CG8013 are positively regulated
by Mirror
The term “fast Mirror response” can also be attributed to the genes dalmatian
and many uncharacterised genes such as CG8066, CG8013 (fig.6.7.). The analysis of
these positively regulated candidate genes hasn’t been carried out yet. However, the
reported expression pattern of the gene dalmatian (Prokopenko et al., 2000) is
promising as it seems to overlap with mirror in the posterior spiracles, in the brain, the
ventral nerve cord and in the PNS.
The sensory neurons of the embryonic PNS can be divided into two major
classes: type I neurons with single dendrites and type II neurons with multiple dendrites.
Type I sensory organs are further classified as external sensory organs, which secrete
cuticular structures from the larval epidermis that detect mechanical or chemical signals
from the environment and as internal chordotonal organs, which form internal
attachments to the larval cuticle that function as stretch receptors. Type II sense neurons
146

Chapter 6: Short heat shock
dalmatian
1400
> 1200
1 1000Bc 800
1 600O)« 400>< 200
0
/ / / / /CG8066
2.2>w<
10009008007006 005 004 003 00200100
0
........................... !| J C _ 1
iL 1
11
------------------- T----- A ----1
. A !
f\0 * / «
/ /
CG8013
2
<
10009008007006005004 00300200100
0
/ y ,\0 9°'y
y
F igure 6,7. The upregulated fast Mirror response genes. The histograms show 3 representative genes of
this group {dalmatian and the unknown genes CG8066, CG8013). Characteristic for the fast Mirror
response genes is that their level o f expression increased during the short heat shock in pK B 12-m irror
embryos compared to the control and that they have a clear tendency o f restoring baseline levels after
recovery. This is most likely what happened for C G 8066 in pK B/yw long hs embryos. Short hs = 30 min
heat shock, long hs = Ih heat shock + 30 minutes recovery at RT, n=3 for each microarray condition.
147

Chapter 6: Short heat shock
are also internal sensory organs and may function as touch- or proprioreceptors
(Bodmer and Jan, 1987; Ghysen and Dambly-Chaudiere, 1986).
Interestingly, removal of the Iroquois complex in the embryo causes aberrant
development of the embryonic PNS, very similar to what has been reported to be a
dalmatian phenotype. Embryos which lack dalmatian function are characterised by
infrequent loss of neurons within the PNS, occasional pathfmding defects and small
round cells that stain darkly with the sensory neuron specific antibody 22C10 (fig.6.8 E;
taken from (Salzberg et a i, 1997). The sensory neurons of the embryonic PNS are
localised in a dorsal (d), a lateral (1) and two ventral (v and v ') clusters as shown by
22C10 staining of stage 16 embryos (fig.6.8.A+C). Individual neurons within a cluster
or entire clusters of sensory neurons are missing in Iroquois mutant embryos.
Misguided axons crossing the segmental border are also seen in these embryos
(fig.6.8.B). Most similar to the described dalmatian phenotype is the appearance of
small round cells without any protrusions (fig.6.8.D).
Another observation should be mentioned in this context. Sensory organ identity
is normally conferred by the bHLH proneural gene atonal or the genes of the AS-C
which act via the neural selector gene cut, a homeobox encoding transcription factor
(Bodmer et a i, 1987). However, a particular neuron of the type II class, the ddal neuron
with extensive dendritic arborizations, contained in the dorsal cluster is formed
independently of this pathway, but requires the activity of the proneural gene absent
MD neurons and olfactory sensilla (amos), a bHLH transcription factor (Brewster et al.,
2001; Goulding et a i, 2000). This cell co-expresses the homeodomain containing
segmentation gene engrailed and the POU domain containing genes pdm l and pdm2.
Indeed, the ddal neuron is easily recognisable by its isolated expression of
Engrailed within each segment (fig.6.8.F). Interestingly, Iroquois mutant embryos lack
148

Chapier 6: Short heal shock
f I %
Figure 6.8. Iroquois mutant embryos exhibit a dalmatian phenotype. A) A wild type embryo at stage 16
stained with M Ah 22C10 shows the well organised dorsal (d), lateral (1) and ventral (v, \ ' ) clusters o f
sensory neurons o f the PNS. B) Iro' mutant embryo lack individual (black arrows) and whole clusters o f
lateral and dorsal sensory neurons (black dotted Une). The clusters are generally less organised and axons
cross occasionally the segmental border (white arrow). Q Higher magnification o f abdominal segments
o f the embryo shown in A and an Iro (D) and dalmatian (H) mutant embryo. D) Small round cells
without axon (arrow) are found in the PNS o f an Iro mutants. E) 22C 10 staining in a dalmatian mutant
embryo to illustrate the small dark rounded cells (figure E is taken from Salzberg e t al„ 1997). F)
Engrailed staining in wt embryo o f stage 16 labels a single cell within each segment, the ddal neuron. G)
These cells are only occasionally detected in embryos which lack Mirror function (white arrow). Anterior
is to the right and dorsal is up in all figures.

Chapter 6: Short, heat shock
expression of Engrailed in this cell, (compare fig.6.8.F with G). If this is also a feature
of the dalmatian phenotype is not known.
Finally, the observed defects in the PNS of mirror loss-of-function embryos
mimic the described dalmatian phenotype. This would be consistent with the
assumption that dalmatian is positively regulated by Mirror. It would be attractive to
carry out an in vivo analysis in Mirror over-expressing and mutant embryos to further
investigate if Mirror acts indeed upstream of dalmatian. Although it is not clear if the
lack of engrailed expression in the ddal neuron of the Iroquois mutants is due to a
failure to activate dalmatian or if this particular cell requires Mirror directly to express
engrailed, this “one-cell” phenotype could provide a relative simple system to analyse
Mirror function in the PNS. In this context, it would be an alternative question to ask, if
Mirror is normally needed to repress atonal in this cell, which in turn would lead to
activity of engrailed and the proneural gene amos and to confer the ddal identity.
6.2.3. fringe, CG2467 and CG10650 are negatively regulated by
Mirror
The new gene expression analysis of Mirror over-expressing embryos using the
short heat shock protocol without recovery period revealed a two fold decrease for the
glycosyltransferase encoding gene fringe compared to the non heat shock pKB 12-mirror
control embryos. Although the expression levels of fringe in the initial long heat shock
experiment were lower in pKB12-mirror embryos than in the control, it was not
statistically significant, fringe transcript levels remain remarkably unchanged in the yw
embryos at the different conditions used (fig.6.9.). The gene fringe has been proposed
150

Chapter 6; Short heat shock
300
fringeg 260
I 200
« 150f 100
250
CG2467 £: 200
r 150m§, 100
y y" y \0'^ .cy
CGI0650a #
yy .■<?
s° V '/Figure 6.9. Genes with a fast response to Mirror over-expression. Fringe, a known target o f Mirror
activity, is significantly downregulated after the short heat shock in pK B l2 -m irro r embryos but not in the
controls and less significant after the long hs. Many unknown genes, such as C G 2467 and C G J0650 also
exhibit a dramatic decrease in response to ectopic Mirror. All these genes have a tendency to restore their
level o f expression during the recovery period. Short hs = 30 min heat shock, long hs = Ih heat shock +
30 minutes recovery at RT, n=3 for each microarray condition.
151

Chapter 6: Short heat shock
to be negatively regulated by Mirror in the Drosophila eye (Dominguez and de Celis,
1998; Yang et al., 1999) and in the oocyte (Jordan et al., 2000; Zhao et al., 2000) where
it modulates Notch activity. The decreased levels offringe expression within 30 minutes
would support the theory that fringe is a direct Mirror target.
Many more, mostly uncharacterised “fast Mirror response” genes were also
identified in the new approach. Characteristic for these genes is that they seem to
display a tendency to restore their baseline level of expression in the long heat shock
experiment after the recovery period. For example the unknown genes CG2467,
CG10650 (fig.6.9) and also the genes CG7526 or CG5699 (not shown) show an even
more dramatic decrease in expression than measured for fringe in short heat shocked
pKBl2-mirror embryos than in the non heat shock control or after the long heat shock
treatment. The ongoing in vivo characterisation of these genes in mutant embryos will
clarify if their expression is really regulated by Mirror.
6.3. The “slow Mirror response” genes
6.3.1. carnation, plutonium and CG4586 are positively regulated
by Mirror
An increase in the relative expression levels which would be characteristic for
“slow Mirror response” genes is observed in pKB 12-mirror embryos for example for
the genes carnation, plutonium or the unknown gene CG4586 (fig.6.10). However,
these genes don’t appear to have consistently altered levels of transcription in long heat
shocked pKB/yw-mirror embryos. These candidates are shown here only to illustrate the
presence of genes in the analysis which would fall under the proposed group. None of
these genes has been investigated further with respect to Mirror function.
152

C hapter 6: Short heat shock
carnation
plutonium
(0csc12>(0
<
350
300
250
200
150
100
50
0
.2>CO
<
400
350
300
250
200150
10050
0
j i
///y/yyCG4586
1200
^ 1000 CO
» 800
CQ
#<
600
4 0 0
200
0
/
I
e f / / /Figure 6.10. The upregulated slow Mirror response genes. The relative expression levels are shown for
the genes plutonium, carnation and the unkown gene C G4586. All three genes show highest levels of
expression in pKB 12-m irror long hs embryos. Levels o f expression in the yw background does not differ
significantly. PKB/yw-mirror embryos don’t show any significant upregulation in the experiment. All
embryos used for the microarray analysis were about embryonic stage 11. Short hs = 30 min heat shock,
long hs = 1 h heat shock + 30 minutes recovery at RT, n=3 for each microarray condition.
153

Chapter 6: Short heat shock
6.3.2. CG15922, CG6900 and gleeful are negatively regulated by
Mirror
Last, there are negatively regulated genes exhibiting a profile, which would
classify them within the group of the “slow Mirror response” genes. Members of this
group have lowest levels of transcription in long hs Mirror over-expressing embryos. A
common feature of this group is that the repressor activity of Mirror is already visible in
short hs Mirror overexpressing embryos. Three representative genes, CG15922,
CG6900 and gleeful should be mentioned for this group (fig.6.11). The in vivo
validation for the uncharacterised gene CGI5922 is currently under way, as well as for
the recently described gene CG4677, which encodes a zinc finger transcription factor
with high sequence similarity to vertebrate Gli proteins, therefore named gleeful (gfl).
gleeful is transcribed in the visceral and somatic mesoderm at stages 11-13 where it acts
downstream of the transcription factor twist to induce somatic muscle development.
Ectopic expression of gleeful in the neuroectoderm causes cells to express a muscle
marker (Furlong et al., 2001). The gene CG6900 is ubiquitously expressed in the wild
type embryo and was eliminated from further analysis.
Finally the gap gene KrUppel (Kr), best known for its segmentation function in
the early embryo, is also found within this group as a potential Mirror target. The
investigation to elucidate whether Mirror regulates Krüppel expression in the embryo is
provided in the next chapter.
154

Chapter 6; Short heat shock
CGI5922
1200
^ 1000 CO
i 8 0 0
« 6 0 0
I* 4 0 0
< 200
0
CG6900
CG4677
(gleeful)
COcBc1O)*
1200 r -
1000
8 0 0
6 0 0
4 0 0
200
0
1 X
Ù/ / /
4 0 0
350
CO 3 0 0c2 25 0c
20 0cO ) 150CO> 100<
50
0
1 _x_
1 % .
^ >f& >(&><&
^ " 9' .■y yFigure 6.11. Downregulated slow Mirror response genes. Three representative genes o f this group are
shown in the histogram, namely the unknown genes CG 15922,C G 6900 and the gene C G 4677 (recently
named gleeful). (Characteristic for the slow Mirror response genes is that their relative amounts o f
transcripts reaches lowest levels in long heat shock Mirror over-exf»-essing embryos. ITie transcript levels
in yw hs embryos remains stable under different conditions used. Short hs = 30 min heat shock, long hs =
Ih heat shock + 30 minutes recovery at R'f, n=3 for each microarray condition.
155

Chapter 6: Short heat shock
Discussion
Heat shock is known to induce the expression of an evolutionary conserved
family of genes encoding the heat shock proteins (Lindquist and Craig, 1988; Nover and
Scharf, 1997). These genes are quickly transcribed in order to produce the heat shock
proteins which function to protect the eukaryotic cell in response to stress inducing
agents such as heat. In Drosophila, the promoter architecture of the gene hsp70 (and
probably other heat shock inducible genes) is primed for the activation under non heat
shock conditions. It is believed that the heat shock promoter sequences are, even before
induction, in a nucleosome-free open configuration and are occupied by several
transcription factors bound to GAGA and TATA box sequences including a RNA
polymerase II complex (Dahmus, 1994; reviewed in Lis, 1998). The hypo-
phosphorylated Pol II molecule has been shown to be in a transcriptionally engaged but
paused stage at the uninduced hsp70 promoter, where it produced short transcripts of
about 21-35 nucleotides (Rasmussen and Lis, 1993; Rougvie and Lis, 1988). It is the
hyper-phosphorylated form of the Pol II which is actively engaged in transcriptional
elongation (Dahmus, 1994; Lu et al., 1991). In addition to the classical heat shock genes
relatively few other genes have been identified which change their relative level of
transcription in response to heat shock (Leemans et al., 2000). By omitting the recovery
period and processing the embryos directly after the heat shock, it became evident that
by far more genes alter their levels of expression in Drosophila embryos when exposed
to a temperature of 36°C. Our analysis identified “slow and fast” heat shock response
genes. Fast heat shock response genes exhibit a significant increase or decrease after 30
minutes of heat shock treatment compared to non heat shocked embryos. Fast heat
shock response genes return to their baseline level of expression after the 30 minutes
recovery, despite the longer (one hour) heat shock treatment and extended accumulation
156

Chapter 6: Short heat shock
of their transcripts. The recovery period was introduced to allow this downregulation of
highly abundant heat shock genes. In contrast, slow heat shock response genes exhibit
their highest levels of transcription only after the long heat shock treatment. Similarly,
potential Mirror target genes can also be classified into slow and fast response genes.
Obviously, it can not be distinguished if the observed “slow” increase (or decrease) in
relative levels of transcripts is indeed due to slow transcriptional activation (or
inhibition) or if these transcripts are more stable than others and accumulated during the
longer heat shock period. Nevertheless, this discrimination illustrates the different
transcriptional behaviour observed for potential Mirror target genes.
Only very few genes from this list have been analysed to date and it is therefore
not possible to definitely conclude if this new approach has indeed been more
successful than the previous attempt. Although the short heat shock protocol hasn’t been
applied to pKB/yw-m/rror embryos, the initial long heat shock results were used to
confirm or disprove if a gene can be classified within the group of up- or down
regulated slow Mirror response genes. To further eliminate false positives, it would
certainly be useful to implement the analysis with data from short heat shocked embryos
of the pKB/yw-m/>ror background. Nevertheless, the combination of different
experimental conditions used in three different genetic backgrounds already provided a
complex data set with several independent intra- and intergenetic controls for Mirror
activity. As a high number of false positives is a general problem of the microarray
analysis, it seems appropriate to apply a stringent method for the identification of
potential Mirror downstream targets. By concentrating only on genes which haven’t
altered their levels of transcription in the yw background, but show consistent increased
or decreased expression in Mirror over-expressing embryos, I was hoping to eliminate
false candidate genes. This raises the question if potential Mirror targets could get lost
157

Chapter 6: Short heat shock
in the analysis. This is likely to be the case, if for example, Mirror has regulatory
function on a gene which also changes its level of transcription in response to heat
shock. Another interesting group of genes, deserves consideration but has not been short
listed for a subsequent in vivo analysis in mirror mutant or over-expressing embryos.
This group consists of genes, for which opposite levels of transcription were detected in
short heat shocked pKB12-wz>ror embryos compared to long heat shocked pKB12-
mirror embryos. The genes gooseberry or glial cells missing would be part of such a
group. Both genes have significant increased levels of transcript in short heat shocked
pKB 12-mirror embryos but were downregulated in long heat shock experiment. This
result is interesting because it would be consistent with the earlier observation that
mirror mutant embryos have decreased levels of gem (or Repo) expression (see chapter
5, fig.5.6+5.7). The fact that over-expression of mirror using the Gal4/UAS system does
not lead to induction of gem expression in vivo might be due to an inhibitory effect on
transcription by large amounts of Mirror protein and could explain why the initial
microarray analysis using long heat shock embryos detected lower levels of gem
transcript.
In addition to the model described above, a preliminary micro array analysis
using Iroquois mutants embryos compared to an “appropriate control” has been carried
out. To do so, a heterozygous Iroquois mutant stock was used, which has a balancer
chromosome carrying a transgene encoding green fluorescent protein (GFP).
Iroquois/GFP balancer collections contain a mixed populations of embryos: one quarter
Iro' homozygotes lacking GFP, half heterozygotes with one copy of GFP, and one
quarter homozygotes for the balancer chromosome with two copies of GFP.
Homozygous Iro' mutant embryos were separated from their siblings using an embryo
sorter and compared to the heterozygotes. Putative homozygous Iro' embryos were
158

Chapter 6: Short heat shock
assessed by in situ hybridisation using a mirror riboprobe. About 90% of the selected
embryos were mirror minus. The earliest time GFP is detectable in balancer embryos is
at stage 9-10 (Casso et al., 2000). Several identified candidate genes from the modified
heat shock subtraction model showed consistent reverse regulated expression in the
absence of Mirror. For example the negatively regulated genes CG2467, CGI0650,
CG7526 and CG5699 from the fast Mirror response group revealed increased levels of
expression in the mutant background. It should also be mentioned that the signal
intensity for gem transcript is reduced in these embryos. This would again be consistent
with the notion of gem being a target of Mirror activity. However, these results would
need to be confirmed by generating replicates of this microarray analysis in order to be
able to apply statistical methods to eliminate false positives. Such a combinatory
microarray analysis of over-expression and loss-of-function using tightly staged
embryos has been used to identify genes which act downstream of the bHLH
transcription factor twist during mesoderm development of the Drosophila embryo. The
bioinformatic analysis revealed that from the 643 genes found to be significantly
different in comparison to stage matched wild type embryos, 81 genes were consistently
up- or down regulated in Twist over-expressing and mutant embryos. The subsequent in
vivo analysis of these genes proved a role for the genes gleeful (gfl) and many
uncharacterised genes in the mesoderm (Furlong et al., 2001). However, it is not known
if these genes are directly regulated by Twist or if they are regulated by additional
factors further downstream in the pathway.
Consistent results in mirror mutant and over-expressing embryos were obtained
for the gene a-Tub84B. Even though tubulin is not the most attractive candidate of
Mirror activity, it would not be the first tubulin isoform which is regulated by a
homeodomain encoding protein. Indeed, it has been reported that beta 3 tubulin at 60D
159

Chapter 6: Short heat shock
{pTubôOD) is directly regulated by two homeodomain transcription factors in the
visceral mesoderm of the Drosophila embryo. The NK homeobox gene tinman has been
proposed to be required for the general activation of pSTubôOD in the visceral
mesoderm (Gasch et al., 1989; Kremser et al., 1999), while the NK homeobox gene
bagpipe (bap) controls expression of pSTubôOD in metameric patches of the trunk
visceral mesoderm. Here, bap activates the FoxF fork head domain gene biniou (bin),
which also regulates expression of pSTubôOD (Zafffan and Frasch, 2002). bap and bin
are coexpressed with the ^3Tub60D in the trunk visceral mesoderm and it has been
shown that they bind to a specific sequence within the intronic enhancer, termed vml.
The combined activity of Bap and Bin is required to mediate enhancer activation
specifically in this area (Zafffan and Frasch, 2002). Unlike bap, which is expressed only
transiently, bin expression continues in the gut musculature until it is fully differentiated
(Zaffran et al., 2001). PTubôOD is known to contribute to the cytoarchitecture of
visceral mesoderm cells, which is essential for the proper functioning of the gut
(Dettman et a l, 1996). In addition, the vml element contains binding sites for the
homeotic factor Ubx, which is involved in regulating the enhancer activity within the
parasegments 6 and 7 (Gasch et a l, 1989; Kremser et a l, 1999). Thus, PTubôOD is
regulated by the combined and independent activity of several transcription factors at
different sites during embryonic development.
160

Chapter 6: Short heat shock
161

Chapter 7: Mirror confers neiirohlast identity^
CHAPTER 7
Mirror confers neuroblast identity
7.1. Mirror regulates Krüppe/expression in the
Drosophila embryo
7.1.1. Introduction to Krüppel
The gap gene Krüppel {Kr), best known for its segmentation function in the
early embryo encodes a zinc finger transcription factor. Krüppel is initially expressed in
a cap at the posterior end and in a broad band that covers about four segment primordia
in the centre of the blastoderm of the embryo (Wieschaus et al., 1984b). The lack of
Krüppel activity causes the deletion of this middle region as well as derivatives of more
posterior segment primordia, which creates a gap of approximately eight segments in
the larval body (Jackie et al., 1988). Gap genes like hunchback, Krüppel, or knirps are
among the earliest genes expressed during development (Nüsslein-Volhard and
Wieschaus, 1980). Their overlapping expression is controlled by the matemal-effect
genes in order to subdivide the embryo along the anterior-posterior axis which has been
studied extensively (Gaul and Jackie, 1987; Gaul et al., 1987; Hoch et al., 1992;
Hülskamp et al., 1990; Jackie et al., 1988; Jacob et al., 1991). AU gap genes are
expressed in specific domains in the blastoderm and all encode transcription factors,
mostly with a zinc finger DNA binding motif (reviewed in Pankratz and Jackie, 1993).
Gap genes induce subsequent expression of the pair-rule genes in double segment
162

Chapter 7: Mirror confers neurohiast identity
intervals resulting in seven symmetrical stripes, which in turn leads to the delineation of
individual segments by the segment polarity genes. Within the segmentation gene
cascade, Krüppel protein functions mainly as a repressor of other gap- and pair-rule
genes (Pankratz and Jackie, 1993). In vitro experiments have shown that Krüppel
protein forms a homodimer when expressed in high concentration. The homodimer acts
exclusively as a transcriptional repressor, whereas the Krüppel monomer acts as a
transcriptional activator, both, in vitro and in Drosophila tissue culture assays. Based on
this observation it has been proposed that Krüppel has repressor activity in the syncytial
blastoderm of the embryo. Outside this central region, where Krüppel expression
gradually decreases, it may function as a transcriptional activator (Sauer and Jackie,
1993). Krüppel, like other gap genes, is also involved in various developmental
processes beyond segmentation. After blastoderm stage Krüppel is involved in the
formation of the Malpighian tubules (Gaul and Weigel, 1990; Hoch and Jackie, 1998;
Knipple et al., 1985), the Bolwig larval photoreceptor organ (Schmucker et a l, 1992), a
specific subset of muscle founder cells and muscles (Ruiz-Gomez et al., 1997) and in
neurogenesis (Hoch et al., 1990; Isshiki et al., 2001; Jacob et al., 1991). Krüppel is also
expressed postembryonically where it plays a role in the expression of fat-body specific
genes in the third-instar larva (Hoshizaki, 1994).
The Krüppel gene consists of two exons which are separated by a 372 bp intron
(Rosenberg et al., 1986; Gaul et al., 1987). Moreover, multiple and independent cis-
acting control elements were identified within 18kb of Krüppel upstream DNA fused to
a lacZ reporter gene, which drive expression at 10 different locations during
embryogenesis. At least two different regions, named NSl and NS2, have been
identified which are necessary for expression of the lacZ transgene in a Krüppel-Wke,
pattern in the developing nervous system (Hoch et al., 1990; Jacob et a l, 1991). The
163

Chapter 7: Mirror confers neurohiast identity
complexity of the expression pattern was further increased when the Krüppel intron was
present in the lacZ constructs. Thus it is suspected that a regulatory sequence for
expression in the CNS lies within the Krüppel intron (Hoch et al., 1990).
7.1.2. Krüppel expression in the developing CNS
The early Krüppel expression in the syncytial blastoderm is well known and its
function as a gap gene has been extensively studied. Although it has been reported that
Krüppel is expressed in various tissues during embryonic development, its detailed
expression pattern with respect to the CNS is less well documented. In order to gain
further insights into the expression pattern of Krüppel during different stages of
development, antibody stainings were carried out on embryos.
By the start of germ band elongation, as the initial expression in the blastoderm
decreases, Krüppel expression expands ventrally into the neuroectoderm. When the first
proneural clusters become resolved at stage 8, Krüppel protein is detected in the nucleus
of cells of the neuroectoderm (flg.7.1.A). At stage 9, a subset of CNS midline cells
(fig.T.l.B), and shortly after, at stage 10, a single neuroblast, namely NB6-1, ceases its
expression of Krüppel protein (fig.T.l.C). All these cells now express Castor, another
zinc finger protein (also known as ming) (Cui and Doe, 1992; Mellerick et al., 1992).
The metameric structure of the embryo also becomes visible, which indicates the
formation of the parasegments. When the germ band is fully extended at stage 11,
Krüppel expression is absent in large segmentally repeated patches of delaminating
neuroblasts (fig.7.1.D). These large domains become restricted while the germ band is
retracting at stage 12 and segments are formed (fig.7.1.E). During subsequent stages in
development, Krüppel is expressed in clusters of cells in each segment, separated from
each other by a 1 -2 cell wide interval, which corresponds to the segmental border.
164

Chapter 7: Mirror confers neurohiast identity'
Figure 7.1. Krüppel expression pattern in the forming CNS. Immunostaining with anti-KrOppel (red) in
wild type embryos from stage 8-17. (A) At stage 8 Krüppel protein is found in almost every nucleus o f
cells o f the neuroectoderm. B) At stage 9, several midline precursor cells lose Krüppel expression (white
arrow). C) Neuroblast 6-1 has lost Krüppel expression at stage 10, giving rise to the T-shape hole within
the Krüppel expressing Neuroblasts. D-F) From stage 11-14 the Krüppel expression pattern is refined as
neuroblasts lose expression in the ventral nerve cord. G+H) After stage 14 the CNS condenses and the
transient hole in the Krüppel expression domains closes again. All panels are ventral views with anterior
to the right.
ILS

Chapter 7: Mirror confers neurohiast identity
The Krüppel-free patches are still visible at the onset of head involution at stage
14 (fig.T.l.F) and become spatially even more restricted after the end of dorsal closure
towards stage 16 (fig.T.l.G). After full condensation of the ventral nerve cord, no
Krüppel-free domains are visible within the CNS (fig.7.1.H).
The antibody staining carried out to detect Krüppel protein in the developing
CNS of the embryo demonstrates that Krüppel expression is extremely complex and
changes rapidly at different stages of development. In addition, the spatio-temporal
expression of Krüppel is heterogeneous, that is, certain areas and cells accumulate
higher levels of protein than others and this varies with time (see fig.T.l.B). This
observation is not new and has been reported earlier (Romani et al., 1996).
Interestingly, several PEST domains have been identified within its protein sequence,
which have been associated with a rapid protein turnover (Jacob et al., 1991).
Furthermore it has been noted previously that KrUppel transcript at blastoderm stage
accumulates homogeneously, while the intensity of its protein staining in this domain is
most intense in the centre and fades gradually at the edges of the stripe (Knipple et al.,
1985). It has been suggested that the protein is posttranscriptionally regulated and a
regulatory region within the intron of the KrUppel transcript has been proposed to be
involved in such a process (Gaul et al., 1987). However, diffusion of the protein into the
syncitial environment could also explain why the Krüppel staining fades towards the
edges of the stripe.
7.1.3. Krüppel and Mirror are expressed in mutually exclusive
domains
Combining an in situ hybridisation using a mirror riboprobe with a Krüppel
166

Chapter 7: Mirror confers neurohiast identity
antibody staining revealed expression patterns in non-overlapping domains. At stage 11
mirror is expressed in a characteristic “maple leaf’ like shape (fig.7.2.A). Krüppel
protein is found exclusively outside this domain (fig.7.2.B). As the dark blue
diformazan/indigo precipitate might mask the fluorescent secondary antibody which
shows the Krüppel expression pattern, double antibody staining for both Mirror and
Krüppel protein were also carried out and showed the same result (fig.7.2.C+D).
In conclusion, it seems that Krüppel expression diminishes to the same extent as
Mirror expression expands in the delaminating neuroblasts, giving rise to a pattern in
which Mirror is surrounded by Krüppel expressing cells. This observation would be
consistent with the notion that Krüppel is negatively regulated by Mirror.
7.1.4. Over-expression of Mirror represses Krüppel in the
embryo
The microarray analysis using the heat shock subtraction model revealed that
heat shock induced over-expression of Mirror in the embryo, at embryonic stage 11,
reduces the amount of Krüppel transcript in a manner characteristic for “slow Mirror
response” genes. Note that the “baseline” level for Krüppel expression in non heat
shock embryos of the three different genetic backgrounds varies considerably,
underlining that the initial attempt to directly compare long heat shocked yw and long
heat shock pYdQ\2-mirror embryos was inappropriate. When each genetic background
is compared separately it becomes evident that the different experimental conditions
have no significant effect on the transcript levels of the Krüppel gene in yw embryos
(fig.7.3). Reduced levels of Krüppel transcript are detectable in short heat shocked
pKB 12-mirror embryos but lowest levels are found in Mirror over-expressing embryos
167

Chapter 7: Mirror confers neurohiast identity'
if.
Figure 7.2. Krüppel and Mirror are expressed in mutually exelusive domains. A) The in situ hybridisation
in a stage 11 embryo shows the characteristic mirror expression in delaminating neuroblasts in a “maple
lea f’ like shape. B) fhe same embryo as in A stained with an anti-KrUppel antibody (green). The
combination o f in situ hybridisation and antibody staining revealed that mirror (black) and Krüppel
(green) are expressed in reciprocal domains. C+D) Complementary expression patterns are also visible in
a stage 12 embryo double stained with anti-Mirror (red) and anti-Kriippel (green) antibody. Krüppel
expression only is show in C. Anterior is to the right in all panels. A+B are dorsal views and C+D are
ventral views.
l a

Chapter 7: M irror confers neurohiast identity
1600
« 800 c ___O) 600 CO
> 400
y
Figure 7.3. Over-expression o f Mirror reduces the level o f Krüppel transcripts in the embryo. The
histogram displays the average signal intensity measured for K rüppel cRNA in the course o f the
microarray analysis in three different genetic backgrounds subjected to heat shock or not. yw embryos
where used as a control and pKB and PKBYW embryos are two different fly strains containing a m irror
transgene under the control o f a heat shock promoter. The average signal intensity for K rüppel cRNA
remains almost unchanged during the different experimental conditions in yw embryos (blue). The signal
intensity o f Krüppel cRNA decreases in Mirror over-expressing embryos achieved by a short heat shock
impulse compared to the non hs embryos of the same genetic background and is even further reduced in
these embryos when hs for a longer period o f time (red). PKBYW embryos (yellow ) also exhibit a
reduction of Krüppel cRNA after the long heat shock treatment, limbryos were collected for 30 min on
apple juice plates at RT and processed 5'^ h A liL (approx. stage 11 ). Short hs = 30min at 36°C; long hs =
Ih at 36“C + 30 min RT. n=3 for each microarray condition.
P-tubulin-rmrror
Figure 7.4. Over-expression o f Mirror downregulates Krüppel expression in the embryo. A) Krüppel in
situ hybridisation in a w ild type embryo at stage 10. B) Expression o f K rüppel is reduced in an em lxyo
ubiquitously over-expressing Mirror at the same stage o f development.
W

Chapter 1: Mirror confers neurohiast identity
after the long heat shock procedure, including a 30 minute recovery period.
To validate in vivo if Krüppel is indeed downregulated in embryos which over
express Mirror, in situ hybridisation experiments were done. Ubiquitous over
expression of Mirror leads to severely reduced KrUppel expression in the embryo (fig.
7.4.).
In addition 5ca-Gal4xUAS-mz>ror embryos were analysed and double labelled
for Krüppel (red) and Castor (green). At embryonic stage 14 the majority of sca-
Gal4xUAS-/?7z>ror embryos exhibit severe morphological defects. The hemi-
neuromeres on both sides of the ventral midline are connected, if at all, only by a thin
stripe of KrUppel expressing cells (fig.7.5. compare A+D). KrUppel staining at this stage
might be generally weaker but is not significantly reduced. Castor staining in these
embryos reveals that the neurogenic regions are separated from each other by a large
gap (fig.7.5. compare B+E and C+F).
To sum up, KrUppel expression is severely reduced in ubiquitously Mirror over
expressing embryos. KrUppel antibody staining in 5ca-Gal4xUAS-mz>ror embryos,
appears abnormal but does not seem to be significantly reduced. Misexpression of
Mirror leads to severe morphological defects in the ventral nerve cord.
170

Chapter 7; M irror confers neurohiast identity
Figure 7.5. Over-expression o f Mirror alters Krüppel expression and leads to morphological defects in
the CNS. A-C) A wild type embryo o f stage 14 demonstrate that Castor (green) and Krüppel (red) are
expressed in overlapping domains in the CNS. D-F) A Mirror over-expressing embryo o f the genotype
sca-Gal4 x GAS-mirror exhibits dramatic deformations o f the developing CNS. Krüppel expression
appears weaker than in wild type and the domain above the ventral midline is thinned out (arrow in D,
compare with A). E) Castor staining reveals a large gap between the hemisegments o f the CNS. Ventral
views with anterior to the right.
171

Chapter 7: Mirror confers neurohiast identity
7.1.5. Krüppel is ectopically expressed in Mirror loss-of-
function embryos
The results described above showed that over-expression of Mirror can alter the
expression pattern of Krüppel in the embryo. The following sections will describe how
Krüppel is ectopically expressed in embryos which lack Mirror function. Initially, in
situ hybridisation was carried out using a Krüppel riboprobe in Iroquois mutant
embryos. About 10% of the embryos at embryonic stage 11 showed ectopic expression
of Krüppel transcript within the entire mirror expression domain (fig.7.6. compare
A+D). By far more frequent is the ectopic expression of Krüppel in only two cells per
segment (fig.7.6. compare B+C with E+F). However, these two cells are not seen
consistently in every segment at stage 11-12. Therefore, segments were counted to
assess the frequency of the “two-cell” phenotype in mutants between stage 11-12. It
became evident that all mutant embryos at this stage exhibited the “two-cell” phenotype
in at least two segments (see table 9). These two Krüppel expressing cells, most likely
neuroblasts, are never seen in wild type embryos at comparable stages of development.
In order to assess which neuroblast is ectopically expressing Krüppel, various
antibody stainings were used to label different neuroblast lineages. As Mirror is
normally expressed in a row of the most anterior neuroblast in a hemisegment but only
in one single neuroblast (NB6-1) positioned further posteriorly, it was speculated that
this could be the Krüppel misexpressing neuroblast. To test this hypothesis, Krüppel
expression was analysed in wild type (flg.7.7.A+B) and Iroquois mutant embryos at
stage 11 (fig.7.7.C+D) and costained with Engrailed (fig.7.7.B+D). According to the
Hyper Neuroblast Map from the Doe lab website. Engrailed is expressed at stage 11 in
NBl-2 at the most anterior position and more posteriorly in NB6-1, NB6-2, NB6-4,
NB7-1, NB7-2 and NB7-4 (fig.7.7.B+D). None of these neuroblasts seems to be the
172

Chapter 7: Mirror confers neurohiast identity
Figure 7.6. Krüppel is ectopically expressed in mirror loss-of-function embryos. Krüppel in situ
hybridisation in wt (A-C) and Iro' embryos (D-F) at stage 11. Ectopic Krüppel expression is occasionally
found in the entire Mirror expression domain (D) and more frequently in only two extra cells per segment
(E and F). In addition, the dorsal view demonstrates an overgrowth o f the amnioserosa in Iroquois'
embryos (compare B with E). Wt and Iro' embryo (C and F, respectively) shown from the ventral side.
Anterior is to the right in all panels.
Table. 9: Penetrance of the two cell phenotype in mirror LOF embryos at stage 11
No. of segments with ectopic Krüppel expression
Percentage (52 embryos counted in total)
I 0%II 15%
III 39%IV 27%
in more than V 19%
m

Chapter 7; Mirror confers neurohiast identity
misexpressing cell (fig.7.7.D). If Iroquois mutant embryos are analysed at earlier stages
the two ectopic Krüppel expressing neuroblasts are regularly found in every segment in
mirror loss-of-function (fig.7.7.E) but not in wild type embryos (inset in fig.7.7.E) at
late stage 10. Before that stage, no difference can be seen between mutant and wild type
embryos when stained for Krüppel protein, which is consistent with the onset of mirror
expression at stage 10. An antibody directed against Castor protein can be used as
marker for NBl-2, NB5-2, NB6-1, NB7-2, and NB7-4 at embryonic stage 10/11.
Costaining for Krüppel and Castor revealed no overlap in the Krüppel misexpressing
cell at this stage (fig.7.7.F). Thus, it was possible to exclude the possibility that NB6-1
or one of the other neuroblasts mentioned above has changed its molecular identity. It
also became clear that the Krüppel misexpressing neuroblasts were located towards the
anterior border of the segment. mirror-lacZ/TM3-lacZ mutant embryos, which express
Beta galactosidase in the mirror pattern, were costained using a Pgal and a Krüppel
antibody. These embryos also maintain Krüppel (red) expression in the two neuroblasts
(fig.7.8.A), which confirms that the phenotype is due to the lack of mirror and is not a
result of the combined loss-of-function of Ara, Caup and Mirr in the Iroquois mutant
embryos. The (3gal staining (green) is detected in mirror-lacZ mutant embryos in NBl-
1, NBl-2, NB2-2, NB2-5, NB3-2 at stage 10/11 (fig.7.8.B). When both channels are
displayed, it is evident that Krüppel is misexpressed in NB2-2 (fig.7.8.C).
Another phenotype of mirror loss-of-function observed in stage 11 embryos is
the previously mentioned overgrowth of the amnioserosa (see chapter 1). In this context
it is worth mentioning that Krüppel is heavily expressed in cells of the amnioserosa in
wild type embryos. Although Iroquois mutant embryos seem to express Krüppel
normally in this tissue, it is intriguing in connection with this phenotype. A double in
situ hybridsation analysis using a Krüppel (red, auto-fluorescent) and mirror (blue)
174
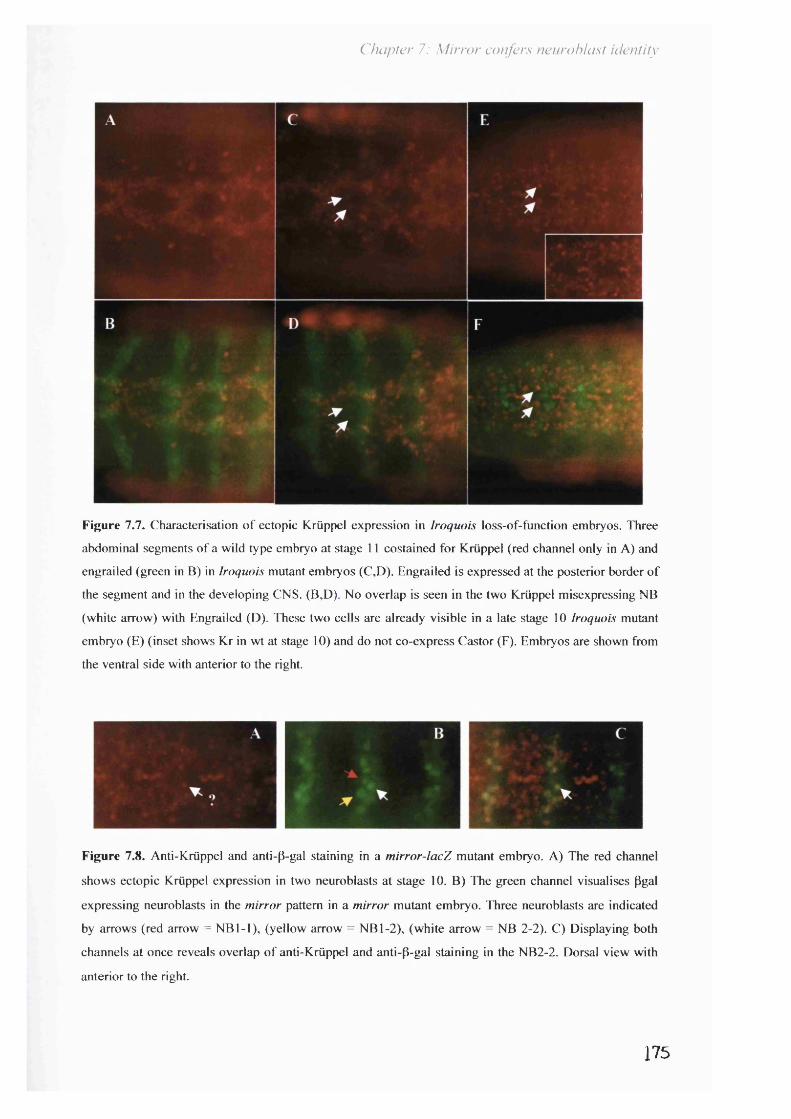
Chapter 7; Mirror confers neuroblast identity
Figure 7.7. Characterisation o f ectopic Kriippel expression in Iroquois loss-of-function embryos. Three
abdominal segments o f a wild type embryo at stage 11 costained for Kriippel (red channel only in A) and
engrailed (green in B) in Iroquois mutant embryos (C,D). Engrailed is expressed at the posterior border o f
the segment and in the developing CNS. (B,D). No overlap is seen in the two Kriippel misexpressing NB
(white arrow) with Engrailed (D). These two cells are already visible in a late stage 10 Iroquois mutant
embryo (E) (inset shows Kr in wt at stage 10) and do not co-express Castor (F). Embryos are shown from
the ventral side with anterior to the right.
Figure 7.8. Anti-Kriippel and anti-^-gal staining in a mirror-lacZ mutant embryo. A) The red channel
shows ectopic Kriippel expression in two neuroblasts at stage 10. B) The green channel visualises pgal
expressing neuroblasts in the mirror pattern in a mirror mutant embryo. Three neuroblasts are indicated
by arrows (red arrow = N B l-1), (yellow arrow = N B l-2), (white arrow = NB 2-2). C) Displaying both
channels at once reveals overlap o f anti-Kriippel and anti-^-gal staining in the NB2-2. Dorsal view with
anterior to the right.
175

Chapter 7: Mirror confers neurohiast identity
antisense probe on an Iroquois mutant collection between stages 11 and 12 also nicely
illustrates this observation. The mutant embryo can easily be distinguished by the lack
of mirror staining (fig.7.9.). The Kriippel expression pattern in the rest of the embryo is
not really clean due to the technically challenging nature of the experiment and the two
NB2-2 cells are not easy to see without additional excitation of the fluorocrome by UV
light.
In later stages of development, around stage 14, Kriippel expression in the CNS
displays a characteristic “bow-tie-like” pattern in wild type embryos. Kriippel
expressing cells crossing the ventral midline appear to connect the larger group of
Kriippel expressing cells of the two hemisegments (fig.7.10.A). These patches are found
in each segment and are usually separated by one or two cells where they are closest
(flg.7.10.A+D). Embryos which lack Mirror function exhibit an irregular Kriippel
expression in the neural tissue (flg.7.10.B+C) and in lateral clusters of muscle precursor
cells (not shown). The CNS phenotype is manifested by extra Kriippel expressing cells
reducing the space between and connecting the metameric domains with each other
(fig.7.10.E).
In conclusion, mutant analysis proved that there is ectopic Kriippel expression
within the Mirror expression domain. Although this does not always expand into the
entire domain, it is consistently seen between stages 10-12 in one neuroblast per
hemisgement, namely NB2-2. This neuroblast delaminates from a medial row 2 cluster
of neuroectodermal cells that also express huckebein Qiuck) at this stage (Bossing et a l,
1996a). Later in development NB2-2 is characterised by the expression of klumpfufi,
castor and seven-up (Broadus and Doe, 1995; Cui and Doe, 1992; Yang et al., 1997). In
addition it has been reported that NB2-2 is rw«^-positive as it delaminates and
throughout the rest of neurogenesis (Dormand and Brand, 1998). The progeny of these
cells normally will generate 2-3 motoneurons that innervate anterior lateral muscles as
176

Chapter 7: Mirror confers neurohiast identity
$
Figure 7.9. Double in situ hybridisation for mirror and Kriippel. The embryo on the top is a wild type and
the one on the bottom is an Iroquois mutant embryo, both around stage 11. Mirror is expressed in its
“maple leaf’ shape in delaminating neuroblasts and in cells o f the midline. Also visible is its expression
in cells o f the lateral epidermis (black). Kriippel (red) is detected in cells o f the compressed amnioserosa.
The Iroquois mutant embryo is recognisable by its overgrown amnioserosa (dotted line) marked by
Kriippel expressing cells (red).
177

Chapter 7: Mirror confers neurohiast identity
-4 s:
< ' • ' k f y: '* :' :- f 7 ' ' ^* 7' '
» '
Figure 7.10. Loss o f Mirror results in aberrant Kriippel expression in the forming CNS at stage 14. A)
Kriippel (red) expression appears in a segmented pattern on the ventral side o f the wild type embryo.
B+C) In Iroquois ' embryos additional cells express Kriippel forming broader expression domains which
are fused together. D) Higher magnification o f wild type Kriippel expressing cells in the ventral nerve
cord. Normally, each Kriippel expression domain is well separated from the neighbouring cluster. E) The
higher magnification o f the same region in an Iroquois mutant embryo illustrates fusion o f adjacent
clusters. Ventral view with anterior to the right.
178

Chapter 7: Mirror confers neurohiast identity'
well as a large number of local interneurons and a smaller number of late-developing
intersegmental intemeurons (Doe, 2003). The abdominal NB2-2 lineage differs from the
thoracic lineage by the absence of a particular glial cell, named the subperineural glial
cell (SPG cell). This cell derives from NBl-1 in the abdominal neuromeres, which
means that thoracic and abdominal SPG cells originate from different progenitors
(Bossing et aL, 1996a; Bossing et al., 1996b). It would be interesting to further
investigate if NB2-2 acquires a different fate in the absence of Mirror activity and if
does, how this affects its progeny.
7.2. The CNS phenotype in /n/rror mutant embryos
7.2.1. Kriippel controls cell fate decisions in the CNS
The widespread spatio-temporal expression pattern of Kriippel in the CNS
suggests an extensive requirement for Kriippel activity throughout neural development.
Indeed, a minigene which rescued the severe segmentation defects of the lack-of-
function mutation Kr^ (Wieschaus et al., 1984a), was missing early and late aspects of
Kriippel expression in the nervous system (Romani et al., 1996). Lack of Kriippel
expression in the nervous system of these embryos increases the number of cells which
express the gene gooseberry-neuro (gsh-n), whereas the excess of Kriippel function
represses gsb-n expression. In addition, it has been shown that an engrailed expressing
medio-lateral cluster of cells in the CNS, which gives rise to serotonergic neurons does
not form in the absence of Kriippel function (Romani et al., 1996). These observations
further suggest a possible role of Krüppel in controlling cell fate decisions in the CNS.
179

Chapter 7: Mirror confers neurohiast identity
7.2.2. Loss of Mirror leads to aberrant Engrailed expression in
the CNS
A medio-lateral (ML) cluster of Engrailed expressing cells in the CNS is either
absent or fails to express Engrailed in Kr^Kr^^"" embryos which lack Kriippel function
in the neural tissue (Romani et ah, 1996). Therefore, it is reasonable to ask, if ectopic
expression of Krüppel, induced by the loss of Mirror in the embryo, leads to ectopic
Engrailed and/or serotonin expressing cells in the ventral nerve cord. Antibody stainings
using Engrailed antibody was carried out on Iroquois mutant and mirror mutant
embryos at stage 16. At this stage Engrailed protein can be found in several discrete
clusters of cells. Within each segment of the CNS, En protein is detectable in three large
ventral midline cells, a cluster of dorsal medial cells and two pairs of small non
neuronal median support cells (MS), just posterior to the median NB. In addition there
are four bilateral clusters of En positive cells, a posterior intermediate group of 4-6 cells
which includes the medio lateral (ML) cluster mentioned above, a posterior lateral (PL)
group of 8-10 cells (fig.7.1 l.A+B) and two pairs of neurons located near the ventral
surface of the anterior region of the segment (Cui and Doe, 1992). Interestingly several
of these clusters are affected by the loss of mirror function in the embryo. In particular
the two MS cells are either absent or have lost their Engrailed expression
(fig.7.1 l.C+D). In addition the ML cluster seems lost and the PL cluster appears larger
than normal and consists now of approximately 14-16 En positive cells (fig.7.1 l.C+D).
It is interesting to note that similar observations were made in lethal o f scute mutant
embryos (Martin-Bermudo et al., 1991).
Thus, loss of Mirror results in aberrant Engrailed expression in the CNS. The
cells of the ML cluster disappear and additional Engrailed expressing cells are
expressed in the PL cluster. However, as the PL cluster appears to be increased by 4-6
180

Chapter 7: Mirror confers neurohiast identity
r Î • "
f * % %k € #
#
i
^ _ » f ; V
\ i *
! < >
' Î 1 %# # $ # Î 9¥
« % \ k t•r ; I 1 < f
Fçure 7.11. Aberrant Engrailed expression in mirror loss-of-function embryos. Stage 16 embryos were
stiined with anti-En using fluorescence (A+C) or DAB labelling (B+D). A+B wild type embryos showing
a 'roup o f 8-10 posterior lateral En positive cells (PL; white arrow) and 4-6 cells in the medial lateral
clister (ML; yellow arrow). B) A pair o f small non-neuronal median support cells (MS; black arrow) is
usially found just posterior to the median NB. C) Iroquois and D) mirror mutant embryo. The two MS
cels are either absent o f fail to express En. The PL cluster seems to contain additional En expressing cells
ani the cells o f the ML cluster are lost. Ventral view with anterior to the right.
181

Chapter 7: Mirror confers neurohiast identity
cells, it might be possible that the ML and PL cluster bave fused and are now sticking
together. If this observation can be directly related to ectopic expression of Kriippel is
unclear.
7.2.3. Eagle expression is lost in Iroquois mutant embryos
To clarify if the missing ML cluster is mislocated or absent in mirror mutant
embryos and if ectopic expression of Krüppel bas an effect on serotonin expressing
cells, antibody stainings using an anti-Eagle and anti-Engrailed antibody were
performed. A subset of cells from the Engrailed expressing ML cluster also express
Eagle (Eg) (fig.7.12.E+G). Eagle encodes a zinc finger protein. It has been proposed
that Eagle acts together with Huckebein, another zinc finger protein, to determine
serotonergic cell fate of the NB7-3 lineage, while Engrailed is more distantly involved
in this process by activating eagle (Dittricb et a i, 1997). During stages 10-11 Eagle is
also expressed in three other neuroblasts, NB2-4, NB3-3 and NB6-3. At stage 14, the
protein can be detected in several types of neural or glial cells (fig.7.12.A), including
interneurons joining to the anterior and posterior commissures (Higasbijima et al.,
1996). Eagle expression in Mirror loss-of-function embryos at the same stage of
development appears reduced (fig.7.12.B). After condensation of the ventral nerve cord
at stage 16, Eagle expression can be found in a group of cells in the thoracic segments
and in only three cells per hemi-neuromere in the abdominal segments (fig.7.12.C).
These clusters appear unevenly spaced and some are lost in Iroquois mutant embryos
(fig.7.12.D). If the ML cluster had shifted towards the posterior lateral cluster, the same
number of Eagle expressing cells should be present in these embryos. This is not the
case, which suggests that at least some Eagle and Engrailed coexpressing cells from the
ML cluster are missing in Iroquois mutant embryos and that the PL cluster is enriched
182

Chapter 7; Mirror confers neurohiast identity
Figure 7.12. Eagle expression is lost in Mirror loss-of-function embryos. A+C) Wild type embryos
stained with anti-Eagle antibody (green) at stage 14 and 16 respectively. B+D) Iroquois mutant embryos
exhibit loss o f Eagle staining in the ventral nerve cord. E+G) Wild type embryo at stage 16 costained with
anti-Engrailed (red) and anti-Eagle (green) antibody. G) Higher magnification o f the first 5 abdominal
segments o f the embryo shown in E. Each segment shows 2-3 En/Eg coexpressing cells (yellow-ish)
within the ML cluster. F+H) Both loss and ectopic Engrailed expression is visible in Iroquois mutant
embryos. When present. Eagle expressing cells are found at their usual location within the ML cluster and
are not seen within the PL group, which has additional engrailed expressing cells. H) Higher
magnification o f the first 5 abdominal segments o f the embryo shown in F. All panels are ventral views
with anterior to the right.

Chapter 7; Mirror confers neurohiast identity
by additional Engrailed expressing cells which must have adopted a new identity
(fig.7.12. F+H).
Thus, ectopic expression of Krüppel in Mirror loss-of-function embryos does not
increase the number of Eagle and therefore serotonin expressing cells. The observed
loss of Eagle expressing cells might be due to Mirror function on yet unidentified target
genes which in turn are also able to promote cell fate changes. Alternatively, it can not
be excluded that potentially shifted Eagle positive cells of the ML cluster died during
development.
7.2.4. Loss of Mirror function leads to defects in the CNS
Most neurons in the ventral nerve cord are interneurons, and many of these
intemeurons have long axons that are organised in a characteristic ladder-like pattern,
showing two longitudinal tracts, which are connected via two commissural tracts per
segment. It has been previously noted that the overall pattern of commissures and
connectives of the CNS is abnormal in mirror mutant embryos (McNeill, unpublished;
see also chapter 6, flg.6.6.F+H). In order to further characterise this observation, stage
14 and 16 Iroquois and wild type embryos were stained with the BP 102 antibody.
Typically the axonal projections cross the midline in one of the commissures before
they extend rostrally or caudally in one of the longitudinal pathways (fig.7.13.A). The
anterior commissure develops after, but contains almost twice as many axon bundles as
does the posterior commissure. Initially the commissures are in close proximity to each
other at the midline and only get separated as a result of cell migration and cell adhesion
events between midline glial cells, the RPl and the RP3 neurons. This separation takes
place normally during embryonic stage 12 (reviewed in Goodman and Doe, 1993;
Klambt and Goodman, 1991) but does not occur in Mirror loss-of-function embryos till
184

Chapter 7: Mirror confers neurohiast identity'
at least stage 14. In addition, the anterior and posterior commissures are usually thinner
in these embryos than in wild type embryos, suggesting that axon projections are
missing (fig.7.13.B). The longitudinal axon pathway is pioneered for much of its length
by the ascending pCC growth cone starting at stage 12. The pCC growth cone appears
to contact and extend directly towards one of the medial rows of longitudinal glial cells,
named LG5 (Jacobs and Goodman, 1989). Finally the pathway becomes complete as
several other growth cones meet and their axons fasciculate (fig.7.13.C). In Iroquois
mutant embryos the longitudinal pathway is often interrupted or does not form to
connect the segmental ganglia in the CNS (fig.7.13.D). A similar phenotype has been
described in embryos with a mutation in the gene longitudinals lacking (lola). In these
embryos the LG5 cell is bom, migrates and divides as normal but the interaction with
growth cones that extend along the longitudinal pathway is perturbed (Giniger et al.,
1994; Seeger et al., 1993). Moreover, Iroquois mutant embryos frequently show a hole
in the CNS (fig.7.13.D).
To conclude. Mirror loss-of-function embryos exhibit several defects during the
development of the CNS. Clusters of Engrailed expressing cells are altered. In particular
cells within the ML cluster are either absent, mislocated or fail to express Engrailed.
Two or three cells of this cluster normally give rise to serotonin expressing neurons.
These cells are usually characterised by their expression of the Eagle protein. A
preliminary attempt to locate this serotonin precursor cells by using an anti-Eagle
antibody suggests that these cells have most likely adopted a different cell fate and
don’t express the antigen anymore or are absent rather than mislocated. The axonal
defects of Mirror loss-of-function embryos resemble those of the lola mutant
phenotype. It would be interesting to investigate if the LG5 has also lost cell adhesion
properties or if it is missing in Mirror mutant embryos. Finally, it should be mentioned
185

Chapter 7: Mirror confers neurohiast identity
*
Figure 7.13. CNS phenotype o f Iroquois mutants. Axon tracts within the ventral nerve cord o f wild type
and Iroquois mutant embryos labelled with mAh BP 102 at stage 14 and 16. A,C) The wild type embryo
demonstrates that axons are normally organised into two longitudinal tracts, which are connected via two
commissural tracts per segment. B,D) /ro-mutant embryos exhibit a loss o f the longitudinal connections
(arrow) o f the CNS and commissures appear thinner (B) than in wild type. The severe defects in the
commissural tracts o f Iro' embryos are usually more outstanding at earlier stages o f development
(compare B and D). In addition, Iro' mutants often have holes in the CNS between the commissures o f
two segments (D). All panels are ventral views o f the embryos and are shown anterior to the right.
Wo

Chapter 7; Mirror confers neurohiast identity^
that a transient hole occurs normally in the epidermis of wild type embryos during the
ventral closure event at stage 12. Closure of this hole implicates cell migration and
adhesion of ventral epidermal cells which seam the midline in a non-random process by
interdigitation from anterior to posterior (reviewed in Martinez Arias, 1993). It might be
that these are the observed holes in the ventral epidermis which failed to close in
Iroquois mutant embryos. Alternatively it might be the same holes, which have been
observed in the disorganised ventral midline of AS-C mutant embryos. In this case, it
has been proposed that the median neuroblast does not develop and leaves a gap at its
location (Cabrera et ah, 1987).
187

Chapter 7: Mirror confers neurohiast identity^
Discussion
The microarray analysis showed that over-expression of Mirror reduces the level
of Kriippel transcript in a manner characteristic for “slow Mirror response” genes.
Downregulation of Kriippel transcript is already detected after 30 minutes of ubiquitous
Mirror activity. The in vivo analysis confirmed expression of Krüppel and Mirror in
mutually exclusive domains in the ventral nerve cord during the embryonic stages 11-
14. Krüppel expression in the CNS is highly dynamic and complex. UAS controlled
over-expression of Mirror reduces Krüppel expression in the neuroectoderm and
individual muscle precursor cells (not shown). Furthermore, loss of Mirror activity leads
to ectopic Krüppel expression in the embryo. The phenotype is variable and is
occasionally manifested in the entire Mirror domain. Krüppel expression is consistently
observed in a single medial row neuroblast per hemisegment, the NB2-2. Several other
defects in the developing CNS were also observed in Mirror loss-of-function embryos,
which might be independent of ectopic Kriippel expression. The expression of
Engrailed and Eagle is altered in these embryos in medial and lateral rows of NBs. It is
noteworthy to mention that medial and lateral rows are also preferentially affected by
the loss of lethal o f scute expression (Martin-Bermudo et al., 1991) or by removal of the
whole AS-C (Jimenez and Campos-Ortega, 1990), which suggests that Mirror and the
AS-C might act in the same pathway. Taken together, this data suggests that Mirror is
required to confer neuroblast identity. Neuroblast identity depends on the combination
of genes it expresses and is tightly correlated with the time and position at which the
neuroblast forms. The identity of a NB can be revealed by the characteristic lineage of
GMC and neurons it will form. Furthermore, local cell-cell interactions have been
shown to contribute to the cell identity of neuroblasts (Goodman and Doe, 1993). This
means that changes in identity could also occur in a non autonomous manner. It was not
188

Chapter 7: Mirror confers neurohiast identity^
possible to determine if NB2-2 or other NBs adopt a new fate in the absence of Mirror
function. It seems that neurons and glia cells are lost in mirror mutant embryos (see also
chapter 4). To assess if loss of Mirror function leads to increased cell death in the
embryo, acridine orange staining were carried out. Acridine orange is a dual-
fluorescence dye that interacts with DNA and RNA and is commonly used as a marker
for apoptotic cell death in Drosophila. Embryos which lack Mirror function showed
increased staining in a segmented pattern at stage 12 and staining did increase further in
later stages (McNeill, unpublished). Thus, cell death could also account for the observed
loss of Eagle and Engrailed positive cells.
The commissural and longitudinal axon pathways of the CNS are disrupted in
Mirror loss-of-function embryos. This indicates that Mirror plays an important role for
the proper formation of the axonal network. Indeed, Mirror is expressed in the median
neuroblast and other midline cells in the anterior part of each segment. It is this area
from which the midline glial cells develop from a set of 2-3 progenitors (Bossing and
Technau, 1994). The midline glial cells migrate along cell processes of the ventral
unpaired median neurons (VUM) to separate anterior and posterior axon commissures
(Klambt and Goodman, 1991). If this migration is blocked, a typical fused commissure
phenotype develops (Stollewerk and Klambt, 1997). This might be the case in Iroquois
mutant embryos at stage 14. However, the separation of the commissures still seems to
take place but in a retarded manner and is often incomplete. In this context, it might be
noteworthy that Krüppel is normally expressed in the midline precursor cells between
the commissures and connectives of the nervous system.
The observed defects in the longitudinal pathway persist in later stages. Again,
the specialised midline cells have an essential role in regulating the neurons that project
their axons on one side (reviewed in Tear, 1999). Specific staining of axon bundles
189

Chapter 7; Mirror confers neurohiast identity^
(fascicles) using Fasciclin I (FasI) and Fasciclin II (FasII) antibodies should be useful to
further characterise the CNS phenotype. FasI is expressed on the surface of a subset of
commissural pathways in the CNS and all sensory axon pathways in the PNS (Zinn et
a i, 1988), whereas FasII specifically labels the medial MPI fascicle and other axons in
the longitudinal pathway (Grenningloh et al., 1991).
During the course of this project, the preferred site for Mirror binding has been
identified using a site selection assay. Mirror binds to a palindromic DNA sequence
(acanntgt), which is different from the previously identified binding site for Araucan
(Aphrodite Bilioni and Helen McNeill, unpublished). In vivo assays have shown that the
palindrome is indeed functional. Repeats of the palindrome fused to a lacZ reporter
construct, which is under the control of a basal promoter, have been injected in flies.
Staining with p-galactosidase of eye imaginai discs revealed repression of the reporter
transgene in the dorsal half of the eye, which correspond to the Mirror expression
domain (Aphrodite Bilioni and Helen McNeill, unpublished). Therefore, the presence of
this palindrome in regulatory elements of genes identified in the microarray screen
would argue in favor of a direct regulation by Mirror. The bioinformatic search to match
potential target genes from the over-expression analysis which contain this site within a
regulatory element is currently under way. Strikingly, three copies of the palindrome are
located in the NS2 element in the central region (within 170bp) of the Krüppel intron.
The NSl element also contains the identified Mirror binding site. NSl and NS2 are
required to drive lacZ expression of a reporter construct in the Krüppel pattern in the
nervous system. Currently in vivo reporter assays using the Krüppel intron with wild
type and mutated palindromes are under way to further strengthen the argument that
Mirror acts directly to shape the Krüppel expression pattern in the developing CNS.
Thus, Krüppel represents the most promising candidate so far to be directly
190

Chapter 7; Mirror confers neurohiast identity
regulated by Mirror,
191

Chapter 1: Mirror confers neurohiast identity^
192

Final remarks
Microarrays were used to analyse the effects on transcription in tightly staged
Drosophila embryos, in which the transcription factor Mirror was ubiquitously over
expressed (by heat shock) for a restricted period of time. Most genes vary as a result of
the heat shock followed by differences caused by the genetic background of the flies. In
addition to the different genetic background of the flies used in this study, it became
obvious that the duration of Mirror activity and probably the amount of produced
protein were crucial for the analysis. For that reason the over-expression protocol
needed to be optimised with the aim to reduce the high number of false positives. A
large number of genes were subsequently identified which change their level of
expression in response to Mirror activity. However, the effect of interest, namely
changes in gene expression due to Mirror activity is the least prevalent effect (see table
X in the appendix). The Correlation Coefficient for each replicate group is summarised
in table XI and their hierachical clustering is illustrated in Figure C in the appendix.
During the course of this project it was attempted to complement the existing
data with an additional microarray analysis using Iroquois mutant embryos.
Unfortunately, this approach failed for technical reasons and needs to be repeated in the
future. The combination of gain and loss-of-function in a microarray screen would be
very useful to narrow down the number of potential candidate genes. But even with
these additional data, the microarray analysis on its own does not allow one to conclude
that these genes are directly regulated by Mirror. The subsequent in vivo analysis of
candidate genes is necessary to shortlist potential Mirror targets for further
investigations. Only after it has been shown that Mirror binds within a regulatory
element of a gene, whose expression is consistently altered in response to loss and gain-
193

of-function of Mirror, would it prove that the regulation is direct. During the course of
this project it has been possible to identify two genes, namely a-Tub84B and Kriippel
which alter their pattern of expression consistently in response to Mirror activity in vivo.
For one of them, Kriippel it has been possible to demonstrate that it is expressed in
mutually exclusive domains to Mirror in the CNS of the Drosophila embryo during
specific stages of development. The regulatory region, necessary for Kriippel expression
in the CNS has been identified (Hoch et al., 1990) and this relative small element
contains several Mirror binding sites (Aphrodite Bilioni and Helen McNeill,
unpublished). Electrophoretic band shift experiments and reporter assays have shown
that this site is functional (Aphrodite Bilioni and Helen McNeill, unpublished). Taken
together these results indicate that the gene Kriippel is a direct target of Mirror activity
during the development of the ventral nerve cord in the Drosophila embryo.
As a final point, it should be mentioned that 18 zinc finger transcription factors
with homology to the Drosophila Kriippel gene have been identified in mammals. This
Krüppel-like family (KLF) of proteins exhibit important tissue- or organ-specific
regulatory functions. They control a diverse range of biological processes, including,
cell growth, cell proliferation, differentiation and development (reviewed in Dang et ah,
2000). For example, KLF9 (also named BTEB) has been suggested to be involved in
neural development (Denver et al., 1999) and KLF2 and KLF6 play a role during the
formation of the lung and the heart (Anderson et a l, 1995; Wani et a l, 1999). In this
context, it would be interesting to investigate if members of the Irx family play a similar
role to regulate one or several genes of the KrUppel-like family in vertebrates in the
same way that Mirror controls Kriippel in Drosophila.
194

195

196

BibJiography
Bibliography
Acampora, D., Mazan, S., Avantaggiato, V., Barone, P., Tuorto, F., Lallemand, Y., Brulet, P. and Simeone, A. (1996). Epilepsy and brain abnormalities in mice lacking the Otxl gene. Nature Genetics 14, 218-222.
Akiyama, ¥ ., Hosoya, T., Poole, A. M. and Hotta, Y. (1996). The gcm-motif: A novel DNA-binding motif conserved in Drosophila and mammals. Proceedings- National Academy o f Sciences USA 93, 14912-14916.
Anderson, K. P., Kern, C. B., Crable, S. C. and Lingrel, J. B. (1995). Isolation of a gene encoding a functional zinc finger protein homologous to erythroid Kruppel-like factor: identification of a new multigene family. Mol Cell Biol 15, 5957-65.
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A. and Struhl, K. (1989). Short Protocols in Molecular Biology.
Azpiazu, N. and Frasch, M. (1993). tinman and bagpipe: two homeo box genes that determine cell fates in the dorsal mesoderm of Drosophila. Genes and Development 7, 1325.
Baker, N. E., Moses, K., Nakahara, D., Ellis, M. C., Carthew, R. W. and Rubin, G.M. (1992). Mutations on the second chromosome affecting the Drosophila eye. J Neurogenet 8, 85-100.
Bao, Z. Z., Bruneau, B. G., Seidman, J. G., Seidman, C. E. and Cepko, C. L.(1999). Regulation of chamber-specific gene expression in the developing heart by Irx4. Science 283, 1161-4.
Baumgartner, S., Bopp, D., Burri, M. and Noll, M. (1987). Structure of two genes at the gooseberry locus related to the paired gene and their spatial expression during Drosophila embryogenesis. Genes Dev 1, 1247-67.
Becker, M. B., Zulch, A., Bosse, A. and Gruss, P. (2001). Irxl and Irx2 expression in early lung development. Mech Dev 106, 155-8.
Bellefroid, E. J., Kobbe, A., Gruss, P., Pieler, T., Gurdon, J. B. and Papalopulu, N.(1998). Xiro3 encodes a Xenopus homolog of the Drosophila Iroquois genes and functions in neural specification. Embo J 11, 191-203.
Bertolino, E., Reimund, B., Wildt-Perinic, D. and Clerc, R. G. (1995). A novel homeobox protein which recognizes a TGT core and functionally interferes with a retinoid-responsive motif. J Biol Chem 270, 31178-88.
Bhat, K. M., van Beers, E. H. and Bhat, P. (2000). Sloppy paired acts as the downstream target of Wingless in the Drosophila CNS and interaction between sloppy paired and gooseberry inhibits sloppy paired during neurogenesis. Development 127, 655-665.
Blair, S. S. (1999). Eye development: Notch lends a handedness. Current Biology 9, R356-R360.
Bo, J. and Wensink, P. C. (1989). The promoter region of the Drosophila alpha 2- tubulin gene directs testicular and neural specific expression. Development 106, 581-7.
197

Bihliography
Bodmer, R., Barbel, S., Sheperd, S., Jack, J. W., Jan, L. Y. and Jan, Y. N. (1987). Transformation of sensory organs by mutations of the cut locus of D. melanogaster. Cell 51, 293-307.
Bodmer, R. and Jan, Y. N. (1987). Morphological differentiation of the embryonic peripheral neurons in Drosophila. Roux' Arch. Dev. Biol. 196, 69-77.
Bossing, T. and Technau, G. M. (1994). The fate of the CNS midline progenitors in Drosophila as revealed by a new method for single cell labelling. Development 120, 1895-906.
Bossing, T., Technau, G. M, and Doe, C. Q. (1996a). huckebein is required for glial development and axon pathfmding in the neuroblast 1-1 and neuroblast 2-2 lineages in the Drosophila central nervous system. Mech Dev 55, 53-64.
Bossing, T., Udolph, G., Doe, C. Q. and Technau, G. M. (1996b). The embryonic central nervous system lineages of Drosophila melanogaster. I. Neuroblast lineages derived from the ventral half of the neuroectoderm. Dev Biol 179, 41-64.
Brand, A. H., Manoukian, A. S. and Perrimon, N. (1994). Ectopic Expression in Drosophila. Methods in Cell Biology, 635.
Brand, M., Jarman, A. P., Jan, L. Y. and Jan, Y. N. (1993). Asense is a Drosophila neural precursor gene and is capable of initiating sense organ formation. Development 119, 1.
Brewster, R., Hardiman, K., Deo, M., Khan, S. and Bodmer, R. (2001). The selector gene cut represses a neural cell fate that is specified independently of the Achaete- Scute-Complex and atonal. Mech Dev 105, 57-68.
Briscoe, J., Pierani, A., Jessell, T. M. and Erieson, J. (2000). A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube. Cell 101, 435-45.
Broadus, J. and Doe, C. Q. (1995). Evolution of neuroblast identity: seven-up and prospero expression reveal homologous and divergent neuroblast fates in Drosophila and Schistocerca. Development 121, 3989-96.
Broadus, J., Skeath, J. B., Spana, E. P., Bossing, T., Technau, G. and Doe, C. Q.(1995). New neuroblast markers and the origin of the aCC/pCC neurons in the Drosophila central nervous system. Mech Dev 53, 393-402.
Bruneau, B. G., Bao, Z. Z., Tanaka, M., Schott, J. J., Izumo, S., Cepko, C. L., Seidman, J. G. and Seidman, C. E. (2000). Cardiac expression of the ventricle- specific homeobox gene Irx4 is modulated by Nkx2-5 and dHand. Dev Biol 217, 266-77.
Bürglin, T. R. (1997). Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals. Nucleic Acids Res 25, 4173-80.
Cabrera, C. V. and Alonso, M. C. (1991). Transcriptional activation by heterodimers of the achaetescute and daughterless gene products. Abstracts o f Papers Presented at the Meeting on Molecular Neurobiology o f Drosophila, 32.
Cabrera, C. V., Martinez-Arias, A. and Bate, M. (1987). The expression of three members of the achaete-scute gene complex correlates with neuroblast segregation in Drosophila. Ce//50, 425-33.
198

Bihliography
Cadigan, K. M., Grossniklaus, U. and Gehring, W. J. (1994). Functional redundancy: The respective roles of the two sloppy paired genes in Drosophila segmentation. Proceedings- National Academy o f Sciences USA 91, 6324.
Campos-Ortega, J. A. (1993). Early Neurogenesis in Drosophila melanogaster. NY: Cold Spring Harbor Laboratory Press.
Campuzano, S. and Modolell, J. (1992). Patterning of the Drosophila nervous system: the achaete-scute gene complex. Trends Genet 8, 202-8.
Carrera, P., Abrell, S., Kerber, B., Walldorf, U., Preiss, A., Hoch, M. and Jackie, H. (1998). A modifier screen in the eye reveals control genes for Kruppel activity in the Drosophila embryo. Proc Natl Acad Sci US A9 5 , 10779-84.
Carroll, S. B. (1990). Zebra patterns in fly embryos: activation of stripes or repression of interstripes? Cell 60, 9-16.
Casso, D., Ramirez-Weber, F. and Kornberg, T. B. (2000). GFP-tagged balancer chromosomes for Drosophila melanogaster. Mech Dev 91, 451-4.
Cavodeassi, F., Diez Del Corral, R., Campuzano, S. and Dominguez, M. (1999). Compartments and organising boundaries in the Drosophila eye: the role of the homeodomain Iroquois proteins. Development 126, 4933-42.
Cavodeassi, F., Modolell, J. and Campuzano, S. (2000). The Iroquois homeobox genes function as dorsal selectors in the Drosophila head. Development 127, 1921-9.
Cavodeassi, F., Modolell, J. and Gomez-Skarmeta, J. L. (2001). The Iroquois family of genes: from body building to neural patterning. Development 128, 2847-55.
Chitnis, A., Henrique, D., Lewis, J., Ish-Horowicz, D. and Kintner, C. (1995). Primary neurogenesis in Xenopus embryos regulated by a homologue of the Drosophila neurogenic gene Delta. Nature 375, 761-6.
Cho, K. O. and Choi, K. W. (1998). Fringe is essential for mirror symmetry and morphogenesis in the Drosophila eye. Nature 396, 272-6.
Christoffels, V. M., Habets, P. E., Franco, D., Campione, M., de Jong, F., Lamers, W. H., Bao, Z. Z., Palmer, S., Biben, C., Harvey, R. P. et al, (2000). Chamber formation and morphogenesis in the developing mammalian heart. Dev Biol 223, 266-78.
Coffman, C. R., Skoglund, P., Harris, W. A. and Kintner, C. R. (1993). Expression of an extracellular deletion of Xotch diverts cell fate in Xenopus embryos. Cell 73, 659- 71.
Cohen, B., McGuffin, M. E., Pfeifle, C., Segal, D. and Cohen, S. M. (1992). apterous, a gene required for imaginai disc development in Drosophila encodes a member of the LIM family of developmental regulatory proteins. Genes Dev 6, 715-29.
Cohen, D. R., Cheng, C. W., Cheng, S. H. and Hui, C. C. (2000). Expression of two novel mouse Iroquois homeobox genes during neurogenesis. Mech Dev 91, 317-21.
Cui, X. and Doe, C. Q. (1992). ming is expressed in neuroblast sublineages and regulates gene expression in the Drosophila central nervous system. Development 116, 943-52.
Dahlsveen, I. (2002). Identification and characterisation of proteins that interact with the Drosophila transcription factor Mirror. Thesis: University College London.
199

Bihliography
Dahmus, M. E. (1994). The role of multisite phosphorylation in the regulation of RNA polymerase II activity. Prog Nucleic Acid Res Mol Biol 48, 143-79.
Dang, D. T., Pevsner, J. and Yang, V. W. (2000). The biology of the mammalian Kruppel-like family of transcription factors. Int J Biochem Cell Biol 32, 1103-21.
de la Calle-Mustienes, E., Glavic, A., Modolell, J. and Gomez-Skarmeta, J. L.(2002a). Xiro homeoproteins coordinate cell cycle exit and primary neuron formation by upregulating neuronal-fate repressors and downregulating the cell-cycle inhibitor XGadd45-gamma. Mech Dev 119, 69.
de la Calle-Mustienes, E., Modolell, J. and Gomez-Skarmeta, J. L. (2002b). The Xiro-repressed gene CoREST is expressed in Xenopus neural territories. Mech Dev 110, 209-11.
Denver, R. J., Ouellet, L., Furling, D., Kobayashi, A., Fujii-Kuriyama, Y. and Puymirat, J. (1999). Basic transcription element-binding protein (BTEB) is a thyroid hormone-regulated gene in the developing central nervous system. Evidence for a role in neurite outgrowth. JBiol Chem 274, 23128-34.
Desplan, C., Theis, J. and O’Farrell, P. H. (1985). The Drosophila developmental gene, engrailed, encodes a sequence-specific DNA binding activity. Nature 318, 630-5.
Dettman, R. W., Turner, F. R. and Raff, F. C. (1996). Genetic analysis of the Drosophila beta3-tubulin gene demonstrates that the microtubule cytoskeleton in the cells of the visceral mesoderm is required for morphogenesis of the midgut endoderm. Dev Biol 111, 117-35.
Diatchenko, L., Lan, Y. F. C., Campbell, A. P., Chenchik, A., Moqadam, F., Huang, B., Lukyanov, S., Lukyanov, K., Gurskaya, N. and Sverdlov, F. D. (1996). Suppression subtractive hybridization: A method for generating differentially regulated or tissue-specific cDNA probes and libraries. Proceedings- National Academy o f Sciences USA 93, 6025-6030.
Diez del Corral, R., Aroca, P., JL, G. m.-S., Cavodeassi, F. and Modolell, J. (1999). The Iroquois homeodomain proteins are required to specify body wall identity in Drosophila. Genes Dev 13, 1754-61.
Dittrich, R., Bossing, T., Gould, A. P., Technau, G. M. and Urban, J. (1997). The differentiation of the serotonergic neurons in the Drosophila ventral nerve cord depends on the combined function of the zinc finger proteins Eagle and Huckebein. Development 124, 2515-25.
Doe, C. Q. (1992). Molecular markers for identified neuroblasts and ganglion mother cells in the Drosophila central nervous system. Development 116, 855-63.
Doe, C. Q. (2003). Neuroblast Lineages. http://www.neuro.uoregon.edu/doelab/nbintro.html
Dominguez, M. and Campuzano, S. (1993). asense, a member of the Drosophila achaete - scute complex, is a proneural and neural differentiation gene. Embo Journal 12, 2049.
Dominguez, M. and de Celis, J. F. (1998). A dorsal/ventral boundary established by Notch controls growth and polarity in the Drosophila eye. Nature 396, 276-8.
Dormand, F. L. and Brand, A. H. (1998). Runt determines cell fates in the Drosophila embryonic CNS. Development 125, 1659-67.
2 0 0

Bihliography
Duboule, D. (1991). Patterning in the vertebrate limb. Curr Opin Genet Dev 1, 211-6.
Egger, B., Leemans, R., Loop, T., Kammermeier, L., Fan, Y., Radimerski, T., Strahm, M. C., Certa, U. and Reichert, H. (2002). Gliogenesis in Drosophila: genome-wide analysis of downstream genes of glial cells missing in the embryonic nervous system. Development 129, 3295-3309.
Fainsod, A., Steinbeisser, H. and De Robertis, E. M. (1994). On the function of BMP-4 in patterning the marginal zone of the Xenopus embryo. Embo Journal 13, 5015.
Finnerty, J. R. and Martindale, M. Q. (1998). The evolution of the Hox cluster: insights from outgroups. Curr Opin Genet Dev 8, 681-7.
Fujise, M., Izumi, S., Selleck, S. B. and Nakato, H. (2001). Regulation of dally, an integral membrane proteoglycan, and its function during adult sensory organ formation of Drosophila. Dev Biol 235, 433-48.
Furlong, E. E. M., Andersen, E. C., Null, B., White, K. P. and Scott, M. P. (2001). Patterns of Gene Expression During Drosophila Mesoderm Development. Science, 1629-1632.
Gasch, A., Hinz, U. and Renkawitz-Pohl, R. (1989). Intron and upstream sequences regulate expression of the Drosophila beta 3-tubulin gene in the visceral and somatic musculature, respectively. Proc Natl Acad Sci U S A ^ 6 , 3215-8.
Gaul, U. and Jackie, H. (1987). Pole region-dependent repression of the Drosophila gap gene Kruppel by maternal gene products. Cell 51, 549-55.
Gaul, U., Seifert, E., Schuh, R. and Jackie, H. (1987). Analysis of Kruppel protein distribution during early Drosophila development reveals posttranscriptional regulation. Cell 50, 639-47.
Gaul, U. and Weigel, D. (1990). Regulation of Kruppel expression in the anlage of the Malpighian tubules in the Drosophila embryo. Mech Dev 33, 57-67.
Gehring, W. J., Qian, Y. Q., Billeter, M., Furukubo-Tokunaga, K., Schier, A. F., Resendez-Perez, D., Affolter, M., Otting, G. and Wuthrich, K. (1994). Homeodomain-DNA recognition. Cell 78, 211-23.
Ghysen, A. and Dambly-Chaudiere, C. (1986). The pattern of sense organs in Drosophila embryos and larvae. Roux' Arch. Dev. Biol. 195, 222-228.
Gibson, G. and Gehring, W. J. (1988). Head and thoracic transformations caused by ectopic expression of Antennapedia during Drosophila development. Development 102, 657-675.
Giebel, B., Stuettem, I., Hinz, U. and Campos-Ortega, J. A. (1997). Lethal of Scute requires overexpression of Daughterless to elicit ectopic neuronal development during embryogenesis in Drosophila. Mechanisms o f Development 63, 75-88.
Giniger, E., Tietje, K., Jan, L. Y. and Jan, Y. N. (1994). lola encodes a putative transcription factor required for axon growth and guidance in Drosophila. Development 120,1385-98.
Glavic, A., Gomez-Skarmeta, J. L. and Mayor, R. (2001). Xiro-1 controls mesoderm patterning by repressing bmp-4 expression in the Spemann organiser. Dev Dyn 222, 368-76.
2 0 1

Bihliography
Gomez-Skarmeta, J., de La Calle-Mustienes, E. and Modolell, J. (2001), The Wnt- activated Xirol gene encodes a repressor that is essential for neural development and downregulates Bmp4. Development 128, 551-60.
Gomez-Skarmeta, J. L., Diez del Corral, R., de la Calle-Mustienes, E., Ferre- Marco, D. and Modolell, J. (1996). Araucan and caupolican, two members of the novel iroquois complex, encode homeoproteins that control proneural and vein-forming genes. Cell 85, 95-105.
Gomez-Skarmeta, J. L., Glavic, A., de la Calle-Mustienes, E., Modolell, J. and Mayor, R. (1998). Xiro, a Xenopus homolog of the Drosophila Iroquois complex genes, controls development at the neural plate. Embo J 17, 181-90.
Gomez-Skarmeta, J. L. and Modolell, J. (2002). Iroquois genes: genomic organization and function in vertebrate neural development. Curr Opin Genet Dev 12, 403-8.
Goodman, C. S. and Doe, C. Q. (1993). Embryonic development of the Drosophila central nervous system. NY.
Goriely, A., Diez del Corral, R. and Storey, K. G. (1999). c-Irx2 expression reveals an early subdivision of the neural plate in the chick embryo. Mech Dev 87, 203-6.
Goulding, S. E., zur Lage, P. and Jarman, A. P. (2000). amos, a Proneural Gene for Drosophila Olfactory Sense Organs that Is Regulated by lozenge. Neuron 25, 69-78.
Grenningloh, G., Rehm, E. J. and Goodman, C. S. (1991). Genetic analysis of growth cone guidance in Drosophila: fasciclin II functions as a neuronal recognition molecule. Cell 67, 45-57.
Grillenzoni, N., van Helden, J., Dambly-Chaudiere, C. and Ghysen, A. (1998). The iroquois complex controls the somatotopy of Drosophila notum mechanosensory projections. Development 125, 3563-9.
Gurskaya, N. O., Diatchenko, L., Chenchik, A., Siebert, P. D., Khaspekov, G. L., Lukyanov, K. A., Vagner, L. L., Ermolaeva, O. D., Lukyanov, S. A. and Sverdlov,E. D. (1996). Equalizing cDNA Subtraction Based on Selective Suppression of Polymerase Chain Reaction: Cloning of Jurkat Cell Transcripts Induced by Phytohemaglutinin and Phorbol 12-Myristate 13-Acetate. Analytical Biochemistry 240, 90-97.
Hafen, E., Levine, M. and Gehring, W. J. (1984). Regulation of Antennapedia transcript distribution by the bithorax complex in Drosophila. Nature 307, 287-9.
Halter, D. A., Urban, J., Rickert, C. and Ner, S. S. (1995). The homeobox gene repo is required for the differentiation and maintenance of glia function in the embryonic nervous system of Drosophila melanogaster. Development 121, 317.
Heberlein, U., Borod, E. R. and Chanut, F. A. (1998). Dorsoventral patterning in the Drosophila retina by wingless. Development 125, 567-77.
Higashijima, S., Shishido, E., Matsuzaki, M. and Saigo, K. (1996). eagle, a member of the steroid receptor gene superfamily, is expressed in a subset of neuroblasts and regulates the fate of their putative progeny in the Drosophila CNS. Development 122, 527-36.
2 0 2

Bihliography
Hinz, U., Giebel, B. and Campos-Ortega, J. A. (1994). The Basic-Helix-Loop-Helix Domain of Drosophila Lethal of Scute Protein is Sufficient for Proneural Function and Activates Neurogenic Genes. Cell 76, 77.
Hirth, F., Hartmann, B. and Reichert, H. (1998). Homeotic gene action in embryonic brain development of Drosophila. Development 125, 1579-1589.
Hoch, M., Gerwin, N., Taubert, H. and Jackie, H. (1992). Competition for overlapping sites in the regulatory region of the Drosophila gene Kruppel. Science 256, 94-7.
Hoch, M. and Jackie, H. (1998). Kruppel acts as a developmental switch gene that mediates Notch signalling-dependent tip cell differentiation in the excretory organs of Drosophila. Embo J 11, 5766-75.
Hoch, M., Schroder, C., Seifert, E. and Jackie, H. (1990). cis-acting control elements for Kruppel expression in the Drosophila embryo. Embo J 9, 2587-95.
Hoppler, S. and Bienz, M. (1994). Specification of a Single Cell Type by a Drosophila Homeotic Gene. Cell 76, 689.
Hoshizaki, D. K. (1994). Kruppel expression during postembryonic development of Drosophila. Dev Biol 162, 133-40.
Houweling, A. C., Dildrop, R., Peters, T., Mummenhoff, J., Moorman, A. F., Ruther, U. and Christoffels, V. M. (2001). Gene and cluster-specific expression of the Iroquois family members during mouse development. Mech Dev 107, 169-74.
Hulskamp, M., Pfeifle, C. and Tautz, D. (1990). A morphogenetic gradient of hunchback protein organizes the expression of the gap genes Kruppel and knirps in the early Drosophila embryo. Nature 346, 577-80.
Isshiki, T., Pearson, B., Holbrook, S. and Doe, C. Q. (2001). Drosophila neuroblasts sequentially express transcription factors which specify the temporal identity of their neuronal progeny. Cell 106, 511-21.
Itoh, M., Kudoh, T., Dedekian, M., Kim, C. H. and Chitnis, A. B. (2002). A role for irol and iro7 in the establishment of an anteroposterior compartment of the ectoderm adjacent to the midbrain-hindbrain boundary. Development 129, 2317-27.
Jackie, H., Gaul, U. and Redemann, N. (1988). Regulation and putative function of the Drosophila gap gene Kriippel. Development 104 Supplement, 29-34.
Jacob, Y., Sather, S., Martin, J. R. and Olio, R. (1991). Analysis of Kruppel control elements reveals that localized expression results from the interaction of multiple subelements. Proc Natl Acad Sci U S A 88, 5912-6.
Jacobs, J. R. and Goodman, C. S. (1989). Embryonic development of axon pathways in the Drosophila CNS. II. Behavior of pioneer growth cones. JNeurosci 9, 2412-22.
Jarman, A. P., Gran, ¥ ., Jan, L. Y. and Jan, Y. N. (1993a). Atonal Is a Proneural Gene That Directs Chordotonal Organ Formation in the Drosophila Peripheral Nervous System. Cell 73, 1307.
Jarman, A. P., Grell, E., Jan, L. Y. and Jan, Y. N. (1993b). atonal, a proneural gene for both chordotonal organs and the eye. In Neurobiology o f drosophila, (ed., pp. 137: Cold Spring Harbor Laboratory.
203

Bihliography
Jarman, A. P., Sun, Y., Jan, L. Y. and Jan, Y. N. (1995). Role of the proneural gene, atonal, in formation of Drosophila chordotonal organs and photoreceptors. Development 121, 2019.
Jimenez, F. and Campos-Ortega, J. A. (1990). Defective neuroblast commitment in mutants of the achaete-scute complex and adjacent genes of D. melanogaster. Neuron 5, 81-9.
Jin, W., Riley, R.M., Wolfinger, R.D., White, K.P., Passador-Gurgel, G., Gibson,G.(2001) The contributions of sex, genotype and age to transcriptional variance in Drosophila melanogaster. Nat Genet 29, 389-399.
Jones, B. W., Fetter, R. D., Tear, G. and Goodman, C. S. (1995). glial cells missing: A Genetic Switch That Controls Glial versus Neuronal Fate. Cell 82, 1013.
Jordan, K. C., Clegg, N. J., Blasi, J. A., Morimoto, A. M., Sen, J., Stein, D., McNeill, H., Deng, W. M., Tworoger, M. and Ruohola-Baker, H. (2000). The homeobox gene mirror links EGF signalling to embryonic dorso- ventral axis formation through notch activation. Nat Genet 24, 429-33.
Jordan, K. C. and Ruohola-Baker, H. (1999). The homeobox gene mirror links EGF signalling to Notch-Fringe mediated border formation. In 16th European Drosophila Research Conference, (ed. Zurich.
Kalfayan, L. and Wensink, P. C. (1982). Developmental regulation of Drosophila alpha-tubulin genes. Cell 29, 91-8.
Kehl, B. T., Cho, K. O. and Choi, K. W. (1998). mirror, a Drosophila homeobox gene in the Iroquois complex, is required for sensory organ and alula formation. Development 125, 1217-27.
Klaes, A., Menne, T., Stollewerk, A., Scholz, H. and Klambt, C. (1994). The Ets transcription factors encoded by the Drosophila gene pointed direct glial cell differentiation in the embryonic CNS. Cell 78, 149-60.
Klambt, C. and Goodman, C. S. (1991). Role of the midline glia and neurons in the formation of the axon commissures in the central nervous system of the Drosophila embryo. Ann N Y Acad Sci 633, 142-59.
Knipple, D. C., Seifert, E., Rosenberg, U. B., Preiss, A. and Jackie, H. (1985). Spatial and temporal patterns of Kruppel gene expression in early Drosophila embryos. Nature 317, 40-4.
Kobayashi, D., Kobayashi, M., Matsumoto, K., Ogura, T., Nakafuku, M. and Sbimamura, K. (2002). Early subdivisions in the neural plate define distinct competence for inductive signals. Development 129, 83-93.
Kopp, A., Muskavitcb, M. A. T. and Duncan, I. (1997). The roles of hedgehog and engrailed in patterning adult abdominal segments of Drosophila. Development 124, 3703-3714.
Krebs, R. A. and Feder, M. E. (1997). Tissue-specific variation in Hsp70 expression and thermal damage in Drosophila melanogaster larvae. J Exp Biol 200 ( Pt 14), 2007-15.
Kremser, T., Gajewski, K., Schulz, R. A. and Renkawitz-Pobl, R. (1999). Tinman regulates the transcription of the beta3 tubulin gene (betaTubbOD) in the dorsal vessel of Drosophila. Dev Biol 216, 327-39.
204

Bihliography
Kudoh, T. and Dawid, I. B. (2001). Role of the iroquois3 homeobox gene in organiser formation, Proc NatlAcad Sci US A9 S , 7852-7.
Lakhotia, S. C. and Prasanth, K. V. (2002). Tissue- and development-specific induction and turnover of hsp70 transcripts from loci 87A and 87C after heat shock and during recovery in Drosophila melanogaster. J Exp Biol 205, 345-58.
Lawrence, P. A. and Morata, G. (1994). Homeobox genes: their function in Drosophila segmentation and pattern formation. Cell 78, 181-9.
Lawrence, P. A., Sanson, B. and Vincent, J. P. (1996). Compartments, wingless and engrailed: patterning the ventral epidermis of Drosophila embryos. Development 122, 4095-4103.
Lawrence, P. A. and Struhl, G. (1996). Morphogens, Compartments, and Pattern: Lessons from Drosophila? Cell 85, 951-962.
Lecaudey, V., Thisse, C., Thisse, B. and Schneider-Maunoury, S. (2001). Sequence and expression pattern of ziro7, a novel, divergent zebrafish iroquois homeobox gene. Mech Dev 109, 383-8.
Lee, H. H. and Frasch, M. (2000). Wingless effects mesoderm patterning and ectoderm segmentation events via induction of its downstream target sloppy paired. Development 127, 5497-508.
Leemans, R., Egger, B., Loop, T., Kammermeier, L., He, H., Hartmann, B., Certa, U., Hirth, F. and Reichert, H. (2000). Quantitative transcript imaging in normal and heat-shocked Drosophila embryos by using high-density oligonucleotide arrays. Proceedings- National Academy o f Sciences USA 97, 12138-12143.
Leemans, R., Loop, T., Egger, B., He, H., Kammermeier, L., Hartmann, B., Certa, U., Reichert, H. and Hirth, F. (2001). Identification of candidate downstream genes for the homeodomain transcription factor Labial in Drosophila through oligonucleotide- array transcript imaging. Genome Biol 2, RESEARCH0015.
Leuzinger, S., Hirth, F., Gerlich, D., Acampora, D., Simeone, A., Gehring, W. J., Finkelstein, R., F urukubo-T okunaga, K. and Reichert, H. (1998). Equivalence of the fly orthodenticle gene and the human OTX genes in embryonic brain development of Drosophila. Development 125, 1703-1710.
Levine, M. and Hoey, T. (1988). Homeobox proteins as sequence-specific transcription factors. Cell 55, 537-40.
Lewis, E. B. (1978). A gene complex controlling segmentation in Drosophila. Nature 276, 565-70.
Lewis, M. T., Ross, S., Strickland, P. A., Snyder, C. J. and Daniel, D. W. (1999). Regulated expression patterns of IRX-2, an Iroquois-class homeobox gene, in the human breast. Cell and Tissue Research 296, 549-554.
Leyns, L., Gomez-Skarmeta, J. L. and Dambly-Chaudiere, C. (1996). iroquois: a prepattem gene that controls the formation of bristles on the thorax of Drosophila. Mech Dev 59, 63-72.
Lindquist, S. and Craig, E. A. (1988). The heat-shock proteins. Annu Rev Genet 21, 631-77.
Lis, J. (1998). Promoter-associated pausing in promoter architecture and postinitiation transcriptional regulation. Cold Spring Harb Symp Quant Biol 63, 347-56.
205

Bihliography
Lu, H., Flores, O., Weinmann, R. and Reinberg, D. (1991). The nonphosphorylated form of RNA polymerase II preferentially associates with the preinitiation complex. Proc Natl Acad Sci U SA 88, 10004-8.
Ma, Q., Kintner, C. and Anderson, D. J. (1996). Identification of neurogenin, a Vertebrate Neuronal Determination Gene. Cell 87, 43-52.
Mann, R. S. and Chan, S. K. (1996). Extra specificity from extradenticle: the partnership between HOX and PBX/EXD homeodomain proteins. Trends Genet 12, 258-62.
Mann, R. S. and Hogness, D. S. (1990). Functional dissection of Ultrabithorax proteins in D. melanogaster. Cell 60, 597-610.
Martin-Bermudo, M. D., Gonzalez, F., Dominguez, M. and Rodriguez, I. (1993). Molecular characterization of the lethal of scute genetic function. Development 118, 1003.
Martin-Bermudo, M. D., Martinez, C., Rodriguez, A. and Jimenez, F. (1991). Distribution and function of the lethal of scute gene product during early neurogenesis in Drosophila. Development 113, 445-54.
Martinez Arias, A. (1993). Development and patterning of the larval epidermis of Drosophila. NY : Cold Spring Harbor Laboratory Press.
Matthews, K. A., Miller, D. F. and Kaufman, T. C. (1990). Functional implications of the unusual spatial distribution of a minor alpha-tubulin isotype in Drosophila: a common thread among chordotonal ligaments, developing muscle, and testis cyst cells. Dev Biol 137, 171-83.
McDonald, M. J. and Rosbash, M. (2001). Microarray analysis and organization of circadian gene expression in Drosophila. Cell 107, 567-78.
McGinnis, W., Levine, M. S., Hafen, E., Kuroiwa, A. and Gehring, W. J. (1984). A conserved DNA sequence in homoeotic genes of the Drosophila Antennapedia and bithorax complexes. Nature 308, 428-33.
McNeill, H., Yang, C. H., Brodsky, M., Ungos, J. and Simon, M. A. (1997). mirror encodes a novel PBX-class homeoprotein that functions in the definition of the dorsal- ventral border in the Drosophila eye. Genes Dev 11, 1073-82.
Mellerick, D. M., Kassis, J. A., Zhang, S. D. and Odenwald, W. F. (1992). castor encodes a novel zinc finger protein required for the development of a subset of CNS neurons in Drosophila. Neuron 9, 789-803.
Milan, M., Weihe, U., Perez, L. and Cohen, S. M. (2001a). The LRR Proteins Capricious and Tartan Mediate Cell Interactions during DV Boundary Formation in the Drosophila Wing. Cell 106, 785-794.
Milan, M., Weihe, U., Tiong, S., Bender, W. and Cohen, S. M. (2001b). msh specifies dorsal cell fate in the Drosophila wing. Development 128, 3263-3268.
Mizuguchi, R., Sugimori, M., Takebayashi, H., Kosako, H., Nagao, M., Yoshida, S., Nabeshima, Y., Sbimamura, K. and Nakafuku, M. (2001). Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype- specific properties of motoneurons. Neuron 31, 757-71.
206

Bihliography
Montalta-He, H., Leemans, R., Loop, T., Strahm, M., Certa, U. and Reichert, H.(2002). Evolutionary conservation of otd/OTX2 transcription factor action: a full genome microarray analysis. Annual Drosophila Research Conference, 746B.
Morata, G. (2001). How Drosophila appendages develop. Nat Rev Mol Cell Biol 2, 89- 97.
Mummenhoff, J., Houweling, A. C., Peters, T., Christoffels, V. M. and Ruther, U.(2001). Expression of Irx6 during mouse morphogenesis. Mech Dev 103, 193-5.
Netter, S., Fauvarque, M. O., Diez del Corral, R., Dura, J. M. and Coen, D. (1998). white+ transgene insertions presenting a dorsal/ventral pattern define a single cluster of homeobox genes that is silenced by the polycomb-group proteins in Drosophila melanogaster. Genetics 149, 257-75.
Nilson, L. A. and Schuepbach, T. (1998). Localized Requirements for windbeutel and pipe Reveal a Dorsoventral Prepattern within the Follicular Epithelium of the Drosophila Ovary, Cell 93, 253-262.
Nogales, E., Wolf, S. G. and Downing, K. H. (1998). Structure of the àa tubulin dimer by electron crystallography. Nature, 199-202.
Nover, L. and Scharf, K. D. (1997). Heat stress proteins and transcription factors. Cell Mol Life Sci SZ, 80-103.
Novitch, B. G., Chen, A. I. and Jessell, T. M. (2001). Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Glig2. Neuron 31, 773-89.
Nusslein-Volhard, C. and Wieschaus, E. (1980). Mutations affecting segment number and polarity in Drosophila. Nature 287, 795-801.
Ogura, K., Matsumoto, K., Kuroiwa, A., Isobe, T., Otoguro, T., Jurecic, V., Baldini, A., Matsuda, Y. and Ogura, T. (2001). Cloning and chromosome mapping of human and chicken Iroquois (IRX) genes. Cytogenet Cell Genet 92, 320-5.
Pankratz, M. J. and Jackie, H. (1993). Blastoderm Segmentation. NY: Cold Spring Harbor Laboratory Press.
Papayannopoulos, V., Tomlinson, A., Panin, V. M., Rauskolb, C. and Irvine, K. D.(1998). Dorsal-Ventral Signaling in the Drosophila Eye. Science, 2031-2033.
Parras, C., Garcia-Alonso, L. A., Rodriguez, I. and Jimenez, F. (1996). Control of neural precursor specification by proneural proteins in the CNS of Drosophila. Embo Journal 15, 6394-6399.
Peri, F., Technau, M. and Roth, S. (2002). Mechanisms of Gurken-dependent pipe regulation and the robustness of dorsoventral patterning in Drosophila. Development 129, 2965-75.
Peters, T., Ausmeier, K., Dildrop, R. and Ruther, U. (2002). The mouse Fused toes (Ft) mutation is the result of a 1.6-Mb deletion including the entire Iroquois B gene cluster. Mamm Genome 13, 186-8.
Peters, T., Dildrop, R., Ausmeier, K. and Ruther, U. (2000). Organization of mouse Iroquois homeobox genes in two clusters suggests a conserved regulation and function in vertebrate development. Genome Res 10, 1453-62.
Pichaud, F. and Casares, F. (2000). homothorax and iroquois-C genes are required for the establishment of territories within the developing eye disc. Mech Dev 96, 15-25.
207

Bihliography
Postlethwait, J. H., Yan, Y. L., Gates, M. A., Home, S., Amores, A., Brownlie, A., Donovan, A., Egan, E. S., Force, A., Gong, Z. et al. (1998), Vertebrate genome evolution and the zebrafish gene map. Nat Genet 18, 345-9.
Primer3: http://www-genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi
Prokopenko, S. N., He, Y., Lu, Y. and Beilen, H. J. (2000). Mutations affecting the development of the peripheral nervous system in Drosophila: A molecular screen for novel proteins. Genetics 156, 1691-1715.
Rasmussen, E. B. and Lis, J. T. (1993). In vivo transcriptional pausing and cap formation on three Drosophila heat shock genes. Proc NatlAcad Sci U S A90, 7923-7.
Robinson-Rechavi, M., Marchand, O., Escriva, H. and Laudet, V. (2001). An ancestral whole-genome duplication may not have been responsible for the abundance of duplicated fish genes. Curr Biol 11, R458-9.
Rodriguez, I., Hernandez, R., Modolell, J. and Ruiz-Gomez, M. (1990). Competence to develop sensory organs is temporally and spatially regulated in Drosophila epidermal primordia. Embo J 9, 3583-92.
Romani, S., Campuzano, S. and Modolell, J. (1987). The achaete-scute complex is expressed in neurogenic regions of Drosophila embryos. Embo J6 , 2085-2092.
Romani, S., Jimenez, F., Hoch, M., Patel, N. H., Taubert, H. and Jackie, H. (1996). Kruppel, a Drosophila segmentation gene, participates in the specification of neurons and glial cells. Mech Dev 60, 95-107.
Rosenberg, U. B., Schroder, C., Preiss, A., Kienlin, A., Cote, S., Riede, J. and Jackie, H. (1986). Structural homology of the product of the Drosophila Kriippel gene W\ûiXenopus transcription factor IIIA. Nature 319, 336-339.
Rougvie, A. E. and Lis, J. T. (1988). The RNA polymerase II molecule at the 5' end of the uninduced hsp70 gene of D. melanogaster is transcriptionally engaged. Cell 54, 795- 804.
Ruiz-Gomez, M., Romani, S., Hartmann, C., Jackie, H. and Bate, M. (1997). Specific muscle identities are regulated by Kruppel during Drosophila embryogenesis. Development 124, 3407-14.
Salzberg, A., Prokopenko, S. N., He, Y., Tsai, P., Pal, M., Maroy, P., Glover, D. M., Deak, P. and Bellen, H. J. (1997). P-element insertion alleles of essential genes on the third chromosome of Drosophila melanogaster: mutations affecting embryonic PNS development. Genetics 147, 1723-41.
Sambrook, J., Fritscb, E. F. and Maniatis, T. (1989). Molecular Cloning: a Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sauer, F. and Jackie, H. (1993). Dimerization and the control of transcription by Kruppel. Nature 364, 454-7.
Scbmucker, D., Taubert, H. and Jackie, H. (1992). Formation of the Drosophila larval photoreceptor organ and its neuronal differentiation require continuous Kruppel gene activity. Neuron 9, 1025-39.
Scbreiber, J., Sock, E. and Wegner, M. (1997). The regulator of early gliogenesis glial cells missing is a transcription factor with a novel type of DNA-binding domain. Proceedings- National Academy o f Sciences USA 94, 4739-4744.
208

Bihliography
Scott, M. P. and Weiner, A. J. (1984). Structural relationships among genes that control development: sequence homology between the Antennapedia, Ultrabithorax, and fushi tarazu loci of Drosophila. Proc Natl Acad Sci U S A S l , 4115-9.
Seeger, M., Tear, G., Ferres-Marco, D. and Goodman, C. S. (1993). Mutations affecting growth cone guidance in Drosophila: genes necessary for guidance toward or away from the midline. Neuron 10, 409-26.
Sharman, A. C. and Brand, M. (1998). Evolution and homology of the nervous system: cross-phylum rescues of otd/Otx genes. Trends in Genetics 14, 211-213.
Skeath, J. B. (1999). At the nexus between pattern formation and cell-type specification: the generation of individual neuroblast fates in the Drosophila embryonic central nervous system. Bioessays 21, 922-931.
Skeath, J. B. and Doe, C. Q. (1996). The achaete-scute complex proneural genes contribute to neural precursor specification in the Drosophila CNS. Curr Biol 6, 1146- 52.
Skeath, J. B., Panganiban, G., Selegue, J. and Carroll, S. B. (1992). Gene regulation in two dimensions: the proneural achaete and scute genes are controlled by combinations of axis-patterning genes through a common intergenic control region. Genes and Development 6, 2606.
Spemann, H. and Mangold, H. (1924). Über Induktion von Embryonanlagen durch Implantation artfremder Organisatoren. Roux' Arch. f. Entw. meek 100, 599-638.
Stollewerk, A. and Klambt, C. (1997). The midline glia cells are required for compartmentalisation of axon commissures in the embryonic CNS of Drosophila. Dev Genes Evol 207, 401-409.
Struhl, G., Barbash, D. A. and Lawrence, P. A. (1997). Hedgehog acts by distinct gradient and signal relay mechanisms to organise cell type and cell polarity in the Drosophila abdomen. Development 124, 2155-2165.
Struhl, G. and White, R. A. (1985). Regulation of the Ultrabithorax gene of Drosophila by other bithorax complex genes. Cell 43, 507-19.
Tan, J. T. T., Korzh, V. and Gong, Z. (1999). Expression of a zebrafish iroquois homeobox gene, Ziro3, in the midline axial structures and central nervous system. Mechanisms o f Development 87, 165-168.
Tear, G. (1999). Axon guidance at the central nervous system midline. Cell Mol Life Sci 55, 1365-76.
Theurkauf, W. E. (1992). Behavior of Structurally Divergent à-Tubulin Isotypes during Drosophila Embryogenesis: Evidence for Post-translational Regulation of Isotype Abundance. Developmental Biology 154, 205.
Van der Hoeven, F., Schimmang, T., Volkmann, A. and Mattel, M. G. (1994). Programmed cell death is affected in the novel mouse mutant Fused toes (Ft). Development 120, 2601.
Walter, M. F., Petersen, N. S. and Biessmann, H. (1990). Heat shock causes the collapse of the intermediate filament cytoskeleton in Drosophila embryos. Dev Genet 11,270-9.
209

Bihliography
Wang, G. F., Nikovits, W., Jr., Bao, Z. Z. and Stockdale, F. E. (2001a). Irx4 forms an inhibitory complex with the vitamin D and retinoic X receptors to regulate cardiac chamber-specific slow MyHC3 expression. JBiol Chem 276, 28835-41.
Wang, X., Emelyanov, A., Sleptsova-Friedrich, I., Korzh, V. and Gong, Z. (2001b). Expression of two novel zebrafish iroquois homologues (zirol and ziroS) during early development of axial structures and central nervous system. Mech Dev 105, 191-5.
Wani, M. A., Wert, S. E. and Lingrel, J. B. (1999). Lung Kruppel-like factor, a zinc finger transcription factor, is essential for normal lung development. J Biol Chem 274, 21180-5.
Wieschaus, E., Nusslein-Volhard, C. and Kluding, H. (1984a). Kruppel, a gene whose activity is required early in the zygotic genome for normal embryonic segmentation. Dev Biol 104, 172-86.
Wieschaus, E., Nusslein-Volhard, C. and Kluding, H. (1984b). Kruppel, a gene whose activity is required early in the zygotic genome for normal embryonic segmentation. Dev Biol 104, 172-86.
Wolff, T. and Ready, D. F. (1991). Role of cell birth and death in pattern formation in the developing Drosophila eye. Abstracts o f Papers Presented at the Meeting on Molecular Neurohiology o f Drosophila, 186.
Yang, C. H., Simon, M. A. and McNeill, H. (1999). mirror controls planar polarity and equator formation through repression of fringe expression and through control of cell affinities. Development 126, 5857-66.
Yang, X., Bahri, S., Klein, T. and Chia, W. (1997). Klumpfuss, a putative Drosophila zinc finger transcription factor, acts to differentiate between the identities of two secondary precursor cells within one neuroblast lineage. Genes Dev 11, 1396-408.
Zaffran, S. and Frasch, M. (2002). The beta 3 tubulin gene is a direct target of bagpipe and biniou in the visceral mesoderm of Drosophila. Mech Dev 114, 85-93.
Zaffran, S., Kuchler, A., Lee, H. H. and Frasch, M. (2001). biniou (FoxF), a central component in a regulatory network controlling visceral mesoderm development and midgut morphogenesis in Drosophila. Genes Dev 15, 2900-15.
Zhang, Y., Ungar, A., Fresquez, C. and Holmgren, R. (1994). Ectopic expression of either the Drosophila gooseberry-distal or proximal gene causes alterations of cell late in the epidermis and central nervous system. Development 120, 1151.
Zhao, D., Woolner, S. and Bownes, M. (2000). The Mirror transcription factor links signalling pathways in Drosophila oogenesis. Dev Genes Evol 210, 449-57.
Zhou, Q. and Anderson, D. J. (2002). The bHLH transcription factors 0LIG2 and OLIGl couple neuronal and glial subtype specification. Cell 109, 61-73.
Zinn, K., McAllister, L. and Goodman, C. S. (1988). Sequence analysis and neuronal expression of fasciclin I in grasshopper and Drosophila. Cell 53, 577-87.
210

Bihliography
211

Appendix
This Appendix contains several aspects of our analysis of our microarray experiments. Included are scatter plots to visualise variability, gene lists sorted after the fold change as done originally in the course of this work as well as an analysis of variance between groups of our complete data sets and raw data of 800 randomly selected genes to give a sampling of the chip data.
212

Analysis of variance between replicates
pkb.60.Rep1
-2 2 6 10 J I 1 1 J L_l_
-2 2 6 10 J I I I I I i _
pkb 60 Rep2
pkb 60 Rep3
yw 60 Repi
>w 60 Rep2
T— I— I— I— I— I— r
2 2 € 101— I— I— I— I— I— r
-2 2 € 10T— I— I— I— I— I— r
•2 2 6 10
■2 2 6 10 J I I I L 1— 1
yw.60.Rep3
- '£f
- CN
<N
Figure A; Long heat shock Scatter plots.Comparison of all 3 chips of yw 60min hs (= long hs) to all 3 chips of pkb-
mirror 60 min hs (= long hs). No gross differences were observed, suggesting that the replicates are comparable. The data centres around the 1 to 1 line, showing that the data has been correctly normalized. The scale indicates natural log of signal intensity.
213

Analysis of variance between replicates
pkb.0 .R ep i
1 1 1 1 1 1 . 1 t 1. 1 1 1 I l l ' l l
pkb 0 Rep2
pkb.0.Rep3
yw 0 R ep i
y yw.0.Rep2
yw.O Rep3
-2 2 6 10 -2 2 € 10 2 2 6 10
Figure B: Non heat shock Scatter plots.Comparison of all 3 chips of yw Omin (= non hs) to all 3 chips of pkb Omin (=
non hs). No gross differences were observed, suggesting that the replicates are comparable. The data centres around the 1 to 1 line, showing that the data has been correctly normalised. The scale indicates natural log of signal intensity.
214

Table I: Initial sorting of upreguiated and downregulated potential Mirror target genes using fold change from the yw long hs versus pKB long hs comparison .
Upreguiated genes Downregulated genesProbe Set ID Fold Common Flybase Probe Set ID Fold Common Flybase
143410_at 54.73334218 w FBgn0003996 151217_at 0.010460389 CG14486 FBgn0040741143388_at 9.981389421 &agr;Tut>84B FBgn0003884 148219_at 0.040458903 CGI 0633 FBgn0035604151577 at 7.831544776 GM04393.3prime-hit 145974_at 0.047413186 CGI 3794 FBgn0031936148137_at 4.782350352 CG14985 FBgn0035482 154326_at 0.067518965 CGI 7322 FBgn0032714
151632_i_at 4.553038672 LD12042.3prime-hit 148741_at 0.076842835 CGI 3485 FBgn0036407151568_at 3.954171351 CG11326 GH27479.3prime-hit 143385_at 0.101877378 &agnlry FBgn0003863147588_at 3.784287337 CG9344 FBgn0034564 148424_at 0.131491314 CGI 3311 FBgn0035929
149907_r_at 3.25569798 CG8066 FBgn0038243 150736_at 0.135848269 CG12885 FBgn0039523141333_at 3.215202318 CG6995 FBgn0039229 152598_at 0.150579578 GstEI FBgn0034335154691_at 3.212474235 CG3502 FBgn0034847 146826_at 0.153506286 CG8579 FBgn0033319152727_at 3.152686379 CG7224 FBgn0031971 144863_at 0.155856054 Kap FBgn0030287154132_at 3.127840999 MRG15 FBgn0027378 145545_s_at 0.156144389 CG12506 FBgn0031276
149906J_at 3.018527863 CG8066 FBgn0038243 146001_at 0.158145864 CGI 2560 FBgn0031974142210_at 2.831904632 HLH54F FBgn0022740 143782_at 0.161221816 Cyp4p1 FBgn0015037142969_at 2.735940603 Cys FBgn0004629 152492_at 0.164481636 CG5741 FBgn0035937154506_at 2.690833379 Srp54 FBgn0024285 146050_at 0.171802812 CG9468 FBgn0032069149494_at 2.543855864 CG7352 FBgn0037581 146183_at 0.176095952 CG7294 FBgn0032284142270_at 2.52363565 CGI 7137 FBgn0032308 152786_at 0.185773699 CG8110 FBgn0035826141282_at 2.472733824 exo70 FBgn0035892 143589_at 0.187550764 Mdr50 FBgn0010241153204_at 2.427982753 CG1463 FBgn0030406 153146_at 0.187784243 CG6530 FBgn0034220148780_at 2.386688881 CGI 8581 FBgn0036466 141473_at 0.19431755 keki FBgn0015399154589_at 2.310998549 CGI 8592 FBgn0031678 142538_at 0.195798283 CG17531 FBgn0034341149857_at 2246386212 CG9599 FBgn0038174 151549_i_at 0.196184935 CG9708 GH21465.3prime-hit152162_at 2240928709 BG:DS02780.1 FBgn0028523 142950_at 0.196948783 BG;DS00941.14 FBgn0028533153571_at 2.237940716 XRCC1 FBgn0026751 144349_at 0.20577898 CGI 3361 FBgn0029532152173_at 2.23171516 RNaseX25 FBgn0010406 141701_at 0.211121171 CG2064 FBgn0033205151851_at 2230804617 BcDNA:GH02419 FBgn0027611 143514_s_at 0.211169194 Ccp84Ab FBgn0004782149603_at 2.223439533 CG5359 FBgn0037773 147336_at 0.217364828 CG6426 FBgn0034162143708_at 2210212523 TfllA-S FBgn0013347 152126_at 0.217415078 BcDNA:GH07626 FBgn0027571152431_at 2.169798897 BcDNA:GH12326 FBgn0028480 151702_r_at 0.220836734 CG7288 LD38070.3prime-hit142541_at 2.14669895 CG14048 FBgn0029613 152833_at 0.222192208 Cyp4p2 FBgn0033395149757_at 2.137925093 CG4381 FBgn0038022 141323_at 022427047 CGI 0200 FBgn0033968153296_at 2.131130144 Inos FBgn0025885 145152_at 0.229468987 CGI 7209 FBgn0030687149016_at 2.09290462 CG6852 FBgn0036820 144220_at 0.232951567 BG;DS07851.3 FBgn0028510141428_at 2.07795766 CGI2026 FBgn0035289 151601_at 0.232989984 CGI 7800 HL02328.3prime-hit142591_at 2.055760747 M6 FBgn0037092 143605_at 0.235923868 Drs FBgn0010381145022_at 2.050669028 CG11164 FBgn0030507 142147_at 0.240117068 CG9466 FBgn0032068154499_at 2.034854057 CGI 0492 FBgn0032748 145366_at 0.243515104 CG12199 FBgn0031016152290_at 2.008888351 BcDNA:GH10229 FBgn0027554 146374_at 0.244870661 CGI 3277 FBgn0032599

Table II; Initial sorting of consistently up- and dowregulated potential Mirror target gene; of the combined pKB and pKByw-mirror data.
Fold change Fold changeProbe Set ID Common in pKB in pKByw Flybase
143410._at w 12.17400669 18.08464526 FBgn0003996143388 _at &agr;Tub84B 7.050864123 34.50667039 FBgn0003884153182 _at CGI 7604 2.619111662 2.615305647 FBgn0038362141266 _at CG14965 2.339370455 2.050401648 FBgn0035414149620._at CG12420 2.12639065 2.258773575 FBgn0037797152083 _at CG15093 0.46068551 0.489924442 FBgn0034390143181 _at Gs2 0.441051193 0.358384361 FBgn0001145141323 _at CG10200 0.440655949 0.234443094 FBgn0033968155011 _at CG5095 0.436119723 0.243101001 FBgn0030825149052 _at CG9449 0.427601257 0.155893354 FBgn0036875141330 _at CG6933 0.426068831 0.326073413 FBgn0036952147749._at CG13545 0.421762112 0.406889239 FBgn0034828151605 _at 0.421520933 0.447397336 HL06604.3prime-hit152155._at CG9338 0.415560704 0.397811018 FBgn0032899150369 _at CGI 8594 0.407177916 0.379817984 FBgn0038973147793._at CG4019 0.402420035 0.480131934 FBgn0034885144283 _at BG:DS05899.3 0.401559907 0.293847275 FBgn0028876150159._at CG7714 0.386813912 0.342198357 FBgn0038645143209 _at Kr 0.383448952 0.474764951 FBgn0001325150161._at CG14302 0.379665366 0.367790434 FBgn0038647147636._at CGI 3492 0.378443513 0.467519575 FBgn0034662149968 _at CG5399 0.376228567 0.452168193 FBgn0038353146181 _at CG7299 0.366858784 0.165898112 FBgn0032282147516._at Obp56d 0.344727831 0.140335267 FBgn0034470144254._at Doci 0.343158356 0.333018874 FBgn0028789141658 _at CG5853 0.337664797 0.351147365 FBgn0032167151793._at CG6483 0.327935365 0.418547328 FBgn0035665147002._at CG9070 0.324330575 0.226655218 FBgn0033604149248 _at CG11370 0.324282438 0.342373501 FBgn0037181146958._at CG12934 0.323935544 0.392768124 FBgn0033541150856 _at CG15515 0.313356362 0.151190801 FBgn0039719153941 _at Fst 0.302436028 0.232231788 FBgn0037724
147127 !5_at CG13323 0.287439223 0.345267261 FBgn0033788143296._at CG10598 0.267223268 0.17925989 FBgn0003057147409 _at CG5765 0.265298075 0.242239532 FBgn0034294141435 _at CG11911 0.265292614 0.131811457 FBgn0031249142733 _at CG7298 0.252671675 0.301385639 FBgn0036948145795._at CG8871 0.234310861 0.186394169 FBgn0031653
143340_f_at Ser99Db 0.219069622 0.33296206 FBgn0003357148182 _at CG7465 0.213176301 0.226150258 FBgn0035551144233 _at BG:DS00810.3 0.202503593 0.265574945 FBgn0028536
143339_ i_at Ser99Db 0.19464327 0.32152288 FBgn0003357143338 f_at Ser99Da 0.178487801 0.335045827 FBgn0003356141369._at CG18030 0.177929755 0.2132279 FBgn0039778
150893_f_at CG2229 0.139005886 0.231483142 FBgn0039777
2iU

ANOVA: A N alysis Of VAriance betw een groups
At the beginning of this project only very few “early” chip experiments were performed which did not allow a thorough analysis of the data. Now at the end of the thesis more information can be added to the nature of the genes selected (or not) during the project and which supports the choice for some genes.
This analysis is used to uncover the main and interaction effects of categorical independent variables on an interval dependent variable. A fixed effects linear model was used, of the form
X=p+Ano vaB i+AnovaAj+( Ano vaD * k)+( Ano vaB * Ano vaA)ij
Where p is the average gene expression over all conditions, AnovaB is the heatshock factor, AnovaA is the transgene factor, AnovaD is the background factor, and (AnovaB*AnovaA) is the interaction between heatshock and pkb. i, j and k index the state of the independent variables.
Unless otherwise stated each of the following tables shows the results of the analysis in the order shown below:
1) Probe Set ID- Affymetrix probe set ID2) FDR- Benjamini and Hochberg False discovery rate (used to rank the lists)3) Fold Change- mean(hs)/mean(hsO) or mean(pkb)/mean(pkbyw)4) P value- Welch t-test pvalue5) AnovaA: The probability of the genes expression changing due to the presence or absence of the pkb transgene. These are changes that seem to in pkb chips- whether with heatshock or not.6) AnovaB: Ano va p-value for heat shock effect. The probability of a gene changing under heatshock. This includes genes changing due to Mirror independent and dependent effects.7) AnovaC (AnovaB * AnovaA) : Anova Mirror heat-shock interaction change. These are the genes that we believe to be good candidates for Mirror targets.8) AnovaD: Change due to genetic background. This is the probability of change due to the yw or pkb background.9) Common-Gene name (March 2003 version Affymetrix annotation).10) Flybase-Flybase gene ID11) Description- A gene description (gene name or product or function)
217

Table III: Differentially expressed genes of yw long hs versus pKB long hs comparison %rnProbe Set ID FDR fold Pvalue Anova A AnovaB AnovaC AnovaD Common Flybase Description
143410_at 0.64539918 54.73333856 0.00105679 1.10885E-08 2.86156E-07 3.52086E-06 0.046356549 w FB:FBgn0003996 white141333_at 0.64539918 3.215202171 0.000314216 1.68507E-11 0.049136085 0.47800224 2.78844E-10 CG6995 FB:FBgn0039229 RNA binding151637_at 0.64539918 2.111482876 0.001310942 8.24663E-06 0.159277862 0.633423983 0.293692576 BDGPiLDI 3050.3prime-hit153388_at 0.64539918 1.758109196 0.001119074 0.027493242 1.10269E-05 0.486105409 0.071018411 CG6329 FB:FBgn0033872148450_at 0.64539918 1.587572275 0.000155179 0.003940869 0.022116942 0.948662249 0.207321573 CG4484 FB:FBgn0035968 transporter154755_at 0.64539918 0.70988419 0.001229177 0.089573793 0.001993034 0.057267535 0.061403218 cas FB:FBgn0004878 castor142242_at 0.64539918 0.703236192 0.00130887 0.315431672 0.037270366 0.504607809 0.775396626 CG11419 FB:FBgn0034231152358_at 0.64539918 0.525447715 0.00116174 2.52081 E-08 0.647645221 0.620084413 3.60864E-05 CG8669 FB:FBgn0032941153600_at 0.64539918 0.488673831 0.000981969 2.81205E-05 0.00025505 0.052718868 8.8577E-06 CG5634 FB:FBgn0039528 structural protein143546 at 0.64539918 0.471598916 0.00105345 7.13389E-05 0.635163151 0.72246655 0.208156555 Shal FB:FBgn0005564 Shaker cognate 1153712_at 0.64539918 0.296326153 0.001303446 1.37871E-05 0.850667567 0.59902244 0.042002254 CGI 2325 FB:FBgn0033557 enzyme146216_at 0.64539918 0.245944564 0.000734282 1.12581E-12 0.000664116 0.024427797 0.806575694 CG6105 FB:FBgn0032325 enzyme148741_at 0.64539918 0.076842829 0.001029965 9.92708E-08 0.025391224 0.561687036 0.045525899 CGI 3485 FB:FBgn0036407145974_at 0.64539918 0.047413186 0.001326708 4.54755E-06 0.122476728 0.218100912 0.242011461 CGI 3794 FB:FBgn0031936148219_at 0.64539918 0.040458904 0.00051387 1.71332E-07 0.111653718 0.049282137 0.139866384 CGI 0633 FB:FBgn0035604 ion channel151217_at 0.64539918 0.010460392 1.53704E-05 8.05186E-08 0.024006342 0.000574846 0.360834159 CGI 4486 FB:FBgn0040741149016_at 0.663586972 2.092904749 0.001617152 9.28399E-05 0.002933437 0.201521437 1.684E-07 CG6852 FB:FBgn0036820147919_at 0.663586972 1.642436132 0.001636915 0.000911309 8.31928E-05 0.119982974 0.016078563 CGI 8321 FB:FBgn0035123141417_at 0.663586972 0.807540395 0.001629421 0.334819148 0.043782134 0.77130812 0.150354876 CGI 0007 FB:FBgn0037971154691_at 0.677893313 3.212474623 0.001800565 1 02047E-09 0.000135368 0.039252287 0.000302503 CG3502 FB:FBgn0034847 peptidase154370_at 0.677893313 0.486651504 0.001866103 8.08919E-06 0.02366416 0.154214669 0.021535539 CG14033 FB:FBgn0040946153235_at 0.677893313 0.420765678 0.001950906 0.000345425 0.003469827 0.212171123 0.008904398 Nop60B FB:FBgn0023184 Nucleolar protein at 6B149757_at 0.700005093 2.137924907 0.002302333 0.216403268 0.003557844 0.038302401 0.014208899 CG4381 FB:FBgn0038022143784_at 0.700005093 0.638561224 0.002275676 0.000206916 0.001855821 0.028661737 0.656407244 Fer2LCH FB:FBgn0015221 Ferritin 2 light chain homologue148424 at 0.700005093 0.131491317 0.002186737 0.100760743 0.001729054 0.319270688 0.021970501 CGI 3311 FB;FBgn0035929152431_at 0.712331033 2.16979878 0.002440493 1.84619E-08 0.810905622 0.657414234 7.39721 E-10 BcDNA:GH12326 FB:FBgn0028480152162_at 0.792000679 2.240928595 0.002821984 1.92815E-09 0.031277912 0.111558279 0.563322862 BG:DS02780.1 FB:FBgn0028523 cell adhesion148137_at 0.84129199 4.782350052 0.00367308 1.20615E-09 0.840641152 0.811234075 0.0002792 CG14985 FB:FBgn0035482149494_at 0.84129199 2.543855679 0.003964439 0.000105964 0.226265949 0.304342691 0.000346828 CG7352 FB:FBgn0037581 motor145810_at 0.84129199 1.819923011 0.003490943 0.000117469 0.407143609 0.032748357 0.366938016 CG5828 FB:FBgn0031682155018_at 0.84129199 1.707111252 0.003777335 0.061501633 0.485013322 0.028096348 0.418982595 CG8213 FB:FBgn0033359 endopeptidase144462_at 0.84129199 1.349386624 0.003613193 0.137356922 0.587895706 0.395870046 0.399758404 CG4985 FB:FBgn0029691 DNA binding143850_at 0.84129199 0.730590062 0.00312289 0.671084855 2.14354E-06 0.180764345 0.196095667 Spx FB:FBgn0015818 Spliceosomal protein on the X153961_at 0.84129199 0.689903034 0.004150543 1.9497E-06 0.000365722 0.001379138 0.005631524 Cks FB:FBgn0010314 Cyclin-dependent kinase subunit153818_at 0.84129199 0.570016034 0.003591872 0.001914072 0.115311049 0.19430257 0.603589075 CG7341 FB;FBgn0036777152829_at 0.84129199 0.406231483 0.003906479 6.33181 E-08 0.001366354 0.003605814 0.001576174 CGI 0860 FB:FBgn0035456141701 at 0.84129199 0.211121166 0.004074719 0.004208794 0.01168294 0.069280583 0.151423555 CG2064 FB:FBgn0033205 enzyme154132_at 0.841658404 3.127840978 0.004910014 2.01845E-07 0.382811213 0.356145865 0.098673062 MRG15 FB:FBgn0027378 DNA binding141606_at 0.841658404 1.835423007 0.004703399 0.000206104 0.000221861 0.047750348 0.795165093 ErolL FB:FBgn0028736 signal transduction152569_at 0.841658404 1.53005984 0.005190438 0.020041855 0.118499368 0.482663229 0.969210973 CGI 8003 FB:FBgn0033565142162_at 0.841658404 1.361212832 0.005139129 0.004970467 0.106417558 0.723792953 0.092791598 Man-ll FB:FBgn0011740 alpha Mannosidase II153593_at 0.841658404 0.79337326 0.004924274 1.931 E-05 0.088936581 0.697827025 0.007426535 vig FB:FBgn0024183 vasa intronic gene

Table IV: Differentially expressed genes from yw long hs versus yw non hs comparison
Probe Set ID FDR Fold P value Anova A AnovaB AnovaC AnovaD Common Flybase Description145503_at 0.166702193 0.361253159 9.66922E-05 0.537314066 0.00034179 0.19908358 0.584888325 CG2761 FB:FBgn0031210142836_at 0.166702193 13.4154288 6.15001 E-05 0.001833765 9.16159E-09 0.00491106 0.44715867 Hsp22 FB:FBgn0001223 Heat shock protein 22141274_at 0.166702193 1.975171469 0.000101448 0.033200591 0.000154482 0.089379535 0.515970319 CG14710 FB:FBgn0037920 nucleic acid binding149782_at 0.166702193 76.54468179 0.000119116 0.467611235 0.001408814 0.732581857 0.152347689 CG6489 FB:FBgn0038059 chaperone149314_at 0.166702193 1.57808488 2.9359E-05 0.865255892 1.39409E-06 0.203200491 0.240856154 CGI 2005 FB:FBgn0037301 DNA repair protein150173_at 0.166702193 0.206334624 8.05638E-05 0.046055453 3.219E-07 0.035588061 0.060836841 CGI 4288 FB;FBgn0038669153105_at 0.166702193 2.181558153 0.00010816 0.040862565 0.00219374 0.497097707 0.252522394 CGI 0681 FB:FBgn0036291153616_at 0.231521068 2.163216392 0.000193003 0.463777997 0.01437799 0.397715978 0.030255436 CGI 0425 FB:FBgn0039304 short-chain dehydrogenase/reductase-like145815_at 0.312830504 0.056311701 0.000521309 0.589994298 4.30322E-05 0.174906886 0.093943132 CG6514 FB;FBgn0031692 ligand binding or carrier154305_at 0.312830504 1.682721348 0.000607925 0.064133007 6.48307E-06 0.084209764 3.92971 E-05 CG3167 FB:FBgn0034962153465_at 0.312830504 1.571607127 0.000657138 0.487270111 0.009634367 0.897586446 0.019345134 Cam FB:FBgn0000253 Calmodulin144827_at 0.312830504 0.536065868 0.000583027 0.196675157 0.000709413 0.730258441 0.006731929 CG15211 FB:FBgn0030234 enzyme142705_at 0.312830504 1.375377916 0.000464378 0.535547124 5.86281 E-05 0.339487332 0.000427785 Pros26 FB:FBgn0002284 Proteasome 26kD subunit141236_at 0.312830504 1.626715403 0.000558329 0.069900569 0.201997203 0.601635584 0.057229732 CG4050 FB:FBgn0020312 motor154274_at 0.312830504 1.612169048 0.000436623 0.181197242 9.28178E-07 0.162137059 0.672152547 CG18610 FB:FBgn0034369142864_at 0.312830504 2.643378729 0.000339452 0.094436457 1.37532E-09 0.360645623 0.724816525 NUCB1 FB;FBgn0023185 calcium binding protein143374_at 0.312830504 2.2225502 0.000346299 0.575556157 8.57266E-05 0.783398849 0.62053872 T-cpI FB:FBgn0003676 Top 1-like141652_at 0.312830504 6.804463239 0.000591453 0.124383483 1.94016E-05 0.553022257 0.92992188 CG5290 FB:FBgn0036772143001_at 0.312830504 2.009017584 0.000670591 0.817300577 1.26305E-05 0.274558636 0.000880883 CG5780 FB:FBgn0032446153306_at 0.330542782 2.802123802 0.000765688 0.273281513 1.96264E-07 0.608117491 0.338690651 CG8863 FB:FBgn0038145 chaperone144865_at 0.330542782 0.532097865 0.000787288 0.101721326 2.80418E-08 0.485551103 0.763937192 CG1597 FB:FBgn0030289 enzyme153833_at 0.342473056 1.357861535 0.000897274 0.688757047 0.222408518 0.234598895 0.004988328 CG3918 FB:FBgn0029873147840_at 0.342473056 1.593099531 0.000878764 0.352625728 0.022159859 0.27848777 0.927218181 CG4049 FB:FBgn0034976 enzyme143198_at 0.348542135 6.381457465 0.000954683 0.276097017 8.05997E-08 0.684463967 0.968543883 Hsp83 FB:FBgn0001233 Heat shock protein 83154439_at 0.355464579 3.887193016 0.001090611 0.993881134 2.83792E-07 0.77519699 0.446500586 CGI 0973 FB:FBgn0036306143723_at 0.355464579 0.105902349 0.001083227 0.117148152 0.006833415 0.045460524 0.861791205 Cyp6a9 FB:FBgn0013771 Cytochrome P45-6a9143835_at 0.355464579 5.712694273 0.00112097 0.776850171 5.5201 E-08 0.439777748 0.034716729 DnaJ-1 FB:FBgn0015657 DnaJ-like-1155036_at 0.355464579 2.576252108 0.001142973 0.247292313 3.28752E-06 0.516728889 0.096643183 CG1416 FB:FBgn0032961142756_at 0.364031746 1.868501069 0.001578597 0.505885503 0.000979415 0.971114493 0.59185886 CG2076 FB:FBgn0030263153583_at 0.364031746 99.8201317 0.001306199 0.000886367 3.52369E-10 0.020689304 0.992789295 Hsp23 FB:FBgn0001224 Heat shock protein 23151499_at 0.364031746 2.039541171 0.001422986 0.628143468 0.012440099 0.59725219 0.09898477 BDGP:GH08192.3prime-hit153137_at 0.364031746 1.917432604 0.001628759 0.626272511 5.76102E-06 0.486432469 0.041645261 CG9177 FB:FBgn0030719 translation initiation factor 5151770_at 0.364031746 1.427527513 0.001387285 0.007332524 5.14452E-07 0.110190864 0.037040123 CG7262 FB:FBgn0038274 structural protein154809_at 0.364031746 1.657914086 0.001647399 0.922481196 7.94415E-05 0.167590024 0.032194599 CG11877 FB:FBgn0039636150490_at 0.364031746 2.641645277 0.001469732 0.256324666 1.05114E-08 0.796063328 0.010709134 CG6198 FB:FBgn0039162152287_at 0.364031746 2.310217868 0.00123738 0.644412029 4.12116E-08 0.900888346 0.042730523 mbfl FB:FBgn0026208 multiprotein bridging factor 1153572_at 0.364031746 1.244610441 0.001520913 0.533853627 0.032241326 0.6939426 0.403216012 CG7706 FB:FBgn0038640 phosphatase inhibitor-like154908_at 0.364031746 1.402081958 0.001605158 0.184893713 0.023676137 0.045059687 0.012542116 CG2982 FB:FBgn0029704151019_at 0.364031746 0.681909789 0.001455352 0.939012036 0.015319402 0.455289987 0.375616939 CG8369 FB:FBgn0040532149401_at 0.367664764 1.574320849 0.001707625 0.210942025 0.00651085 0.593374477 0.299596148 CGI 0272 FB:FBgn0037444153731_at 0.387276505 32.95753243 0.001844833 0.188802979 1.64061 E-07 0.462013035 0.62177769 Hsp26 FB:FBgn0001225 Heat shock protein 26149319_at 0.3966391 3.98254529 0.002495104 0.923536097 0.000170905 0.793324847 0.00462514 CG2676 FB:FBgn0037309 galactose-specific C-type lectin-like

Table V: Differentially expressed genes from pKB long hs versus pKB non hs comparison
Probe Set ID FDR Fold Change P value Anova A AnovaB AnovaC AnovaD Common Flybase Description149688_at 0.318778312 0.611844609 0.000911203 0.162637597 0.000377679 0.592395405 0.0249506 CG5281 FB:FBgn0037902152720_at 0.318778312 2.158851461 2.1505E-05 0.166499753 3.6468E-08 0.253391392 0.237424538 CG15187 FB:FBgn0037438 actin binding151754_at 0.318778312 0.725846971 0.000949084 0.446201639 0.032077715 0.764527413 0.021583487 BDGP:SD07967.3prime-hit154500_at 0.318778312 1.302260767 0.000609522 0.35125626 0.89178232 0.161421685 0.378323448 alpha-Spec FB:FBgn0003470 alpha Spectrin145638_at 0.318778312 2.159961571 0.000305294 0.177219138 4.73502E-05 0.727917496 0.290449319 CGI 5390 FB:FBgn0031419153677_at 0.318778312 0.543371878 0.000124729 0.034026271 3.09625E-07 0.014643825 0.007740021 CG2109 FB:FBgn0037330 structural protein of ribosome150348 at 0.318778312 2.154673384 0.000690775 0.263497615 9.72249E-06 0.685450424 0.267908071 CGI 3863 FB:FBgn0038936147775 at 0.318778312 0.630938012 0.000698147 0.001636992 0.005795641 0.676939125 0.412158185 CG9812 FB:FBgn0034860141723_at 0.318778312 0.611890831 0.000438715 0.032530611 9.43202E-05 0.928870822 0.236213309 CGI 2263 FB:FBgn0034346 enzyme143410_at 0.318778312 12.33586641 0.000430949 1.10885E-08 2.86156E-07 3.52086E-06 0.046356549 w FB:FBgn0003996 white149914_at 0.318778312 0.407633377 0.000878393 0.974867407 0.000124452 0.778615904 0.833848903 smp-30 FB:FBgn0038257 Senescence marker protein-3153751_at 0.318778312 1.571943074 0.000461023 0.807292453 0.002498662 0.402596049 0.292258798 CG2116 FB:FBgn0030003 nucleic acid binding153839_at 0.318778312 2.64324045 0.000443046 0.466954107 1.28123E-07 0.399520303 0.708803122 nia FB:FBgn0026629 nebula143313_at 0.318778312 1.544731218 0.000846289 0.000463379 0.011475997 0.282031326 0.000147126 Pros29 FB:FBgn0003150 Proteasome 29kD subunit144192_at 0.318778312 0.605747017 0.000208425 0.259449108 0.007122392 0.792834288 0.427346909 Hey FB:FBgn0027788 Hairy/E(spl)-related with YRPW motif153327_at 0.318778312 1.24858863 0.000109498 0.772684814 0.005458464 0.077349321 0.845605768 TXBP181-like FB:FBgn0026326 motor144169_at 0.318778312 2.316636318 0.00091088 0.115621248 0.017764372 0.998931453 0.153437974 CHIP FB:FBgn0027052 Hsp70-interacting protein14271Q_at 0.318778312 8.865948323 0.000835738 0.02100142 0.000457085 0.675943894 0.033608848 CGI 3761 FB:FBgn0029628154114_at 0.318778312 0.652586945 0.000913023 0.071191785 0.000249111 0.479442384 0.462022611 CG2818 FB:FBgn0031566143367_at 0.318778312 3.496769357 0.000664337 0.373256432 9.83009E-07 0.608458924 0.009508639 Sry-alpha FB:FBgn0003510 Serendipity alpha154415_at 0.323572782 2.511266677 0.001043827 0.090315455 0.000923923 0.999294223 0.781892837 CG3279 FB:FBgn0035156 motor147365_at 0.323572782 1.590937934 0.001064149 0.690733601 1.3821 IE-05 0.163908927 0.219577386 CG6522 FB:FBgn0034223 transcription factor144024_at 0.356112758 1.626982538 0.001229876 3.44488E-07 0.008787379 0.517502733 0.198978569 PRL-1 FB;FBgn0024734 prenylated protein tyrosine phosphatase150155_at 0.358781533 1.762838317 0.001375242 0.767405284 0.002530686 0.306329679 0.002985049 CG7700 FB;FBgn0038637146280_at 0.358781533 1.653455711 0.002395772 0.484281632 0.003877683 0.893616612 0.070874706 CG6583 FB:FBgn0032420143209_at 0.358781533 0.385108831 0.002220311 0.005454367 8.03366E-07 0.003249209 0.017168123 Kr FB;FBgn0001325 Kruppel143000_at 0.358781533 1.642313317 0.0033734 0.000329594 3.14502E-05 0.813009729 0.097795761 CG5525 FB;FBgn0032444 chaperone141550_at 0.358781533 1.162756154 0.001935239 0.068746963 0.036960308 0.467182643 0.96319183 CG2970 FB:FBgn0034936 stomatin-like141359 at 0.358781533 0.715500662 0.002374821 0.021745618 0.000462434 0.21839572 0.022431187 CG3702 FB:FBgn0031590141776_at 0.358781533 2.264854346 0.002930713 0.076181553 0.014266981 0.888492159 0.081399095 CG12428 FB:FBgn0039543 peroxisomal carnitine 0-octanoyltransferase141695 at 0.358781533 2.121755979 0.003318872 0.37806663 0.000133504 0.67100348 0.00495909 CG8241 FB:FBgn0033898 RNA binding141726_at 0.358781533 1.391226064 0.003009081 0.026443127 0.005566095 0.025578606 3.69666E-05 alien FB:FBgn0013746 alien153172_at 0.358781533 0.505232706 0.003336432 0.004671517 0.001271118 0.801080266 0.864303551 CG9524 FB:FBgn0032079155053_at 0.358781533 2.733482483 0.001762274 0.006738455 3.84897E-06 0.066474316 0.514910897 CG8295 FB:FBgn0034051 Myelodysplasia/myeloid leukemia factor154298_at 0.358781533 2.32407443 0.002161404 0.214523054 0.012161375 0.992366584 0.006451226 RhoBTB FB:FBgn0036980 RHO small GTPase154816_at 0.358781533 0.559658682 0.002185333 0.398350886 0.006164261 0.412615207 0.169198658 Actn FB:FBgn0000667 alpha actinin154406 at 0.358781533 2.369247722 0.003104229 0.027080085 2.12108E-06 0.654840883 0.924806226 CG8229 FB:FBgn0033356143314_at 0.358781533 1.923552415 0.002997334 0.001829697 0.000832218 0.30180444 7.1495E-05 Pros35 FB:FBgn0003151 Proteasome 35kD subunit153947_at 0.358781533 2.737884819 0.002652362 0.040165035 0.157084161 0.427891277 0.273731643 twe FB:FBgn0002673 twine154560_at 0.358781533 2.089903743 0.002163736 0.002502153 2.40617E-07 0.464831004 0.074525553 CG17153 FB:FBgn0036248154659_at 0.358781533 1.415116309 0.003104956 0.996666253 3.32747E-06 0.071989852 0.003376187 Cafi FB:FBgn0015610 Chromatin assembly factor 1 subunit142314_at 0.358781533 1.697390408 0.002889193 0.000490327 2.95241 E-05 0.098978122 0.004151025 CG7188 FB:FBgn0035871

Table VI: Differentially expressed genes from pKByw long hs versus pKByw non hs comparison
Probe Set ID FDR FoldChange P value Anova A AnovaB AnovaC AnovaD Common Flybase Description145043 at 0.106049154 0.059213652 3.2862E-05 0.600384416 1.06951 E-05 0.567252962 0.022964539 CGI 2480 FB FBgn0030537 mucin-like proten
143465 f at 0.106049154 0.307525364 2.55038E-05 0.253173667 0.029978711 0.988732822 0.001030006 LysC FB FBgn0004426 Lysozyme C145872_at 0.106049154 0.629568977 4.81778E-05 0.689625123 0.353954145 0.758996806 0.031691649 CG5972 FB FBgn0031781 structural protein148653_at 0.106049154 0.501867624 5.05176E-05 0.577921941 0.001413503 0.313641245 7.91874E-05 CG10418 FB FBgn0036277 LSM2; U6 snRNP core protein147217_at 0.220011099 0.371625969 0.000191874 0.996443911 0.003283477 0.963703042 0.431904696 CGI 8373 FB FBgn0033956148261_at 0.220011099 0.452456781 0.000267069 0.999193897 1.81932E-06 0.111409569 7.76123E-08 CG6610 FB FBgn0035675 LSM5; U6 snRNP core protein150955_at 0.220011099 0.550695189 0.000360468 0.000124295 4.572E-07 0.001566472 0.373953615 CG2191 FB FBgn0039873 sodium/iodide symporter153430_at 0.220011099 0.71268344 0.000393017 0.566287895 0.405712392 0.731933272 0.002248421 CGI 0536 FB FBgn0033456154785_at 0.220011099 0.611695232 0.000164289 0.071982443 0.007614191 0.878384238 0.146245685 CGI 3626 FB FBgn0039212148095_at 0.220011099 0.552133368 0.000287885 0.798147728 0.007720981 0.467076458 0.041610137 CGI 7737 FB FBgn0035423144283_at 0.220011099 0.294558331 0.000378174 0.396946267 1.30586E-06 0.012322149 0.601926167 BG:DS05899.3 FB FBgn0028876 cell adhesion154323_at 0.220011099 0.538165188 0.000376602 0.607897509 0.000119369 0.039671548 0.000579513 CG6617 FB FBgn0030944145902_at 0.220011099 0.458254621 0.000181344 0.778575769 0.009568827 0.106801364 1.25049E-05 CG9548 FB FBgn0031822149208_at 0.220011099 0.351617245 0.000288514 0.815954393 0.002645445 0.427920225 0.065521851 CGI 4572 FB FBgn0037128145626 at 0.227867218 0.612719397 0.000434188 0.991129197 0.523293069 0.593267879 0.066438842 CGI 5387 FB FBgn0031403155112_at 0.288384916 0.635108441 0.000824251 0.049208211 0.275989552 0.479469168 0.000133499 CG7217 FB FBgn0038570148544_at 0.288384916 2.383952563 0.000716438 1.91846E-05 7.71298E-09 0.003742426 1.23548E-05 CG6327 FB FBgn0036115 amino-acid permease-like144254_at 0.288384916 0.332400986 0.000654378 0.605572755 2.26481 E-09 0.002323325 0.222621288 Dorsocross FB FBgn0028789 DNA binding142193_at 0.288384916 0.553888879 0.00076774 0.002775662 1.26392E-07 0.040845612 0.000903467 pnt FB FBgn0003118 pointed153501_at 0.288384916 0.683505644 0.000734305 0.187133934 0.001052719 0.471568927 0.002083979 CGI 3098 FB FBgn0032053149933_at 0.288384916 1.356128095 0.000655087 0.090819594 0.067994083 0.031971949 0.005384914 CG6966 FB FBgn0038286 enzyme inhibitor143410_at 0.288384916 17.73239245 0.000818256 1.10885E-08 2.86156E-07 3.52086E-06 0.046356549 w FB FBgn0003996 white143196_at 0.293826222 8.698157471 0.000874795 0.080481097 6.98419E-05 0.197949957 0.019822824 Hsp67Bc FB FBgn0001229 Heat shock gene 67Bc141203_at 0.298755086 0.564901684 0.001654616 0.010429663 0.066239514 0.477083302 0.297440362 CG2947 FB FBgn0029676 chaperone143109_at 0.298755086 1.940426696 0.001801129 0.544620898 7.44769E-07 0.120808028 0.335360235 Cp18 FB FBgn0000357 Chorion protein 18153589_at 0.298755086 1.853241828 0.001806717 0.570796587 0.021121436 0.599164024 0.306730637 CG12713 FB FBgn0036536
143466_f_at 0.298755086 0.44334963 0.001240897 0.248728756 0.089616216 0.781906248 0.038080858 LysD FB FBgn0004427 Lysozyme D143183_at 0.298755086 0.582385988 0.002021295 3.23569E-07 3.74967E-10 6.58712E-08 0.00038421 gsb FB FBgn0001148 gooseberry146674_at 0.298755086 1.864001514 0.002244373 0.376229461 2.27802E-05 0.782931902 0.299871282 CG11212 FB FBgn0033068 transmembrane receptor154792_at 0.298755086 0.708026063 0.001671161 0.1070915 0.022801196 0.417629851 0.875654006 pnut FB FBgn0013726 peanut147534_at 0.298755086 2.5377605 0.001258033 0.097053824 0.000781995 0.772514639 0.846238026 CG8908 FB FBgn0034493 ATP-binding cassette transporter147816_at 0.298755086 0.558008126 0.002044461 0.99115856 0.014716929 0.83982739 0.116564412 CG5594 FB FBgn0034929 sodium/potassium/chloride symporter153358_at 0.298755086 0.752377355 0.001694015 0.358255828 0.028653595 0.75873303 0.458987297 Nmd3 FB FBgn0023542 Nonsense-mediated mRNA 3143254_at 0.298755086 0.622863784 0.001642479 0.097093887 3.43273E-08 0.395027798 0.586648612 HLHm7 FB FBgn0002633 E(spl) region transcript m7145061_at 0.298755086 15.22199974 0.002244188 0.023496458 0.001124268 0.033753789 0.016132992 CG5228 FB FBgn0030561 structural protein149924_at 0.298755086 1.683827631 0.001532494 0.132179791 0.009560593 0.641437622 0.02139953 CG3641 FB FBgn0038270150308_at 0.298755086 0.807380239 0.00203458 0.935955264 0.422985027 0.614377153 0.189364232 CG6056 FB FBgn0038875 clathrin-associated protein146132_at 0.298755086 0.609211227 0.001699643 9.03427E-08 0.62355205 0.09646777 0.049895772 CG5676 FB FBgn0032200154601_at 0.298755086 0.496558435 0.001197999 0.050344591 0.036882532 0.071530083 0.83701355 CG4155 FB FBgn0038499 RNA polymerase II transcription factor B-like147436_at 0.298755086 0.417824276 0.001166177 0.087556192 0.000545206 0.384833169 0.401529323 CGI 7524 FB FBgn0034337 glutathione transferase141428_at 0.298755086 1.976830051 0.001194283 1.53546E-06 0.000655849 0.055060596 0.084510222 CGI 2026 FB FBgn0035289154533_at 0.298755086 1.233726136 0.002277042 0.573452397 0.001027038 0.304945993 0.10288322 CG11802 FB FBgn0030346

Table VII: Differentially expressed genes of the combined pKB and pKByw-mirror data. Probe Set ID's the false discovery rate and the different Anova results are shown, followed by the Flybase ID, the gene name and a description.
Probe Set ID FDR Anova A AnovaB AnovaC AnovaD Flybase Gene Name Description143209_at 0.358781533 0.005454367 8.03E-07 0.003249209 0.017168123 FB:FBgn0001325 Kr Kruppel154406_at 0.358781533 0.027080085 2.12E-06 0.654840883 0.924806226 FB:FBgn0033356 CG8229143091_at 0.358781533 0.128286554 2.27E-05 0.005935035 0.507107676 FB:FBgn0000233 btd buttonhead152355_at 0.358781533 0.002345151 6.31 E-06 0.228633163 0.000486449 FB:FBgn0015828 TfllEalpha Transcription factor IIEalpha142469_at 0.365273892 0.211290203 1.52E-05 0.070177934 0.198937257 FB:FBgn0000307 chif chiffon154404_at 0.365273892 4.69E-07 1.60E-06 0.448010234 0.24776182 FB:FBgn0033021 CG10417 protein phosphatase154633_at 0.365273892 0.89960828 0.122886738 0.626823226 0.838722959 FB:FBgn0002715 mei-8332 meiotic 8332154323_at 0.365273892 0.607897509 0.000119369 0.039671548 0.000579513 FB:FBgn0030944 CG6617147556_at 0.370723804 0.413822889 0.042076645 0.007616024 0.008663859 FB:FBgn0034525 CGI 3435 ligand binding or carrier148717_at 0.370723804 0.141120731 0.009368702 0.196499582 0.45519033 FB:FBgn0036372 CGI 0083 cell cycle regulator154728_at 0.370723804 0.423449887 0.912619552 0.59041096 0.002748133 FB:FBgn0032236 CG5108 structural protein of ribosome153589_at 0.370723804 0.570796587 0.021121436 0.599164024 0.306730637 FB:FBgn0036536 CG12713141731_at 0.370723804 0.014390516 1.45E-06 0.83282134 0.331450703 FB:FBgn0039176 CGI 3610 transporter153283_at 0.370723804 0.141002836 0.000340992 0.151011079 0.005896283 FB:FBgn0019644 ATPsyn-b ATP synthase, subunit b142270_at 0.377344616 0.139359306 0.004880238 0.090812716 0.017140015 FB:FBgn0032308 CG17137 ion channel146639_at 0.377344616 0.723483559 0.719015387 0.818408532 0.324125471 FB:FBgn0033017 CGI 0465 ion channel147768_at 0.37935578 0.580878124 0.551864903 0.596268986 0.139837512 FB:FBgn0034851 CG11079153828_at 0.37935578 0.643362012 0.000900054 0.81456443 0.182598639 FB:FBgn0032136 CGI 5828 ligand binding or carrier142193_at 0.386321455 0.002775662 1.26E-07 0.040845612 0.000903467 FB:FBgn0003118 pnt pointed153759_at 0.388782172 0.436989908 0.368222816 0.220728281 0.016806998 FB:FBgn0036854 CG9750151788_at 0.403308595 0.474306771 6.91 E-07 0.070298388 0.043574956 FB:FBgn0038546 CG7379 enzyme152110_at 0.409245406 0.549377983 0.619589312 0.300759586 0.100340303 FB:FBgn0038806 CG5412 dihydrofolate reductase153100_at 0.409245406 0.242716985 0.003199678 0.136125359 0.044789217 FB:FBgn0038583 CG7183154422_at 0.413388371 3.86E-05 0.001888009 0.006670284 0.172925048 FB:FBgn0035130 CG6994 cell adhesion143523_at 0.419448126 0.045267908 1.09E-06 0.30183783 0.006960329 FB:FBgn0004862 bap bagpipe143363_at 0.420668327 0.000356722 0.00242535 0.981006283 0.731393025 FB:FBgn0003483 spn-E spindle E152195_at 0.420668327 0.869480071 0.04866774 0.685016893 0.030393894 FB:FBgn0011715 Snri 8nf5-related 1154099_at 0.420668327 0.002999334 4.70E-05 0.062797802 0.001829973 FB:FBgn0000257 car carnation147221_at 0.420668327 0.543849181 1.29E-05 0.295774718 0.012027107 FB:FBgn0033961 CGI 2859153498_at 0.420668327 0.085327955 0.526694741 0.195841618 0.544003382 FB:FBgn0027897 BcDNA:LD03471150741_at 0.420668327 0.379098648 0.012729814 0.762484901 0.204965588 FB:FBgn0039531 CG5611 enzyme147919_at 0.420668327 0.000911309 8.32E-05 0.119982974 0.016078563 FB:FBgn0035123 CGI 8321155104_at 0.420668327 0.383507076 0.00351346 0.727593902 0.033338345 FB:FBgn0034451 CG11242 motor142999_at 0.422142515 0.096443098 0.046976844 0.061919107 0.893603267 FB:FBgn0010280 TafllO TBP-associated factor IlkD151063_at 0.422142515 0.078374742 5.21 E-05 0.031797191 0.0001984 FB:FBgn0040575 CGI 5922154428_at 0.423022245 0.180778931 0.241445567 0.737037701 0.012166021 FB:FBgn0038411 CG6963150083_at 0.424136246 0.972494526 0.559788664 0.581376671 0.848678884 FB:FBgn0038522 CG18139153182_at 0.424822491 0.124524497 0.005884591 0.015618716 0.015303255 FB:FBgn0038362 CGI 7604 motor143307_at 0.437929229 0.038562941 0.002790242 0.298576093 0.049198254 FB:FBgn0003114 plu plutonium153395__at 0.437929229 0.153683754 0.018185214 0.29922991 0.050857581 FB:FBgn0000229 bsk basket154073_at 0.437929229 0.869434987 0.000584883 0.887559283 0.01415254 FB:FBgn0035524 CG11583150248_at 0.45175177 0.005917197 0.165264949 0.091110001 0.285571881 FB:FBgn0038780 CG5060

Table VIII: Differentially expressed genes from yw short hs versus yw non hs comparison
Probe Set ID FDR Fold P value Anova A AnovaB AnovaC AnovaD Common Flybase Description147928_at 0.039473049 2.417620033 1.88034E-05 0.231724001 5.3731 E-06 0.25070096 0.323704208 CG6905 FB FBgn0035136 cell cycle regulator152518_at 0.039473049 2.11396771 1.18807E-05 0.721855416 2.20381 E-06 0.361886614 0.31102141 CG8231 FB FBgn0030681 T-complex protein 1, zeta-subunit142836 at 0.039473049 63.93765878 1.07686E-05 0.001833765 9.16159E-09 0.00491106 0.44715867 Hsp22 FB FBgn0001223 Heat shock protein 22147373_at 0.039473049 1.579893789 1.2889E-05 0.978005321 5.39364E-09 0.297588365 0.102084166 CG4878 FB FBgn0034237 translation initiation factor 3-beta142644_s_at 0.039473049 0.632642886 1.52167E-05 0.047267485 0.015347419 0.960927343 0.65358041 CGI 7552 FB FBgn0037211149937_at 0.053286876 0.673724869 3.80757E-05 9.25048E-05 0.001896614 0.406862444 0.002882855 CG6912 FB FBgn0038290148159_at 0.053286876 1.895885186 3.7762E-05 0.037666071 3.83504E-05 0.448388325 0.969054592 CGI 5010 FB FBgn0035516 enzyme154846_at 0.068766567 2.088415055 5.73259E-05 0.08437703 0.010638037 0.858940348 0.000238436 CGI 7952 FB FBgn0034657151578_s_at 0.085044662 6.462936029 0.000111216 0.793039145 1.2332IE-07 0.196460535 0.114150969 CG14897 BDGP:GM04711.3prime-hit motor143657_at 0.085044662 1.545873464 0.000113624 0.057223145 7.77727E-06 0.181639032 0.184256108 OstStt3 FB FBgn0011336 Ollgosaccharyl transferase 3149586_at 0.085044662 1.302003394 9.60777E-05 0.010984353 0.005881159 0.466997787 0.416974787 CG8495 FB FBgn0037752 ribosomal protein S29-llke154274_at 0.085044662 1.855148314 0.00012082 0.181197242 9.28178E-07 0.162137059 0.672152547 CG18610 FB FBgn0034369142982_at 0.085044662 1.706981243 0.000133342 0.181238339 2.72154E-06 0.470460593 0.013846744 Pros26.4 FB FBgnOO15282 Proteasome 263 subunit subunit 4151049_at 0.085044662 4.202776501 0.00013314 0.008871092 5.36876E-06 0.361065133 0.148291486 CG14873 FB FBgn0040562148718_at 0.085044662 1.932895064 0.000141792 0.000820475 0.000152081 0.143626529 0.177570672 CGI 0741 FB FBgn0036373146769_at 0.085123729 3.257513016 0.000162198 0.014943721 7.56358E-08 0.400570334 0.823223913 CGI 553 FB FBgn0033224154058_at 0.085123729 3.280122389 0.000154481 0.232747267 0.000307811 0.564550707 0.050283989 CG5295 FB FBgn0036449143835_at 0.086474938 21.5315268 0.000175467 0.776850171 5.5201 E-08 0.439777748 0.034716729 DnaJ-1 FB FBgn0015657 DnaJ-llke-1145560_at 0.086474938 0.53254105 0.00018537 0.274290427 0.004951883 0.758939513 0.046116701 CG4726 FB FBgn0031307 sodium/phosphate cotransporter142167_at 0.093870032 1.322075227 0.000234759 0.499625051 0.062831224 0.271231048 0.727925411 CGI 7259 FB FBgn0031497 serlne-tRNA llgase-llke152862_at 0.093870032 0.461382046 0.000227986 0.006700506 8.22996E-06 0.395963782 0.01339704 CG5805 FB FBgn0039223 mitochondrial carrier protein-like143197_at 0.093870032 238.3428133 0.000225485 0.107355065 2.54453E-08 0.321151823 0.064025476 Hsp68 FB FBgn0001230 Heat shock protein 68142977_at 0.099660391 3.681058904 0.000261109 0.002413891 2.5892E-10 0.585419427 0.00813454 CG7033 FB FBgn0030086 T-complex protein 1, beta-subunit142683_s_at 0.101921114 1.583977704 0.000279169 0.022828558 2.27871 E-06 0.705722161 0.939176982 Amy-p FB FBgn0000079 Amylase proximal147521_at 0.10342693 0.059595618 0.000296028 0.069736883 0.000234881 0.159094257 0.190217686 CGI 3874 FB FBgn0034475 ligand binding or carrier151463_at 0.10342693 1.877116564 0.000307928 0.412288916 0.009623009 0.737233815 0.044727232 CG6890 FB FBgn0041571149260_s_at 0.109849726 1.334307938 0.000423159 0.017456739 0.000154727 0.096419451 0.364883406 CG10712 FB FBgn0037201 DNA binding151770_at 0.109849726 1.602871476 0.000480441 0.007332524 5.14452E-07 0.110190864 0.037040123 CG7262 FB FBgn0038274 structural protein149782_at 0.109849726 87.30469298 0.000459553 0.467611235 0.001408814 0.732581857 0.152347689 CG6489 FB FBgn0038059 chaperone141541_at 0.109849726 0.249885895 0.000420538 0.335768614 0.000235887 0.305041425 0.081206692 CGI 3868 FB FBgn0034501149908_at 0.109849726 15.32258472 0.000497117 0.040641584 8.62575E-08 0.291443428 0.020173855 CGI 8485 FB FBgn0038245153308_at 0.109849726 1.502595159 0.000427467 0.390163263 0.005288782 0.925832295 0.707900837 bio FB FBgn0000181 bicaudal154694_at 0.109849726 2.366333861 0.000387469 0.015363626 1.16865E-06 0.481303728 0.755562453 CG3428 FB FBgn0036005151467_at 0.109849726 1.407743459 0.000392627 0.007432247 2.46494E-05 0.036216023 0.173194935 BDGP:GH01059.3prime-hit145386 at 0.109849726 1.465614818 0.000374598 0.138174658 0.000262123 0.278545842 0.060504337 CGI 2234 FB FBgn0031051 transmembrane receptor152720_at 0.109849726 1.911604429 0.000473528 0.166499753 3.6468E-08 0.253391392 0.237424538 CG15187 FB FBgn0037438 actin binding153821 at 0.109849726 1.536127248 0.000493351 0.649433211 0.000629362 0.629479107 0.950924018 CGI 6975 FB FBgn0032475 transcription factor151287_at 0.109849726 0.482029347 0.000391074 0.221129688 0.00030491 0.782181086 0.324161382 CG11051 FB FBgn0040813142544_at 0.109849726 2.035873245 0.000478663 0.493087839 0.002961331 0.128062099 0.744230363 CG7372 FB FBgn0036522 nucleic acid binding154285_at 0.112701518 1.481763868 0.000583382 2.85962E-06 0.004294493 0.13356103 0.000109255 CG5522 FB FBgn0034158 guanine nucleotide releasing prote153583_at 0.112701518 117.5730207 0.000578905 0.000886367 3.52369E-10 0.020689304 0.992789295 Hsp23 FB FBgn0001224 Heat shock protein 23145994_at 0.112701518 2.455920286 0.000590305 0.706748117 2.31243E-07 0.633750821 0.794356684 CGI 7973 FB FBgn0031963

Table IX: Differentially expressed genes from pKB short hs versus pKB non hs comparison
#7«S
Probe Set ID FDR Fold Change P value Anova A AnovaB AnovaC AnovaD Common Flybase Description144031 .at 0.075742 1.733046783 0.000110947 0.782686074 0.000187972 0.060350712 0.059285986 I s FB:FBgn0024920 thymidylate synthase151121 .at 0.075742 0.63996906 0.000152466 0.352289267 0.068284684 0.980437381 0.447036434 CGI 3255 FB:FBgn0040636154524, at 0.075742 1.335973638 7.71012E-05 0.000444705 0.000583617 0.063598618 0.366370352 CGI 6728 FB:FBgn0033539 GAP-like protein144626..at 0.09816586 1.670073776 0.000491005 0.015815546 0.000144356 0.058486049 0.059955383 CG4564 FB:FBgn0029916150068. at 0.102203868 2.745671472 0.000559888 0.087923704 0.008096956 0.015125925 0.945878401 CG5419 FB:FBgn0038502151831. at 0.114750462 1.439890018 0.000752841 0.001605109 0.853874119 0.189372025 0.068021425 CG3459 FB:FBgn0033489 transcription factor143011..at 0.114750462 0.243794533 0.00076841 0.280223432 0.49887903 0.094449905 0.659995442 CG4822 FB:FBgn0031220 ATP-binding cassette tra152004..at 0.114750462 1.837187185 0.000731191 0.053949514 7.74465E-06 0.005588149 0.292982044 cli FB:FBgn0000320 Clift142830..at 0.115142346 2.064244808 0.001160413 0.000534226 0.01199117 0.003184698 0.679664559 kis FB:FBgn0001316 klarsicht154588. at 0.115142346 0.460478032 0.001158918 0.935632596 0.228415223 0.555210064 0.045298947 LamC FB:FBgn0010397 Lamin C141683 .at 0.115142346 1.545264903 0.001034748 0.004306775 0.048132505 0.217121037 0.119775429 CG5874 FB:FBgn0038872147230. at 0.115142346 1.379278006 0.00135038 0.458958764 0.056731745 0.645652428 0.643970848 CGI 0265 FB:FBgn0033990154506. at 0.115142346 1.924735401 0.001399738 2.70913E-12 0.00136448 0.000177782 0.052537545 Srp54 FB:FBgn0024285 splicing factor, SRp54151780..at 0.115142346 1.441218824 0.001136295 0.161398959 0.081000424 0.027629648 0.097904535 CG7993 FB:FBgn0038585150910 .at 0.115142346 1.773526328 0.001294762 0.008954388 0.002285539 0.217733658 0.05027885 CGI 349 FB:FBgn0039802 enzyme143472..at 0.115142346 1.447161428 0.001667745 0.183486347 0.539530356 0.409014634 0.200051947 Ten-m FB:FBgn0004449 Tenascin major146215 at 0.115637842 0.45457612 0.001776501 0.712667339 0.216513652 0.4910666 0.12973793 CG6093 FB:FBgn0032323 signal transduction141413. at 0.117130401 1.957417397 0.002227458 0.001468659 0.011486615 0.045647325 0.000314615 CG4699 FB:FBgn0038364141498..at 0.117130401 1.281196659 0.002181329 0.039708493 0.273457618 0.417794942 0.453169899 brk FB:FBgn0024250 brinker154017. at 0.117130401 1.689500764 0.002034762 0.058611014 0.001083068 0.447497949 0.385692614 BcDNA:LD21504 FB:FBgn0027526141708. at 0.117130401 1.429635773 0.002193197 7.13891 E-05 0.120602215 0.000170341 0.011281782 CG7971 FB;FBgn0035253 RNA binding protein150478. at 0.117130401 1.78886902 0.001889446 0.753666992 0.010266637 0.187366891 0.522196383 CG5410 FB:FBgn0039140 cell cycle regulator143955..at 0.117130401 0.061145757 0.001915735 0.764713619 0.002572595 0.218651364 0.045888365 CGI 7467 FB:FBgn0022341150379. at 0.117130401 1.287331482 0.001893169 0.347826904 0.269104288 0.717492507 0.182081789 CG6937 FB:FBgn0038989 RNA binding protein-like155047. at 0.117511761 1.416261104 0.002281102 0.016363193 0.014853379 0.340858664 0.020226037 Tbp FB:FBgn0003687 TATA binding protein145452 at 0.11827877 2.058748406 0.002420905 0.000477494 0.001073247 0.003694237 0.046315385 CGI 5449 FB:FBgn0031146147788. at 0.119184247 0.745744311 0.002462081 0.872358685 0.030103725 0.502963708 0.005170583 CG5365 FB:FBgn0034875152887..at 0.120852318 0.68321302 0.002631869 0.372606599 0.013062259 0.279385308 0.239932395 CG11388 FB:FBgn0034959152295 .at 0.122268232 1.445813076 0.00278114 0.117644098 0.002728594 0.348263745 0.573427554 BcDNA:GH10333 FB:FBgn0027553153823 .at 0.124327998 1.697213354 0.003242567 5.1092E-06 0.000467862 0.00011823 0.534401873 CG8073 FB;FBgn0033377 phosphomannomutase-li154073. at 0.124327998 1.255476831 0.002993688 0.869434987 0.000584883 0.887559283 0.01415254 CG11583 FB:FBgn0035524
144630_s5_at 0.124327998 1.698203837 0.003107824 0.00719183 4.32776E-05 0.003136761 0.006000246 CG4575 FB:FBgn0029920 transcription factor142201 .at 0.125355962 0.374440477 0.00334402 0.372899722 0.012795936 0.30452151 0.826557051 CG4363 FB:FBgn0034663152614. at 0.126858769 0.089058273 0.003465259 0.98093162 0.048413337 0.438413489 0.430137318 CG14021 FB:FBgn0031702154901..at 0.126858769 0.700472558 0.003405913 2.56966E-05 0.254792963 0.050298266 0.111809615 CG7319 FB:FBgn0036197154594 .at 0.126858769 1.301701054 0.003493154 0.002642963 0.012363142 0.199366624 0.110300691 Rnp4F FB:FBgn0014024 RNA-binding protein 4F154747. at 0.12790434 0.430987598 0.003813066 0.457380641 0.023482381 0.344786696 0.197649098 CGI 5106 FB:FBgn0034406 enzyme146919. at 0.12790434 0.660919656 0.003757554 0.250879371 0.005760579 0.102964209 1.49 IE-05 CGI 2921 FB;FBgn0033480151406 at 0.129518783 2.004383626 0.00528486 0.06121643 0.006825067 0.540862245 0.168566421 CGI 2647 FB:FBgn0040939151797 at 0.129518783 0.440352619 0.004657022 0.00486931 0.001202968 0.661152039 0.10097151 CG10611 FB:FBgn0032820 enzyme153324 .at 0.129518783 1.279015605 0.004722115 0.012286862 0.009544186 0.126598019 0.013968609 Su(fu) FB:FBgn0005355 Suppressor of fused143207 at 0.129518783 1.242648638 0.004768359 0.000133276 0.004653 0.587924881 0.012788287 kn FB:FBgn0001319 knot141737..at 0.129518783 1.990067025 0.004102536 0.004555148 0.000335936 0.038117067 0.161539425 CG8491 FB:FBgn0036912 transcription factor

Table X: Number of significant genes in each of the Anova categories.
Sig Level AnovaB AnovaA AnovaDAnovaC Expected0.05 4103 1906 2295 440 4560.01 2642 999 1121 100 91
0.001 1352 435 382 21 90.0001 670 202 133 4 1
For the 100 genes of interest atp<.01Sig Level lAnovaB AnovaA AnovaD
0.01 71 60 300.001 55 40 13
0.0001 41 25 8
For the 21 genes of interest at
Sig Level AnovaB AnovaA AnovaD0.001
0.000114 11 4 10 11 3
Anova A and D are very similar, supporting the belief that the attempt to cross pkb into the yw background was unsuccessful. This data also suggests that the number of true Mirror targets induced by ectopic Mirror and detected in this experimental paradigm is small (Anova C). The four highly significant genes in this category are the genes: white, gooseberry and the uncharacterised genes CG8800 and CGI 677. In the course of this work it could be shown that gooseberry expression can be repressed by ectopic Mirror protein in vivo. However,no alterations in gooseberry expression were observed using mirror loss-of-function embryos (see page 123). The gene white is highly upregulated probably due to the presence of the miniwhite gene within the same cassette as the mirror heatshock construct. The two uncharacterised genes have not yet been analysed. The gene Kriippel which is most likely a true Mirror target can be found within the 100 genes of significant level 0.01 in the Anova C category. Several other genes of this group have been analysed but don't seem to be real Mirror targets. Increasing the number of replicates, to allow a more powerful statistical analysis could be helpful to identify true target genes.
225

Table XI; Correlation Matrix.Normalised Signal intensity measurements were filtered to exclude
measurements from genes that were called Absent in at least 22/24 cases across all chips. This reduced the list from 14010 to 8397 elements. A matrix was constructed using Pearson's Product Moment Correlation Coefficient (R) applied to the mean of the measurements in each replicate group. YWO = yw non hs, YW30 = yw short hs, YW60 = yw long hs, PKBO = pKB non hs, PKB30 = pKB short hs, PKB60 = pKB long hs, PKB YWO = pKByw non hs, PKB YW 60 = pKByw long hs.
YW30 PKB60 PKBO PKB30 PKBYW60 PKBYWO YW60 YWOYW30PKB60PKBOPKB30PKBYW60PKBYWOYW60YWO
1 0.957118 0.937421 0.983516 1 0.92994 0.958299
1 0.933075 1
0.9688264 0.934438 0.970792 0.939709 0.966622 0.92607 0.983247 0.925745
0.9139972 0.971714 0.946342 0.979336 0.9633094 0.914354 0.95535 0.915474
1 0.928884 0.961741 0.919962 1 0.947244 0.986343
1 0.956067 1
Tree Diagram for 8 Variables Single Linkage
1-Pearson r
YW 30
PKB3U
PKBBO
YweoPKBYWBO
PKBO
PKBYWO
0.010 0.015 0.020 0.025 0.030
Linkage Distance
0.035 0.040 0.045
Figure C. Clustering Based Correlation Matrix.Hierarchical clustering was performed on the table of 1-R distance
measurements using Single Linkage clustering. The Tree Diagram illustrates that experiments cluster according to the different treatments and that the distance between each replica group is very small. YWO = yw non hs, YW30 = yw short hs, YW60 = yw long hs, PKBO = pKB non hs, PKB30 = pKB short hs, PKB60 = pKB long hs, PKBYWO = pKByw non hs, PKBYW 60 = pKByw long hs.
226

Table XII : Raw data of 800 randomly selected genes
Probe Set ID
ywnonhsi
yw yw yw yw yw yw pKU pKH pKU pKU pKU pKb pKU pKU pKU non hs non hs short hs yw short yw short long hs long hs long hs non hs non hs non hs short hs short hs short hs long hs long hs long hs
2 3 1 h s 2 h s 3 1 2 3 1 2 3 1 2 3 1 2 3PKBYW non hs 1
PKBYW non hs 2
PKBYW non hs 3
PKBYW long hs 1
PKBYW long hs 2
PKBYW long hs 3 Flybase
150092_at 115.8 155.8 69.7 107.4 90.4 153.9 103.2 78 122.8 142.1 84.5 101.2 162.2 61 89 15.2 94.1 103.7 139.4 112 96.1 216 316.8 104.6 FB FBgn0038532154940_at 616.9 634.6 819.4 878.4 900.4 835.4 838.7 865.1 775.2 710.4 753.8 666.7 918.7 848.8 754.1 817.9 759.1 636.8 578.9 617.4 578.1 562.4 539.9 557.4 FB FBgn0036487150641_at 241.6 196.5 333.7 282.8 284.5 257.7 454.4 157.7 258.7 468.2 420.3 236.5 192.3 193.4 207.6 370.2 249.4 199.6 237 186.5 257.9 165.9 250.7 351.1 FB FBgn0039404147605_at 376.3 313.9 309.7 149.5 315.1 261.2 518.7 297.4 227.8 689.7 383.1 376.7 332.1 341.6 322.4 616.3 408.3 551.9 221.2 210.3 215 260.3 267.2 289.6 FB FBgn0034600148372_at 6.7 18.7 6.7 20.1 7.9 13.3 15.2 7.1 7.9 8 13.4 6.3 13.3 9.1 18.1 8.2 6.5 8 9.1 7.5 4.8 20 11.7 13 FB FBgn0035834147162_at 17.9 13.6 7.7 9.3 18.6 10.5 12.2 9.3 5.8 14 10.1 16.3 10 16.5 10.4 9.6 6.2 13.3 6.5 5 6.2 15.9 63.7 13.5 FB FBgn0033861147104_at 420.8 377.9 464.1 263.6 271.9 273.5 560.5 362.4 420.9 427.5 310.1 257 221.2 248.3 141.3 363.2 322.6 380.9 397.2 417.3 426.2 260.7 246 410.1 FB FBgn0033754149377_at 35.3 19.8 148.7 11.4 59.6 76.4 19.3 18.1 28.6 178.5 27.8 84.8 79.9 63.8 25.3 40.3 14.8 25.2 16 11.1 50.6 36.7 30.2 33.3 FB FBgn0037411150320_at 129 16.2 34.7 168.1 172.1 161.6 180.3 102.4 62 52.4 32.5 88.7 71.2 61.6 212.5 63.3 115 125.5 60 120.6 46 80.9 119.5 83.6 FB FBgn0038895143647_at 53.4 37.1 75 75.4 53.3 76.2 56.6 52.1 6.1 73.2 28.7 19.7 51.7 57.7 91.3 66.1 46.8 39.9 24.5 41.9 38.2 55.6 34.7 4.2 FB FBgnOO11281151599_at 144.5 223.3 213.3 324.6 248.4 284.6 138.1 130.6 223.7 238.3 297.1 236.5 448.3 586.8 437.4 262 229.1 241.7 255.5 272 301.6 657.3 717.1 332.8 BDGP:HL01913.3155001_at 229.3 275.6 226.2 217.4 197.8 221.6 223.8 287.2 308.9 265.7 229.4 139.7 181.1 178.8 169.1 232.6 351.8 340.1 266.3 242.9 337.2 316.8 237.9 450.6 FB FBgn0033677147236_at 86.7 137.2 160.5 63.5 97.4 138.3 237.8 138.8 114.9 98.1 162.5 73.7 37.4 76.5 30.6 139.5 150.2 185.2 93.6 35.4 99.2 119.1 61.5 81.1 FB FBgn0034001147489_at 167 129.4 233.7 235.9 304.9 202.6 160.5 365 195 113 162.4 35.9 203 205.8 259.4 284.2 271.1 281.3 132.5 218.7 137.7 177.9 318.8 156.1 FB FBgn0034433148079_at 171 364.8 25.6 210.1 180.1 149.3 54.1 18.4 63.8 19.8 177 200.2 20.3 19 8.2 10 71.7 66.5 282.8 158 236.7 161.3 86.7 428.4 FB FBgn0035398154827_at 1418 1407 800.2 1928.6 2226.3 2298.4 1863.5 1703.8 2162 761.6 956.4 1303 2179 2154.7 1958.1 2104.8 1828.4 1759.2 1086.7 1279.1 1083.2 1606.1 2211.8 2589.7 FB FBgnOO16756142139_at 467.7 432.8 220 274.1 300.9 228.7 420.2 162.4 192.4 351.2 437.6 546 173.1 305.6 249.6 296.8 194 244.7 656.2 363.2 745.8 390.2 204.4 667.4 FB FBgn0029549152002_at 244.6 455.7 321.8 208.1 207 216.5 274 131.5 336.6 354.4 324.1 322.1 184.5 140 203.1 239.9 281.8 240.7 547.5 440.5 441.2 300.1 280 332.5 FB FBgn0004880154398_at 147.3 212.5 312.1 166.9 211.6 186.2 221.7 246.8 248.7 271.5 246.9 166.7 173.2 216 204.8 169.7 228.5 115.7 194.7 236.7 202.7 279.7 349.1 149.4 FB FBgn0034002154365_at 438.3 608 639.6 884.4 730.9 561.8 726.4 1013.5 754.3 748.3 735.1 640 862.9 653 780.1 995.5 1012.8 928.9 680 663.4 640.8 1046.4 1193.9 834.5 FB FBgn0039733141294_at 61 64.3 202.6 89.4 84.3 128.7 76.3 170.6 216.1 72 97.4 87.2 89.2 59.9 63.2 161.3 135.4 112.8 42.3 121.6 24.9 69.1 99.9 58.3 FB FBgn0035945142981_at 196.4 218.1 165.4 199.8 148.3 215.3 130.9 283.5 228.6 165.9 167.2 203.7 145.6 173.1 208.6 154.3 238.7 220.8 124.8 202.5 208.5 211.2 98 219.8 FB FBgn0029778152730_at 179.2 266.6 142.7 134.8 145.1 155.3 135.8 243 269.2 106.6 135 136.3 73.3 117.2 74.2 138.9 176.5 224.6 156.3 210.6 161 214.6 200 285.1 FB FBgn0038344153684_at 1972 2181 2265 2071.4 2113.5 2072.2 1813 3134.8 2798.4 1371 1307 1432 1698 1618.9 1597.9 1912.6 2514.1 2228.4 1610 1595.2 1534.9 1357 1861.7 1560.3 FB FBgn0000250154023_at 2117 1589 2265 2488.4 2569.1 2641.7 2747.7 2132.5 1948 2389 2038 2223 2690.9 2955.1 2777.4 2561.9 2274.2 2447.6 2076.2 1727.6 2101.3 1954.9 2099.8 2196.1 FB FBgn0015283146667_at 12.8 22.5 35.7 37.4 39.2 47.7 12.9 14.8 13.8 22.4 8.3 105.5 47.3 14.4 61.9 19.9 16.7 57.1 9.8 12.6 9.6 87 24.7 16.1 FB FBgn0033056147553_at 53.7 83.1 85.6 16 74.7 63.5 32.9 47.1 42.2 81 46.4 52.3 48.5 54 39 47.5 52.7 101.3 47.6 93 48.7 97.2 87.7 49 FB FBgn0034520153304_at 107.1 70.9 74.3 69.3 95.6 76.7 85.7 77.9 79.6 28.7 79.7 47.3 56.2 31.6 98.9 26.9 78.3 64.2 42 54.2 52.7 139.8 59.9 78.2 FB FBgn0026263146515_at 2.8 53.5 6.5 5.7 3.1 1.2 7.2 4.7 1.3 8.4 5 3.8 5.2 2.4 2.4 5.7 2.4 9.3 5.5 2.4 2.3 4.7 7.8 2 FB FBgn0032824145179_at 1307 1172 1293 1603.3 1530 1705 1200.4 1551.3 1483 1040 1216 1292 1708.7 1432.2 2094 1398.5 1614.6 1157.5 1361.9 1391.7 1467.6 1361.3 1599 1333.5 FB FBgn0030738144595_at 2929 2516 2664 2609.7 2942.7 2387.7 2680.8 2025.4 2084.6 3374 4152 3997 3451.3 3010.7 2805.1 2428.5 2875.3 2356.1 3529 2890 3667.9 2047.7 1721.3 3002.9 FB FBgn0029868147122_at 716.5 483 461.5 453.4 577 416.1 528.8 645.1 508.6 164.4 605 440.8 348 485.9 273.9 460.3 426.8 477.6 317 385 333.3 363.1 432 302.3 FB FBgn0033781151054_at 12.9 7.3 3.9 24 3 37.1 2.9 2.5 3.7 2.9 1.3 13.9 3.6 4.8 39.3 16.3 2.6 26.6 21.7 3.1 23.4 50.3 4.2 1.1 FB FBgn0040567149814_at 472.7 471 354.2 574 463.2 643.9 470.8 493.1 419.9 465.8 457 528.6 621.8 559.2 633.2 544.6 488.6 494.7 473 464.8 385.5 466.5 511.3 475.9 FB FBgn0038111147273_at 552.6 489.3 584.6 599.6 697.4 629.8 819 424.6 356.5 751.6 733.1 674.6 717.1 702.4 806.7 744.3 507.3 446.5 438.7 625.5 542.9 578.8 414.7 411.9 FB FBgn0034065149585_at 10.6 8.5 18.1 4 12.1 21.1 9.9 8 6.2 19.8 11.8 7.3 8 11.1 5.4 13.5 5.6 14.5 7.2 5.2 5.7 17.5 10.9 45.4 FB FBgn0037751149616_at 5.7 18.3 4.6 4 21.6 16.3 12.5 4.3 28.3 10.7 15.3 11.4 34.2 43.5 45.3 70 13.2 22.4 9.5 17.3 23.4 13.8 11.2 16.4 FB FBgn0037789142033_at 88.2 8.8 119.6 37.2 95.9 96.1 98.1 72.5 21.4 114.4 60.2 82.7 77.8 68.6 59.7 157.1 47.6 57.1 36.1 130.6 51.5 10.4 144.7 69.8 BDGP;LD32106.3141630_at 314.7 323.7 215.9 242.1 290.1 317.2 331.9 299.5 478.1 231.1 249 306.9 260.2 253.3 334.4 504.5 306.6 383.1 310.1 161.5 195.8 415.2 458.9 379.9 FB FBgn0034728149455_at 17.1 10.4 13.3 42.9 24.5 33.4 7 5.7 17.8 6.3 19.1 3.3 7.3 62.1 55.7 4.1 2.9 32.1 17.7 21.4 29.1 8.8 19.8 7 FB FBgn0037514145085_at 4.6 62.1 32.4 9.7 50.4 11.3 13.8 4.1 11.7 47.5 10.7 5.8 17.5 8.2 8.1 38.3 4.6 6.2 18.8 36.1 4.6 25.3 35.8 37.2 FB FBgn0030590149320_at 112.5 71.5 32.5 31.1 125.3 35.1 42.7 30.6 23.2 107.9 33.5 38.4 107.9 24.5 47.1 17.9 28.6 75 31.9 26.8 53.8 38.8 77.3 39.7 FB FBgn0037310148660_at 26 29.1 25.7 20.6 11.2 6.8 21.3 47.3 32.3 21 19.8 25.4 15.8 39 42.7 13.6 35.5 19.9 72.8 62.4 46.5 71 10.3 57.6 FB FBgn0036287152340_at 115.4 183.8 203.2 135 163 91.8 140,5 252.8 209.8 238.8 166.2 162.8 136 133.4 175.3 127.1 241.8 289.3 144 188 166.1 234.6 215.4 161.6 FB FBgn0004837153280_at 786.9 1229 1305 1050 1095.5 1040.3 910.4 1304.3 1222.7 1086 660.2 725.7 1217.2 1053.4 912.2 1186.1 1134.1 1053.3 843 1143.5 923.3 814.3 645.5 772.9 FB FBgn0030468148305_at 10.1 8.7 7.3 5.7 6 9.9 19.5 2.6 10 7 28.4 10.9 13.5 80.3 11.8 7.7 11.1 18.8 11 11.5 15.7 14.3 8 3.7 FB FBgn0035739
142713_at 796.9 846.4 1141 793.9 711.2 772.4 622.7 897 952.6 724 497.6 549.2 582.7 636.7 642.8 478.6 600.2 512.4 581.3 666.2 591 526.5 712.9 457.5 FB FBgn0015664153119_at 111.3 112.4 119.4 130.2 72.5 53 59.1 136.8 163.6 172.2 122.9 122.4 80.4 158.9 120.4 71.7 102.5 130.9 59.5 65.7 95.2 108.4 86.1 99.8 FB FBgn0038167
145615_at 21.3 64 93.9 7.2 110.7 24.6 127.2 24.9 45.4 49.3 50 90.6 20.8 46.9 51.6 101.5 22.7 32.2 35 54.8 9.4 39.2 87.5 10.4 FB FBgn0031384

Probe SetID
ywnonhsi
yw
2
ywnon hs
3
ywshort hs1 yw short
h s2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
pKB
1
pKB non hs
2
pKB
3
p K B
short hs 1
p K B
short hs 2
p K B
short hs 3
p K B
long hs 1
p K B
long hs 2
pKBlong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
153575_at 641.9 478 824.1 395.3 430.2 448.4 398.8 428.4 398 686.3 572.8 568.8 547.6 450.7 461.4 452.8 460.7 450.9 767.5 666.9 791.3 512.1 571.8 675.8 FB:FBgn0035148142971_at 445 588.8 883.9 681.7 553.3 486 281 534 486.4 863.8 467.7 628.7 733.1 640.4 610.4 235 540.8 355.9 585.8 577.8 502.7 474.2 390.8 283.8 FB:FBgn0003638142976_at 41.4 162 116.5 130.3 84.4 25.6 61.4 114.5 136 213.7 76.3 62.3 144.1 30.1 145.2 34.8 42.4 118 16.8 42.1 97.7 165.2 159 89.8 FB:FBgn0032260143249_at 8725 17396 4365 15298 15745.1 16558.6 18312 10395 10588 4445 11374 13645 10820.5 22690.5 13778.4 29956 12047 12140 10043.1 17650.4 10720.8 26808.9 23497.4 11245.8 FB:FBgn0002622141414_at 125.2 184.2 199.8 109.2 96.3 84.8 133.3 175.7 123.9 210.3 188.8 152.8 121 129.9 75.7 179.4 137.1 97.8 153.5 176.5 143.4 120.9 107.5 140.1 FB:FBgn0031428146923_at 18.3 129.4 114.4 78.1 126.4 107.9 10.9 100.1 84.8 88.5 54.8 91.5 102.2 70.7 166.8 16.7 97.1 24 168.2 179.4 104.9 162.6 288.3 134.8 FB:FBgn0033488152853_at 23.7 67.7 45.4 3.7 30.1 26.3 30.1 30.4 24.2 51.3 3.9 64.2 17.8 9.8 26.1 19.3 2 44.4 6.4 3.9 34.8 35.6 27.9 2.4 FB:FBgn0038664144143_at 1657 1850 1153 1528.6 1918.2 1942.5 1136 1509.9 1640.2 735.4 1632 1531 1604 1645 1420.9 1483.2 1659.1 1597.5 1414.6 1147.2 1541.1 1328.8 1460.5 2412.8 FB:FBgn0026415152598_at 723.5 444 361.8 536.1 514.1 459.6 1175.5 1182.1 1303.1 276.6 632.9 421.4 125.1 167.9 97.4 128.6 223.6 248.8 736.5 580.3 490.9 346.6 269 1112.3 FB:FBgn0034335150120_at 322.6 291.4 332.2 315.5 347.1 394.6 535 388.4 259 432.8 344.8 313.7 358.4 319.1 450.9 515.7 347.2 529 364.2 448.4 392.7 421.6 421.9 439.6 FB:FBgn0038582143888_at 113.9 127.5 147.9 131.1 153.6 188 131.1 73 99.9 215.3 125 145.9 115.3 211.7 90.3 166.8 57.5 134.4 111.4 200.6 115.6 194.8 82.7 76.4 FB:FBgn0017424148585_s_at 65.1 43.2 91.3 60.1 99 79.2 158.3 32.4 135.4 58.2 117.4 123.3 109.5 76.8 171.6 67.1 126 43.7 105.4 113.9 30.3 36.3 107.7 123.8 FB:FBgn0036176151939_at 647.3 549 858.5 636.3 622.2 578.8 1217.9 606.8 625.7 726.1 544.4 484.2 577.2 510.9 601.5 923.6 564.6 663 539.8 850.5 559.6 590.4 884.9 375.3 FB:FBgn0033459143694_at 11.5 16.7 26.1 37 19.4 43.9 45 32.8 34.3 10.1 15.4 37.8 31 53.3 24 27.2 16.8 15.3 10.5 11.6 12.1 35.1 49.1 7.1 FB:FBgn0011820145163_at 248.8 336.5 330.4 186.8 231.4 201.7 307.7 344.5 311.1 502.4 405.5 375.9 346.4 255.1 304.8 281 427.4 593.9 245.5 286.6 262 334.9 267.4 162.8 FB:FBgn0030711147097_at 105.3 46.9 24.3 59.8 59.4 99.4 60.9 76.1 13.9 59 32.8 25 111.2 99 106.5 14.5 94.2 52.9 56 100.3 84.4 145.3 81.8 55 FB:FBgn0033746153476_at 6 39.9 3.9 8.1 43.7 7.4 7.5 39.4 47.7 11.5 8.8 12.8 5.1 10.5 29.4 98.6 8.9 12.9 11.2 10.4 4.2 16.7 74.8 9.6 FB:FBgn0036907154320_at 263.1 226 370.4 350.6 409.5 391.1 310.8 208.2 274.6 379.6 268.6 317.3 385.5 322.8 476.8 368.3 216.1 203.8 317 205.2 273.3 377.9 331.6 403.5 FB;FBgn0001291143534_at 32.2 38.6 7.5 10.7 7.6 5.7 10.9 9.6 16.9 12.9 9.3 12.5 5.4 15.8 3.5 3.7 10.7 9.4 26.2 38.1 42.7 10.5 19.6 56.9 FB:FBgn0004910144072_at 305 234.3 386.3 313.8 366.3 423.4 302.6 244 348.6 374.5 331.5 396.4 280.2 365.5 251.9 194.4 355.1 393.8 275.6 347 288.7 381.8 367.8 415.6 FB:FBgn0025621146279_at 7.2 7.5 11.5 8 31.8 10.5 19.6 2.1 13 10.6 2.9 33.5 48.4 1.7 59 2.3 17.2 5 3.9 21.8 13.8 3.5 5.5 1.2 FB:FBgn0032419147672_at 33.4 24.8 21.5 14.6 67.7 48.3 30.1 12.8 6 21.2 76 7.4 19.7 6.4 7.7 6 76.2 27.8 34.1 28.5 4.5 22.6 11.8 16.1 FB:FBgn0034721153689_at 274.1 274.6 254.4 272.6 352.5 355 268 577.1 499.5 219.2 228 310.1 391 356.4 377.4 503 487 488.4 148.5 221.3 195.5 282.4 329.6 265.5 FB:FBgn0037106149285_at 3.5 8.8 12.1 10.8 8 6.6 6.4 3.3 4.7 10.7 11.1 14.8 9.5 13.9 16.9 14.2 3.8 26.3 5.1 26.3 6.1 7.6 44.7 6.9 FB:FBgn0037260151364_at 134.1 68.5 43 110.7 116.9 82.8 102.7 22.7 77.5 65.6 88.4 76.8 43.9 89.8 93.6 51.6 40.9 51.9 48.4 113.1 45.6 108.6 111.9 30.6 FB;FBgn0040891143909_at 26.7 63.6 56.4 5.2 4.8 58.9 8.4 8.4 20.7 33.4 44.8 2.7 12 19.7 4.3 7.7 42.8 15.7 4.3 24.2 13.5 23.5 4.4 4.2 FB;FBgn0019985145685_at 349.8 291.4 297.4 265.5 355.3 263.8 328.2 636 500.3 252.4 260.6 227.9 269.8 228.7 306.2 375.4 531.1 470.1 229 446.8 288.7 300.5 423.4 251 FB;FBgn0031491148090_at 60.9 126.4 24 113.5 149.8 159.1 22.1 96.3 112.2 114.2 90.1 222.5 220.1 210.1 343 11.1 261.7 40.2 216.3 223.4 135.2 154.9 215.9 115 FB:FBgn0035416149531_at 6.3 14.6 5.9 100.3 107.2 125.1 11.6 9.9 61.4 67.5 13.2 88.3 54.4 65.1 110.9 13.8 21.2 20.2 29.4 66.7 38.5 18.6 101.3 27.3 FB;FBgn0037641154450_at 127.2 170.5 125.4 152.3 108.8 114.4 124.6 181.9 141.6 115.8 132.8 165.5 176.1 123.1 163.9 138.2 208.2 165.5 237.8 184 193.5 98.8 120.4 116.8 FB:FBgn0015331148747_at 77 173.6 47.6 13.3 31.9 18.6 102.5 84.4 29.9 62.8 177 200.3 67.4 27.5 60.7 123.3 49.9 19.7 54.4 95.4 81.8 121.4 46.7 127.3 FB:FBgn0036419150105_at 188.7 357.1 309.1 377.3 324.2 367.8 230.5 170.3 241.1 514.7 501.2 402.8 479.5 483.1 427.2 264.8 328.4 271.9 337 534.9 307.6 714.5 682.7 148.2 FB:FBgn0038555146944_at 33.4 20.5 9.6 30.2 16.3 20.8 4.8 24.3 18 6.7 3.9 42 58.5 34.4 11.6 10.8 33.6 26.7 6 3.3 6.9 10.4 9.6 30.5 FB;FBgn0033516142784_at 2257 1897 2088 2305.7 2266.3 2367.9 1908.7 2221.2 1988.8 2937 2328 2692 2989.2 2602.6 2799.6 1823.9 2108.4 1957.1 2472 1905.2 2194.1 2369.3 2283.6 1951.8 FB:FBgn0037621151395_at 161.5 170.7 230.3 226.9 224.8 264 271.3 224 116.9 155.4 211.1 169.1 104.3 325.3 114 273.4 253.3 150.6 162.4 149.5 137 151.8 292.4 224.8 FB:FBgn0040926152823_at 50.4 14.5 74 59.9 63.5 24.5 17.1 19.8 37.7 164.5 60.5 66 54.1 80.6 45.2 75.8 63.7 28.7 34 53.6 7.5 9.7 63.4 10.6 FB;FBgn0036403141427_at 301.5 171.7 274.3 188.6 197 162.4 182 286.5 275.5 328.7 323 313.8 256.8 287.6 177.5 294.7 274 346.2 177 165.3 230.2 296.7 320.3 189.1 FB:FBgn0031098155110_at 1580 1420 1630 2056.9 2030.4 2204.5 2520.5 1671.5 1946.8 1699 1905 1738 2325.7 2246.1 2190.4 2796.6 1760.1 1802.1 1397.4 1835.5 1533.9 1001.6 1561.7 1578.7 FB;FBgn0003449147244_at 12 61.7 16.3 18.8 41.1 52 145.4 65.6 14.7 69.9 58.8 72.4 63.8 65.3 68 114.2 22.1 68.2 61 78.2 30.1 133.6 73.5 71 FB;FBgn0034012149712_at 23.7 49.1 18.7 12 23 13.1 31.5 57.9 21.5 49.7 122.4 31.6 25 11.2 28.2 15.9 18 24.9 23.8 44.9 40.8 36.1 24.4 16.6 FB:FBgn0037956152259_at 171.1 176.6 224.8 135 186.4 208.7 168 311.6 275.2 279.5 189.3 149.7 139.5 178 128 245 260.7 344.5 149.2 144.8 169.9 254.1 243.4 229.1 FB:FBgn0030671149607_at 202 186 126.4 194 215.3 175.9 160.8 151.7 144.3 102.4 201.8 190.9 103.9 116.7 118.6 157.8 174.3 164.2 174.2 140.4 233.9 100.8 195.7 224.8 FB:FBgn0037779141644_at 1643 1933 1523 1225.1 1085.5 1109.6 1047.2 1014.4 2128.7 1554 1648 1877 1405.9 1360.9 1348.5 1203.5 1626.6 1458.4 1731 1545 1630.8 2024.1 2091.6 1868.9 FB:FBgn0001309147583_at 52.2 19.8 17.2 77.2 18 31.3 23.1 16.7 17.9 57.5 18.2 20.7 82.6 128.9 60.4 46.5 40.3 20.7 52.6 39.6 57 17.1 30 17.7 FB:FBgn0034559153330_at 2001 2416 2963 2132.7 1803 1797.3 2756.3 2725.3 2304.4 2682 2571 2374 2770.8 2498.9 2648.3 3017.5 2429.9 2387.6 2101.3 2126.4 2220.5 2160.4 2344.7 2275.2 FB:FBgn0038964154148_at 2046 1890 2364 1917.9 2114.4 1815.6 1767.1 2274.2 2338 2151 1780 2214 2009.5 1739.9 1852.4 1819.6 2267.5 2517.8 1899.8 1831.4 2070.1 1494.7 1854.4 1946.4 FB:FBgn0036974148738_at 380.3 560.1 648.4 934.4 1003.6 937 281.6 587.8 759.1 585.3 452.6 412.2 933.8 632.7 744.1 197.7 614.3 517.5 395.6 547.7 296.2 975.7 1034.5 239.1 FB:FBgn0036399153673_at 784.3 884.9 568.9 1085.2 841.6 694.5 1165 958.7 719.4 484.3 874.5 970.1 829.7 733.3 726.2 1284.7 1171.5 1089.4 891.2 981.5 1123.5 744 817.1 1636.9 FB;FBgn0000083149847_at 66.8 42.2 54.4 47 48.1 81.1 19 54.8 30.4 90 112.5 32.1 70.1 110 126 122.1 20.7 42.4 88.4 16.9 17.1 61.2 32.1 25.9 FB:FBgn0038158142509_at 306.2 275.5 349.5 303.2 365.1 291 337.9 314.3 351.3 392 298.5 343.9 429.5 263.9 474.5 489.3 447.5 325.6 356.8 242 345.6 568.5 434.8 336.5 FB;FBgn0033846152028_at 179.5 189.3 293.2 177.7 158.4 194.9 154.2 237.2 204.4 296.2 166.6 263.2 173.3 188.8 96.9 242.3 278.6 197.2 207.5 233.6 233.9 123.8 270.7 225.7 FB:FBgn0033010146277_at 47.9 110.5 66.2 8.7 44.8 22.6 12.2 42.3 54 57.5 10.6 8.2 35.6 60.3 28.3 31.7 39.4 64.5 41.3 70.1 13.7 21.2 58.7 13.5 FB:FBgn0032417147043_at 27.7 36.2 31.9 47.3 26.2 68.1 53.6 7.4 42.3 57.9 64.6 5.8 42.4 6.2 33.7 47.8 36.6 44.8 35.4 46.2 11.6 19.7 50.1 58.7 FB;FBgn0033659150352_at 70.5 78.7 104.2 77.1 51.5 71.9 52.1 17.4 63.3 121.9 46.6 128.8 63.7 77.5 93.8 33.7 66.5 79.1 32.8 82.4 66.9 14.4 31.9 93.8 FB:FBgn0038943
151590_s_at 85.8 76.3 34.5 160.3 82.4 119.7 36.5 28.3 50.4 39.6 34.6 79.5 145.8 110.1 161.7 33.3 66.8 59.3 46 37.3 52.2 150 130.3 63.1 BDGP;GM09104.:

Probe SetID
ywnonhs1
ywnon hs
2
ywnon hs
3
ywshort hs
1yw short
hs 2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
pKb
1
pKb
2
pKb
3
pKbshort hs
1
pKbshort hs
2
pKbshort hs
3
pKblong hs
1
pKblong hs
2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
142929_at 29.5 46.2 4.1 7.7 21.4 33.7 8 1.5 8.4 4.3 1.5 3.6 35 12.9 5 11.7 12.7 8.2 17.3 4.5 23.2 21.2 38.8 12.3 FB:FBgn0038798150432_at 126.5 143.7 101.7 130.5 151.8 129.2 63.6 113.6 113.7 137.5 152.6 136.6 148.6 160.9 161.9 66.8 110.6 127.7 106.6 2223 124.6 224 138.4 76 FB:FBgn0039062151699_at 276.7 139.4 94.8 300.4 297.3 464.3 217.7 157.8 210.7 30.6 142.9 239.3 202.2 312.4 250.5 142.2 115.6 220.1 171.2 213.6 197.8 280.1 235 215.5 BDGP:LD34994.3prime-hit141704_at 1093 1387 879.7 852.5 722.8 924.1 702.3 1256.9 1417.9 621.7 671.9 783.8 647.7 588 612.8 595.2 1164.6 978.2 1024.7 968 1028.5 806.5 760.1 1030.2 FB:FBgn0032456154370_at 417.8 487.8 380.4 596.4 627.7 647.3 496.9 586.9 597.7 164.9 258.3 235.9 307 250.7 245.8 235.4 319.4 309.2 180.2 613,8 316.2 383 348.9 323.9 FB:FBgn0040946143840_at 73.1 19 29 13.8 17 21.8 78.3 19.7 64.7 17.7 6.7 13.7 26.5 28.6 17.9 18.5 56.6 59.6 8.4 9.7 6.6 17.3 11.1 10.3 FB:FBgn0015770150177_at 24.2 79 73.6 16.9 13.6 34.6 21.2 16.8 29 22.2 133.7 80.9 8.2 38.6 5.7 11.5 72.8 16.2 49.5 15.1 147.4 128 118.2 303.4 FB:FBgn0038676152672_at 10.3 29.6 24.4 3.7 38.7 41.6 44.1 23.1 32.8 12 16.3 21 13.6 37.8 33.9 10.4 18.2 27.3 23.6 10.1 8.8 65.4 4.4 38.8 FB:FBgn0035214142951_at 98.4 109.2 117.1 85.2 49 87.9 77.9 94.2 53.3 106.8 128.9 85.8 50.7 80.9 60.4 90.1 54.1 69.4 64 101.3 76.1 94.3 8.5 22.8 FB:FBgn0028534150964_at 443.5 472.6 466.1 444.8 522.4 494.4 277.1 763.8 793.4 391.4 457.6 394.4 480.3 423.3 385.6 214.4 599.2 628.2 358.9 566.8 416 376.4 483.5 258.3 FB:FBgn0039888153794_at 215.5 317.9 325.4 243.4 235.8 234.7 310.8 288.4 321.3 283.8 243.1 189.6 186.3 184.2 135.9 254.1 310.9 238 249.3 335.8 266.3 193.3 242.1 331.7 FB:FBgn0039179154638_at 290.6 359.1 402.2 260.2 280.7 265.5 193 403.3 374.1 312.5 303.6 234.9 226.9 298.9 252 198.4 355.8 439.2 319.6 285.3 341.6 257.3 284.5 260.7 FB:FBgn0031062150318_at 140.5 72.2 128.5 109.7 94.1 147.3 120.8 104.6 126.7 41.4 98 108.9 103.8 102.3 117.9 39.2 80.6 185.3 129.1 125.6 127.5 29 8 15.6 126.4 FB:FBgn0038893152042_at 19.3 5.4 7.8 10.8 12.1 5.1 12.2 47.5 57.3 9.7 8.2 12.6 6.4 24.6 5.4 8 7 44 20.6 42.8 46.5 49.4 15.7 7.6 73.9 FB:FBgn0032032148508_at 45.3 104.3 90.6 12.3 29.8 12.1 7.9 15.8 64.8 28.1 14.5 12.6 16.1 14,1 11.7 33.1 18.1 16.3 6.1 33 31.5 35.6 33.3 12.8 FB:FBgn0036069147112_at 792.9 636.6 748.8 700.7 833 640 794.3 622.6 934.3 683.2 668.6 698.7 659.2 919,7 738.7 595.8 623.8 716 860.9 7734 677.9 841.5 724.2 595.4 FB:FBgn0033766148620_at 71.6 28.7 41.3 18.8 20.1 35.3 131.1 21.8 77.6 72.4 112.9 35.7 12.3 97.5 13 84.9 23.9 18.5 134.1 117 4 177.5 31.4 18 50.2 FB:FBgn0036227143095_at 18.5 30 51.4 10.5 14.9 30.5 17.5 22.6 9.9 9.5 26.9 76.8 30.3 9.8 8.9 146.2 12.7 34.7 37.4 10 25.9 25.3 16.4 23.4 FB:FBgn0000274141358_at 15208 20231 9239 22586 21578.8 24358.5 23654 15316 16210 9775 16537 18358 17961.2 30163.9 22828 40144 16673 25803 16046.3 21735.8 15085.2 27772.1 28558.8 17256.3 FB;FBgn0031035146480_at 424.8 365.3 604.1 462.2 417.8 432.6 716.9 481.3 478.4 534 482.3 398.5 420.7 329.2 481.8 574 344.2 451.9 388.1 4173 453.9 450.5 408 419.2 FB:FBgn0032761150292_at 30.5 35.5 34.4 8.7 30.8 9.7 31 20.7 20.3 12.8 89.3 4.8 13.1 36.7 60.7 58.9 5.4 6.9 23.5 27 21.5 35.8 46.1 28.9 FB:FBgn0038849141527_at 146.6 314.9 117.4 144.8 75.8 127.2 92 104.8 92.2 35.4 122.6 140.4 42.4 106.9 117.4 66.1 103.7 115 149.5 121 6 187.1 136.4 125.7 364.5 FB:FBgn0023479141846_at 47.8 5.3 2.9 43.3 46.2 6.3 57.4 8.5 22.2 5 26.6 24.8 7.2 42.4 27.5 33.3 23.4 7.9 2.8 4.8 2.4 35.8 30.6 24.1 BDGP:GH07296.3prime-hit154517_at 427.9 675.4 1043 579.4 446.4 440 668.1 987.1 694.2 1089 627.3 607.2 509.9 495.3 517.3 578.8 1050.7 536.6 421.1 7557 555.9 618.5 668 709.4 FB:FBgn0029687152400_at 23.2 19.6 25.7 13.3 16.6 77.2 21.6 24.5 25.6 20.5 12.6 118 44.8 27.5 9.5 47.8 11.8 31.9 13.8 10.6 10.6 28.4 23.6 15.7 FB:FBgn0033794144697_at 19.7 24.6 32.7 20.5 10.7 14.3 28.2 10.8 11.3 27.9 11.2 15.5 9.9 13.5 12.6 15 25.2 20.8 13.4 20.8 12.2 34.9 20.8 14.5 FB:FBgn0030032153311_at 343.1 454.1 519.9 585.6 440.5 585.4 578.4 741.6 970.1 436.9 331 324.9 477.6 483.6 467.4 466.2 779.2 810.1 544.9 447.4 456.2 641.6 594.2 1093.5 FB:FBgn0001220148216_at 1054 1146 1590 1205 1113.4 955.2 960 2051.8 2146.2 1367 1035 971.4 1049.2 841.8 916.6 1054.3 2229.4 2283.2 1003.3 1062.5 890.9 693.2 924.5 635.7 FB;FBgn0035601149862_at 10.2 11.1 3.1 29 7.3 5.2 22.2 38.3 4 4 10.3 12.5 2.8 13.7 22.7 16.1 34.9 12.2 20.2 5.9 4.4 9.7 7.5 19.3 FB:FBgn0038186142401_at 20.5 34.5 8.8 44.6 12 6.9 14.1 21 5.5 15.5 12.7 71.1 50.4 4.5 19.8 95.6 89 15.1 87.9 57.6 72.9 22.7 6.1 101.1 FB:FBgn0028428152742_at 14.8 57.3 17.7 24.4 16.9 10.2 71.2 10.7 24.6 30.2 28.7 25.7 8 13.8 9 13.8 39.7 7 4.2 15.6 13.8 19.1 10.9 59.4 FB;FBgn0037381153368_at 495.2 501.5 256.7 376.9 301.9 418.7 251.1 205.6 182.8 352.5 481.2 497.2 254.9 382.8 320.5 338.9 284.9 214.2 539.7 404 1 535.5 353.5 261 842.9 FB:FBgn0026721145463_at 199.7 40 76.7 167.8 83.8 144.7 82.6 106.7 57.9 187.4 36.3 136.9 133 248.2 186.8 180.7 164.6 79.4 159.6 140.2 133.3 93.5 254 208.2 FB:FBgn0031162147878_at 19.7 71.4 27.5 22.9 18.3 12.2 45.9 25.1 22.3 27.9 28 8 23.8 21.3 29.4 66.9 41.1 23.9 31.8 12.4 18 8.1 16.6 25.3 20 FB:FBgn0035042148182_at 5967 3676 1537 1159.9 1274.2 1801.3 2452 1061.5 1185.5 958.5 4309 5224 493.3 1159.3 894 536 509.5 660.2 7564.9 7441.8 9241.5 1670.4 1778.4 2507.8 FB:FBgn0035551144177_at 98.6 162.8 124.6 72.5 47.7 131.4 227.4 103.4 126.8 212,3 154.6 146.6 85.4 27.5 37.4 63.9 129.9 158 187.5 220,3 179.8 206.2 180.1 258.3 FB:FBgn0027343146150_at 13 34.8 34.6 18.2 24.4 35.5 13.4 52.8 19.7 36 25 27.6 32.3 34.7 26 29.4 11.3 16.9 32.5 19.6 12.9 34.7 29.5 15.3 FB:FBgn0032233141707_at 544.1 623.7 653.7 549.1 496.1 570.7 421.8 434.6 419.7 543.3 523 582.9 531.5 523 500 315.3 461.6 633.5 522.2 585.4 498.6 594.9 652.7 476.6 FB:FBgn0011570144490_at 13 10.5 45.6 7.2 8.6 6.6 9.8 30.1 9.9 38.2 19.8 52 24.5 24.1 20.1 32.3 31.8 29.4 2.2 19.9 10.4 37.7 2.8 46.6 FB:FBgn0029724144839_at 275.5 425 241.8 298.8 287 256.3 282.6 226 221.8 266.5 341.9 316.6 322 308.8 297.6 224 307.2 363.3 499.6 385.6 527.9 481.3 359.3 860.6 FB:FBgn0030255146183_at 520.4 488.2 422.3 137.3 165.8 85 740 3 241.3 203.9 222.7 99.1 397.6 63.7 135.9 141.6 36.5 89.7 70.3 588.5 793.2 1195.5 194.5 178.3 414.4 FB;FBgn0032284148143_at 45.2 180.4 220 71.8 98.5 159.1 199.3 62.4 136 267.9 102.9 69.8 47.9 127.3 44.1 188.1 48.9 174.9 129.5 45.2 31 51.7 123 165.7 FB:FBgn0035491150628_at 33.2 34.4 22 8 57.3 49.7 15.9 13.2 45.1 30.5 21.1 16.7 32 26.6 37.4 8.4 12.2 11.9 79.6 53.2 46.7 12.9 41.8 70 FB:FBgn0039390143390_at 7328 7674 7352 6752.7 6614.3 6295.8 4172.8 5693.1 6877.2 7060 6746 7026 6457.7 5201.5 6452.6 4672.1 6131.1 6221 7325.4 6387.3 6675.1 3706 6408.9 5925.1 FB:FBgn0003887
154825_at 251.9 212.8 181.5 169.3 213.2 183.8 265.9 258.2 320.7 219.1 185.9 194.2 167.7 132.5 140.4 192.9 354.6 293.9 122.6 1684 97.7 141.9 112.4 229.7 FB:FBgn0034689152796_at 895.5 1147 864.1 893.5 698.2 902.6 863.5 1531 682.3 906.4 985.3 1278 1268.1 1037 1201.5 1009.6 832.5 1092.4 921.7 803.3 802.4 752.3 967.1 870.7 FB:FBgn0034975150849_at 115.6 119.7 85.3 98 95.9 91.3 27.9 148.2 129.1 109.2 39.7 89.8 85.1 43.6 78.5 24.9 106.4 103.7 119.8 100 1 80.2 111.3 118.4 81.3 FB:FBgn0039706147062_at 10.3 13.7 20 18 14.1 24.2 11.1 16.1 15 14 12.6 15,8 16.9 18.2 26.5 13.7 12.6 18.6 14 12.9 5.2 18.5 41 14.2 FB:FBgn0033693154283_at 872.7 622.7 800.5 1009.3 1019 990.1 869.9 1086.9 858.5 1083 1052 1171 1599.3 1236.3 1581 1514.6 1347.2 1121.4 667.6 762 2 676.8 653.7 749.3 839.1 FB:FBgn0030365
148884_at 192.8 211.8 405 123.7 43 9 46 7 189.4 43.5 34.1 67.2 48 3 181 111.9 265 193.-1 79.7 266.6 130.2 184.9 107 9 4 4 69.7 52 5 143.9 FB.FBgn0036610
152638_at 821.1 773.3 571.8 642.1 637.3 576.5 697.1 757.5 1082.6 622.4 817.4 843.5 759 628.7 612.6 901.1 966.1 799 734.1 961.7 850.5 1111 1223.7 1339.4 FB:FBgn0034987
145388_at 164.2 137.1 201.5 201.5 132.1 208.7 148.2 201.6 200 8 273.7 117 4 130 3 184 1 221 4 244 9 82 4 269 175 4 190 7 2138 190 1 123 4 199 ? 128 9 FR FBan0031054
141697_at 132.4 116.5 133.8 85.9 100.8 74.3 87.6 107.3 84.7 134.6 50.2 92 93.4 93.5 126.1 97.2 117.7 76 138.4 126.2 191 126.5 57.9 245.6 FB:FBgn0010225
154600_at 76.2 50.7 70.2 31.6 33.5 31.1 144.4 93.1 133.4 35.2 27 30 32.3 34.7 29.6 292.6 133.2 36.2 35.1 1135 103.2 33.5 130 47.7 FB:FBgn0038548
147245_at 8.9 208.8 14.6 61.7 67.7 34.3 13 22.4 27.5 16.7 18.6 20 85.3 32.7 19.1 22.6 20.8 14.4 107.5 109 16.1 31 130.1 22.4 FB:FBgn0034013

Probe SetID
ywnonhs1
ywnon hs
2
ywnon hs
3
ywshort hs
1yw short
h s2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
p b non hs
1
pKbnon hs
2
pKbnon hs
3
pKbshort hs
1
pKbshort hs
2
pKbshort hs
3
pKblong hs
1
pKblong hs
2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
153763_at 265.9 293.7 312.3 262.2 185.7 203.6 293.4 306.6 423.1 223 268.5 222.3 190.5 233.2 209.7 275.4 294 228.6 359.5 330.1 377.6 254.6 274.9 520.7 FB:FBgn0035807151120_at 46.5 74.9 103.5 199 135.5 182 36.4 169.6 107.7 140.5 52.1 51.9 146.2 127.5 107.5 11.4 155.8 125.2 57.1 139.2 70.4 125.3 85 71.6 FB:FBgn0040635145068_at 26.8 137.2 23.7 65.1 76.2 22.7 10.6 9.4 23.6 98.5 83.6 141.6 94 137.4 16.5 17.4 51.4 16.7 40.8 104.8 123.4 70 157.8 79.7 FB:FBgn0030568148478_at 5.1 7.4 25.5 3.9 33.2 6.2 4.9 2 2 25.7 37.8 4.7 65.1 7.4 3 26.5 11.1 17.7 8.5 2.2 4.3 2.7 46.9 4.8 1.7 FB:FBgn0036019150845_at 62.2 87.4 5.7 39.2 80.5 63 69.7 21.1 7.2 67.6 78.3 86.6 72 115.6 155.2 32.8 14 54.2 25.5 69.3 29.9 126.2 55.6 58.4 FB:FBgn0039699141793_at 149.8 179.6 155.7 145.3 110 132.4 197.4 116.7 91.5 184.8 176.6 132.1 64.3 72.5 92.8 192.8 102.8 106.9 143.2 136.7 134.9 120.2 152.1 251.4 FB:FBgn0034698154956_at 649.9 465.4 618.2 804.7 807.3 536.6 320 392.3 481.3 818.9 808.4 664,6 827.1 847.2 513.4 706.4 581.9 577.5 394.1 418.6 266.8 237.6 428.4 214 FB:FBgn0039073151114_at 71.9 40.2 76.8 35.4 10.8 65 57.7 49 33.1 25 83.4 15.5 73 103.7 65.2 22.1 52.9 63 8.6 43.6 60 21.2 24.1 50 FB:FBgn0040628154974_at 1033 1186 1495 1682.5 1982.7 1791 1227.5 1771.8 1636.9 1291 1394 1216 1721.1 1677.7 1783.8 1067.1 1447.7 1399.6 1551.6 1442.6 996.9 1655.1 1414.6 1122.8 FB:FBgn0036389
152045_at 164.1 129.6 39.9 31.9 77.5 39.3 11.8 26.1 9.7 15.7 12.6 203 13.5 15.7 70.7 32.1 80.1 24.3 210.9 179.9 130.1 27 15.7 157.3 FB:FBgn0015032
145768_at 9.5 22.9 61.9 39.2 8.1 7.2 19.3 6.2 10 11 3.8 7.1 34.9 15 8.2 10.7 14.5 23.1 8.1 36.2 4.2 13 51.1 4.8 FB:FBgn0031616
143984_at 7.6 10.9 13.9 11.2 19.2 20.1 21.8 2.3 4.6 12.5 7.8 3.2 5.6 18 12 71.5 7.3 3.7 9.8 5.7 8 11.5 8.9 7.1 FB:FBgn0023415
142732_at 71.7 75 22.9 21.4 16.7 31.5 127.7 30.5 15,7 43 181.4 145.1 37.6 34.6 32.4 23.2 25.8 20.3 144.8 125.4 122.6 32.4 46 51.5 FB:FBgn0036947146316_at 99.1 105.5 189.2 211.8 235.2 271.9 98 246.6 250.2 149.7 142.2 106.5 243.3 199.6 255.6 121.5 268.7 230 176.1 193 206.8 226.8 196.2 210.5 FB;FBgn0032485147365_at 191.4 202.2 136.8 506.8 542.6 379.1 214 216.8 375.7 211.7 202.9 228.4 378.3 303.6 412.4 286.5 349.9 339.3 206.8 247.5 120.8 206.3 342.8 85.5 FB:FBgn0034223151857_at 278.1 220.9 424 240.2 346.4 215.6 514.6 321.4 248.1 399.4 282.8 2845 165 259.6 264.6 507.5 295.7 297.6 300.1 395.4 358.2 325.2 239.2 349.6 FB:FBgn0036286142662_at 1964 110.4 111 55.7 44.5 53.2 122.1 101.2 37.3 86.7 97.4 162.3 3.4 33.5 16.1 85.2 59.6 66.3 176.6 118.7 230.7 123.6 28.9 185 FB:FBgn0038613142547_at 100.6 104.3 118.9 119.9 105.2 118.6 15.2 82.4 34.8 54.8 77.7 43.1 13.2 19.4 74.2 18 25.3 75.8 56.4 66.4 88 138.2 117.2 38.4 FB:FBgn0034175154839_at 681.9 570.8 601.2 800.6 852.8 869.2 804.7 1082.9 844.1 626.8 628.6 557.7 1100.8 974.3 853.8 700.3 908.2 491.6 742.6 542 672.9 554.6 899 580 FB:FBgn0037255154724_at 5.4 7.6 12.1 69.8 71.8 78.1 6 23.4 66.4 25.7 16.5 11.4 51.4 98 8 35.9 9.9 86.6 67.9 28 97.9 18.4 75.1 76.3 8.2 FB:FBgn0037445144779_at 142.2 344.6 75.6 109 168.9 159 26.7 386 86.9 23.3 132.8 61.7 73.5 80.5 29.7 50.5 38.7 75.4 166.6 162.4 158.6 107.7 125.1 359.2 FB:FBgn0030171
153977_af 118.5 83.7 46.9 78.1 82.9 108.7 20.7 39.5 35.1 58.3 135.7 170.3 8 8 116.9 59.2 17.4 67.2 70.4 123.2 57.9 86.7 75.4 36.3 204.1 FB:FBgn0022770142837_at 43 83.1 46.7 187.6 102.5 139.8 168.7 58.8 88.4 104.8 85.7 81.3 195.5 174.2 249.9 87.3 121.5 136.1 119.4 71.4 79.3 185.7 112.2 173.5 FB;FBgn0001228148636_at 49.6 15.5 23.5 13.8 11.7 11.9 38.2 20.6 12.9 19.1 12.9 13.5 26.3 11.7 27.1 18.4 13.7 7.5 16 32.3 5.5 27.2 29.2 13.7 FB:FBgn0036259145471_at 303.2 438.4 360 365.1 478.3 379.5 163.8 407.8 438.9 175.8 312.9 230.4 220.1 146.2 225.4 74.8 242.5 153.8 358.7 478.7 213.8 457.1 402.4 304.9 FB:FBgn0031170142056_at 27 18.7 46.5 51.8 29.4 34.7 76.6 53.5 50.2 32.1 39.4 45.4 26.3 4.8 20.9 24.6 40.7 44.9 22.3 57.4 21.5 35.3 16.5 25.6 BDGP:LD39243.3prime-hit144223_at 71.2 95.7 142.7 110.6 107.2 109 122.2 147.6 97.4 167.2 138.4 140.1 72.5 86.1 151.7 91.2 71.5 109.9 118.1 105.6 92 88.6 128.7 93.6 FB:FBgn0028515154712_at 703.9 369.7 598.3 824.8 730.1 787.4 788 795.1 673.7 730.7 635.2 651.5 982.2 675.3 919.9 901.5 601.1 544.7 454.8 454 451.6 471.8 597.4 391.1 FB:FBgn0030871
149054_at 10.6 36.7 10.6 11.2 49.1 19.9 30.4 15.3 20.4 20.6 13.4 15.3 17.2 20.7 26.2 43.1 25.3 20.4 18.3 21.4 13.2 76.1 73.8 11.2 FB:FBgn0036877
151626_at 19.3 140.3 9.8 32.6 67.5 82.9 17.3 23.7 14.4 17.1 22.8 21.3 21.8 13.3 13 28.1 20.6 20.9 116.3 27.9 13.3 31.9 20.2 103.7 BDGP:LD10305.3prime-hit
152979_at 102.1 64.3 79 113.2 166.5 122 77.6 98.7 85.8 105 129.8 92.2 99.5 115.6 964 64.5 101.7 117.3 104.4 100 125.8 117.5 118.3 126.9 FB:FBgn0037267
154490_at 217.3 328.7 155.8 369.3 354.3 385 277.4 298.6 334.9 196.3 234.8 214.4 366.5 252.5 255.1 195.2 342.8 306.5 317 409.5 333.2 253.4 416.9 294 FB:FBgn0031118146363_at 2.4 4.1 1.4 22.8 3.8 2.3 8.3 5.7 1.2 1.9 4.9 44 12.9 6.6 3.4 13.9 5.5 3.8 17 0.7 9.3 39.8 6.1 11.7 FB:FBgn0032585153261_at 73 113.5 109.9 106.7 105.8 103.1 93.2 106.7 83.7 70.6 63.7 35.8 83.5 70.2 82.7 105.6 107.9 67.6 112.5 74.2 108.4 65.2 77.2 139.8 FB:FBgn0034230145970_at 238 320.2 220.7 268 242.4 302.3 168.7 167.3 175.2 67.3 92.1 90.5 30.9 19.5 85.7 62.4 153.1 71.1 138.1 148.6 128.1 192.9 130.9 121.2 FB;FBgn0031931142423_at 64.8 90 120.6 25 75 43.9 22.3 29.1 57.4 48.4 105.3 117.1 30.2 23 87.4 53.7 36.8 28.5 120.8 188.1 137.5 20.3 80.7 27.2 FB:FBgn0039844144776_at 94.7 49.8 79.6 22.7 81.6 34.1 48.9 40.3 17.1 27.3 20.5 79.3 25.9 26.1 73 16.2 57.9 51.6 69.7 50.2 61.4 60.5 67.4 89.8 FB:FBgn0030167
147780_at 108.8 59.6 89.7 39 16 118.9 62.5 57.8 42.1 34.1 77 54 16.6 37 4 80.9 43.4 63.8 37.6 115 51.5 25.6 51.3 140.3 126.9 FB:FBgn0034865
150993_at 641.1 861.1 1226 573.4 512.3 493.2 538.3 741.9 691.7 979.7 659.9 742.1 694.3 594.9 558.9 576 691.8 707.9 866.6 755.1 999.9 683.3 831.3 592.8 FB:FBgn0039929152306_at 255.3 188.5 197.5 239.5 144.5 148.9 191.9 163.8 170.8 249.7 133.8 213.9 108.1 61.7 92.7 110.1 159 209.2 213 137.1 178.2 128.1 24.5 159.9 FB:FBgn0030531148446_at 83.9 169.5 69.6 168.4 92.7 121.1 38.1 77.4 50.4 66.5 62.9 139.6 37.4 70.9 100.3 104.7 69.1 49.6 75.8 101.4 81 87.6 83.4 92.9 FB:FBgn0035963146621_at 9367 12191 9907 10903 9829.6 11053.4 12369 10335 10743 9272 9394 9866 10291.1 12568.9 9318.1 16936 10894 12715 10526.8 11192.8 11341.5 10980.4 13271,1 9406 FB:FBgn0032987
143098_at 13.2 85.3 17.3 44.6 14.8 58.1 62.9 17.2 66.8 17.6 10.2 25.6 15.6 29.1 12.6 115.4 6 12.4 39.3 8.7 18.6 34.1 10.7 53.3 FB:FBgn0000278
145932_af 25.8 16.6 15.5 94 21.1 65.6 39.9 5.5 16 35.9 40.1 22.4 75 57.4 22.1 9.3 15.1 122.5 38 17.6 50.4 55.7 22.4 158.9 FB:FBgn0031870149740_at 28.2 122.9 238.1 129.2 61.7 26.6 148.4 34.5 62.8 142.6 134.9 63.5 36.2 98 61.7 247.5 97.8 146.7 110.1 53.8 115.6 104.1 18.7 68.5 FB:FBgn0038001150492_at 584.5 471.9 337.7 877 788.5 624.4 351.5 470.4 519.6 481.3 581.2 616.2 799.9 612.8 732.5 237.3 363.6 482.6 514.3 696.9 420.2 787.9 757.7 414.9 FB:FBgn0039166153719_at 481.6 564.5 317.5 649.9 472.9 421.2 291.7 671.7 686.8 365.1 355.4 493.6 542.1 327.1 464.2 312.3 343.6 224.3 500 855.5 594.9 526.6 506.7 308.9 FB:FBgn0030505154836_at 244.8 141.6 226.4 288.6 326.4 291.1 236 9 321.7 185 33.6 194.5 148.6 39 22.9 15.2 46.6 245.2 196.5 123.2 65.7 112.5 113.8 38.6 241.3 FB:FBqn0034021
142458_at 11.5 32.8 75.3 32.9 41.2 14.6 33.9 25 53.8 60.9 15.1 18.2 6.1 10.3 14.1 88.8 38.6 10.4 28.2 11.8 6.5 17.5 24 58.9 FB:FBgn0038448
148511_at 42.6 54.3 44.5 36.5 53.4 41.6 4.8 48.5 48.9 45.3 45.9 25.9 40 52.2 45.9 74.3 55.8 51 56.8 25.8 35.5 18.8 67.7 20.6 FB:FBgn0036072
144670_at 10.5 20.7 18.2 25.5 83 30 27.7 23.3 40.7 25 24.6 19.3 9 6 24.7 97.9 31.1 30.6 109 6.5 47.9 8 34.5 32.1 13 FB:FBgn0029987
149329_at 286.2 360.2 153.8 306 319.7 396.5 132.6 74 8 25.3 204 271.1 292.9 166.8 68.1 143.5 164 2 134 93 1 311.1 134 9 215 7 313 9 142 1 611 8 FB FB gn0037323
150353_at 91.8 25.2 25.9 20.5 78.8 43.6 92.7 131.6 78.6 23.4 57.8 102.5 112 71.4 104.9 62.3 161.2 80.5 83.7 95.7 17.6 41.3 30.2 83.8 FB:FBgn0038944
141781_at 338.1 266.7 370.5 348.9 412.9 414.5 418.6 294.7 290.6 475.8 567.5 529.9 674.9 420.6 641.5 655.4 481.5 267.7 470.2 355.2 376.5 458 421.8 324.6 FB:FBgn0038578
151315_at 4534 6482 1976 1473.2 2156.2 1596.3 3640.3 1414.5 1413.4 1134 2345 5073 793.6 1115.8 892.9 3062.9 1458 1565.9 6304.3 1592.7 2876.9 1246.5 1121.5 9932.2 FB:FBgn0040842

Probe SetID
ywnonhs1
ywnon hs
2
ywnon hs
3
ywshort hs1 yw short
hs 2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
pKB1
pKbnon hs
2
pKbnon hs
3
pKbshort hs
1
pKbshort hs
2
pKbshort hs
3
pKblong hs
1
ph.blong hs
2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
154762_at 183.1 154.9 253 205.3 161.6 193.5 186.7 218,9 283.2 187.6 154.6 227.2 215.6 270.1 195.4 139.2 235.5 197.3 208.9 202.2 155.1 244.9 199 166.9 FB:FBgn0005198144798_at 10.6 92.5 72.9 94.2 19.1 61.6 139.4 67.2 41.6 95 42.5 105.4 19 52.3 51.4 131.6 16.7 154.2 11.5 5.5 11.9 49.1 102.3 11.3 FB:FBgn0030200150980_at 1117 859.9 838 800.3 707.1 736.3 689.7 691.8 737.9 1073 1013 872.1 856.8 606.6 720.7 944.1 783.4 760.9 886 788.6 1066.7 727.1 671.2 797.5 FB:FBgn0039909154547_at 406 443.6 477.1 412.9 254.1 273.6 307.6 568 616.6 323.6 231 358.9 255.4 178.7 275.9 348.6 523.2 469.3 310.8 446.2 365.1 282.7 310.4 282.6 FB:FBgn0030890152236_at 33.9 15.5 1.3 9.8 9.7 3.5 11.2 2.3 1.4 16.1 1.8 3.1 0.7 7.5 18.1 39.1 7.7 29.1 14.5 16.6 12.8 43 5.5 26.2 FB:FBgn0030776152510_at 342.1 404.5 378.6 617.1 495.2 530.7 324.1 236.8 487.7 373.9 437.5 380.4 606.6 511.7 678.4 287.2 359.8 409 426.9 516.3 397.2 539.5 684.4 374.5 FB:FBgn0038178150547_at 1737 1564 1845 1590.2 1539.2 1779.5 2434.2 1811.4 1807.1 2266 2201 1468 2051.7 2078.8 1910.7 2418.2 1898.3 2480.4 1513.3 1696.3 1594.5 1353.8 1301.5 1781.2 FB:FBgn0039262152027_at 23.9 66.5 5.6 15.1 9.7 39.3 4.7 2.7 4.1 6.8 51.3 78.7 8.1 4.4 5.3 12.3 5.3 8.9 75.3 58.4 74.8 30.3 28.3 111 FB:FBgn0035076150909_at 12 17.2 50.6 11 12.6 20.7 17.9 8.5 13.3 65.3 14.6 16.3 9 73.6 10.8 12.5 16 9.2 27.9 23.8 19 15.2 69.5 70.6 FB:FBgn0039799144505_at 473.9 639.1 569.1 465.7 526.1 354.9 211.4 799.5 728 660.1 533.2 364.2 488.3 418.8 4164 223.7 962.8 554.2 402.5 337.7 433.6 643.7 578.6 223.9 FB:FBgn0029742150798_at 8.9 9.3 10.8 19.6 11.1 45.7 85.7 4.1 6.8 11.1 49.7 10 59.3 15.3 26 18.1 7.6 61.8 3.4 15.7 7.2 51.9 7.5 7.2 FB:FBgn0039622151963_at 52.6 53.5 36.7 6.8 59 52.4 61.7 66.5 53.3 21.5 49.9 29.2 23.4 10.9 37.4 62.2 41.8 37.2 56.1 46.7 76 76 8.9 95 FB:FBgn0031589141805_at 576.3 431.6 628.6 614.6 550.3 559.4 742.9 740.1 495.2 674.3 680.5 603 776.1 647.7 621.5 726.2 638.9 715.5 434.1 559.6 437.8 583.4 481 519.6 FB:FBgn0024754154953_at 426.1 500.7 532.2 452.3 521.9 538.4 505.5 498.3 615 592.9 498.8 558.2 578.2 539.2 632.7 465.3 479.6 623.1 503.7 480.6 473.8 594.1 638.2 581.9 FB:FBgn0028529145073_at 56.4 33.2 20.2 32.4 44.5 56.9 29.7 14.5 18.9 35.2 39.1 28.2 23.8 16.3 40.1 11.5 34.4 36.8 43 23.7 77 15 89.1 29.2 FB:FBgn0030574148072_at 114.6 129.5 49.5 127.2 54.5 119.5 78.2 129.1 18.3 110.4 20.7 22.8 150.7 63.4 75.7 26.3 105.6 107.3 16.1 122.7 66.7 145 36.2 24.7 FB:FBgn0035381142534_at 105.9 180.2 164.2 210.4 202.7 205 48.9 227.5 223.9 131.5 105.4 107 208.6 222 264 163.3 241.7 197.8 146.8 265.3 164.6 274.3 269 96,3 FB:FBgn0038754144129_at 149.7 89.1 180.1 164.4 148.9 171.8 135.5 185.1 124.2 178.3 206.5 176.5 146.8 258.1 181.8 98.6 162.9 161.6 85 101.1 71.5 213.2 188.5 133.8 FB:FBgn0026387154183_at 448.4 384.4 474.3 330.9 430 394 396.6 536.2 476.6 512.4 514.2 473.9 439.9 479.9 350 841.8 682.5 560.4 401.7 323.4 467.3 234.9 197.8 483.2 FB:FBgn0025720146269_at 94.9 159.3 66.7 69.8 68.6 18.5 45.2 80.9 61.5 118 95.9 84.4 24.7 106.7 55.8 11.6 86.9 105.4 84 90.6 57 114.1 67.1 106.8 FB:FBgn0032405142552_at 23.3 9.8 39.3 49.7 51.9 59.3 69.1 20.3 40.4 26.4 39.9 49.2 81.5 40.3 24.9 8.6 11.2 81.9 4.7 9.8 12.6 6.8 42.3 17.9 FB:FBgn0003751143076_at 252.2 255.7 263.9 406 386.6 462.2 558.3 294.6 342.7 444.7 710.5 450.3 574.7 629.7 451 806.3 372.2 418.6 242.5 222.5 354.5 313.4 151.9 258.8 FB:FBgn0000109153331_at 143.3 144 200.8 143.9 112.9 119.6 98.7 191 474.7 207.4 189.6 194 91.3 128,9 90.4 207.6 212.7 191.9 128.3 142 163.7 175.2 189.9 139.9 FB:FBgn0036812154532_at 677.6 440 716 711.2 690.2 634.6 1031 689.5 584.9 749.3 538.7 575.6 966.7 718.6 849.7 790.6 480.5 609.8 545.8 550.8 546.9 766.8 682.9 519.8 FB:FBgn0000629151968_at 186.1 172.8 280.8 143 172.2 167 103.1 150 189.4 254 7 239.6 192.1 239 270.1 152.3 118.3 142.1 131.5 177 137.2 162.6 244.9 288.5 160.9 FB:FBgn0033376152773_at 44.2 72.6 63.6 42,3 15.3 22 55.7 56.8 61.7 127.2 24 9 157.5 44.7 103.5 101.7 18.3 53.3 102.6 48.5 11.5 46 82.8 25.2 49.4 FB:FBgn0029611142918_at 1804 2279 2390 1885 2067.4 1902.7 2100.9 1641.9 1559.5 3219 2517 2339 2314.6 2278.3 2352.2 2463.5 1773.4 1393.9 2832,4 2493.8 3126.2 1954.8 1753.3 1567.4 FB:FBgn0040897148018_at 17.7 6.7 33.3 5 5.2 45.9 57.3 6.4 8.6 18 27.3 24.8 27.5 11.9 10.6 38.9 9.2 17.3 20.5 21 4.7 63.6 38.2 8.2 FB:FBgn0035299148125_at 403.4 354 295.2 374.8 378.6 371.5 279.5 280.7 362.1 313.6 351.9 395.8 333.3 388.2 297.4 254.5 258.3 398.4 365.1 229.8 293.8 399.3 420.1 487.7 FB:FBgn0035465142282_at 39.9 20.7 22.6 28.6 87.8 29 14.6 28.5 23 12.4 20.9 20.4 72.5 37.1 26.7 10.1 29 42.3 58.2 55.5 104.7 40.1 31.7 128.3 FB:FBgn0032797154546_at 112.5 89.4 104.8 158.1 157.7 113 107.8 208.7 171 63.5 83.3 76.7 33.4 87.5 47.6 19.8 155.8 189.2 60.8 32.1 126.3 108 73.4 115.9 FB:FBgn0037807150659_at 67.3 95.5 136 66.2 78.9 65.3 19.7 43.7 59.3 148.4 106.4 58,2 158.4 61.7 95.9 34.1 42.7 48.9 14.3 41.5 89.8 90.4 147.1 94.3 FB:FBgn0039428150015_at 178.5 300 296.1 160.3 171 259.2 285 7 459.6 312.5 189.7 276.8 220.1 364.5 178.6 184.8 206.1 308.2 347.8 165.6 176.1 205.4 69.4 182.8 167.6 FB:FBgn0038428148992_at 23.3 16.7 21.8 53.3 18.4 26.6 66.6 69.8 5.9 8 82.2 14.9 31.3 20.9 80.1 45.1 24 4 18.9 15.9 102.3 12.7 94.3 49.2 62.2 FB:FBgn0036784153670_at 113.4 169.7 179.1 117.3 210 139.2 130.5 149.8 235.9 105.3 118.9 108.1 110.7 175.8 148.1 115.4 118.4 214.9 123.1 97.7 104.9 212 181.9 111.2 FB:FBgn0026616154781_at 125.7 201.8 111.7 119.7 101.7 141.2 49.7 15.8 43.2 177.8 145.3 101.2 84.4 71.8 97.8 66.4 93.4 71.1 219.8 63.7 133.6 57.6 29.4 190.8 FB:FBgn0040667154994_at 89.6 100.8 132.9 83.4 97.8 95.9 70.5 174.7 125.9 101.6 56.9 88.6 101 88.7 24.5 60.4 138 99.9 85.4 106.1 61.9 32.2 35.4 102.6 FB:FBgn0034640146393_at 22.7 24.2 21.1 10.5 10.1 32.7 21.5 21.3 29.1 12.1 13.2 6.9 11.6 20.3 12.7 25 18.3 18.3 18.5 27 8.6 17.4 20.6 11 FB:FBgn0032625151385_at 16.9 78 120.1 27.7 58.9 8.5 64.3 53 83.5 35.5 32 51 33.6 51.2 57.3 10.2 63.7 78.6 88.7 60.2 70.6 93.1 91.4 43.1 FB:FBgn0040916146046_at 424.1 392.8 413.2 328.6 350.4 373.4 447.6 366.8 312.4 598.9 537.9 552.4 453.4 473.8 497.1 201.4 314 454.3 521.9 614.4 444.5 646.7 557.4 446.1 FB:FBgn0032063149011_at 790.2 665.5 815 900.9 1011.9 1026.9 1349.2 873.4 767.7 1097 850.7 745.6 1215.1 1183.5 988.5 1483.6 933.3 1057 768 813.6 658.2 738.5 972 789.6 FB;FBgn0036811145363_at 30 183.9 96.4 54.1 22.1 83.5 57.6 85.6 38.9 224.4 81 202.7 113.8 61.2 56.1 27.5 93.1 135 111.2 95.6 139.5 32.9 166.4 163.5 FB:FBgn0031010152750_at 16.1 16.9 23 10.3 30.9 11.4 12.9 10.4 14.8 19.6 17.8 18.4 13.3 57.7 11 48.3 23.6 54.8 14.1 14.3 9.4 17.7 26.8 14.5 FB:FBgn0034937144596_at 1691 1687 1521 2496.6 2436.3 2010 939.8 1464.7 1159.2 1554 1230 1583 2236 2252.7 1972.6 918.3 1472.3 1037.3 2363.4 1932.7 1801.2 1487.7 1561.3 1170.3 FB;FBgn0029869142166_at 2.9 10.4 9.5 9.2 7.5 6.4 12.6 8.2 12.4 9 6.9 5.2 7.2 13.9 8.8 50.1 24.2 9.2 6.9 5.9 23 12.7 15 15.6 FB:FBgn0031496143923_at 22.9 120.8 34.1 27.9 32.9 73.4 117.9 89.3 122.4 20.6 33.6 14.6 23.8 42.8 56.9 123.4 103.4 57.2 19.9 24.9 21.2 23.3 19.1 62.7 FB:FBgn0020379145177_at 1478 1572 1676 1446.5 1481.9 1527.4 1897.6 1212.7 1174 2632 1681 1667 1348.1 1273.3 1379.8 2417.8 1196.1 1562.6 1489.5 1174.3 1743.6 1100.8 971.5 1192.2 FB:FBgn0030733148733_at 28 22.7 35.8 21.4 57.4 33.6 22.7 13.7 18.4 29.3 17.2 29.8 64.1 174 115.5 17.7 8.7 14.5 12.5 10.5 21.7 27.3 15.8 18 FB;FBgn0036391145963_at 26.7 69.1 26.1 104.9 93.9 72.9 20.5 23.9 22.6 16.8 66.9 19.4 173.9 139.9 102.4 35.3 26.7 17.8 43.6 70 50.5 40.9 89,9 26.7 FB:FBgn0031922U 908ë_a{ 54.g 32 9 43.2 26 8 18 9 98.8 26.2 22.6 16 6 27.2 f ë S i 9 i 56.6 29.1 2 i.7 29 13.4 24.4 11.4 67.6 75.6 38.8 37.3 54.6 FB:FBgn0036934151311_at 9.2 8.7 6.9 20.5 25 14.3 10.7 3.9 16 3 49.8 10.5 37.1 17.8 15.5 11.4 17.2 12.1 44.7 11.4 16.9 g.3 11.7 13.6 10.7 FB.regn0040838145884_at 15.1 15.8 20.4 17.6 17.8 20.5 16.5 7.7 9.4 14.8 9 1 19 1 17.2 36.2 16.5 17.6 9.2 15 2 113 14 9 7 7 25 9 ?R 8 R ? FB FB gn003170/1
142863_at 624.3 673.4 1089 843.9 862.3 1015.8 676.1 811.2 768.2 1096 1044 876 1254.5 1095.4 1220.5 550.1 727.7 882 1015.4 634.6 875.3 943.1 966.6 595.4 FB:FBgn0032216151398_at 634.8 600.9 814.9 755.9 839 765.2 813.2 836.6 804.6 899.1 758.7 629.9 920.3 879.5 876.3 1295.7 811.1 889.3 618.3 814.7 504.1 583.6 684 708.8 FB:FBgn0040929153356_at 54.4 60 128.8 119.5 88 101.8 49.6 127.2 104.1 163.9 131 100.1 111.3 31.9 24.2 338.9 129.1 109.4 100 121.1 145 203.5 33.6 156.1 FB:FBgn0032445

Probe SetID
ywnonhs1
ywnon hs
2
yw
3
ywshort hs
1yw short
h s2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
pKb
1pKb
2
pKb
3
pKbshort hs1
pKbshort hs
2
pKbshort hs
3
pKb long hs
1
pKblong hs
2
pKb long hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
146488_at 6.4 9.3 18.2 16.3 9.2 13.3 15 5.7 6.7 11.4 10.3 9.5 5.4 10.8 9.5 9 21.1 22.1 7.2 7 7.1 13.7 12.8 8.5 FB:FBgn0032772145950_at 7.9 22.8 16.4 7.1 11.1 11.7 11.5 6.9 20 16.5 12.3 47.8 5.5 17 55.7 12.3 8.5 7 18.4 20 28.2 52.4 16.1 31.7 FB:FBgn0031902154381_at 326.3 284 453.5 310.4 271.5 314.8 298 424.8 391.3 251 354.4 315 247.2 276.8 263.9 187.4 428.8 451.3 184 184.1 299.4 411.5 340.5 429.3 FB:FBgn0036357151183_at 26.3 44 19.9 22.5 38.7 28.3 45.7 23.4 17.2 35.5 15.2 27.4 37.1 29.4 40.2 40.1 18.2 46.9 16.4 52.8 33.3 48.3 65.9 43.1 FB:FBgn0040706143038_at 269.8 254.2 474.6 392.1 379.5 369.3 135.1 314.8 335.9 529.7 341.4 404.5 516.6 548.1 529.1 143 286.1 350.4 459.6 570 307.5 375 715.2 175.6 FB:FBgn0037207152248_at 20.6 12.6 60.5 47 19.6 13.3 36.4 58.6 28.2 13.9 19.7 24.4 15.4 15.1 48.3 69.1 23.4 12.7 31.8 36.5 6.8 73.9 14.6 4.5 FB:FBgn0028520154198_at 4141 3486 3337 4411.8 4416.7 5095.4 5154.1 4096.1 4148.8 3537 4065 4027 5167 4961.3 5306.1 5554.8 4439.2 4851.4 4331 3590.6 4054.9 4011.4 4578.5 4251.6 FB:FBgn0033902144475_at 46.1 60.9 11.3 23.1 12.4 30.7 32.5 36.5 13.4 5.9 62.6 46.1 7.2 48 26.5 45.3 20 74.9 9.2 7.5 21.3 50.1 42.3 12.2 FB:FBgn0029705144607_at 173.8 266.5 246.8 405.3 417.7 461 395.7 247.4 339.3 133.9 419.1 309.9 512.9 487.6 434.1 470.1 383.1 264.4 288.3 308.1 380 390.6 593.9 459 FB:FBgn0029887153553_at 261.4 280 477.6 296.4 299.5 181 255.4 299 304.8 458.2 208.5 331.1 274 348.2 318.2 210.8 291.1 299.3 237.2 284.7 195.3 262.2 298.3 224 FB:FBgn0029525142179_s_at 266.8 234.9 357.5 250.6 287.7 273.8 358.6 360.6 318.2 335.4 244.6 260.8 302 308.1 348.8 335.9 279.8 302.7 290.8 223.7 241.6 252.7 308.9 385.3 FB:FBgn0028495146994_at 50.3 104.4 82.7 147.4 90.7 26.5 153.4 121 84.1 78.8 167.7 176.1 103.1 73 116.1 151.9 34.7 40.2 159.5 127.6 77.1 32 201.5 187 FB:FBgn0033596150385_at 843.4 778.7 665 880.6 903.2 786.8 651.4 587.7 604.2 701.3 1048 614.1 730.5 804 652.5 590.1 531 584.3 634.5 766.5 691.5 452.9 512.4 662.5 FB:FBgn0038996152739_at 84.7 36.6 65.4 104.9 132 98.8 168 26.5 92.8 64.9 65.6 93.2 48.2 79.4 26.4 95.1 47.5 186.3 65.4 22.7 76 167.4 64.7 116.7 FB:FBgn0029859154882_at 423.9 386.4 453.1 400.7 570.6 607.7 233.2 194.6 301.7 494.9 294.2 379.8 523.8 721 692.6 377.4 290.8 205.6 391.4 259.8 264.7 462.9 528.6 207.7 FB:FBgn0034041149276_at 20.7 51.2 29.6 47.3 12.9 9.4 19 11.1 18.7 34.3 24.5 34.6 4 54 64 37.9 44.6 14.3 32 42.9 56.7 26.3 20.9 45.5 FB:FBgn0037246149887_at 3.3 4.8 5.2 3.1 3.4 4.3 6.9 5.8 4.2 5.2 3.1 12.1 3.1 5.3 2.7 5 5.2 7.7 2.2 2 3.2 14.5 3.8 3.7 FB:FBgn0038218152578_at 2.1 4.8 3.4 14.7 8.2 35.3 2.8 26.1 1.7 24.4 3.9 14.5 18.8 11.3 16.8 8.8 28.9 7.7 10.4 4.9 5.7 6 47.9 2.3 FB:FBgn0037003144320_at 13.9 17.4 12.6 14.1 12.3 15.4 18.1 11.7 8.4 12.4 8.2 11.2 11 9.7 11.7 8 11.4 16 23.8 17.9 11.8 12.2 17.3 14.5 FB:FBgn0028936145953_at 20.6 21.3 26.2 7.7 88.7 71.8 63.1 42 10.9 18 54.4 64.6 35.3 16.3 73.3 17.5 25.9 12.5 11.6 65.2 69 79.8 35.8 61.7 FB:FBgn0031906150109_at 78.5 65.4 90.5 109.3 50 38 63.6 15.4 70.8 25.7 53.4 57.4 13.7 68.4 15.2 91.5 9.5 65 74 51.8 63.4 86.4 97.3 109.7 FB;FBgn0038559146450_at 101.1 45.2 62 19.8 17.1 25.7 26.4 20.2 20 78 35.7 29 28.1 29.5 36.5 32.8 49.9 87.8 58.6 63.6 105.4 128.1 147.1 130.5 FB;FBgn0032707150642_at 435.9 348.5 359.2 313.4 422.9 396.6 479.4 410.8 326.8 361.4 670.9 358 337.1 292.1 351 577.3 400.9 416.9 362.9 306.2 257.5 435.8 296.8 319.7 FB;FBgn0039405144473_at 58 35 41.9 125.5 32.7 57.1 61.1 44.1 39.9 54.9 47.9 68.5 51.9 164.6 33.9 83.5 42.4 54.9 95.9 70.7 83.3 82.2 25.6 42.7 FB;FBgn0029702147600_at 96.9 124.4 36.8 66.9 25.2 27.9 31.1 26 106.5 30.9 40.3 44.5 24.5 26.7 21.6 112.6 43.8 69.6 64.1 27.4 83.7 28.5 100.8 136 FB:FBgn0034587148086_at 299.7 266.5 354.9 377.2 425.1 454.6 272.2 305.1 371.3 358.9 337.4 353 441 255.6 460.1 127.8 468.9 275.8 292.2 429.2 231.5 306.8 578.6 203.2 FB:FBgn0035411144437_at 389.2 222.2 469.7 220.9 240.8 383.9 281.8 209.6 206.2 436.3 460.3 353.8 415.6 489.3 390 274.3 170.1 251.9 154 396.9 286.2 185.8 250.5 282.4 FB:FBgn0029657150621_at 10.7 27.3 6.5 12.5 35 2.7 22.2 15.6 6.5 14.5 11.1 15.3 9.6 62.1 12.8 35.2 14.4 52.5 20.5 12.6 61.4 23.5 41.1 18 FB;FBgn0039380151215_at 7547 6977 4682 6361.6 6607.9 6884.9 4615.3 5016.4 8148.4 4482 6935 7081 5011.9 4654.2 4861.4 5220.8 6808 5848.3 6347.3 8485.1 6529.1 6608.4 7636.5 9933.5 FB:FBgn0040739143603_i_at 1511 781.3 1911 132.3 147.5 767.9 1457.7 423.6 370.9 975.2 2958 779.4 239.1 126.7 239.9 106.1 255.7 337.4 894.1 1388.9 1724 861.2 796.6 678.8 FB:FBgn0010359149401_at 600.7 743.3 702 573.9 787.6 1105.3 1009.7 942.4 860.3 767.1 622 476.5 1005.4 1065.3 781.8 1145.7 1112.3 638.2 701.6 890.4 749.5 1121.8 1494.7 893.6 FB:FBgn0037444149438_at 23.4 81.2 67.7 105.8 96.4 92 55.4 84.9 50 54.3 65.5 94.9 19.5 61.2 98.7 105.7 58.5 83.2 33.2 6 36.2 56.2 137.8 12 FB:FBgn0037495151351_at 128.4 191.6 119.9 158.1 167.6 159.4 158.4 146.4 61.9 191 115.7 302.7 187.6 230.4 98.1 138.6 148.5 213.2 170.9 165.3 135.3 184.7 151.7 158.9 FB;FBgn0040878144995_at 246.4 115 251.8 403 263.5 239.3 310 195.2 228.1 163 190.8 202.7 217.4 388.4 201.5 247.9 177.7 68.8 306.1 248.9 245.5 356.1 199.1 267.8 FB;FBgn0030464151429_at 51.7 41.7 32.9 7.5 9.1 50.7 5.9 30 4.2 28.3 85.5 67.7 40.2 29.5 27.9 8.5 48 18.2 69 82.2 97.5 50.5 47.6 54.5 FB:FBgn0040968142761_at 11.8 19 14.9 17.3 9.5 39.8 17.6 29.6 15.8 28.8 17.7 16.8 15.4 39.3 17 63.7 12.3 26.9 15.5 18.6 12 12.5 23.4 23.3 FB:FBgn0032040142769_at 190.2 121.2 179 301 79.8 195.6 127.4 150.9 86.7 197 219.8 252.1 132.5 218.2 140.7 96.8 88.2 126 171.7 209.7 196.4 322.8 216.8 259.8 FB:FBgn0036678147864_at 68.1 13.5 18 78.5 30.4 101.4 13.5 26.5 3.8 14.5 8.1 40.9 76.1 90.9 22.8 15.2 19.4 126.7 38.1 65.7 5.7 119.9 26.6 43.6 FB:FBgn0035014146623_at 385.4 515.6 684.3 516.1 444.9 483.7 231.6 573.6 754.6 561.8 352 429.3 548 507.5 507.9 231.7 552.4 538.5 586.2 753.4 464.6 475.5 572.4 335.9 FB:FBgn0032989143573_at 2728 2637 5258 2774.6 3127.8 2588 3693.6 3357.7 2520 6742 4887 4386 4042.4 4625.6 4014.5 3740.8 3405.3 3012.8 3781.4 4032.3 3536.6 5118.7 4651.1 3447.1 FB:FBgn0005672154013_at 465.9 571.2 608 722.6 640.6 613.4 496.5 823.6 631.7 466.7 456.9 445.5 739.6 614.6 681.2 634.7 887.2 798.5 329 619.1 377.9 566.4 407.2 403 FB:FBgn0031148151935_at 1069 1545 1344 1054.8 952.6 1000.9 568 1388.9 1820.8 950.1 920.9 794.3 1062.5 960.4 945.7 621.5 1442.4 1155.3 1163.2 1587.7 1221 865.9 1041.8 945.9 FB;FBgn0003717151057_at 37.4 23.5 60.1 82.6 45.5 65.6 133.7 40.3 40.1 20.8 51.4 69.5 32.5 26.5 36.7 28.1 38.2 99.9 33.2 44.6 37.1 41.1 106.8 63.4 FB:FBgn0040570143668_at 320.7 424.3 724.2 388.6 455.4 444 487.3 397.8 442 211.5 155.9 179.1 170.7 264.5 111 205.6 215.5 120.8 314.3 357.8 412.4 237.5 313.8 283.5 FB; FBgnO011638150038_at 55.3 97.4 88.3 71.8 122.7 94.6 54.7 84.1 60.9 125.3 78.1 63.8 72.3 110.2 110.5 111.9 76.6 102.9 61.4 71.5 32 130.2 105.7 99.1 FB:FBgn0038458151227_f_at 83.7 98.6 339.5 177.2 175.8 176.8 42 159.9 125.8 421 113 147.2 234.1 144.7 160.5 15.9 120.1 128.1 101.4 251.7 88 262.1 204.5 76.6 FB:FBgn0040750152822_at 682.3 693.3 728.4 625.4 556.6 758.3 625.9 1002.1 805.6 544.2 609.2 598.1 582 505 457.8 735.5 994.7 823.4 520.6 678.6 598.7 638.3 576.6 744.1 FB:FBgn0029891153880_at 1498 1577 2297 1569.1 1751.6 1470.4 2337.4 2667.7 2253.3 1414 1805 1638 1208.5 905.5 1376.2 3065.9 2938.9 3386.7 1077.4 1420.1 1102 1313.7 1014.7 1212.4 FB;FBgn0026380153110_at 5008 4106 3975 4094.4 4850.5 5109.6 3831.9 5173.8 5511.9 4191 4805 5733 6240.1 5420.7 5629.8 4307.2 5481.6 5990.7 4213.7 4012.2 4089 4941.4 5823.8 5437.3 FB:FBgn0025582149234_at 176.6 218.3 285.4 161.8 183.6 201.8 86.7 176.8 163 321.7 237.6 175.6 186.1 255.7 212.4 77.5 221.6 131.6 227.7 195 114.3 256.5 336.4 80.9 FB:FBgn0037167145205_at 95 247.1 118.8 97.7 103.2 73.9 159.6 58.4 56.2 180.5 168.4 146.2 92 26.8 56 28.5 104 92.6 260.4 133.9 133 106.6 55.2 90.6 FB:FBgn0030781150061_at 27.3 18.8 19.9 71.3 23.5 15.5 56.9 13.6 13.8 15.5 24.1 20.2 37.6 18.3 23.5 16.6 91.4 10.8 18.2 30.8 12.9 20 13.4 14 FB:FBgn0038493150755_at 7.2 24.7 3.5 5.5 7.1 4.7 6.7 4.7 7.9 7.6 4 11.9 4 10.9 7.7 9.5 3 48.4 4.5 2.2 0.9 5.1 2.7 3.1 FB:FBgn0039551141455_at 214.7 213.8 251.1 341 205.2 174.2 132.8 257.2 286.1 226.8 178.7 225.4 368.8 312.1 405.8 294.2 373.3 392.5 201.5 250 226.8 277.9 313.7 176.1 FB:FBgn0032886142414_at 387.1 294.6 319.7 72.7 104.2 72.1 55 59.8 311.2 263.8 407.4 273.4 51.5 69.8 65.6 34.3 155.8 176.5 57.5 384.3 610.6 79.6 49.8 416.1 FB:FBgn0039801

Probe SetID
ywnonhs1
yw
2yw
3
ywshort hs1 yw short
h s2yw short
hs 3
ywlong hs1
ywlong hs2
ywlong hs
3
pKbnon hs1
pitb
2pKb
non hs 3
pKbshort hs
1
pKbshort hs2
pKbshort hs
3
pitb long hs
1
pKblong hs2
pKb long hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
148759_at 32.8 64.5 58.7 106.9 44.4 142.3 20.5 25.7 48.2 25.3 15.6 31.2 51.6 124.3 17.5 25.3 78.5 63.8 39.3 54.8 11.1 14.5 140.9 108.5 FB;FBgn0036437145987_at 144.4 127.3 102.5 102 95.3 137.6 93 87.5 73.2 82.4 110.6 136.7 113 103.4* 114.4 77.4 118 112.7 87.8 156.5 74.7 194.9 135.6 85 FB;FBgn0031955145955_at 705.4 782.4 636.5 796.1 758.6 870.7 375.2 530.5 493.1 506.9 1055 784.7 596 578.1 661.2 563.8 590.7 431.7 1326.4 970 1313.1 595.5 671.5 929.4 FB:FBgn0031912149199_at 97.2 22.1 24.5 25.2 61.8 20.4 17.3 31.8 17.9 35.4 40.4 60 90.2 95.5 85.4 33 21.7 36.3 23.5 61.3 19 65.5 59.2 87.5 FB:FBgn0037115143406_at 378.6 266.8 250.1 274.8 213.9 215 210.1 125.9 171.8 441.2 255.8 275.4 113.5 306.4 210.7 224.6 222.6 316.2 165 710.9 192.2 404.4 303.9 300.6 FB;FBgn0003983145214_at 117.9 51.9 62 162.7 33.7 249.8 287.6 116.6 73.3 16.6 60.1 52.1 88.4 164.3 35.9 42.2 27.4 90.7 144 149.7 138.2 143.9 38.1 32.3 FB:FBgn0030792147239_at 23.1 28.2 35.3 38.7 14.8 60 11.3 3.5 20.3 69.1 26.8 46.1 81.6 5 6.9 28.9 6.4 27.6 5.6 27 4.5 40.4 47.3 26.4 FB:FBgn0034005151463_at 293 364 313.4 517.7 559.6 601.2 210.1 288.4 363.6 337.3 215.6 324.4 509.8 429.5 620.4 194.5 255.8 368.3 373 548.4 162.9 727.9 760.9 209.9 FB:FBgn0041571146868_at 1200 1196 1309 1026.2 1113 1051.2 1606.7 1251.5 1270.1 1692 1091 1236 980.1 868.7 1158.8 1246.2 1099.5 1457.8 1138.3 1149.9 1065.9 998.9 962.3 1177.8 FB;FBgn0033384145663_at 187.6 175.2 235.5 109.2 188.2 182.7 145.7 239.9 261.7 176.7 113.8 209 242 211.3 202.3 56.1 256.6 159.3 177.3 169.6 207.3 246.1 211.8 131 FB:FBgn0031452149922_at 11.9 9.7 83.2 54.1 14.3 12.5 14.3 14.2 3 66 13.5 17.2 10.5 88.2 14 7.8 10.5 5.8 52.6 8.4 5.4 21.1 44 11.3 FB:FBgn0038266153094_at 234.8 261.2 188.7 382.6 478.3 489.8 352.3 314.9 280.7 257.7 210.7 286.7 454.6 423.5 449.6 384.6 245.1 233.1 259.3 254.7 202 431 439.9 332.9 FB:FBgn0005654149741_at 13.3 23 10.7 15.5 15.9 23.7 28.4 40 15.2 12.1 11.5 26.3 12 21.8 22.1 82.9 20 22.4 13 8.7 17.7 58.2 38 19.9 FB:FBgn0038002147737_at 293.1 168.7 225.7 171.6 299.1 257.4 268.3 402.6 323.8 226.2 222.7 183.5 208.4 215.6 228.2 308.4 371.6 365.8 186 195.2 247.1 200.1 264.7 256.6 FB:FBgn0034814149366_at 119 29.7 89.1 11.7 42.4 12.3 27.9 7.8 16.8 17.8 7.3 113.7 15.5 18.3 11.8 16.1 34 11 153.9 91.6 101.1 20.3 16 72.1 FB:FBgn0037400150743_at 39.2 76.9 38.1 54.8 14.7 56.9 41.3 12.6 24.4 29.1 6.1 12.6 45.7 25.1 7.2 8.1 7.4 61 28.9 35.3 55.7 12.6 8.5 31.9 FB:FBgn0039533143661_at 225.9 153.4 195.9 92.6 20.6 92.9 205.1 81.3 9.3 107.4 51.5 219.5 8.4 7.7 134.3 210.6 80.2 92.4 160 183.4 312.4 105.8 38.4 109.5 FB;FBgn0011556144898_at 173.5 540.2 127.9 161.3 211.2 194.7 83.8 37.5 74.4 63.1 304.6 300.6 32.1 90.4 109 54.5 50.9 88.4 281.4 49.5 172 120.4 20.6 365.1 FB;FBgn0030340146380_at 8.8 62.2 6.7 15.5 63.5 18.3 23.6 9.4 8.5 15.6 8.2 9 11.6 23.9 20.5 35.8 9.8 41.1 10.6 5.5 7.1 15.6 19.7 57.2 FB:FBgn0032607149038_at 15.8 31.8 18.1 19.4 34.3 17.9 9.4 6.7 40.6 18.1 23.9 8.5 15.4 21.8 19.7 28.6 10.6 16.7 36.5 53.4 10 22 11.9 52.9 FB:FBgn0036860149519_at 85 87.5 132.1 46.7 25.7 82.9 311.4 155.7 152.1 115.4 189.1 86.6 54.7 11.8 41.4 43.7 130.5 257.1 110.5 87.2 147.2 81.7 71.7 146.7 FB:FBgn0037613151758_at 240.6 276.5 357.5 234 130.2 172.8 108.4 169.8 214.1 352.2 226.2 232.8 259.3 248.9 219.6 150.8 174.9 208.6 274.3 288 331.2 145.6 201.4 210.6 BDGP:SD09884.3prime-hit150328_at 11.2 6.7 9.3 2.5 2 25.7 40.7 28.5 4.1 17.9 21 2 23.1 2.1 2.2 17.1 24.8 1.9 13.4 2 26.6 23.3 3.4 1.8 FB:FBgn0038905150467_at 983.5 1032 579.4 749.2 607.2 653.8 1209.4 752.2 1022.1 523.6 846.2 797.3 773 683.6 780.7 1053.6 853.8 1348.9 915.9 784.9 1059.1 747.3 546.7 945.4 FB;FBgn0039112142038_at 119.8 92.6 144.6 115.2 98.4 117.7 110.6 161.2 210.5 142.9 209.9 129 107 46.1 28.8 212.5 184.6 210.4 172.6 201.2 103.1 207.5 241 242.7 BDGP:LD33778.3prime-hit141911_at 16.4 58.5 17.2 18.8 67.8 40.1 17 14.5 18.6 13.7 21.1 22.6 15.8 46.9 26.3 25.2 41.6 27.4 19.7 21.8 18.9 19.6 44.1 13.5 BDGP:GH24095.3prime-hit151981_at 1117 877.7 1338 1157.8 1576.7 1557.8 1040.7 800 830.5 1376 973.3 977.9 1333.7 1461 1077.3 605.5 341 343 1005.8 867.4 841.2 803.3 697.9 481.4 FB:FBgn0003430150903_at 351.4 344.6 305.1 322.1 289.1 276.9 181.5 281.4 218.9 343.8 327.2 446.7 317.3 320.7 321.3 266.8 279 403.2 346.9 273.1 204.9 478.7 444.5 503.6 FB:FBgn0039789153051_at 426.1 448.9 357 428.6 500.3 507.7 355.2 380.8 513.8 395.6 411.6 330.9 445.7 472 453.5 293.6 461.6 302.6 384.2 404.3 385.3 500.8 655.4 634.8 FB:FBgn0032622143472_at 245.7 339.8 313.1 257.1 285.4 311.2 177.9 230.7 532 286.7 247.9 282.5 345.5 336.4 346.2 214.6 369.1 332.9 329.3 368.5 329.8 335.4 410.2 399 FB:FBgn0004449147096_at 13.3 23.6 38.2 10.8 20.1 4.1 11.7 9.3 36.3 5 36.8 6 7.4 6.3 5.7 19.7 16.8 9 36.1 16.3 14.1 12.9 8.2 12.7 FB;FBgn0033744153885_at 192.2 128.5 45.6 11 16.5 13.1 9.2 19.6 9 28.4 79.2 59.2 4.5 7 8.9 23.2 20.5 9.3 89.2 92.5 111.3 24.1 6.4 81.2 FB;FBgn0032279151719_s_at 112.7 214.5 92.6 113.4 95.4 81.1 140.8 194.4 178.6 75.2 102.7 95.7 79.9 72.8 44.4 155.3 185.3 81.7 136 134.2 88.2 114.5 68.7 132.1 BDGP:LD46228.3prime-hit148889_at 156.7 127.2 122.6 85.8 103.2 87.2 46.6 95.6 51.4 150.9 51.1 147.9 151.5 156.4 134.2 62.5 127.6 143.5 152.8 146.8 70.3 81.5 104.7 89 FB;FBgn0036618154389_at 272.7 310.3 281.7 224.5 203.9 219.5 120.6 425.4 450.1 224.8 187.2 223.2 142.1 100.9 99.7 192.3 377 306.9 216.9 328.6 217.7 205.1 201.6 223.1 FB;FBgn0031233143048_at 399.1 276.7 252.4 547.8 685.7 567.3 259.5 380.4 340.2 221.3 379.9 311.6 594.4 510.8 486.2 233.8 355.9 298.3 241.9 408.6 239.7 667.4 659.9 447.3 FB:FBgn0034615144417_at 8.1 22.2 51.8 47.6 85 17.2 18.7 15 14.2 35.6 35.5 25.3 22 35.3 19.6 7.9 48.3 14.1 22.5 28.9 66.5 151.8 54 59.7 FB:FBgn0029637145417_at 48.3 84 10.6 19.6 22.6 83 11.9 33.6 49.4 36.1 17.3 26.6 34.5 61.1 39.9 39.7 52.7 22.8 21.6 15 30 134.9 131.4 67.6 FB:FBgn0031100148851_at 299.7 386.2 519.3 199.7 316.4 413.5 324.5 613.7 400.9 852.3 439.8 610.7 688.4 918.2 661.3 205.6 713.2 206.9 267 1060.7 361 699.4 422.6 487 FB;FBgn0036576151487_at 228.8 248.6 280.2 153.1 178.3 120.5 79.4 190.2 224.6 63.2 75.4 13.1 70.6 105.7 63.9 96.3 96.9 107.7 108.5 107.5 94.8 93.6 96.2 89.2 BDGP;GH04717.3prime-hlt153796_at 1720 1853 1423 3143.8 3025 2553.1 2168 1463 1427 2015 1512 1210 3086.4 2741.3 2688.2 2269.8 1348.3 1565.5 1878.3 1891.7 1860 1705.5 2898 1648.5 FB:FBgn0010501143889_at 439.7 311.2 394.5 356.6 580.6 274.4 544.2 482.5 330.1 317.8 274.9 244.4 429.4 393.4 331 417.2 348.8 361.2 252.1 325.9 311 390 509.5 362.5 FB;FBgn0017430147668_at 7.3 12.2 28.9 12.3 13.1 8.7 8.6 10.5 8.7 21 6.1 14.1 7.7 11.9 5.1 5.8 5.2 11.7 6.2 10.8 3 23.9 5.7 10.8 FB:FBgn0034714149112_at 30.6 75.6 43.3 12.7 55.5 84.3 63 76.1 79 59 47.3 35.4 43.3 66.6 128.1 92.7 82.1 47.3 57.9 74.5 20.6 66 96.8 5.1 FB:FBgn0036966151847_at 309.8 323.8 291.5 140.6 187 147.2 370 220.4 174.5 309.5 261.8 387.2 234.5 238.5 194.5 294.1 294.6 117.9 274.3 306.3 338.5 239 336.2 426.9 FB;FBgn0028504143632_at 531.3 581.2 626.2 682.7 631.8 594.4 446.5 752.6 738.6 500.4 436.2 562.6 547.4 596.8 692.9 492.5 782.7 777.1 467.7 600.3 519.5 432.3 527.6 536.5 FB:FBgn0010747149024_at 84.7 82.6 186.1 16.4 61.4 29.6 106.3 34.7 55.4 89.3 207.6 69.6 21.1 18 26 94 11.5 93 125.5 276.3 382.6 83 65.3 60.2 FB:FBgn0036833141219_at 458.5 458.8 421 339.8 448.3 499.5 288.6 401.4 486.8 207.4 292.8 208.5 232 137.5 266.4 190.9 334.4 287.3 208.6 244.9 211.3 242.7 182.4 296.1 FB:FBgn0032517142511_at 98.7 69.9 87.2 89.5 87.6 96.8 49.3 151.5 149.5 121.4 99.1 90.1 54.5 87.7 51.9 70 123 193.7 114 98.8 77.1 184.3 154.8 77.9 FB:FBgn0035591144065_at 504.1 336.3 769.9 271.3 438.1 373.4 812.4 456.6 580.8 549.8 523.3 463.8 368 6 460.1 374.2 892.1 663 828.2 387.3 416.7 497.5 487.9 380 424.2 FB:FBgn002S4691484Q2_§t 173:9 162:8 186 207 217 179.9 213.4 210.3 144.3 296.8 377.8 238.6 222.1 224.4 230.9 215.9 196.2 174.9 209.9 223.6 186.5 296.6 173.3 202.8 FB:FBgn0035888143456_at 144.8 185.4 143.7 147 110 148.1 52.7 58.5 173.1 127.5 98.4 104.5 103.3 118.7 111.1 103.9 124.7 135.1 104 138.4 109.5 91.1 136.8 72.1 FB:FBan0004390141979_r_at 4.7 3.4 7.7 29.6 8.6 12 11.3 5.8 10.4 16.7 23.9 2.3 12.7 7.2 6.8 17 39.8 58 31.4 0.9 10.2 17.6 6.6 17.3 BDGP:LD21735.3prime-hit
147073_at 6.9 16.6 12.2 6 5.2 7.4 9.3 7 6.1 8.9 7.9 8.1 6.9 7.9 7 25.4 6.6 7.3 5.7 4.5 7.2 13.6 9.6 6 FB;FBgn0033706148299_at 71.8 85.6 34.3 13.8 21.8 15.2 99.4 16 7 12.8 10.7 57.9 22 19 11.9 137.2 37.9 62.8 36.6 55.6 58 7.5 24.6 29 FB:FBgn0035731

yw yw yw yw yw yw yw pKb pKB pKb pKb pKb pKb pKb pKb pKbProbe Set non non hs non hs short hs yw short yw short long hs long hs long hs non hs non hs non hs short hs short hs short hs long hs long hs long hs PKBYW PKBYW PKBYW PKBYW PKBYW PKBYW
ID hs1 2 3 1 hs 2 hs 3 1 2 3 1 2 3 1 2 3 1 2 3 non hs 1 non hs 2 non hs 3 long hs 1 long hs 2 long hs 3 Flybase ID
148596_at 11 50.7 12.7 9.4 13.3 29.7 15.9 60.1 17 12.3 7.8 16.5 10.6 5.7 10.5 19.8 8.7 15.6 27.4 40.9 24.5 75.5 66.3 4.9 FB FBgn0036195142954_at 218.8 219.4 336.9 143.3 79.1 107.3 191.7 195.7 81 422.7 446.1 204.4 242.3 143.8 191.7 402.2 272.9 250.1 248 267.1 342 342.6 69.1 277.8 FB FBgn0034290149633_at 1556 996.2 1285 2464.2 2354.9 2299.1 3549.1 2139.1 1656.2 1229 1456 1320 2322.7 2440.8 2283.8 4174.5 2128.6 2118 1190.3 1200.2 1180.5 1530.7 1942.8 2214.6 FB FBgn0037819147959_at 904.2 647.2 902.1 501.8 688.3 537.9 1144 592.4 546.1 1407 809.5 1018 542.1 510.4 601 1739.6 870.6 716.4 786.1 733.3 786.3 569.6 587.6 991.2 FB FBgn0035186149417_at 5 15.3 3.9 1.3 4.1 2.1 1.8 1.3 3.3 2.7 12.1 3.5 5.7 1.3 10 4.7 3 9 6.6 4.4 1.1 23.2 2.3 5.4 FB FBgn0037463143574_at 226.5 186.4 194.7 154.3 113.2 159.5 73.8 128.3 162.5 208.5 175.6 131.1 103.4 98 55.5 149.4 116.7 74.4 90.5 191 111.5 98.1 130.9 176.1 FB FBgn0005677150939_at 7.2 8.7 34.6 41.1 3.8 4.2 10.6 4.8 5.4 15.7 6.2 11.9 33.1 5.5 6 15.1 10.7 6.6 6.8 2.6 2.4 29.4 12 28.5 FB FBgn0039842153635_at 148.1 106.1 195.3 107.7 117.1 746 152.4 143.3 184.5 107.4 66.8 106.5 98.1 62 55.7 121.4 69.4 92.3 28.4 103 72.1 108.4 111 78.2 FB FBgn0035627145725_at 114.4 80.9 62.6 114.8 16.2 84 88.5 79.8 61.2 91.5 113.3 108.3 103.7 80.5 128.7 192.5 22.6 86.6 123.3 135.4 107.2 79.5 137.9 134.3 FB FBgn0031553145308_at 68.8 72 103.4 56.9 114.3 155.3 211.3 145.3 197.3 42.6 89.6 91.3 35.5 68.2 60.2 143 113.8 129.1 76.2 85.6 66.8 200.1 248.8 128.6 FB FBgn0030931154683_at 253.9 282.8 310.2 168.6 290.9 238 309 314.9 356.4 337.7 330.8 336.7 267.9 287.2 186.1 369.3 330.3 301.2 347.3 232.1 269 202.4 227.7 266.8 FB FBgn0037608146360_at 12.5 16.8 4.8 24.1 27 3.6 60.4 10.3 12.2 20.6 22 58.5 9.5 13.2 23.2 40.9 26 10 18 25.9 15.8 55.8 4.9 6.5 FB FBgn0032574148326_at 30.4 11 8.6 60 15.4 7.1 27.8 15.9 9.7 12.4 27.9 13.5 19.3 20.5 16.5 16.3 11.8 7.8 13.2 11.1 13.4 16 16.5 21.7 FB FBgn0035768150997_at 42.3 70.4 111.8 79.1 71.4 88.1 34.8 163.7 156.3 107.6 47 76.2 127.6 93.3 100.2 30.4 130.3 92 84.9 204.7 67.6 117.3 105.4 49.8 FB FBgn0039933143627_at 221.1 500.4 360 265.2 199 202.2 257 191.6 177 271.7 300.7 314.1 140.9 70.9 141.2 141.3 163.9 100.9 359.6 455.3 516.2 412.4 223.5 692.8 FB FBgnOO10434154739_at 295.1 238.3 346.2 394.8 409.9 423.5 516.4 438.3 384.3 450.6 265.5 333.7 320.1 357.7 361.5 573.9 334 287.7 306.3 191.4 262.4 392.8 505.8 409 FB FBgn0039144154867_at 97.1 173.1 265.4 83 104.2 118.3 82.1 128.5 162.8 106.1 81.6 47.3 109 62.8 53.2 97.8 92.8 56.8 120.9 58.9 89.5 56.2 98.1 50.5 FB FBgn0037471143846_at 245 143.9 187.9 101.2 252.1 183 138.4 294.3 264.5 124.9 55.8 176.3 73.4 121.5 146 121.4 249.1 230.3 114.6 201.3 128.9 125.5 277.3 193.9 FB FBgn0015800148039_at 390.5 174.6 91.1 187.5 180.8 238 137 185.3 161.1 91.8 192.8 117 237.8 124.2 221.2 222.2 160.5 310.1 281.8 299 249.6 248.3 131.5 208.4 FB FBgn0035332153317_at 8303 7897 6963 10816 13698.2 11465.7 11499 7861.4 7848.1 8263 11507 10781 15211.6 14154.1 13899.9 12922 9754.1 12473 8398.1 8274.4 8783.2 10874.8 10787.5 10785.4 FB FBgn0025366148608_at 14499 15419 8087 16817 17346 18008.1 21773 15667 14830 6610 12109 12823 13213 22730.7 15340.8 15969 14010 15577 14091.5 20112.8 12799.8 22022 23287.1 15356.2 FB FBgn0036213152060_at 460.5 349 294.1 549.3 527.3 518.7 1007.4 454.9 383.3 462.7 556.1 560 640.7 665.6 610 1081.1 379.5 474.4 327 338.1 402.7 683.1 563.5 663.4 FB FBgn0016762147740_at 1468 874.7 847.2 217.4 186.2 242.7 1788.5 545.9 434.3 525.2 394.6 1211 148.5 213.9 192.2 459.1 291.5 300 746.1 1352.3 1570.9 397.4 327.6 613.8 FB FBgn0034819150299_at 400.3 432.8 409.2 321.7 306.9 390.8 492.9 517.8 329.4 568.5 389.7 441.2 353.5 482.4 322.4 741.1 434.7 468.9 339.9 295.7 447.6 492 347.5 549.2 FB FBgn0038861151092_at 52 51.9 48.8 49.9 64.2 64.4 36.8 41.3 58.7 41.4 44.8 57.6 46.1 76.1 62.9 61.8 53.2 65.5 50.6 32.1 27 72.1 64.4 31.2 FB FBgn0040607141598_at 251 324.4 432.6 179 203.7 136.7 222.9 264.8 331.6 348.1 293.2 226.4 148.2 172 171.5 176.4 212.2 212.9 246.9 307.8 270.5 232.3 210 234.4 FB FBgn0032731152419_at 35.9 33.5 28.3 37.3 6.7 19.5 37 40.6 28.8 31.9 48.2 67.4 16.8 5.9 5.3 24.1 5.5 37.4 80.7 21.5 50.1 10.2 3.9 117.5 FB FBgn0028481144925_at 12.4 23.2 56.6 38.4 6.4 14.6 12 10.4 19.1 35.4 18.2 31.4 14.4 6.3 20.6 22.3 20 9.1 9.9 14.7 45 18.4 21 46.9 FB FBgn0030378144946_at 521.7 272.9 400.5 558.2 561.5 573.6 460.9 337 432.4 386.1 430.5 459 546.2 628.2 678.6 474.6 458.7 390.6 269 397.6 247.4 662.2 807.6 318 FB FBgn0030400153182_at 31.9 80.4 90.4 40.1 49.9 44.6 39.6 70.7 64.6 34.2 68.3 35.3 92.6 76.5 66.1 103.2 145.5 85.1 19.7 27.2 35.5 116.8 68.7 52.5 FB FBgn0038362153738_at 1477 1289 1285 1133.5 1168.9 1127 1265.3 1182.9 1254.2 1128 1433 1179 1299.1 1045.1 1004.1 1204.1 1144.8 945.7 1147.7 1341.3 1091.6 1083.5 1276.1 1101.1 FB FBgn0024947150973_at 7 63.8 2.4 1.8 4.2 4.5 5.5 6 3.4 3.8 24.4 13.8 4 1 2.5 2.2 3 6 7.7 2 8.6 3.2 7.5 29.3 5.6 FB FBgn0039899146714_at 132.1 158.6 170.6 140.9 94.9 141.6 175.2 131.8 153.7 196.4 147.7 190.6 143.5 162.1 122.7 188.4 144.4 133.8 163.8 135.1 121.1 162.1 195.1 127.2 FB FBgn0033133151430_at 3.6 3.8 5 4.5 17.6 8.5 3.9 6.7 31.6 7 8 12.1 3.5 5.3 7.2 11.4 15.3 8.9 3.6 3.9 6.4 18.8 34.3 3.3 FB FBgn0040969148302_at 28.5 334.9 30.6 178.8 178.4 124.3 87.3 22.6 21.9 26.6 133.9 118 14.4 25.1 66.9 29.1 17.9 51.5 238.7 34.3 97.4 107 34.2 519.2 FB FBgn0035736143975_at 165,5 220.1 364.6 170.7 121.1 200 241.3 282.5 237.3 287.9 273.7 226.2 198.4 191.4 202.2 316.5 247.4 365.7 197.1 202.2 217.3 216.4 222.1 240.8 FB FBgn0023171143070_at 2.5 10.7 38.8 2.3 7,1 3.9 25.5 44.7 3.7 7.9 26.9 7.7 4 5 5.2 3.9 19.2 7 7.5 2.9 3.1 7.1 19.5 2.6 7.1 FB FBgn0000093149445_at 20,7 4.5 21.2 2 23.9 4 45.9 20.1 35 53.5 6.3 46 1 52.1 19.2 11.1 48.4 24.9 3.2 11.2 24.4 27.6 38.7 39.7 36.5 FB FBgn0037502146991_at 31.5 13 66.1 11.7 12.9 4.7 5.7 5.5 5 33.9 15.9 14.8 19.3 10.7 11.7 29.8 30.7 4.4 10.5 17.2 3.5 17.6 8.7 8.9 FB FBgn0033593145737_at 21.5 15.6 21.1 22.4 8.7 20.7 13.5 23.3 18.7 28.5 15.6 81.7 17.6 70.5 51.1 42.8 33.1 24.8 27.8 18.3 15.5 34.6 37.3 57.2 FB FBgn0031569149151_at 91.8 161.5 98.2 61 80.2 64.6 93.5 213 100.5 120.6 108.9 83 116.2 77 59.7 107.5 138.6 129 44 101.7 74.4 21.3 12.2 110.4 FB FBgn0037031142263_at 884.9 967 1218 1030.1 1230.1 833.7 1073.9 1030.1 917 1188 924.7 923.5 1114.7 944 1042.6 820 1266.9 998.9 1004.1 1407.2 913.8 1356.3 1359.6 1218.6 FB FBgn0035075143376_at 56.9 129.6 89 69.7 83.1 63.7 81 66.5 41.1 48.8 34.2 89.5 73.1 59.9 90.1 109.3 61.6 81 75.2 73.7 47.3 43.7 63.6 63.3 FB FBgn0003715141540_at 622.9 345.8 468 643.6 622 578.8 520 480.7 468.8 637.2 479.5 606.1 529.8 626.8 472.1 605.3 402.7 480.1 553.6 430.6 382.5 710.3 568.2 315.8 FB FBgn0033890142576_at 426.4 651.2 786.2 556.2 605.5 707.4 626 810 683 773.2 521.8 541.9 817.7 946.2 765.6 1111.7 1040.8 676.2 670.2 582.2 525.8 923.9 822.1 756.4 FB FBgn0030175151142_at 34 10.4 5.3 27.4 37.2 11.1 15.4 6.4 12.8 6.6 29.3 12.9 11.4 47.5 17.3 8.7 11.1 38.5 34.3 34.8 3.8 18 23.5 49.6 FB FBgn0040659153909_at 393.6 413.8 427.6 278.1 406.8 314.4 398.3 371 354.5 400.2 454.5 422.4 414.9 371.2 523.4 440.8 419.5 474.2 354.1 367.9 355.7 434.5 480.6 421.1 FB FBgn0030177145269_at 463.2 404.9 385.5 254.9 260.2 267.3 573.5 286 232.7 1188 1101 1012 715.3 776.8 739.9 741 708.6 518 432.6 321.2 446.9 401.9 433.4 588 FB FBgn0030882147830_at 186.1 111.8 124.1 77.1 137.6 103.2 135.2 89.9 148.2 132.9 115.5 138.6 96 152.5 101.9 138.9 131.3 141 158.9 136.6 134.1 168.1 195.4 195.1 FB FBgn0034958150248_at 75.7 102.3 71.2 13.9 97.1 106.6 38.3 51.8 49.4 81.7 85.9 100 87.1 144.3 166.3 69.6 65.4 61.8 103.7 79.7 71.7 142.2 147.8 101.5 FB FBgn0038780151327_âl 301.9 323.1 188.8 268.1 207 190.G 204.1 289.2 190.5 260 4 315 254.5 199.7 266.1 295.5 369 286 243.5 239.3 222 172.6 379.7 287.6 258.6 FB FBgn0040854143362_at 93.3 88.8 119.5 128.5 111.8 86.2 97.7 149.8 103 1 79 8 39.3 117 80 2 137 1 16 5 127 84 5 131 7 79.9 60 9 102 3 98 70 7 63 7 FB FBon0003480149710_r_at 869.8 1979 736.1 543 496.2 1416.6 891.8 564.1 434.1 634.8 474.9 294.6 369.7 1046.8 710.2 535.8 341.3 592.8 1779.9 1703.5 1168.9 784.9 777.3 637.2 FB FBgn0037953143678_at 249.1 456.9 153 58.6 110.8 32.8 186.4 18.2 20.1 25 36.8 200.3 11.2 7 9.5 29.4 10 18.3 387.8 98.6 200.5 32.1 140.7 508.7 FB FBgn0011695146138_at 270.4 259.5 177.7 214 191.2 120.8 186.1 103.5 115.9 283.9 190.9 289.1 90.2 141.8 139.3 103.8 177.3 189 176.7 175.5 230.3 191.9 127.9 301.9 FB FBgn0032213

Probe SetID
ywnonhsi
ywnon hs
2
ywnon hs
3
ywshort hs
1yw short
hs 2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
p K bnon hs
1
p i \ b
2
p K B
3
pKB short hs
1
p K B
short hs 2
PK Bshort hs
3
p K B
long hs 1
p K B
long hs 2
p K B
long hs 3
PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYW long hs 3 Flybase ID
149123_at 1830 1869 1560 1736.6 1989.5 1690.3 1690.1 1511.6 1729.1 1924 2022 1934 1814.1 1940.1 1767.7 1562.5 1568 1558.2 2228.7 1930 1791.4 2261.8 2331.9 2310.2 FB:FBgn0036983150364_at 110.9 153.1 149.5 136.1 112.4 154.7 86.7 283.1 200.8 79.8 155.9 180.5 109.6 87.5 189.2 27.5 176.6 113.2 127 193.8 139.7 151.1 210 113.5 FB:FBgn0038962152906_at 102.6 147.6 84.2 149.1 132.1 160.2 193.5 80.1 169.3 74.8 124.3 104.9 145.6 178.4 183.1 274.4 126.1 180.9 99,4 74.2 113.4 226.5 210.7 245.7 FB:FBgn0037375143416_at 201.1 157.2 79.9 98.8 89.9 140 91.5 90.3 62.2 87 133.5 290.7 50.8 28.5 119 8 34.1 145.7 170.5 134 108.2 134.9 127.8 42.2 28.2 FB:FBgn0004034145601_at 24.1 64.9 10.5 104.7 59.7 80.1 35.7 14.3 17 18.2 68.1 55.6 62,4 51.8 30.8 48.3 36.9 9 10.2 57.9 36.8 70.9 29.2 10 FB:FBgn0031366150892_at 157.8 255.3 283.9 363 194.6 353.4 178 275.1 279.3 562.6 257.6 323.2 325.1 421.1 487.3 467.8 570.5 493.7 227.3 349.3 229.4 332.2 327.1 403.7 FB:FBgn0039775150895_at 49 108 37.9 91.4 193.3 113.8 102.8 24.3 34 94 106.7 101.4 82.3 89,1 107.8 23.4 68.2 122.3 44.6 75 31.1 141.7 69.1 15.4 FB:FBgn0039781145899_at 384.4 282.8 394.3 512.2 387 355 241.9 448.5 427.7 500.4 411.4 384.4 539.8 507,6 596.4 188.4 512.1 424.7 432.6 381.8 344.8 595 394.3 338.4 FB:FBgn0031814154843_at 730.5 688.3 928,8 704.5 543.9 636.4 1005.8 1020.3 1000.9 1096 977.3 661.1 1080.5 906,8 960.5 1563.2 1439.8 1080 915.5 781.6 955.8 1457.1 1615.5 1270.6 FB:FBgn0035850144957_r_at 1127 1455 570.3 1361 1334.3 1583.8 859.7 778.9 1742.1 1057 1527 1033 1241.3 1751.1 1500.8 1198.8 1231.1 1366.9 1621.1 1575.1 1567.2 1245.4 2175.6 1449.7 FB:FBgn0030415152639_at 14.1 11.3 13 63.5 72.2 27.2 101.7 8.4 15.5 30.4 44.6 26.1 86.8 10.6 54.3 57.8 7.1 17.1 51.4 10.3 12.8 46 21.3 36.3 FB:FBgn0036319144868_at 1525 1258 1313 970 831.2 794.8 1365.5 848.4 780,5 1595 1501 1162 1122.7 784,5 863.3 1039.4 1054.4 1199.1 1120.8 882.7 1391.4 762.8 775.1 813.2 FB:FBgn0030292144531_at 805.8 364.6 431.5 472.3 436.4 621.9 401.1 335.3 287.6 637.8 394.6 594.4 338.9 486.9 554 7 350.9 225.8 295.1 191 421 332 667 611.3 495.4 FB:FBgn0029780153014_at 220.4 385 314.9 264.2 191.6 170.5 307.8 257 265.9 494.6 507.5 332.1 230.6 230,1 251.2 450.6 329.1 352.1 370,9 320.7 421 167,9 189.8 171,7 FB:FBgn0035408148282_at 154.4 59.2 161.7 152.3 136.1 84.6 162.3 124.7 61 153.7 134.8 136.3 107.5 78,3 159.6 61.9 193.5 103.2 60,4 67.5 108.3 171.6 146 163,6 FB:FBgn0035703145297_at 1009 923.9 938.6 733.6 959.3 929 980.4 998.8 1016.3 1099 957.5 856.7 740.6 881,7 884.9 937.3 770.6 935.9 719,1 821.6 880.3 814.6 703,1 771,4 FB:FBgn0030915146451_at 12.4 52.9 12.3 41.2 31 15.1 34.4 47 30.7 73.6 47.1 11 39.2 35.6 9.7 7.9 20.8 10.3 25 17.7 8.4 57.3 49.3 57.2 FB:FBgn0032708143183_at 1051 858.9 991.6 1095.2 1264.6 942.7 1085.2 758.6 830.5 1563 1527 1663 2435 2585 2581.9 695.7 598.2 639.9 1350.8 1202.9 1100.8 768.8 791.3 722.4 FB:FBgn0001148146117_at 21.5 12 12.7 9.8 8.6 62.7 18.8 11.9 16.6 15.7 80.6 19.6 54.3 8.8 81.4 10.7 3.3 21.7 52.1 7.2 12.4 16.8 14.2 8.6 FB:FBgn0032176152671_at 61.8 11.1 43.7 22.7 42.8 41.5 5.6 30.5 28 41.1 4.3 63.1 356 15 87.9 35.1 16.1 60.5 28.1 37.3 5 73.5 16.8 5.6 FB:FBgn0032353145040_at 12.1 9 22.5 20.9 8.7 15.8 15.9 12.8 14.9 18,3 11.7 18.3 17 12.5 20.5 18 15.5 28.1 22.8 7.6 10.9 23.9 18.8 12.2 FB:FBgn0030534143602_at 593.1 104 868.8 56.8 40.1 70.3 810.4 49.2 109 95.9 1479 120.1 50 6 47.4 63.8 50.3 44.9 57.1 196 709.3 561.9 84.5 160.4 81.7 FB:FBgn0010357149943_at 18.4 10.2 15.1 25.6 5,5 4.8 20.4 38.1 4.9 15.2 9.6 21.1 32.9 49.3 64.4 4.4 15.3 15.3 7.1 4.2 7.4 6.8 5.3 4.4 FB:FBgn0038309146233_at 97.6 46.6 126.2 14.4 20.4 24.5 81.3 36.7 64.8 112.8 172.9 151.7 18,9 31.4 67.7 66.4 34 16.4 83.3 115.8 198.2 107.2 55.3 109.9 FB:FBgn0032349149525_at 12.6 10 11 8.2 14.8 15.2 7.1 10.6 9.6 16.1 11.4 10.6 11.2 48.6 44.4 15.1 9.7 13.5 12.6 8.2 9.4 21 11.6 9.9 FB:FBgn0037629143128_at 131.4 135 22.5 112.5 95.3 126.9 39.8 66.4 75.4 37.6 97.6 101.8 109.4 102.8 143.9 161.6 103.1 43.4 54.6 102.9 65.3 112.1 35.1 114.7 FB:FBgn0000479151473_i_at 46.2 83.8 30.1 24.4 96 2.4 138.9 44 1.1 43.2 76.1 76.9 121.7 79.3 80.2 105.4 97 39.2 46 58.9 4.5 84.4 52.6 45.1 BDGP:GH01240.3prime-hit151868_at 266.5 296.7 456.7 272 267 167.2 261.7 385.6 302.5 222.9 274.8 327.8 279.2 270.4 298.2 214 280.2 445.9 306.4 247.5 270 194.5 336,7 346.4 FB:FBgn0027605152125_at 675 578.1 546.6 827.6 768 958,9 1013.6 695.9 733.5 599 854.1 939.1 1093.7 1035 1036 1769 1217.2 1042.1 899.4 851.8 855.5 962.8 872.7 948.5 FB:FBgn0039560146960_at 183.7 171.4 177.4 167.8 165 219.5 174.5 151.5 113.5 185.9 139.8 160.1 117.5 134.8 133.6 44.1 107.9 139.2 169 148.9 163.2 215 117.7 187.5 FB:FBgn0033543149169_at 437.9 344.1 265.5 260.2 239.1 280.2 298.5 216.1 236 213.1 362.9 353.6 133.4 194.9 168.5 265.3 224.4 213.4 467.1 340.7 577.1 397.6 180 467.8 FB;FBgn0037065147615_at 929.8 774.1 908.1 1525.1 1343.1 1582 787.9 1029.8 1100.7 1118 1504 1056 1970.9 1905.5 1931.4 1411.7 1143.6 1166 1051.3 1090.6 937.9 1594.4 1388.2 973.6 FB:FBgn0034611146767_at 163.7 105.8 113,5 52.4 12.8 6.8 82.6 23.7 62.6 158.7 165.7 188.1 68.3 19.7 79.7 126.2 100.5 68.1 202.5 123.7 222.3 81.9 64.4 121.2 FB:FBgn0033221144390_at 3.4 14 6 8.4 32.6 8.9 4.9 15.1 4.7 32 5 7.8 17.4 13.5 1.5 5.4 2.3 6.8 14.9 6 2 13.8 6.6 1.2 FB:FBgn0029591141646_at 59.2 76.8 95,9 64.2 52.4 65 48.7 59.2 32.2 44.3 59 16.2 11.1 6.4 3.8 20.3 64.8 73.4 52,3 69.3 69.1 45.5 53.6 33.3 FB;FBgn0032775149598_at 462.6 626.5 184.6 951.8 1277.1 1396.4 115.7 223.6 395.5 222.6 274.5 171.6 484.1 738 679.9 267 186.3 274.2 447.1 593.2 379.3 1014.2 1017.4 221.7 FB:FBgn0037768145065_i_at 45.8 48.4 49.1 62.4 55.5 62.1 6.4 32.3 17.1 51.5 28.4 74.5 24.8 61.1 44.3 88.5 40 82.3 36.5 42.8 8.4 79.2 48,2 70.6 FB:FBgn0030566143028_at 51.6 59.6 107.5 45.5 112.1 110.9 56.4 90 57 71.5 72.8 72.6 53.1 69.7 4.7 25.9 26.6 70.9 26.2 41.4 47.4 161.3 19 57.7 FB:FBgn0035311154589_at 119.5 164.6 122.9 132.8 121.9 149.9 153.8 162.3 145.7 350.2 355.2 364.4 413.9 286.6 407.4 455.3 404 282.4 199.8 233.5 156.6 175 180,6 154 FB;FBgn0031678152410_at 211.3 62.6 79.1 78.9 79.5 62.6 12.8 97.1 99.1 148 8 243 9 2476 238.6 177 171 111.6 235.9 143 147.3 200.5 126.6 113.5 109 79.4 FB;FBgn0015573153604_at 130.6 164.6 198.4 76.7 118.6 101.5 224.2 159.4 217.3 139.6 122.4 135.5 95.7 108.6 84.8 147.6 203.9 210 87.9 119.7 103.7 120 119 114,5 FB:FBgn0037933154717_at 34 44,3 35.3 46.5 58.9 4.9 71 6 26.5 45.5 29.7 39.6 13 7 15 26.1 31,9 54.6 24.5 61.9 25.3 18.9 21.3 39.5 37.8 44.8 FB:FBgn0033840151516_at 2.4 8.2 1.9 4.3 1.7 3.2 2.2 1.2 3.4 34 2.6 36.1 4.7 2.8 14.4 5.4 4.8 22.3 1.1 0.9 2 3.6 2.4 2.3 BDGP:GH 12815,3prime-hit148597_at 89.5 117.6 58.1 79.2 83.7 70.3 28.1 135.5 117.2 161 177.4 98.3 89.8 81.3 130.3 67.3 162.6 172.8 63.7 114 96.9 147.3 94.5 115.6 FB:FBgn0036196154694_at 298.1 305 354.3 758.2 669 664 6106 463.5 610.9 392,1 369.7 405.8 718.6 717.4 821.6 722.3 584.8 858.1 454.2 274.5 331.3 937 992.2 557.7 FB:FBgn0036005146939_at 17.8 17.3 18.8 71.1 16.7 59.4 18.3 13.6 48.8 70 14.4 12.6 30 13.3 84.3 71.2 13.8 69.3 9.2 7.4 20.4 46.6 13.2 51.9 FB:FBgn0033511150132_at 4.2 21.9 8.5 32.9 9.5 6.9 10.2 4.6 4.9 10,6 8.1 12.8 5.2 8.4 4.9 9.8 4 28.5 11.4 3.8 7.3 7.9 9 71.8 FB:FBgn0038604147263_at 359.1 354.6 373.4 515.8 353.6 419 412.9 296.4 293.9 154.6 348.4 301.5 425.1 493.5 423.6 175.4 349.5 313.4 419.9 496.4 389.3 583.7 403.7 402.9 FB:FBgn0034043154845_at 104.5 250.3 278.7 204.8 222.7 198.6 226.4 301.7 269 269 172.6 171.9 24-0.9 204.8 205.3 293.7 215.5 249.3 159 201.6 149.2 249.9 194.1 195.4 FB:FBgn0030874154875_at 136.2 148.6 176.9 97.9 192 206.8 178.3 176.6 224,2 106.6 141.2 182.4 45.6 239.8 171.7 123.6 220 189 79,3 174.3 192.3 216.4 132.5 202.1 FB:FBgn0024975143264_at 3679 3221 1715 1876.4 1575.6 1875.7 3208.2 925 957.5 1034 2183 2727 331.8 745.2 492.1 1354.4 854.7 1098.4 4163.8 2550 4648.1 1339.8 1167.2 4203.4 FB:FBgn0002772143565_at 462.4 701.8 610.8 499.9 306.6 464.8 291.2 533 574 488.2 521.4 435.1 513.5 466.9 460.6 304 470.5 347.4 169.2 416.2 173.7 188.4 173.2 152.8 FB:FBgn0005640
147446_at 24.1 15.5 40 35.8 19 42.9 12.6 34.7 13.6 37.6 59.9 28.8 55.8 30.2 157.5 30.2 36.8 45.9 119 118.5 77.9 21.8 36.8 40.2 FB,FBgn0034359
J48236_at _15.3 28.9 32.5 3.7 7.9 8.7 7.7 7,2 4.4 200 258.3 246.1 234.6 294.4 342.9 174.2 166.6 159.6 197.4 96.3 267,3 91.4 165.9 207,6 FB;F^n0035638

Probe SetID
yw
hs1
yw
2yw
non hs3
ywshort hs1 yw short
h s2yw short
hs 3
ywlong hs1
ywlong hs2
ywlong hs
3
pKb
1pKU
2pKb
3
pKbshort hs1
p K bshort hs2
pKbshort hs
3
pKb long hs 1
pKblong hs2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
145832_at 92.5 93.6 53.4 11.9 79.4 66.8 105.3 56.2 19.3 124.8 149.5 31.1 39.4 100.3 87.2 28.6 76.3 26.5 62.7 14.4 92.7 87.5 130.5 58.9 FB:FBgn0031720151585_at 268 449.4 505.3 444.3 379.3 402.2 268.9 288.1 547.2 497.4 376.2 365.9 508.6 250.9 394.4 262 401.1 350.7 424.9 338.9 299.6 297.9 500.7 272.5 BDGP:GM06601.3prime-hit151139_at 4.9 6.5 5.1 5.4 6.3 10.9 8.4 13.2 3.3 5.4 6.4 4.4 4.9 9.2 6.3 9.6 8 8 4.7 3.8 4.2 10.9 10.1 4.9 FB;FBgn0040655143687_at 835 583.2 749.6 742.8 637.2 725 407.7 527.9 497.1 784.2 645.6 667.7 691.8 565.1 640.3 429.2 610.7 503.8 720.1 617.3 760.8 337.4 538.1 490.1 FB:FBgn0011745147954_at 227.3 337.9 332.9 425.8 383.5 473.5 240.9 501.4 391.1 388.3 144.5 225.3 219.3 282.3 367.6 146.8 343.8 287.2 278.1 366.5 293 423.2 339.6 189.3 FB:FBgn0035180147976_at 67.5 8.9 11.9 54.7 22.9 25.6 21 9.2 16.3 26.1 75 11.2 107.9 53.8 21.2 62 38.8 110.2 22.8 17.9 15.3 20.3 28.1 37.6 FB:FBgn0035218145945_at 28 80.8 24.8 45.1 66.7 44.5 5.3 35.8 64.1 5 33.3 17.2 67.6 16.1 53.7 11.7 24.5 86.3 37.1 64.9 20.8 36.4 90.7 16.3 FB:FBgn0031892149010_at 86.5 70.1 91.4 90.8 56.6 92.2 105 191.6 128.7 75.6 154.3 90.7 238.8 125.8 131.7 280.8 221.9 185.2 48.1 56.6 52.8 182.1 235.9 58.5 FB:FBgn0036810141792_at 251.2 127.3 258.1 192.7 215.7 163.6 148.9 253 195.6 380.4 221.6 153.3 244.6 269.3 290.3 372.5 391 446.1 256.1 154.6 184.9 93.3 131.1 156.5 FB:FBgn0034748152610_at 49.6 76.3 77.1 48.8 76.8 75.5 51.1 77.8 73.4 56.3 44.5 63.2 58.9 100.2 68.1 44.3 65.1 61 47.8 46.6 44.3 82.7 71.3 36.6 FB:FBgn0036611152316_at 187.8 195.2 138.8 188.1 270.4 240.7 25.6 110.2 74.9 220.7 151.6 172.2 97.6 196.5 195.2 124.1 184.3 22.6 138.6 149.9 149.1 140.8 170.7 124.3 FB:FBgn0037023152365_at 2139 1675 1339 2151.6 2363.5 2263.8 1396.8 1906.5 1928.5 2132 1920 1766 2590.2 2376.2 2077.5 1091.2 1771.4 2007.4 1715.9 1514.2 1787.9 1653.7 2073.7 1601 FB:FBgn0028690142607_at 440 359 263.1 367.8 328.3 306.2 268.1 616 501.6 318.7 301.9 356.7 367.4 307.5 394.3 324.9 424.6 462.6 382.6 369.4 374.6 376.4 307.9 226.2 FB:FBgn0033390141347_at 242.3 223 237.4 273.5 372.6 231.3 329.3 253.5 205.4 343.8 313.1 347.9 321.9 334.4 464.7 433.8 315.9 367.3 298.9 287.1 237 355.6 392.2 284.2 FB;FBgn0030360143663_at 254.1 446.4 542.2 234.5 210.4 163.8 287.9 328.4 315.5 838.6 755.4 543.4 262.8 360.5 264.7 344.2 600.4 650.8 612.6 750.4 959.8 464.8 411 1032 FB:FBgn0011584145458_at 94.6 74.1 96.5 121.9 83.5 129.5 114.7 107 60.2 106.6 121.9 163.7 153.7 74.2 99.5 31.2 94.3 200.2 100.4 88.5 62.3 122.7 125.3 50.7 FB:FBgn0031154148131_at 457.6 257.1 196.6 308.1 387.9 388.1 379.9 353.6 311.4 234.7 304.7 365.4 356.4 229.3 276.5 254 178.3 405.9 196.5 238.4 244.3 349.9 264.6 98.4 FB:FBgn0035475142717_at 316 304.9 671.7 475.1 407 521.7 1153.7 775.3 644.9 614.9 312.1 268.4 472.9 560.8 618.3 1024.8 756.2 799.9 367.5 330.4 320.3 552.9 532 631.8 FB;FBgn0003741144203_at 574.1 552.9 652.5 594.1 506.5 566.6 678.7 733.6 552.2 724.1 580 610.7 672.2 641.4 579.5 737.1 586.3 703.6 452.6 468.9 626.9 360.2 504 611.1 FB;FBgn0028398150754_at 42.8 20.6 73.4 7.1 30.2 52.5 83.7 47.4 37.3 55.1 56.6 39.7 26.9 73.3 15.6 11.6 27.4 21.2 34.8 51.8 28.6 20.5 81.4 37.4 FB:FBgn0039550153727_at 21.8 81.8 43.9 28.5 51.6 4.6 160.1 83.5 53 36.1 13.7 66.9 42.2 41.7 16.4 17.7 85.7 79.6 54.9 45.6 56 93.2 81.2 98.9 FB:FBgn0028962154174_at 2801 1991 2306 2143.2 2092.8 2174.7 2607.4 2451.1 1767.9 3026 2362 3576 2576.8 2787.6 3031.2 3356.1 2092.6 1972 2359.7 2182.3 1875.4 1818.4 2397.6 1557.1 FB:FBgn0030943144099_at 3.1 6.3 17.3 4 9.8 27.3 3.5 7.3 1.9 8.6 4.7 4 7.4 34.3 2.2 65.5 2.7 5.5 4.5 5.5 10 20.3 36.4 3.1 FB:FBgn0025821149496_at 13.4 22.9 18.8 12.2 15.7 26.4 9.2 13.4 24.3 21.5 19.7 11.3 15.9 32.9 9.9 21.3 14.7 15.9 19.8 8.6 58.3 41.9 18.8 10.5 FB;FBgn0037584149628_at 336.3 281.3 202 361.6 332 361.8 205.7 286.2 345.1 285.5 279.5 311.2 424.1 299.9 268 122.1 322.9 285.4 297.7 333.4 267.5 386.2 361.1 430.5 FB;FBgn0037808150152_at 6.9 63.2 18.8 9.3 6.8 6.1 9.4 7.6 5.4 24.8 13.6 19.5 15.4 21.1 5.7 9.7 4.7 6.5 9.3 3.6 3.3 7.8 5.3 7.7 FB;FBgn0038632151527_at 29.5 67.8 73.9 123.4 136.6 93 100.6 49.5 114.9 72.7 111.3 35.9 60.6 74.6 189.8 178.3 92.2 28.2 90.4 17.4 15.1 125.6 25.1 27.4 BDGP:GH14535.3prime-hit144051_at 388.2 388.8 389.6 452.4 371.6 269.7 343.8 516.5 781.8 361.6 297.1 382.2 277.8 331 314.7 274.6 449.5 390.8 431.5 545.7 623.1 348.5 529.1 399.9 FB:FBgn0025186147388_at 2254 2320 1628 4230.5 4410 4463.7 3028.7 2676.3 3915.5 1618 2033 2414 4509.3 5325.9 4259.8 3309.4 3166.5 3124.7 2850.6 2109.3 3295.2 2147.2 2548.5 3539.4 FB:FBgn0034258151557_i_at 190.9 334.1 241 369.3 221.8 259.5 66.2 132.8 240.4 197.3 249.8 223.3 337.6 339.1 204.5 143.7 207.1 189 321.2 166.6 267.4 378.7 459.7 150.6 BDGP:GH23604.3prime-hit145916_at 205.3 123.7 159.3 217.1 219 217.8 140.7 229.7 250.1 89.6 201.8 148.5 205.5 223 218.5 55 208.7 268.5 167.1 114.2 171.7 269.3 206.3 134.2 FB:FBgn0031840142655_at 75.6 68.3 123.9 28.6 106.4 22.6 85.7 79.6 64 96.7 88 28.4 29.2 35.1 62.3 84.3 87.4 117.6 44.5 54.1 57.2 140.7 129.9 81.2 FB:FBgn0038395150789_at 8 10.2 7.5 64 5.9 5.2 13 7 12.2 8.9 9.3 8.1 5.8 10 14.7 10.7 9.3 7.9 10.8 10 18 7.7 20.1 11.8 FB:FBgn0039609142946_at 252.1 68.5 257.1 163.3 162.3 184.6 303.6 193.2 121.3 290.6 161.2 207.1 255.5 234 173.2 284.1 134.9 128.1 133.4 196.9 205.6 139.4 155.2 169.4 FB:FBgn0020129152047_at 237.5 227.5 296.6 232.3 213 173.9 208.3 223.2 222.6 359.6 231 224 233.7 201.1 215.2 330.8 388.8 282.6 151 190.5 207.4 135 214.1 204.3 FB:FBgn0027583147552_at 191.8 164.1 226.1 99.5 140.9 151.9 207.2 187.7 107.4 178.5 132.4 137 76.5 31.4 108.5 198.8 145.7 131 335.6 286.3 376.8 81 47.3 171.7 FB:FBgn0034518148114_at 53.7 118.7 53.3 85.9 82.6 77.8 64.5 116.4 121.9 127.6 126.4 105.7 48 71.9 114.9 83.9 117.6 134.6 62.3 117.4 54.8 113.6 59.6 65.9 FB:FBgn0035449148233_at 289.9 501.3 449.5 378.9 376.6 247.2 454.7 294.9 482.5 547.4 378.6 445.4 318.9 338.2 471.3 531.8 506 501.2 552.1 575.9 589.4 585.5 571.2 715.6 FB:FBgn0035626149520_at 270 318.3 267.7 313.1 315.3 437.2 260 592.7 496.3 34.9 42.7 80.8 152.8 90.2 304.1 264.1 659.1 373.8 186.3 292.8 53.3 53.4 236.5 145.3 FB:FBgn0037614143628_at 737.5 662 971 786.4 685.2 618.9 978.9 847.8 686.7 1150 1009 768.3 712.9 672 509.2 1209.5 671.4 909.3 618.6 502 685.1 692.1 433.4 579.1 FB:FBgn0010438150360_at 111.6 227 67.3 110.6 62.4 194.5 50.8 53.5 125.5 37.6 53.9 145.5 44.6 91.2 65.4 71.9 137.5 78.3 109.2 68.8 138.3 136.4 90.7 255.5 FB;FBgn0038958151260_at 295.9 305 572.8 296.4 345.5 301.2 593.6 462.7 469.6 529.3 264.1 327.4 393.4 419.8 320.1 510.7 547 532.3 289.2 253 309.1 368.3 317.2 374.1 FB:FBgn0040789154350_at 223.5 203.3 244.2 181.6 165.1 165.1 203 181.5 159.6 239.2 240.9 226.5 111.8 130.7 227.9 255.3 260.8 212.8 160.2 224.3 196.9 86 93 122.9 FB:FBgn0033735145762_at 142.2 74.5 168.2 149.7 143 109.6 132.8 216.8 227.3 245.8 155.2 156.2 237.6 238 189.8 209.6 180.5 73.2 80.6 101.6 113.6 276.2 391.9 149.7 FB:FBgn0031608153154_at 399.1 345.3 378.2 329.6 256.7 353.1 194.5 342.8 351.6 423 330.5 464.7 337.5 295 450.3 189.5 374.1 389.4 384 397.1 400.7 357.4 405 366.2 FB;FBgn0037877148838_at 3.9 42.2 13.5 21.5 8.7 17 3.4 18.2 17.1 11.2 6.8 50.3 40.9 17.6 39.3 59.8 13 21.5 5.5 7 13.7 5.6 63.6 26.9 FB:FBgn0036552154289_at 1004 829.1 850.6 1203.2 1128 1055.4 759.4 990.4 1011.6 886.7 1003 803.3 1178.5 1229.5 1217 729.7 989.2 937.3 882.1 811.3 936.8 829 1031.1 827.4 FB:FBgn0032640150736_at 101.2 151.2 238.9 218.8 153.5 168.5 113.6 192.8 140.5 104.1 40.2 32.9 15.3 43.8 42.2 11.6 20.2 38.1 153.9 147.7 178.7 103.8 160.5 133.5 FB:FBgn0039523146752_at 7.4 50.2 37.4 53.7 56.8 9.6 83 50.1 98 37.5 11.4 14.3 14.2 25.6 9.6 41.5 8.3 44.4 6.7 46.3 51.1 15.3 10.9 8.7 FB:FBgn0033196150259_at 177.1 192.4 135.2 142.8 99.1 140.1 73.9 120.5 108.2 190.9 149.1 184.1 116 164 140.8 33.6 144.9 123.1 142.6 78.7 110.5 137.5 190.4 109.2 FB:FBgn0038792142211_at 239.1 241.1 277.6 274.8 241.1 231.9 330.2 215.3 176.9 317 208.7 288.5 335.5 282.7 277.3 252.1 245.6 149.8 237.6 284.4 311.1 358.2 277.2 234.9 FB:FBgn0034392150750_at 12 69.7 76.8 43.6 18.6 21 19.5 35.6 10 84.4 26.2 18.3 12.9 42 17.9 26.6 62.1 11.5 51.4 13 7 24.6 17.2 20.7 FB:FBgn0039546
142371_at 80.1 82.6 90.6 102.5 21.5 105.5 32.4 43.8 37.9 101.2 79.5 50.3 82.5 103.7 188.7 4.2 83.8 51.6 45.5 93.2 72.1 216.2 131.2 17.5 FB:FBgn0016684
150246_at 21.7 10.7 10.6 7.5 9.5 10.7 13.5 14.5 15.3 15.9 12.1 10.2 12 13.3 13.4 8.4 11.6 11.6 55.4 6.8 9.8 16.8 14.4 9.3 FB:FBgn0038778
155030_at 656.7 638.1 704.3 707.7 658.7 728.6 480.5 578 670.5 631.1 607.7 516.7 744.5 624.6 585.6 679.7 581.9 512.6 542.9 570.6 568.7 614.5 548.5 485.5 FB;FBgn0027786

Probe Set 10yw
hs1
yw non hs
2
yw non hs
3
yw short hs
1yw short
hs2yw short
hs 3
yw long hs
1
yw long hs
2
yw long hs
3
p K H
1
p K B
2
pKb
3
p K B
short hs 1
pKU short hs
2
pKUshort hs
3
pKU long hs 1
pKblong hs2
pivb long hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase 10
154778_at 483.9 578.2 568.3 364 389.3 413.7 331.7 755.6 594.3 325 409.5 307.6 369.4 282.5 326 318 649.9 630.1 305.6 375.6 336.6 428 425.9 260.2 FB:FBgn0035321151461_r_at 62 108.3 94.6 88.5 81.3 38.8 100.8 81.6 94.1 84.2 78.3 96 136.1 99.9 93.3 152.6 129.8 92.8 60.6 82.4 74.9 81.8 83.3 62 FB:FBgn0041005143411_at 168.6 207.1 255.8 239.2 289.2 211.4 183.1 92.9 154.8 240.5 158.6 170.2 134.5 200.5 150.7 65.9 203.8 72.5 212.9 349.7 270.4 452.3 440.5 211.8 FB:FBgn0003997146415_at 62.9 108.2 39.4 96.3 93.1 85 18.2 55.8 24.4 112.2 34.8 11.7 30.5 27 52.9 143.3 8.3 20.9 51.5 47.1 19.2 113.1 32.1 95.9 FB:FBgn0032654141922_at 19.7 12.5 5.2 49.8 54.3 7.1 54.3 9.2 6 11.2 8.6 7 7.4 31.4 8 6.2 7.6 16.7 7.8 16.3 6.2 10 14.6 5.8 BDGP:GM02310.3prime-hit144604_at 170.1 147.1 200.7 202.1 183.4 291.7 147.9 175.9 204.6 188.3 196.3 135.2 187.7 195.4 202.2 121.7 177.5 205.9 127.4 109.9 150.6 321.9 300.7 218.3 FB:FBgn0029884152483_at 36.1 13.3 33.3 48 34.9 13.2 34 20.9 49.1 15.8 38.1 32.4 19.9 19 16.7 39.1 11.6 35.7 16.7 15.9 18.5 19.8 9.4 13.5 FB:FBgn0035915141896_at 20.2 8.9 14.2 5.3 42.1 10.2 7.8 7.2 7.3 6.8 13 20.2 30.9 51 33.4 6.6 38.2 6.3 10.2 4.5 6.7 18.2 27 3.2 BDGP:GH 17661.3prime-hit146094_at 143.8 99.3 86.3 125.6 133.7 129.4 27.3 98 56.1 150.2 121.8 162.9 122.8 84.8 157.9 14.2 90.7 115.6 91.9 131.1 81.2 159.8 100.7 119.7 FB:FBgn0032129141374_at 94.6 56.8 120.8 73.5 43.8 70 54.4 85.3 40.7 157.4 48.1 97.5 87.6 68.4 53.7 155.3 47.9 103.8 53 35.7 36.8 143.7 57.2 63.9 FB:FBgn0012042143272_at 2.1 7.4 1.8 18.6 2.4 1.4 13.3 2.9 9.4 7.3 4.8 18.6 1.6 15.8 23.2 5.7 1.4 5.9 15.4 12 0.6 4.7 23.6 0.9 FB:FBgn0002855150995_at 28 60.7 31.1 8.6 99.9 69.8 78.5 50.6 36.2 63.8 30.8 49.3 27.5 38.7 25.7 114.9 12.4 7.1 37.5 61 51.7 89.7 38.6 22.1 FB:FBgn0039931150724_at 80.5 33.2 21.9 162.9 90 149.4 230.7 130.9 76.3 72.6 153.1 36.2 155.3 75.3 29.7 105 218.2 68 79.8 103.4 152.4 127.5 80.6 128.2 FB:FBgn0039510149080_at 314.3 236.4 298.7 283.7 277.4 242.7 421.5 382.2 270.6 358 318.2 252.6 328.5 302.8 295.5 449.9 294.7 341.1 299.9 381.5 296.4 345.2 387.1 333.5 FB:FBgn0036922143713_at 228.1 139.8 272.8 367.9 431.7 336.5 314.3 240.4 200.7 276 266 378.6 518.7 419.1 645.7 705.4 205.1 418.1 263.8 313.7 247.4 174.1 333.9 213 FB:FBgn0013531153745_at 11.3 23.3 50.4 9.3 18.9 18.5 35.1 24 13.3 12.8 63.5 15.2 60.5 90.4 73.7 26.2 16.3 15.4 6.1 10.9 28.2 25.7 68.1 17.8 FB:FBgn0037542149268_at 44.1 22.1 13.7 29.8 23.4 7 33.7 76.9 35.5 32.6 8.4 79.1 33.2 10.2 71.5 72.3 2.6 43.3 17.8 21.2 14.8 93.7 68.6 3.6 FB:FBgn0037223147022_at 97.4 168.2 278.2 73.3 59 49.9 139.4 84.3 90.8 56 25.3 6.4 20 18.7 56.7 58.9 13.3 80.8 64.6 96.3 97.3 127.4 95.2 66.3 FB:FBgn0033634151862_at 70.6 25.3 91.3 54.2 29.3 77.1 60.4 61.3 53.7 70.4 57.5 84.2 51.7 48.3 54.4 49.2 58.8 60.1 68.3 61.4 9.4 99 65 69.2 FB:FBgn0023533144290_at 19.6 31.2 118.3 19 16 16.6 17.9 21.9 40.1 99 13.6 13.7 20.2 37.7 76.5 17.3 55.7 21.6 11.4 21.1 14.9 34.8 28.2 23.4 FB:FBgn0028888149219_at 148.4 37.4 110.6 27 80.5 123.4 93.2 37.5 70.2 89.2 141.1 109.4 102.6 195.6 112.7 113.6 64.7 32.6 64.5 111.1 112.6 109.3 169.5 144.9 FB:FBgn0037147152165_at 2.7 8.9 0.3 4.2 6.9 1.8 5.3 2 9.4 11.8 1.6 2.4 1.8 17 1.5 4.5 8 9.3 10.8 4.4 2.6 4.5 1.4 5.2 FB:FBgn0037158151410_at 4958 4173 4840 4194.5 4216.5 3923.5 3883.5 3308.3 2954 5527 6614 4935 4298.9 4671.9 4182.8 4223 2862.9 3918.5 4395 3668.9 4505.8 2195.8 2714.6 2818.4 FB:FBgn0040943141570_at 321.1 278.4 258.9 389.8 347.6 351.8 373.4 297.4 282.3 286.1 281.7 314.1 398.4 348.3 416.7 308.2 297.1 369.3 305.2 294.4 350.9 214.6 294.2 266.6 FB:FBgn0039175154448_at 877.5 911.4 793.4 903.4 807.5 749 605.9 1119.3 1542.4 776.5 1385 824.2 1041.4 920.4 1051.4 763.7 1051.1 1334.9 738.2 1111.5 921 514.5 824.4 784 FB:FBgn0037834142037_at 116.9 60.6 85.2 54.8 60.5 54.5 86.5 151.8 116.9 134.7 83.9 79.3 34.2 35.2 56.8 232.3 134.7 109.9 76.1 111.5 42.6 75.2 110.2 67 BDGP:LD33386.3prime-hit146254_at 107 67.2 23.2 30 31.7 25.3 25.6 22.6 25.6 21.6 24.5 40.5 20 16.2 21.4 16.3 16.1 15.9 27.6 120.1 105.2 25.1 135.2 16.6 FB:FBgn0032379154412_at 380.6 435 491.3 429.7 376.9 304.2 220.3 692.3 812.6 451.4 293 263.4 406 332.5 417.7 217.1 682.7 491 449.4 529.2 346.8 362.4 341.3 238.3 FB:FBgn0010550150934_at 374.8 382.3 313 311.8 300.3 300 366.3 293 353.7 400.8 323 329.6 359 228.5 308.9 275.1 341.9 506.4 364.6 336.1 337.9 279 244.4 226.4 FB:FBgn0039835151503_at 179.5 230 165.4 228.2 146.4 128.7 272.2 259.1 214.7 316.5 376.9 383.9 332.5 330.4 253.5 490.9 476.1 397.4 272.8 219.9 244.4 273.8 234.2 392.1 BDGP;GH 10173.3prime-hit141341_at 12.7 43.3 7.2 4.2 21.5 54.7 9.6 5.2 15.3 51.6 6.8 11.9 18.6 46.4 14.9 27 14.6 43.7 14.8 7.9 12.9 10.4 9.6 10.2 FB:FBgn0033848148275_at 57.7 42.8 111.4 9.8 16.7 8.2 10.4 3.1 9 117.9 36.5 78.5 25.3 18.4 15.2 9.2 10.8 15.4 7.9 30.9 3.5 10.2 54.7 17.2 FB:FBgn0035694151800_at 136.4 118.7 111.4 19.2 189.6 31.7 109.6 44.1 51.5 60.4 38.2 45.3 55.2 177.2 47.4 43 52.7 21.2 195.3 175.8 331.6 80.2 70.6 181.1 FB:FBgn0034229152696_at 19.2 32 18.5 13.1 17 20.1 13.4 90.9 35.2 14.9 12.3 8.8 38.3 11.9 11.1 10.9 28.4 17.5 13.1 14.7 10.1 20.3 7 11.5 FB:FBgn0030066147808_at 8.5 13.3 13 14.7 4.9 8.1 10.4 11.1 12.3 11.7 10.4 4.5 11.2 9.2 6.6 12.7 10.9 14.4 5.9 6.7 8 14.1 10.6 8.2 FB:FBgn0034910150829_at 28 28.4 26.1 10.4 23 9.9 45.1 37.1 72.2 64.7 23.1 19.5 34.6 27.6 15.4 135.8 69.8 25.4 31.9 18.8 78.9 28.4 34.9 76.9 FB:FBgn0039676146352_r_at 5948 4202 2869 3984.1 4338 4646 3892.7 3364.1 3671.3 3799 4703 5784 3009 4347.4 2703.7 4032.9 3634.6 3260.6 5191.8 4702.4 4538.6 4679.2 3918.7 5498.8 FB:FBgn0032538149474_at 86 114.3 86 77.4 141 114.3 113.7 95.3 78.2 114.6 159 82.2 203.6 123.8 88.2 132.2 159 166.5 114.5 131.5 148.1 151.2 125.7 166.2 FB:FBgn0037548149745_at 6.3 34.9 8.5 5.4 4.2 7.2 4.9 8.7 10.1 5.3 8.5 10.1 5.8 15.7 90.1 6.6 13.3 15.3 7.2 15.7 45.9 19.1 19.1 6.4 FB:FBgn0038007142393_at 122.9 99.7 77.7 187 214.5 179.6 71.7 144.9 130.8 100 168.4 153.7 162 190.4 197.5 87.5 127.6 65 114.7 137.7 122.9 72.4 197.3 26.1 FB;FBgn0039026153016_r_at 50.3 61.6 91.7 71.9 75.4 39.1 46.6 69.7 89.7 92.2 47.7 89.6 127.7 124.8 67.9 92.6 67.7 58.5 61.5 79.7 112.9 133.9 81.6 72.2 FB:FBgn0037269154378_at 277.1 297.5 349.3 288.5 318.2 288.9 267.2 340.9 273 383.1 264.2 275.9 234.9 335 268.8 276.4 359.3 260.5 256.4 254.4 211.5 323.9 347.5 175.3 FB:FBgn0031314141882_at 114.9 21.4 33.4 82.3 43.1 136.6 174.1 118.7 122.8 71.8 49.4 37.2 99.9 145.5 29.3 181.8 38.6 97 43.4 110.6 116 240.2 69.8 30.3 BDGP:GH14349.3prime-hit153450_at 571.5 707.9 533.9 327.4 371.8 210.6 474.3 635 728.8 662.3 592.5 481.8 330.6 343.3 373.4 340.5 710 755.8 424.8 233.2 464.6 412.2 473.4 434.9 FB:FBgn0030724146294_at 8.8 14.6 9.3 6.9 8.1 39.5 11.2 25.9 41.3 12.8 13.9 55.1 13.3 31.2 8.7 61.7 10.1 35.9 11.4 31.7 13.8 24.2 23.6 29.6 FB:FBgn0032441150195_at 309.5 268.4 302.2 227.8 104.2 161 92 297.7 349.6 223.4 146.3 154.2 151.4 175.6 190.6 296.8 416.8 343.8 302.9 217 270.8 199.2 121.9 147 FB;FBgn0038704153599_at 532.7 388 721.7 336.2 351.9 168.5 395.1 601.7 547.9 563.2 409.2 490.8 356.1 238.4 311.6 339.4 416.1 330.3 367.2 535.6 423.7 492 472.6 402.1 FB:FBgn0036305144788_at 27.3 13.7 58.4 52.2 42.4 63.2 52.2 24.4 44 13.2 51.6 9.1 21.7 12.7 40.8 10 24.9 18.6 56.6 11.7 16.6 89.3 9.8 38.2 FB:FBgn0030185145855_at 86.5 35.3 55.5 90 87.1 79.6 66.1 73.8 41.7 66.6 80.7 78.4 108.9 96.7 62.2 15 52.6 87.3 85.5 69.9 47.3 117.4 140.3 69.2 FB:FBgn0031750151851_at 344.1 372 616.6 514.1 653.2 518.4 364.4 284.8 387.2 1268 1194 1007 1202 1378.9 1227.8 1090 718.9 658.8 1062.2 1302.6 1080.1 748.4 729 732.9 FB:FBgn0027611143434_at 24.9 27.1 32.3 42.6 24.8 28.8 127 26.2 24 27.8 19.4 27 25.1 27.5 14.5 16.3 24.4 23.1 25.2 18 15 40.7 39.5 16.5 FB:FBgn0004173145247^at 4.7 5.4 7.2 49.6 7.8 22.3 11 11.9 7.4 4.5 5.4 23.4 7.4 17 4.3 27.2 12.3 6.5 6 7.4 15.1 12.5 12.9 13.9 FB:FBgn0030844145361_at 7.5 20.4 25.4 7.2 53.5 23.1 70 21.9 9.7 13.5 61.6 11.9 54.3 31.1 71.6 16.1 6.4 12 40.2 5.9 9.6 29.2 15.3 41.4 FB:FBgn0031005146031_at 9.5 14.5 9.7 12.9 7.8 7 2.8 5.7 7.4 1.7 25.2 34.5 8.1 43.4 3.9 16.7 7.9 9.7 16 1.1 28.3 46.7 44 17.5 FB:FBgn0032038147047_at 26.8 68.8 18 27 12.8 21.2 22.6 22.1 13.6 14.5 13.8 29.3 12.3 22.4 20.6 31 10.6 12.6 92.7 17.8 31.4 16.3 14,4 21.1 FB:FBgn0033667

Probe SetID
yw
hsi
yw
2yw
non hs3
ywshort hs1 yw short
h s2yw short
hs 3
ywlong hs1
ywlong hs2
ywlong hs
3
pKU
1pKU
2PK.B
3
pKbshort hs1
pKbshort hs2
pKbshort hs
3
pKblong hs1
pKblong hs2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
150184_at 21 43.8 10.8 24.1 50.3 70.6 100.4 29.1 13.6 57.2 11.3 20.8 47.8 52.4 81.8 50.2 16.3 60.3 33.4 23.7 17.3 95.9 30.5 66.7 FB:FBgn0038691151486_at 49.5 9.8 52.2 245.7 219.2 383.4 18.1 41.5 27.5 67.9 7.4 63.5 174 192.9 217.7 102 65.2 50.1 43.8 44.7 19.5 55.5 59.6 60.1 BDGP:GH04518.3prime-hit150499_at 247.7 223.6 139.2 278.1 173.7 215.7 218.6 176.2 238 118 194.5 199.9 151.2 131 102.7 36.7 200.1 75.4 329.3 456.6 457 186.3 224.7 326.5 FB FBgn0039178150762_at 436.4 406.4 470 415.7 269.2 265.7 184.7 232.6 187.2 401.3 465.2 426.9 115.8 267.4 105.5 106 284.4 170.2 652.7 389.3 801.4 226.7 319.6 608 FB FBgn0039567152389_at 21.3 27.4 19.6 20.4 13.1 20.3 20.5 28.1 15.4 64.8 28.8 59.1 18.3 20.3 46 26.2 18.2 26.1 19.7 16.7 35.8 30.3 15.4 25.7 FB FBgn0038208141509_at 170.4 163.6 193.9 205.1 171.8 201.2 180.6 256.1 374.6 172.3 191.5 164.9 189.8 187.5 149.3 253.3 237.1 185.3 134.8 163.2 229.6 252.7 231.5 172.5 FB FBgn0035145149188_at 5623 5454 5152 4966.6 4997.9 4273.6 5096.8 3540.4 3988.1 5596 7219 5544 6369.2 5987.1 4971 4992.4 3611.8 5623.4 5517.3 4107.4 6252.8 2179.3 2281.7 3556.1 FB FBgn0037097148987_at 4.7 7 30.5 7.2 6.7 8.2 10.1 7.2 12.5 24.9 7.3 10.1 31.9 15.5 2.2 4.8 9.4 11.3 3.9 7.4 49.1 7.9 13 2 FB FBgn0036778152411_at 245.7 339.7 273.4 311.6 291.1 343.2 585.9 392 533.2 349.6 351.2 264.1 426 475.6 380.5 542.8 565.5 711.7 347.1 310.4 410.9 318.5 445.6 640.2 FB FBgn0037439143545_at 45.9 51.6 23 66.8 2.4 49.2 24.1 18.7 3.5 11.1 1.3 27.4 22.3 66.4 35.7 31.1 7 12.9 10.7 31.5 12.9 12.5 27.9 33.1 FB FBgn0005563151268_i_at 19.4 101.5 10.1 67.8 58.9 5.5 28.2 3.2 12.4 34.2 13.1 52.7 50.5 56.1 38.6 132.3 40.5 67.3 112.8 5.9 16.5 38.1 3.7 48 FB FBgn0040798141931_at 368 246 331.8 262.2 277.2 174 325.3 208.1 211 436.7 252.1 332.3 276.4 336.7 248 364.8 239.3 235.8 204.2 471.3 219.9 320.7 324.7 204.8 BDGP:GM09283.3prime-hit146848_at 27.9 31.6 39.8 108.6 155.3 43.5 147.7 59.7 89.4 30.3 42.3 15.7 32 95.4 35.9 183.8 27.6 32 60.2 29.4 20.8 138.9 114.9 107.4 FB FBgn0033353154944_at 425.9 536.1 392.8 620.2 629.8 444.6 365.5 411.9 340.2 422.6 358.8 488.3 502.7 517.1 514.4 266.2 315.9 378.1 458.9 579.3 431.2 536.1 477.5 598.5 FB FBgn0030856149384_at 10.8 82.4 38.8 10.5 18.7 16.4 50.7 11.7 8.9 24.9 8.2 36.8 61.4 29.8 46 7.4 11.2 21 16.6 12.5 10.8 10.9 19 12.2 FB FBgn0037418149527_at 30 56.9 37.1 64 65.6 35.4 32.2 32.8 49.3 39.1 32.1 57.7 45 34.5 100 112.4 39.8 98.2 23.8 60 18.8 40.7 121.8 28 FB FBgn0037631145059_at 6.5 34.2 12.5 8.2 10.3 14 17.1 15 10.5 21.5 52.3 14.4 15.9 13.9 14 17.8 18.8 23.7 9.8 41.8 8.6 20.9 11.2 23.4 FB FBgn0030559153366_at 162.6 148.6 221.2 71.5 103.9 98 87.1 141.1 184 155 120.6 122.2 55 72.1 73.7 30.3 141.6 164.3 182.2 135.4 208.4 107.2 105.1 168.7 FB FBgn0036316148893_at 93.9 27.7 142.3 278.4 293.8 316 201.7 210.6 152 134.3 50.2 40.8 363.1 313 492.6 198.1 168.7 199.9 41.8 67.2 46.5 97.9 83.8 67.4 FB FBgn0036626153409_at 7730 6900 7080 12155 12021.2 11990.2 9366.7 8435.4 9662.1 6913 9081 9217 11929.9 16099.5 13325.8 10818 8133.4 9462.1 7829.5 9478.4 7556.9 10538.2 11925.6 10390.2 FB FBgn0004587149450_at 96.1 67.4 108.2 94.1 86.4 59.8 71.2 33.9 109.8 101.7 93.7 89.1 57.2 81.4 110.9 22.3 112.4 36.3 114.2 64.2 27.5 197.1 69.1 80.4 FB FBgn0037507146977_at 111.9 187.8 65.5 218.2 136.8 268.3 143.5 74.4 133.5 58.1 86 104.8 160.1 149 234.5 219.5 123.5 86.4 159.5 241.2 213.8 236.3 174.7 187 FB FBgn0033575146291_at 14 15.7 72.6 14.4 12.3 9.9 51.7 7.9 4.2 17 8.9 31.6 13.8 26.4 25 8.9 9 28.3 11 8.3 16.7 23 99.9 10 FB FBgn0032438141949_at 66.8 48.8 20.1 25.8 9.3 12.8 13.6 66.1 47.3 16.2 60.9 71 44.8 74.4 13.3 83.9 12.3 117.6 30.2 94.3 23.1 13.6 61 92.6 BDGP:LD04933.3prime-hit153367_at 657.9 669.4 941.8 412.6 563.9 363.4 496.4 455.3 230.3 903.5 483.2 554.9 468 459.6 469.6 379.2 332.5 244.8 964.2 619.3 860.4 423.3 280.4 489.5 FB:FBgn0037977143214_at 261.5 206.7 234.1 274.5 281.8 206.1 203.3 261.2 281.7 353 233.9 253.1 490.5 463.9 661.8 468.5 486.2 258.8 414.5 274.4 402 463.8 603.2 458.3 FB:FBgn0001965144268_at 6.4 3 6.1 16.4 48.7 4.6 5.7 15.9 2.4 5.2 16 8 6.3 72.9 6.1 3.3 10.4 20 20.8 22.9 7.5 15.2 4 8.3 FB:FBgn0028857150978_at 12.2 17 18.9 23.8 19.1 15.1 14 17.9 14.9 18.8 17.9 20.8 19.1 22.5 23 28.6 13.2 31.4 13.6 11.9 10.6 22.8 17.9 14.5 FB;FBgn0039906152921_at 939.4 858.4 848.1 1150.8 1193 1184.3 678.7 1032.3 1106.6 773.9 1098 1108 1619.2 1795 1850.7 635.2 829.2 781.4 832.3 880.7 800.7 889.8 1193.7 912.9 FB:FBgn0039581143447_at 219.3 172.8 64.2 152.3 184.5 136.3 38.7 165.6 161.9 89.4 123.9 102.3 159 159.2 203.9 76.1 104.5 177.7 119.6 130.3 110.6 113.9 98.3 55.4 FB:FBgn0004367154935_at 986 708.3 751.8 793.6 911.4 929.2 1405.8 872.1 1128.8 705.8 903.4 729 641.6 638.8 878.2 878.7 903 1159.9 947.6 735.2 804 836.1 859.6 1026.8 FB:FBgn0034727145485_at 196.4 188.4 176.4 143.9 106.8 140.3 103.5 269.1 205.9 128.3 181 213.7 111.4 62.2 114.2 113.5 188.4 378.3 151.1 126.9 119.8 188.7 117 125.6 FB:FBgn0031188150718_at 227.6 273.9 163.4 169.5 328.3 538.3 255.6 162.4 141.9 96.8 99.5 220.5 175.6 289.2 372.6 88 207.2 159.4 295.9 241.7 163.3 247.5 307.7 210.8 FB;FBgn0039497149226_at 42.3 93.8 57.2 40.9 38.9 53.2 76.4 16.1 19.9 152 117.8 78 110.7 117.6 24.6 100.3 69.9 12.8 77.9 63.6 34.8 56.2 32.1 44.6 FB:FBgn0037157148205_at 6108 4095 2095 2006.1 1965.5 1870 3521.1 1414.9 1486.2 2016 2731 7040 1200.7 2086.1 1571.1 1785.3 1173 1413.5 7430.6 7535 11124.4 2311.4 1699.5 3336.2 FB:FBgn0035582144082_at 305.2 398.6 223.2 273 306.3 343.3 186.2 493.9 357.3 103.3 263.9 210.3 245.7 199.4 208.1 190 460.3 251.8 323.4 247 242.3 187 307.8 254.7 FB:FBgn0025641150290_at 86.6 28.7 8.9 20.5 16.9 17.6 20 14.9 3.6 11.8 10.4 11.8 9.3 21.3 10 17.3 4.3 14 9.5 15.1 5.1 19.8 9.4 17 FB:FBgn0038847144333_at 54.9 37.3 40.2 33.9 59.7 34.3 65.8 8.9 27.6 113.1 64.8 98.4 56.2 26.3 65.7 46.1 45.9 100.7 45.7 48.4 35 72.5 84.3 40.1 FB:FBgn0028961152344_at 446.2 349.7 383.4 565 428.7 433.9 309 301.8 378.1 411.3 306.1 436.6 452.1 442.3 432.3 311.2 326.5 434.9 324.4 307 279.8 634.3 437.4 372 FB;FBgn0027795144178_at 195.2 183.8 103.5 219.6 170.8 283.7 179.7 99 87.7 41.5 178.6 254.8 241.5 298.6 205.4 190.5 37.1 139.6 99.1 143.3 100.4 175.2 216.6 160.3 FB:FBgn0027358141794_at 122.1 154.7 138.9 133.2 130.6 162.7 95.4 110.6 196.9 135.5 115.3 116.1 206 119.9 160.1 184.6 129.5 192.7 120.2 143 190.1 136.7 142.3 113.9 FB:FBgn0036021145490_at 23 36 18 6.6 10.5 11.8 13.4 6.8 4.1 9.1 7.7 13.8 28.6 11.6 14.6 10.6 8.2 4.9 21.7 15 14.7 79.2 11.6 8.5 FB:FBgn0031196147965_at 22.9 16.4 20 9 89.4 9.4 14.2 12.1 12.8 60.5 79.9 17 64.7 98.1 27.4 83.7 75.2 91.6 89 14.6 38.7 168.8 88.4 116.3 FB:FBgn0035196149232_at 222.5 166.2 106.7 38.2 198.4 150.8 53.3 57.9 96.6 185.3 223 194.9 117 96.5 109.4 150.3 83.7 112.3 201.9 156.5 204.7 203.5 150.1 167.1 FB:FBgn0037165154128_at 259.3 253.5 313.9 283.5 342.4 383.6 209.7 241.3 300.7 187.5 229.7 262.5 333.7 382.9 323.5 231.2 239.7 229.4 417.2 281.3 386.6 184.4 332 356.7 FB:FBgn0012051141314_at 1669 1027 461.1 125.2 126.4 164.5 315.6 196.4 160 229.3 158 750.3 58.2 120.9 89.8 151.2 190.1 197.8 1957.2 656.4 1718.4 126.3 163.5 943.5 FB;FBgn0020638142745_at 74.7 135.7 112.2 128.5 75.4 169.6 124.1 77.8 146.1 129.7 133.5 94.5 103.5 105.1 165.7 204.2 146.9 107.5 148 103 93 212.8 214.7 118 FB:FBgn0002905150790_at 521.1 438.7 64.3 282.2 409.3 472.7 511.3 460.2 470.6 172.6 351.5 519.3 450.1 413.3 386.6 274.5 516.2 388.1 412.2 277 459.7 382.6 475.2 93.9 FB;FBgn0039610143141_at 220.4 224.7 348.9 380.9 178.6 145.6 173.5 220.5 248.6 380.6 213.2 281 258.1 389 339.8 245.4 303.5 244.8 211.3 260.1 216.5 292.6 329 232 FB:FBgn0000552145581_at 630.2 762.3 487.9 2037 1918.6 1912.3 479.5 476.8 665.4 966.8 1018 1054 2224.4 2215.4 2571.6 638.3 914.7 586.1 1145.6 1444.4 661.7 2152.2 2242.9 666.1 FB:FBgn0031338
144306_at 1534 1299 944.3 694.7 867.9 741.5 1249.5 491.5 360 599 705.5 1788 269.5 455.7 522.4 812.5 500 495.5 1431.7 911.8 1186.9 672.7 532.5 1312 FB:FBgn0028915142297_at 14.9 88.6 113.3 38.4 21.7 50.5 104.3 42.9 55.5 44.1 98.3 118.7 101.1 38.1 79.5 64.4 75.7 67.1 47.5 18.4 47.2 96 81.3 67.6 FB:FBgn0033494
143737_at 49.5 79 40 9.6 45.7 27 56.1 30.8 4.5 8.7 37.7 73.7 3.0.9 71 7.3 47.1 8.7 28.3 90.4 56.5 68.9 2.8 24.8 38.4 FB;FBgn0013995
146287_at 31.8 46.3 29.7 83.1 57.8 77 14.9 18.6 12 16.5 32.7 10.9 31.2 6.6 19.1 30.5 31 13.8 4.6 5.9 19.6 52.3 46.1 15.1 FB:FBgn0032434
153115_at 514.9 452.2 188.7 315.4 479.1 298.1 290 218.4 405.9 210.9 472 458.7 385 334.5 261.9 299.6 444.4 507.2 475.1 307.1 357.3 253.4 382.6 432.9 FB;FBgn0037891

Probe SetID
ywnonhs1
ywnon hs
2
ywnon hs
3
ywshort hs
1yw short
hs 2yw short
hs 3
ywlong hs
1
ywlong hs
2
ywlong hs
3
pKb1
pKbnon hs
2
pKbnon hs
3
pKbshort hs
1
pitb short hs
2
p itb short hs
3
ph.blong hs
1
pKblong hs
2
pKblong hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
144464_at 376,5 232.4 268.3 231 414.9 403.3 287 309.3 187.7 206.5 251.5 273 325.8 355.5 358.1 252.4 255.8 123.5 328.8 211.7 249.8 336.4 434.8 149.9 FB:FBgn0029693149253_at 50.1 54.6 63.9 5.4 23.8 87.3 81.1 45.3 60.5 94.4 60.6 35.9 48.5 40.9 53.9 44 83.1 94.1 61.9 42.5 69.4 84.1 82.3 77.7 FB:FBgn0037190146179_at 50 73 18 83.3 86.4 51.7 25.5 37.8 19.5 10.2 95.9 30 121.3 48.2 26.6 73.1 31.3 19.6 17.8 98.9 32.1 41.2 135 37.8 FB:FBgn0032280147152_at 115.5 138.9 80.1 128.5 155.5 87.9 121.6 123.8 68.2 90.8 90.6 106.4 116.5 178.4 127.6 91.5 101.8 73.8 54.7 110.6 51.9 255.3 96.3 63.7 FB:FBgn0033838148404_at 630.1 523.4 609.3 855.6 878.8 1014.5 868.6 485 783.9 663.5 748.9 758 902.8 974.9 1076.2 695.7 503.4 628.5 515 525.3 603.6 893.5 960.5 633.1 FB:FBgn0035898150063_at 74.3 59.1 36 86.2 56.7 103.9 108.8 95 78.5 105.4 26.7 77.8 60.7 93.3 19.1 53 56.5 112.9 116.6 54 42.3 31.3 141.1 52.3 FB:FBgn0038495151589_s_at 10.3 87.7 65 68.9 39.6 41.6 71.1 26.3 28.6 41.3 44 78.2 32.4 42.5 51.1 64.4 23.7 47.2 44.5 37.2 16.7 44.2 58.4 31 BDGP:GM09007.3prime-hit154054_at 197.2 291.2 260 198.6 127.9 96.2 201.8 228.6 203.4 166.7 172.2 241.3 142.6 236.8 59.6 118 317.8 267.1 156 251.8 165.8 160.3 148.6 200.8 FB:FBgn0037638145280_at 12.1 65.9 18.8 15.6 62.2 34.1 39.4 60.7 16.1 29.2 18.9 57.7 20.3 26.1 28.9 19.8 21.4 75 13.8 13.6 19.8 18.3 60.7 20.9 FB:FBgn0030898149906J_ a t 167.6 95.5 50 103.4 123.3 95.8 304.5 110.7 102 233.8 338.7 341.1 818.8 778.6 785.1 912.6 300.9 398.5 122.4 117.8 127.5 15.4 96 109.6 FB:FBgn0038243141800_at 292.8 191 392.3 380.6 414.5 285.7 367.5 230.8 176.2 297.2 379.2 416.4 485.3 474.6 540.8 510 251.9 300.6 246.2 221 254.9 271.8 351.1 322.9 FB:FBgn0035149144309_at 612.4 373.4 529.2 657.2 770.4 694.2 862.9 602.1 588.9 533.6 474.9 566.8 759.6 709.6 862.6 798.6 606.7 664.1 389 6 403.9 359.2 413 595.3 302.5 FB:FBgn0028919152132_at 157.5 92.8 86.2 276.4 175.4 228.8 116 154.7 221.2 78.9 43.6 82 108 117.5 136.9 106.3 66.4 56.8 50.9 23.5 46.1 197.9 219 134.7 FB:FBgn0035403148652_at 11.2 65.4 11.4 13.5 13.7 10.5 10.9 17 6.1 9.3 33 26.6 10.2 27.1 46.2 8.2 36 40.8 29.6 7.8 7.2 10.3 87.9 13.3 FB:FBgn0036276150059_at 242.4 144.9 205 255.2 188 97.8 262.1 353.2 216 234.7 152.8 219.6 228.7 123.9 183.3 328.6 212.8 219.4 139.6 226.1 203.9 59.3 246.8 204.5 FB:FBgn0038491145318_at 41 19.9 18.2 24.4 25.1 21.8 25.2 13 13.5 19.3 29.8 81.3 17.2 17 4.9 23 7.9 7.1 15 6.8 43.3 20.2 18.2 21.7 FB:FBgn0030947147281_at 5.4 5.2 30.9 8.3 6.9 15 6.2 16 17.6 15.8 11.4 15.3 21.7 4.8 28.9 10.8 8.4 24.4 2.4 1.8 10.4 4 6.3 3.8 FB:FBgn0034077141250_at 361.7 414 500.9 285.6 315.6 320.4 206.6 498.9 530.9 363.7 293.3 319.9 268 302.1 248 295 363.2 382.6 331.1 378.2 421.8 292.9 337.7 357.8 FB:FBgn0036850143388_at 80 123.8 74.8 248.9 95.6 269.3 69.8 95.1 51.7 196.3 88.6 75.1 3415.7 3712.6 4989.2 330.8 1117.3 1068.5 109.9 173.1 61.2 7173.4 6653.6 1240.8 FB:FBgn0003884147324_at 86.3 171.1 129.4 210.7 186.4 154.1 60.4 74.8 115.5 98.6 76.6 70 114.8 199.2 124.9 24.8 94.4 80.3 148.7 162.9 67.4 162.2 139.3 101.2 FB:FBgn0034149141741_at 122.9 76.5 176.8 130.4 96.7 20.7 12.5 65.9 76.8 96.9 93.7 54 67.4 118 97.8 200.5 121.5 111.3 38.1 67.4 93.6 103 97.1 101.2 FB:FBgn0031490151879_at 186.1 201.7 201.2 250 294.2 273.7 235.1 326.3 274.4 178.3 217.9 224.9 340 470.8 301.9 275.8 320.1 576.4 176.5 199 273.7 324.8 278.1 217.1 FB:FBgn0027602152820_at 311.7 294.8 461.1 230.5 231.8 295.8 406.7 373.9 403.3 451.4 401 432.2 242.6 216.8 253.1 421.5 314.6 516.5 208.1 226.9 290.2 262.3 298.6 200.2 FB:FBgn0030802147642_at 2.5 12.7 6.8 4.6 3.4 8.5 8.1 4 40.1 4.2 5.5 13.1 12 4.6 10.5 11.1 4.8 7.8 3.2 7 8.1 16.6 8.1 4.6 FB:FBgn0034670153780_at 66.7 77.4 150.3 66.7 119.8 81.2 147 86.3 140.1 82.1 97 91.4 55.7 89.8 44.2 193.9 72.5 124.2 47 38.5 90.2 38.5 48.9 95.7 FB:FBgn0033206146381_at 9.3 32.2 35.5 47.4 8.9 28.4 7.9 13.6 42.9 17.2 53.3 13.2 32.3 52 64.7 8.1 8.5 7.1 55.4 26.6 29.8 8.5 5.1 9.1 FB:FBgn0032608154049_at 160.4 191.2 90.1 158.4 158.8 134.3 127.7 62.4 73.4 101 223.4 217.1 106 103.7 96.7 145.6 91.9 111.5 182.4 140.1 159.8 149.2 123.8 172.7 FB:FBgn0036444142382_at 75.6 118.5 279 100 142 125.3 75.7 95.2 43.1 182.3 69.8 131.6 74.7 5.5 116 115.2 46.4 42.5 115.6 99.2 73 56.4 97.6 90.9 FB;FBgn0032936144014_at 44.7 80.1 91.9 49.3 111.4 72.7 45.8 61.3 68.1 103.4 75.4 87.9 57.9 101 45.4 38 67 63.1 56.3 77.8 81.2 154.2 96 38.6 FB;FBgn0024249146981_at 282.7 215.2 113 233.7 262.9 275.3 98.5 132.7 318.3 283.3 191.6 259.7 251.5 340 258.3 362.5 347.7 200.5 225.3 292.7 210.8 285.7 254.5 180.1 FB:FBgn0033580150206_at 15 10.2 38.4 12.1 15.7 10.1 31.1 14.4 24.3 22.6 119.8 7.1 17 21.5 132.9 18.2 17.7 40.3 52.1 12.9 11.1 46.1 51.8 14.2 FB:FBgn0038718151609_at 282.5 420.8 335.6 268 292 243.2 338.5 285.6 590 215.1 199.2 316.2 288.8 301.2 260.5 287.6 336 259.6 355.6 379 385.5 436.1 416.8 577.4 BDGP:LD01876.3prime-hit143686_at 8.9 12.6 16.1 15.8 11.8 5.2 13 34 8.4 26.5 4.9 19.6 3.3 11.8 51.8 23.7 24.7 7.1 4.5 3.8 12.9 75 18.2 9.5 FB:FBgn0011743142381_at 367.5 380.5 667.9 430.2 496.5 434.2 644.4 462.8 395.1 847 445.3 587.8 505.5 592.3 622.1 624.9 662.3 462.3 592.4 516.8 699.6 506.3 429.3 495.2 FB:FBgn0032465146136_at 20.4 37.8 11.1 5.8 5 6.5 43.4 8.4 19.9 4 10.5 48.1 2.9 2.2 27.1 7.4 2.8 27.7 12.2 17.8 20.7 5.8 2 2.5 FB:FBgn0032209149029_at 7.7 11.5 11.3 16.3 10.8 43.4 41.1 19.9 15.9 13.7 9.2 13.9 8.9 12.6 28.6 12.9 12.6 23.2 12.5 10.7 10.1 27.2 14.2 7.7 FB:FBgn0036843151149_at 13.5 22.1 46.2 15.6 16.4 10.5 11.9 18.5 17.6 26.8 5.6 24.1 22.9 65.8 31.4 16 16.2 8.2 20 10.9 18.5 35.7 30 9.6 FB;FBgn0040669151247_at 32.4 18.4 180.8 42.9 74.1 49.7 38.3 70.9 67.2 244.3 12.2 97.9 86.5 70.7 75.3 166.2 52.9 113.8 41.9 38.2 53 172.8 68.6 47 FB;FBgn0040772153974_at 262.4 196.7 164.8 187.7 107.8 189.9 266.8 188.9 255 187.6 187.7 182.2 367.9 195.1 251.6 304.5 253 220.8 145.8 155.1 193.1 114.3 189.5 173.4 FB:FBgn0034109147393_at 93.8 96.5 144 88.6 103.9 82.1 69.8 128.5 159.2 108.8 129.2 70.6 35.3 119.1 148 86.7 185.9 139.7 118.5 81 96 149.6 212.2 141.9 FB:FBgn0034265154666_at 227.1 236.8 324.5 233.1 204.3 251.9 112.9 195.2 238 427 435.4 341.2 465.5 442 330.5 244.9 329.2 332.2 300.7 173.1 286.7 428 502 170.8 FB:FBgn0031540146548_at 203.6 223 242.6 207.4 225.8 242.4 158.3 194.1 211.8 205.8 163.8 303.5 165.3 234.5 178.4 131 210.5 169.5 253.5 265.1 162.1 199.8 263.2 205.8 FB:FBgn0032874153584_at 675.5 457.7 764.7 482.6 510.3 556.3 468.8 649.9 561.8 899.5 901.8 800.4 656.4 604 519.9 506.9 946.6 985.8 619.6 619.7 701.6 698.2 578.9 501 FB:FBgn0027894154410_at 523.2 666.7 525.7 299.2 242.1 277.6 496 526.7 433 585.7 396.4 595.2 308.8 255.8 324.5 485.6 677 401.1 764.5 627.8 713.4 346.6 370.7 770.4 FB:FBgn0035147141249_at 197.3 178.3 132.2 211.4 187.2 142.4 237.4 242.7 281.3 218.1 136.1 190.2 263.8 276.3 342.7 321.7 323.7 296.5 182.9 195.9 171 173 249.9 205.4 FB:FBgn0033740148612_at 23.5 43.5 43.1 60.8 41.6 63.4 16.2 52.4 32.9 69.7 9.8 35.6 12.8 61.2 77 57.6 30.3 42.5 34.1 54.2 23 47.6 11.8 84.3 FB:FBgn0036217153130_at 136.7 192.9 132.9 137.4 139.7 174.3 140.4 260.7 243.6 112.7 88.2 114.1 158.3 123.7 141.5 133.3 185.3 185.9 86.6 111.7 141 164.8 103.5 137.1 FB:FBgn0030744147136_at 221.6 176.3 262.8 208.7 201.1 217.9 483.8 276.2 173.4 149.1 191.9 188.6 217.8 180.9 149.2 258.1 198.8 175.7 182.5 231.7 164.9 294.5 227.8 215.2 FB:FBgn0033815145632_af 22.9 14 34.2 15.8 14.1 33.9 15.1 14.1 18.5 21.1 12.4 14.8 15.2 14.3 12.9 19 12.8 28.8 9.7 10.6 10.8 21.1 24.3 9.3 FB:FBgn0031411151282_at 78.4 85.5 92.8 82.1 60 116.1 103 81.2 80.2 144.1 130.8 132.2 141.2 149.1 135.4 69.6 97.9 112.5 43.8 83.5 71.1 140.4 87.3 89.1 FB:FBgn0040807141867_at 52 42.2 133 6 69 38 48.6 119.4 31.3 29.5 67.3 32.7 31.1 30.7 59.2 44.3 62.7 25.6 83.2 31.9 60 24.1 103.5 30.2 38.4 BDGPGH10642.3prime-hit148345_at 17.4 41.5 3.7 9.5 5.9 10.3 5.2 5.9 5.3 12.1 19.7 7.2 8.9 16.3 5.9 15.6 5.2 6 2 5.3 5.3 11.9 12.2 7.7 FB:FBgn0035796149306_at 58.6 156J1 47.-1 21.9 88.3 45.6 23.1 92.2 108 1 17.7 124 4 22.1 21.7 190 27.3 33.2 39.1 33.4 11.7 16.8 15.5 36.7 99.4 104.9 FB:FBgn0037291149306_at 58.6 156.3 47.4 21.9 88.3 45.6 23.1 92.2 108.1 17.7 124.4 22.1 21.7 190 27.3 33.2 39.1 33.4 11.7 16.8 15.5 36.7 99.4 104.9 FB:FBgn0037291154577_at 213.1 277,7 344.4 228.9 246.3 144.8 238.4 353.8 394.5 207.9 231 211.5 187.1 152.2 248.3 292.9 452.5 337 303.2 364.8 238.6 317.9 265.6 291.1 FB:FBgn0031573143007_at 662.4 745.3 681.9 750.1 802.6 681.7 980.1 884.5 658 605.2 979.5 527.2 906.8 715.2 724.1 601.4 607.8 539.2 596.7 662.1 545.2 745 4 595.2 775.7 FB:FBgn0001961

yw yw yw yw yw yw yw pKb pKb pKb pKb pKb pKb pKb pKbProbe Set non hs non hs short hs yw short yw short long hs long hs long hs non hs non hs non hs short hs short hs short hs long hs long hs long hs PKBYW PKBYW PKBYW PKBYW PKBYW PKBYW
ID hsi 2 3 1 hs2 hs 3 1 2 3 1 2 3 1 2 3 1 2 3 non hs 1 non hs 2 non hs 3 long hs 1 long hs 2 long hs 3 Flybase ID149025_at 140 407.2 266.7 147.6 222.1 42.1 37.3 38.8 315.5 212.7 184.2 50.1 45.4 64.3 46.7 40.7 161.7 288.3 141.5 172.3 231.1 310.6 250.5 218.6 FB FBgn0036835148264_at 18.9 75.3 16.6 20.7 17.9 35.8 63.3 15 17.1 14.4 24.3 21.4 96.8 57.8 26.4 12.1 43.4 95.3 17.8 57.7 12.2 31.7 16.2 35.5 FB FBgn0035678155038_at 585.6 432.5 294.3 311.9 229.5 194.8 144.4 631.2 558.5 348.8 360.4 327.1 233.5 279.1 216.6 336.9 406 376.8 302.8 293.9 266 224.1 340.4 281.5 FB FBgn0025549148805_at 70.1 134.4 145 116.1 163.5 100 131.5 207.5 180.7 276.6 150 128.2 94.4 272.5 157.1 30.9 236.9 163.9 173.1 185.7 158.9 181.1 342.8 173.2 FB FBgn0036502145482_at 49.4 47.4 60 59.4 66.6 55.6 72.2 82.6 45.3 45.5 81.4 57.3 38.2 63.8 122.2 18.3 36.9 10.2 47.1 67.8 56.8 128.6 84.5 9.8 FB FBgn0031184151945_at 25.7 67.2 106.2 83.1 65.5 48.7 84.2 66.3 6.8 104.3 33 76.9 34.7 62.3 38.4 112.7 55.7 34.8 24.1 26.4 5.5 23 62.9 8.7 FB FBgn0038171141623_at 292.6 170.7 223.5 307.2 309.3 371.8 152.9 186.8 144.1 321.9 246.3 162.2 202.4 273.2 285.1 173.1 186.7 398.1 171.8 324.6 382.3 270.8 410.5 215.3 FB FBgn0011285153401_at 161.3 129.4 19.2 49.8 19.5 25.1 140.4 87.2 12.3 58.4 96 25.3 9.2 12.1 46.9 22 16.6 17.6 69.4 24.3 88.3 125.7 19 18.1 FB FBgn0031940147117_at 28.4 17.1 50 15.8 25.1 60.3 18 82.9 121.2 27.2 24.7 26.8 11.9 14.6 18 43.5 31.6 25.9 8.8 9.9 27.6 14.5 10.8 12.7 FB FBgn0033773149974_at 159.5 130.6 292.2 134.2 136.1 102 98.1 162.4 83.3 225.1 157.5 160.6 129.5 146.7 46.9 249.6 138.1 207.5 183.6 124.1 110.4 99.7 80.4 29.9 FB FBgn0038363143195_at 9.2 15.2 8.2 56.1 6.4 28.4 68.1 15.2 32.9 24.9 8.8 6.8 58.9 110.9 132.2 64.2 9.6 53 5.5 5.1 6.6 121.1 106.2 52.2 FB FBgn0001227152530_at 281.3 209 295.2 336.4 368.5 358.6 218.1 325.1 275.4 289.9 310.2 252.4 487.7 338.2 399.9 380.9 239.4 232.1 268.1 271.6 192.2 355.9 483.7 267.8 FB FBgn0015270142308_at 10 21.2 10.2 9.4 9.1 18.9 9.2 11.9 6.8 9.9 12.2 12.7 12.5 20 15.6 20.6 12.4 17.5 8.3 8.3 15.5 17.1 14.7 16.7 FB FBgn0032073142221_at 2.5 32.6 38.5 9.4 11 8.5 4.2 8 7.3 10.5 19.2 8.9 5.4 34.8 9.2 13.6 5.7 3.5 6.2 33.3 5.1 12.6 31.9 32.5 FB FBgn0015038144026_at 149.1 177.7 239.4 198.8 179.2 188.6 281.7 227.8 191.6 158.7 128.5 156.4 43.7 108.4 141.9 198.6 129.6 259.5 130.9 80.5 231.5 59.6 98.6 144.4 FB FBgn0024841147607_at 185.3 148.9 101.1 192.8 184.4 167.7 123.4 245.7 172.8 141.6 136.8 120.4 104.1 216.2 167.1 76.8 207.1 141.1 161 157.7 107 158.2 160.6 125 FB FBgn0034603149187_at 2166 2338 1985 1135.3 1327.8 1239.4 2124.7 2264.6 2173.9 1373 1797 1505 798.6 820.2 775.5 1964.7 2113 2018.9 2233.7 1572.4 2500.9 873.1 1031 2539.7 FB FBgn0037096152120_at 448.7 544.7 382.2 433.9 388.3 350.2 563.6 498.2 378.1 969.3 803.8 964.3 608.1 566.7 747.3 978.1 700.7 543.6 856 1340.3 1412.9 409.3 597.5 1418.7 FB FBgn0038194148148_at 8.4 13.5 25 18.1 29.9 6.8 9.2 4.2 19.9 37.2 29.2 5.3 6.1 26.2 18.7 6.4 8.5 14.8 43 52 12.5 31.6 20 17.5 FB FBgn0035503141815_at 691.4 509.8 598.6 491.5 405.9 557.9 676.1 692.5 643.1 271.1 680.9 485.2 241 282.8 257.3 479.6 352.2 468.1 296.9 290.3 234.7 358.6 303.3 343.4 FB FBgn0035438144688_at 87.8 49.8 95 99.9 63.3 45.4 117.4 80.5 41.7 222 48.6 39.1 14.8 72.7 97 40.9 42 42.9 23.2 39.2 50.3 58.7 103.7 52.8 FB FBgn0030019146203_at 34.3 52 54.1 23.3 23.6 28.9 70.4 30.6 66.9 36.2 15.2 16.1 18 54.7 28 37.7 14.5 34.8 18.1 25.4 34.4 29 23.9 19.4 FB FBgn0032311147895_at 28 71.2 150.8 103.8 17.1 12.8 12.1 42.7 55.3 186.2 59.5 21.4 17.1 104.1 16.1 17.4 15.2 23.3 48.2 7.5 41.1 66.2 18.7 30.3 FB FBgn0035071154470_at 355.2 229.4 235.6 332.9 292.2 338.4 232.1 195 222.8 270.4 293.8 512.8 366.6 404.3 409.5 163.1 359.3 201.2 281 244.3 312.8 245.7 394.9 228 FB FBgn0033859141875_at 16 65.6 36.2 38.6 80.8 63.9 67.7 40.1 3.6 46.5 24.9 67.1 28.2 42.4 84.5 47.2 33.9 45.2 34.2 88.1 45.9 41.1 53.5 76.7 BDGPrGH 12726.3prime-hit151130_at 7.2 3.2 8 3.1 2.4 12.1 9.4 2.3 14.4 7.1 17.3 3.3 2.7 7.9 11.4 3.4 30.6 5.3 3 1.9 6.6 3.9 21.5 2.3 FB:FBgn0040646146832_at 276.1 191.4 179.1 131.1 147.8 148.5 340 228.5 126.8 69.2 579.9 63.2 117.2 165 43.7 121.7 192.9 99.8 205.1 400.1 360.5 242.2 290.3 277.2 FB:FBgn0033328154465_at 72 113.4 109.2 101.6 41.9 18.5 72.5 116.1 114.4 51.2 81.2 86.8 65.6 71.5 45.5 91.6 62.9 105.9 68.4 78 77.2 99.4 42.4 66.6 FB:FBgn0038268144023_at 60.6 81.4 145.6 68.5 99.8 59.2 10.4 136.2 120.7 79 73.7 51.5 59.8 76.1 62.7 72.6 105.3 150.4 35.6 42.3 55.4 11.9 32.2 51.6 FB:FBgn0024732151961_at 2.7 43.8 13.6 41.9 4.6 50.7 36.1 10 22.3 6.7 5.4 2.5 16 2.9 31.4 35.1 3.3 36.6 1.8 4.4 59.6 10.2 26.6 3.6 FB:FBgn0031327150227_at 14.5 66.3 36.7 10.5 62.3 33.3 8.3 11.3 31 59.9 25.1 8.7 39.1 34.9 58 34.1 10.9 18.7 9.8 78.4 39.5 23.9 63.8 100.7 FB:FBgn0038744149944_at 29.6 50.1 60.4 118.5 142.4 163.6 21.2 109.2 97.8 47.2 56.4 14.3 145 53.2 120.9 39.6 124.9 200 55.2 126.5 39.9 174.9 205.8 89.3 FB:FBgn0038310154050_at 248.6 237 336 390 350.7 373.5 278 317.1 341.2 450.1 228.2 191.8 320 452.1 328 279.9 300.3 365.8 226 195.7 276.6 428.1 340.4 327.4 FB:FBgn0027518154085_at 1397 1058 1288 1428.3 1516.7 1499.7 1262 1462.3 1161.3 1360 1575 1193 1958.5 1973.3 1669.6 1773.7 1619.9 1459.4 919.3 906.5 988 851.1 1150.5 863.4 FB:FBgn0000566150565_at 472.3 237.8 194.8 130.3 20.4 68.2 1161.9 249 175.8 131 136 234.5 56.7 117.6 92.9 164.4 133.8 149.2 607.8 679.1 1225.5 203.4 150.9 610.8 FB:FBgn0039297153258_at 125.3 23.9 80.6 68.3 97.4 18.1 135 117.2 176.4 28.9 44.9 36.6 75.8 61.2 39.3 73.8 241.1 201.5 114.5 81.8 147.9 31.7 100.8 105.7 FB:FBgn0037999143670_at 296.7 515 408.3 168.4 188.4 221.9 148.9 423.5 532.3 332.6 252.6 251.3 230.5 169.1 170.1 238.1 517 561.5 297.2 314.1 373.4 249.1 153.6 257.6 FB:FBgn0011648154923_at 1008 744.1 1004 1265.4 1260.9 1239.8 1152 1052.9 933 1010 1154 1025 1551.3 1489.8 1451.4 1833.3 878.6 861.5 846.9 805.6 775.5 664.4 725.7 618.7 FB:FBgn0034646143883_at 80.8 10.2 70.4 59.9 36.4 42.4 34.5 6.2 80.2 13.4 50.6 9.9 13.2 84.6 17 14.6 20.6 25.7 36.5 60.1 39.8 23.7 15 12.9 FB:FBgn0016920145382_at 22.2 17.8 53 17.2 11.4 8.2 15.1 68.2 68.8 23.2 33.9 5.7 17.4 17.1 16.8 52.4 13 19.3 58.3 16.5 62 69 57.4 8.9 FB:FBgn0031045153198_at 320.1 250.8 207.3 382 181.4 311.6 570.5 263 290.3 372.9 417.2 343.5 345.3 286.1 216.3 502.8 439.1 309.9 347.2 266.1 260.8 280 214 362.3 FB:FBgn0031708155114_at 557.1 838.8 797.9 776.7 980.7 1024.3 677.3 734.3 679.4 1077 930.8 665.3 1109 1111.9 1037.8 762.4 817.1 598.3 834.3 905.5 779.8 870.5 922.7 758 FB;FBgn0037362144461_at 87.7 138.6 25 15.7 21.1 16.5 27.4 16.9 32.5 34.9 26.5 100.8 9.9 82.5 8.6 48.5 122.3 27.9 163.9 17.9 18.7 13.3 26.7 156.8 FB:FBgn0029690143546_at 217.2 193.6 168.9 127.9 174.7 202.3 190.1 204.1 161.8 143.7 155.9 132.3 79.1 116.7 112.2 82 102 91.1 135.9 146.5 113.3 138.1 157.3 155.4 FB:FBgn0005564155093_at 17.4 89.6 13.9 9.7 56 65.6 53 90.5 58 31 33.5 85.3 9 6.6 26.5 47.5 12.4 46.4 85.8 3.7 48.4 74.9 50.7 66 FB:FBgn0030414144938_at 298.8 248.6 191.3 407.7 325.3 481.8 195.3 250.9 293.4 187.8 309.1 206.7 320.7 428.8 293.9 242.4 307 290 268.4 214.5 160.3 401.8 310.1 245.2 FB:FBgn0030391149938_at 192.4 75.8 37.1 16.4 16.5 14.4 190.1 73.7 21.3 22.8 50.8 156.9 16.7 21.3 27.8 81.5 26.5 88.5 145.5 28.5 123.5 42.1 33.7 193.6 FB:FBgn0038291147120_at 61.2 10.6 14.4 53.1 7.5 69.4 13 14.7 43.6 24.6 22.8 36 15.7 24.8 20.8 19.6 15.4 8.5 10.1 13.7 13.4 27 67.6 8.8 FB:FBon0033776148623_at 74.5 12.8 75.1 26.9 16.1 18.8 63.6 87.2 26.1 86.6 90.4 78.7 16.2 27.5 27.7 26.4 20.5 105.5 43.6 76.6 70.9 20.5 89.2 86.5 FB:FBgn0036230153536_at 360.5 265.9 447.2 272.9 506.4 404.1 322.3 298.2 273.5 306.4 338.8 395.3 400.9 386.8 369 376.5 316.2 245.8 353 210.1 421.4 207.5 241.3 346 FB:FBgn0024978142793_at 79.3 96.8 92.3 55.9 49 47.5 52.8 41.7 70.2 45 34.2 43 3.8 2.2 44 4.9 44.6 63.3 112 120.8 105.1 42.4 24.5 44.5 FB:FBgn0004624
141246_at 1268 1486 1035 1414.9 1357 1423.1 1951.8 877.1 1229 1459 1242 1605 1601.3 1454.7 1247.1 2256.1 692.4 870.4 1208 1287.2 1069.6 1534.9 1301.2 1741.7 FB:FBgn0036314149269_at 581 861 411.8 840.6 409.4 480.6 70.2 49.8 203.5 146 779.3 510 34.1 177.6 64 65 176.7 202 708 175.9 313.8 144.5 103.4 1304.6 FB:FBgn0037225149269_at 581 861 411.8 840.6 409.4 480.6 70.2 49.8 203.5 146 779.3 510 .34.1 177.6 64 65 176.7 202 708 175.9 313.8 144.5 103.4 1304% FB;FBgn0037229149534_at 149.3 152.4 115.2 119.8 74.2 128.5 67.4 74.2 54.8 240.1 199.7 133.1 90.3 105.1 72 114.4 121.9 97.3 151.9 129.6 144 174.4 131.9 277.6 FB:FBgn0037649149989_at 13 71.5 61.1 64.3 69.8 78.1 32.7 9.9 44.6 8.6 100.2 34.7 89.2 20.5 34.9 40.5 75.2 43.1 79.7 45.9 8.5 68.6 70.9 69.8 FB;FBgn0038386

Probe SetID
ywnonhs1
ywnon hs2
yw
3
ywshort hs1 yw short
h s2yw short
h s3
ywlong hs
1
ywlong hs2
ywlong hs
3
pKb
1pKb
2pKb
3
pKbshort hs1
pKbshort hs2
pKbshort hs
3
p K blong hs1
pKblong hs2
pKb long hs
3PKBYWnon hs 1
PKBYWnon hs 2
PKBYWnon hs 3
PKBYWlong hs 1
PKBYWlong hs 2
PKBYWlong hs 3 Flybase ID
149743_at 2.4 32.2 13.4 34.2 31.9 22.6 52.8 16.3 17.5 17.8 4.4 29.2 34 44.6 5.7 59.1 22.6 18.4 18 40 11.1 34.2 4.1 17 FB:FBgn0038005150852_at 288.2 177.9 286.4 198.1 275.7 263.2 325.4 391.8 238.1 296.9 166.4 247.9 209.7 110.9 236.7 214.1 291.4 127.5 201.6 292.5 260.4 46.9 182 156.2 FB:FBgn0039711147093_at 51.1 22.5 19.6 44.9 32.3 15 13.7 46 12.2 22.3 24.2 58.3 65.5 15.2 44.8 18.4 17.5 88.3 17.9 32.4 13.2 116.1 21.2 2.8 FB:FBgn0033733148697_at 56.7 12.8 12.5 28 5.7 10.9 9.7 25.7 30.1 15.2 12.8 15.3 10.2 32.7 21.9 24.4 6.1 56.5 16.3 35.6 16.5 9.8 18.6 30.3 FB:FBgn0036344153454_at 182.6 134.7 116.4 180.4 224.1 233.7 93.7 35.4 99.9 133 309.6 266 192.6 259.6 168.9 209.3 125.6 168.9 174.4 151.6 128.5 39.2 36.5 111.7 FB:FBgn0027794150612_at 49.2 13.5 26.3 12.1 46.5 18.5 7.2 11.6 14.6 15 11.6 39.3 19.9 55.2 33.3 10.6 7.8 9.5 5.7 30.9 13.6 17.4 11.8 32 FB:FBgn0039368148634_at 2057 1616 1759 1668.7 1965.6 1816.3 1711.3 1439.7 1517.9 1329 1783 1678 1599.4 1685.5 1535.8 1409 1268.7 2053.9 1917.5 1213.6 1972.8 794.3 1204.4 1475.6 FB:FBgn0036256149493_at 3.4 10.3 5.7 23.6 2.3 5.2 5.6 25.4 6.1 7.7 5.5 13 17.8 8.8 3.7 7.3 7.9 9.6 4.9 4.1 6.9 28 81.8 3.6 FB:FBgn0037579146941_at 10.8 22.6 39.6 28.5 26.5 77.8 13.5 15.7 15.2 45.7 35 37.2 35.9 23.4 33.8 43.6 51.1 16.2 21.2 17.1 16.5 20.7 23.4 14.1 FB:FBgn0033513142158_at 2100 2199 2209 1366.4 1273.8 1451.7 2014.2 2709.9 2104 1743 1475 1588 1015.6 1034.5 881.5 1560.2 2178.9 1624.3 2310.7 2155 2256.2 1263.2 1343.7 2070.6 FB:FBgn0038278142699_at 289.6 175 325.1 266 262.2 300.6 421.6 230.6 166.4 276 246.5 288.3 253.6 246 344.4 442.8 320.1 278.1 237.8 233.3 296.5 389.8 382.4 388.1 FB:FBgn0037730145295_at 113.2 19.8 25.1 59.3 93.2 133.6 17.1 17.6 28.1 37.9 28.1 32.5 116.8 37.7 19.6 45.3 71.2 31.4 76 20.8 24.8 44.2 68.3 33.6 FB:FBgn0030913153795_at 2988 2832 2844 2921.7 2623.5 2961.8 2225.8 3653.7 3097.8 2571 2897 3390 4543.5 3982.5 4010.5 3217.3 3982.3 3878.8 2631.3 2738.3 2244.7 2997.1 3235.1 2603.8 FB;FBgn0027338155154_at 446.3 461 491.1 267.7 319.2 308.7 303.4 491.4 594.4 364.6 221.8 234.5 187.4 159 236.9 274.5 278.3 343.8 267.7 485.5 393.2 493.8 467.5 264.3 FB:FBgn0035617142408_at 468.4 514.1 357.2 576.7 523.6 368.9 384.8 469.1 350.8 349.2 485 432.1 574.1 469 217 369.7 617.1 732.3 329.3 688.1 532.3 346.5 292.3 412.6 FB:FBgn0036623147070_at 331.2 115.3 79.2 173.2 150.2 130.8 178.5 66.2 158.1 369.1 307.4 260.4 115.7 177.9 213 19.1 241.7 148 144.8 233.3 130.5 352 362.2 283.7 FB:FBgn0033703148364_at 31.5 47.9 13.1 43.9 33.8 8 34.5 15.6 17.1 20.8 25.2 25.7 59.2 10.9 66.1 9.2 7.5 35.1 11.1 23.9 3.6 32.5 3.9 24.1 FB:FBgn0035823151562_at 58.7 56.1 54.3 46.4 12 4.9 43.4 7.4 50.9 43.5 28.6 36.7 47.5 26 44.4 9.5 49 46.7 44.2 50 54.6 34.4 32.8 49.1 BDGP:GH26019.3prime-hit144539_at 150.4 99 37.2 121.5 126 83.9 56.5 112.7 46.8 92.8 31.6 93.3 120.5 38.1 73.2 183.4 93.1 56.1 157.3 66.8 99.4 98.1 50.5 88.5 FB:FBgn0029789150816_at 409.1 567.2 477.5 452.8 453 407.9 571 564.9 455.7 296.8 377 269 266.1 244.4 252.2 407.1 350.9 476.3 459.5 453.1 490 299.8 369.6 431.4 FB;FBgn0039653154617_at 839.6 670.2 663.9 845.2 862.1 607.1 676.6 658.6 815.5 641.6 866.1 870.8 861.9 714.7 798.5 682.4 679.2 876 674.6 648.4 648.8 916 965.2 864.1 FB;FBgn0036104152032_at 1684 1588 1945 2592.2 2756 3005.1 2063.5 1622.2 1860.7 1524 2472 1548 3371.2 3532.6 3002.6 2330.7 1840.3 1190 1589.3 2107 1559.5 2557.9 3490.7 1861.9 FB:FBgn0022986151190_at 14.2 8.6 22.8 23.4 14 11.6 12.1 9.8 10.3 15.2 10.3 14.2 22.3 16.6 16.6 9.5 9.4 8 8.6 9 28.6 23 13 8.6 FB;FBgn0040713151649_at 48.5 109.2 86.9 165 117 87.1 56.4 102.9 40.5 143.1 113.4 139.1 84.2 162.9 88.9 29.4 59.7 162.4 158.4 67.2 72.9 184.1 67.3 95.1 BDGP:LD18295.3prime-hit149647_at 3.2 11.4 28.8 25.5 4.4 33.6 27.5 15.8 47.2 41 7 5.9 35.8 32.1 27.3 64.5 10.5 16 18.5 36 27 67.6 49.1 3.3 FB;FBgn0037837151399_at 186 111.4 167.5 205.5 239 210 236.8 212.5 208.2 180.2 187.3 215 101 223.1 184.3 284.2 215.6 209.6 212.9 154.3 166.7 233.8 254.5 205.2 FB:FBgn0040930150103_at 11.2 20.4 39.8 87.8 86.1 21.1 11.6 29.9 20.2 15.7 8.4 10 152 129.3 148.2 34.1 80.4 32.1 34.7 80.1 7.5 30.4 21.4 16.7 FB:FBgn0038553147174_at 28.9 60.3 67.2 11.4 44.7 13.3 6.4 32.6 42.3 45.1 7 32.5 44.8 54 68.1 20.9 11.3 10.8 26 21.1 12.9 10 15 6.1 FB:FBgn0033880153203_at 103.7 172.1 120.8 28.5 27.5 86.3 36.3 133.3 182 97.5 125 105.4 22.5 76.9 18.4 85.5 87.2 128.4 152.3 152.2 107.7 11.9 128.7 167.5 FB:FBgn0039156154693_at 255.2 293.5 176.2 137.7 153.6 110.3 240.7 324.5 263.7 280 270.3 278.5 200.6 238.6 221 423.8 379.4 433.2 176.7 268.3 233.9 251.5 173.9 248.4 FB:FBgn0031587141951_at 57.6 36.9 41 6.8 89.5 49.1 52.8 30 38.7 31.2 44.9 54.6 82.8 70 65.7 151.5 54.6 31.9 24.7 49.6 48.3 89.1 101 94.8 BDGP:LD05703.3prime-hlt142084_at 250 99.8 194.4 212.2 238 183.9 120.7 180.8 175.1 193 169.9 232.9 156.9 198.2 187.9 320 171.8 36 163.2 270 93.5 204.4 275.2 157.9 BDGP:LD46437.3prime-hit148045_at 20.1 171.4 101.2 203.7 186.2 278.4 89.5 73.7 115.6 26.5 31.8 42.6 265.8 131.6 92.8 178.7 169.6 124 29.3 73.6 50.5 85.9 129.7 178.2 FB;FBgn0035339151015_at 5.2 17.1 8.5 8.1 24.3 62.2 61.8 43.7 25.9 17.2 3.2 9.7 10.1 14.9 26.7 15.6 6 16.9 3.1 39.2 2.9 10.3 10.2 3.4 FB:FBgn0040528151374_at 9.3 28.5 30.3 9.2 10.9 8.5 6.2 29.9 12.2 18.2 41.8 42.5 7 9.8 5.5 23.1 5.9 17.4 20.8 5.9 7.4 73.3 15.9 10.8 FB:FBgn0040903148998_s_at 24.9 25.2 222.7 27 43.4 32.3 31.2 695.8 654.2 16.7 326.7 46.4 315.1 72.9 50.3 657.8 720.4 917.4 13.6 19.4 23.1 26.7 36.4 16.5 FB:FBgn0036791




















