
Functional domains of the human estrogen receptor
11
Cell. Vol. 51, 941-951. December 24, 1987, Copyright 0 1987 by Cell Press Functional Domains of the Human Estrogen Receptor Vijay Kumar, Stephen Green, Gary Stack, Meera Berry, Jia-Rui Jin, and Pierre Chambon Laboratoire de Genetique Moleculaire des Eucaryotes du CNRS Unite 184 de Biologie Moleculaire et de Genie Genetique de I’ INSERM lnstitut de Chimie Biologique, Faculte de Medecine 11, Rue Humann 67085 Strasbourg Cedex, France Summary Two domains of the human estrogen receptor, respon- sible for hormone binding (region E) and tight nuclear binding (region C), are essential for the receptor to ac- tivate efficiently the transcription of estrogen-respon- sive genes. Region D, which joins the DNA- and hor- mone-binding domains, can be altered without affecting activation. Deletion of the N-terminal domain (region A/B) has no effect on activation of a reporter gene containing a vitellogenin estrogen-responsive element (ERE) and the HSV-tk promoter, whereas it severely im- pairs activation of the human pS2 gene promoter. De- letion of most or all of the hormone-binding domain leads to only about 5% constitutive transcriptional ac- tivity, yet these mutants appear to bind efficiently to an ERE in vivo. Apparently, region C recognizes the ERE of target genes, and the hormone-binding domain plays an essential role for efficient activation of tran- scription. Introduction The process by which gene transcription is regulated dur- ing eukaryotic development involves interaction of tfans- acting factors with specific &-acting elements located within and around genes. Steroid hormone receptors rep- resent a class of such trans-acting factors that stimulate transcription by binding to specific DNA elements termed hormone response elements (HRE). It has been demon- strated in the case of glucocorticoid-responsive (for re- view, see Yamamoto, 1985) and estrogen-responsive genes (Klein-Hitpass et al., 1986; Seiler-Tuyns et al., 1986; our unpublished data for human pS2 gene) that such ele- ments have the properties of enhancers, in that they exert their action in an orientation-independent manner when placed at variable distances upstream or downstream from homologous or heterologous promoter elements. In the case of steroid receptors, the stimulation of gene transcription is strictly dependent upon the receptor bind- ing to its cognate hormone, and therefore these receptors represent inducible enhancer factors. Steroid hormone * Present address: New Haven Hospital, Department of Clinical Pathology, 20 York Street, Yale, Connecticut 06504. receptors thus provide a useful model system to study the mechanism by which enhancers work. A human estrogen receptor (ER) cDNA sequence con- taining the complete open reading frame (Walter et al., 1985; Green et al., 1986) when expressed in the ER- negative human cervical cell line HeLa produces a 66 kd protein (Green et al., 1986) that binds estradiol with wild- type affinity (Kd = 0.5 nM) and associates tightly with the nucleus only in the presence of hormone (Kumar et al., 1986). Furthermore, cotransfection of the wild-type ER cDNA expression-vector HE0 with an estradiol-responsive reporter gene demonstrated the functionality of the cloned receptor in activating gene transcription (Druege et al., 1986; Green and Chambon, 1987). A comparison of the chicken and human estrogen receptors (ERs) (Krust et al., 1986) revealed two regions, termed C and E, that are not only highly conserved between chicken and man (100% and 94% amino acid identity, respectively; see Fig- ure 4) but are also conserved between the different ste- roid and related nonsteroid nuclear receptors (for reviews, see Green and Chambon, 1986, and Gronemeyer et al., 1987a). Such a high degree of conservation suggests that these regions perform important functions that are com- mon to all of these receptors. To assess these functions, we constructed a series of human ER deletion mutants using site-directed mutagenesis (Kumar et al., 1986). Tran- sient expression of these mutants in HeLa cells, followed by measurement of hormone binding, showed that region E corresponds to the hormone-binding domain (Kumar et al., 1986). That region C may correspond to the DNA- binding domain was indicated by its absolute requirement to achieve tight nuclear binding. This putative DNA- binding domain is characterized by its high content of cys- teine, lysine, and arginine. Moreover, the general structure of this region is highly conserved among other members of the nuclear receptor family (see Green and Chambon, 1987, and Gronemeyer et al., 1987a, for reviews; and Arriza et al., 1987) and may fold into a structure similar to the DNA-binding “fingers” proposed for eukaryotic transcrip- tion factor TFIIIA (Weinberger et al., 1985 Krust et al., 1986). Hydrophilic region D, which is conserved neither in sequence nor in length among the different nuclear re- ceptors and is only poorly conserved for the ER from different species (Krust et al., 1986; Koike et al., 1987; Weiler et al., 1987; for review, see Gronemeyer et al., 1987a), may act as a hinge (Kumar et al., 1986) between the DNA- and hormone-binding domains. Besides the DNA- and hormone-binding domains, it is speculated that a third domain exists that modulates gene transcription, possibly by interaction with other transcrip- tion factors. We have investigated in our study whether both the DNA- and hormone-binding domains are essen- tial for the receptor to stimulate the transcription of estradiol-inducible genes, and have attempted to localize the putative domain(s) responsible for the activation of gene transcription.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Functional domains of the human estrogen receptor

Cell. Vol. 51, 941-951. December 24, 1987, Copyright 0 1987 by Cell Press
Functional Domains of the Human Estrogen Receptor
Vijay Kumar, Stephen Green, Gary Stack, Meera Berry, Jia-Rui Jin, and Pierre Chambon Laboratoire de Genetique Moleculaire des Eucaryotes du CNRS Unite 184 de Biologie Moleculaire et de Genie Genetique de I’INSERM lnstitut de Chimie Biologique, Faculte de Medecine 11, Rue Humann 67085 Strasbourg Cedex, France
Summary
Two domains of the human estrogen receptor, respon- sible for hormone binding (region E) and tight nuclear binding (region C), are essential for the receptor to ac- tivate efficiently the transcription of estrogen-respon- sive genes. Region D, which joins the DNA- and hor- mone-binding domains, can be altered without affecting activation. Deletion of the N-terminal domain (region A/B) has no effect on activation of a reporter gene containing a vitellogenin estrogen-responsive element (ERE) and the HSV-tk promoter, whereas it severely im- pairs activation of the human pS2 gene promoter. De- letion of most or all of the hormone-binding domain leads to only about 5% constitutive transcriptional ac- tivity, yet these mutants appear to bind efficiently to an ERE in vivo. Apparently, region C recognizes the ERE of target genes, and the hormone-binding domain plays an essential role for efficient activation of tran- scription.
Introduction
The process by which gene transcription is regulated dur- ing eukaryotic development involves interaction of tfans- acting factors with specific &-acting elements located within and around genes. Steroid hormone receptors rep- resent a class of such trans-acting factors that stimulate transcription by binding to specific DNA elements termed hormone response elements (HRE). It has been demon- strated in the case of glucocorticoid-responsive (for re- view, see Yamamoto, 1985) and estrogen-responsive genes (Klein-Hitpass et al., 1986; Seiler-Tuyns et al., 1986; our unpublished data for human pS2 gene) that such ele- ments have the properties of enhancers, in that they exert their action in an orientation-independent manner when placed at variable distances upstream or downstream from homologous or heterologous promoter elements. In the case of steroid receptors, the stimulation of gene transcription is strictly dependent upon the receptor bind- ing to its cognate hormone, and therefore these receptors represent inducible enhancer factors. Steroid hormone
* Present address: New Haven Hospital, Department of Clinical Pathology, 20 York Street, Yale, Connecticut 06504.
receptors thus provide a useful model system to study the mechanism by which enhancers work.
A human estrogen receptor (ER) cDNA sequence con- taining the complete open reading frame (Walter et al., 1985; Green et al., 1986) when expressed in the ER- negative human cervical cell line HeLa produces a 66 kd protein (Green et al., 1986) that binds estradiol with wild- type affinity (Kd = 0.5 nM) and associates tightly with the nucleus only in the presence of hormone (Kumar et al., 1986). Furthermore, cotransfection of the wild-type ER cDNA expression-vector HE0 with an estradiol-responsive reporter gene demonstrated the functionality of the cloned receptor in activating gene transcription (Druege et al., 1986; Green and Chambon, 1987). A comparison of the chicken and human estrogen receptors (ERs) (Krust et al., 1986) revealed two regions, termed C and E, that are not only highly conserved between chicken and man (100% and 94% amino acid identity, respectively; see Fig- ure 4) but are also conserved between the different ste- roid and related nonsteroid nuclear receptors (for reviews, see Green and Chambon, 1986, and Gronemeyer et al., 1987a). Such a high degree of conservation suggests that these regions perform important functions that are com- mon to all of these receptors. To assess these functions, we constructed a series of human ER deletion mutants using site-directed mutagenesis (Kumar et al., 1986). Tran- sient expression of these mutants in HeLa cells, followed by measurement of hormone binding, showed that region E corresponds to the hormone-binding domain (Kumar et al., 1986). That region C may correspond to the DNA- binding domain was indicated by its absolute requirement to achieve tight nuclear binding. This putative DNA- binding domain is characterized by its high content of cys- teine, lysine, and arginine. Moreover, the general structure of this region is highly conserved among other members of the nuclear receptor family (see Green and Chambon, 1987, and Gronemeyer et al., 1987a, for reviews; and Arriza et al., 1987) and may fold into a structure similar to the DNA-binding “fingers” proposed for eukaryotic transcrip- tion factor TFIIIA (Weinberger et al., 1985 Krust et al., 1986). Hydrophilic region D, which is conserved neither in sequence nor in length among the different nuclear re- ceptors and is only poorly conserved for the ER from different species (Krust et al., 1986; Koike et al., 1987; Weiler et al., 1987; for review, see Gronemeyer et al., 1987a), may act as a hinge (Kumar et al., 1986) between the DNA- and hormone-binding domains.
Besides the DNA- and hormone-binding domains, it is speculated that a third domain exists that modulates gene transcription, possibly by interaction with other transcrip- tion factors. We have investigated in our study whether both the DNA- and hormone-binding domains are essen- tial for the receptor to stimulate the transcription of estradiol-inducible genes, and have attempted to localize the putative domain(s) responsible for the activation of gene transcription.
Cell 942
vit-tk-CAT
pS2-CAT
vit-tk-Globin
pS2-Globin
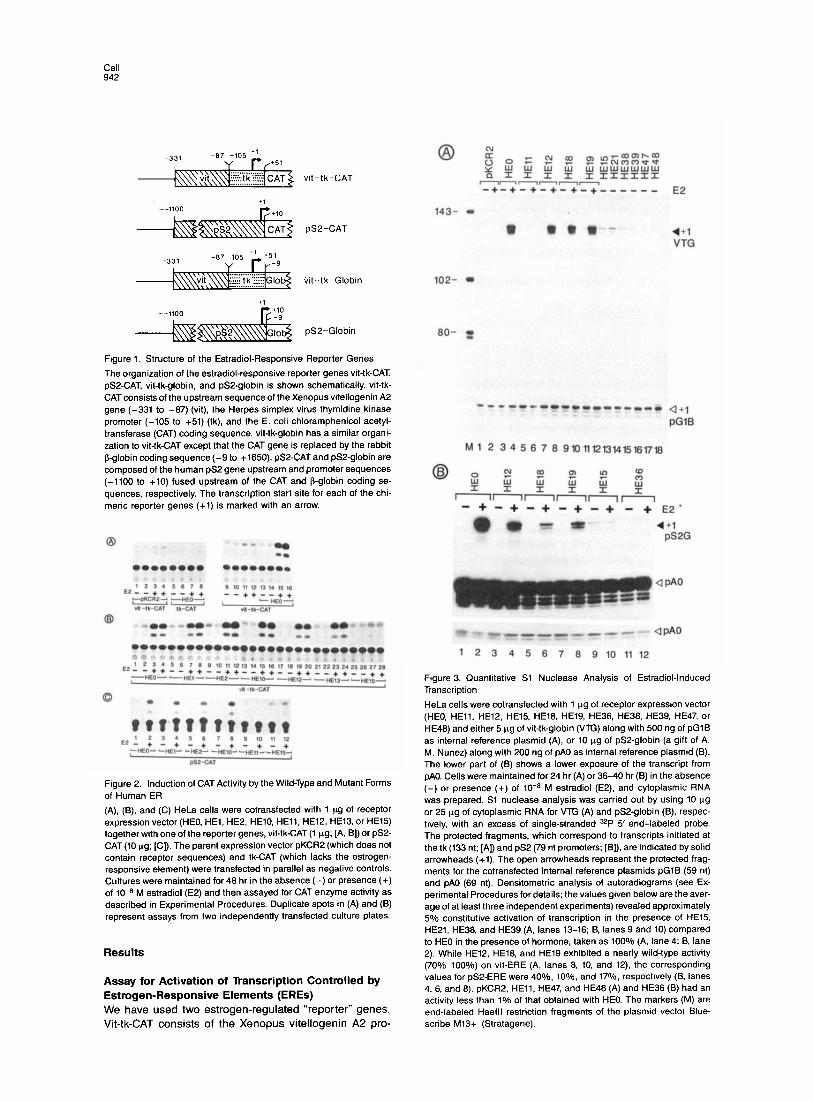
Figure 1. Structure of the Estradiol-Responsive Reporter Genes
The organization of the estradiol-responsive reporter genes vit-tk-CAT, pSP-CAT, vit-tk-globin. and pS2-globin is shown schematically. vit-tk- CAT consists of the upstream sequence of the Xenopus vitellogenin A2 gene (-331 to -87) (vit), the Herpes simplex virus thymidine kinase promoter (-105 to +51) (tk), and the E. coli chloramphenicol acetyl- transferase (CAT) coding sequence. vit-tk-globin has a similar organi- zation to vit-tk-CAT except that the CAT gene is replaced by the rabbit 6-globin coding sequence (-9 to +1650). pSP-CAT and pS2-globin are composed of the human pS2 gene upstream and promoter sequences (-1100 to +10) fused upstream of the CAT and 8-globin coding se- quences, respectively. The transcription start site for each of the chi- merit reporter genes (+l) is marked with an arrow.
Figure 2. Induction of CAT Activity by the Wild-Type and Mutant Forms of Human ER
(A), (B), and (C) HeLa cells were cotransfected with 1 trg of receptor expression vector (HEO, HEl, HE2, HEIO, HEll, HE12, HE13, or HE15) together with one of the reporter genes, vit-tk-CAT (1 ug; [A, B]) or pS2- CAT (10 ug; [Cl). The parent expression vector pKCR2 (which does not contain receptor sequences) and tk-CAT (which lacks the estrogen- responsive element) were transfected in parallel as negative controls. Cultures were maintained for 48 hr in the absence (-) or presence (+) of 10-s M estradiol (E2) and then assayed for CAT enzyme activity as described in Experimental Procedures. Duplicate spots in (A) and (B) represent assays from two independently transfected culture plates.
Results
Assay for Activation of Transcription Controlled by Estrogen-Responsive Elements (EREs) We have used two estrogen-regulated “reporter” genes. Vit-tk-CAT consists of the Xenopus vitellogenin A2 pro-
-I---- -+-+-+-+-+-+ -m-m -- E2
143- *
a a a a-- 4+1 VTG
102- *
80- t
M 1 2 3 4 5 6 7 8 9lOll12131415161718
-+-+-+-+-+ -+ E2
4+1 pS2G
Q PAO
I 2 3 4 5 6 7 8 9 10 11 12
Figure 3. Quantitative Sl Nuclease Analysis of Estradiol-Induced Transcription
HeLa cells were cotransfected with 1 trg of receptor expression vector (HEO, HEll, HE12. HE15. HE18, HE19, HE36, HE38, HE39. HE47, or HE48) and either 5 Fg of vit-tk-globin (VTG) along with 500 ng of pGlB as internal reference plasmid (A), or IO vg of pS2-globin (a gift of A. M. Nunez) along with 200 ng of pA0 as internal reference plasmid (B). The lower part of (B) shows a lower exposure of the transcript from pA0. Cells were maintained for 24 hr (A) or 36-40 hr (B) in the absence (-) or presence (+) of lo-* M estradiol (E2), and cytoplasmic RNA was prepared. Sl nuclease analysis was carried out by using 10 vg or 25 ug of cytoplasmic RNA for VTG (A) and pS2-globin (B), respec- tively, with an excess of single-stranded 32P 5’ end-labeled probe. The protected fragments, which correspond to transcripts initiated at the tk (133 nt; [A]) and pS2 (79 nt promoters; [B]), are indicated by solid arrowheads (+l). The open arrowheads represent the protected frag- ments for the cotransfected internal reference plasmids pGlB (59 nt) and pA0 (69 nt). Densitometric analysis of autoradiograms (see Ex- perimental Procedures for details; the values given below are the aver- age of at least three independent experiments) revealed approximately 5% constitutive activation of transcription in the presence of HE15, HE21, HE38, and HE39 (A, lanes 13-16; B, lanes 9 and 10) compared to HE0 in the presence of hormone, taken as 100% (A, lane 4; B, lane 2). While HE12, HE18. and HE19 exhibited a nearly wild-type activity (70%-100%) on vit-ERE (A, lanes 8, 10, and 12) the corresponding values for pSP-ERE were 40%, IO%, and 17%, respectively (B, lanes 4,6. and 8). pKCR2, HEll, HE47, and HE48 (A) and HE36 (B) had an activity less than 1% of that obtained with HEO. The markers (M) are end-labeled Haelll restriction fragments of the plasmid vector Blue- scribe M13+ (Stratagene).

Functional Domains of the Human Estrogen Receptor 943
Table 1. Relative Ability of Human ER Mutants to Stimulate Expression of Vit-tk-CAT
Deleted
Relative CAT Activity (a/o)
Amount of Reporter Plasmid (Kg)
Mutant Amino Acids 0.01 0.1 1.0 5.0
HE0 100 100 100 100 HE1 (9-62) 116[97-1521 (4) 107 103[65-1491 (5) HE2 (67-131) 130[95-1931 (3) 75 116[66-148) (6) HE3 (132-198) 2 (O-41 (2) HE4 (199-264) 0 (2) 4 HE5 (265-330) - 0 (2) 0 HE6 (331-396) 0 0 0 (3) 0 HE7 (397-462) 5 (2-71 (2) 1 HE6 (463-528) 0 (2) 1 HE9 (527-588) 0 (2) 1 HE10 (141-170) 103[ 16-l 721 (4) 59 99 173-1261 (4) HE1 1 (185-251) 0 0 (2) 0 (4) 0 HE12 (271-300) 76[30-1001 (3) 105 120[75-1941 (8) HE13 (553-595) 190[42-3031 (3) 93[54-1541 (5) HE14 (l-281) 0 (2) 2 HE15 (282-595) 1312-231 (4) 1P-11 (2) 4 (O-41 (4) 712-l l] (3) HE16 (262-595) 6 0 311-41 (2) HE17 (251-595) - 6
;10]190-2241 (3) 2 PI (2)
HE18 (l-139) 35119-503 (2) 53 90 HE19 (1-178) 75[56-941 (2) 54 137[103-1891 (3) 115 HE20 (1-184 and
251-595) 2 0 HE21 (341-595) 3 8[3-121 (2) HE22 NA 52 129 93 97 HE23 NA 82 104 119 84 HE24 NA 44 104 95 81 HE25 NA 40 60 121 114
The table is a compilation of data from 19 independent experiments in each of which a subset of mutants was compared to HEO. In each case, duplicate plates of HeLa cells were cotransfected with 1 ng of vit-tk-CAT and four different amounts of receptor expression plasmid. Cells were cultured for 48 hr in the presence of 10-s M estradiol, and CAT activity was measured. The values are expressed as the percentage of CAT activi- ty obtained with the wild-type ER, HEO. The range of values is indicated in brackets and the number of independent observations is shown in parentheses. At least two different plasmid preparations for each construction were tested in this study. The positions of the deleted amino acids are indicated in parentheses at the left. NA. not applicable.
moter upstream region (-331 to -87, which contains an ERE) fused to the Herpes simplex virus thymidine kinase (tk) promoter (-105 to +51), downstream of which lies the E. coli chloramphenicol acetyltransferase (CAT) gene (Klein-Hitpass et al., 1986; Figure 1). pSBCAT was con- structed from the human estrogen-responsive gene pS2 (Masiakowski et al., 1982; Brown et al., 1984; Jeltsch et al., 1987) by insertion of the ERE-containing promoter region (m-1100 to +lO) upstream of the CAT gene. The CAT gene was also replaced by a promoterless 6-globin gene (see Figure 1) to allow quantitative Sl nuclease RNA de- terminations. Transcriptional activation of these reporter genes was analyzed in transient assays by transfection into HeLa cells together with vectors expressing the wild- type or mutated human ER.
Stimulation of gene transcription from the vit-tk reporter genes was dependent upon the presence of HE0 and the vitellogenin A2 gene ERE (Figures 2A and 3A). Similar results were obtained when the pS2 reporter genes were used (Figure 2C and Figure 38, and data not shown). Half- maximal stimulation of vit-tk-CAT required approximately 200 pM estradiol (data not shown), and all assays were performed with 10e8 M estradiol. When 1 wg of vit-tk-CAT or 10 ug of pSBCAT was used, maximal activity was ob- tained with approximately 100 ng and 250 ng, respec-
tively, of the human ER expression vector HE0 (data not shown).
Region C, Essential for Activation of Transcription, Determines the Specific Recognition of the Responsive Element The deletion mutants HE3 and HE4, which lack part of re- gion C, were unable to activate gene transcription (Table 1 and Figure 4). Furthermore, HEll, which lacks the core of region C, failed to stimulate transcription of either the vitellogenin or pSBCAT reporter genes (Tables 1 and 2 and Figure 2C, lane 10). The stimulatory activity of HE11 was less than 1% of that of HEO, as determined by Sl nuclease analysis of RNA synthesized from the vit-tk- globin reporter gene (Figure 3A, lanes 3-6). Thus the 66 amino acid-long highly conserved core from region C is essential for activation of transcription.
To demonstrate that ER region C is responsible for the specific recognition of the responsive element of an estro- gen target gene, we performed a region C exchange ex- periment, analogous to that which allowed us to establish that region C of the human glucocorticoid receptor (GR) determines the specific recognition of the responsive ele- ment of the glucocorticoid-inducible MMTV-CAT reporter gene (Green and Chambon, 1987). Region C cassettes,

Cl?ll 944
..__ HE9 HE9 HE10 HE10 “El, HE,, HE12 HE12 HE14 HE14 HE13 HE13 HE21 HE21 HE.39 HE39 HE36 HE38 HElri HE15 HE16 HE16 HE17 HE17 HE47 HE47 HE46 HE48 HE36 HE36 HE16 HE36 HE19 HE20
HE19 HE20
+ +
+ +
Table 2. Relative Ability of Human ER Mutants to Stimulate Expression of pSP-CAT
Receptor Mutant Relative CAT Activity (%)
HE0 100 HE1 115 [80-1501 (2) HE2 59 [55-681 (3) HE10 118 [70-l 671 (2) HE11 0 (3) HE12 37 [28-431 (5) HE13 84 (1) HE15 6 [3-lo] (7) HEld 9 [6-l l] (3) HE19 17 [l l-251 (6) HE21 4 [4.3-4.51 (2) Duplicate plates of HeLa cells were cotransfected with pSP-CAT (10 pg) and receptor expression vector (1 frg). Culture conditions and CAT assays were performed as described in Table 1, and CAT activity is expressed as the percentage of that obtained with HEO. The range of values is indicated in brackets and the number of experiments per- formed is shown in parentheses.
corresponding to the 66 amino acid core region, were created in both the human ER (ERCAS) and GR (GRCAS) by using site-directed mutagenesis to create HE63 and HG7 (Figure 5) which stimulated transcription to the same extent as the wild-type ER and GR (data not shown). The cassettes were exchanged between the two receptors. The ER chimera ER-GRCASl bound es-
Vit ERE ps2 m,: --
: +
+
+
+ -10%
-5% -5 % -5%
+ +
+ +
t + +
Figure 4. Human ER Deletion and Insertion Mutants
The human (hER) sequence has been divided into six regions (A-F) based on its homology with the chicken estrogen receptor (cER), and the percentage of amino acid identity in each region is indicated in parentheses at the top (see Krust et al., 1986, for further details). The N terminus of the 595 amino acid long human ER is positioned at the left. The human ER de- letion mutants (HE1 to HE21, HE38, HE39, HE47, and HE48) and insertion mutants (HE22 to HE25, HE55, and HE56) (see also Table 1 and Experimental Procedures) are shown sche- matically below the hERIcER comparison. A solid black bar represents the portion of human ER retained in the deletion mutants, with the deleted portion shown as a gap. The insertion mutants are represented by the number of amino acids inserted between residues 281 and 282. The thin vertical lines represent the boundaries of the six regions (A-F); the dotted lines, the position of the 66 amino acid core of region C. The table at the right depicts the abil- ity of the receptor mutants to bind estradiol (cy- toplasmic), to be tightly bound to the nucleus in the presence of estradiol (nuclear), and to activate transcription of either the vitellogenin (Vit ERE) or pS2 (pS2 ERE) reporter genes when expressed transiently in HeLa cells. A plus sign represents wild-type activity and a mi- nus sign means no activity. Values that fall between these two extremes are indicated as percentage of wild-type activity and are repre- sentative of values obtained by quantitative Sl nuclear anaiyses, similar to those shown in Fig- ure 3. NA, not applicable: ND, not determined.
tradiol, while the GR chimera GR-ER.CASl bound the synthetic glucocorticoid dexamethasone (data not shown). Transfections were performed in HeLa cells (which con- tain very low levels of transcriptionally active endogenous GR; see transfections with the parent vector pKCR2 in Fig- ure 5, lanes l-3) by using either the MMTV-CAT or vit-tk- CAT reporter genes. Stimulation of vit-tk-CAT was seen only with HE0 in the presence of estradiol (Figure 5, lanes 23 and 24) and with the chimeric receptor GR-ER.CASl in the presence of dexamethasone (Figure 5, lanes 20 and 21). No stimulation was observed with the ER-GRCASl chimeric receptor (Figure 5, lanes 17 and 18), demonstrat- ing that region C of the ER is responsible for specific tar- get gene recognition. In agreement with our previous report (Green and Chambon, 1987), stimulation of MMTV- CAT was observed with both the GR expression vector HGl in the presence of dexamethasone (Figure 5, lanes 11 and 12) and the chimeric receptor ER-GR.CASl in the presence of estradiol (Figure 5, lanes 5 and 6), whereas no stimulation (compared with pKCR2) was observed with GR-ER.CASl (Figure 5, lanes 7-9). These results were confirmed at the RNA level by using quantitative Sl nu- clease analysis in similar cotransfection experiments in which MMTV-CAT and vit-tk-CAT were replaced by MMTV- globin and vit-tk-globin (Figure 1). In this assay, the chi- merit receptors ER-GR.CASl and GR-ER.CASl were ap- proximately 50% as efficient at stimulating transcription as HGl and HEO, respectively (data not shown).

Functronal Domains of the Human Estrogen Receptor 945
Et?-GR.CASl ~~ Lw1 OR-ER.CASl
.Q -+--++ ___- -_ E2 -+--++----++
MMTV-CAT vit-tk-CAT
Figure 5. Construction of Chimenc Receptors and Their Selective Inductron of Steroid-Responsive Genes The structures of HEO, HE63, ER-GR.CASl, HGl, HG7, and GR- ER.CASl are shown schematically. The numbers above each recom- binant refer to the position of amino acids. The putative DNA-binding domams and hormone-binding domains of the human ER (HEO) and GR (HGi) are symbolized with ER and E2, and GR and DEX. respec- tively. The Kpnl and BamHl restriction endonuclease sites were intro- duced at the extremities of the putative DNA-binding domains of HE0 and HGl to create HE63 and HG7, which contain the ER.CAS and GR.CAS, respectively (Indicated by shading). The cassettes were ex- changed between HE63 and HG7 to construct the chimeric receptors ER-GRCASl and GR-ER.CASl. The chimeric receptors, HEO, HGl. and pKCR2 (1 ng each) were cotransfected in HeLa cells together with either the estrogen-responsrve (vit-tk-CAT) or the glucocorticoid-re- sponsive (MMTV-CAT) reporter genes (1 ug each). Cultures were main- tained for 48 hr in the absence (-) or presence (+) of 10-s M estradiol (E2) or 1O-7 M dexamethasone (DEX) before CAT activity was mea- sured
Region E Is Essential for Efficient Activation of Transcription Receptors containing large deletions within hormone- binding domain E (HE5 to HE9, Kumar et al., 1986; see Figure 4) failed to activate vit-tk-CAT transcription, whereas HE13, which lacks the less conserved C-terminal region F, had wild-type activity (Table 1 and Figure 2B, lanes 23 and 24). These results suggest that hormone- binding domain E is indispensable for gene activation, but do not exclude the possibility that the only function of the unoccupied hormone-binding domain is to prevent the receptor from binding to the ERE and/or activating tran- scription. Mutants HE15 HE16, HE17, HE38, and HE39- which lack regions E and F-and HE21-which lacks a large portion of region E and all of region F-were pre- pared to investigate these possibilities (Figure 4). Ir- respective of the presence of estradiol, HE15, HE21, HE38, and HE39 only stimulated vit-tk-CAT (Table 1 and Figure 28, lanes 25 to 28) and vit-tk-globin (Figure 3A, lanes 13 to 16) to a level approximately 5% of that ob-
18.4- 0
“‘3-, 2 3 4 56 7 8 9%112131415
Figure 6. Expression of Human ER Deletion Mutants In Vivo
HeLa cells, transfected with the wild-type (HEO) and/or mutant human ER receptor expression vectors, were metabolically labeled with [ssS]methionine. The expressed receptors were immunoprecipitated from cytoplasmic extracts of these cells with the monoclonal antibody H226 (for HEO, HE15, HE16. HE17, and HE21) or H222 (for HE18 and HE19), and then separated on a 12% SDS-polyacrylamide gel. The ex- pression of HE16 in lane 13 is not representative since equal levels of expression for HE16 and HE0 were seen in similar independent ex- periments. The position of the wild-type ER and of various mutant pro- teins is indicated by solid arrowheads, while the open arrowheads rep- resent proteolytic fragments of the receptor. The wild-type and mutant receptor proteins appeared to undergo a specific cleavage in HeLa cells near a point corresponding to the border of regions C and D. A proteolytic fragment of -28 kd (lanes l-4) was observed with the N-terminal-specific monoclonal antibody H226, while an -38 kd poly- peptide could be seen with the C-terminal-specific monoclonal anti- body H222 (lanes 8-9). Note, however, that no proteolytic fragments were observed in crude nuclear preparations in similar experiments (lanes 11.13. and 15). Lane M (lane 10) represents the mol. wt. markers (68 kd, BSA; 43 kd, ovalbumin; 25.7 kd, a-chymotrypsinogen; 18.4 kd, 8-lactoglobulin; and 14.3 kd, lysozyme).
served under identical conditions with HEO, whereas HE16 and HE17 were totally inactive (Table 1; Sl nuclease data not shown). Experiments performed with the same C-terminal ER mutants and the pS2 reporter genes gave similar results (Table 2 and Figure 2C, lanes 11 and 12; Figure 38, lanes 9 and 10). No stimulation of the vitelloge- nin reporter genes could be detected by using HE20 (see Figure 4) a severely deleted ER mutant containing the highly conserved 66 amino acids of region C (Table 1 and data not shown). As expected from the total inability of HE1 1 to stimulate transcription (see above), the low consti- tutive activity of HE15 HE21, HE38, and HE39 required the integrity of region C, since both HE36 (HE21 deleted for the core of region C; Figure 4) and HE26 (HE21 bear- ing in region C the Cys-*His mutations present in HE27; see Green and Chambon, 1987) were unable to stimulate transcription (Figure 3B, lanes 11 and 12; Figure 4; and Sl nuclease data not shown).
In HE47 and HE48, we deleted regions of the ER that correspond to two regions proposed to be important in

Cell 946
@ HE0 (Song) + Competitor (500ng) a3 0 Figure 7. Quantitative Sl Nuclease Analysis of I HEOtSOng) +
Y L I I I
4 Y,k?kr;lW
HE0 (50ng) HEO-Medicated Vit-tk-Globin Transcription in
PYYrrrY GG - +++++++E2 - -+ --_-- *CR2
en-no00 the Presence of Various Competitor Receptors E2 ‘- - + +‘~~~~~~~ ‘0.2 1 5 ktgl
- - -9 c.“?OO 0 ooii 1 HElS(lg) (A) HeLa cells were cotransfected with 50 of ng
‘;G’ - - ‘;;G HE0 and 500 ng of competitor expression vec- -122- m tor (pKCR2, HE15, HE16, HE17, HE21, HE26, or
.1 r-110- *r HE36) in the presence of 1 pg of vit-tk-globin (VTG) and 200 ng of the internal reference plas- mid pG1B. Cells were maintained for 36-40 hr
m-go- - in the absence (-) or presence (+) of lo-* M estradiol (E2), and cytoplasmic RNA was pre-
r-76-m pared. Sl nuclease analysis was performed
--67-W with 10 ng of cytoplasmic RNA, and the den-
a+1PW@4P r*.bu--n--a+i sitometric analysis of the suppression of tran-
12 3 4 5 6 7 6 9101112131416161716 pa,a , s 3 M M 1 2 3 4 5 6 7 6 pG’s scription (VTG, +l) was performed with refer- ence to pGlB as an internal standard (see Experimental Procedures). About 90% (aver-
age value of three independent experiments that gave between 88% and 92% decreases as estimated by densitometric scanning of the autoradio- grams) suppression of transcription was observed with HE15 and HE21, while the level of transcription in the presence of pKCR2, HE16, HE17, HE26, and HE36 was between 92% and 100% when taking HE0 alone as 100%. (6) HeLa cells were cotransfected with 50 ng of HE0 and increasing amounts of VTG plasmid (200 ng, 1 Kg, and 5 ug). pGlB (200 ng) was also added as an internal control. Cells were maintained for 36-40 hr in the presence of 10-s M estradiol, cytoplasmic RNA was isolated, and Sl nuclease analysis was performed as in (A). (C) Competition with increasing amounts of HE15. HeLa cells were cotransfected with HE0 (50 ng) and pKCR2 (2 pg) or increasing amounts of HE15 (50 ng, 150 ng, 500 ng, and 1 pg. as indicated). pGlB (200 ng) was transfected as the internal reference plasmid. RNA was isolated after cells were grown for 36 hr in the absence or presence of estradiol (10-s M). A lo-fold excess (500 ng) of HE15 gave a 90% decrease of the transcriptional activity observed with HE0 alone. The markers (M) are end-labeled Mspl restriction fragments of pBR322.
preventing the GR from activating gene transcription in the absence of the hormone (see Discussion). Both HE47 and HE48 were unable to activate any of the ER reporter genes (see Figure 3A, lanes 17 and 18; and data not shown).
Possible explanations as to why little or no stimulation was observed when using these mutants with deletion of or within the hormone-binding domain (HE15 HE16, HE17, HE20, HE21, HE38, HE39, HE47, and HE48) are that they are unstable or unable to enter the cell nucleus, and there- fore unable to interact with the ERE. The use of human ER monoclonal antibodies (H222 and H226) showed that these receptor mutants (except for HE20, which does not contain the epitopes for these antibodies) are expressed and have the expected size (Figure 6 and data not shown). In addition, HE15, HE16, and HE17(theother mutants were not tested) could be detected in the nucleus (our unpub- lished immunohistochemistry data). As shown in Figure 6, mutants HE15 (lanes 2-4) HE16 (lanes 5, 12, and 13) and HE17 (lanes 6, 14, and 15) are all expressed in amounts not too different from HE0 (see legend to Figure 6). Simi- lar results were obtained with HE21, HE38, HE39, HE47, and HE48 (data not shown). HE20, when expressed in a coupled in vitro transcription/translation assay (Kumar et al., 1986) produced a protein of the expected size (data not shown).
The Hormone-Binding Domain Is Not Required for Recognition of the ERE Alternative explanations as to why mutants HE15, HE16, HE17, HE21, HE38, and HE39 activate gene expression poorly, or not at all, are that they lack a domain located in regions D or E that is important for activation of transcrip- tion, and/or that they have a weak affinity for the ERE. Competition experiments performed in HeLa cells by
cotransfection of these mutants with HE0 and vit-tk-CAT (data not shown) or vit-tk-globin (Figure 7A) showed that HE15 HE21, HE38, and HE39 could efficiently “repress” the ability of HE0 to stimulate the expression of the reporter gene (Figure 7A, compare lanes 7 and 8 and lanes 13 and 14 with lanes 3 and 4; and data not shown). No significant competition was seen with HE26 (Figure 7A, lanes 15 and 16) HE36 (lanes 17 and 18) the parent vector pKCR2 (lanes 5 and 6) HE16 (lanes 9 and lo), or HE17 (lanes 11 and 12). These results suggest that the poor transcriptional activators HE15, HE21, HE38, and HE39 bind efficiently to the ERE in vivo and therefore pre- vent stimulation with HEO, whereas the noncompeting receptors mutated within the DNA-binding domain (HE26 and HE36) cannot bind to the ERE. The other noncompet- ing mutants, HE16 and HE17, which contain deletions ei- ther just at the border or within region C (see Figure 4) and cannot stimulate transcription, may also be incapable of binding to the ERE. To exclude the possibility that these “repressing” receptor mutants compete for a limiting tran- scription factor, we performed transfections using a fixed quantity of HE0 (50 ng, identical to that used in Figure 7A) with increasing amounts of vit-tk-globin (200 ng, 1 pg, and 5 pg; see Figure 78). That more RNA was synthesized when 5 pg of vit-tk-globin compared with 1 pg was used suggests that no transcription factor was limiting under these conditions. It is noteworthy that a lo-fold excess of competitor DNA resulted in an approximately 90% de- crease in activation of transcription by HE0 (see Figures 7A and 7C and their legends), which suggests that the truncated HE15 and HE21 receptors have an affinity for the ERE similar to that of the wild-type ER. It appears, therefore, that the presence of the intact hormone-binding domain of the human ER is essential for efficient stimula- tion of gene transcription, but not for specific recognition of the ERE.

Functional Domains of the Human Estrogen Receptor 947
Differential Effect of N-Terminal Deletions on Vitellogenin and pS2 EREs It was previously speculated that region A/B may be im- portant for activating gene transcription (Krust et al., 1986; Kumar et al., 1986). Cotransfections of mutants containing deletions in this region (HEl, HE2, and HElO) with vit-tk- CAT demonstrated that they were in fact able to activate transcription (Table 1 and Figure 2B, lanes 5-16). Similar results were obtained with pSBCAT, but HE2 had only ap- proximately 60% the stimulatory activity of HE0 (Table 2 and Figure 2C, lanes 3-8).
Two deletion mutants, which removed almost all (HE18) or all (HE19) of region A/B (Figure 4), were constructed to eliminate the possibility that this region may contain sev- eral functionally redundant elements. Both HE18 and HE19 bound hormone with the same affinity as HE0 (Fig- ure 4 and data not shown). “Tight nuclear binding” deter- minations showed that HE18 had nuclear levels approxi- mately 40% those of HEO, while the value for HE19 was about 70% (data not shown). Each of these mutants stimu- lated transcription to the same extent as HE0 with vit-tk- CAT (Table 1) or vit-tk-globin (Figure 3A; compare lanes 10 and 12 with lane 4). On the other hand, pSBCAT expres- sion was stimulated only by approximately 10% with HE18 and 17% with HE19 (Table 2 and Figure 38, compare lanes 6 and 8 with lane 2), indicating that these mutants have different effects on the two types of estrogen-responsive reporter genes. The same relative decrease was observed when between 250 ng and 2 ug of the receptor expression vectors was used (data not shown), suggesting that this effect cannot simply be ascribed to a lower affinity of the ER mutants for the pS2 ERE.
Thus the presence of region A/B is not absolutely re- quired for stimulation of transcription, but may be nec- essary to obtain maximal stimulation with some estrogen- responsive genes.
Does Region D Act as a Hinge? One deletion mutant (HE12) and six insertion mutants (HE22 to HE25, HE55, and HE56) were constructed to in- vestigate what effect varying the length of region D has upon stimulation of transcription (Figure 4). HE12 contains a deletion of 30 amino acids that are relatively poorly con- served between the human (Green et al., 1986), rat (Koike et al., 1987) chicken (Krust et al., 1986), and Xenopus (Weiler et al., 1987) ERs. This mutant receptor bound es- tradiol with the same affinity as HEO, but the amount of the receptor-hormone complex that bound tightly to the nu- cleus in the presence of 1O-8 M estradiol was only about 20% that observed with HE0 (data not shown). HE12 stimulated vit-tk-CAT and vit-tk-globin transcription equally as well as HE0 (Table 1 and Figure 3A, compare lane 8 with lane 4), whereas only approximately 40% stimulatory activity was obtained with the pS2 reporter genes (Table 2 and Figure 3B, compare lane 4 with lane 2), again indi- cating that alterations within the human ER has a different effect upon the two estrogen-responsive reporter genes.
The insertion mutants that contain between 3 and 14 ad- ditional amino acids in the middle of region D (HE22 to HE25, HE55, and HE56; see Figure 4 and Experimental
Procedures) displayed wild-type characteristics for es- tradiol binding (Figure 4 and data not shown). Their ability to stimulate vit-tk-CAT and vit-tk-globin transcription was not significantly different from that of HE0 (Table 1, Figure 4, and data not shown). Note that mutants HE25, HE55, and HE56 contain 1, 2, or 5 proline residues within the in- sertion.
Thus the length of region D (39 amino acids) may be al- tered considerably in both length (between the deletion of 30 amino acids and the insertion of 14 amino acids) and composition without a loss of receptor function.
Discussion
Mutational analyses of the human ER showed that the conserved region E contains the hormone-binding do- main, and that the highly conserved region C is essential for tight nuclear binding of the receptor (see Kumar et al., 1986). We attempted here to define those regions that are responsible for two other essential properties of the recep- tor: specific recognition of the ERE and activation of gene transcription.
Region C Recognizes the Estrogen-Responsive Element Region C of the human GR recognizes specifically the GRE present in the long terminal repeat (LTR) of MMTV, and we postulated that region C of the ER may have an analogous function (Green and Chambon, 1987). We demonstrate here, using a new set of chimeric recom- binants, that region C of the ER is indeed responsible for the specific recognition of the ERE within target genes. “Specific recognition” refers here to recognition leading to the specific activation of target genes. Since region C is rich in basic amino acids and has a sequence reminiscent of the DNA-binding “fingers” (see Berg, 1986, for refer- ences), it is probable that this region interacts directly with the ERE in vivo. That no additional factors are involved in this interaction is suggested by the observation that ho- mogeneous preparations of the progesterone receptor (PR) can bind to its responsive element (Bailly et al., 1986). Moreover, results of the competition experiments suggest that the hormone-binding domain is not required for region C to exist in a conformation capable of efficiently interacting with the ERE.
Multiple Domains Are Required for Efficient Activation of Transcription Mutations within region C result in receptors incapable of activating transcription, most probably because they can- not bind to the ERE. The results obtained with mutants in which hormone-binding domain E or the A/B region have been deleted suggest that region C may also contain an element involved in activation of transcription. However, mutants that lack the hormone-binding domain only stimulate transcription poorly (approximately 5% of the level of the wild-type receptor; see Figure 4) even though competition experiments (Figure 7) suggest that they bind efficiently to the ERE in vivo. Thus, in the absence of the hormone-binding domain the truncated receptor activates

Cell 948
transcription poorly, yet appears to bind to the ERE as effi- ciently as the intact hormone-receptor complex. The poor constitutive stimulatory activity of these truncated ER mu- tants is in apparent contrast to results obtained with the rat (Godowski et al., 1987) and human (Hollenberg et al., 1987) GRs where various mutants, with deletion of or within region E, activate MMTV-CAT transcription to a level between 10% and 120% that observed with the wild- type GR. In our hands, using quantitative Sl nuclease analysis, the human GR mutant HG3, which is truncated from amino acid 532 at the border of regions D and E (equivalent to the ER mutant HE39). activates transcrip- tion from the reporter gene MMTV-globin to a level 20% of that achieved with the wild-type GR (data not shown). Using MMTV-CAT as a reporter gene, Hollenberg et al. (1987) found 10% activity for an identical truncation in the human GR. We note, however, that an analogous mutation at the border of regions D and E of the chicken PR results in less than 1% constitutive activity of that observed with the wild-type PR when MMTV-globin is used as a reporter gene (Gronemeyer et al., 1987b). In an attempt to produce ER deletion mutants that would exhibit higher levels of constitutive activity, we constructed HE47 and HE48. HE48 bears a deletion in the hormone-binding domain from amino acid 315 to 478, which is equivalent to the dele- tion present in a human GR mutant (A532-697) that ex- hibits 120% constitutive activity (Hollenberg et al., 1987). HE47 is deleted for a segment of the hormone-binding do- main that is well conserved among the members of the receptor gene family (Krust et al., 1986; Gronemeyer et al., 1987a), and has been proposed to be important in prevent- ing activation of transcription in the absence of the hor- mone (G. Ringold, personal communication). Neither HE47 nor HE48 is able to stimulate transcription of either the vitellogenin or pS2 reporter genes.
In contrast to this hormone-binding domain require- ment, region A/B does not appear to have a general role in activation of transcription by the ER, since its deletion reduces expression of the pS2 but not of the vitellogenin reporter genes. Although there are no apparent sequence similarities within the corresponding region of the various members of the steroid receptor family (Krust et al., 1986; Gonemeyer et al., 1987a), this region seems to be impor- tant also for activation of transcription by both the GR and the PR. Mutagenesis within this region of the human (Giguere et al., 1986; Hollenberg et al., 1987) and rat (Miesfeld et al., 1987) GRs results in up to a 9.5% decrease in activation of transcription, whereas the chicken PR is inactivated by deletion of region A/B (Gronemeyer et al., 1987b).
None of the insertions nor the deletion made within re- gion D of the ER prevented the receptor from stimulating transcription. These results, which support our sugges- tion that region D acts as a hinge between domains C and E, indicate that a functional receptor does not require that region D adopt a precise structure. We note, however, that a single Pro-+Arg mutation in the corresponding region of v-e&A inactivated this member of the nuclear receptor family (Damm et al., 1987). It is interesting to note that the insertion of proline residues (HE25 HE55, HE56) that
would be expected to disturb greatly the structure of re- gion D does not inactivate the ER, and that the human mineralocorticoid receptor contains a stretch of eight pro- lines in between the DNA- and hormone-binding domains (Arriza et al., 1987).
How Does the Estrogen Receptor Function? Binding of the hormone to domain E of the estrogen receptor is required to achieve both tight nuclear binding (Kumar et al., 1986) and activation of transcription of target genes. Presumably, the action of the hormone binding in- volves a modification in the structure of the receptor that allows it to bind to the ERE, since genomic footprinting ex- periments have shown that the GR occupies the GRE only in the presence of hormone (Becker et al., 1986). Studies on the GR have also led to the proposal that the only func- tion of the hormone-binding domain is to “mask” the DNA- binding domain and/or the domain responsible for activa- tion of the transcription machinery (Godowski et al., 1987; Hollenberg et al., 1987). Our results, which indicate that ER mutants lacking the hormone-binding domain interact with the ERE in vivo as efficiently as the wild-type hor- mone-receptor complex, are compatible with the idea that the presence of an unoccupied hormone-binding domain results in the masking of the DNA-binding domain. Mask- ing may correspond to the direct physical masking of re- gion C by region E, or to a structural constraint imposed by region E that maintains the DNA-binding domain in an inactive form, or to the association of the unoccupied receptor with other proteins (such as the 90 kd heat shock protein; see Groyer et al., 1987; Green and Chambon, 1987) resulting in an inactive 8s receptor complex. Note that none of the present deletions within the hormone- binding domain of the ER nor alteration in the length of region D can relieve this putative “masking” effect,
Masking of the DNA-binding domain cannot, however, be the only function of the hormone-binding domain, since ER mutants lacking this domain only activate tran- scription poorly. There are several possibilities to explain how the hormone-binding domain could play a more posi- tive role in activation of gene transcription. It may interact directly with some component of the transcription ma- chinery or influence the ability of other regions of the receptor to interact with these components. Alternatively, the activation function of the hormone-binding domain could be related to the property of the ER to form a 55 homodimer complex in the presence of the hormone (for references, see Gordon and Notides, 1986). Linstedt et al. (1986) have shown that the dimer binds one molecule of the H222 monoclonal antibody and two molecules of the H226 monoclonal antibody, while the monomer form binds one molecule of each. We have previously shown the H226 epitope to be located in region A/B, just N-ter- minal to the DNA-binding domain, and that of H222 to be located in the hormone-binding domain (Kumar et al., 1986). Therefore, dimerization may occur through interac- tion of the hormone-binding domains, resulting in the masking of one of the epitopes for H222. Since the EREs of both the Xenopus and chicken vitellogenin genes ap- pear to be palindromic (Klein-Hitpass et al., 1986; Seiler-

Functional Domains of the Human Estrogen Receptor 949
Tuyns et al., 1986; our unpublished data), the ER may need to dimerize in order to activate transcription effi- ciently. If so, this may explain why ER mutants lacking the hormone-binding domain, and therefore presumably act- ing as monomers, only stimulate transcription poorly.
It is puzzling that the presence of region A/B is required for efficient activation of the pS2 reporter gene, but not for activation of the vitellogenin reporter gene. As noted above, the analogous region of the human (Gigu&re et al., 1986; Hollenberg et al., 1987) and rat (Miesfeld et al., 1987) GRs is also necessary for efficient stimulation of tran- scription from the MMTV LTR. Interestingly, in the latter case, as for the pS2 gene, the receptor is acting on a “nat- ural” promoter region, whereas the vit-tk reporter genes are highly chimeric. This suggests that the requirement for region AIB may be dependent on the nature of the acti- vated promoter. Alternatively, it may be dependent on the nature of the ERE. In this respect, we note that the pS2 gene ERE does not contain the perfect palindromic se- quence present in the vitellogenin gene EREs (M. Berry, A. M. Nunez, and l? Chambon, unpublished data). It should also be stressed that the present studies have been performed using “transient” assays in which the acti- vated reporter genes are not integrated in the genome, and therefore not in a “normal” chromatin environment.
In conclusion, although the role of region E in binding the hormone and of region C in interacting with the ERE appears clear, our studies have failed to identify a discrete region(s) responsible for efficiently activating the tran- scription machinery. Whether this reflects the complexity of the activating domain(s) or the nature of the mechanism by which enhancer factors stimulate transcription remains to be established.
Experimental Procedures
Construction of Human ER Mutants Site-directed mutagenesis was utilized to construct the present series of human ER deletion mutants (see also Kumar et al., 1986). Single- stranded DNA templates were prepared by using either the viral vector M13mplE or the plasmid vector Bluescribe M13+ (Stratagene) contain- mg the 1.8 kb insert that has the ER cDNA complete open reading frame (ORF) (Green et al., 1986). The design of the mutagenesis oligo- nucleotide primers was such that they allowed precise deletions to be made as well as the introduction of either a 3’stop codon in HE16, HE38, and HE39 or a Kozak Sconsensus leader sequence (SCCACC- ATGG-3’; Kozak. 1986) in HE18 and HE19 without alteration of the reading frame of the ORF (see Table 1 for position of deletions). The mutants were screened by using the 5’ end-labeled mutagenesis oli- gonucleotides as probes, and all the constructions were verified by dtdeoxy sequencing. HE15 was constructed from HE22. and HE17 from HE28 (Green and Chambon, 1987), by introduction of an in-frame stop codon after the unique Xhol site at amino acids 281 and 251, respectively. HE15 has an in-frame sequence for three extra amino acids (SerArg-Val) between residue 281 and the stop codon. HE20 was prepared from the ER-CAS (Green and Chambon. 1987) by addition of a Kozak consensus sequence upstream of the Kpnl site (amino acid 185) and an in-frame stop codon after the Xhol site (amino acid 250). Mutants HE21, HE26, and HE36 were constructed respectively from HEO, HE27 (Green and Chambon. 1987), and HE11 by cleaving at the unique Hindlll site in the ORF (located at amino acid 340) and ligating oligonucleotides containing an in-frame stop codon. All mutant cDNAs possessed EcoRl sites at their extremities, which facilitated their clon- ing Into the eukaryotic expression vector pKCR2 (Breathnach and Harris, 1983)
The insertion mutants were constructed after creation of a unique
Xhol site between residues 281 and 282 of the human ER, which mtro- duced three amino acids (HE22). Oligonucleotides corresponding to different amino acid sequences were then introduced into this Xhol site to generate HE23-HE25, HE55, and HE56. The amino acids (stngle-letter code) inserted were as follows: HE22, SRA; HE23, SRALALAIDISRA; HE24, SRGMGMGIDISRA; HE25, SRPIDSLLSLL- SRA; HE55, SRAPPIDISRA; HE56, SRAPPPPPIDISRA. The se- quence of each of the mutants was verified before cloning into pKCR2.
Construction of the Human GR Expression Vectors and the Chimeric Receptors The human GR expression vector HGl was constructed from the cDNA clones isolated by M. V. Govindan, M. Devic, and U. Stropp (Govindan et al., 1985. and our unpublished data). The two internal EcoRl sites in the 2331 nucleotide long ORF (Hollenberg et al., 1985) were destroyed by site-directed mutagenesis without alteration of the protein sequence. A translation consensus leader sequence, YCCACCAn;G-3’ (Kozak, 1986), with an EcoRl site was also intro- duced at the 5’end by site-directed mutagenesis. The resulting 2.7 kb cDNA fragment contained 9 nucleotides upstream of the ATG codon, an in-frame insertion coding for four additional amino acids (IleAla- ArgAla) between the residues 500 and 501, and 386 nucleotides down- stream of the TGA stop codon. This fragment was inserted at the EcoRl site of the eukaryotic expression vector pKCR2 (Breathnach and Harris, 1983) to obtain HGI. The level of MMTV LTR-GRE stimulation with HGl was similar to that observed with HGO, the human GR ex- pression vector containing the wild-type ORF, cloned at the BamHl site in the pKCR2 vector (Breathnach and Harris, 1983) after repair of the ends and addition of BamHl linkers (data not shown). HG3 was con- structed by site-directed mutagenesis of HGl, inserted in the Blue- scribe M13+ vector, by creation of a stop codon after amino acid 532.
The chimeric receptors ER-GRCASI and GR-ERCASl (see Fig- ure 5) were also constructed by mutagenesis as described previously (Kumar et al., 1986; Green and Chambon, 1987). The cDNA inserts of HE0 and HGI were cloned into the Bluescribe M13+ vector in which the Kpnl and BamHl sites were selectively removed from its polylinker. The oligonucleotides used for creating the Kpnl and BamHl sites withm the putative DNA-binding domains of human ER and GR were 30 nucleotides in length, with 12 nucleotides on either side of the new site being complementary to the cDNA sequence. Both the Kpnl and BamHl oligonucleotides were used together during mutagenesis, and clones that were positive with both probes were isolated. The Kpnl and BamHl sites allowed the exchange of this region as a cassette (ER.CAS or GRCAS) to yield, finally, ER-GR.CASl and GR-ERCASl.
Construction of Reporter Plasmids The reporter plasmids vit-tk-CAT (Klein-Hitpass et al., 1986) and MMTV-CAT (Cat0 et al., 1986) were kindly provided by G. Ryffel and H. Ponta, respectively. The MMTV-globin reporter gene was con- structed and made available to us by our colleagues studying the progesterone receptor (Gronemeyer et al., 1987b). The pSP-CAT re- porter plasmid was constructed by replacement of the Sall-Hindlll fragment of the chicken collagen a2 promoter region from pCOL-CAT (Herbomel et al., 1984) with a Pvull-Hinfl fragment (position -1100 to +lO) of the pS2 promoter and 5’ flanking sequences (Jeltsch et al., 1987). Similarly, vit-tk-globin and pS2-globin plasmids were con- structed by substitution of the SV40 early promoter region of the pA0 plasmid (Zenke et al., 1986). A 400 bp Hindlll-Bglll fragment from the vit-tk-CAT plasmid containing the Xenopus vitellogenin A2 (vit) up- stream sequence (-331 to -87) and the Herpes simplex virus thymi- dine kinase (tk) promoter (-105 to +51) was inserted upstream of globin coding sequences, in the case of vlt-tk-globtn. pS2-globin was constructed by insertion of the 1.1 kb Pvull-Hinfl fragment encom- passing the 5’flanking and promoter region (-1100 to +lO) of the pS2 gene.
Cytoplasmic and Nuclear Estrogen Binding Assays Plates (90 mm) of HeLa cells at 30%-50% confluence were transiently transfected with 10 pg of receptor expression vector purified on two CsCl gradients by using the calcium phosphate coprecipitabon tech- nique of Banerji et al. (1981). The cells were grown for 48 hr in asteroid- free medium, cell extracts were prepared, and the cytosolic receptor content was measured by single saturation assay (Green et al., 1986).

Cell 950
The affinity (K,,) of the receptor mutants was calculated from Scatch- ard plot analysis with a multipoint assay.
Nuclear binding capabilities of the human ER mutants were deter- mined from similarly transfected HeLa cells in the presence of 10m9 M and/or lo-* M [3H]estradiol (101 Cilmmol; Amersham) as described earlier (Kumar et al., 1988). and the nuclear binding sites were calcu- lated from the radioactivity bound tightly to the nuclei.
lmmunoprecipitation of Estrogen Receptor For immunoprecipitation of the human ER mutants, subconfluent HeLa cells were transfected as above with IO ug of DNA per 90 mm plate, grown in the absence of estradiol for 40 hr, and metabolically labeled for 3 hr at 37% in 5 ml of methionine- and serum-deficient Dulbecco’s medium supplemented with [ssS]methionine (10 Q/ml medium; 1200 Cilmmol; Amersham). Cell extracts, prepared by 0.5% Nonidet P-40 treatment, were processed for immunoabsorption with human ER monoclonal antibodies H222 or H228 (Abbott Laboratories), electro- phoresed, and visualized by fluorography as described earlier (Green et al., 1986).
CAT Assay CAT enzyme assays were performed as described by Gorman et al. (1982). Briefly, subconfluent HeLa cells were cotransfected in duplicate with different amounts of the receptor expression vectors and either the estradiol-responsive reporter plasmids vit-tk CAT (1 pg) or pSP-CAT (10 ug), or the glucocorticoid-responsive reporter plasmid MMTVCAT (1 ug). In each case, the carrier DNA (Bluescribe M13+) was added to a total of 20 ug per 90 mm culture plate. Cells were cultured for 48 hr in the absence or presence of either 10-s M estradiol or lo-’ M dexa- methasone, and cell extracts were prepared by five cycles of freeze- thawing followed bycentrifugation at 10,000 x g for 15 min. The protein content of the samples was determined by Bradford assay, and CAT assay was performed by using either IO ug of protein extract (vit-tk-CAT and MMTV-CAT) or 50 ug of protein extract (pS2CAT) with 0.1 uCi of [14C]chloramphenicol (53 mCi/mmol; Amersham) for 1 hr at 37%. The acetylated and nonacetylated forms of [%]chloramphenicol were separated by thin-layer chromatography, excised, and quantitated by liquid scintillation counting. CAT activity was calculated as the percent- age of that observed with HEO.
Quantitative Sl Nuclease Analysis Quantitative St analysis was performed by the procedure of Zenke et al. (1986). The single-stranded DNA probes were prepared by primer extension of a single-stranded DNA template of M13mp19 containing the 1142 bp Hindlll-Dral fragment from vit-tk-globin. or of M13mp18 containing the 605 bp BamHI-BamHI fragment from pS2-globin. A [y-s2P] 5’ end-labeled synthetic oligonucleotide, 5’-GCACCATTCTGT- CTGTTTTGGG-3’, complementary to +39 to +60of the rabbit 5-globin gene, was used in each case for priming the template. The primer ex- tension products were cleaved with EcoRI. and the 212 nucleotide- and 190 nucleotide-long DNA probes for vit-tk-globin and pS2-globin, respectively, were purified by electrophoresis on a 6% polyacrylamide gel containing 8.3 M urea and were recovered by electroelution.
HeLa cells, transfected with various receptor recombinants and the reporter plasmids vit-tk-globin or pS2-globin (see Figure I), were main- tained for 36-40 hr in the absence or presence of 10-s M estradiol. pA0 (Zenke et al., 1986) or pGlB (Wasylyk and Wasylyk, 1986) were also cotransfected as internal reference plasmids. The cytoplasmic RNA was prepared by lysing of the cells with 0.5% Nonidet P-40 (Grou- dine et al., 1981) and hybridized with an excess of probe either at 65% in 10 mM PIPES (pH 6.5) and 400 mM NaCI, or at 42% in the above buffer containing 50% (v/v) formamide and 1 mM EDTA. After St nuclease digestion (100 units at 25% for 2 hr; Appligene), the pro- tected DNA fragments were subjected to electrophoresis on a 6% se- quencing gel. Quantitation was performed by densitometric scanning of autoradiograms corresponding to different exposures of the same gel from at least three independent experiments. Signals two orders of magnitude lower than those with HE0 could be measured. The in- tensity of transcription signals from internal reference plasmid pGlB or pA0 was used to correct any variation in transfection efficiencies.
Acknowledgments
We are grateful to A. M. Nunez, G. Ryffel, and Ii. Ponta for providing
pS2-globin, vit-tk-CAT, and MMTV-CAT, respectively; to M. Govindar,, M. Devic, and U. Stropp for providing the hGR cDNA clones; and to Abbott Laboratories for providing the H222 and H226 monoclonal anti- bodies. We also thank V. L. Kumar for helpful discussion, E. Scheer and I. lssemann for technical assistance, A. Staub and F. Ruffenach for synthesizing the oligonucleotides, the tissue culture lab for media and cells, and C. Werle and B. Boulay for preparing the figures. This research was supported by INSERM. CNRS (A. I. V), the INSERM (grants CNAMTS), the Ministere de la Recherche et de la Technologie, the Fondation pour la Recherche Medicale, and the Association pour la Recherche sur le Cancer. G. Stack was supported by the Rockefeller Foundation and the National Institutes of Health.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received September 2, 1987; revised October 22, 1987
References
Arriza, J. L., Weinberger, C., Cerelli, G., Glaser, T. M., Handelin, B. L., Housman, D. E.. and Evans, R. M. (1987). Cloning of human miner- alocorticoid receptor complementary DNA: structural and functional kinship with the glucocorticoid receptor. Science 237, 268-275.
Bailly, A., Le Page, C., Rauch, M., and Milgrom, E. (1988). Sequence- specific DNA binding of the progesterone receptor to the uteroglobin gene: effects of hormone, antihormone and receptor phosphorylation. EMBO J. 5, 32353241.
Banerji, J., Rusconi. S.. and Schaffner, W. (1981). Expression of a 6-globin gene is enhanced by remote SV40 DNA sequences. Cell 27, 299-308.
Becker, f? B.. Gloss, B., Schmid, W., Strahle, U., and Schutz, G. (1986). In viva protein-DNA interactions in a glucocorticoid response element require the presence of the hormone. Nature 324, 686-688. Berg, J. M. (1986). More metal-binding fingers, Nature 379, 264-265.
Breathnach, R., and Harris, B. A. (1983). Plasmids for the cloning and expression of full-length double-stranded cDNAs under control of the SV40 early or late gene promoter. Nucl. Acids Res. 17, 7119-7136.
Brown, A. M. C., Jeltsch, J. M., Roberts, M.. and Chambon, P (1984). Activation of pS2 gene transcription is a primary response to estrogen in the human breast cancer cell line MCF-7. Proc. Natl. Acad. Sci. USA 81, 6344-6348.
Cato. A. C. B., Miksicek, Ft.. Schijtz, G., Arnemann, J., and Beato, M. (1986). The hormone regulatory element of mouse mammary tumour virus mediates progesterone induction. EMBO J. 5, 2237-2240.
Damm, K., Berg, H., Graf, T., and Vennestrom, B. (1987). A single point mutation in erbA restores the erythroid transforming potential of a mu- tant avian erythroblastosis virus (AEV) defective in both erbA and erbB oncogenes. EMBO J. 6, 375-382.
Druege, P M., Klein-Hitpass, L., Green, S.. Stack, G., Chambon, P.. and Ryffel, G. U. (1988). Introduction of estrogen-responsiveness into mammalian cell lines. Nucl. Acids Res. 74, 9329-9337.
Giguere. V., Hollenberg, S. M., Rosenfeld, M. G.. and Evans, R. M. (1986). Functional domains of the human glucocorticoid receptor. Cell 46, 645-652.
Godowski, f? J., Rusconi, S., Miesfeld, R., and Yamamoto, K. R. (1987). Glucocorticoid receptor mutants that are constitutive activators of tran- scriptional enhancement. Nature 325, 365-388.
Gordon, M. S.. and Notides, A. C. (1986). Computer modeling of es- tradiol interactions with the estrogen receptor. J. Steroid Biochem. 25, 177-181.
Gorman, C. M.. Moffat, L. F., and Horward, B. H. (1982). Recombinant genomes which express chloramphenicol acetyltransferase in mam- malian cells. Mol. Cell. Biol. 2, 1044-1051.
Govindan, M. V., Devic, M., Green, S., Gronemeyer, H., and Chambon, P (1985). Cloning of the human glucocorticoid receptor cDNA. Nuci. Acids Res. 13. 8293-8304.
Green, S., and Chambon, l? (1988). A superfamily of potentially onco- genie hormone receptors. Nature 324, 615-617.

Functional Domains of the Human Estrogen Receptor 951
Green, S., and Chambon. P. (1967). Oestradiol induction of a gluco- corticoid-responsive gene by a chimaeric receptor. Nature 32.5, 75-78.
Green, S., Walter, P., Kumar, V., Krust, A.. Bornert, J. M., Argo& P., and Chambon, P (1986). Human oestrogen receptor cDNA: sequence, ex- pression and homology to verb-A. Nature 320, 134-139.
Gronemeyer, H., Green, S., Kumar, V., Jeltsch, J. M., and Chambon, P (1987a). Structure and function of the oestrogen receptor and other members of the nuclear receptor family. In Steroid Receptors and Dis- orders: Cancer, Bone and Circulatory Disorders, P J. Sheridan, K. Blum, and M. C. Trachtenberg, eds. (New York: Marcel Dekker Inc.), in press.
Gronemeyer, H., Turcotte, B., Ouirin-Stricker, C., Bocquel, M. T., Meyer, M. E., Krozowski, Z., Jeltsch, J. M., Lerouge, T., Gamier, J. M., and Chambon. F? (1987b). The chicken progesterone receptor: sequence, expression and functional analysis. EMBO J., in press,
Groudine, M., Peretz, M., and Weintraub, H. (1981). Transcriptional regulation of hemoglobin switching in chicken embryos. Mol. Cell. Biol. I, 281-288.
Grayer, A., Schweizer-Grayer, G.. Cadepond. F., Mariller, M., and Baulieu, E. E. (1987). Antiglucocorticosteroid effects suggest why ste- roid hormone is required for receptors to bind DNA in viva but not in vitro. Nature 328, 624-626.
Herbomel, l?, Bourachot, B., and Yaniv, M. (1984). Two distinct en- hancers with different cell specificities coexist in the regulatory region of polyoma. Cell 39, 653-662.
Hollenberg, S. M., Weinberger, C., Ong, E. S., Cerelli, G., Oro. A., Lebo, R., Thompson, E. B., Rosenfeld, M. G., and Evans, R. M. (1985). Primary structure and expression of a functional human glucocorticoid receptor cDNA. Nature 378, 635-641.
Hollenberg, S. M., Gig&e, V., Segui, P., and Evans, R. M. (1967). Colocalization of DNA-binding and transcriptional activation functions in the human glucocorticoid receptor. Cell 49, 39-46.
Jeltsch, J. M., Roberts, M., Schatz, C., Garnier, J. M., Brown, A. M. C.. and Chambon, P. (1987). Structure of the human oestrogen-responsive gene pS2. Nucl. Acids Res. 15, 1401-1414.
Klein-Hitpass, L., Schorpp, M., Wagner, U., and Ryffel, G. U. (1986). An estrogen-responsive element derived from the 5’flanking region of the Xenopus vitellogenin A2 gene functions in transfected human cells. Cell 46, 1053-1061.
Koike, S., Sakai, M., and Muramatsu, M. (1987). Molecular cloning and characterization of rat estrogen receptor cDNA. Nucl. Acids Res. 15, 2499-2513.
Kozak, M. (1986). Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 44, 283-292.
Krust, A., Green, S., Argos, P.. Kumar, V., Walter, P, Bornert, J. M., and Chambon, P (1986). The chicken oestrogen receptor sequence: ho- mology with v-erbA and the human oestrogen and glucocorticoid receptors. EMBO J. 5, 891-897.
Kumar, V., Green, S., Staub, A., and Chambon, P (1986). Localisation of the oestradiol-binding and putative DNA-binding domains of the hu- man oestrogen receptor. EMBO J. 5, 2231-2236.
Linstedt, A. D., West, N. B., and Brenner, R. M. (1986). Analysis of monomeric-dimeric states of the estrogen receptor with monoclonal antiestrophilins. J. Steroid Biochem. 24, 677-686. Masiakowski, P.. Breathnach, R., Bloch, J., Gannon, F., Krust, A., and Chambon, P (1982). Cloning of cDNA sequences of hormone-regu- lated genes from the MCF-7 human breast cancer cell line. Nucl. Acids Res. 70, 7895-7903. Miesfeld, R.. Godowski, t? J., Maler, B. A., and Yamamoto, K. R. (1987). Glucocorticoid receptor mutants that define a small region sufficient for enhancer activation. Science 236, 423-427.
Seiler-Tuyns, A., Walker, P, Martinez, E., Merillat, A. M., Givel, F., and Wahli, W. (1986). Identification of estrogen responsive DNA sequences by tranSient expression experiments in a human breast cancer cell line. Nucl. Acids Res. 14, 8755-8770.
Struhl, K. (1987). Promoters, activator proteins, and the mechanism of transcriptional initiation in yeast. Cell 49, 295-297.
Walter, P., Green, S., Greene, G., Krust, A., Bornert, J. M.. Jeltsch,
J. M., Staub, A.. Jensen, E., Scrace. G., Waterfield. M., and Chambon, P (1985). Cloning of the human estrogen receptor cDNA. Proc. Natl. Acad. Sci. USA 82, 7889-7893.
Wasylyk, C., and Wasylyk, 8. (1986). The immunoglobulin heavy-chain B-lymphocyte enhancer efficiently stimulates transcription in non- lymphoid cells. EMBO J. 5, 553-560.
Weiler, I. J., Lew, D., and Shapiro, D. J. (1987). The Xenopus laevis estro- gen receptor: sequence homology with human and avian receptors and identification of multiple estrogen receptor messenger ribonucleic acids. Mol. Endo. 7, 355-362.
Weinberger, C., Hollenberg, S. M., Rosenfeld, M. G.. and Evans, R. M. (1985). Domain structure of human glucocorticoid receptor and its rela- tionship to the v-erbA oncogene product. Nature 378, 670-672.
Yamamoto, K. R. (1985). Steroid receptor regulated transcription of specific genes and gene networks. Ann. Rev. Genet. 19, 209-252.
Zenke. M., Grundstrom, T., Matthes, H., Wintzerith. M.. Schatz, C., Wildeman, A., and Chambon. P (1986). Multiple sequence motifs are involved in SV40 enhancer function. EMBO J. 5. 387-397.




















