
Gender differences and estrogen effects in parkin null mice
15
*Department of Neurobiology, Hospital ‘Ramo ´n y Cajal’ and CIBERNED, Madrid, Spain Department of Neurology, Hospital ‘Ramo ´n y Cajal’ and CIBERNED, Madrid, Spain Estrogens have neurotrophic effects on midbrain dopamine (DA) neurons, promoting the growth of tyrosine hydrox- ylase positive (TH+) neurites, increasing the TH expression in the neonate mouse midbrain and enhancing differenti- ation and survival of dopaminergic neurons derived from human neuronal stem cells (Reisert et al. 1987; Beyer and Karolczak 2000; Sawada and Shimohama 2000; Ivanova and Beyer 2003; Kishi et al. 2005). In addition, in vitro and in vivo models of Parkinson’s disease (PD), estrogens protect DA cells from toxic compounds such as MPTP (Disshon and Dluzen 1997; Callier et al. 2000; Bains et al. 2007), meta-amphetamine (Gajjar et al. 2003), and 6-OH- DA (Dluzen 1997). Estrogen-mediated neuroprotective effects may have clinical relevance in PD, as this disease occurs 1.5 times more frequently in men than in women (Fahn and Sulzer 2004; Wooten et al. 2004; Haaxma et al. 2007); and in women, the age at onset of PD correlates with the duration of the fertile life (Ragonese et al. 2006; Rocca et al. 2008). Cellular responses to estrogen are tightly controlled. A large number of estrogen receptor (ER) interacting proteins, which behave as co-activators or co-repressors, modify the ERa transcriptional activity (Metivier et al. 2003; Reid et al. Received March 14, 2008; revised manuscript received June 18, 2008; accepted July 8, 2008. Address correspondence and reprint requests to Dr M. A. Mena, Departamento de Investigacio ´ n, Hospital Ramo ´n y Cajal, Ctra. de Col- menar, Km. 9, Madrid 28034, Spain. E-mail: [email protected] Abbreviations used: AKT, serine/threonine kinase protein; BRCA1, breast cancer suppressor protein; CHIP, C-terminal Hsc-interacting protein; DA, dopamine; E2, estradiol; E6-AP, E6-associated protein; EFP, estrogen responsive finger protein; ERa, estrogen receptor-a; ERK, extracellular signal-regulated kinase; GFAP, glial fibrillary acidic pro- tein; GPx, glutathione peroxidase; GSK-3, glycogen synthase kinase-3; HSP, heat-shock protein; MAP-2, microtubule associated protein 2a+2b; MAPK, mitogen-activated protein kinase; PD, Parkinson’s disease; PI3K, phosphatidylinositol 3¢-kinase; PK)/), parkin null mice; RING, really interesting new gene; SDS, sodium dodecyl sulfate; TH, tyrosine hydroxylase; WT, wild-type. Abstract Estrogens are considered neurotrophic for dopamine neurons. Parkinson’s disease is more frequent in males than in females, and more prevalent in females with short reproduc- tive life. Estrogens are neuroprotective against neurotoxic agents for dopamine neurons in vivo and in vitro. Here, we have investigated the role of estrogens in wild-type (WT) and parkin null mice (PK)/)). WT mice present sexual dimor- phisms in neuroprotective mechanisms (Bcl-2/Bax, chaper- ones, and GSH), but some of these inter-sex differences disappear in PK)/). Tyrosine hydroxylase (TH) protein and TH+ cells decreased earlier and more severely in female than in male PK)/) mice. Neuronal cultures from midbrain of WT and PK)/) mice were treated with estradiol from 10 min to 48 h. Short-term treatments activated the mitogen-activated protein kinase pathway of WT and PK)/) neurons and the phosphatidylinositol 3¢-kinase/AKT/glycogen synthase kinase-3 pathway of WT but not of PK)/) cultures. Long-term treat- ments with estradiol increased the number of TH+ neurons, the TH expression, and the extension of neurites, and de- creased the level of apoptosis, the expression of glial fibrillary acidic protein, and the number of microglial cells in WT but not in PK)/) cultures. The levels of estrogen receptor-a were elevated in midbrain cultures and in the striatum of adult PK)/) male mice, suggesting that suppression of parkin changes the estrogen receptor-a turnover. From our data, it appears that parkin participates in the cellular estrogen response which could be of interest in the management of parkin-related Parkinson’s disease patients. Keywords: C-terminal Hsc-interacting protein, dopamine neurons, estradiol, estrogen receptor-a, glial cells, heat-shock protein70, mitogen-activated protein kinase, Park-2 gene, Parkinson’s disease and gender differences, phosphatidyl- inositol 3¢-kinase/AKT/glycogen synthase kinase-3. J. Neurochem. (2008) 106, 2143–2157. JOURNAL OF NEUROCHEMISTRY | 2008 | 106 | 2143–2157 doi: 10.1111/j.1471-4159.2008.05569.x ȑ 2008 The Authors Journal Compilation ȑ 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157 2143
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Gender differences and estrogen effects in parkin null mice

*Department of Neurobiology, Hospital ‘Ramon y Cajal’ and CIBERNED, Madrid, Spain
�Department of Neurology, Hospital ‘Ramon y Cajal’ and CIBERNED, Madrid, Spain
Estrogens have neurotrophic effects on midbrain dopamine(DA) neurons, promoting the growth of tyrosine hydrox-ylase positive (TH+) neurites, increasing the TH expressionin the neonate mouse midbrain and enhancing differenti-ation and survival of dopaminergic neurons derived fromhuman neuronal stem cells (Reisert et al. 1987; Beyer andKarolczak 2000; Sawada and Shimohama 2000; Ivanovaand Beyer 2003; Kishi et al. 2005). In addition, in vitroand in vivo models of Parkinson’s disease (PD), estrogensprotect DA cells from toxic compounds such as MPTP(Disshon and Dluzen 1997; Callier et al. 2000; Bains et al.2007), meta-amphetamine (Gajjar et al. 2003), and 6-OH-DA (Dluzen 1997). Estrogen-mediated neuroprotectiveeffects may have clinical relevance in PD, as this diseaseoccurs 1.5 times more frequently in men than in women(Fahn and Sulzer 2004; Wooten et al. 2004; Haaxma et al.2007); and in women, the age at onset of PD correlateswith the duration of the fertile life (Ragonese et al. 2006;Rocca et al. 2008).
Cellular responses to estrogen are tightly controlled. Alarge number of estrogen receptor (ER) interacting proteins,which behave as co-activators or co-repressors, modify theERa transcriptional activity (Metivier et al. 2003; Reid et al.
Received March 14, 2008; revised manuscript received June 18, 2008;accepted July 8, 2008.Address correspondence and reprint requests to Dr M. A. Mena,
Departamento de Investigacion, Hospital Ramon y Cajal, Ctra. de Col-menar, Km. 9, Madrid 28034, Spain. E-mail: [email protected] used: AKT, serine/threonine kinase protein; BRCA1,
breast cancer suppressor protein; CHIP, C-terminal Hsc-interactingprotein; DA, dopamine; E2, estradiol; E6-AP, E6-associated protein;EFP, estrogen responsive finger protein; ERa, estrogen receptor-a; ERK,extracellular signal-regulated kinase; GFAP, glial fibrillary acidic pro-tein; GPx, glutathione peroxidase; GSK-3, glycogen synthase kinase-3;HSP, heat-shock protein; MAP-2, microtubule associated protein 2a+2b;MAPK, mitogen-activated protein kinase; PD, Parkinson’s disease;PI3K, phosphatidylinositol 3¢-kinase; PK)/), parkin null mice; RING,really interesting new gene; SDS, sodium dodecyl sulfate; TH, tyrosinehydroxylase; WT, wild-type.
Abstract
Estrogens are considered neurotrophic for dopamine neurons.
Parkinson’s disease is more frequent in males than in
females, and more prevalent in females with short reproduc-
tive life. Estrogens are neuroprotective against neurotoxic
agents for dopamine neurons in vivo and in vitro. Here, we
have investigated the role of estrogens in wild-type (WT) and
parkin null mice (PK)/)). WT mice present sexual dimor-
phisms in neuroprotective mechanisms (Bcl-2/Bax, chaper-
ones, and GSH), but some of these inter-sex differences
disappear in PK)/). Tyrosine hydroxylase (TH) protein and
TH+ cells decreased earlier and more severely in female than
in male PK)/) mice. Neuronal cultures from midbrain of WT
and PK)/) mice were treated with estradiol from 10 min to
48 h. Short-term treatments activated the mitogen-activated
protein kinase pathway of WT and PK)/) neurons and the
phosphatidylinositol 3¢-kinase/AKT/glycogen synthase kinase-3
pathway of WT but not of PK)/) cultures. Long-term treat-
ments with estradiol increased the number of TH+ neurons,
the TH expression, and the extension of neurites, and de-
creased the level of apoptosis, the expression of glial fibrillary
acidic protein, and the number of microglial cells in WT but not
in PK)/) cultures. The levels of estrogen receptor-a were
elevated in midbrain cultures and in the striatum of adult
PK)/) male mice, suggesting that suppression of parkin
changes the estrogen receptor-a turnover. From our data, it
appears that parkin participates in the cellular estrogen
response which could be of interest in the management of
parkin-related Parkinson’s disease patients.
Keywords: C-terminal Hsc-interacting protein, dopamine
neurons, estradiol, estrogen receptor-a, glial cells, heat-shock
protein70, mitogen-activated protein kinase, Park-2 gene,
Parkinson’s disease and gender differences, phosphatidyl-
inositol 3¢-kinase/AKT/glycogen synthase kinase-3.
J. Neurochem. (2008) 106, 2143–2157.
JOURNAL OF NEUROCHEMISTRY | 2008 | 106 | 2143–2157 doi: 10.1111/j.1471-4159.2008.05569.x
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157 2143

2003; Tateishi et al. 2004; Zhang et al. 2006). The protea-some is involved in the degradation (Alarid et al. 1999;Nawaz et al. 1999a), transactivation (Lonard et al. 2000),and in the complete process of estrogen-dependent transcrip-tion (Lonard et al. 2000; Reid et al. 2003). The activation ofother nuclear receptors is also coupled with degradation ofthese proteins by the ubiquitin-proteosome pathway (Boudj-elal et al. 2000; Dace et al. 2000). Each nuclear receptor co-activator degradation is promoted by a specific E3 ubiquitinligase (Yan et al. 2003).
Parkin, an enzyme E3 ligase, which deficiency causesautosomal recessive PD and ubiquinates several candidatesubstrate proteins. Parkin targets its substrates for proteo-somal degradation collaborating with the ubiquitin-conju-gating enzyme UbcH7 (Shimura et al. 2000), a proteinwhich has been described as a co-activator for steroidhormone receptors (Verma et al. 2004). Parkin has a RING-IBR-RING (really interesting gene – in between ringfingers) motif in the C-terminus which was predicted tobe involved in the regulation of gene expression (Morett andBork 1999). Other RING type E3 ligases are required forthe degradation and activation of the transcription estrogendependent, such as murine double minute 2 [implicated inbreast cancer (Saji et al. 2001)] or E6-associated protein(E6-AP) in Angelman syndrome (Nawaz et al. 1999b) andmore recently a breast cancer suppressor protein (BRCA1)and estrogen responsive finger protein (EFP) (Eakin et al.2007; Nakajima et al. 2007).
The interactions between parkin and the estrogen systemand the role of parkin in the estrogen effects on DA neuronsare unknown. In this study, we have investigated the effectsof estradiol on neuronal-enriched midbrain primary culturesfrom E-13 embryos of parkin null (PK)/)) and wild-type(WT) mice as well as the gender differences in thedopaminergic system of adult PK)/) mice.
Materials and methods
MaterialsThe culture reagents B27 supplement, Neurobasal TM medium with
and without Phenol Red, fetal calf serum, penicillin–streptomycin,
pyruvate, and glutamine were obtained from GIBCO-Life Technol-
ogies (Paisley, UK). Trypan blue and bovine serum albumin were
purchased from Sigma (Madrid, Spain). All other agents were of the
highest purity commercially available from Merck (Darmstatdt,
Germany) or Sigma. Details about the antibodies used in this study
are provided in the Supporting information (Appendix S1).
Transgenic animalsParkin-null mutant (PK)/)) mice and their corresponding WT
littermate controls were obtained from heterozygous parkin)/+intercross, as previously described (Itier et al. 2003; Casarejos et al.2006; Menendez et al. 2006; Rodriguez-Navarro et al. 2007).
Heterozygous parkin)/+ were obtained by crossing homozygous
PK)/) from our colony with C57Bl/6 mice from Charles River
(Barcelona, Spain). The colony has been refreshed with new mice for
the last 6 years and the clinical phenotype and genotype has remained
stable. The genotype of each animal was defined as described below
(Itier et al. 2003; Casarejos et al. 2006; Menendez et al. 2006). Theoriginal PK)/) founders of the colony were generously donated by
Aventis Pharma SA laboratory (Vitry-sur-Seine, France) (Itier et al.2003). Procedures using laboratory animals were in accordance with
the European Union Directives. All efforts were made to minimize
the number of animals used and their suffering. In vivo studies were
performed in 5-, 18-, and 24-month-old male and female mice, six for
each experimental group. The estrous phase was induced in the
females by 3 days of exposure to male urine prior to killing.
Brain dissection and tissue preparationAfter decapitation, brain parts were dissected, according to Carlsson
and Lindqvist (1973) and the areas that contain a rich density of DA
terminals (limbic system and striatum) or DA cell bodies (midbrain),
were frozen on dry ice and used for further analysis. The striatum was
sonicated (VibraCell, level 2 for 30 s, Sonics and Materials Inc.,
Danburg, CT, USA) in eight volumes (w/v) of 50 mM potassium
phosphate, pH 7.2, 1 mM EDTA, and then centrifuged at 10 000 gfor 20 min at 4�C. The supernatant was recovered and 100 lL were
used for enzymatic activity measurement. The remaining supernatant
was carried to 0.4 N perchloric acid by addition of 4 N perchloric acid
and then centrifuged at 10 000 g for 20 min at 4�C. That supernatantwas used for GSH determination.
The pellet, with the proteins, was neutralized (w/v = 1/9) with
0.75% Na2CO3, 2% sodium dodecyl sulfate (SDS), 0.25 mM
phenylmethylsulfonyl fluoride, 10 mg/mL leupeptin, 2 mg/mL
aprotinin, 10 mg/mL pepsin, and then sonicated and centrifuged at
10 000 g for 30 min at 4�C. The supernatant was used for protein
determination by bicinchoninic acid assay and for electrophoresis
analysis.
GSH levels and glutathione peroxidase activityTotal GSH and GSSG levels were measured according to Tietze
(1969) and Griffith (1980), respectively. Glutathione peroxidase
(GPx) activity was measured according to Flohe and Gunzler
(1984). For details see Supporting information (Appendix S1).
Midbrain neuronal-enriched culturesMidbrain neuronal cultures were obtained from the ventral mesen-
cephalon of 13 day gestation embryos and processed as previously
described (Casarejos et al. 2005). The cultures were treated for 48 h
with estradiol (diluted in ethanol) to study its transcriptional effects
and, for less than 1-h, to see rapid estrogenic effects. The doses of
estradiol used are in the range of the physiological concentrations
of this hormone in mice plasma (Islamov et al. 2002). For detailssee the Supporting information (Appendix S1).
Neuronal-enriched cultures grown in B27 supplement/neurosabal
TM serum-free defined medium for 7–8 days in vitro are formed by
70–80% neurons [microtubule associated protein 2a+2b (MAP-2+)],
of which around 2–3% are dopaminergic (TH+), and 10–15% glia,
mostly astrocytes (glial fibrillary acidic protein; GFAP+), glial
progenitors (A2B5), and oligodendrocytes (O1+). The rest of the
cells are neuronal progenitors (nestin+ cells). Midbrain neuronal-
enriched cultures from PK-knockout mice have similar proportions
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2144 | J. A. Rodrıguez-Navarro et al.

of the different cell types with the exception of a significant
increment of microglial cells and glial progenitors (Casarejos et al.2005).
ImmunocytochemistryThe cell cultures were fixed with 4% p-formaldehyde, washed in
0.1 M phosphate-buffered saline, pH 7.4, permeabilized with
ethanol-acetic acid (19 : 1), and incubated at 4�C for 24 h with
primary antibodies diluted in phosphate-buffered saline containing
10% fetal calf serum. DA neurons were characterized by immuno-
staining with a mouse anti-TH antibody (1 : 500) and astrocytes
with a rabbit anti-GFAP antibody (1 : 500). To detect all neurons in
the culture, a mouse anti-neuronal nuclei (Neu N) antibody (1 : 10)
and mouse monoclonal anti-MAP-2 antibody (1 : 250) were used.
To study the effects of estradiol in WT and PK)/) midbrain cultures
on ERa and pERK location, anti-ERa (1 : 50) and anti-pextracel-
lular signal regulated kinase (ERK) (1 : 100) antibodies were used.
Fluorescein- and rhodamine-conjugated secondary antibodies were
employed to visualize positive cells under fluorescent microscopy
and Fast Red under optical microscopy. For microglial cell
identification, cultures were fixed with 4% p-formaldehyde–1%
glutaraldehyde and incubated at 4�C for 24 h with isolectin B4
peroxidase-labeled diluted in Tris-buffered saline/1% Triton X-100;
the positive cells were developed with a DAB-system (LSAB2
system; Dako, Glostrup, Denmark) and visualized under optical
microscopy. The number of immunoreactive cells was counted in
one-seventh of the total area of the cover slides. The cells were
counted in pre-defined parallel strips using a counting reticule
inserted into the ocular.
The intensity and the area of fluorescence of GFAP+, MAP-2,
and ERa immunostaining were measured in 20 pre-defined fields for
each slide, processed with the automatic software Image-Pro 6.2
(Media Cybernetics, Silver Spring, MD, USA) and expressed as
arbitrary units or intensity (integrated optical density). For all
immunocytochemical methods, negative controls were tested by
incubation without the primary antibody in each case.
Apoptosis detectionApoptosis was measured by light microscopic morphological
features and DNA staining with bis-benzimide (Hoechst 33342).
Briefly, cultures were fixed in 4% p-formaldehyde, nuclei were
stained with bis-benzimide added in the anti-fading solution
(3 · 10)6 M final concentration) and counted in 10 pre-defined
fields that represent 1/14 of the cover slide area; apoptotic cells were
identified by condensation and fragmentation of chromatin.
Western blot analysisPrimary midbrain cultures were rinsed, scraped, and homogenized
with a sonicator in buffer containing 20 mM Tris–HCl, pH 7.4,
10 mM potasium acetate, 1 mM dithiothreitol, 1 mM EDTA, 1 mM
phenylmethylsulfonyl fluoride, benzamidine, leupeptin, aprotinin,
and pepstatin 1 mM each, 100 mM NaF, 20 mM sodium molibdate,
20 mM b-glycerophosphate, and then centrifuged at 12 000 g for
30 min at 4�C. The supernatant was used for protein determination
by the bicinchoninic acid protein assay kit and for electrophoretical
separation. Samples from cultures or from the mice striatum (30 lg)were added to the SDS sample-loading buffer, electrophoresed in
10% SDS–polyacrylamide gels and then electroblotted to 0.45 lm
nitrocellulose membranes, as described by Rodriguez-Navarro et al.(2007).
Histological studiesThe animals used for these studies were anesthetized intraperitone-
ally with a mixture (5 : 4 : 1) of ketamine (50 mg/mL), diazepam
(1 mg/mL), and atropine (1 mg/mL) and perfused with 4%
p-formaldehyde. The whole brain was immersed in p-formaldehyde
for 24 h and then included in paraffin, sectioned in the microtome at
a thickness of 4 lm, and stained for hematoxylin/eosine (H&E) and
Nissl’s stain. TH antibody was used for immunohistochemistry.
Mouse monoclonal anti-TH antibody, from Chemicon (Madrid,
Spain) was diluted 1 : 2000 and anti-rabbit secondary antibodies
were from Dako. The number of TH immunoreactive neurons in the
substantia nigra was counted in an Olympus Bx51 stereological
microscope (Bellarup, Denmark) using a Cast Grid software
(Interactivision, Silkeborg, Denmark). DA cells were quantified in
eight to nine regularly spaced sections in each mouse brain, covering
the entire surface of the antero-posterior extent of the substantia
nigra. Counting of cells was performed independently by two blind
observers but it was not performed by stereology.
Statistical analysisThe results were statistically evaluated with different tests. Signif-
icant differences between the four experimental groups were
analyzed with one and two way ANOVA followed by the Newman–
Keuls multiple comparison test or Bonferroni test, respectively, as a
post hoc evaluation. Differences were considered statistically
significant when p < 0.05. The analysis of data was performed
using the Prism 4 GRAPHPAD Software (San Diego, CA, USA).
Results
Survival proteins and chaperones in striatum of male andfemale WT and PK)/) miceIt has been reported that the expression of antiapoptoticproteins, such as Bcl-2, in the brain is increased by estrogens(Garcia-Segura et al. 1998; Dubal et al. 1999). We have alsofound that female WTmice have greater Bcl-2/Bax ratios thanWT males. Young, 5-month-old, female PK)/) mice lose theneuroprotective effect of this pro-survival shift of proteins andhave similar Bcl-2/Bax ratios to male PK)/) mice butsignificantly lower than female WT (Fig. 1a), suggesting thatthe pro-survival effect of gender observed inWT disappears inPK)/) mice. These gender and genotype differences in theBcl-2/Bax ratios persist in 18-month-old mice (Fig. S1a).
C-terminal Hsc-interacting protein (CHIP) and heat-shockprotein (HSP70) cooperate with parkin. Both proteins areimplicated in the unfolding protein stress response. HSP70levels are higher in males than in females (Olazabal et al.1992). We found higher levels of HSP70 and CHIP instriatum of male WT mice than in females (Fig. 1b and c),suggesting that males developed compensatory changes to anincreased protein stress. With respect to PK)/) mice, theinter-gender differences in striatal levels of these chaperones
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2145
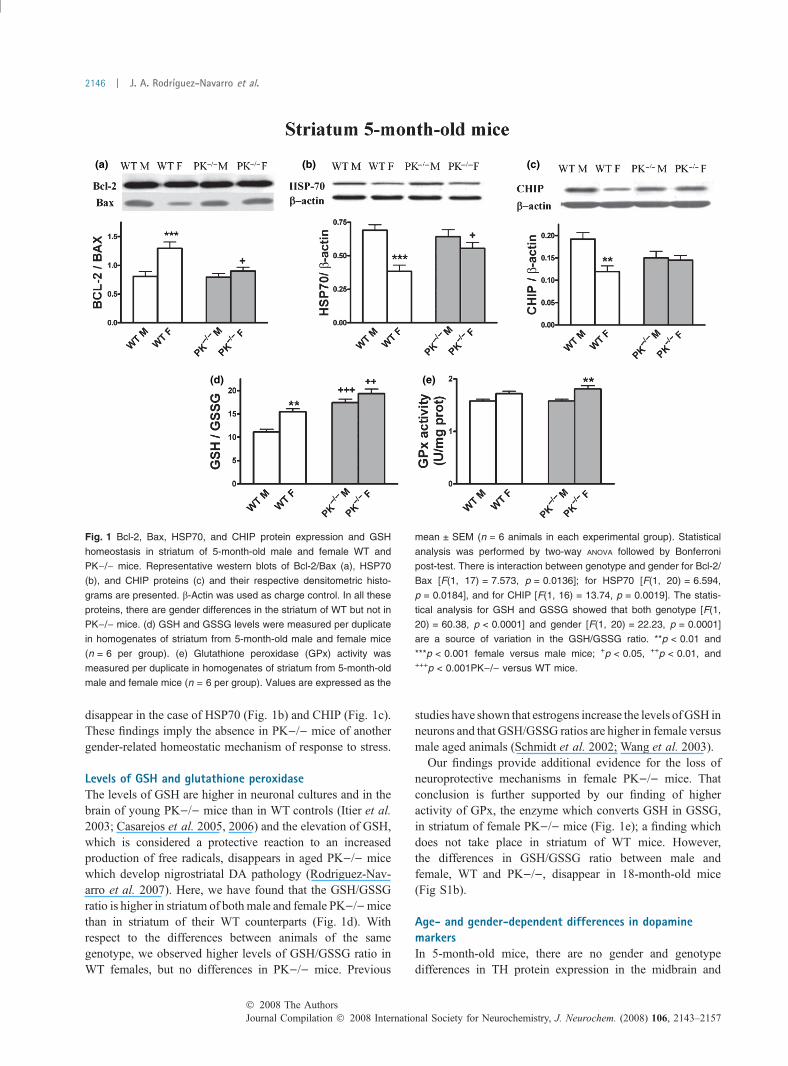
disappear in the case of HSP70 (Fig. 1b) and CHIP (Fig. 1c).These findings imply the absence in PK)/) mice of anothergender-related homeostatic mechanism of response to stress.
Levels of GSH and glutathione peroxidaseThe levels of GSH are higher in neuronal cultures and in thebrain of young PK)/) mice than in WT controls (Itier et al.2003; Casarejos et al. 2005, 2006) and the elevation of GSH,which is considered a protective reaction to an increasedproduction of free radicals, disappears in aged PK)/) micewhich develop nigrostriatal DA pathology (Rodriguez-Nav-arro et al. 2007). Here, we have found that the GSH/GSSGratio is higher in striatum of bothmale and female PK)/)micethan in striatum of their WT counterparts (Fig. 1d). Withrespect to the differences between animals of the samegenotype, we observed higher levels of GSH/GSSG ratio inWT females, but no differences in PK)/) mice. Previous
studies have shown that estrogens increase the levels ofGSH inneurons and that GSH/GSSG ratios are higher in female versusmale aged animals (Schmidt et al. 2002; Wang et al. 2003).
Our findings provide additional evidence for the loss ofneuroprotective mechanisms in female PK)/) mice. Thatconclusion is further supported by our finding of higheractivity of GPx, the enzyme which converts GSH in GSSG,in striatum of female PK)/) mice (Fig. 1e); a finding whichdoes not take place in striatum of WT mice. However,the differences in GSH/GSSG ratio between male andfemale, WT and PK)/), disappear in 18-month-old mice(Fig S1b).
Age- and gender-dependent differences in dopaminemarkersIn 5-month-old mice, there are no gender and genotypedifferences in TH protein expression in the midbrain and
(a)
(d) (e)
(b) (c)
Fig. 1 Bcl-2, Bax, HSP70, and CHIP protein expression and GSH
homeostasis in striatum of 5-month-old male and female WT and
PK)/) mice. Representative western blots of Bcl-2/Bax (a), HSP70
(b), and CHIP proteins (c) and their respective densitometric histo-
grams are presented. b-Actin was used as charge control. In all these
proteins, there are gender differences in the striatum of WT but not in
PK)/) mice. (d) GSH and GSSG levels were measured per duplicate
in homogenates of striatum from 5-month-old male and female mice
(n = 6 per group). (e) Glutathione peroxidase (GPx) activity was
measured per duplicate in homogenates of striatum from 5-month-old
male and female mice (n = 6 per group). Values are expressed as the
mean ± SEM (n = 6 animals in each experimental group). Statistical
analysis was performed by two-way ANOVA followed by Bonferroni
post-test. There is interaction between genotype and gender for Bcl-2/
Bax [F(1, 17) = 7.573, p = 0.0136]; for HSP70 [F(1, 20) = 6.594,
p = 0.0184], and for CHIP [F(1, 16) = 13.74, p = 0.0019]. The statis-
tical analysis for GSH and GSSG showed that both genotype [F(1,
20) = 60.38, p < 0.0001] and gender [F(1, 20) = 22.23, p = 0.0001]
are a source of variation in the GSH/GSSG ratio. **p < 0.01 and
***p < 0.001 female versus male mice; +p < 0.05, ++p < 0.01, and+++p < 0.001PK)/) versus WT mice.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2146 | J. A. Rodrıguez-Navarro et al.

striatum of WT and PK)/) (Fig. 2a and b). However,18-month-old female PK)/) mice have lower TH expressionthan their corresponding WT. Decreased TH expression in18-month-old female PK)/) mice is higher in midbrain thanin striatum (Fig. 2c and d).
We found a loss of TH+ cells in the substantia nigra ofmale PK)/) mice versus WT at 24 months (Rodriguez-Navarro et al. 2007), but the DA cell loss is higher in24-month-old female PK)/) mice versus their respectiveWT and male, respectively (Fig. 2e and f).
The loss of the neuroprotective mechanisms in femalePK)/) mice induces an earlier and more pronouncedreduction of TH protein and TH+ cells in female than inmale PK)/) mice.
Reproductive deficits in PK)/) miceTo further determine if parkin plays a role in the response toestrogens, we have studied the effects of estradiol onneuronal-enriched midbrain primary cultures. We killedpregnant female mice so as to obtain the E-13 neuronalcultures. We have studied the percentage of pregnant mice(16.1 ± 1.7% for WT and 11.45 ± 1.8 for PK)/)) and thenumber of fetus per pregnancy (8.3 ± 0.4 in WT and6.3 ± 0.4 in PK)/)) and they are significantly lower inPK)/) than in the corresponding age-matched WT mice.
Midbrain neuronal-enriched cultures do not respond toestradiolEstradiol increases the number of TH+ neurons around 50%of baseline in WT but not in PK)/) cultures (Fig. 3a and b),suggesting that estradiol is neurotrophic for DA neurons ofWT mice but not for those of PK)/). The increased numberof TH+ neurons after treatment with estradiol could be relatedto increased neurogenesis of DA neurons and to enhancementof TH expression in DA neurons which do not express THstrongly at this stage of development. Similarly, the levels ofTH protein were increased by treatment with estradiol incultures from WT but not in those from PK)/) (Fig. 3c). Theeffect of estradiol on TH+ cells from WT midbrain cultureswas dose dependent and optimal at 10 nM (Fig. 3b).
The differential response of WT and PK)/) mice toestradiol is not restricted to DA neurons. Neurites frommidbrain neurons (MAP-2+ cells) are increased by treatmentwith E2, 10 nM, for 48 h in WT but not in PK)/) cultures(Fig. 4a and b). The treatment with E2 significantlydecreases the percentage of apoptotic cells in the WT butnot in PK)/) cultures (Fig. 4c).
Parkin null glial cells do not respond to estradiolPrevious studies have reported effects of estrogens in fetal ratglial cultures (Garcia-Ovejero et al. 2005) and enhancementof neurite outgrowth by repression of GFAP expression(Rozovsky et al. 2002). In this study, we have found that thetreatment with estradiol for 48 h produces a dose-dependent
reduction of the area of GFAP immunoreactive cells up to50% of baseline in WT but not in PK)/) midbrain cultures(Fig. 5a and b). We obtained similar findings with regard tothe number of microglial cells from midbrain cultures(Fig. 6a) and to striatum of 5-month-old mice (Fig. 6b).The percentage of microglia was greater in both midbraincultures and striatum from PK)/) mice than in those fromWT. Interestingly, we observed that the treatment withestradiol reduced the percentage of microglia in midbrain ofWT but not of PK)/) cultures (Fig. 6a). Therefore, in theabsence of parkin, the proportion of cells with pro-inflam-matory effects is increased and there is a lack of thesuppressing effects of estrogens over microglia.
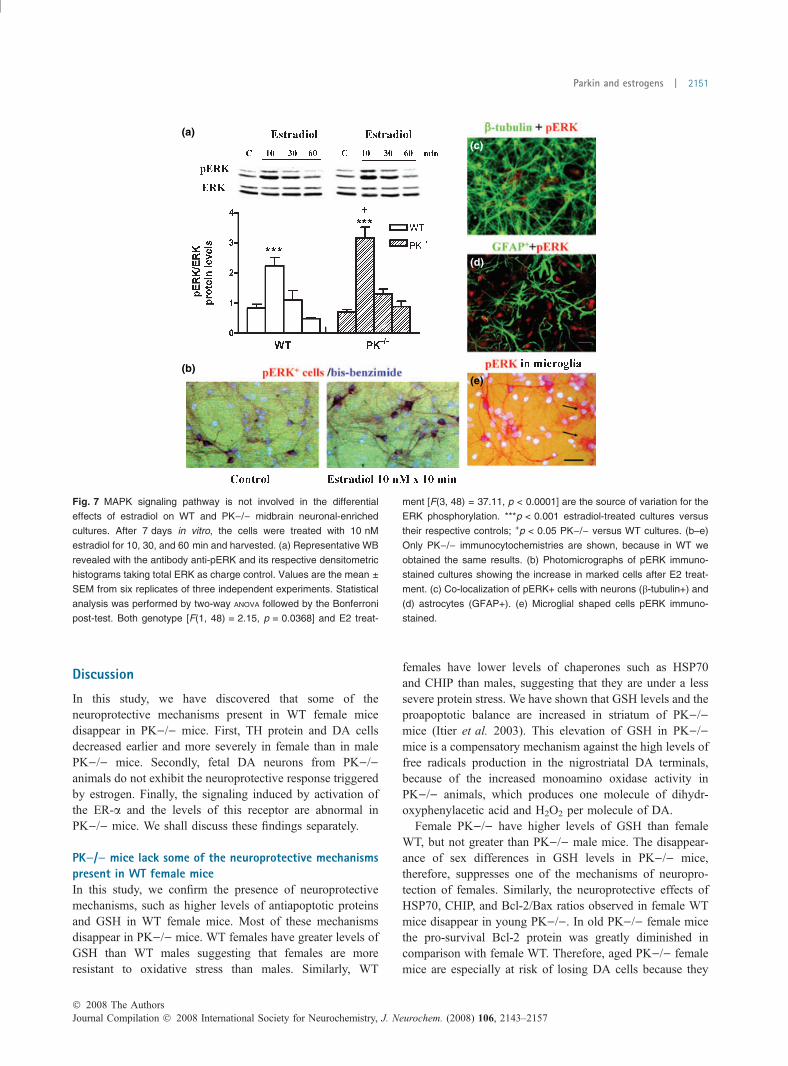
The MAPK signaling pathway is not involved in thedifferential effects of estradiol on WT and PK)/) culturesWe have examined the putative cytoplasmic mechanismsimplicated in the differential effects of estradiol on WT andPK)/) neuronal cultures. Rapid activation by estradiol ofextracellular signal-regulated kinase 1/2 signaling has beendescribed in other neuronal models (Setalo et al. 2002;Belcher et al. 2005). In Fig. 7a, we show that treatment withestradiol produces a rapid phosphorylation (activation) ofERK 10 min after E2, in both WT and PK)/), and of aslightly higher magnitude in PK)/). Phosphorylation ofERK takes place in the neurons and microglia (Fig. 7b–e),and the kinetics and the location of the pERK are similar inWT and PK)/) cultures. Therefore, abnormalities of ERKphosphorylation do not appear to be the key explanation forthe absence of effects of E2 in PK)/).
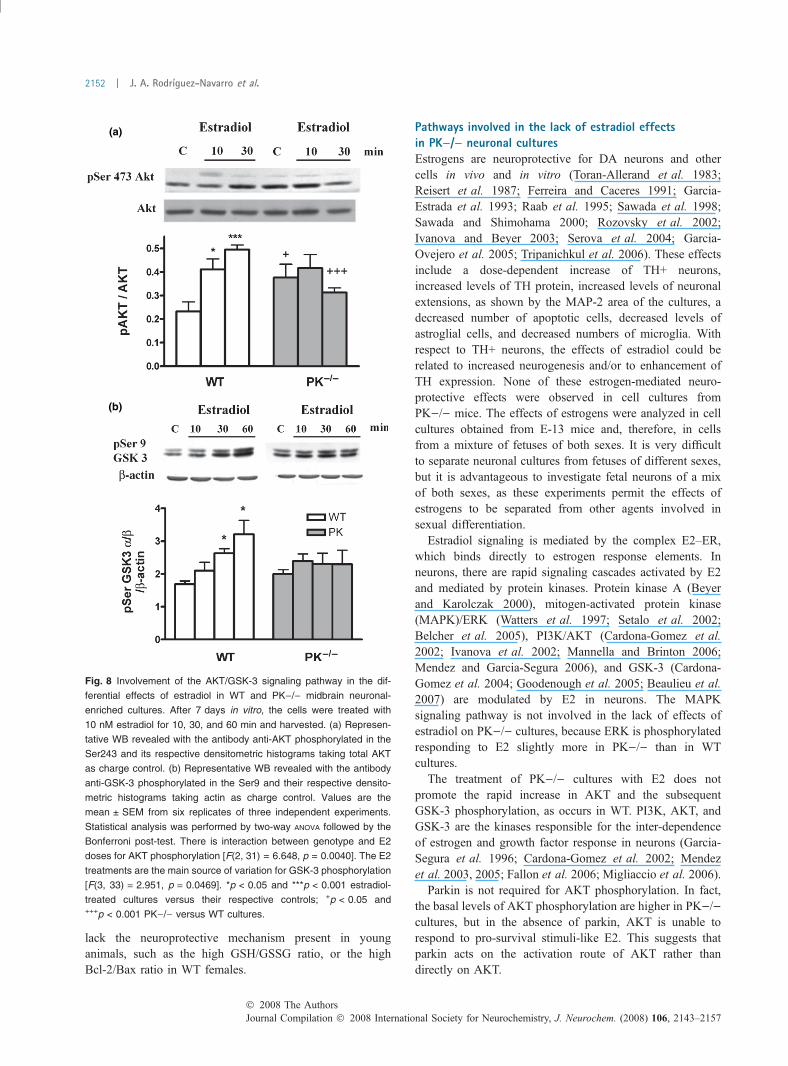
Lack of AKT/GSK-3 phosphorylation in response toestradiol in PK)/) culturesSeveral studies suggested that the neuroprotective role of E2in cultured neurons is mediated by the phosphatidylinositol3¢-kinase/AKT/glycogen synthase kinase-3 (PI3K/AKT/GSK3) signaling pathway (Honda et al. 2000; Cardona-Gomez et al. 2002, 2004; Wilson et al. 2002). The baselinelevel of phosphorylation of AKT is higher in PK)/) than inWT neuronal-enriched midbrain cultures (Fig. 8a), suggest-ing a compensatory survival response to the lack of parkin.The treatment with 10 nM E2 promotes a rapid increase inthe phosphorylation levels of AKT in the WT but not inPK)/) cultures (Fig. 8a). This effect is followed by thephosphorylation of the Ser9 residue of GSK-3 which inhibitsthe basal kinase activity of GSK-3. The differences weresignificant in the WT but not in PK)/) cultures (Fig. 8b).Therefore, parkin should be involved in the AKT activationmediated by the ERs.
Estrogen receptor-a levels are increased in absence ofparkinThe levels of ERa, as quantified by western blot, are higherin the striatum and midbrain of female WT than in males but
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2147
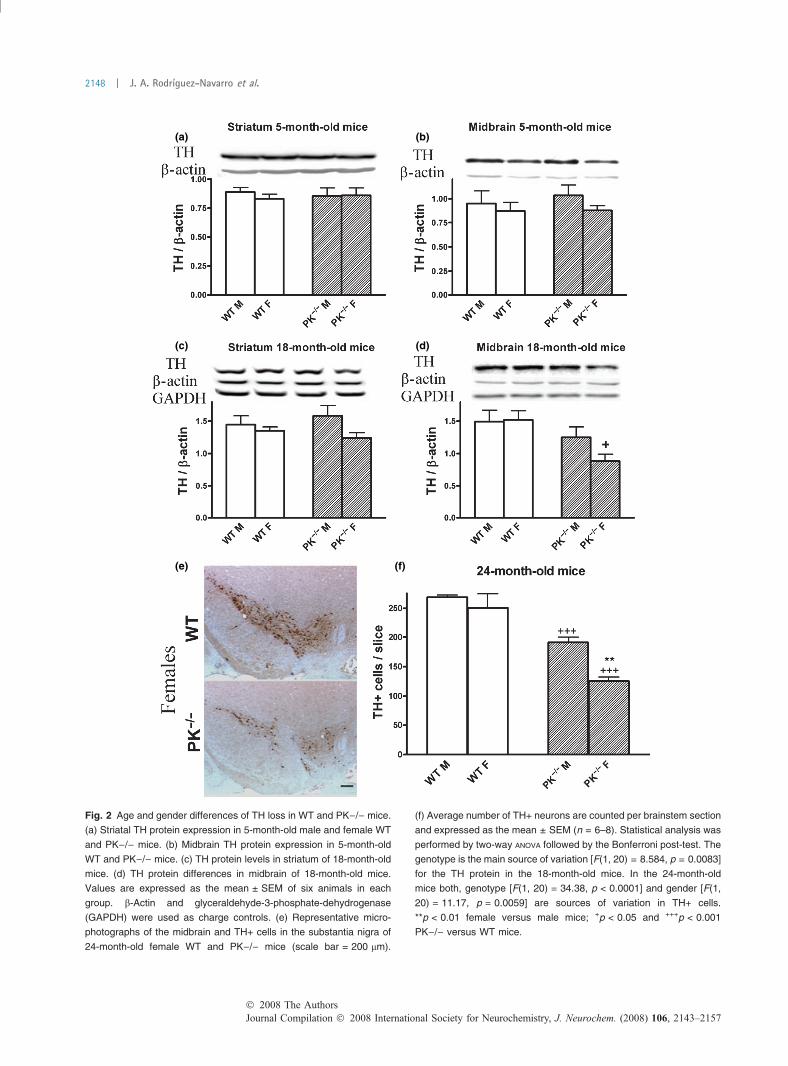
(a) (b)
(c) (d)
(e) (f)
Fig. 2 Age and gender differences of TH loss in WT and PK)/) mice.
(a) Striatal TH protein expression in 5-month-old male and female WT
and PK)/) mice. (b) Midbrain TH protein expression in 5-month-old
WT and PK)/) mice. (c) TH protein levels in striatum of 18-month-old
mice. (d) TH protein differences in midbrain of 18-month-old mice.
Values are expressed as the mean ± SEM of six animals in each
group. b-Actin and glyceraldehyde-3-phosphate-dehydrogenase
(GAPDH) were used as charge controls. (e) Representative micro-
photographs of the midbrain and TH+ cells in the substantia nigra of
24-month-old female WT and PK)/) mice (scale bar = 200 lm).
(f) Average number of TH+ neurons are counted per brainstem section
and expressed as the mean ± SEM (n = 6–8). Statistical analysis was
performed by two-way ANOVA followed by the Bonferroni post-test. The
genotype is the main source of variation [F(1, 20) = 8.584, p = 0.0083]
for the TH protein in the 18-month-old mice. In the 24-month-old
mice both, genotype [F(1, 20) = 34.38, p < 0.0001] and gender [F(1,
20) = 11.17, p = 0.0059] are sources of variation in TH+ cells.
**p < 0.01 female versus male mice; +p < 0.05 and +++p < 0.001
PK)/) versus WT mice.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2148 | J. A. Rodrıguez-Navarro et al.
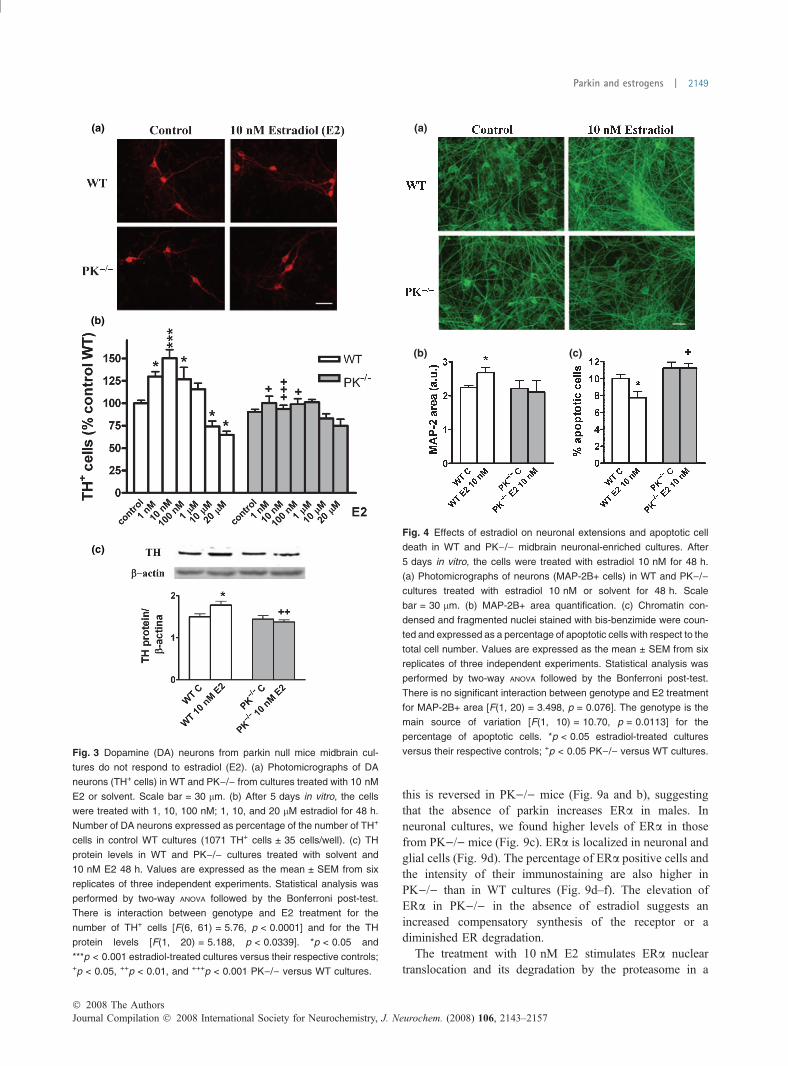
this is reversed in PK)/) mice (Fig. 9a and b), suggestingthat the absence of parkin increases ERa in males. Inneuronal cultures, we found higher levels of ERa in thosefrom PK)/) mice (Fig. 9c). ERa is localized in neuronal andglial cells (Fig. 9d). The percentage of ERa positive cells andthe intensity of their immunostaining are also higher inPK)/) than in WT cultures (Fig. 9d–f). The elevation ofERa in PK)/) in the absence of estradiol suggests anincreased compensatory synthesis of the receptor or adiminished ER degradation.
The treatment with 10 nM E2 stimulates ERa nucleartranslocation and its degradation by the proteasome in a
(a)
(b)
(c)
Fig. 3 Dopamine (DA) neurons from parkin null mice midbrain cul-
tures do not respond to estradiol (E2). (a) Photomicrographs of DA
neurons (TH+ cells) in WT and PK)/) from cultures treated with 10 nM
E2 or solvent. Scale bar = 30 lm. (b) After 5 days in vitro, the cells
were treated with 1, 10, 100 nM; 1, 10, and 20 lM estradiol for 48 h.
Number of DA neurons expressed as percentage of the number of TH+
cells in control WT cultures (1071 TH+ cells ± 35 cells/well). (c) TH
protein levels in WT and PK)/) cultures treated with solvent and
10 nM E2 48 h. Values are expressed as the mean ± SEM from six
replicates of three independent experiments. Statistical analysis was
performed by two-way ANOVA followed by the Bonferroni post-test.
There is interaction between genotype and E2 treatment for the
number of TH+ cells [F(6, 61) = 5.76, p < 0.0001] and for the TH
protein levels [F(1, 20) = 5.188, p < 0.0339]. *p < 0.05 and
***p < 0.001 estradiol-treated cultures versus their respective controls;+p < 0.05, ++p < 0.01, and +++p < 0.001 PK)/) versus WT cultures.
(a)
(b) (c)
Fig. 4 Effects of estradiol on neuronal extensions and apoptotic cell
death in WT and PK)/) midbrain neuronal-enriched cultures. After
5 days in vitro, the cells were treated with estradiol 10 nM for 48 h.
(a) Photomicrographs of neurons (MAP-2B+ cells) in WT and PK)/)cultures treated with estradiol 10 nM or solvent for 48 h. Scale
bar = 30 lm. (b) MAP-2B+ area quantification. (c) Chromatin con-
densed and fragmented nuclei stained with bis-benzimide were coun-
ted and expressed as a percentage of apoptotic cells with respect to the
total cell number. Values are expressed as the mean ± SEM from six
replicates of three independent experiments. Statistical analysis was
performed by two-way ANOVA followed by the Bonferroni post-test.
There is no significant interaction between genotype and E2 treatment
for MAP-2B+ area [F(1, 20) = 3.498, p = 0.076]. The genotype is the
main source of variation [F(1, 10) = 10.70, p = 0.0113] for the
percentage of apoptotic cells. *p < 0.05 estradiol-treated cultures
versus their respective controls; +p < 0.05 PK)/) versus WT cultures.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2149
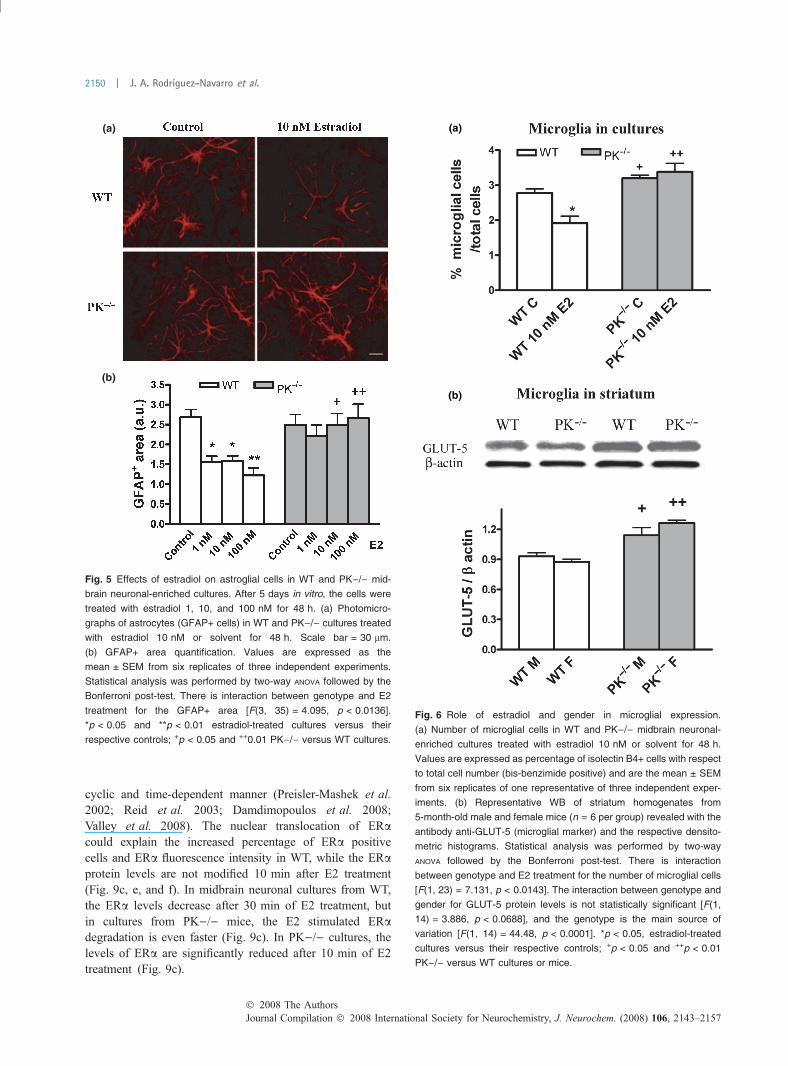
cyclic and time-dependent manner (Preisler-Mashek et al.2002; Reid et al. 2003; Damdimopoulos et al. 2008;Valley et al. 2008). The nuclear translocation of ERacould explain the increased percentage of ERa positivecells and ERa fluorescence intensity in WT, while the ERaprotein levels are not modified 10 min after E2 treatment(Fig. 9c, e, and f). In midbrain neuronal cultures from WT,the ERa levels decrease after 30 min of E2 treatment, butin cultures from PK)/) mice, the E2 stimulated ERadegradation is even faster (Fig. 9c). In PK)/) cultures, thelevels of ERa are significantly reduced after 10 min of E2treatment (Fig. 9c).
(a)
(b)
Fig. 5 Effects of estradiol on astroglial cells in WT and PK)/) mid-
brain neuronal-enriched cultures. After 5 days in vitro, the cells were
treated with estradiol 1, 10, and 100 nM for 48 h. (a) Photomicro-
graphs of astrocytes (GFAP+ cells) in WT and PK)/) cultures treated
with estradiol 10 nM or solvent for 48 h. Scale bar = 30 lm.
(b) GFAP+ area quantification. Values are expressed as the
mean ± SEM from six replicates of three independent experiments.
Statistical analysis was performed by two-way ANOVA followed by the
Bonferroni post-test. There is interaction between genotype and E2
treatment for the GFAP+ area [F(3, 35) = 4.095, p < 0.0136].
*p < 0.05 and **p < 0.01 estradiol-treated cultures versus their
respective controls; +p < 0.05 and ++0.01 PK)/) versus WT cultures.
(a)
(b)
Fig. 6 Role of estradiol and gender in microglial expression.
(a) Number of microglial cells in WT and PK)/) midbrain neuronal-
enriched cultures treated with estradiol 10 nM or solvent for 48 h.
Values are expressed as percentage of isolectin B4+ cells with respect
to total cell number (bis-benzimide positive) and are the mean ± SEM
from six replicates of one representative of three independent exper-
iments. (b) Representative WB of striatum homogenates from
5-month-old male and female mice (n = 6 per group) revealed with the
antibody anti-GLUT-5 (microglial marker) and the respective densito-
metric histograms. Statistical analysis was performed by two-way
ANOVA followed by the Bonferroni post-test. There is interaction
between genotype and E2 treatment for the number of microglial cells
[F(1, 23) = 7.131, p < 0.0143]. The interaction between genotype and
gender for GLUT-5 protein levels is not statistically significant [F(1,
14) = 3.886, p < 0.0688], and the genotype is the main source of
variation [F(1, 14) = 44.48, p < 0.0001]. *p < 0.05, estradiol-treated
cultures versus their respective controls; +p < 0.05 and ++p < 0.01
PK)/) versus WT cultures or mice.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2150 | J. A. Rodrıguez-Navarro et al.
Discussion
In this study, we have discovered that some of theneuroprotective mechanisms present in WT female micedisappear in PK)/) mice. First, TH protein and DA cellsdecreased earlier and more severely in female than in malePK)/) mice. Secondly, fetal DA neurons from PK)/)animals do not exhibit the neuroprotective response triggeredby estrogen. Finally, the signaling induced by activation ofthe ER-a and the levels of this receptor are abnormal inPK)/) mice. We shall discuss these findings separately.
PK)/) mice lack some of the neuroprotective mechanismspresent in WT female miceIn this study, we confirm the presence of neuroprotectivemechanisms, such as higher levels of antiapoptotic proteinsand GSH in WT female mice. Most of these mechanismsdisappear in PK)/) mice. WT females have greater levels ofGSH than WT males suggesting that females are moreresistant to oxidative stress than males. Similarly, WT
females have lower levels of chaperones such as HSP70and CHIP than males, suggesting that they are under a lesssevere protein stress. We have shown that GSH levels and theproapoptotic balance are increased in striatum of PK)/)mice (Itier et al. 2003). This elevation of GSH in PK)/)mice is a compensatory mechanism against the high levels offree radicals production in the nigrostriatal DA terminals,because of the increased monoamino oxidase activity inPK)/) animals, which produces one molecule of dihydr-oxyphenylacetic acid and H2O2 per molecule of DA.
Female PK)/) have higher levels of GSH than femaleWT, but not greater than PK)/) male mice. The disappear-ance of sex differences in GSH levels in PK)/) mice,therefore, suppresses one of the mechanisms of neuropro-tection of females. Similarly, the neuroprotective effects ofHSP70, CHIP, and Bcl-2/Bax ratios observed in female WTmice disappear in young PK)/). In old PK)/) female micethe pro-survival Bcl-2 protein was greatly diminished incomparison with female WT. Therefore, aged PK)/) femalemice are especially at risk of losing DA cells because they
(a)
(b)(e)
(d)
(c)
Fig. 7 MAPK signaling pathway is not involved in the differential
effects of estradiol on WT and PK)/) midbrain neuronal-enriched
cultures. After 7 days in vitro, the cells were treated with 10 nM
estradiol for 10, 30, and 60 min and harvested. (a) Representative WB
revealed with the antibody anti-pERK and its respective densitometric
histograms taking total ERK as charge control. Values are the mean ±
SEM from six replicates of three independent experiments. Statistical
analysis was performed by two-way ANOVA followed by the Bonferroni
post-test. Both genotype [F(1, 48) = 2.15, p = 0.0368] and E2 treat-
ment [F(3, 48) = 37.11, p < 0.0001] are the source of variation for the
ERK phosphorylation. ***p < 0.001 estradiol-treated cultures versus
their respective controls; +p < 0.05 PK)/) versus WT cultures. (b–e)
Only PK)/) immunocytochemistries are shown, because in WT we
obtained the same results. (b) Photomicrographs of pERK immuno-
stained cultures showing the increase in marked cells after E2 treat-
ment. (c) Co-localization of pERK+ cells with neurons (b-tubulin+) and
(d) astrocytes (GFAP+). (e) Microglial shaped cells pERK immuno-
stained.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2151
lack the neuroprotective mechanism present in younganimals, such as the high GSH/GSSG ratio, or the highBcl-2/Bax ratio in WT females.
Pathways involved in the lack of estradiol effectsin PK)/) neuronal culturesEstrogens are neuroprotective for DA neurons and othercells in vivo and in vitro (Toran-Allerand et al. 1983;Reisert et al. 1987; Ferreira and Caceres 1991; Garcia-Estrada et al. 1993; Raab et al. 1995; Sawada et al. 1998;Sawada and Shimohama 2000; Rozovsky et al. 2002;Ivanova and Beyer 2003; Serova et al. 2004; Garcia-Ovejero et al. 2005; Tripanichkul et al. 2006). These effectsinclude a dose-dependent increase of TH+ neurons,increased levels of TH protein, increased levels of neuronalextensions, as shown by the MAP-2 area of the cultures, adecreased number of apoptotic cells, decreased levels ofastroglial cells, and decreased numbers of microglia. Withrespect to TH+ neurons, the effects of estradiol could berelated to increased neurogenesis and/or to enhancement ofTH expression. None of these estrogen-mediated neuro-protective effects were observed in cell cultures fromPK)/) mice. The effects of estrogens were analyzed in cellcultures obtained from E-13 mice and, therefore, in cellsfrom a mixture of fetuses of both sexes. It is very difficultto separate neuronal cultures from fetuses of different sexes,but it is advantageous to investigate fetal neurons of a mixof both sexes, as these experiments permit the effects ofestrogens to be separated from other agents involved insexual differentiation.
Estradiol signaling is mediated by the complex E2–ER,which binds directly to estrogen response elements. Inneurons, there are rapid signaling cascades activated by E2and mediated by protein kinases. Protein kinase A (Beyerand Karolczak 2000), mitogen-activated protein kinase(MAPK)/ERK (Watters et al. 1997; Setalo et al. 2002;Belcher et al. 2005), PI3K/AKT (Cardona-Gomez et al.2002; Ivanova et al. 2002; Mannella and Brinton 2006;Mendez and Garcia-Segura 2006), and GSK-3 (Cardona-Gomez et al. 2004; Goodenough et al. 2005; Beaulieu et al.2007) are modulated by E2 in neurons. The MAPKsignaling pathway is not involved in the lack of effects ofestradiol on PK)/) cultures, because ERK is phosphorylatedresponding to E2 slightly more in PK)/) than in WTcultures.
The treatment of PK)/) cultures with E2 does notpromote the rapid increase in AKT and the subsequentGSK-3 phosphorylation, as occurs in WT. PI3K, AKT, andGSK-3 are the kinases responsible for the inter-dependenceof estrogen and growth factor response in neurons (Garcia-Segura et al. 1996; Cardona-Gomez et al. 2002; Mendezet al. 2003, 2005; Fallon et al. 2006; Migliaccio et al. 2006).
Parkin is not required for AKT phosphorylation. In fact,the basal levels of AKT phosphorylation are higher in PK)/)cultures, but in the absence of parkin, AKT is unable torespond to pro-survival stimuli-like E2. This suggests thatparkin acts on the activation route of AKT rather thandirectly on AKT.
(a)
(b)
Fig. 8 Involvement of the AKT/GSK-3 signaling pathway in the dif-
ferential effects of estradiol in WT and PK)/) midbrain neuronal-
enriched cultures. After 7 days in vitro, the cells were treated with
10 nM estradiol for 10, 30, and 60 min and harvested. (a) Represen-
tative WB revealed with the antibody anti-AKT phosphorylated in the
Ser243 and its respective densitometric histograms taking total AKT
as charge control. (b) Representative WB revealed with the antibody
anti-GSK-3 phosphorylated in the Ser9 and their respective densito-
metric histograms taking actin as charge control. Values are the
mean ± SEM from six replicates of three independent experiments.
Statistical analysis was performed by two-way ANOVA followed by the
Bonferroni post-test. There is interaction between genotype and E2
doses for AKT phosphorylation [F(2, 31) = 6.648, p = 0.0040]. The E2
treatments are the main source of variation for GSK-3 phosphorylation
[F(3, 33) = 2.951, p = 0.0469]. *p < 0.05 and ***p < 0.001 estradiol-
treated cultures versus their respective controls; +p < 0.05 and+++p < 0.001 PK)/) versus WT cultures.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2152 | J. A. Rodrıguez-Navarro et al.
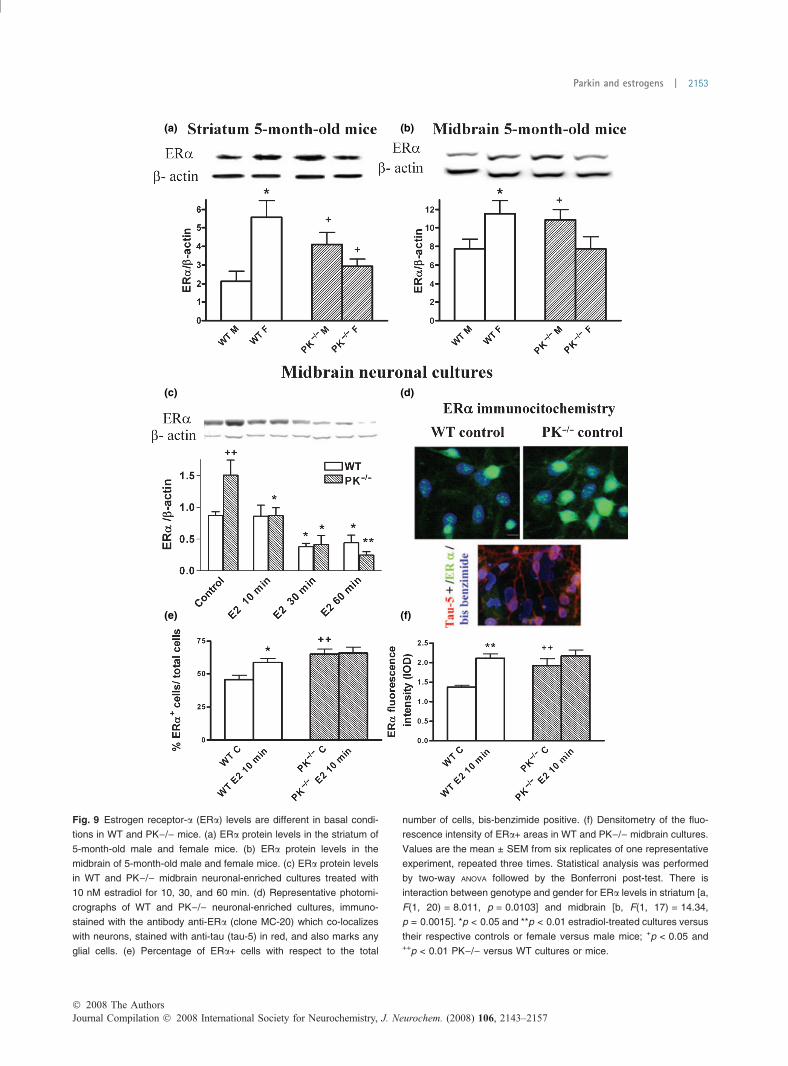
(a) (b)
(c) (d)
(e) (f)
Fig. 9 Estrogen receptor-a (ERa) levels are different in basal condi-
tions in WT and PK)/) mice. (a) ERa protein levels in the striatum of
5-month-old male and female mice. (b) ERa protein levels in the
midbrain of 5-month-old male and female mice. (c) ERa protein levels
in WT and PK)/) midbrain neuronal-enriched cultures treated with
10 nM estradiol for 10, 30, and 60 min. (d) Representative photomi-
crographs of WT and PK)/) neuronal-enriched cultures, immuno-
stained with the antibody anti-ERa (clone MC-20) which co-localizes
with neurons, stained with anti-tau (tau-5) in red, and also marks any
glial cells. (e) Percentage of ERa+ cells with respect to the total
number of cells, bis-benzimide positive. (f) Densitometry of the fluo-
rescence intensity of ERa+ areas in WT and PK)/) midbrain cultures.
Values are the mean ± SEM from six replicates of one representative
experiment, repeated three times. Statistical analysis was performed
by two-way ANOVA followed by the Bonferroni post-test. There is
interaction between genotype and gender for ERa levels in striatum [a,
F(1, 20) = 8.011, p = 0.0103] and midbrain [b, F(1, 17) = 14.34,
p = 0.0015]. *p < 0.05 and **p < 0.01 estradiol-treated cultures versus
their respective controls or female versus male mice; +p < 0.05 and++p < 0.01 PK)/) versus WT cultures or mice.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2153

Estrogen receptors in PK)/) miceWild-type female mice have higher levels of ERs in striatumthan males but these inter-gender differences disappear inPK)/). Both inhibition and constitutive activation of GSK-3 inhibit the transcriptional activity of ERa (Medunjaninet al. 2005). ERa phosphorylation by GSK-3 regulatesits localization, degradation, and transcriptional activity(Azuma et al. 2004; Callige et al. 2005; Mendez andGarcia-Segura 2006; Grisouard et al. 2007). Therefore, thelack of GSK-3 phosphorylation in response to E2 in PK)/)cultures could be implied in the lack of estrogen responseand in the differences observed in the localization and levelsof ERa.
Estrogen receptor-a interacts with HSP70 and tubulin(Azuma et al. 2004) which are two proteins related to parkinfunction. Microtubular stabilization impairs ER-dependenttranscription (Manavathi et al. 2006), so the absence ofparkin could affect ER transcription because of its effect ontubulin and microtubules. Moreover, parkin could be impli-cated in the nuclear transport of ERa because it degradesRanBP2 (Um et al. 2006), a constituent of the nuclear porecomplex with small ubiquitin-related modified protein(SUMO) E3 ligase activity. The SUMOylation of ERaregulates its transcriptional activity in presence of E2.
In addition, the suppression of an E3 ubiquitin ligase couldmodify the ER degradation through the reduction of itsdegradation by the proteasome. We have found that ERalevels are increased in neuronal cultures and in the striatumand midbrain of males in absence of parkin and E2. In thisway, the CHIP–HSP70 complex degrades ERa through theproteasome, preferentially in its unliganded or misfoldedform, and CHIP elimination promotes ERa accumulation(Tateishi et al. 2004; Fan et al. 2005). CHIP and HSP70form a complex in vitro and in vivo with parkin, modifyingits ubiquitination ability (Imai et al. 2002; Petrucelli et al.2004); parkin could be part of the CHIP–HSP70 complexdegrading unliganded ERa.
Estradiol stimulates ERa degradation by the proteasomein a cyclic and time-dependent manner, and this process isneeded for its transcriptional activity (Preisler-Mashek et al.2002; Reid et al. 2003). In a recent report, Valley et al.(2008) have described that after initial E2 treatment, thehalf-life of ERa is shortened from 3 to 1 h, but increasesover the time of treatment achieving a half-life of 6 h after72 h of treatment. They have explained these phenomenaas evidence of cell adaptation to chronic stimulation,because of the accumulation of a proteasome-resistantphosphorylated form of ERa. This could explain theincreased levels of ERa found in WT female striatumand in cultures after 48 h of E2 treatment (data not shown).In neuronal cultures from PK)/) mice, the levels of ERaare significantly reduced after 10 min of E2 treatment, evenfaster than in WT, which suggests that parkin could beimplied in the degradation of the unliganded but not in the
E2 bound form of ERa. Other E3 ligases, implied in E2stimulated ERa degradation, could be over-expressed inPK)/) mice as a compensation of the lack of parkinfunction.
Several E3 ubiquitin ligases have been implicated in ERadegradation, some with RING domains similar to parkin(Nawaz et al. 1999b; Smith et al. 2002; Kinyamu andArcher 2003; Tateishi et al. 2004, 2006; Burger et al. 2005,2006; Fan et al. 2005; Agirre et al. 2006; Eakin et al. 2007;Lu et al. 2007; Nakajima et al. 2007). The first E3 ligaseimplicated in ER transcription and co-activation was E6-AP,responsible for the Angelman syndrome, whose expressionin breast and ovary tumors is decreased, as happens withparkin, which is proposed as a tumor suppressor (Cesariet al. 2003; Denison et al. 2003; Gao et al. 2005).Furthermore, we have observed a decreased number offetuses and percentage of pregnant females in PK)/)mice; the ablation of E6-AP in mice causes reproductivedeficits and the response to E2 is impaired as we observein PK)/) (Nawaz et al. 1999a,b; Smith et al. 2002; Gaoet al. 2005).
Recently, other RING type E3 ligases implicated in cancer,such as the BRCA1 and the EFP have been proposed as E3ligases for ERa (Eakin et al. 2007; Nakajima et al. 2007).Mutants of EFP and BRCA1 lacking the RING domainprolonged the half-life of ERa and inhibited its ubiquitina-tion and transcriptional response to estradiol. Our resultssuggest that parkin could act in the same way as these RINGtype E3 ligases.
In conclusion, we have observed that parkin is involved inthe estrogen response at various levels. Our studies suggestthat the pharmacological manipulation of the signalingpathways related to the neurotrophic effects of estrogen,could provide a therapeutic tool to restore the protectiveeffects of estrogens in patients with mutations of parkin.
Acknowledgements
This study was supported in part by grants from the Spanish
Ministry of Health, FIS CP04/180, FIS 2004/PI040360; FIS 2007/
0037; CIBER 2006/05/0059, and CAM 0202/2006. The authors
thank I. Rodal and M. Serrano for excellent technical assistance and
Mrs Claire Marsden for editorial help.
Supporting information
Additional supporting information may be found in the online
version of this article.
Fig. S1 Bcl-2 and Bax protein expression and GSH homeostasis
in striatum of 18-month-old male and female WT and PK)/) mice.
Appendix S1 Supplementary material and methods
Please note: Blackwell Publishing are not responsible for the
content or functionality of any supporting materials supplied by the
authors. Any queries (other than missing material) should be
directed to the corresponding author for the article.
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2154 | J. A. Rodrıguez-Navarro et al.

References
Agirre X., Roman-Gomez J., Vazquez I. et al. (2006) Abnormal meth-ylation of the common PARK2 and PACRG promoter is associatedwith downregulation of gene expression in acute lymphoblasticleukemia and chronic myeloid leukemia. Int. J. Cancer 118, 1945–1953.
Alarid E. T., Bakopoulos N. and Solodin N. (1999) Proteasome-mediatedproteolysis of estrogen receptor: a novel component in autologousdown-regulation. Mol. Endocrinol. 13, 1522–1534.
Azuma K., Horie K., Inoue S., Ouchi Y. and Sakai R. (2004) Analysis ofestrogen receptor alpha signaling complex at the plasma mem-brane. FEBS Lett. 577, 339–344.
Bains M., Cousins J. C. and Roberts J. L. (2007) Neuroprotection byestrogen against MPP+-induced dopamine neuron death is medi-ated by ERalpha in primary cultures of mouse mesencephalon.Exp. Neurol. 204, 767–776.
Beaulieu J. M., Gainetdinov R. R. and Caron M. G. (2007) The Akt-GSK-3 signaling cascade in the actions of dopamine. TrendsPharmacol. Sci. 28, 166–172.
Belcher S. M., Le H. H., Spurling L. and Wong J. K. (2005) Rapidestrogenic regulation of extracellular signal-regulated kinase 1/2signaling in cerebellar granule cells involves a G protein- andprotein kinase A-dependent mechanism and intracellular activationof protein phosphatase 2A. Endocrinology 146, 5397–5406.
Beyer C. and Karolczak M. (2000) Estrogenic stimulation of neuritegrowth in midbrain dopaminergic neurons depends on cAMP/protein kinase A signalling. J. Neurosci. Res. 59, 107–116.
Boudjelal M., Wang Z., Voorhees J. J. and Fisher G. J. (2000) Ubiquitin/proteasome pathway regulates levels of retinoic acid receptorgamma and retinoid X receptor alpha in human keratinocytes.Cancer Res. 60, 2247–2252.
Burger A. M., Gao Y., Amemiya Y., Kahn H. J., Kitching R., Yang Y.,Sun P., Narod S. A., Hanna W. M. and Seth A. K. (2005) A novelRING-type ubiquitin ligase breast cancer-associated gene 2 cor-relates with outcome in invasive breast cancer. Cancer Res. 65,10401–10412.
Burger A., Amemiya Y., Kitching R. and Seth A. K. (2006) Novel RINGE3 ubiquitin ligases in breast cancer. Neoplasia 8, 689–695.
Callier S., Morissette M., Grandbois M. and Di Paolo T. (2000) Ste-reospecific prevention by 17beta-estradiol of MPTP-induceddopamine depletion in mice. Synapse 37, 245–251.
Callige M., Kieffer I. and Richard-Foy H. (2005) CSN5/Jab1 is involvedin ligand-dependent degradation of estrogen receptor {alpha} bythe proteasome. Mol. Cell. Biol. 25, 4349–4358.
Cardona-Gomez G. P., Mendez P. and Garcia-Segura L. M. (2002)Synergistic interaction of estradiol and insulin-like growth factor-Iin the activation of PI3K/Akt signaling in the adult rat hypothal-amus. Brain Res. Mol. Brain Res. 107, 80–88.
Cardona-Gomez P., Perez M., Avila J., Garcia-Segura L. M. and Wan-dosell F. (2004) Estradiol inhibits GSK3 and regulates interactionof estrogen receptors, GSK3, and beta-catenin in the hippocampus.Mol. Cell. Neurosci. 25, 363–373.
Carlsson A. and Lindqvist M. (1973) Effect of ethanol on the hydrox-ylation of tyrosine and tryptophan in rat brain in vivo. J. Pharm.Pharmacol. 25, 437–440.
Casarejos M. J., Solano R. M., Menendez J., Rodriguez-Navarro J. A.,Correa C., Garcia de Yebenes J. and Mena M. A. (2005) Differ-ential effects of l-DOPA on monoamine metabolism, cell survivaland glutathione production in midbrain neuronal-enriched culturesfrom parkin knockout and wild-type mice. J. Neurochem. 94,1005–1014.
Casarejos M. J., Menendez J., Solano R. M., Rodriguez-Navarro J. A.,Garcia de Yebenes J. and Mena M. A. (2006) Susceptibility to
rotenone is increased in neurons from parkin null mice and isreduced by minocycline. J. Neurochem. 97, 934–946.
Cesari R., Martin E. S., Calin G. A. et al. (2003) Parkin, a geneimplicated in autosomal recessive juvenile parkinsonism, is acandidate tumor suppressor gene on chromosome 6q25-q27. Proc.Natl Acad. Sci. USA 100, 5956–5961.
Dace A., Zhao L., Park K. S., Furuno T., Takamura N., Nakanishi M.,West B. L., Hanover J. A. and Cheng S. (2000) Hormone bindinginduces rapid proteasome-mediated degradation of thyroid hor-mone receptors. Proc. Natl Acad. Sci. USA 97, 8985–8990.
Damdimopoulos A. E., Spyrou G. and Gustafsson J. A. (2008) Ligandsdifferentially modify the nuclear mobility of estrogen receptorsalpha and beta. Endocrinology 149, 339–345.
Denison S. R., Wang F., Becker N. A., Schule B., Kock N., PhillipsL. A., Klein C. and Smith D. I. (2003) Alterations in the commonfragile site gene Parkin in ovarian and other cancers. Oncogene 22,8370–8378.
Disshon K. A. and Dluzen D. E. (1997) Estrogen as a neuromodulator ofMPTP-induced neurotoxicity: effects upon striatal dopamine re-lease. Brain Res. 764, 9–16.
Dluzen D. (1997) Estrogen decreases corpus striatal neurotoxicity inresponse to 6-hydroxydopamine. Brain Res. 767, 340–344.
Dubal D. B., Shughrue P. J., Wilson M. E., Merchenthaler I. and WiseP. M. (1999) Estradiol modulates bcl-2 in cerebral ischemia: apotential role for estrogen receptors. J. Neurosci. 19, 6385–6393.
Eakin C. M., Maccoss M. J., Finney G. L. and Klevit R. E. (2007)Estrogen receptor {alpha} is a putative substrate for the BRCA1ubiquitin ligase. Proc. Natl Acad. Sci. USA 104, 5794–5799.
Fahn S. and Sulzer D. (2004) Neurodegeneration and neuroprotection inParkinson disease. NeuroRx 1, 139–154.
Fallon L., Belanger C. M., Corera A. T. et al. (2006) A regulatedinteraction with the UIM protein Eps15 implicates parkin in EGFreceptor trafficking and PI(3)K-Akt signalling. Nat. Cell Biol. 8,834–842.
Fan M., Park A. and Nephew K. P. (2005) CHIP (carboxyl terminus ofHsc70-interacting protein) promotes basal and geldanamycin-in-duced degradation of estrogen receptor-alpha.Mol. Endocrinol. 19,2901–2914.
Ferreira A. and Caceres A. (1991) Estrogen-enhanced neurite growth:evidence for a selective induction of Tau and stable microtubules.J. Neurosci. 11, 392–400.
Flohe L. and Gunzler W. A. (1984) Assays of glutathione peroxidase.Methods Enzymol. 105, 114–121.
Gajjar T. M., Anderson L. I. and Dluzen D. E. (2003) Acute effects ofestrogen upon methamphetamine induced neurotoxicity of thenigrostriatal dopaminergic system. J. Neural Transm. 110,1215–1224.
Gao X., Mohsin S. K., Gatalica Z., Fu G., Sharma P. and Nawaz Z.(2005) Decreased expression of e6-associated protein in breast andprostate carcinomas. Endocrinology 146, 1707–1712.
Garcia-Estrada J., Del Rio J. A., Luquin S., Soriano E. and Garcia-Segura L. M. (1993) Gonadal hormones down-regulate reactivegliosis and astrocyte proliferation after a penetrating brain injury.Brain Res. 628, 271–278.
Garcia-Ovejero D., Azcoitia I., Doncarlos L. L., Melcangi R. C. andGarcia-Segura L. M. (2005) Glia-neuron crosstalk in the neuro-protective mechanisms of sex steroid hormones. Brain Res. BrainRes. Rev. 48, 273–286.
Garcia-Segura L. M., Duenas M., Fernandez-Galaz M. C., Chowen J. A.,Argente J., Naftolin F. and Torres-Aleman I. (1996) Interaction ofthe signalling pathways of insulin-like growth factor-I andsex steroids in the neuroendocrine hypothalamus. Horm. Res. 46,160–164.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2155

Garcia-Segura L. M., Cardona-Gomez P., Naftolin F. and Chowen J. A.(1998) Estradiol upregulates Bcl-2 expression in adult brainneurons. Neuroreport 9, 593–597.
Goodenough S., Schleusner D., Pietrzik C., Skutella T. and Behl C.(2005) Glycogen synthase kinase 3beta links neuroprotection by17beta-estradiol to key Alzheimer processes. Neuroscience 132,581–589.
Griffith O. W. (1980) Determination of glutathione and glutathionedisulfide using glutathione reductase and 2-vinylpyridine. Anal.Biochem. 106, 207–212.
Grisouard J., Medunjanin S., Hermani A., Shukla A. and Mayer D.(2007) Glycogen synthase kinase-3 protects estrogen receptoralpha from proteasomal degradation and is required for fulltranscriptional activity of the receptor. Mol. Endocrinol. 21,2427–2439.
Haaxma C. A., Bloem B. R., Borm G. F., Oyen W. J., Leenders K. L.,Eshuis S., Booij J., Dluzen D. E. and Horstink M. W. (2007)Gender differences in Parkinson’s disease. J. Neurol. Neurosurg.Psychiatry 78, 819–824.
Honda K., Sawada H., Kihara T., Urushitani M., Nakamizo T., AkaikeA. and Shimohama S. (2000) Phosphatidylinositol 3-kinasemediates neuroprotection by estrogen in cultured cortical neurons.J. Neurosci. Res. 60, 321–327.
Imai Y., Soda M., Hatakeyama S., Akagi T., Hashikawa T., Nakayama K.I. and Takahashi R. (2002) CHIP is associated with Parkin, a generesponsible for familial Parkinson’s disease, and enhances itsubiquitin ligase activity. Mol. Cell. 10, 55–67.
Islamov R. R., Hendricks W. A., Jones R. J., Lyall G. J., Spanier N.S. and Murashov A. K. (2002) 17Beta-estradiol stimulatesregeneration of sciatic nerve in female mice. Brain Res. 943,283–286.
Itier J. M., Ibanez P., Mena M. A. et al. (2003) Parkin gene inactivationalters behaviour and dopamine neurotransmission in the mouse.Hum. Mol. Genet. 12, 2277–2291.
Ivanova T. and Beyer C. (2003) Estrogen regulates tyrosine hydroxylaseexpression in the neonate mouse midbrain. J. Neurobiol. 54, 638–647.
Ivanova T., Mendez P., Garcia-Segura L. M. and Beyer C. (2002)Rapid stimulation of the PI3-kinase/Akt signalling pathway indeveloping midbrain neurones by oestrogen. J. Neuroendocrinol.14, 73–79.
Kinyamu H. K. and Archer T. K. (2003) Estrogen receptor-dependentproteasomal degradation of the glucocorticoid receptor is coupledto an increase in mdm2 protein expression. Mol. Cell. Biol. 23,5867–5881.
Kishi Y., Takahashi J., Koyanagi M., Morizane A., Okamoto Y., Hor-iguchi S., Tashiro K., Honjo T., Fujii S. and Hashimoto N. (2005)Estrogen promotes differentiation and survival of dopaminergicneurons derived from human neural stem cells. J. Neurosci. Res.79, 279–286.
Lonard D. M., Nawaz Z., Smith C. L. and O’Malley B. W. (2000) The26S proteasome is required for estrogen receptor-alpha and coac-tivator turnover and for efficient estrogen receptor-alpha transac-tivation. Mol. Cell. 5, 939–948.
Lu Y., Amleh A., Sun J., Jin X., McCullough S. D., Baer R., Ren D., LiR. and Hu Y. (2007) Ubiquitination and proteasome-mediateddegradation of BRCA1 and BARD1 during steroidogenesis inhuman ovarian granulosa cells. Mol. Endocrinol. 21, 651–663.
Manavathi B., Acconcia F., Rayala S. K. and Kumar R. (2006) Aninherent role of microtubule network in the action of nuclearreceptor. Proc. Natl Acad. Sci. USA 103, 15981–15986.
Mannella P. and Brinton R. D. (2006) Estrogen receptor protein inter-action with phosphatidylinositol 3-kinase leads to activation ofphosphorylated Akt and extracellular signal-regulated kinase 1/2 in
the same population of cortical neurons: a unified mechanism ofestrogen action. J. Neurosci. 26, 9439–9447.
Medunjanin S., Hermani A., De Servi B., Grisouard J., Rincke G. andMayer D. (2005) Glycogen synthase kinase-3 interacts withand phosphorylates estrogen receptor alpha and is involved inthe regulation of receptor activity. J. Biol. Chem. 280,33006–33014.
Mendez P. and Garcia-Segura L. M. (2006) Phosphatidylinositol 3-ki-nase and glycogen synthase kinase 3 regulate estrogen receptor-mediated transcription in neuronal cells. Endocrinology 147,3027–3039.
Mendez P., Azcoitia I. and Garcia-Segura L. M. (2003) Estrogen receptoralpha forms estrogen-dependent multimolecular complexeswith insulin-like growth factor receptor and phosphatidylinositol3-kinase in the adult rat brain. Brain Res. Mol. Brain Res. 112,170–176.
Mendez P., Cardona-Gomez G. P. and Garcia-Segura L. M. (2005)Interactions of insulin-like growth factor-I and estrogen in thebrain. Adv. Exp. Med. Biol. 567, 285–303.
Menendez J., Rodriguez-Navarro J. A., Solano R. M., Casarejos M. J.,Rodal I., Guerrero R., Sanchez M. P., Avila J., Mena M. A. and deYebenes J. G. (2006) Suppression of Parkin enhances nigrostriataland motor neuron lesion in mice over-expressing human-mutatedtau protein. Hum. Mol. Genet. 15, 2045–2058.
Metivier R., Penot G., Hubner M. R., Reid G., Brand H., Kos M. andGannon F. (2003) Estrogen receptor-alpha directs ordered, cyclical,and combinatorial recruitment of cofactors on a natural targetpromoter. Cell 115, 751–763.
Migliaccio A., Castoria G., Di Domenico M. et al. (2006) Crosstalkbetween EGFR and extranuclear steroid receptors. Ann. NY Acad.Sci. 1089, 194–200.
Morett E. and Bork P. (1999) A novel transactivation domain in parkin.Trends Biochem. Sci. 24, 229–231.
Nakajima A., Maruyama S., Bohgaki M., Miyajima N., Tsukiyama T.,Sakuragi N. and Hatakeyama S. (2007) Ligand-dependent tran-scription of estrogen receptor alpha is mediated by the ubiquitinligase EFP. Biochem. Biophys. Res. Commun. 357, 245–251.
Nawaz Z., Lonard D. M., Dennis A. P., Smith C. L. and O’Malley B. W.(1999a) Proteasome-dependent degradation of the human estrogenreceptor. Proc. Natl Acad. Sci. USA 96, 1858–1862.
Nawaz Z., Lonard D. M., Smith C. L., Lev-Lehman E., Tsai S. Y., TsaiM. J. and O’Malley B. W. (1999b) The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nuclear hormonereceptor superfamily. Mol. Cell. Biol. 19, 1182–1189.
Olazabal U. E., Pfaff D. W. and Mobbs C. V. (1992) Sex differences inthe regulation of heat shock protein 70 kDa and 90 kDa in therat ventromedial hypothalamus by estrogen. Brain Res. 596, 311–314.
Petrucelli L., Dickson D., Kehoe K. et al. (2004) CHIP and Hsp70regulate tau ubiquitination, degradation and aggregation. Hum.Mol. Genet. 13, 703–714.
Preisler-Mashek M. T., Solodin N., Stark B. L., Tyriver M. K. and AlaridE. T. (2002) Ligand-specific regulation of proteasome-mediatedproteolysis of estrogen receptor-alpha. Am. J. Physiol. Endocrinol.Metab. 282, E891–E898.
Raab H., Pilgrim C. and Reisert I. (1995) Effects of sex and estrogen ontyrosine hydroxylase mRNA in cultured embryonic rat mesen-cephalon. Brain Res. Mol. Brain Res. 33, 157–164.
Ragonese P., D’Amelio M. and Savettieri G. (2006) Implications forestrogens in Parkinson’s disease: an epidemiological approach.Ann. NY Acad. Sci. 1089, 373–382.
Reid G., Hubner M. R., Metivier R., Brand H., Denger S., Manu D.,Beaudouin J., Ellenberg J. and Gannon F. (2003) Cyclic, protea-some-mediated turnover of unliganded and liganded ERalpha on
Journal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157� 2008 The Authors
2156 | J. A. Rodrıguez-Navarro et al.

responsive promoters is an integral feature of estrogen signaling.Mol. Cell. 11, 695–707.
Reisert I., Han V., Lieth E., Toran-Allerand D., Pilgrim C. and Lauder J.(1987) Sex steroids promote neurite growth in mesencephalictyrosine hydroxylase immunoreactive neurons in vitro. Int. J. Dev.Neurosci. 5, 91–98.
Rocca W. A., Bower J. H., Maraganore D. M., Ahlskog J. E., GrossardtB. R., de Andrade M. and Melton Iii L. J. (2008) Increased risk ofparkinsonism in women who underwent oophorectomy beforemenopause. Neurology 103, 98–114.
Rodriguez-Navarro J. A., Casarejos M. J., Menendez J., Solano R. M.,Rodal I., Gomez A., Yebenes J. G. and Mena M. A. (2007) Mor-tality, oxidative stress and tau accumulation during ageing in par-kin null mice. J. Neurochem. 103, 98–114.
Rozovsky I., Wei M., Stone D. J., Zanjani H., Anderson C. P., Morgan T.E. and Finch C. E. (2002) Estradiol (E2) enhances neurite out-growth by repressing glial fibrillary acidic protein expression andreorganizing laminin. Endocrinology 143, 636–646.
Saji S., Okumura N., Eguchi H., Nakashima S., Suzuki A., Toi M.,Nozawa Y. and Hayashi S. (2001) MDM2 enhances the function ofestrogen receptor alpha in human breast cancer cells. Biochem.Biophys. Res. Commun. 281, 259–265.
Sawada H. and Shimohama S. (2000) Neuroprotective effects of estra-diol in mesencephalic dopaminergic neurons. Neurosci. Biobehav.Rev. 24, 143–147.
Sawada H., Ibi M., Kihara T., Urushitani M., Akaike A. and ShimohamaS. (1998) Estradiol protects mesencephalic dopaminergic neuronsfrom oxidative stress-induced neuronal death. J. Neurosci. Res. 54,707–719.
Schmidt A. J., Krieg J. C. and Vedder H. (2002) Differential effectsof glucocorticoids and gonadal steroids on glutathione levelsin neuronal and glial cell systems. J. Neurosci. Res. 67, 544–550.
Serova L. I., Maharjan S., Huang A., Sun D., Kaley G. and SabbanE. L. (2004) Response of tyrosine hydroxylase and GTPcyclohydrolase I gene expression to estrogen in brain catechol-aminergic regions varies with mode of administration. BrainRes. 1015, 1–8.
Setalo G. Jr, Singh M., Guan X. and Toran-Allerand C. D. (2002)Estradiol-induced phosphorylation of ERK1/2 in explants of themouse cerebral cortex: the roles of heat shock protein 90 (Hsp90)and MEK2. J. Neurobiol. 50, 1–12.
Shimura H., Hattori N., Kubo S. et al. (2000) Familial Parkinson diseasegene product, parkin, is a ubiquitin-protein ligase. Nat. Genet. 25,302–305.
Smith C. L., DeVera D. G., Lamb D. J., Nawaz Z., Jiang Y. H., BeaudetA. L. and O’Malley B. W. (2002) Genetic ablation of the steroidreceptor coactivator-ubiquitin ligase, E6-AP, results in tissue-selective steroid hormone resistance and defects in reproduction.Mol. Cell. Biol. 22, 525–535.
Tateishi Y., Kawabe Y., Chiba T., Murata S., Ichikawa K., Murayama A.,Tanaka K., Baba T., Kato S. and Yanagisawa J. (2004) Ligand-dependent switching of ubiquitin-proteasome pathways for estro-gen receptor. EMBO J. 23, 4813–4823.
Tateishi Y., Sonoo R., Sekiya Y. et al. (2006) Turning off estrogenreceptor beta-mediated transcription requires estrogen-dependentreceptor proteolysis. Mol. Cell. Biol. 26, 7966–7976.
Tietze F. (1969) Enzymatic method for quantitative determinationof nanogram amounts of total and oxidized glutathione:application to mammalian blood and other tissue. Anal. Biochem.27, 502–522.
Toran-Allerand C. D., Hashimoto K., Greenough W. T. and Saltarelli M.(1983) Sex steroids and the development of the newborn mousehypothalamus and preoptic area in vitro: III. Effects of estrogen ondendritic differentiation. Brain Res. 283, 97–101.
Tripanichkul W., Sripanichkulchai K. and Finkelstein D. I. (2006)Estrogen down-regulates glial activation in male mice following1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine intoxication. BrainRes. 1084, 28–37.
Um J. W., Min D. S., Rhim H., Kim J., Paik S. R. and Chung K. C.(2006) Parkin ubiquitinates and promotes the degradation ofRanBP2. J. Biol. Chem. 281, 3595–3603.
Valley C. C., Solodin N. M., Powers G. L., Ellison S. J. and Alarid E. T.(2008) Temporal variation in estrogen receptor-alpha protein turn-over in the presence of estrogen. J. Mol. Endocrinol. 40, 23–34.
Verma S., Ismail A., Gao X., Fu G., Li X., O’Malley B. W. and NawazZ. (2004) The ubiquitin-conjugating enzyme UBCH7 acts as acoactivator for steroid hormone receptors. Mol. Cell. Biol. 24,8716–8726.
Wang H., Liu H. and Liu R. M. (2003) Gender difference inglutathione metabolism during aging in mice. Exp. Gerontol. 38,507–517.
Watters J. J., Campbell J. S., Cunningham M. J., Krebs E. G. and DorsaD. M. (1997) Rapid membrane effects of steroids in neuroblastomacells: effects of estrogen on mitogen activated protein kinase sig-nalling cascade and c-fos immediate early gene transcription.Endocrinology 138, 4030–4033.
Wilson M. E., Liu Y. and Wise P. M. (2002) Estradiol enhances Aktactivation in cortical explant cultures following neuronal injury.Brain Res. Mol. Brain Res. 102, 48–54.
Wooten G. F., Currie L. J., Bovbjerg V. E., Lee J. K. and Patrie J. (2004)Are men at greater risk for Parkinson’s disease than women?J. Neurol. Neurosurg. Psychiatry 75, 637–639.
Yan F., Gao X., Lonard D. M. and Nawaz Z. (2003) Specific ubiquitin-conjugating enzymes promote degradation of specific nuclearreceptor coactivators. Mol. Endocrinol. 17, 1315–1331.
Zhang H., Sun L., Liang J., Yu W., Zhang Y., Wang Y., Chen Y., Li R.,Sun X. and Shang Y. (2006) The catalytic subunit of theproteasome is engaged in the entire process of estrogenreceptor-regulated transcription. EMBO J. 25, 4223–4233.
� 2008 The AuthorsJournal Compilation � 2008 International Society for Neurochemistry, J. Neurochem. (2008) 106, 2143–2157
Parkin and estrogens | 2157




















