
Estrogen receptors and endocrine diseases: lessons from estrogen receptor knockout mice
Upload
independentCategory
view
2download
0
105
Estrogen receptor-related rece
ptors in the killifish Fundulusheteroclitus: diversity, expression, and estrogenresponsivenessA M Tarrant, S R Greytak1, G V Callard1 and M E HahnBiology Department, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts 02543, USA
1Department of Biology, Boston University, Boston Massachusetts 02215, USA
(Requests for offprints should be addressed to A M Tarrant; Email: [email protected])
Abstract
The estrogen receptor-related receptors (ERRs) are a group of nuclear receptors that were originally identified on the
basis of sequence similarity to the estrogen receptors. The three mammalian ERR genes have been implicated in diverse
physiological processes ranging from placental development to maintenance of bone density, but the diversity, function,
and regulation of ERRs in non-mammalian species are not well understood. In this study, we report the cloning of four
ERR cDNAs from the Atlantic killifish, Fundulus heteroclitus, along with adult tissue expression and estrogen
responsiveness. Phylogenetic analysis indicates that F. heteroclitus (Fh)ERRa is an ortholog of the single ERRa
identified in mammals, pufferfish, and zebrafish. FhERRba and FhERRbb are co-orthologs of the mammalian ERRb.
Phylogenetic placement of the fourth killifish ERR gene, tentatively identified as FhERRgb, is less clear. The four ERRs
showed distinct, partially overlapping mRNA expression patterns in adult tissues. FhERRa was broadly expressed.
FhERRba was expressed at apparently low levels in eye, brain, and ovary. FhERRbb was expressed more broadly in
liver, gonad, eye, brain, and kidney. FhERRgb was expressed in multiple tissues including gill, heart, kidney, and eye.
Distinct expression patterns of FhERRba and FhERRbb are consistent with subfunctionalization of the ERRb paralogs.
Induction of ERRamRNA by exogenous estrogen exposure has been reported in somemammalian tissues. In adult male
killifish, ERR expression did not significantly change following estradiol injection, but showed a trend toward a slight
induction (three- to five-fold) of ERRa expression in heart. In a second, more targeted experiment, expression of ERRa in
adult female killifish was downregulated 2$5-fold in the heart following estradiol injection. In summary, our results indicate
that killifish contain additional ERR genes relative to mammals, including ERRb paralogs. In addition, regulation of ERRa
expression in killifish apparently differs from regulation in mammals. Together, these featuresmay facilitate determination
of both conserved and specialized ERR gene functions.
Journal of Molecular Endocrinology (2006) 37, 105–120
Introduction
The estrogen receptor-related receptors (ERRs) aremembers of the nuclear receptor superfamily oftranscription factors (Giguere et al. 1988). Three ERRgenes have been identified in mammals, ERRa(NR3B1), ERRb (NR3B2), and ERRg (NR3B3)(Giguere et al. 1988, Chen et al. 1999, Hong et al.1999, Heard et al. 2000). ERRs and estrogen receptors(ERs) have overlapping affinities for co-activators andDNA-binding sites, but differ markedly in ligandbinding and activation (Vanacker et al. 1999a,b, Giguere2002). Unlike ERs, ERRs do not bind estradiol, andhave been reported to be either constitutively active(Hong et al. 1999, Xie et al. 1999, Greschik et al. 2002) oractivated by an unidentified ligand (Vanacker et al.1999b). While high-affinity ERR agonists have notbeen identified, some ER ligands, including4-hydroxytamoxifen and diethylstilbestrol, can
Journal of Molecular Endocrinology (2006) 37, 105–1200952–5041/06/037–105 q 2006 Society for Endocrinology Printed in Great Britain
antagonize ERR activity (Coward et al. 2001, Tremblayet al. 2001a,b).
The functions and target genes of ERRs are not yetwell understood. ERRa helps to regulate bone growthand maintenance by binding to the osteopontinpromoter, a gene target that is shared with ERa(Bonnelye et al. 1997b, 2001, Vanacker et al. 1999a).ERRa has also been shown to repress activity ofPPARg co-activator 1a (PGC-1a), a co-activator thatinteracts with PPARs (peroxisome proliferator-activatedreceptors) to regulate gluconeogenesis and adaptivethermogenesis (Ichida et al. 2002). ERRa-null mutantmice are essentially normal with reduced body weightand peripheral fat deposits, which supports thehypothesis that ERRa helps to regulate energeticmetabolism and fat storage (Luo et al. 2003). Incontrast to the mild ERRa knockout phenotype,ERRb-null mutants die during development due todefects in placental formation (Luo et al. 1997).
DOI: 10.1677/jme.1.01976Online version via http://www.endocrinology-journals.org

A M TARRANT and others $ Killifish estrogen receptor-related receptors106
ERRb-null mutants rescued from embryonic lethalityexhibit behavioral abnormalities and reduced numbersof germline cells (Mitsunaga et al. 2004). The role ofERRg has not been elucidated through knockoutexperiments, but high expression has been noted indifferentiating neural tissues (Hermans-Borgmeyer et al.2000). Other proposed target genes for ERRs includelactoferrin, aromatase, small heterodimer partner,endothelial nitric oxide synthase, SULT2A1, and thyroidhormone receptor-a (Yang et al. 1996, Vanacker et al.1998, Zhang & Teng 2000, Sanyal et al. 2002, Sumi &Ignarro 2003, Seely et al. 2005).
In mice, ERRa and ERRg are broadly expressed inadult and embryonic tissues (Bonnelye et al. 1997a,Shigeta et al. 1997, Heard et al. 2000, Hermans-Borgmeyer et al. 2000). In contrast, ERRb has morelimited expression, most notably in a subset of placentalcells during early embryonic development and indeveloping germ cells (Pettersson et al. 1996, Luo et al.1997, Mitsunaga et al. 2004). In the adult, ERRb isexpressed at low levels in a few tissues including kidney,heart, testis, hypothalamus, hippocampus, cerebellum,and prostate (Giguere et al. 1988, Pettersson et al. 1996).ERRa expression can be upregulated by estrogenexposure in some mammalian tissues (Shi et al. 1997,Shigeta et al. 1997, Liu et al. 2003), but regulation of ERRexpression is not well understood.
Examination of teleost genomic databases hasrevealed that fishes contain additional diversity ofERR genes as compared with mammals: six ERR geneshave been identified in the Japanese pufferfish Takifugurubripes (‘fugu’), five in the spotted green pufferfishTetraodon nigroviridis (‘tetraodon’), and five in thezebrafish Danio rerio (Bardet et al. 2002, Bertrand et al.2004). A genome duplication within the teleost lineage(Amores et al. 1998, Taylor et al. 2001, Christoffels et al.2004, Jaillon et al. 2004, Postlethwait et al. 2004) mayaccount for some of the additional ERR diversity, but ithas also been suggested that the ERRd identified inzebrafish and fugu has been secondarily lost frommammals and tetraodon (Bardet et al. 2002).Additional diversity of ERR genes within the teleostlineage is of evolutionary interest, but also provides anopportunity to gain mechanistic insight into mamma-lian ERR genes. In particular, the duplication, degener-ation, complementation hypothesis predicts that themultiple functions of a gene (e.g. a mammalian ERR)may be partitioned between duplicated co-orthologs(Force et al. 1999, Lynch & Force 2000).
The few published studies of ERR function in fishesand invertebrates have provided insight into evolution-ary biology and novel aspects of ERR function. Forexample, in situ hybridization showed that ERRs aredevelopmentally expressed in a segmented pattern inboth the amphioxus (single ERR) and the zebrafish(ERRa, ERRbb, and ERRga) hindbrain, which indicates
Journal of Molecular Endocrinology (2006) 37, 105–120
that a structure similar to a segmented hindbrainpredated the divergence of invertebrates andvertebrates (Bardet et al. 2005b). Knockdown of ERRaexpression using morpholino antisense oligonucleo-tides in zebrafish indicated a novel role for ERR inregulating morphogenic movement during gastrula-tion (Bardet et al. 2005a). ERRs of zebrafish and humanare similar with respect to ligand binding andtransactivation (Bardet et al. 2004); however, furtherinvestigation is needed to understand ERR signaling inteleosts. For example, ERR expression patterns havenot been described in adults of any teleost species andregulation of teleost ERR expression (e.g. in responseto estradiol exposure) has not been described.
In this study, we report the cloning, adult tissue-expression patterns and estrogen responsiveness of ERRcDNAs in the Atlantic killifish, Fundulus heteroclitus.F. heteroclitus has been used as a model species for severalrecent studies of endocrine disruption. In particular,laboratory and natural populations exposed to environ-mental contaminants show altered levels of sex steroids(Dube & MacLatchy 2001, Hewitt et al. 2002, MacLatchyet al. 2003, Boudreau et al. 2004, Greytak et al. 2005),thyroid hormones (Zhou et al. 2000, Carletta et al. 2002),and aromatase mRNA (Greytak et al. 2005). Given thecross-talk between mammalian ERs and ERRs (Vanackeret al. 1999a, Giguere 2002), and the hypothesizedregulatory role of ERRa in aromatase and thyroidreceptor expression (Vanacker et al. 1998, Yang et al.1998), elucidation of ERR signaling may provide insightinto endocrine regulation and disruption in this modelspecies. Specific objectives of this study were: (1) todetermine whether F. heteroclitus contained duplicatedco-orthologs of any mammalian ERR gene, which couldprovide insight into gene function, (2) to compareadult tissue expression patterns of killifish ERR cDNAswith expression patterns reported for mammalianERRs, and (3) to determine whether estradiol exposureaffects killifish ERR cDNA expression, especially theexpression of ERRa.
Materials and methods
Animals and RNA isolation
For cloning of ERR and determination of tissue-specificexpression patterns, F. heteroclitus (Atlantic killifish ormummichog) were trapped in salt marshes surround-ing Scorton Creek on Cape Cod, MA, USA in May andJune 2003. Fish were reproductively active, with matureeggs visible in the ovaries. Three adult male fish andthree adult female fish were anesthetized with MS-222and killed via cervical transection. Liver, gonad, brain,eye, kidney, gill, gut, heart, and spleen were dissectedand pooled for the three fish of a given sex. Total RNA
www.endocrinology-journals.org

Killifish estrogen receptor-related receptors $ A M TARRANT and others 107
was isolated from tissues using RNA STAT-60 (Tel-Test,Inc.). A negative control consisted of a sham RNAextraction with no tissue added.
Two experiments were conducted to determine theeffects of estradiol exposure on ERR mRNA transcriptexpression. In both the experiments, reproductivelyregressed adult F. heteroclitus were collected fromScorton Creek and injected intra-peritoneally withestradiol (5 mg/g body weight, as a 1 mg/ml solution insesame oil) or with a vehicle control (sesame oil). Bothconcentration and method of exposure are predictedto produce a high spike in plasma estradiol concen-tration that is cleared rapidly (Pankhurst et al. 1985)and a robust induction of vitellogenesis in male fish(Pait & Nelson 2003).
In the first experiment, male F. heteroclituswere injectedin November 2003 and sacrificed after 2 or 5 days. Theorgans were immediately flash-frozen, and the total RNAwas extracted subsequently from organs of individual fishusing Tri-reagent (Sigma). Thirteen fish were injected intotal (6 with estradiol and 7 with vehicle), giving a samplesize of 3–4 fish per time point within a treatment.
In the second experiment, adult female F. heteroclituswere injected in January 2005 and sacrificed after2 days. The organs were flash-frozen as in the previousexperiment and total RNA was extracted from organs ofindividual fish with STAT-60. Twenty five fish wereinjected in total (nZ12, estradiol treatment; nZ13,vehicle control).
Reverse transcriptase (RT)-PCR
cDNA was synthesized from 3 mg total RNA usingrandom hexamers and the Omniscript cDNA SynthesisKit (Qiagen). Degenerate oligonucleotide primers,ERRf1 and ERRr1, were designed based on highlyconserved regions of the fugu-predicted ERR genes(Table 1). PCRs with these degenerate primers resultedin two different cDNAs (441 bp F. heteroclitus (Fh)ERRaand 432 bp FhERRba) when used with Advantage2Polymerase (BD Biosciences Clontech) with thefollowing cycling conditions in a Perkin-Elmer Gen-eAmp 2400 thermocycler: 95 8C/60 s (95 8C/30 s,65 8C/45 s, 68 8C/45 s) for 35 cycles, 68 8C/60 s.Additional degenerate primers were targeted towardother ERR genes predicted from the fugu genome.These reactions were conducted using AmpliTaq GoldPolymerase (Applied Biosystems, Framingham, MA,USA). A 398 bp cDNA fragment (FhERRbb) wasamplified using ERRf2 and ERRr1 under the followingconditions: 94 8C/5 min (94 8C/15 s, 62$5 8C/15 s,72 8C/30 s), 72 8C/5 min, followed by (94 8C/15 s,62 8C/15 s, 72 8C/30 s) for 10 cycles, 72 8C/5 min.ERRf3 and ERRr1 amplified a 324 bp fragment(FhERRgb) at 95 8C/10 min (94 8C/15 s, 63$5 8C/15 s, 72 8C/30 s) for 35 cycles, 72 8C/10 min.
www.endocrinology-journals.org
Rapid amplification of cDNA ends (RACE) and
amplification of full-length PCR products
5 0-/3 0-RACE reactions were performed using a SMARTRACE cDNA amplification kit (BD Biosciences Clon-tech). Briefly, adapter-ligated, oligo(dT)-primed cDNAwas produced from brain or liver total RNA. Gene-specific primers were used with adapter primers in PCR.To most RACE PCR, 5% dimethylsulfoxide (DMSO)was added. Touchdown PCRs cycling conditions wereused according to the manufacturer’s instructions withprimers shown in Table 1. For FhERRa 5 0-RACE, primerERRAr1 and nested primer ERRAr2 were used togenerate a partial 5 0-RACE product. To obtain the 5 0
end of FhERRa, primer ERRAr3 was used as a nestedprimer. For FhERRa 3 0-RACE, primer ERRAf1 was used.For FhERRba 5 0-RACE, a partial 5 0-RACE product wasobtained using primer ERRB1r1. To obtain a complete5 0 sequence, additional fragments were amplified usingnested primers ERRB1r2 and ERRB1r3. For FhERRba3 0-RACE and FhERRbb 5 0-RACE, primers ERRB1f1 andERRB2r1 were used, respectively, in two rounds of PCRwith nested adapter primers. For FhERRbb 3 0-RACE,primer ERRB2f1 and nested primer ERRB2f2 wereused. For FhERRgb 5 0-RACE, primer ERRGr1 andnested primer ERRGr2 were used. For FhERRgb3 0-RACE, primer ERRGf1 and nested primer ERRGf2were used.
Once full-length RACE products were obtained,additional gene-specific primers (Table 1) weredesigned within the untranslated regions to amplifyfull-length PCR products for each gene. All full-lengthproducts were amplified using Advantage2 Polymerasewith 5% DMSO added to the reactions. FhERRa,FhERRbb, and FhERRgb were amplified from cDNAmade from brain total RNA using the following cyclingconditions: 94 8C/1 min, 35–37 cycles of (94 8C/s,65 8C/10 s, 68 8C/2 min), 72 8C/7 min. FhERRba wasamplified from cDNA made from brain poly-AC RNAusing the following cycling conditions: 94 8C/1 min, 40cycles of (94 8C/s, 64 8C/10 s, 69 8C/2 min), 72 8C/7 min. During analysis of 5 0-RACE products and full-length cDNA clones for FhERRa, an apparent frameshift was noted in the sequence. When these sequenceswere aligned with the other fish ERRa sequences, itappeared that the FhERRa clones might be missing asection coding for 16 amino acid residues. Usingspecific primers, we amplified a 300 bp product thatoverlapped previous sequences and contained anadditional 56 bp in the frame shift region. The 300 bpproduct and the 56 bp insert had 68 and 80% GCcontent respectively. The high GC content of thisregion is likely to have resulted in the secondarystructure leading to errors in RT-PCR. The completepredicted cDNA sequence is thus a composite offull-length clones with the 56 bp region inserted; the
Journal of Molecular Endocrinology (2006) 37, 105–120

Table 1 Oligonucleotide sequences of primers. In degenerate primers S=C or G, W=A or T, R=A or G
Name Oligonucleotide sequence
ApplicationAmplification of partial cDNA ERRf1 5 0-TCCTCTGAGGCCTGCAARGC-30
ERRf2 5 0-TCCATGCCSAAGAGRCTG-30
ERRf3 5 0-GTCAAGTGCCTGGCTGTGG-3 0
ERRr1 5 0-WGGRATGTGYTTGGCCCAGC-30
5 0 RACE of FhERRa ERRAr1 5 0-CCAGCTCGCGGTCGGCAAGGTCAC-3 0
ERRAr2 5 0-GCACTCGTTGGACGCAGGGCAGC-3 0
ERRAr3 5 0-CACACCAGACACAGCCTCTTGGG-3 0
3 0 RACE of FhERRa ERRAf1 5 0-CACCACCCTGTGTGACCTTGCCG-30
5 0 RACE of FhERRba ERRB1r1 5 0-GCTTTTTTGCTGGTGGAGGAAGAGTG-30
ERRB1r2 5 0-GGGTTGAACATCGGAGGTGAGTC-3 0
ERRB1r3 5 0-TCACAAGAGGCGACCCCGTAGTG-3 0
3 0 RACE of FhERRba ERRB1f1 5 0-GATGCTGAGAATGGGTCTTACCTG-30
5 0 RACE of FhERRbb ERRB2r1 5 0-GGGTTGAACATCGGAGGTGAGTC-3 0
3 0 RACE of FhERRbb ERRB2f1 5 0-TGCTCAAAGAAGGGGTTCGTCTGG-3 0
ERRB2f2 5 0-GGAGGCTGGACACAGAAAACAACC-30
5 0 RACE of FhERRgb ERRGr1 5 0-ACCACCTTGTTTTCCACTGCGTC-30
ERRGr2 5 0-CCAGACGGACGCCTTCTCTCAAC-3 0
3 0 RACE of FhERRgb ERRGf1 5 0-TGTTGAGAGAAGGCGTCCGTCTG-30
ERRGf2 5 0-CGACATCAAGGCTCTGACCACGC-3 0
Cloning of FhERRa ORF ERRAFLf 5 0-CAGGCACACCTGACCTTTGAGTG-3 0
ERRAFLr 5 0-CCCCAATCCCATCTTTATGTCCT-3 0
Cloning of FhERRba ORF ERRB1FLf 5 0-AACAGAAAGCCTGCACGAGT-30
ERRB1FLr 5 0-CGTCTCTGACTGATGATG C-3 0
Cloning of FhERRbb ORF ERRB2FLf 5 0-CCAGGCTTTGTTGTCCAAAT-3 0
ERRB2FLr 5 0-GGGATCCACACAATGAGGAG-30
Cloning of FhERRgb ORF ERRGFLf 5 0-TCCTTGAGACTGACTGACTGC-30
ERRGFLr 5 0-CTAAAATGGACGAATACACCG-30
QPCR ERRAQf 5 0-GAGGGAGTACGTCTCGACAGAG-30
ERRAQr 5 0-CACGATGATGTTGGAAGAACC-3 0
ERRBaQf 5 0-GCATATCCCAGGTTTTTCCAC-3 0
ERRBaQr 5 0-AAACACAATGCTCAGCACCAG-30
ERRBbQf 5 0-TGGTGCGCAAGTACAAGAAG-30
ERRBbQr 5 0-CCTCTATGTGCATGGAGTCTG-30
ERRGbQf 5 0-CAGGGTAACATCGAATACAGC-30
ERRGbQr 5 0-GACGCCTTCTCTCAACATGC-30
FhACrt-F 5 0-TGGAGAAGAGCTACGAGCTCC-30
FhACrt-R 5 0-CCGCAGGACTCCATTCCGAG-3 0
FhEF1-F 5 0-GGGAAAGGGCTCCTTCAAGT-30
FhEF1-R 5 0-ACGCTCGGCCTTCAGCTT-3 0
FhVtg-F 5 0-GAGGATCTGTGCTGATGCAGTTGTG-3 0
FhVtg-R 5 0-GGGTAGAAGGCAGTCTTTCCCAGG-3 0
ORF, open reading frame; QPCR, quantitative real-time PCR
A M TARRANT and others $ Killifish estrogen receptor-related receptors108
location of the insert is marked in Fig. 1. The insert iswell conserved among fishes (not shown) but notbetween fishes and mammals.
Cloning and sequencing
All PCR products were cloned into pGEM-T Easy(Promega). PCR products were sequenced by theUniversity of Maine DNA Sequencing Facility (Orono,ME, USA) or at the Bay Paul Center Sequencing Facility(Marine Biological Laboratory, Woods Hole, MA, USA).Both strands from multiple clones were sequenced toensure accuracy. DNA sequences were analyzed,assembled, and translated using the Wisconsin Package
Journal of Molecular Endocrinology (2006) 37, 105–120
(GCG, Accelrys, Burlington, MA, USA) and BioeditSequence Alignment Editor software (Hall 1999).
Phylogenetic analysis
F. heteroclitus ERR-deduced amino acid sequences werealigned with previously reported ERR sequences fromfishes, mammals, and Drosophila melanogaster usingClustal X 1$81 with default parameters (for accessionnumbers see Table 2). Gaps and the highly variableA/B domain were excluded from phylogenetic analysis.The aligned amino acid sequences were used tocreate phylogenetic trees using maximum parsimonyand distance (minimum evolution) criteria with
www.endocrinology-journals.org

Figure 1 Alignment of F. heteroclitus ERR amino acid sequences. Deduced amino acidsequences of four ERR genes cloned from F. heteroclitus were aligned with three human ERRgenes using ClustalW within Bioedit. Identical residues are shaded. Sixteen amino acid residuesshown in boldface indicate a portion of the FhERRa sequence that was obtained from a separatePCR product. See Results for further information. The start codon of FhERRbb is not known;three potential translation initiation sites are underlined. The DBD (C domain) is enclosed inbrackets, and the start of the LBD (E/F domain) is indicated by an arrow. All GenBank accessionnumbers are given in Table 2.
Killifish estrogen receptor-related receptors $ A M TARRANT and others 109
www.endocrinology-journals.org Journal of Molecular Endocrinology (2006) 37, 105–120

Table 2 ERR gene names, synonyms, and accession numbers. Ensembl predicted proteins from the following databases: human31$35d, Takifugu build 2c, release 31$2f, Tetraodon version 7, Danio release 31$4d. Other Ensembl predicted proteins are abbreviatednames and correspond to older assemblies for comparison with previously published literature (i.e. Bertrand et al. 2004)
Official name Trivial name(s)Pubmed accessionnumber (protein)
Ensembl predictedprotein
‘Other’ Ensemblpredicted protein
SpeciesHuman NR3B1 ERRa, ESRRA NP_004442 ENSP00000000442
NR3B2 ERRb, ESRRB,ERR2, ESRL2
NP_004443 ENSP00000261532
NR3B3 ERRg, ERR3 NP_001429 ENSP00000354584Takifugu ERRa N/A SINFRUP
00000138848SIN67383
ERRba N/A SINFRUP00000140232
SIN72315
ERRbb N/A SINFRUP00000141263
FRUP141263/62880
ERRga N/A SINFRUP00000162788
SIN57192
ERRgb (b/g) N/A SINFRUP00000130211
FRUP130211(51057)
ERRd N/A SINFRUP00000134462
FRUP134462
Tetraodon ERRa N/A GSTENP00020389001
GST20389
ERRba N/A GSTENP00032328001
GST32328
ERRga N/A GSTENP00032849001
GST32848
ERRgb (b/g) N/A GSTENP00030242001
FS563
Danio ERRa, esrral (zfin) AAS66634 ENSDARP00000050433
ERRbb esrrb (zfin) AAS66635 ENSDARP00000040417
ERRga esrrg (zfin) AAS66636 ENSDARP00000005364
ERRgb, ERRb/gesrrgl (zfin)
AAS66638 ENSDARP00000002838
ERRd esrrd (zfin) AAS66637 ENSDARP00000013201
Fundulus ERRa DQ241376 N/AERRba DQ241377 N/AERRbb DQ241378 N/AERRgb (b/g) DQ241379 N/A
Drosophila ERR NP_729340 CG7404 (flybase)
A M TARRANT and others $ Killifish estrogen receptor-related receptors110
PAUP*4.0b10 software (Swofford 2003). The D. melano-gaster ERR was used as the outgroup. Trees wereconstructed with a heuristic search strategy, and branchswapping and tree-bisection reconnection wererepeated to obtain bootstrapping values from 1000replicates.
In an attempt to further resolve the ERRb andERRg clades, a second alignment was created usingfish and mammalian ERRa, ERRb, and ERRgsequences (i.e., no ERRd or invertebrate ERRsequences). Phylogenetic trees were created usingparsimony and minimum evolution criteria, as pre-viously, using ERRa genes as the outgroup. Inaddition, a maximum likelihood tree was constructed
Journal of Molecular Endocrinology (2006) 37, 105–120
using Phylip 3.64 (Felsenstein 2004). Weights corre-sponding to 1000 bootstrap replicates were generatedby SEQBOOT (within Phylip), and a g-law parameter,a, was estimated by PHYML (Guindon et al. 2005).Maximum likelihood trees were constructed using theJones, Taylor and Thornton (JTT) substitution model(Jones et al. 1992) as implemented in ProML (withinPhylip) with a g distribution of rates between sites(four categories). Consensus trees were created byCONSENSE and rooted with the ERRa sequences.Alternative tree topologies were compared with themaximum likelihood consensus tree using the -Shimodaira–Hasegawa test (SH-test; Shimodaira &Hasegawa 1999), as implemented in ProML.
www.endocrinology-journals.org

Killifish estrogen receptor-related receptors $ A M TARRANT and others 111
Nomenclature
We have named teleost co-orthologs (e.g. FhERRba andFhERRbb) to be consistent with zebrafish nomencla-ture rules (Sprague et al. 2001). Similar nomenclaturehas been applied to duplicated teleost ER genes(Hawkins & Thomas 2004).
Quantitative real-time RT-PCR (qPCR)
F. heteroclitus ERR splice sites were predicted bycomparing genomic sequences for human, fugu, andzebrafish ERRs with cDNA sequences (human, zebra-fish) or gene predictions (fugu). Splice sites weregenerally well conserved among species and amongvarious ERR genes within a species (data not shown).Primers for ERRa, ERRba, ERRbb, ERRgb, and b-actin(Table 1) were designed with one primer spanning apredicted exon–exon junction to avoid amplification ofgenomic DNA. Primers for EF-1 (Bears et al. 2006) andvitellogenin (Garcio-Reyero et al. 2004) were taken frompublished studies. cDNA was synthesized from 2 mgtotal RNA using random hexamers and the OmniscriptcDNA Synthesis Kit (Qiagen). In the tissue-distributionstudy, cDNA was diluted in a ratio of 1:3 in ERRbb andEF-1 assays. qPCR was performed using the iQ SYBRGreen Supermix (Bio-Rad) and reactions were run inan iCycler iQ Real-Time PCR Detection System(BioRad). The PCR mixture consisted of the following:11 ml molecular biology grade distilled water, 12$5 ml iQSYBR Green Supermix, 0$25 ml 5 0-primer (10 mM),0$25 ml 5 0-primer (10 mM), and 1 ml cDNA.
In the analysis of tissue distribution of ERRs, the PCRconditions for FhERRa and FhERRbb were:95 8C/3 min, 95 8C/15 s, 66 8C/1 min, 40 cycles. PCRconditions of other genes were identical except thatannealing/extension temperatures were adjusted tomaximize the amplification of the specific product:FhERRba (64 8C), FhERRgb (67$9 8C), EF-1 (60 8C). Atthe end of each PCR cycle, the PCR products weresubjected to melt-curve analysis to ensure that only asingle product was amplified. For both males andfemales, each of the nine tissues was represented by asingle cDNA derived from pooled total RNA from threefish. There were three technical replicates (qPCR well)per sample per gene. Expression data were quantifiedbased on threshold cycle (Ct) values and the –2DDCt
method (Livak & Schittgen, 2001). b-Actin expressionwas highly variable among tissues (data not shown), sofor each ERR gene, Ct values were normalized to EF-1(Bears et al. 2006). Relative mRNA expression for eachgene was calculated as the fold change compared withthe tissue with the lowest Ct (i.e. data were normalizedsuch that the tissue with the highest expression was setequal to one).
www.endocrinology-journals.org
For analysis of qPCR data from dosing experiments, astandard curve for each ERR gene was generated byserially diluting plasmids containing a full-length copy ofeach gene from 103 to 108 molecules/ml. The PCRconditions for FhERRa, FhERRgb, and Fh-b-actinwere: 95 8C/3 min, 95 8C/15 s, 66 8C/1 min, 40 cycles.PCR conditions for FhERRbb were nearly identical:95 8C/3 min, 95 8C/15 s, 64 8C/1 min, 40 cycles. At theend of each PCR cycle, the PCR products were subjectedto melt-curve analysis to ensure that only a single productwas amplified. The number of molecules/ml for eachgene of interest in each RNA sample was calculated fromthe standard curve. ERR expression was presented asunnormalized, and the b-actin expression is shown forcomparison. The data were transformed via the naturallogarithm to obtain a normal distribution (Shapiro–Wilktest and visual inspection of normal probability plots) andequality of variance (visual inspection of residuals) foreach gene. Gene expression in estradiol-treated andcontrol tissues were compared using two-tailed t-tests.
Results
Cloning and phylogenetic analysis of killifish ERRs
Using RT-PCR and 5 0/3 0 RACE, full-length cDNA anddeduced amino acid sequences were determined forfour ERR genes in F. heteroclitus (Fig. 1).
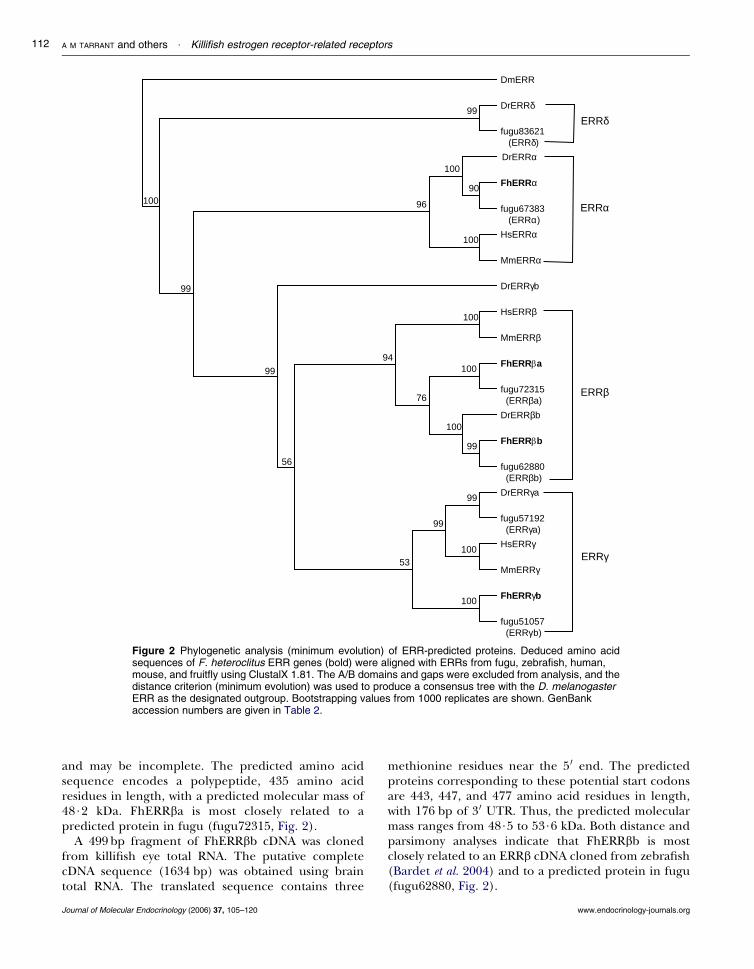
Identical 441 bp fragments of one ERR cDNA(FhERRa) were cloned initially from total RNA derivedfrom killifish liver, brain, kidney, and heart. The full-length ERRa cDNA sequence is 1617 bp in length,including 131 bp 50 untranslated sequence, an openreading frame of 1302 bp, and 185 bp 30 untranslatedsequence including a poly-AC tail. The predicted aminoacid sequence encodes a polypeptide 415 amino acidresidues in length with a predicted molecular mass of45$8 kDa. Phylogenetic analysis using distance(minimum evolution, Fig. 2) or parsimony (not shown)criteria clearly indicate that FhERRa is closely relatedto ERRa genes found in mammals and other speciesof fish.
Each of the mammalian and teleost species includedin this study contains a single ERRa gene. In contrast,the teleost ERRb genes form two clades, which togetherare a sister group to the mammalian ERRb genes(Bertrand et al. 2004, Fig. 2). We have identified twoERRb genes in F. heteroclitus (FhERRba and FhERRbb);these genes are apparent co-orthologs of the mamma-lian ERRb genes. Fragments (429 bp) of FhERRbacDNA were cloned from killifish brain and heart totalRNA, and the full-length cDNA was cloned from brainpoly-AC RNA. The sequence is 1656 bp long, including185 bp of 5 0 untranslated sequence, an open readingframe of 1305 bp, and 166 bp 3 0 untranslated sequence.Available sequence for the 3 0 UTR lacks a poly-AC tail
Journal of Molecular Endocrinology (2006) 37, 105–120
DmERR
DrERRδ
fugu83621 (ERRδ)
99
DrERRα
FhERRα
fugu67383 (ERRα)
90
100
HsERRα
MmERRα
100
96
DrERRγb
HsERRβ
MmERRβ
100
FhERRβa
fugu72315 (ERRβa)
100
DrERRβb
FhERRβb
fugu62880 (ERRβb)
99
100
76
94
DrERRγa
fugu57192 (ERRγa)
99
HsERRγ
MmERRγ
100
99
FhERRγb
fugu51057 (ERRγb)
100
53
56
99
99
100
ERRδ
ERRα
ERRβ
ERRγ
Figure 2 Phylogenetic analysis (minimum evolution) of ERR-predicted proteins. Deduced amino acidsequences of F. heteroclitus ERR genes (bold) were aligned with ERRs from fugu, zebrafish, human,mouse, and fruitfly using ClustalX 1.81. The A/B domains and gaps were excluded from analysis, and thedistance criterion (minimum evolution) was used to produce a consensus tree with the D. melanogasterERR as the designated outgroup. Bootstrapping values from 1000 replicates are shown. GenBankaccession numbers are given in Table 2.
A M TARRANT and others $ Killifish estrogen receptor-related receptors112
and may be incomplete. The predicted amino acidsequence encodes a polypeptide, 435 amino acidresidues in length, with a predicted molecular mass of48$2 kDa. FhERRba is most closely related to apredicted protein in fugu (fugu72315, Fig. 2).
A 499 bp fragment of FhERRbb cDNA was clonedfrom killifish eye total RNA. The putative completecDNA sequence (1634 bp) was obtained using braintotal RNA. The translated sequence contains three
Journal of Molecular Endocrinology (2006) 37, 105–120
methionine residues near the 5 0 end. The predictedproteins corresponding to these potential start codonsare 443, 447, and 477 amino acid residues in length,with 176 bp of 3 0 UTR. Thus, the predicted molecularmass ranges from 48$5 to 53$6 kDa. Both distance andparsimony analyses indicate that FhERRbb is mostclosely related to an ERRb cDNA cloned from zebrafish(Bardet et al. 2004) and to a predicted protein in fugu(fugu62880, Fig. 2).
www.endocrinology-journals.org

Killifish estrogen receptor-related receptors $ A M TARRANT and others 113
A 324 bp fragment of a fourth ERR cDNA, FhERRgbwas cloned from killifish heart cDNA. The cDNAsequence corresponding to the complete coding regionwas obtained by RACE and PCR with gene-specificprimers using brain total RNA. The sequence includes132 bp of 5 0 UTR, 1320 bp coding region, and a partial3 0 UTR of 88 bp. The predicted protein contains 439amino acid residues and the predicted molecular massis 48$3 kDa. Phylogenetic analysis using distancecriterion (minimum evolution) indicates that FhERRgbis a form of ERRg and an ortholog of a predictedprotein in fugu (fugu51057, Fig. 2), but inclusion ofFhERRgb and fugu51057 in the ERRg clade has arelatively low bootstrap support (Fig. 2). Parsimony-based analysis of the same alignment indicates thatthese two fish ERRs form a sister group to the ERRb andERRg clades (not shown), as described for zebrafish(Bardet et al. 2004).
To further investigate the evolutionary relationships,we aligned the full-length vertebrate ERRa, ERRb, andERRg sequences and constructed phylogenetic treeswith distance, parsimony, and maximum likelihoodcriteria (Fig. 3). As in the previous analysis, placementof FhERRgb, fugu51057, and DrERRgb wereequivocal. The parsimony-based analysis placed thethree genes within the ERRg clade (with bootstrapsupport of 54%), but there were polytomies within theclade. The distance tree placed all the three genes assister to the ERRb and ERRg clades, in contrast to theprevious distance analysis (which included ERRds andthe ERR from D. melanogaster Fig. 2), which placed thefish genes within the ERRg clade. The maximumlikelihood consensus tree was consistent with a fish-specific duplication of ERRg, with low bootstrapsupport (Fig. 3), which is similar to the results froma previous study (Bertrand et al. 2004). However, anSH-test showed that the topology indicated by themaximum likelihood tree was not significantly betterthan two alternative topologies: (1) FhERRgb,fugu51057, and DrERRgb grouping outside the cladeformed by mammalian ERRs and teleost ERRga genes(SH-test, PZ0$45) and (2) DrERRgb grouping outsidethe other fish and mammalian ERRgs (PZ0$155). Themaximum likelihood tree was also unable to resolvethe branching patterns within the ERRb clade.
Possible explanations for the ‘outgroup topology’(i.e. fish ERR genes grouping outside the ERRb andERRg clades) include an ancient duplication, orartifacts due to differences in evolutionary rate orsaturation (Van de Peer et al. 2003). We used theprogram AsaturA (Van de Peer et al. 2002) with arange of cut-off values, substitution matrices, anddistance correction methods to explore the effectsof amino acid saturation on the topology of treesmade with distance and parsimony criteria. In nocase did the tree topology provide clear evidence for
www.endocrinology-journals.org
two groups of fish ERRg co-orthologs of mammalianERRg genes, as would be expected if fish ERRgdiversity resulted from a duplication in the teleostlineage. The results from these parsimony- anddistance-based analyses contradict both the predictionsfrom a teleost genome duplication and the weaklysupported results from maximum likelihood analysis.Thus, the evolutionary history of FhERRgb and relatedgenes in fugu and zebrafish remains unresolved. Wehave provisionally named FhERRgb based on thehypothesis that this gene resulted from a teleost-specific duplication of an ancestral ERRg, as suggestedby Bertrand et al. (2004).
Tissue-specific expression
FhERRa, FhERRba, FhERRbb, and FhERRgb tran-scripts were measured by qPCR in tissues from maleand female fish. F. heteroclitus ERR genes showeddistinct, partially overlapping expression patterns(Fig. 4). FhERRa was widely expressed and detectablein all tissues studied. FhERRba was expressed at lowlevels in brain, female eye, and ovary. FhERRbb wasdetected primarily in gonad, eye, brain, and male liver,whereas FhERRgb was detected primarily in kidney, eye,heart, and gill. Males and females showed somedifferences in ERR expression including the ovarian,but not testicular, expression of ERRba. Since eachtissue was represented by a single pooled sample, amore detailed study is needed to determine the sex-specific expression patterns.
Effects of estradiol exposure on ERR expression
In the first experiment, male fish were injected withestradiol, or a vehicle control, and transcriptexpression of ERRa, ERRbb, and ERRgb wasquantified in several tissues using qPCR (Fig. 5).Two days after exposure, expression of ERRa, ERRbb,and ERRgb was not significantly affected by estradioldosage for any tissue (PO0$05), although there was ahigh degree of variability among the samples. Inparticular, 2 days after injection, a single fishdisplayed relatively high levels of ERRa expressionin heart, testis, and gill. Because the same fishshowed elevated b-actin expression in some tissues,statistical analysis was repeated on unnormalized data,but this did not result in any significant differences.The greatest trend toward induction was observed forERRa expression in heart tissue. In comparing themean transcript levels, FhERRa expression was2$6-fold higher in E2-treated normalized heart tissuesrelative to the control. The fish with the highestERRa expression had transcript levels 5$3-fold greaterthan the control mean transcript level. A power
Journal of Molecular Endocrinology (2006) 37, 105–120

MmERRα
HsERRα
99
DrERRα
fugu67383(ERRα)
FhERRa
61
97
93
DrERRγa
fugu57192(ERRγa)
54
DrERRγb
FhERRγb
fugu51057(ERRγb)
97
70
37
MmERRγ
HsERRγ
92
53
DrERRβb
FhERRβb
fugu62880(ERRβb)
62
70
HsERRβ
MmERRβ
98
44
FhERRβa
fugu72315(ERRβa)
95
48
ERRα
ERRγ
ERRβ
Figure 3 Maximum likelihood analysis of ERRa-, ERRb-, and ERRg-predicted proteins. Deduced aminoacid sequences of F. heteroclitus ERR genes (bold) were aligned with ERRs from fugu, zebrafish,human, and mouse using ClustalX 1.81. The A/B domains and gaps were excluded from analysis, andmaximum likelihood criterion was used to produce a consensus tree that was rooted with the ERRasequences. Bootstrapping values from 1000 replicates are shown. GenBank accession numbers aregiven in Table 2.
A M TARRANT and others $ Killifish estrogen receptor-related receptors114
analysis demonstrated that to detect a threefoldinduction in FhERRa expression in heart (aZ0$05,1KbZ0$80), a sample size of 12 fish per treatment wouldhave been needed. In the experiment conducted in thisstudy (nZ3), a 15-fold induction would have beendetected with a power of 0$874 (aZ0$05). Thus, thehigh inter-individual variability of ERR expressionprecludes the detection of modest differences inexpression. We also measured FhERRa and b-actinexpression in fish 5 days after exposure to E2 or a vehicle
Journal of Molecular Endocrinology (2006) 37, 105–120
control. We detected no effect of E2 exposure on FhERRaexpression in these fish (data not shown). In a relatedstudy using qPCR with the same tissues, we have detecteda twofold induction of cytochrome P450 aromatase B(AroB) expression in brain and greater than 100-foldinduction of vitellogenin in the liver (SR Greytak, AMTarrant, ME Hahn & GV Gallard, unpublished obser-vations). Induction of AroB and vitellogenin by estradioldemonstrates that the fish were effectively exposed andnormally responsive to estradiol.
www.endocrinology-journals.org

0
0·2
0·4
0·6
0·8
1·0
Liver Gonad Eye Brain Kidney Gill Gut Heart Spleen
Liver Gonad Eye Brain Kidney Gill Gut Heart Spleen
Liver Gonad Eye Brain Kidney Gill Gut Heart Spleen
Liver Gonad Eye Brain Kidney Gill Gut Heart Spleen0
0·2
0·4
0·6
0·8
1·0
0
0·2
0·4
0·6
0·8
1·0
0
0·2
0·4
0·6
0·8
1·0
Nor
mal
ized
rel
ativ
e ex
pres
sion
Nor
mal
ized
rel
ativ
e ex
pres
sion
A B
DC
Figure 4 Tissue-specific expression of F. heteroclitus ERRs. Relative expression of four F. heteroclitus ERRs ((A) ERRa, (B) ERRba,(C) ERRbb, (D) ERRgb) was measured in cDNAs from adult male (shaded bars) and female fish (open bars) by qPCR, as described inMaterials and methods. Expression of each ERR gene was normalized to EF-1. Relative mRNA expression for each tissue isrepresented as the fold change relative to the expression in the highest-expressing tissue (thus setting maximum relative expressionequal to one). For each sex, relative expression was calculated from the mean of three technical replicates from a single cDNA poolderived from three fish.
Killifish estrogen receptor-related receptors $ A M TARRANT and others 115
In a second more targeted experiment, ERRatranscript expression was quantified in the heartsfrom female fish injected with estradiol or a vehiclecontrol (Fig. 6). Two days after exposure, expression ofERRa was downregulated 2$5-fold in heart relative tothe control (PZ0$001). Liver from a subset of these fishshowed a greater than 100-fold induction of vitello-genin (data not shown), demonstrating the effective-ness of the estrogen exposure.
Discussion
ERR diversity in F. heteroclitus
Examination of genomic databases and cloning effortshave demonstrated that teleost fish have additionaldiversity of ERR genes relative to mammals (Maglichet al. 2003, Bardet et al. 2004, Bertrand et al. 2004, thisstudy). Increased diversity of nuclear receptors andother genes has been attributed to frequent geneduplication or to a genome duplication within theteleost lineage (Robinson-Rechavi et al. 2001a,b, Tayloret al. 2003). In zebrafish, the five ERR genes are each onseparate chromosomes, as predicted from version 4 of
www.endocrinology-journals.org
the genome assembly. (In version 5 of the assembly, thezebrafish ERRb is on a scaffold that has not beenmapped to a chromosome.) In tetraodon, two genes(GSTENG00030324001, an ortholog of FhERRba, andGSTENG00030242001, an ortholog of FhERRgb) areboth on chromosome 14, but they are separated byapproximately 800 kb. Thus, the additional ERRdiversity observed in teleosts cannot be explained byrecent tandem duplication events.
In the present study, we have identified four ERR genesfrom F. heteroclitus. These genes are predicted to beorthologs of four of the six ERR genes predicted from thefugu genome. Without a fully sequenced genome, it is notpossible to know whether we have identified the fullcomplement of ERR genes in F. heteroclitus. In our cloningefforts, we did not identify orthologs of ERRga or ERRdgenes, which are present in pufferfish genomes andexpressed in zebrafish embryos (Bardet et al. 2004,Bertrand et al. 2004). These genes may have been lostfrom F. heteroclitus or may have been difficult to detect,possibly due to the low expression in the adult tissuesexamined. Among the four killifish ERRs we identified,FhERRa is an ortholog of the single ERRa identified inmammals, pufferfish, and zebrafish. FhERRba and
Journal of Molecular Endocrinology (2006) 37, 105–120

0·0E+00
5·0E+03
1·0E+04
1·5E+04
2·0E+04
2·5E+04
3·0E+04
0·0E+00
5·0E+03
1·0E+04
1·5E+04
2·0E+04
2·5E+04
3·0E+04
Liver Heart Testis Gill
Liver Heart Testis Gill
0·0E+00
5·0E+04
1·0E+05
1·5E+05
2·0E+05
2·5E+05
3·0E+05
0·0E+00
2·0E+07
4·0E+07
6·0E+07
8·0E+07
Liver Heart Testis Gill Brain
Liver Heart Testis Gill Brain
A
B
C
D
Figure 5 Effects of estradiol treatment on ERR expression in F. heteroclitusmales. Adult male F. heteroclitus were injected intra-peritoneally with avehicle control (open bars) or with 5 mg/g body weight E2 (shaded bars) andsacrificed after 2 days. Tissue-specific expression of: b-actin (A), ERRa (B),ERRbb (C), and ERRgb (D) was measured by qPCR. Error bars representS.D. (nZ3 fish). No significant effects (aZ0$05) of E2 treatment weredetected (see the text for further details).
A M TARRANT and others $ Killifish estrogen receptor-related receptors116
Journal of Molecular Endocrinology (2006) 37, 105–120 www.endocrinology-journals.org

0·0E+00
1·0E+04
2·0E+04
3·0E+04
4·0E+04
Control E2
FhERRα
Control E2
Actin
0·0E+00
2·0E+06
4·0E+06
6·0E+06
8·0E+06
1·0E+07
Figure 6 Effects of estradiol treatment on ERRa expression in F. heteroclitus female heart. Adult female F.heteroclituswere injected intra-peritoneally with a vehicle control (open bars) or with 5 mg/g body weight E2
(shaded bars) and sacrificed after 2 days. Expression of ERRa and b-actin was measured by qPCR. Errorbars represent S.D. (nZ12 estradiol treatment, nZ13 vehicle control). ERRa expression was 2$5-foldlower in estradiol-treated fish relative to fish injected with a vehicle control (PZ0$001; see the text forfurther details).
Killifish estrogen receptor-related receptors $ A M TARRANT and others 117
FhERRbb are co-orthologs of the mammalian ERRb, asindicated by parsimony and distance analysis. Similarly,duplicated ERRb genes have been identified in the fugu,tetraodon, and medaka genomes (Bertrand et al. 2004,AM Tarrant, unpublished data). In contrast, only a singleERRb, similar to FhERRbb, is present in the zebrafish(Bardet et al. 2004); however, the tree topology isconsistent with the loss of an ERRba-like gene from thezebrafish lineage.
The phylogenetic placement of FhERRgb is less clear.Fugu and zebrafish each have one gene that groupsclearly as an ERRg and a second gene that groups as anERRg or within a sister group to the ERRb and ERRgclades, depending on the analysis. Our likelihoodanalysis and a previously published likelihood analysis(Bertrand et al. 2004) are consistent with the hypothesisthat there has been a duplication of ERRg within theteleost lineage. However, bootstrap support for thegrouping of two fish ERRg genes is low in both analyses.We have tentatively identified these groups of fish genesas ERRga and ERRgb. FhERRgb is an apparentortholog of this second group of fish genes.
Translation of the FhERRbb cDNA sequence revealedthree methionine residues near the N-terminus of thepredicted protein, and it is not clear which of theseresidues represents the translation initiation site(s).The third methionine aligns with the predicted startsite of most other ERRs, including HsERRb andFhERRba, and this methionine has an adenine at the–3 position, the most conserved position in the Kozakconsensus sequence (Kozak 1987). We did not detectmultiple potential start codons of FhERRba, andsimilarly did not detect multiple potential start codonsin genomic sequences corresponding to other fishERRb genes. The predicted ERRb sequence for the
www.endocrinology-journals.org
chimpanzee (GenBank accession number XP510082)similarly has an additional methionine upstream of thepredicted start site for most ERRb genes, and thesequence near the N terminus is highly similar toFhERRba. It is possible that multiple initiation sites areused for some ERRb genes. Indeed, apparent isoformsof HsERRg that use different initiation sites have beendescribed based on cDNA sequences obtained fromdifferent tissues (Heard et al. 2000). Further, appli-cation of several specific antibodies revealed thatmammalian glucocorticoid-receptor mRNAs producemultiple functional isoforms that differ primarily in theN-termini and have distinct expression patterns andfunctional properties (Lu & Cidlowski 2005). Thebiological significance of multiple predicted start siteswithin the FhERRbb sequence is currently unknown.
Tissue-specific expression
This study contains the first description of spatialpatterns of ERR-transcript expression in adult fish. Likeits mammalian ortholog, FhERRa was broadlyexpressed and detectable in all tissues. FhERRba wasapparently expressed at low levels in the eye, brain, andovary. FhERRbb was detected primarily in the liver,gonad, eye, brain, and kidney, and FhERRgb in the eye,kidney, gill, and heart. While expression patterns weresimilar between males and females, some differencesare apparent, such as expression of ERRba in femaleeye and ovary. In the initial tissue comparison, thetissues from male and female fish are each representedby a single cDNA, and therefore, a more detailedanalysis with multiple independent samples throughoutthe reproductive cycle will be needed to robustlycompare expression patterns.
Journal of Molecular Endocrinology (2006) 37, 105–120

A M TARRANT and others $ Killifish estrogen receptor-related receptors118
The ERR expression patterns indicate the utility ofF. heteroclitus and other fish models for the character-ization of ERR function in future. For example,FhERRba and FhERRbb have distinct spatial expressionpatterns, which may indicate subfunctionalization ofthe co-orthologs. Thus, F. heteroclitus may serve as aparticularly useful model for dissecting the ERRbfunction. Mammalian ERRb helps regulate placentaldevelopment and primordial germ cell proliferation.Expression of FhERRba in ovary is particularly interest-ing in this respect and is consistent with some role ofERRb in killifish reproduction. The role of ERRb inteleost development is unknown and would also beinteresting to study, particularly given the differences inextraembryonic tissues between fishes and mammals.To give a second example, ERRg function remainspoorly characterized in any organism, and mouseknockout phenotypes have not been described. Theadditional diversity of ERRg-like genes in teleosts, suchas zebrafish, may facilitate characterization of functionthrough knockdown experiments. In addition,FhERRgb expression in gill might indicate uniquefunction relative to mammalian ERRs.
Effects of estradiol dosage on ERR expression
In the first experiment, we detected no significanteffects of exposure to E2 (5 mg/g body weight) on ERRgene expression in any tissue. We did observesubstantial variability among individual fishes and atrend towards a slight induction (three- to five-fold) ofERRa expression in heart. In a more targetedexperiment with female fish, we observed a highlysignificant 2$5-fold downregulation of ERRa expressionin heart following estrogen exposure. This downregula-tion contrasts with reports of ERRa induction byestrogens in some mammalian tissues (Shigeta et al.1997, Liu et al. 2003).
The effect of estrogen exposure on mammalianERRa expression is primarily mediated throughmultiple steroid hormone-response element half-sitesthat are conserved between the human and mouseERRa gene promoters (Liu et al. 2003). While fish ERRgene promoters have not yet been characterized fully, inpreliminary searches of teleost genomic databases, wehave not identified any of the predicted ER responseelements upstream of fish ERR genes (data not shown).Downregulation of ERRa in female heart followingexposure to estradiol may indicate an importantdifference in regulation of expression between teleostsand mammals. This difference warrants further inves-tigation and may provide an opportunity to identifyestrogen-independent pathways of ERR expression. Forexample, ERRa expression in some mouse tissuesdisplays circadian rhythmicity (Horard et al. 2004).
Journal of Molecular Endocrinology (2006) 37, 105–120
In conclusion, we have identified four ERR genes inF. heteroclitus. Phylogenetic analysis of our sequencesand other teleost ERRs indicates that fishes possessadditional diversity of ERRs relative to mammals andspecifically that F. heteroclitus contains co-orthologs ofERRb. Further, characterization of the duplicated genesmay provide insight into conserved or teleost-specificfunctions of ERRb. Downregulation of ERRa in thefemale heart by estradiol in our study also suggests thatERR expression is regulated differently in fishes andmammals.
Acknowledgements
We thank Ann Michelle Morrison for F. heteroclitus RNAsamples used to measure tissue-specific expression.Sibel Karchner, Diana Franks, and Brad Evans assistedwith the development of qPCR assays. Jed Goldstone,Rob Jennings, and Ken Halanych provided advice onmaximum likelihood analysis.
Funding
This research was supported by the National Institutesof Health under Ruth L Kirschstein National ResearchService Award (F32 ES013092-01) from the NationalInstitute of Environmental Health Sciences, the SewardJohnson Foundation and the Superfund Basic ResearchProgram (P42ES007381). The authors declare thatthere is no conflict of interest that would prejudicethe impartiality of this work.
References
Amores A, Force A, Yan Y, Joly L, Amemiya C, Fritz A, Ho R,Langeland J, Prince V, Wang Y et al. 1998 Zebrafish hox clustersand vertebrate genome evolution. Science 282 1711–1714.
Bardet P-L, Horard B, Robinson-Rechavi M, Laudet V & Vanacker J-M2002 Characterization of oestrogen receptors in zebrafish (Daniorerio). Journal of Molecular Endocrinology 28 153–163.
Bardet P-L, Obrect-Pflumio S, Thisse C, Laudet V, Thisse B &Vanacker J-M 2004 Cloning and developmental expression offive estrogen-receptor related genes in the zebrafish. DevelopmentGenes and Evolution 214 240–249.
Bardet P-L, Horard B, Laudet V & Vanacker J-M 2005a The ERRaorphan nuclear receptor controls morphogenetic movementsduring zebrafish gastrulation. Developmental Biology 281 102–111.
Bardet P-L, Schubert M, Horard B, Holland L, Laudet V, Holland N &Vanacker J-M 2005b Expression of estrogen-receptor relatedreceptors in amphioxus and zebrafish: implications for theevolution of posterior brain segmentation at the invertebrate-to-vertebrate transition. Evolution and Development 7 223–233.
Bears H, Richards J & Schulte P 2006 Arsenic exposure alters hepaticarsenic species composition and stress-mediated gene expression inthe common killifish (Fundulus heteroclitus). Aquatic Toxicology 77257–266.
www.endocrinology-journals.org

Killifish estrogen receptor-related receptors $ A M TARRANT and others 119
Bertrand S, Brunet F, Escriva H, Parmentier G, Laudet V &Robinson-Rechavi M 2004 Evolutionary genomics of nuclearreceptors: from twenty-five ancestral genes to derived endocrinesystems. Molecular Biology and Evolution 21 1923–1937.
Bonnelye E, Vanacker J, Spruyt N, Alric S, Fournier B, Desbiens X& Laudet V 1997a Expression of the estrogen-related receptor 1(ERR-1) orphan receptor during mouse development.Mechanisms of Development 65 71–85.
Bonnelye E, Vanacker J, Dittmar T, Begue A, Desbiens X, Denhardt D,Aubin J, Laudet V & Fournier B 1997b The ERR-1 orphan receptoris a transcriptional activator expressed during bone development.Molecular Endocrinology 11 905–916.
Bonnelye E, Merdad L, Kung V & Aubin J 2001 The orphan nuclearestrogen receptor-related receptor a (ERRa) is expressedthroughout osteoblast differentiation and regulates bone formationin vitro. Journal of Cell Biology 153 971–983.
Boudreau M, Courtenay S, MacLatchy D, Berube C, Parrott J & Van derKraak G 2004 Utility of morphological abnormalities during early-life development of the estuarine mummichog, Fundulus heteroclitusas an indicator of estrogenic and antiestrogenic endocrinedisruption. Environmental Toxicology and Chemistry 23 415–425.
Carletta M, Weis P & Weis J 2002 Development of thyroidabnormalities in mummichogs, Fundulus heteroclitus, from apolluted site. Marine Environmental Research 54 601–604.
Chen F, Zhang Q, McDonald T, Davidoff M, Bailey W, Bai C,Liu Q & Caskey C 1999 Identification of two hERR2-relatednovel nuclear receptors utlizing bioinformatics and inversePCR. Gene 228 101–109.
Christoffels A, Koh E, Chia J, Brenner S, Aparicio S & Venkatesh B2004 Fugu genome analysis provides evidence for a whole-genomeduplication early during the evolution of ray-finned fishes. MolecularBiology and Evolution 21 1146–1151.
Coward P, Lee D, Hull MV & Lehmann JM 2001 4-Hydroxytamoxifenbinds to and deactivates the estrogen-related receptor g. PNAS 988880–8884.
Dube M & MacLatchy D 2001 Identification and treatment of a wastestream at a bleached-kraft pulp mill that depresses a sex steroid inthe mummichog (Fundulus heteroclitus). Environmental Toxicology andChemistry 20 985–995.
Felsenstein J 2004 PHYLIP (Phylogeny Inference Package) version 3.64,Department of Genome Sciences and Department of Biology,University of Washington, Seattle, WH (distributed by the author;http://evolution.genetics.washington.edu/phylip.html).
Force A, Lynch M, Pickett F, Amores A, Yan Y & Postlethwait J 1999Preservation of duplicate genes by complementary, degenerativemutations. Genetics 151 1531–1545.
Garcio-Reyero N, Raldua D, Quiros L, Llaveria G, Cerda J, Barcelo D,Grimalt J & Pina B 2004 Use of vitellogenin mRNA as a biomarkerfor endocrine disruption in feral and cultured fish. Analytical andBioanalytical Chemistry 378 670–675.
Giguere V 2002 To ERR in the estrogen pathway. Trends in Endocrinologyand Metabolism 13 220–225.
Giguere V, Yang N, Segui P & Evans R 1988 Identification of a new classof steroid hormone receptors. Nature 331 91–94.
Greschik H, Wurtz J-M, Sanglier S, Bourguet W, van Dorsselaer A,Moras D & Renaud J-P 2002 Structural and functional evidence forligand-independent transcriptional activation by the estrogen-related receptor 3. Molecular Cell 9 303–313.
Greytak S, Champlin D & Callard G 2005 Isolation and characterizationof two cytochrome P450 aromatase forms in killifish (Fundulusheteroclitus): differential expression in fish from polluted andunpolluted environments. Aquatic Toxicology 71 371–389.
Guindon S, Lethiec F, Duroux P & Gascuel O 2005 PHYML Online–aweb server for fast maximum likelihood-based phylogeneticinference. Nucleic Acids Research 33 W557–W559.
Hall TA 1999 Bioedit: a user-friendly biological sequence alignmenteditor and analysis program for Windows 95/98/NT. Nucleic AcidsSymposium Series 41 95–98.
www.endocrinology-journals.org
Hawkins MB & Thomas P 2004 The unusual binding properties of thethird distinct teleost estrogen receptor subtype ERba are accom-panied by highly conserved amino acid changes in the ligandbinding domain. Endocrinology 145 2968–2977.
Heard D, Norby P, Holloway J & Vissing H 2000 Human ERRg, a thirdmember of the estrogen receptor-related receptor (ERR) subfamilyof orphan nuclear receptors: tissue-specific isoforms are expressedduring development and in the adult. Molecular Endocrinology 14382–392.
Hermans-Borgmeyer I, Susens U & Borgmeyer U 2000 Developmentalexpression of the estrogen receptor-related receptor g in thenervous system during mouse embryogenesis. Mechanisms ofDevelopment 97 197–199.
Hewitt L, Smyth S, Dube M, Gilman C & MacLatchy D 2002 Isolation ofcompounds from bleached kraft mill recovery condensatesassociated with reduced levels of testosterone in mummichog(Fundulus heteroclitus). Environmental Toxicology and Chemistry 211359–1367.
Hong H, Yang L & Stallcup M 1999 Hormone-independenttranscriptional activation and coactivator binding by novel orphanreceptor ERR3. Journal of Biological Chemistry 274 22618–22626.
Horard B, Rayet B, Triqueneaux G, Laudet V, Delaunay F &Vanacker J-M 2004 Expression of the orphan nuclear receptorERRa is under circadian regulation in estrogen-responsivetissues. Journal of Molecular Endocrinology 33 87–97.
Ichida M, Nemoto S & Finkel T 2002 Identification of a specificmolecular repressor of the nuclear coactivator PGC-1a. Journal ofBiological Chemistry 277 50991–50995.
Jaillon O, Aury J-M, Brunet F, Petit J-L, Stange-Thomann N, Mauceli E,Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A et al. 2004 Genomeduplication in the teleost fish Tetraodon nigroviridis reveals the earlyvertebrate proto-karyotype. Nature 431 946–957.
Jones DT, Taylor WR & Thornton JM 1992 The rapid generation ofmutation data matrices from protein sequences. Computer Appli-cations in the Biosciences 8 275–282.
Kozak M 1987 An analysis of 5 0-noncoding sequences from 699vertebrate messenger RNAs. Nucleic Acids Research 25 8125–8148.
Liu D, Zhang Z, Gladwell W & Teng C 2003 Estrogen stimulatesestrogen-related receptor alpha gene expression through con-served hormone response elements. Endocrinology 144 4894–4904.
Livak KJ & Schittgen TD 2001 Analysis of relative gene expression datausing real-time quantitative PCR and the 2(-DDC(T)) method.Methods 25 402–408.
Lu N & Cidlowski J 2005 Translational regulatory mechanismsgenerate N-terminal glucocorticoid receptor isoforms with uniquetranscriptional target genes. Molecular Cell 18 331–342.
Luo J, Sladek R, Bader J-A, Matthyssen A, Rossant J & Giguere V 1997Placental abnormalities in mouse embryos lacking the orphannuclear receptor ERR-b. Nature 388 778–782.
Luo J, Sladek R, Carrier J, Bader J-A, Richard D & Giguere V 2003Reduced fat mass in mice lacking orphan nuclear receptor estrogen-related receptor a. Molecular and Cellular Biology 23 7947–7956.
Lynch M & Force A 2000 The probability of duplicate genepreservation by subfunctionalization. Genetics 154 459–473.
MacLatchy D, Courtenay S, Rice C & Van der Kraak G 2003Development of a short-term reproductive endocrine bioassay usingsteroid hormone and vitellogenin end points in the estuarinemummichog (Fundulus heteroclitus). Environmental Toxicology andChemistry 22 996–1008.
Maglich J, Caravella J, Lambert M, Willson T, Moore J & Ramamurthy L2003 The first completed genome sequence from a teleost fish(Fugu rubripes) adds significant diversity to the nuclear receptorsuperfamily. Nucleic Acids Research 31 4051–4058.
Mitsunaga K, Araki K, Mizusaki J, Morohashi K-I, Haruna K, Nakagata N,Giguere V, Yamamura K-I & Abe K 2004 Loss of PGC-specificexpression of the orphan nuclear receptor ERR-b results in reductionof germ cell number in mouse embryos.Mechanisms of Development 121237–246.
Journal of Molecular Endocrinology (2006) 37, 105–120

A M TARRANT and others $ Killifish estrogen receptor-related receptors120
Pait A & Nelson J 2003 Vitellogenesis in male Fundulus heteroclitus(killifish) induced by selected estrogenic compounds. AquaticToxicology 64 331–342.
Pankhurst N, Stacey N & Peter R 1985 An evaluation of techniquesfor the administration of 17b-estradiol to teleosts. Aquaculture 52145–155.
Pettersson K, Svensson K, Mattsson R, Carlsson B, Ohlsson R &Berkenstam A 1996 Expression of a novel member of estrogenresponse element-binding nuclear receptors is restricted to theearly stages of chorion formation during mouse embryogenesis.Mechanisms of Development 54 211–223.
Postlethwait J, Amores A, Cresko W, Singer A & Yan Y 2004Subfunction partitioning, the teleost radiation and the annotationof the human genome. Trends in Genetics 20 481–490.
Robinson-Rechavi M, Marchand O, Escriva H & Laudet V 2001a Anancestral whole-genome duplication may not have been respon-sible for the abundance of duplicated fish genes. Current Biology 11R458–R459.
Robinson-Rechavi M, Marchand O, Escriva H, Bardet P-L, Zelus D,Hughes S & Laudet V 2001b Euteleost fish genomes arecharacterized by expansion of gene families. Genome Research 11781–788.
Sanyal S, Kim J-Y, Kim H-J, Takeda J, Lee Y-K, Moore D & Choi H-S 2002Differential regulation of the orphan nuclear receptor smallheterodimer partner (SHP) gene promoter by orphan nuclearreceptor ERR isoforms. Journal of Biological Chemistry 277 1739–1748.
Seely J, Amigh K, Suzuki T, Mayhew B, Sasano H, Giguere V, Laganiere J,Carr B & Rainey W 2005 Transcriptional regulation of dehydroe-piandrosterone sulfotransferase (SULT2A1) by estrogen-relatedreceptor a (ERRa). Endocrinology 146 3605–3613.
Shi H, Shigeta H, Yang N, Fu K, O’Brian G & Teng C 1997 Humanestrogen receptor-like 1 (ESRL1) gene: genomic organization,chromosomal localization, and promoter characterization. Genomics44 52–60.
Shigeta H, Zuo W, Yang N, DiAugustine R & Teng C 1997 Themouse estrogen receptor-related orphan receptor alpha 1:molecular cloning and estrogen responsiveness. Journal ofMolecular Endocrinology 19 299–309.
Shimodaira H & Hasegawa M 1999 Multiple comparisons of log-likelihoods with applications to phylogenetic inference. MolecularBiology and Evolution 16 1114–1116.
Sprague J, Doerry E, Douglas S, Westerfield M & Group TZ 2001The zebrafish information network (ZFIN): a resource forgenetic, genomic and developmental research. Nucleic AcidsResearch 29 87–90.
Sumi D & Ignarro L 2003 Estrogen-related receptor a1 up-regulatesendothelial nitric oxide synthase expression. PNAS 100 14451–14456.
Swofford D 2003 PAUP*: Phylogenetic Analysis Using Parsimony (and OtherMethods). Version 4.0b8. Sinauer Associates, Sunderland, MA.
Taylor J, Van de Peer Y, Braasch I & Meyer A 2001 Comparativegenomics provides evidence for an ancient genome duplicationevent in fish. Philosophical Transactions of the Royal Society B BiologicalSciences 356 1661–1679.
Journal of Molecular Endocrinology (2006) 37, 105–120
Taylor J, Braasch I, Frickey T, Meyer A & Van de Peer Y 2003 Genomeduplication, a trait shared by 22,000 species of ray-finned fish.Genome Research 13 382–390.
Tremblay G, Bergeron D & Giguere V 2001a 4-Hydroxytamoxifen is anisoform-specific inhibitor of orphan estrogen-receptor-related(ERR) nuclear receptors b and g. Endocrinology 142 4572–4575.
Tremblay G, Kunath T, Bergeron D, Lapointe L, Champigny C,Bader J-A, Rossant J & Giguere V 2001b Diethystilbestrolregulates trophoblast stem cell differentiation as a ligand oforphan nuclear receptor ERRb. Genes and Development 15833–835.
Van de Peer Y, Frickey T, Taylor JS & Meyer A 2002 Dealing withsaturation at the amino acid level: a case study based on ancientlyduplicated zebrafish genes. Gene 295 205–211.
Van de Peer Y, Taylor JS & Meyer A 2003 Are all fishes ancientpolyploids? Journal of Structural and Functional Genomics 2 65–73.
Vanacker J-M, Bonnelye E, Delmarre C & Laudet V 1998 Activation ofthe thyroid hormone receptor a gene promoter by the orphannuclear receptor ERRa. Oncogene 17 2429–2435.
Vanacker J-M, Pettersson K, Gustafsson J-A & Laudet V 1999aTranscriptional targets shared by estrogen receptor-relatedreceptors (ERRs) and estrogen receptor (ER)a, but not by ERb.EMBO Journal 18 4270–4279.
Vanacker J-M, Petterson K, Gustafsson J-A & Laudet V 1999bTranscriptional activities of the orphan nuclear receptor ERR alpha(estrogen receptor-related receptor-alpha). Molecular Endocrinology13 764–773.
Xie W, Hong H, Yang N, Lin R, Simon C, Stalcup M & Evans R 1999Constitutive activation of transcription and binding of coactivatorby estrogen-related receptors 1 and 2. Molecular and CellularEndocrinology 13 1594–1604.
Yang C, Shigeta H, Shi H & Teng C 1996 Estrogen-related receptor,hERR1, modulates estrogen receptor-mediated response ofhuman lactoferrin gene promoter. Journal of Biological Chemistry 2715795–5804.
Yang C, Zhou D & Chen S 1998 Modulation of aromatase expression inthe breast tissue by ERR a-1 orphan receptor. Cancer Research 585695–5700.
Zhang Z & Teng C 2000 Estrogen receptor-related receptor-a1interacts with coactivator and constitutively activates the estrogenresponse elements of the human lactoferrin gene. Journal ofBiological Chemistry 275 20837–20846.
Zhou T, John-Alder H, Weis J & Weis P 2000 Endocrine disruption:thyroid dysfunction in mummichogs (Fundulus heteroclitus) from apolluted habitat. Marine Environmental Research 50 393–397.
Received in final form 1 May 2006Accepted 8 May 2006Made available online as an Accepted Preprint 9 May 2006
www.endocrinology-journals.org
Copyright © 2022 FDOKUMEN




















