
Induction of hTERT Expression and Telomerase Activity by Estrogens in Human Ovary Epithelium Cells
Upload
independentCategory
view
2download
0
The Role of Estrogens and Estrogen Receptors in Normal ProstateGrowth and Disease
Gail S. Prins1 and Kenneth S. Korach2
1 Professor, Department of Urology, University of Illinois at Chicago, 820 S Wood St, Chicago, IL 60612,gprins @uic.edu
2 Director, Environmental Disease and Medicine Program; Chief, Laboratory of Reproductive andDevelopmental Toxicology, NIEHS/NIH, 111 Alexander Drive, Research Triangle Park, NC 27709,[email protected]
Keywordsprostate; prostate cancer; estrogens; estradiol; estrogen receptor
Estrogens have significant direct and indirect effects on prostate gland development andhomeostasis and have been long suspected in playing a role in the etiology of prostatic diseases.Direct effects are mediated through prostatic estrogen receptors alpha (ERα) and beta (ERβ)with expression levels changing over time and with disease progression. The present reviewexamines the evidence for a role of estrogens and specific estrogen receptors in prostate growth,differentiation and disease states including prostatitis, benign prostatic hyperplasia (BPH) andcancer and discusses potential therapeutic strategies for growth regulation via these pathways.
Estrogens in the Male and Effects on the Prostate GlandWhile low levels of circulating estrogens are present throughout life in males, there are twotime periods, during in utero development and aging, when males are exposed to relativelyhigher levels of circulating estradiol which have been shown to impact the prostate gland. Inaddition, estrogens may be produced locally within the prostate via conversion of testosteroneto 17β-estradiol by aromatase expressed within the prostate stroma [1,2], thus estrogen actionin the prostate may occur independent of serum levels of this steroid.
During the third trimester of in utero development in humans, rising maternal estradiol anddeclining fetal androgen production result in an elevated estrogen/testosterone (E/T) ratio. Thisrelative increase in estradiol has been shown to directly stimulate extensive squamousmetaplasia within the developing prostatic epithelium which regresses immediately after birthwhen estrogen levels rapidly decline[3,4]. Although the natural role for estrogens duringprostatic development is unclear, it has been proposed that excessive estrogenization duringprostatic development may contribute to the high incidence of BPH and prostatic carcinomacurrently observed in the aging male population [5]. African-American men have a two-foldincreased risk of prostatic carcinoma as compared to their Caucasian counterparts and it hasbeen suggested that this is related, in part, to elevated levels of maternal estrogens during earlygestation in this population [6,7]. Indicators of pregnancy estrogen levels such as length of
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptSteroids. Author manuscript; available in PMC 2009 March 1.
Published in final edited form as:Steroids. 2008 March ; 73(3): 233–244.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

gestation, pre-eclampsia and jaundice indicate a significant correlation between higherestrogen levels and prostate cancer risk [8,9]. In addition, maternal exposure todiethylstilbestrol (DES), a potent synthetic estrogen agonist, during pregnancy was found toresult in more extensive prostatic squamous metaplasia in male offspring than observed withmaternal estradiol alone [10]. While prostatic metaplasia eventually resolved following DESwithdrawal, ectasia and persistent distortion of ductal architecture remained [11]. This has leadto the postulation that men exposed prenatally to DES may be at increased risk for prostaticdisease later in life, although this has not been borne out in limited population studies conductedto date [12]. However, extensive studies with rodent models predict marked abnormalities inthe adult prostate including chronic inflammatory cell infiltration, prostatic intraepithelialneoplasia and increased susceptibility to adult-onset carcinogenesis as a result of earlyestrogenic exposures [13-15]. Although DES use during pregnancy is no longer practiced, therecent realization that certain environmental chemicals have potent estrogenic activities [16]has lead to a renewed interest in evaluating the effects and roles of exogenous estrogens duringprostatic development [17].
The relative levels of free circulating estrogens increase as men age which can increaseestrogenic action in the prostate gland. Bioavailable testosterone levels decline in the agingmale due to decreased production by the testis and increased sex hormone binding globulin(SHGB) levels which combine to lower free circulating testosterone [18]. However, circulatinglevels of free estradiol remain constant in the aging male due to an age-related increase in bodyweight and adipose cells which express high levels of aromatase and peripherally convertandrogens to estrogens [19]. The net result is a significant increase in the E/T ratio allowingthe balance between androgen and estrogen regulation of prostate growth to shift towardsestrogen dominance. It has been proposed that increased estrogenic stimulation of the prostatein the aging male may lead to reactivation of growth and subsequent neoplastic transformation[20,21].
Estrogens have significant direct effects on the adult prostate gland as well and have long beensuspected in the etiology of prostatic disease [22,23]. In 1936, Dorothy Price demonstratedthat estrogens given to adult rodents leads to hyperplasia, squamous metaplasia andkeritinization of the prostate epithelium [24]. Long-term exposure of adult rats tosupraphysiologic but non-pharmacologic levels of estradiol and physiologic levels oftestosterone leads to prostatic intraepithelial neoplasia (PIN) in the dorsolateral lobe andcarcinoma in the periurethral ducts of Noble rats and this is used as a model for estrogen-induced adenocarcinoma of the prostate [25]. Estrogen-induced aberrations in prostateepithelial growth have also been observed in dogs, monkeys and humans with results varyingaccording to species and experimental conditions [26]. In addition to epithelial effects,estrogens induce a preferential stimulation of prostate stromal cell proliferation, consequently,combined administration of estrogens with androgens has been used to experimentally induceBPH in dogs [26,27]. In humans, the estradiol:DHT ratio increases moderately within normalprostate epithelial and stromal cells upon aging, however, the increased ratio is massive withinBPH tissue which directly implicates estradiol in the disease process [28].
Estrogen effects on the prostate gland may also be indirectly mediated through alterations inother serum hormones. Estrogens stimulate the pituitary release of PRL and some, but not all,of estrogenic effects have been attributed to direct PRL action on the prostate [29-31].Furthermore, estradiol negatively feeds back on the hypothalamic-hypophyseal-testicular axis,blocking lutinizing hormone (LH) secretion and testicular steroidogenesis of androgens (i.e.chemical castration). This feedback regulation was the basis for high-dose estrogen therapy ofprostate cancer for several decades [32]. Despite these indirect effects, there is ample evidencethrough hormone-controlled studies and with in vitro approaches to clearly document that many
Prins and Korach Page 2
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

of the estrogenic effects on the prostate are directly mediated through prostatic expression ofestrogen receptors [33-35].
Estrogen ReceptorsEstrogen action is mediated through specific estrogen receptors (ER) which are members of alarge family of nuclear transcription factors found ubiquitously throughout the animalkingdom. Using a classification system based on the cytochrome P450 family [36] estrogensreceptors (ERα and ERβ) are the sole members of the NR3A subgroup, ERα and ERβ beingNR3A1 and NR3A2, respectively [37].
The human ESR1 (ERα) cDNA was first cloned in 1985 [38] and has since been isolated inmultiple species from humans to fish [39]. The encoded ERα proteins are 595 and 599 aminoacids in length in human and mice, respectively, with an approximate molecular weight of 66kDa (Figure 1A) [38,40]. Numerous naturally occurring variants of the ESR1 mRNA in normaland neoplastic tissues of several species have been described but the existence of correspondingproteins remains controversial [41,42].
A second ER gene termed ESR2 (ERβ) was discovered in 1996 from rat [43] and human tissues[44] and has also been cloned in multiple species. Unlike the androgen or progesteronereceptors, ERα and ERβ are not isoforms, but are encoded by separate genes possibly by geneduplication on different chromosomes, and therefore are distinct receptor forms. Translationof the ESR2 mRNA yields a receptor of 549 amino acids in rodents and 530 amino acids inhumans, each with an approximate molecular weight of 60−63 kDa. Therefore, ERβ is slightlysmaller than ERα with the majority of this difference in the N-terminus. A number of varianttranscripts of the ESR2 gene have been described [45]; however; unlike ERα, it appears thatthese non-ligand binding isoforms (ERβ2, ERβ4, ERβ5) co-exist with the ligand-bindingERβ1 in certain tissues where they can heterodimerize with ERβ1 and modulate itstranscriptional activities.
ERs are modular in both structure and function [39,46,47] and are composed of 6 functionalmodules; an N-terminal (NTD) or A/B domain, the DNA-binding (C) domain, a hinge (D)region, ligand-binding (E) domain and a unique C-terminal F domain of unknown function(Figure 1 B). The NTD (A/B domain) harbors the transcriptional activation function 1 (AF-1)domain and specifies the cell and promoter-specific activity of the receptor as well as co-receptor protein interactions. Phosphorylation of the A/B domain is the most well characterizedpost-translational modification and occurs via the actions of multiple intracellular signalingpathways such as MAPK and PKA pathways at specific sites [39,47]. The greatest structuraldisparity between ERα and ERβ lies within the A/B domain which exhibits only 24% homologybetween the two receptors and is ∼90 nucleotides shorter in the ERβ as compared to ERβ[48] (Figure 1). This divergence likely accounts for the many functional differences that havebeen revealed from comparative studies of the two ER forms including coactivator interactionswith A/B domain as well as ER-protein interactions, such as AP-1, which modify genetranscription through nonclassical pathways [49-51]. Notably, the AF-1 domain of ERβ is notconstitutive and depends on ligand-activated AF-2 action whereas the ERα AF-1 can functionindependent of AF-2 and ligand. Consequently, certain antagonists (e.g. tamoxifen) that exhibitsome agonist-like properties when bound to ERα exhibit no such agonist activity with ERβ[49,51]. The value in differential responses lies with the potential to selectively modulateestrogen action using SERMs that behave differently for ERα and ERβ in a tissue-specificmanner. Overall, ERβ tends to be a less effective transcriptional activator compared to ERαin vitro [44,49,52].
Prins and Korach Page 3
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The DNA binding domain (DBD) or C domain is the region of the receptor that recognizes andbinds to cis-acting enhancer DNA sequences known as estrogen response elements (ERE)located within regulatory regions of target genes. It is the most highly conserved region (>95%homology) of the ERα and ERβ which accounts for the similar affinity for the same EREs[43,53]. The functionality of the C domain is provided by a motif of two zinc-fingers encodedby separate exons, each composed of four cysteine residues that complex with a single Zn2+
ion. The “P-box” (proximal) encodes the first zinc-finger and confers specificity sequences onthe ER protein for EREs and forms a “recognition helix” [39]. The “D-box” (distal) encodesthe second zinc-finger which is involved in spacer sequence and receptor dimerization [39].In addition, ERα can induce gene expression when bound as a monomer to an ERE half-sitein close proximity to Sp1 binding sites due to cell context-specific interactions of proteins withspecific amino acids on Zn fingers 1 and 2 DBD [54].
The hinge region or D domain of the ERs harbors a nuclear localization signal that influencescellular compartmentalization of the receptor. The ligand-binding (LBD) or E domain is ahighly structured, multifunctional region that serves to specifically bind estrogens and providefor ligand-dependent transcriptional activity [39]. An Activation Function-2 (AF-2) domainlocated in the C-terminus of the E domain mediates this latter function and is subject to post-translational receptor modifications [39]. Also harbored within the E domain is a strongreceptor dimerization interface. Although there is only ∼ 60% homology in the primarysequence of the LBD of ERα and ERβ, comparative crystal structure studies of liganded andun-liganded LBDs indicate a highly conserved structural arrangement of 12 α-helices whichserve as a docking station for agonists and antagonists and a co-activator/repressor recruitmentsite [43,55]. The LBD of ERα and ERβ binds the endogenous hormone estradiol, estrone andestriol with similar affinity (ERα = 0.1 nM; ERβ = 0.4 nM) and exhibits equal affinity for DES[56,57]. However, due to divergence in homology, ERα and ERβ also exhibit measurabledifferences in their affinity for other endogenous steroids and xenoestrogens [57-59]. Forexample, ERβ tends to exhibit a stronger affinity for certain phytoestrogens (e.g. genestein andcoumestrol) [57,58]. Recent advances have now permitted the generation of ER-selective non-steroidal ligands [49,59,60] that exploit as well as illustrate differences between the LBDs ofERα and ERβ and provide for pharmacological tools to discern the overall function of eachER.
Among the sex steroid receptors, the well-defined F domain is only found in ERs. Studies haveindicated a role for the F-domain in receptor stability and co-activator recruitment [61] and inrecent findings, in modulating the activational state of AF-2 by affecting the conformationalequilibrium of the LBD and selectively modulating its response to ligands [62].
Estrogen Receptor Mechanisms of ActionThe ligand/ERE-dependent or “classic” model of estrogen action states that the ER resides inthe nucleus but is sequestered in a multi-protein inhibitory complex in the absence of hormone.Upon hormone binding a conformational change occurs in the ER, transforming it to an“activated” state that homodimerizes, shows increased phosphorylation, and binds to an EREwithin target gene promoters. The ligand/ERE-bound receptor complex interacts with thegeneral transcription apparatus via co-regulatory proteins to promote transcription of the targetgene. This classic estrogen receptor mechanism is dependent on the functions of both AF-1and AF-2 domains of the receptor which synergize via the recruitment of co-activator proteins,most notably the p160 family members. It is generally believed that the DNA-bound receptor/co-activator complex facilitates chromatin remodeling and formation of a stable transcriptionpre-initiation complex. When acting via a classic ERE-driven mechanism in vitro, ERαhomodimers and ERα:ERβ heterodimers tend to be stronger activators of transcriptioncompared to ERβ1 homodimers [44,45,49,52]. In breast cancer cell lines, ERβ transfection
Prins and Korach Page 4
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

into ERα positive cells inhibited ligand-induced ERα-dependent transcription whereas ERαtransfection had little effect on ERβ transactivation [63]. Furthermore, recent findings inprostate cancer cell lines show that non-ligand binding ERβ isoforms (ERβ2, ERβ4, ERβ5)can heterodimerize with ERβ1 under the stimulation of estrogens, but not phytoestrogens, andenhance ERβ1 transactivation in a ligand-dependent manner [45].
There is ample evidence that ERs can be activated in a ligand-independent manner viaintracellular second messenger and signaling pathways. This activation throughphosphorylation is referred to as receptor crosstalk and it permits induction of ER target genesin the absence of steroid ligand [64,65]. Polypeptide growth factors phosphorylate ERα-mediated gene expression via the mitogen activated protein kinase (MAPK) pathway and theG-protein/cyclic-adenosine monophosphate (cAMP)/ protein kinase A (PKA) pathway. Whilegrowth factors are able to mimic the effects of estradiol in the rodent uterus via this estradiol-independent ERα activation [66,67], such studies in the prostate are not as yet well described.
Ligand-activated ERα can also stimulate the expression of genes that lack a conspicuous HREwithin their promoter [68,69]. This mechanism of HRE-independent ER activation ispostulated to involve a “tethering” of the ligand-activated ER to other transcription factors thatare directly bound to DNA via their respective response elements [68,69]. Therefore, ERαacting in this fashion, may be better defined as a co-regulator rather than a direct actingtranscription factor. Estradiol/ERα regulation of several genes, including ovalbumin,collagenase, insulin-like growth factor-1 and cyclin D is believed to occur via a tethering ofthe receptor to a DNA-bound AP-1 (Fos/Jun) complex with the gene promoter [69]. A similarERE-independent mechanism of ERα action has been documented for genes that possess aGC-rich region or Sp1 binding site within the promoter where ERα enhances the actions of abound Sp1 complex [68,69]. How much of this type of gene regulation is involved in prostategrowth and stimulation is unknown at the present time.
Estrogen Receptor Expression in the Prostate GlandOriginal studies with ligand binding assays, sucrose density gradients and autoradiographyidentified specific estrogen binding sites in both epithelial and stromal cell fractions of theprostate gland in different species and these were assumed to be a single ER. With the discoveryof ERß in addition to ERα, the localization and relative contributions of each receptor typerequired reanalysis. Results demonstrated that for the most part, ERα and ERß are expressedin different cellular compartments of the prostate gland with ERα localized primarily toprostatic stromal cells and ERß primarily expressed in prostate epithelium. This differentiallocation as well as differential affinity of the two ERs for ligands, enhancers and co-activatorsmay explain the diverse biological functions of estrogens within the prostate gland and mayalso be exploited for regulation of prostate disease.
ERαERα is localized primarily to the stromal cells of the adult prostate gland in man, dogs, monkeysand rodents [70-74]. Immunohistochemical analysis reveals, however, that ERα expression isheterogeneous in stromal cells, i.e. only a portion of the cells are ERα positive while manyremain ERα negative. Studies in rodent prostate glands have shown a relatively high percentageof stromal cells express ERα mRNA and protein during perinatal morphogenesis and thisproportion significantly declines thereafter suggesting a specific role for ERα in prostatedevelopment [73,75,76]. A decline in expression with puberty suggests that androgens maynormally suppress ERα expression, a finding that has been borne out in direct studies [77,78].In humans, ERα has been consistently observed in stromal cells during fetal development[79]. However, while one report restricts ERα protein to only stromal cells [76], a recent reportindicates the presence of ERα in fetal prostatic utricle and periurethral epithelium during mid-
Prins and Korach Page 5
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

to-late gestation [79]. Importantly, squamous metaplasia, observed in all developing humanprostates during the third trimester, is directly associated with epithelial ERα in the periurethralducts and stromal ERα in the peripheral prostatic acini [79].
It is believed that stromal proliferation, a hallmark response to estrogen treatment in mostspecies, may be mediated through stromal ERα. In humans, there is evidence for an increasedaccumulation of estradiol in nuclei of stromal cells in BPH specimens [80] suggesting thatelevated ERα in stromal cells may be involved in the etiology of BPH. Of interest, epithelialcells in prostatic periurethral ducts have also been found to consistently express ERα in bothnormal and BPH tissue [70]. Since this is the prostatic region which forms BPH, it is possiblethat epithelial ERα in that specific region are involved. In rodents, periductal mesenchymaland smooth muscle cells, which are in close proximity to epithelial cells, express ERα whileinterductal fibroblasts are ERα negative [73]. The close proximity of ERα positive stromalcells to epithelial cells allows for paracrine effects of estrogens on prostate epithelium. Indeed,work with ERα knock out mice (αERKO) demonstrated that estradiol-induced squamousmetaplasia in adult prostates is mediated through ERα [81]. Similarly, using αERKO andβERKO mice, neonatal estrogenization of the prostate, which includes stromal hyperplasia,epithelial PIN lesions and inflammatory cell infiltration, was shown to be mediated throughstromal ERα [82]. It is noteworthy, however, that deletion of ERα in transgenic mice did notproduce a marked phenotype in the prostate suggesting that ERα's role in the prostate gland isnot necessary for normal growth and function [83,84].
Recent reports have shown that in prostatic carcinoma, the ERα gene is methylated leading tosilencing of this gene, loss of ERα transcription and ERα protein [85,86]. While comparativedata with normal prostate specimens was not available, it was noted that the incidence ofERα gene methylation and silencing increased with progression of prostatic disease from BPHto low grade and to high grade cancer. Thus it was proposed that ERα may have a tumorsuppressor role in the prostate gland and loss of its expression may be an early event in prostaticdisease. Interestingly, ERα expression has been observed in some prostate cancer cell lines[86] as well as in hormone refractory and metastatic lesions suggesting its re-emergence ascancer progresses [87] although this has not been consistently seen in all studies [88]. It is alsonoteworthy that prostate cancer risk has been associated with genetic polymorphisms in theERα gene particularly within Japanese and African American populations implicating apotential causal relationship between ERα mediated estrogenic action and prostate cancer[89,90].
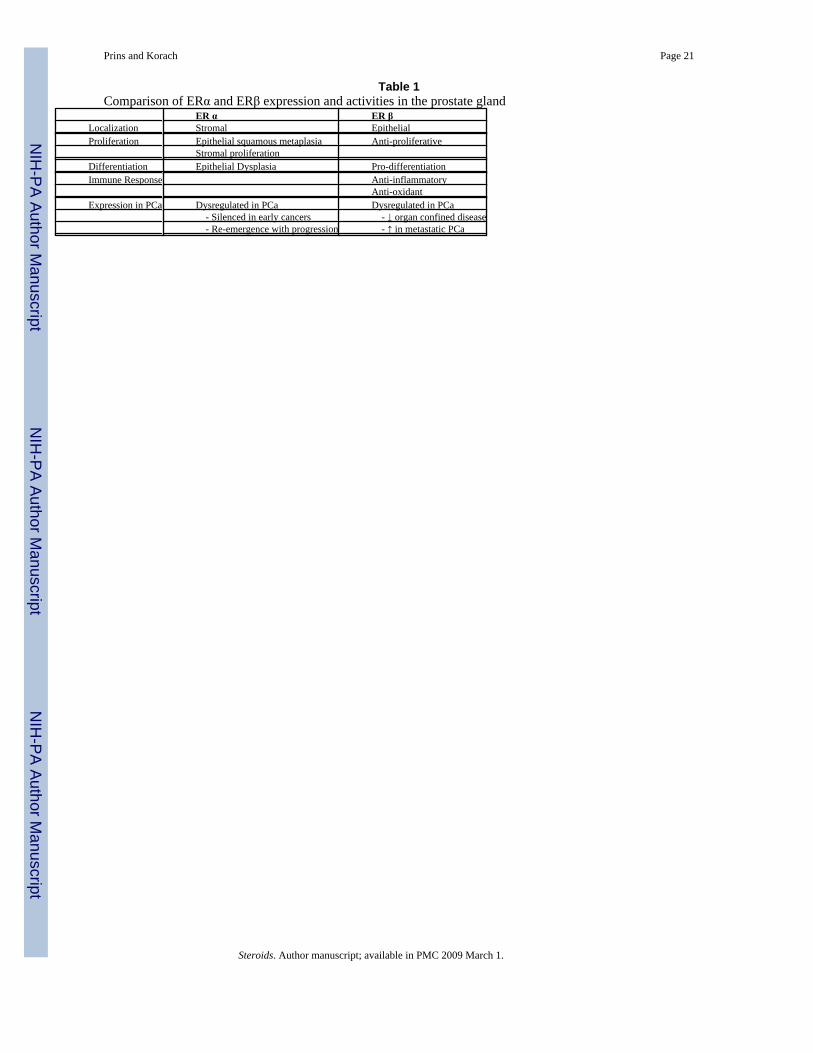
In summary, there is evidence across multiple species that estrogens acting through stromalERα may contribute at some level to the etiology of the most prevalent prostatic diseasesincluding chronic prostatitis, BPH, carcinogenesis and cancer progression (Table 1).
ERβSince ERβ was originally cloned from a rat prostate cDNA library, it was not surprising to findthat the rat prostate expressed this receptor at levels comparable to those found in other high-expressing reproductive organs such as the ovary, endometrium and testis [43,91]. In the ratand murine prostate, ERβ mRNA and protein are primarily localized to differentiated luminalepithelial cells, which may preclude formation of ERα:ERβ heterodimers in this organ [82,92,93]. Expression of ERβ is low at birth, increases as epithelial cells cytodifferentiate andreaches maximal expression with onset of secretory capacity at puberty which suggests a rolefor ERβ in the differentiated function of the rodent prostate [92]. In the adult, an ERβ expressiongradient is observed with low proximal levels and increased expression distally which maycontribute to heterogeneity in differentiation and function along the ductal length [92]. Incontrast to ERα, androgens up-regulate expression of ERβ in the rodent prostate gland whileestrogens to not autoregulate ERβ [92,94,95].
Prins and Korach Page 6
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

It is noteworthy that the developmental pattern for ERβ in the human prostate differs markedlyfrom the rodent. As early as fetal week 7, ERβ is expressed throughout the urogenital sinusepithelium and stroma and this strong expression is maintained in most epithelial and stromalcells throughout gestation suggesting the involvement of ERβ and estrogens in morphogenesisand differentiation [79]. While this pattern is maintained postnatally for several months,ERβ expression declines thereafter with a noticeable decrease in adluminal cells at puberty[76]. In the adult human prostate, ERβ mRNA expression is low relative to testicularexpression, again showing a divergence from the rodent prostate gland [44,91]. Reports varyon ERβ localization in the human prostate which may be a function of antibodies used inimmunohistochemical assays. While some have shown that ERβ is expressed by basalepithelial cells with lower stromal cell expression [74,88], others have shown high expressionof ERβ in both basal and luminal epithelial cells of the adult human prostate [96,97]. In normaland tumorigenic human prostate epithelial cell lines, ERβ is expressed at high levels whileERα is typically absent [86,98]. In response to in vitro estrogen exposure, estrogen regulatedgenes (progesterone receptor, pS2) are activated again pointing towards a role for ERß in thedifferentiated function of the prostatic cell. Together, these findings indicate that, in the prostateepithelium, ERβ may be the key mediator of estrogen-induced events. The prostate gland alsoexpresses ERβ isoform variants which have been shown to act as either constitutive activators,transcription enhancers or dominant negative regulators of estrogen action which furthercomplicates estrogenic action within this gland [45,99,100].
Putative roles of ERβWhile ERβ is the predominant estrogen receptor expressed in the adult prostate gland, its rolehas not yet been clearly established. As stated above, indirect evidence exists for a role ofERβ in the differentiated state of the prostate epithelium. A recent study using βERKO miceshowed a shift in basal, intermediate and luminal epithelial cell markers in the prostate towardsa less differentiated gland which supports this purported role [101]. As a counterpart to thehypothesis that ERß plays a role in epithelial differentiation, it has also been suggested thatERß has an antiproliferative role in the prostate and participates as a brake for androgenicstimulation of prostate growth [102]. Indirect evidence for this role was suggested by thehyperplastic and dysplastic adult prostate epithelium with reduced ERβ expression followingneonatal estrogen exposure [92]. Direct studies for a role of ERβ in prostate proliferation usingβERKO mice have yielded conflicting results. While some studies show epithelial hyperplasiawith increased BrdU labeling in the βERKO prostates [103], this was not supported insubsequent studies using the same mice [82,104] or different βERKO models [105]. Morerecently, direct evidence has been presented for an antiproliferative role of ERβ. Prostateepithelium in aromatase knock out mice (ArKO) becomes hyperplastic with age due tohormonal imbalance. Using tissue recombinants of ArKO neonatal seminal vesiclemesenchyme and adult ArKO anterior prostate epithelium grafted under the renal capsule, thehyperplastic epithelium was reverted to the normal state with the administration of an ERβspecific agonist [106].
Studies in our laboratories suggest that ERβ may also play an immunomodulatory role in theprostate gland. βERKO (n=18) and wild-type (n=18) mice were aged to one year of age andblinded histologic analysis by two independent investigators was performed for the entireglandular complex using serial sections (>100 sections/prostate complex). Aging associatedchanges were noted at similar incidence in the ventral, dorsal and lateral lobes of both genotypeswhich consisted of reduced secretions, flattened epithelium and degenerated apoptotic cellswithin ductal lumens as is typical in the aged murine prostate. Proliferative and apoptotic scoreswere not different for the two genotypes. However, a feature observed in βERKO but not inwild-type prostates was abundant-to-massive lymphoid aggregates which were at timesassociated with reactive (proliferative) epithelium consistent with injury-repair cycles (Figure
Prins and Korach Page 7
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

2). T-cell infiltration throughout the prostatic complex was blindly scored as 0 (absent), 1 (rare),2 (focal), 3 (abundant) and 4 (massive) for the two genotypes. As shown in Figure 2, the percentof wild-type mice in each category revealed that rare or focal T-cell infiltration were presentin 50% of animals while the remaining 50% of wild-type mice had no inflammatory cellspresent. In marked contrast, T-cell infiltration scores for the βERKO mice showed that the vastmajority (89%) of mice possessed prostatic inflammatory cells with 44% of cases presentingwith abundant or massive immune cell infiltration (p< 0.005; Chi-square test). Thus we proposethat ERß may normally play an immunoprotective role in the prostate gland perhaps limitingtissue damage or modulating expression of stimulus for immune cell infiltration. Estrogens arewidely known to affect the development and regulation of the immune system and have beenshown to exert potent anti-inflammatory effects [107,108]. The present data suggests that theanti-inflammatory effects at the level of the prostate gland may be mediated through ERß. Thishypothesis is supported by a study which demonstrated that an ERβ-selective ligand was ableto prevent inflammatory bowel disease in a rat model [109]. Such an approach may holdpromise for treatment of prostatitis which is the most prevalent of prostatic diseases.Furthermore, there is increasing evidence for a link between chronic prostatic inflammationand prostate cancer etiology [110] suggesting a potential protective role for prostatic ERβ inthis regard.
ERβ has also been proposed to have an anti-oxidant function. ERβ can bind the electrophile/antioxidant response element (EpRE) and is a more potent activator at the EpRE element thanERα [111]. Thus ERß is capable of inducing genes that encode chemoprotective detoxificationenzymes (quinone reductase, glutathione S-transferase) and may play an active role inprotecting prostate epithelial cells from carcinogens by detoxifying electrophiles. This isparticularly relevant since GST-pi has been shown to be critical in protecting against prostatecancer through genome damage initiated by inflammatory cells and carcinogens [112].
ERβ in prostate cancerDynamic changes in ERβ expression have been observed during the progression of prostatecancer which suggests that estrogen action through ERβ may play an important role in prostatecarcinogenesis, metastasis and perhaps, androgen independence. Most reports on ERβexpression concur that levels decline in localized prostate cancer with increasing grade fromPIN through low to high Gleason scores [88,97,113,114]. This loss of ERβ expression in organconfined prostate cancer has been shown to be epigenetically regulated by progressivehypermethylation of ERβ promoter CpG islands causing transcriptional silencing [115]. Arecent study suggests that this promoter hypermethylation may be preceded by loss of AP-2expression in prostate cancer cells which permits methylation at a critical AP-2 binding site inthe ERβ promoter [116]. This expression pattern fits with the proposed antiproliferative andpro-differentiation function of prostatic ERβ with its loss permitting unregulated growth andde-differentiation of prostatic epithelium. In support of ERβ as a putative tumor suppressor, arecent study using adenoviral vectors found that ERβ expression in prostate cancer cell linesinhibited growth and invasiveness suggesting that loss of ERβ in higher grade tumors permitsproliferation and eventual metastasis [117].
Counter to this concept, ERβ expression remerges as prostate cancer metastasizes to distancesites with 100% of osseous and non-osseous metastatic cells expressing ERβ to varying degrees[96,118,119]. ERβ promoter analysis revealed complete hypomethylation of the three 5’ CpGislands in the ERβ 5’ flanking region in metastatic prostate cancer which permits high ERβgene expression at these distant sites. Thus if ERβ is considered antiproliferative, it is unclearhow high ERβ expression in metastatic disease permits uncontrolled proliferation withmetastatic spread. Since one study reported that localized prostate cancers which retain ERβin the primary tumor were associated with a higher rate of recurrence [114], it is possible that
Prins and Korach Page 8
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ERβ expression in metastatic cells may be a function of a selective advantage of a subset ofERβ-retaining cells in the primary tumor to metastasize. However, this possibility must bereconciled with evidence that ERβ can inhibit migration and invasion in prostate cancer cells[117,120]. Thus it is currently unclear whether ERβ functions has an antiprolferative role inprostate cancer or whether it promotes metastasis and growth at distant sites. Whatever thecase, the important feature is the strong ERβ expression in metastatic, androgen-independentprostate cancer. This suggests that the metastatic cells are targets of estrogen action and thusmay be potential targets for therapeutic interventions with antiestrogenic agents or moreeffective ERβ selective antagonists.
In summary, evidence has been shown for a variety of roles for prostatic epithelial ERβ whichare pro-differentiation, anti-proliferative, anti-inflammatory and as an inducer of anti-oxidantgenes. While loss of ERβ may contribute to prostate cancer progression in organ confineddisease, strong re-emergence of ERβ at metastatic sites implicates a potential role in androgen-independent progression (Table 1).
Estrogen agonists, antagonists, modulators as therapeutic agents forprostate cancer
Due to the above considerations concerning the effects of estrogens on prostate growth andcarcinogenesis and the expression of ERs in prostate stromal and epithelial cells, the use ofestrogen/antiestrogen therapy may have efficacy for the treatment of prostate cancer. Thusmany estrogens including phytoestrogens, DES and 2-methoxyestradiol as well as selectiveestrogen receptor modulators (SERMs) including raloxifene, ICI 182,780, trioxifene andtorimifene have been shown to affect prostate tumor growth through a variety of mechanisms[20,120-126]. While initial clinical trials using tamoxifene and toremifene proved to beunremarkable for the treatment of prostate disease [127,128], more recent Phase II studiesindicate that the toremifene may in fact effectively block development and progression ofclinical prostate cancer and further clinical trials are underway [126,129]. In is unclear atpresent whether these agents act primarily through antagonism of ERα or ERβ or whether theydifferentially modify the actions of both receptors. Future studies with ERα and ERβ specificantagonists and agonists [109] as well as the development of third generation SERMs mayprovide insight into the specific prostatic ER which is the most effective target for therapeuticuse.
Another possible site for treatment of prostate cancer is through inhibition of estradiolproduction using aromatase inhibitors [130,131]. Recent work has shown high expression ofaromatase in prostate cancer with alternate promotor utilization which suggests thatintraprostatic production of estradiol may contribute to progression of this disease [132].Despite this, clinical trials with aromatase inhibitors failed to show efficacy of global estrogenreduction as an effective prostate cancer therapy [133,134].
Therapeutic basis for natural estrogenic products as chemopreventiveagents
In addition to pharmaceutical approaches, there is considerable evidence that phytoestrogensmay modulate prostate growth which has been the basis for herbal supplements and dietarymodulation for the treatment of abnormal prostate growth. Genestein [135,136], resveratrol[137] and soy [138] have all been shown to have beneficial effects and consumption of theseproducts has been inversely correlated with prostate cancer risk [138-140]. High prostaticexpression of ERβ may explain why phytoestrogens (genestein, apigenin, coumestrol) arebeneficial to prostate health since these compounds have been found to bind to ERß with anaffinity up to 10 times higher than for ERα [57,141]. Thus if ERβ is in fact anti-proliferative
Prins and Korach Page 9
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

in early stage prostate cancer, specific activation of this receptor may be the basis for thebeneficial effects of these natural products. However, since clinical trials evaluating theusefulness of these dietary approaches have been limited, caution in their usage must be issueduntil their safety and effectiveness has been clearly demonstrated.
ConclusionsA considerable amount of evidence now exists that estrogens play an important role in prostatedevelopment and homeostasis with different actions being mediated by the stromal ERα andepithelial ERβ. Evidence also indicates that estrogen action mediated through the separatereceptors may contribute to the etiology and progression of multiple prostate diseased states.These findings provide new avenues and alternative approaches for the treatment of prostatediseases including prostate cancer with novel therapies directed at estrogen receptors orestrogen metabolism. Since the two types of estrogen receptors may play distinct and perhapsopposing roles in many diseases of the prostate including cancer progression, it is possible thatreceptor-specific agonists and antagonists may provide the most beneficial therapeuticstrategies in future clinical trials.
REFERENCES1. Matzkin H, Soloway MS. Immunohistochemical evidence of the existence and localization of
aromatase in human prostatic tissues. Prostate 1992;21:309–314. [PubMed: 1281323]2. Risbridger GP, Bianco JJ, Ellem SJ, McPherson SJ. Estrogens and prostate cancer. Endocr Relat Cancer
2003;10:187–191. [PubMed: 12790781]3. Zondek T, Mansfield MD, Attree SL, Zondek LH. Hormone levels in the fetal and neonatal prostate.
Acta Endocr 1986;112:447–456. [PubMed: 3751459]4. Wernet N, Kern L, Heitz PH, Bonkhoff H, Goebbels IR, Seitz G, Inniger R, Remberger K, Dhom G.
Morphological and immunohistochemical investigations of the utriculus prostaticus from the fetalperiod up to adulthood. Prostate 1990;17:19–30. [PubMed: 1696712]
5. Rajfer J, Coffey DS. Sex steroid imprinting of the immature prostate. Invest Urol 1978;16(3):186–190.[PubMed: 711410]
6. Henderson BE, Bernstein L, Ross RK, Depue RH, Judd HL. The early in utero oestrogen andtestosterone environment of blacks and whites: potential effects on male offspring. Br J Cancer 1988;57(2):216–218. [PubMed: 3358915]
7. Platz EA, Giovannucci E. The epidemiology of sex steroid hormones and their signaling and metabolicpathways in the etiology of prostate cancer. Journal of Steroid Biochemistry and Molecular Biology2004;92:237–253. [PubMed: 15663987]
8. Ekbom A, Wuu J, Adami HO, Lu CM, Lagiou P, Trichopoulos D, Hsieh C. Duration of gestation andprostate cancer risk in offspring. Cancer Epidemiol Biomarkers Prev 2000;9(2):221–223. [PubMed:10698486]
9. Ekbom A, Hsieh CC, Lipworth L, Wolk A, Ponten J, Adami HO, Trichopoulos D. Perinatalcharacteristics in relation to incidence of and mortality from prostate cancer. British Med J1996;313:337–341.
10. Driscoll SG, Taylor SH. Effects of prenatal maternal estrogen on the male urogenital system. ObstetGynecol 1980;56(5):537–542. [PubMed: 7432722]
11. Yonemura CY, Cunha GR, Sugimura Y, Mee SL. Temporal and spatial factors in diethylstilbestrol-induced squamous metaplasia in the developing human prostate. II. Persistent changes after removalof diethylstilbestrol. Acta Anatomica 1995;153:1–11. [PubMed: 8560954]
12. Giusti RM, Iwamoto K, Hatch EE. Diethylstilbestrol revisited: A review of the long-term healtheffects. Ann Int Med 1995;122(10):778–788. [PubMed: 7717601]
13. Arai, Y.; Mori, T.; Suzuki, Y.; Bern, HA. Long-term effects of perinatal exposure to sex steroids anddiethylstilbestrol on the reproductive system of male mammals.. In: Bourne GHaD, JF., editor.International Review of Cytology. 84. Academic Press, Inc.; New York, New York: 1983. p. 235-268.
Prins and Korach Page 10
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

14. Prins, GS. Developmental estrogenization of the prostate gland.. In: Naz, RK., editor. Prostate: Basicand Clinical Aspects. C. R. C. Press; Boca Raton: 1997. p. 247-265.Chapter 10
15. Prins GS, Huang L, Birch L, Pu Y. The role of estrogens in normal and abnormal development of theprostate gland. Annals of the New York Academy of Sciences 2006;1089:1–13. [PubMed: 17261752]
16. McLachlan JA. Environmental signaling: what embryos and evolution teach us about endocrinedisrupting chemicals. Endocrine Reviews 2001;22(3):319–341. [PubMed: 11399747]
17. Ho SM, Tang WY, Belmonte J, Prins GS. Developmental exposure estradiol and bisphenol A (BPA)increases susceptibility to prostate carcinogenesis and epigenetically regulates phosphodisesterasetype 4 variant (PDE4D4) in the rat prostate. Cancer Research 2006;66(11):5624–5632. [PubMed:16740699]
18. K JM, Vermeulen A. The decline of androgen levels in elderly men and its clinical and therapeuticimplications. Endocrin Rev 2005;26:833–875.
19. Vermeulen A, Kaufman JM, Goemaere S, van Pottelberg I. Estradiol in elderly men. Aging Male2002;5(2):98–102. [PubMed: 12198740]
20. Ho SM. Estrogens and antiestrogens: Key mediators of prostate carcinogenesis and new therapeuticcandidates. Journal of Cell Biochemistry 2004;91:491–503.
21. King KJ, Nicholson HD, Assinder SJ. Effect of increasing ratio of estrogen:androgen on proliferationof normal and human prostate stromal and epithelial cells and the malignant cell line LNCaP. Prostate2006;66:105–114. [PubMed: 16114065]
22. Thomas JA, Keenan EJ. Effects of estrogen on the prostate. Journal of Andrology 1994;15(2):97–99.[PubMed: 8056642]
23. Castagnetta, LAM.; Carruba, G. Non-reproductive Actions of Sex Steroids. Chichester; Wiley: 1995.Human prostate cancer: A direct role for oestrogens.; p. 269-289.
24. Price D. Normal development of the prostate and seminal vesicles of the rat with a study ofexperimental postnatal modifications. Am J Anat 1936;60:79–127.
25. Leav I, Ho S, Ofner P, Merk F, Kwan P, Damassa D. Biochemical alterations in sex hormone-inducedhyperplasia and dysplasia of the dorsolateral prostates of Noble rats. J Nat'l Canc Inst 1988;80(13):1045–1053.
26. Coffey D, Walsh P. Clinical and experimental studies of benign prostatic hyperplasia. Urol Clin NoAm 1990;17(3):461–475.
27. DeKlerk D, Coffey D, Weing L, McDermott I, Reiener W, Robinson C, Scott W, Strandberg J, TalalayP, Walsh P, et al. Comparison of spontaneous and experimetally induced canine prostatic hyperplasia.J Clin Invest 1979;64:842–849. [PubMed: 89121]
28. Krieg M, Klotzl G, Kaufmann J, Voigt K. Stroma of human benign prostatic hyperplasia: preferentialtissue for androgen metabolism and oestrogen binding. Acta Endocr 1981;96:422–432. [PubMed:6163294]
29. Lee C, Prins GS, Henneberry MO, Grayhack JT. Effect of estradiol on the rat prostate in the presenceand absence of testosterone and pituitary. J Androl 1981;2(6):293–299.
30. Gilleran JP, Putz O, De Jong M, De Jong S, Birch L, Pu Y, Huang L, Prins GS. The Role of Prolactinin the Prostatic Inflammatory Response to Neonatal Estrogen. Endocrinology 2003;144(5):2046–2054. [PubMed: 12697713]
31. Lane K, Leav I, Ziar J, Bridges R, Rand W, Ho SM. Suppression of testosterone and estradiol-17beta-induced dysplasia in the dorsolateral prostate of Noble rats by bromocriptine. Carcinogenesis 1997;18(8):1505–1520. [PubMed: 9276623]
32. Huggins C, Hodges CF. Studies on prostatic cancer. I. The effect of castration, of estrogen, and ofandrogen injection on serum phosphatases in metastic carcinoma of the prostate. Can Res1941;1:293–297.
33. Prins GS. Neonatal estrogen exposure induces lobe-specific alterations in adult rat prostate androgenreceptor expression. Endocrinology 1992;130(6):3703–3714. [PubMed: 1597166]
34. Nevalainen MT, Harkonen PL, Valve EM, Ping W, Nurmi M, Martikainen PM. Hormone regulationof human prostate in organ culture. Can Resdearch 1993;53:5199–5207.
35. Jarred R, Cancilla B, Prins GS, Thayer K, Cunha GR, Risbridger G. Evidence that estrogens directlyalter androgen-regulated prostate development. Endocrinology 2000;141(9):3471–3477. [PubMed:10965920]
Prins and Korach Page 11
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

36. Nebert DW, Adesnik M, Coon MJ, Estabrook RW, Gonzalez FJ, Guengerich FP, Gunsalus IC,Johnson EF, Kemper B, Levin Wl, et al. The P450 gene superfamily: recommended nomenclature.DNA 1987;6:1–11. [PubMed: 3829886]
37. Committee NRN. A unified nomenclature system for the nuclear receptor superfamily. Cell1999;97:161–163. [PubMed: 10219237]
38. Walter P, Green S, Greene G, Krust A, Bornert J-M, Jeltsch J-M, Staub A, Jensen E, Scrace G,Waterfield M, et al. Cloning of the human estrogen receptor cDNA. Proceedings of the NationalAcademy of Science, USA 1985;82:7889–7893.
39. Laudet, V.; Gronemeyer, H. The nuclear receptor factsbook. Academic Press; San Diego: 2002.40. White R, Lees JA, Needham M, Ham J, Parker M. Structural organization and expression of the mouse
estrogen receptor. Mol Endocrinol. Mol Endocrinol 1:735−744 1987;1:735–744.41. Miksicek RJ. Steroid receptor variants and their potential role in cancer. Sem Cancer Biol 1994;5:369–
379.42. Sluyser M. Mutations in the estrogen receptor gene. Hum Mutat 1995;6:97–103. [PubMed: 7581411]43. Kuiper GGJM, Enmak E, Pelto-Huikko M, Nilsson S, Gustaffson JA. Cloning of a novel estrogen
receptor expressed in rat prostate and ovary. Proc Natl Acac Sci USA 1996;93:5925–5930.44. Mosselman S, Polman J, Dijkema R. ER B: identification and characterization of a novel human
estrogen receptor. FEBS 1996;392:49–53.45. Leung YK, Mak P, Hassan S, Ho SM. Estrogen receptor (ER)-beta isoforms: a key to understanding
ER-beta signaling. Proceedings of the National Academy of Science 2006;103(35):13162–13167.46. Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, Blumberg B, Kastner P,
Mark M, Chambon P, et al. The nuclear receptor superfamily: the second decade. Cell. 83:835−839.Cell 1995;83:835–839. [PubMed: 8521507]
47. Aranda A, Pascual A. Nuclear hormone receptors and gene expression. Physiol Rev 2001;81:1269–1304. [PubMed: 11427696]
48. Ueda T, Bruchovsky N, Sadar MD. Activation of the androgen receptor N-terminal domain byinterleukin-6 via MAPK and STAT3 signal transduction pathways. J Biol Chem 2002;277:7076–7085. [PubMed: 11751884]
49. Katzenellenbogen BS, Montano MM, Ediger TR, Sun J, Ekena K, Lazennec G, Martini PG,McInerney EM, Delage-Mourroux R, Weis K, Katzenellenbogen JA. Estrogen receptors: selectiveligands, partners, and distinctive pharmacology. Recent Prog Horm Res 2000;55(163−193;discussion 194−165)
50. Delaunay F, Pettersson K, Tujague M, Gustafsson JA. Functional differences between the amino-terminal domains of estrogen receptors alpha and beta. Mol Pharmacol 2000;58:584–590. [PubMed:10953052]
51. Watanabe T, Inoue S, Ogawa S, I, shii Y, Hiroi H, Ikeda K, Orimo A, Muramatsu M. Agonist effectof tamoxifen is dependent on cell type, ERE-promoter context, and estrogen receptor subtype:Functional difference between estrogen receptors ? and ? Biochem Biophys Res Comm1997;236:140–145. [PubMed: 9223441]
52. Cowley SM, Parker MG. A comparison of transcriptional activation by ER alpha and ER betaJ. SteroidBiochem Mol Biol 1999;69:165–175.
53. Kuiper GGJM, Gustafsson J-A. The novel estrogen receptor-b subtype: potential role in the cell- andpromoter-specific actions of estrogens and anti-estrogens. FEBS Lett 1997;410:87–90. [PubMed:9247129]
54. Kim K, Thu N, Saville B, Safe S. Domains of estrogen receptor alpha (ERalpha) required for ERalpha/Sp1-mediated activation of GC-rich promoters by estrogens and antiestrogens in breast cancer cells.Mol Endocrinol 2003;17:804–817. [PubMed: 12576490]
55. Shiau AK, Barstad D, Radek JT, Meyers MJ, Nettles KW, Katzenellenbogen BS, KatzenellenbogenJA, Agard DA, Greene GL. Structural characterization of a subtype-selective ligand reveals a novelmode of estrogen receptor antagonism. Nature Structural Biology 2002;9(5)
56. Stoica A, Saceda Ma, A, Solomon HB, Fenster BD, Martin MB. Regulation of estrogen receptor-alpha gene expression by 1, 25-dihydroxyvitamin D in MCF-7 cells. Journal of Cell Biochemistry1999;75:640–651.
Prins and Korach Page 12
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

57. Kuiper GGJM, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson JA.Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptorsα and B. Endocrinology 1997;138(3):863–870. [PubMed: 9048584]
58. Kuiper GGJM, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT, van der Burg B,Gustaffsson J-A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β.Endocrinology 1998;139(10):4252–4263. [PubMed: 9751507]
59. Harris HA, Bapat AR, Gonder DS, Frail DE. The ligand binding profiles of estrogen receptors alphaand beta are species dependent. Steroids 2002;67:379–384. [PubMed: 11958794]
60. Harris HA, Katzenellenbogen JA, Katzenellenbogen BS. Characterization of the biological roles ofthe estrogen receptors, ERalpha and ERbeta, in estrogen target tissues in vivo through the use of anERalpha-selective ligand. Endocrinology 2002;143:4172–4177. [PubMed: 12399409]
61. Montano MM, Muller V, Trobaugh A, Katzenellenbogen BS. The carboxy-terminal F domain of thehuman estrogen receptor: role in the transcriptional activity of the receptor and the effectiveness ofantiestrogens as estrogen antagonists. Mol Endocrinol 1995;9:814–825. [PubMed: 7476965]
62. Koide A, Zhao C, Naganuma M, Abrams J, Deighton-Collins S, Skafar DF, Koide S. Identificationof Regions within the F Domain of the Human Estrogen Receptor {alpha} that Are Important forModulating Transactivation and Protein-Protein Interactions. Molecular Endocrinology2007;21:829–842. [PubMed: 17185393]
63. Gougeleta A, Mueller SO, Korach KS, Renoir JM. Oestrogen receptors pathways to oestrogenresponsive elements: The transactivation function-1 acts as the keystone of oestrogen receptor (ER)alpha-mediated transcriptional repression of ERalpha. Journal of Steroid Biochemistry andMolecular Biology 2007;104(3−5):110–122. [PubMed: 17478088]
64. Ignar-Trowbridge DM, Nelson KG, Bidwell MC, Curtis SW, Washburn TF, McLachlan JA, KorachKS. Coupling of Dual Signaling Pathways: Epidermal Growth Factor Action Involves the EstrogenReceptor. Proc Natl Aca Sci 1992;89:4658–4662.
65. Coleman KM, Smith CL. Intracellular signaling pathways: nongenomic actions of estrogens andligand-independent activation of estrogen receptors. Front Biosci 2001;6:D1379–1391. [PubMed:11578956]
66. Klotz DM, Hewitt SC, Ciana P, Raviscioni M, Lindzey JK, Foley J, Maggi A, DiAugustine RP,Korach KS. Requirement of estrogen receptor-alpha in insulin-like growth factor-1 (IGF-1)-induceduterine responses and in vivo evidence for IGF-1/estrogen receptor cross-talk. J Biol Chem2002;277:8531–8537. [PubMed: 11751931]
67. Klotz DM, Hewitt SC, Korach KS, Diaugustine RP. Activation of a uterine insulin-like growth factorI signaling pathway by clinical and environmental estrogens: requirement of estrogen receptor-alpha.Endocrinology 2000;141:3430–3439. [PubMed: 10965916]
68. Safe S, Kim K. Nuclear receptor-mediated transactivation through interaction with Sp proteins. ProgNucleic Acid Res Mol Biol 2004;77:1–36. [PubMed: 15196889]
69. Kushner PJ, Agard DA, Greene GL, Scanlan TS, Shiau AK, Uht RM, Webb P. Estrogen receptorpathways to AP-1. Journal of Steroid Biochemistry and Molecular Biology 2000;74:311–317.[PubMed: 11162939]
70. Schulze H, Claus S. Histological localization of estrogen receptors in normal and diseased humanprostates by immunocytochemistry. Prostate 1990;16:331–343. [PubMed: 1695369]
71. Schulze H, Barrack E. Immunocytochemical localization of estrogen receptors in the normal maleand female canine urinary tract and prostate. Endocrinology 1987;121(5):1773–1783. [PubMed:3665847]
72. Brenner RM, West N, McClellena M. Estrogen and progestin receptors in the reproductive tract ofmale and female primates. Biol Reprod 1990;42:11–19. [PubMed: 2178696]
73. Prins GS, Birch L. Neonatal estrogen exposure up-regulates estrogen receptor expression in thedeveloping and adult rat prostate lobes. Endocrinology 1997;138(5):1801–1809. [PubMed: 9112371]
74. Tsurusaki T, Aoki D, Kanetake H, Inoue S, Muramatsu M, Hishikawa Y, Koji T. Zone-dependentexpression of estrogen receptors alpha and beta in human benign prostatic hyperplasia. J Clin EndoMetab 2003;88(1333−1340)
Prins and Korach Page 13
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

75. Tilley WD, Horsfall DJ, Skinner JM, Henderson DW, Marshall VR. Effect of pubertal developmenton estrogen receptor levels and stromal morphology in the guinea pig prostate. Prostate 1989;15:195–210. [PubMed: 2678040]
76. Adams JY, Leav I, Lau KM, Ho SM, Pflueger SM. Expression of estrogen receptor beta in the fetal,neonatal, and prepubertal human prostate. Prostate 2002;52(1):69–81. [PubMed: 11992621]
77. Kruithof-Dekker IG, Tetu B, A JPJ, Van der Kwast TH. Elevated estrogen receptor expression inhuman prostatic stromal cells by androgen ablation therapy. J Urol 1996;156:1194–1197. [PubMed:8709345]
78. Tilley W, HJorsfall D, McGee M, Alderman J, Marshall V. Effects of ageing and hormonalmanipulations on the level of oestrogen receptors in the guinea-pig prostate. Journal of Endocrinology1987;112:139–144. [PubMed: 3819628]
79. Shapiro E, Huang H, Masch RJ, McFadden DE, Wilson EL, Wu XR. Immunolocalization of estrogenreceptor alpha and beta in human fetal prostate. J Urology 2005;174(5):2051–2053.
80. Kozak I, Bartsch W, Krieg M, Voight K. Nuclei of stroma: site of highest estrogen concentration inhuman benign prostatic hyperplasia. Prostate 1982;3:433–438. [PubMed: 6183651]
81. Risbridger GP, Wang H, Young P, Takeshi K, Wong YZ, Lubahn D, Gustafsson JA, Cunha G.Evidence that epithelial and mesenchymal estrogen receptor-α mediates effects of estrogen onprostatic epithelium. Developmental Biology 2001;229:432–444. [PubMed: 11150243]
82. Prins GS, Birch L, Couse JF, Choi I, Katzenellenbogen B, Korach KS. Estrogen imprinting of thedeveloping prostate gland in mediated through stromal estrogen receptor α: studies with αERKO andβERKO mice. Cancer Research 2001;61:6089–6097. [PubMed: 11507058]
83. Eddy EM, Washburn TF, Bunch DO, Goulding EH, Gladen BC, Lubahn DB, Korach KS. Targeteddisruption of the estrogen receptor gene in male mice causes alteration of spermatogenesis andinfertility. Endocrinology 1996;137(11):4796–4805. [PubMed: 8895349]
84. Donaldson KM, Tong SY, Washburn T, Lubahn DB, Eddy EM, Hutson JM, Korach KS.Morphometric study of the gubernaculum in male estrogen receptor mutant mice. J Andrology1996;17(2):91–95.
85. Li L, Chui R, Nakajima K, Oh BR, Au HC, Dahiya R. Frequent methylation of estrogen receptor inprostate cancer: correlation with tumor progression. Cancer Research 2000;60:702–706. [PubMed:10676656]
86. Lau KM, LaSpina M, Long J, Ho SM. Expression of estrogen receptor (ER)-α and ER-β in normaland malignant prostatic epithelial cells: regulation by methylation and involvement in growthregulation. Cancer Res 2000;60:3175–3182. [PubMed: 10866308]
87. Bonkhoff H, Fixemer T, I. H, Remberger K. Estrogen receptor in prostate cancer and premalignantlesions. Am J Pathol 1999;155(2):641–647. [PubMed: 10433957]
88. Leav I, Lau KM, Adams J, NcNeal J, Taplin M, Wang J, Singh H, Ho SM. Comparative studies ofthe estrogen receptors beta and alpha and the androgen receptor in normal human prostate gland,dysplasia, and in primary and metastatic carcinoma. Am J Pathol 2001;1:79–92. [PubMed: 11438457]
89. Hernandez J, Balic I, Johnson-Pais TL, Higgins BA, Torkko KC, Thompson I, Leach RJ. Associationbetween estrogen receptor alpha gene polymorphism and risk of prostate cancer in black men. JUrology 2006;175:523–527.
90. Tanaka Y, Sasaki M, Kaneuchi M, Shiina H, Igawa M, Dahiya R. Polymorphisms of estrogen receptoralpha in prostate cancer. Mol Carcinogenesis 2003;37:202–208.
91. Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, Fried G, Nordenskjold M,Gustafsson J. Human estrogen receptor β -gene structure, chromosomal localization, and expressionpattern. JCEM 1997;82(12):42584265.
92. Prins GS, Marmer M, Woodham C, Chang WY, Kuiper G, Gustafsson JA, Birch L. Estrogen receptor-β messenger ribonucleic acid ontogeny in the prostate of normal and neonatally estrogenized rats.Endocrinology 1998;139(3):874–883. [PubMed: 9492016]
93. Makela S, Strauss L, Kuiper G, Valve EM, Salmi S, Santti R, Gustasson JA. Differential expressionof estrogen receptors alpha and beta in adult rat accessory sex glands and lower urinary tract. MolCell Endocrinol 2000;164:109–116. [PubMed: 11026563]
94. Chang W, Prins G. Estrogen receptor-β: Implications for the prostate gland. Prostate 1999;40:115–124. [PubMed: 10386472]
Prins and Korach Page 14
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

95. Asano K, Maruyama S, Usui T, Fujimoto N. Regulation of estrogen receptor alpha and beta expressionby testosterone in the rat prostate gland. Endocrin Journal 2003;50(3):281–287.
96. Fixemer T, Remberger K, Bonkoff H. Differential expression of estrogen receptor beta in humanprostate tissue, premalignant changes and in primary, metastaic and recurrent prostaticadenocarcinoma. Prostate 2003;54:79–87. [PubMed: 12497580]
97. Fujimura T, Takahashi S, Urano T, Ogawa S, Ouchi Y, Kitamura T, Muramatsu M, Inoue S.Differential expression of estrogen receptor beta (ERbeta) and its C-terminal truncated splice variantERbetacx as prognostic predictors in human prostatic cancer. Biochem Biophys Res Commun2001;289(3):692–699. [PubMed: 11726203]
98. Hanstein B, Liu H, Yancisin M, Brown M. Functional Analysis of a Novel Estrogen Receptor BetaIsoform. Mol Endocrinol 1999;13:129–137. [PubMed: 9892018]
99. Maruyama K, Endoh H, Sasaki-Iwaoka H, Kanou H, Shimaya E, Hashimoto S, Kato S, KawashimaH. A novel isoform of rat estrogen receptor beta with 18 amino acid insertion in the ligand bindingdomain as a putative dominant negative regular of estrogen action. Biochem Biophjys Res Commun1998;246:142–147.
100. Petersen D, Tkalcevic G, Koza-Taylor P, Turi T, Brown T. Identification of Estrogen Receptor Beta2: A Functional Varient of Estrogen Receptor Beta Expressed in Normal Rat Tissues. Endocrinology1998;139:1082–1092. [PubMed: 9492041]
101. Imamov O, Morani A, Shim GJ, Omoto Y, Thulin-Andersson C, Warner M, Gustafsson JA. Estrogenreceptor beta regulates epithelial cellular differentiation in the mouse ventral prostate. Proceedingsof the National Academy of Science 2004;101:9375–9380.
102. Weihua Z, Makela S, Andersson LC, Salmi S, Saji S, Webster JI, Jensen EV, Nilsson S, Warner M,Gustafsson JA. A role for estrogen receptor beta in the regulation of growth of the ventral prostate.Proceedings of the National Academy of Science 2001;98(11):6330–6335.
103. Weihua Z, Warner M, Gustaffson J. Estrogen receptor beta in the prostate. Mol Cell Biol 2002;193:1–5.
104. Couse JF, Curtis-Hewitt S, Korach KS. Receptor null mice reveal contrasting roles for estrogenreceptor alpha and beta in reproductive tissues. J Steroid Biochem Mol Biol 2000;74(5):287–296.[PubMed: 11162937]
105. Dupont S, Krust A, Gansmuller A, Dierich A, Chambon P, Mark M. Effect of single and compoundknockouts of estrogen receptors alpha (ERalpha) and beta (ERbeta) on mouse reproductivephenotypes. Development 127(19):4277–4291. [PubMed: 10976058]200
106. McPherson SJ, Ellem SJ, Simpson ER, Patchev V, Fritzemeier KH, Risbridger GP. Essential rolefor estrogen receptor beta in stromal-epithelial regulation of prostatic hyperplasia. Endocrinology2007;148(2):566–574. [PubMed: 17068134]
107. McMurray RW. Estrogen, prolactin, and autoimmunity: actions and interactions. Internat'lImmunopharm 2001;1:995–1008.
108. Yellayi S, Teuscher C, Woods JA, Welsh TH Jr. Tung KS, Nakai M, Rosenfeld CS, Lubahn DB,Cooke PS. Normal development of thymus in male and female mice requires estrogen/estrogenreceptor-alpha signaling pathway. Endocrine 2000;12(3):207–213. [PubMed: 10963039]
109. Harris HA, Albert LM, Leathurby Y, Malamas MS, Mewshaw RE, Miller CP, Kharode YP, MarzolfJ, Komm BS, Winneker RC, et al. Evaluation of an estrogen receptor-beta agonist in animal modelsof human disease. Endocrinology 2003;144(10):4241–4249. [PubMed: 14500559]
110. DeMarzo AM, Marchi VL, Epstein JI, Nelson WG. Proliferative inflammatory atrophy of theprostate implications for prostatic cacinogenesis. American Journal of Pathology 1999;155(6):1985–1992. [PubMed: 10595928]
111. Montano M, Jaiswal A, Katzenellenbogen B. Transcriptional regulation of the human quinonereductase gene by antiestrogen-liganded estrogen receptor-α and estrogen receptor-α. Fertility andSterility 1998;273:25443–25449.
112. Nelson WG, DeWeese TL, DeMarzo AM. The diet, prostate inflammation, and the development ofprostate cancer. Cancer Metast Rev 2002;21(1):3–16.
113. Pasquali D, Rossi V, Esposito D, Abbondanza C, Puca GA, Bellastella A, Sinisi AA. Loss of estrogenreceptor beta expression in malignant human prostate cells in primary cultures and in prostate cancertissues. J Clin Endo Metab 2001;86:2051–2055.
Prins and Korach Page 15
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

114. Horvath LG, Henshall SM, Lee CS, Head DR, Quinn DI, Makela S, Delprado W, Golovsky D,Brenner PC, O'Neill G, et al. Frequent loss of estrogen receptor-beta expression in prostate cancer.Cancer Res 2001;61:5331–5335. [PubMed: 11454669]
115. Zhu X, Leav I, Leung Y, Wu M, Liu Q, Gao Y, McNeal JE, Ho SM. Dynamic regulation of estrogenreceptor-beta expression by DNA methylation during prostate cancer development and metastasis.Am J Pathol 2004;164(6):1883–1886. [PubMed: 15161624]
116. Zhang X, Leung YK, Ho SM. AP-2 regulates the transcription of estrogen receptor (ER)-beta byacting through a methylation hotspot of the 0N promoter in prostate cancer cells. Oncogene.2007Epub ahead of print(May 21.)
117. Cheng J, Lee EJ, Madison LD, Lazennec G. Expression of estrogen receptor beta in prostatecarcinoma cells inhibits invasion and proliferation and triggers apoptosis. FEBS Lett 2004;566:169–172. [PubMed: 15147889]
118. Lai JS, Brown LG, True LD, Hawley SJ, Etzioni RB, Higano CS, Ho SM, Vessella RL, Corey E.Metastases of prostate cancer express estrogen receptor-beta. Urology 2004;64:814–820. [PubMed:15491740]
119. Wilson J. The pathogenesis of benign prostatic hyperplasia. Am J Med 1980;68:745–756. [PubMed:6155068]
120. Guerini V, Sau D, Scaccianoce E, Rusmini P, Ciana P, Maggi A, Martini PG, Katzenellenbogen BS,Martini L, Motta M, et al. The androgen derivative 5alpha-androstane-3beta, 17beta-diol inhibitsprostate cancer cell migration through activation of the estrogen receptor beta subtype. Cancer Res2005;65(12):5445–5453. [PubMed: 15958594]
121. Garcia GE, Wisniewski HG, Lucia MS, Arevalo N, Slaga TJ, Kraft SL, Strange R, Kumar AP. 2-Methoxyestradiol inhibits prostate tumor development in transgenic adenocarcinoma of mouseprostate: role of tumor necrosis factor-alpha-stimulated gene 6. Clin Cancer Research 2006;12:980–988.
122. Schulz P, Bauer HW, Brade WP, Keller A, Fittler F. Evaluation of the cytotoxic activity ofdiethylstilbestrol and its mono- and diphosphate towards prostatic carcinoma cells. Cancer Research1988;48(2867−2870)
123. Kim IY, Kim BC, Seong, Lee DK, Seo JM, Hong YJ, Kim HT, Morton RA, Kim SJ. Raloxifene, amixed estrogen agonist/antagonist, induces apoptosis in androgen-independent human prostatecancer cell lines. Cancer Res 2002;62:5365–5369. [PubMed: 12235008]
124. Neubauer BL, McNulty AM, Chedid M, Chen K, Goode RL, Johnson MA, Jones CD, Krishnan V,Lynch R, Osborne HE, et al. The selective estrogen receptor modulator trioxifene (LY133314)inhibits metastasis and extends survival in the PAIII rat prostatic carcinoma model. Cancer Res2003;63:6056–6062. [PubMed: 14522935]
125. Corey E, Quinn JE, Emond MJ, Buhler KR, Brown LG, Vessella RL. Inhibition of androgen-independent growth of prostate cancer xenografts by 17beta-estradiol. Clin Cancer Research 2002;8(4):1003–1007.
126. Raghow S, Hooshdraran MZ, Katiyar S, Steiner MS. Toremifene prevents prostate cancer in thetransgenic adenocarcinoma of mouse prostate model. Cancer Research 2002;62:1370–1376.[PubMed: 11888907]
127. Bergan RC, Reed E, Myers CE, Headlee D, Brawley O, Cho HK, Figg WD, Tompkins A, LinehanWM, Kohler D, et al. A Phase II study of high-dose tamoxifen in patients with hormone-refractoryprostate cancer. Clin Cancer Research 1999;5(9):2366–2373.
128. Stein S, Zoltick B, Peacock T, Holroyde C, Haller D, Armstead B, Malkowicz SB, Vaughn DJ.Phase II trial of toremifene in androgen-independent prostate cancer: a Penn cancer clinical trialsgroup trial. Am J Clin Oncol 2001;24(3):283–285. [PubMed: 11404501]
129. Steiner MS, Pound CR. Phase IIA clinical trials to test the efficacy and safety of Toremifene in menwith high-grade prostatic intraepithelial neoplasia. Clin Prostate Cancer 2003;2(1):32–33.[PubMed: 15046681]
130. Ellem SJ, Risbridger GP. Aromatase and prostate cancer. Minerva Endocrinol 2006;31:1–12.[PubMed: 16498360]
131. Narashimamurthy J, Rao AR, Sastry GN. Aromatase inhibitors: a new paradigm in breast cancertreatment. Curr Med Chem Anticancer Agents 2004;4(6):523–534. [PubMed: 15579017]
Prins and Korach Page 16
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

132. Ellem SJ, Schmitt JF, Pedersen JS, Frydenberg M, Risbridger GP. Local aromatase expression inhuman prostate is altered in malignancy. J Clin Endocrinol Metab 2004;89(5):2431–2441.
133. Smith MR, Kaufman D, George D, Oh WK, Kazanis M, Manola J, Kantoff PW. Selective aromataseinhibition for patients with androgen-independent prostate carcinoma. Cancer 2002;95(9):1864–1868. [PubMed: 12404279]
134. Santen RJ, Petroni GR, Fisch MJ, Myers CE, Theodorescu D, Cohen RB. Use of the aromataseinhibitor anastrozole in the treatment of patients with advanced prostate carcinoma. Cancer 2001;92(8):2095–2101. [PubMed: 11596025]
135. Wang J, Eltoum IE, Lamartiniere CA. Dietary genistein suppresses chemically induced prostatecancer in Lobund-Wistar rats. Cancer Letters 2002;186(11−18)
136. Mentor-Marcel R, Lamartiniere CA, Eltoum A, Greenberg N, M. Elgavish A. Dietary genisteinimproves survival and reduces expression of osteopontin in the prostate of transgenic mice withprostatic adenocarcinoma (TRAMP). J Nutr 2005;135(5):989–995. [PubMed: 15867270]
137. Jones SB, DePrimo SE, Whitfield ML, Brooks JD. Resveratrol-induced gene expression profiles inhuman prostate cancer cells. Cancer Epidemiol Biomarkers Prev 2005;14:596–604. [PubMed:15767336]
138. Lee MM, Gomez SL, Chang JS, Wey M, Wang RT, Hsing AW. Soy and isoflavone consumptionin relation to prostate cancer risk in China. Cancer Epidemiol Biomarkers Prev 2003;12:665–668.[PubMed: 12869409]
139. Ozasa K, Nakao M, Watanabe Y, Hayashi K, Miki T, Mikami K, Mori M, Sakauchi F, Washio M,Ito Y, et al. Serum phytoestrogens and prostate cancer risk in a nested case-control study amongJapanese men. Cancer Sci 2004;95(1):65–71. [PubMed: 14720329]
140. Manson MM, Farmer PB, Gescher A, Steward WP. Innovative agents in cancer prevention. RecResults Cancer Res 2005;166:257–275.
141. Mak P, Leung YK, Tang WY, Harwood C, Ho SM. Apigenin suppresses cancer cell growth throughERbeta. Neoplasia 2006;8(11):896–904. [PubMed: 17132221]
Prins and Korach Page 17
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
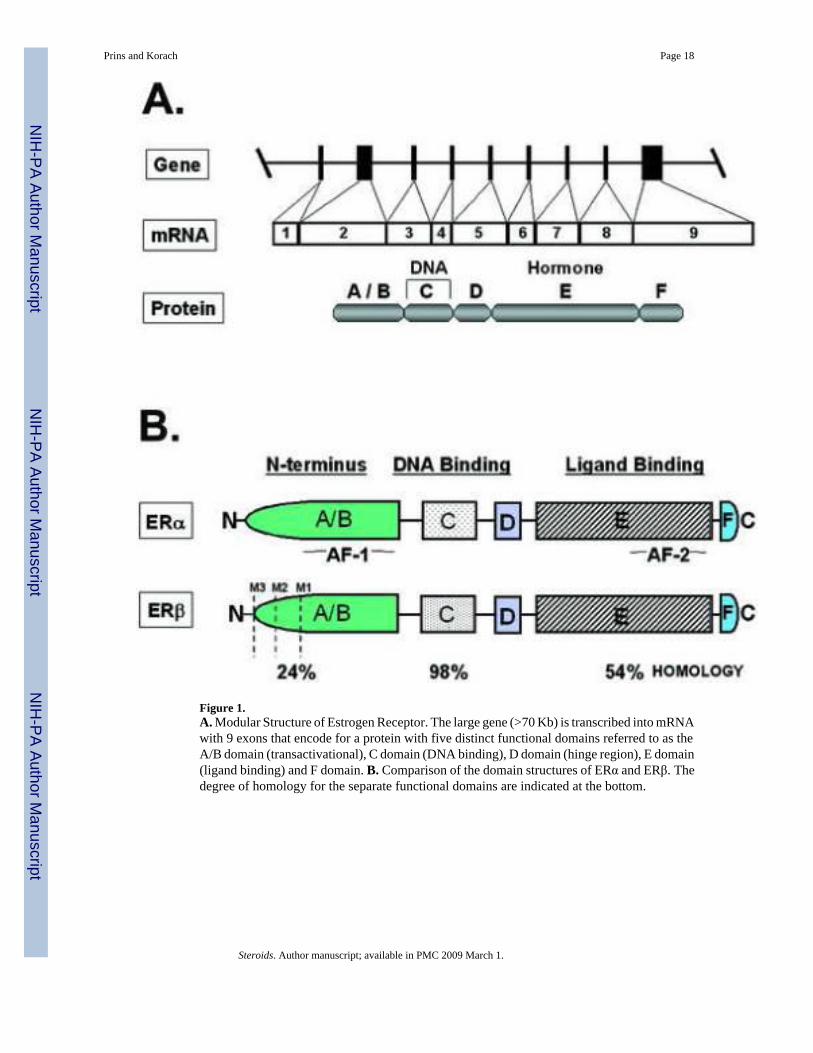
Figure 1.A. Modular Structure of Estrogen Receptor. The large gene (>70 Kb) is transcribed into mRNAwith 9 exons that encode for a protein with five distinct functional domains referred to as theA/B domain (transactivational), C domain (DNA binding), D domain (hinge region), E domain(ligand binding) and F domain. B. Comparison of the domain structures of ERα and ERβ. Thedegree of homology for the separate functional domains are indicated at the bottom.
Prins and Korach Page 18
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2.A. Incidence of wild-type and βERKO mice presenting with T-cell infiltration within theprostatic complex of at one year of age. T-cell infiltration throughout the prostatic complexwas blindly scored as 0 (absent), 1 (rare), 2 (focal), 3 (abundant) and 4 (massive) for the twogenotypes. Overall incidences between the genotypes was shown to be significant at P < 0.005using a Chi-square test for trend. B. Inflammatory cell infiltration in the ventral prostate lobesof wild-type (with 0 and 2 Score) and βERKO (with 3 and 4 Score) mice at 1 year of age. Wild-type prostates were either free of lymphoid infiltration (0) or contained rare or focal (2) T-cellinfiltration. In contrast, focal, abundant (3) and massive (4) stromal lymphocytic infiltrate wereroutinely observed in the βERKO prostates. As infiltration became abundant, evidence for
Prins and Korach Page 19
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

diapadesis in the epithelium was observed (arrows). In several instances, lymphoid aggregateswere associated with reactive, proliferative epithelium in the immediate vicinity (arrowheads).Magnification = 40×.
Prins and Korach Page 20
Steroids. Author manuscript; available in PMC 2009 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Prins and Korach Page 21
Table 1Comparison of ERα and ERβ expression and activities in the prostate gland
ER α ER βLocalization Stromal EpithelialProliferation Epithelial squamous metaplasia Anti-proliferative
Stromal proliferationDifferentiation Epithelial Dysplasia Pro-differentiationImmune Response Anti-inflammatory
Anti-oxidantExpression in PCa Dysregulated in PCa Dysregulated in PCa
- Silenced in early cancers - ↓ organ confined disease - Re-emergence with progression - ↑ in metastatic PCa
Steroids. Author manuscript; available in PMC 2009 March 1.
Copyright © 2022 FDOKUMEN




















