
Activation of Estrogen Receptor-A by the Anion Nitrite
10
2008;68:3950-3958. Cancer Res David J. Veselik, Shailaja Divekar, Sivanesan Dakshanamurthy, et al. by the Anion Nitrite α Activation of Estrogen Receptor- Updated version http://cancerres.aacrjournals.org/content/68/10/3950 Access the most recent version of this article at: Cited Articles http://cancerres.aacrjournals.org/content/68/10/3950.full.html#ref-list-1 This article cites by 46 articles, 14 of which you can access for free at: Citing articles http://cancerres.aacrjournals.org/content/68/10/3950.full.html#related-urls This article has been cited by 4 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications Research. on July 9, 2014. © 2008 American Association for Cancer cancerres.aacrjournals.org Downloaded from Research. on July 9, 2014. © 2008 American Association for Cancer cancerres.aacrjournals.org Downloaded from
Transcript of Activation of Estrogen Receptor-A by the Anion Nitrite

2008;68:3950-3958. Cancer Res David J. Veselik, Shailaja Divekar, Sivanesan Dakshanamurthy, et al.
by the Anion NitriteαActivation of Estrogen Receptor-
Updated version
http://cancerres.aacrjournals.org/content/68/10/3950
Access the most recent version of this article at:
Cited Articles
http://cancerres.aacrjournals.org/content/68/10/3950.full.html#ref-list-1
This article cites by 46 articles, 14 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/68/10/3950.full.html#related-urls
This article has been cited by 4 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Activation of Estrogen Receptor-A by the Anion Nitrite
David J. Veselik,1Shailaja Divekar,
1Sivanesan Dakshanamurthy,
2Geoffrey B. Storchan,
1
Jasmine M.A. Turner,2Kelly L. Graham,
2Li Huang,
2
Adriana Stoica,2,3and Mary Beth Martin
1,2,3
Departments of 1Biochemistry and Molecular and Cellular Biology, 2Oncology, and 3Human Sciences, Georgetown University,Washington, District of Columbia
Abstract
In this study, the ability of nitrite and nitrate to mimic theeffects of estradiol on growth and gene expression wasmeasured in the human breast cancer cell line MCF-7. Similarto estradiol, treatment of MCF-7 cells with either 1 Mmol/Lnitrite or 1 Mmol/L nitrate resulted in f4-fold increase in cellgrowth and 2.3-fold to 3-fold increase in progesteronereceptor (PgR), pS2, and cathepsin D mRNAs that wereblocked by the antiestrogen ICI 182,780. The anions alsorecruited estrogen receptor-A (ERA) to the pS2 promoter andactivated exogenously expressed ERA when tested in transientcotransfection assays. To determine whether nitrite or nitratewas the active anion, diphenyleneiodonium was used toinhibit oxidation/reduction reactions in the cell. The abilityof diphenyleneiodonium to block the effects of nitrate, but notnitrite, on the induction of PgR mRNA and the activation ofexogenously expressed ERA suggests that nitrite is the activeanion. Concentrations of nitrite, as low as 100 nmol/L,induced a significant increase in PgR mRNA, suggesting thatphysiologically and environmentally relevant doses of theanion activate ERA. Nitrite activated the chimeric receptorGal-ER containing the DNA-binding domain of GAL-4 and theligand-binding domain of ERA and blocked the binding ofestradiol to the receptor, suggesting that the anion activatesERA through the ligand-binding domain. Mutational analysisidentified the amino acids Cys381, His516, Lys520, Lys529,Asn532, and His547 as important for nitrite activation of thereceptor. [Cancer Res 2008;68(10):3950–8]
Introduction
Over the past several decades, there has been a continuousincrease in the incidence rate of breast cancer in the United Statesand worldwide (1, 2). Screening practices and access to mammog-raphy are thought to have contributed to the increased incidencerate during the 1980s, but changes in screening and mammographydo not explain the increasing incidence rate before the 1980s or theincreasing rates in countries with low rates of screening. Thereason for the continuous increase is not known, but it has beensuggested that there is an increased exposure to known risk factorsor that there are new unidentified risk factors for the disease (2, 3).Family history is one risk factor (4). However, only 5% of breastcancers are believed to be truly hereditary (4), suggesting that mostcases are due to factors such as lifestyle and the environment.
Endocrine status is a prominent risk factor for breast cancer andincludes early age at menarche and late age at menopause (5),increasing age at first full-term pregnancy (6) and oral contra-ceptives in specific subgroups of women (7). Postmenopausalobesity is also a risk factor for the disease (3) due to thearomatization of adrenal estrogen precursors in adipose tissue.These well-established risk factors are estimated to account foronly f40% of breast cancer (8). Childhood exposure to ionizingradiation (9) and alcohol consumption (10) are also risk factors.However, these modest risk factors do not account for theremaining cases supporting the hypothesis that there are newunidentified risk factors.Because estrogens play an important underlying role in the
etiology of breast cancer, it has been suggested that environmentalexposures that mimic the effects of estrogens may be potentialrisk factors. However, the role of environmental estrogens is notclear. Early ecological and epidemiologic studies showed thatAsian populations that consume diets rich in phytoestrogens(natural estrogens) have a lower incidence of breast cancer, raisingthe possibility that phytoestrogens may be effective chemo-preventive agents. However, most epidemiologic studies do notshow a protective effect (reviewed in ref. 11). While phytoestrogensare thought to be protective, xenoestrogens (synthetic estrogens)are thought to increase the risk of breast cancer (12). However,most exposure studies do not show a clear correlationbetween xenoestrogens, such as polychlorinated biphenyls and/orp,p’-dichlorodiphenyl dichloroethylene, and the disease (reviewedin ref. 13). In addition to phytoestrogens and xenoestrogens, wehave identified a new potent class of environmental estrogens(14–18) called metalloestrogens. These metals fall into twosubclasses: metal/metalloid anions that include arsenite, selenite,and vanadate and bivalent cations that include cadmium, cobalt,copper, nickel, chromium, lead, mercury, and tin. In this study, weshow that the ubiquitous anions, nitrite and nitrate, mimic thefunctions of estradiol at environmentally relevant doses. Similar toestradiol, the anions induced the growth and expression ofestrogen-regulated genes in breast cancer cells and activatedestrogen receptor-a (ERa) in transient transfection experiments.The ability of the anions to mimic the effects of estradiol seems tobe due to the interaction of nitrite with the ligand-binding domainof the receptor.
Materials and Methods
Cell CultureMCF-7 cells were maintained in improved minimal essential medium
(IMEM; Biofluids, Inc.) and 5% fetal bovine serum (FBS). Cells (3 � 104)
were then plated into a six-well plate in 4 mL IMEM supplemented with 5%charcoal-stripped calf serum (CCS). At 40% confluence, the medium was
changed to phenol red–free IMEM with 5% CCS. Two days later, cells were
treated. After treatment, cells were counted with a Coulter counter.
Requests for reprints: Mary Beth Martin, Lombardi Comprehensive CancerCenter, Research Building, 3970 Reservoir Road NW, Washington, DC 20007. Phone:202-687-3768; E-mail: [email protected].
I2008 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-07-2783
Cancer Res 2008; 68: (10). May 15, 2008 3950 www.aacrjournals.org
Research Article
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

ImmunoblotsMCF-7 cells were lysed in 300 AL lysis buffer (400 mmol/L KCl, 50 mmol/L
Tris (pH 7.5; 1% NP40, 0.5% Na deoxycholate) in the presence of protease
inhibitors (Roche). Fifty micrograms of protein were loaded onto a 7.5%
polyacrylamide gel. Gels were run for 1.5 h at 150 V and transferred for 3 h at
100 V. The membranes were blocked, washed, and incubated with the
primary antibody progesterone receptor (PgR) antibody 1294, DakoCyto-
mation, Inc., and actin antibody, Santa Cruz Biotechnology). The membrane
was then washed and incubated with the secondary antibody (goat anti-
mouse IgG + IgM alkaline phosphatase, Amersham Biosciences). After
washing, the membrane was incubated with ECF substrate, scanned on the
Storm imaging system, and quantified using ImageQuant.
Chromatin ImmunoprecipitationProtein and DNA were cross-linked with 1% formaldehyde for 5 min at
25jC, washed in PBS containing protease inhibitors, collected in lysis
buffer [50 mmol/L Tris (pH 8.0), 10 mmol/L EDTA, 1% SDS], and disrupted
by sonication. The lysate was precleared by incubating with protein
G agarose/salmon sperm DNA and incubated first with ERa antibody
(H-184, Santa Cruz Biotech, Inc.) and then with protein G agarose/salmon
sperm DNA. After centrifugation, the agarose pellet was washed once with a
low salt buffer [20 mmol/L Tris (pH 8.0), 2 mmol/L EDTA, 150 mmol/L
NaCl, 0.1% sodium deoxycholate, 1% Triton X-100], twice with a high salt
buffer [20 mmol/L Tris (pH 8.0), 2 mmol/L EDTA, 500 mmol/L NaCl, 0.1%
sodium deoxycholate, 1% Triton X-100], once with a LiCl buffer [10 mmol/L
Tris (pH 8.0), 1 mmol/L EDTA, 250 mmol/L LiCl, 0.5% sodium deoxycholate,
05% NP40], and twice with a TE buffer [10 mmol/L Tris (pH 8.0), 1 mmol/L
EDTA]. The DNA/protein complex was eluted (1% SDS, 0.1 mol/L NaHCO3),
the crosslink was reversed with 5 mol/L NaCl, and the protein was digested
with proteinase K. DNA was purified with PCR purification kit (Qiagen). For
the PCR assay, the forward primer (5¶-GGCCATCTCTCACTATGAAT-CACTTCTGC-3¶), the reverse primer (5¶-GGCAGGCTCTGTTTGCTTAAA-GAGCG-3¶; ref. 19), 5 AL of DNA, and 33 cycles of amplification were used.
Real-Time Reverse Transcription–PCRThe RNA was isolated using the Trizol method. DNA was degraded with
DNase I. For the reverse transcriptase reaction, each 70 AL reaction
contained 7 AL of 10� Taqman RT buffer [500 mmol/L KCl, 100 mmol/LTris-HCl (pH 8.3)], 15.4 AL 25 mmol/L MgCl2, 14 AL deoxynucleotide
triphosphates, 3.5 AL random hexamers, 1.4 AL RNase inhibitor, 1.75 ALMuLV reverse transcriptase, and 1 Ag RNA. The mixture was incubated in athermal cycler for 10 min at 25jC, 30 min at 48jC, and 5 min at 95jC. Forthe real-time PCR reaction, each 25 AL reaction contained 12.5 AL UniversalMaster Mix, 1.25 AL of 20� Assay on Demand, and 5 AL cDNA. Sampleswere run on the 7900HT, and the data were analyzed by the 2�DDCt methodusing the SDS 2.1 software (Applied Biosystems).
Transfection AssaysCOS-1 cells (75 � 103) were plated into a 12-well plate in 1 mL of phenol
red–free IMEM supplemented with 5% CCS. At 50% confluence, the cellswere transfected with Fugene 6 and 2 Ag of DNA (1 ER:10 reporter:0.4
h-galactosidase). A/B-GAL and GAL-ER (20, 21) and the ER mutants C381A,C417A, C447A, H516A, N519A, K520A, E523Q, K529A, C530A, N532D,and
D538N (22–24) are described elsewhere. The ERa mutant H547A wasgenerated using a QuikChange site-directed mutagenesis kit. After
treatment, the amount of CAT activity was measured using the CAT
enzyme assay (Promega). Heat-inactivated cell lysate (100 AL) wasincubated with [14C]chloramphenicol and n-butryl CoA for 1 h at 37jC.The reaction product was extracted with mixed xylenes, and the amount of
radioactivity was measured. For the h-galactosidase assay, 8.0 mmol/L
O-nitrophenyl-b-D-galactopyranosidase were added to the cell lysate andincubated at 37jC for 2 h. The absorbance was read at 414 nm.
Binding AssayHuman recombinant ERa (4 � 10�9 mol/L; PanVera Corp.) was
incubated on ice for 1 h with sodium nitrite. [3H]Estradiol (10�8 mol/L)was added in the presence or absence of a 200-fold molar excess of
diethylstilbestrol and incubated for 2 h at 37jC. Dextran-coated charcoal
was added to each sample and incubated for 15 min at room temperature.The samples were centrifuged, and the supernatant was counted in a liquid
scintillation counter. Specific binding was calculated by subtracting
nonspecific binding from total binding.
Molecular Modeling of the Nitrite Ion into Potential BindingSitesModel construction, visualization, and analysis were performed with
Insight II (Accelrys, Inc.) and Sybyl 7.2 (Tripos, Inc.) in conjunction with the
molecular mechanics/dynamics package DISCOVER (Accelrys, Inc.).
Molecular mechanics was used to minimize the energy of the molecule
and to approach the global minimum energy state. Molecular dynamics wasthen performed on these molecules until they reached dynamic equilibrium.
Both techniques treat the molecules as a collection of interaction centers
(atoms) held together by parameterized intramolecular forces and are used
to represent the bonds between atoms and the van der Waals forcesbetween nonbonded atoms. Within the molecular dynamics simulation, the
molecules adjust their conformations to optimize the bond lengths and
angles to achieve a near ‘‘minimum energy’’ least strained structure,attained by a compromise minimum distortion. The force field is the sum of
the various contributions derived from a potential. The force field used was
the all-atom ‘‘consistent valence force field,’’ which includes explicit
hydrogens.Model building. The potential binding sites of nitrite were analyzed
based on the crystal structure of the ligand-binding domain of ERa(PDB: 3ERD; ref. 25), the chemical properties of nitrite, and the mutational
data. The oxyanion motif and the histidine, lysine, arginine, and cysteineside-chain residues were used in the identification of the binding sites.
Models were constructed with nitrite docked manually into different
binding sites. The quality of the models was checked by comparison of
coordination geometry with the representative models of nitrite-bindingproteins from the PDB database. Binding models were energy minimized
followed by 25 ps molecular dynamics simulations using DISCOVER. In
vacuo molecular dynamics simulations followed by energy minimizationson such dimolecular complex produced trigonal coordination geometries.
The hydrogen bond distances between nitrite and the side chains ranged
from 2.5 to 3.0 A.
Energy minimization. A series of algorithms was used for minimization,including an initial steepest descent stage followed by a conjugate gradient
stage converging to within the final mean square derivative threshold of
0.001 kcal mol�1 A�1. The efficient Newton-Raphson algorithm was avoided
because of instability when applied to the adopted energy expression.Periodic boundary conditions were applied using the explicit image
convention.
Molecular dynamics. A dynamic simulations protocol, consisting of amolecular dynamics run at different temperatures followed by minimiza-
tion, was found to be most effective for the dynamic behavior and binding
mode prediction of the nitrite ion. Crystallographic coordinates of ERa wereused as starting points, but no constraints were imposed during thesimulation, so that the simulated structures depend only on the applied
force field and not on the initial geometries. Insight II/DISCOVER, using the
consistent valence force field, was used in the molecular dynamics
simulations. Periodic boundary conditions, with bonds across theboundaries in the chain direction, were used in these dynamic simulations.
The Ewald summation method was used in the calculation of nonbonded
interactions. The simulations were performed at 298 K under constantpressure and temperature assembly conditions and consisted of 10,000
molecular dynamics steps in 1 fs intervals. The effect of cutoff distances was
examined at rc = 12.0 A. All other simulation variables were kept on default.
All systems were equilibrated under the simulation conditions for at least10 ps before collecting the trajectories for subsequent analysis.
Results
Effects of nitrite and nitrate on estrogen responses in MCF-7cells. To determine whether nitrite and/or nitrate have estrogen-like effects, the ability of the anions to replace estradiol in the
Nitrite Activates ERa
www.aacrjournals.org 3951 Cancer Res 2008; 68: (10). May 15, 2008
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
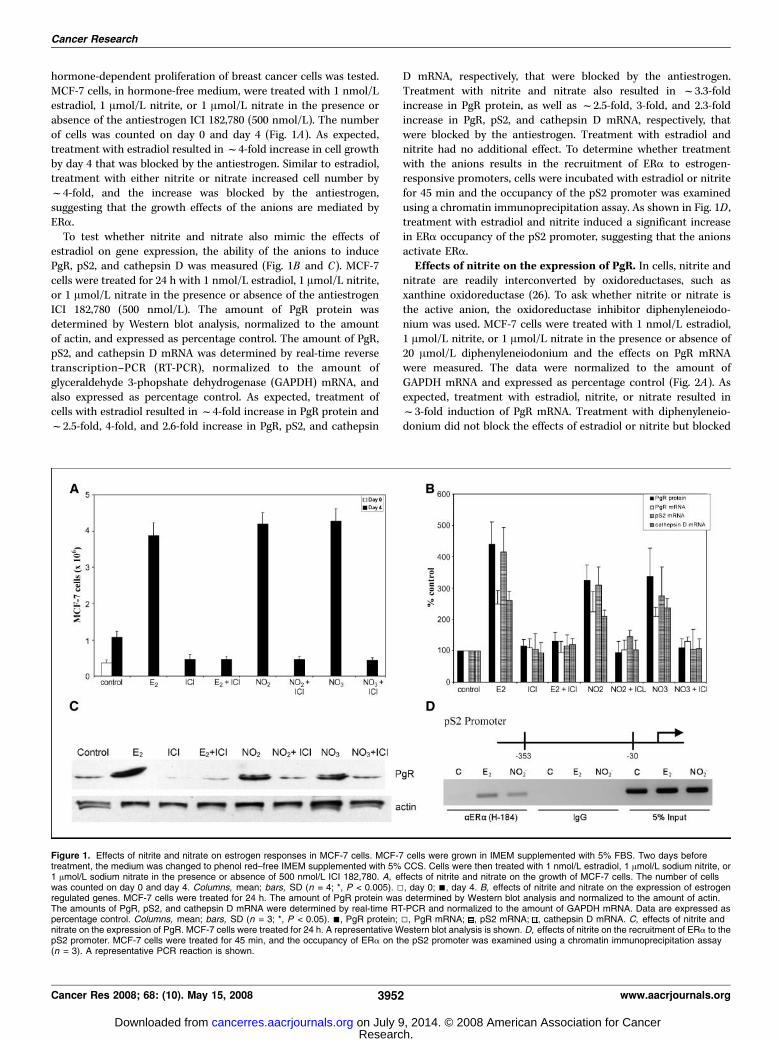
hormone-dependent proliferation of breast cancer cells was tested.MCF-7 cells, in hormone-free medium, were treated with 1 nmol/Lestradiol, 1 Amol/L nitrite, or 1 Amol/L nitrate in the presence orabsence of the antiestrogen ICI 182,780 (500 nmol/L). The numberof cells was counted on day 0 and day 4 (Fig. 1A). As expected,treatment with estradiol resulted inf4-fold increase in cell growthby day 4 that was blocked by the antiestrogen. Similar to estradiol,treatment with either nitrite or nitrate increased cell number byf4-fold, and the increase was blocked by the antiestrogen,suggesting that the growth effects of the anions are mediated byERa.To test whether nitrite and nitrate also mimic the effects of
estradiol on gene expression, the ability of the anions to inducePgR, pS2, and cathepsin D was measured (Fig. 1B and C). MCF-7cells were treated for 24 h with 1 nmol/L estradiol, 1 Amol/L nitrite,or 1 Amol/L nitrate in the presence or absence of the antiestrogenICI 182,780 (500 nmol/L). The amount of PgR protein wasdetermined by Western blot analysis, normalized to the amountof actin, and expressed as percentage control. The amount of PgR,pS2, and cathepsin D mRNA was determined by real-time reversetranscription–PCR (RT-PCR), normalized to the amount ofglyceraldehyde 3-phopshate dehydrogenase (GAPDH) mRNA, andalso expressed as percentage control. As expected, treatment ofcells with estradiol resulted inf4-fold increase in PgR protein andf2.5-fold, 4-fold, and 2.6-fold increase in PgR, pS2, and cathepsin
D mRNA, respectively, that were blocked by the antiestrogen.Treatment with nitrite and nitrate also resulted in f3.3-foldincrease in PgR protein, as well as f2.5-fold, 3-fold, and 2.3-foldincrease in PgR, pS2, and cathepsin D mRNA, respectively, thatwere blocked by the antiestrogen. Treatment with estradiol andnitrite had no additional effect. To determine whether treatmentwith the anions results in the recruitment of ERa to estrogen-responsive promoters, cells were incubated with estradiol or nitritefor 45 min and the occupancy of the pS2 promoter was examinedusing a chromatin immunoprecipitation assay. As shown in Fig. 1D ,treatment with estradiol and nitrite induced a significant increasein ERa occupancy of the pS2 promoter, suggesting that the anionsactivate ERa.
Effects of nitrite on the expression of PgR. In cells, nitrite andnitrate are readily interconverted by oxidoreductases, such asxanthine oxidoreductase (26). To ask whether nitrite or nitrate isthe active anion, the oxidoreductase inhibitor diphenyleneiodo-nium was used. MCF-7 cells were treated with 1 nmol/L estradiol,1 Amol/L nitrite, or 1 Amol/L nitrate in the presence or absence of20 Amol/L diphenyleneiodonium and the effects on PgR mRNAwere measured. The data were normalized to the amount ofGAPDH mRNA and expressed as percentage control (Fig. 2A). Asexpected, treatment with estradiol, nitrite, or nitrate resulted inf3-fold induction of PgR mRNA. Treatment with diphenyleneio-donium did not block the effects of estradiol or nitrite but blocked
Figure 1. Effects of nitrite and nitrate on estrogen responses in MCF-7 cells. MCF-7 cells were grown in IMEM supplemented with 5% FBS. Two days beforetreatment, the medium was changed to phenol red–free IMEM supplemented with 5% CCS. Cells were then treated with 1 nmol/L estradiol, 1 Amol/L sodium nitrite, or1 Amol/L sodium nitrate in the presence or absence of 500 nmol/L ICI 182,780. A, effects of nitrite and nitrate on the growth of MCF-7 cells. The number of cellswas counted on day 0 and day 4. Columns, mean; bars, SD (n = 4; *, P < 0.005). 5, day 0; n, day 4. B, effects of nitrite and nitrate on the expression of estrogenregulated genes. MCF-7 cells were treated for 24 h. The amount of PgR protein was determined by Western blot analysis and normalized to the amount of actin.The amounts of PgR, pS2, and cathepsin D mRNA were determined by real-time RT-PCR and normalized to the amount of GAPDH mRNA. Data are expressed aspercentage control. Columns, mean; bars, SD (n = 3; *, P < 0.05). n, PgR protein; 5, PgR mRNA; , pS2 mRNA; , cathepsin D mRNA. C, effects of nitrite andnitrate on the expression of PgR. MCF-7 cells were treated for 24 h. A representative Western blot analysis is shown. D, effects of nitrite on the recruitment of ERa to thepS2 promoter. MCF-7 cells were treated for 45 min, and the occupancy of ERa on the pS2 promoter was examined using a chromatin immunoprecipitation assay(n = 3). A representative PCR reaction is shown.
Cancer Research
Cancer Res 2008; 68: (10). May 15, 2008 3952 www.aacrjournals.org
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

the effects of nitrate on the induction of PgR, suggesting that nitriteis the anion that activates ERa.To determine the concentrations of nitrite that induce the
expression of PgR, MCF-7 cells were treated for 24 h with nitritedoses ranging from 0.01 nmol/L to 10 Amol/L. The effects on PgRmRNA were measured using the real-time RT-PCR assay (Fig. 2B).Treatment with 100 nmol/L, 1 Amol/L, or 10 Amol/L nitriteresulted in f2.3-fold, 3.7-fold, and 4.1-fold induction, respectively,indicating that, at physiologically and environmentally relevantconcentrations, nitrite has estrogenic activity.
Effects of nitrite and nitrate on the activity of ERA. To testthe ability of nitrite and nitrate to activate ERa, COS-1 cells weretransiently cotransfected with wild-type ERa and an estrogenresponse element-CAT reporter construct. The transfected cellswere treated for 24 h with 1 nmol/L estradiol, 1 Amol/L nitrite, or1 Amol/L nitrate in the presence or absence of the antiestrogenICI 182,780 (500 nmol/L) or the oxidoreductase inhibitordiphenyleneiodonium (20 Amol/L).CAT activity was measured,normalized to the amount of h-galactosidase activity, and
expressed as percentage control. Treatment with estradiol resultedin f3-fold to 4-fold increase in CAT activity that was blocked bythe antiestrogen. Similar to estradiol, treatment with either nitriteor nitrate resulted inf3-fold increase in CAT activity that was alsoblocked by the antiestrogen (Fig. 3A), demonstrating the ability ofthe anions to activate exogenously expressed ERa. The oxidore-ductase inhibitor had no effect on the induction of CAT activity byestradiol or nitrite but blocked the induction of CAT activity bynitrate (Fig. 3B) providing further support that nitrite is the anionthat activates ERa.
Effects of nitrite and nitrate on the activation of ERAmutants. To identify the region of ERa activated by nitrite,transient transfection assays were conducted using chimericreceptors (20, 21). To determine whether the anion activates thereceptor through the NH2 terminus, the A/B-GAL chimeracontaining the A/B domain of ERa fused to the DNA-bindingdomain of the yeast transcription factor GAL4 was used. Toestablish whether the anions activate through the COOH terminus,the GAL-ER chimera was used. The latter chimera contains theDNA-binding domain of GAL4 fused to the ligand-binding domainof ERa. In these assays, the chimeric receptors were transientlycotransfected with a GAL4-CAT reporter construct into COS-1cells and the cells were treated for 24 hours with 1 nmol/L
Figure 2. Effects of nitrite on estrogen responses in MCF-7 cells. MCF-7cells were grown and treated as described in Fig. 1. Total mRNA was isolated,and the amount of PgR mRNA was determined by real-time RT-PCR. Datawere normalized to the amount of GAPDH mRNA and expressed as percentagecontrol. A, effects of diphenyleneiodonium on PgR induction by nitrite andnitrate. MCF-7 cells were treated for 24 h with 1 nmol/L estradiol, 1 Amol/Lsodium nitrite, or 1 Amol/L sodium nitrate in the presence or absence of20 Amol/L diphenyleneiodonium (DPI). Columns, mean; bars, SD (n = 3;*, P < 0.05). B, dose effect of nitrite on the expression of PgR. MCF-7 cells weretreated with 0.01 nmol/L, 0.1 nmol/L, 1 nmol/L, 10 nmol/L, 0.1 Amol/L,1 Amol/L, or 10 Amol/L sodium nitrite for 24 h. Points, mean; bars, SD (n = 3;P < 0.05 for 0.1–10 Amol/L).
Figure 3. Effects of nitrite and nitrate on the activity of ERa. COS-1 cells weregrown in phenol red–free IMEM supplemented with 5% CCS and transientlycotransfected with wild-type ERa and an estrogen response element-CATreporter construct. The transfected cells were treated for 24 h with 1 nmol/Lestradiol, 1 Amol/L sodium nitrite, or 1 Amol/L sodium nitrate in the presence orabsence of 500 nmol/L ICI 182,780 (A ) or 20 Amol/L diphenyleneiodoniumchloride (B). CAT activity was measured, normalized to the amount ofh-galactosidase activity, and expressed as percentage control. Columns, mean;bars, SD (n = 3; *, P < 0.05).
Nitrite Activates ERa
www.aacrjournals.org 3953 Cancer Res 2008; 68: (10). May 15, 2008
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from
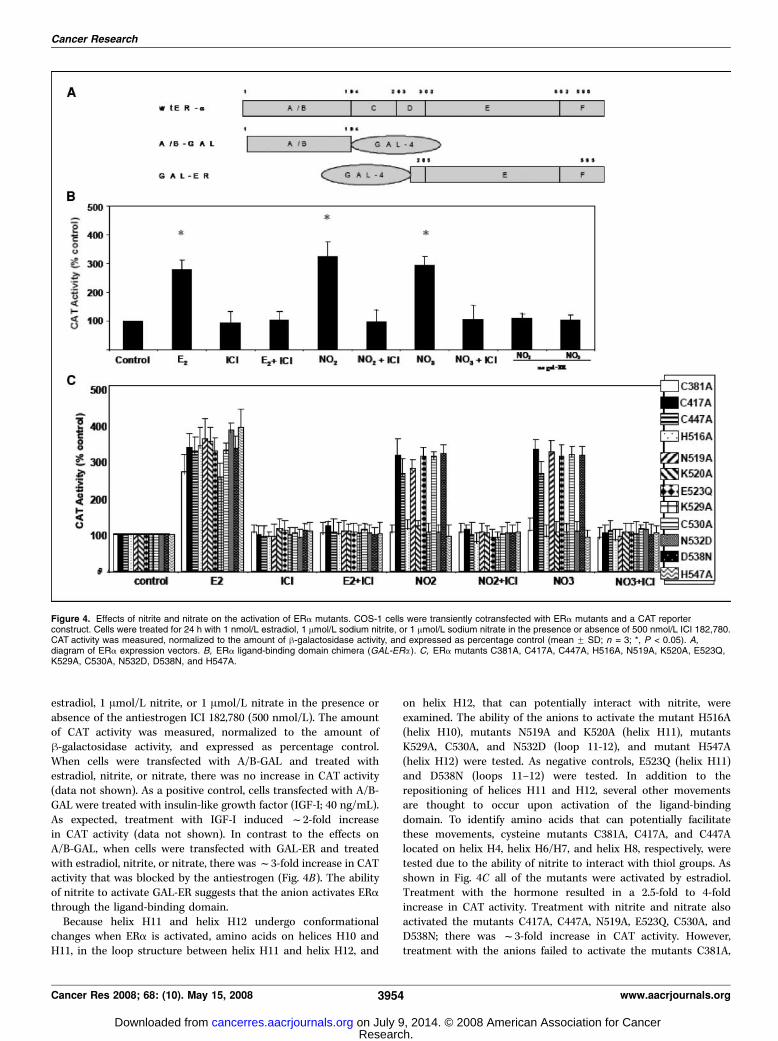
estradiol, 1 Amol/L nitrite, or 1 Amol/L nitrate in the presence orabsence of the antiestrogen ICI 182,780 (500 nmol/L). The amountof CAT activity was measured, normalized to the amount ofh-galactosidase activity, and expressed as percentage control.When cells were transfected with A/B-GAL and treated withestradiol, nitrite, or nitrate, there was no increase in CAT activity(data not shown). As a positive control, cells transfected with A/B-GAL were treated with insulin-like growth factor (IGF-I; 40 ng/mL).As expected, treatment with IGF-I induced f2-fold increasein CAT activity (data not shown). In contrast to the effects onA/B-GAL, when cells were transfected with GAL-ER and treatedwith estradiol, nitrite, or nitrate, there wasf3-fold increase in CATactivity that was blocked by the antiestrogen (Fig. 4B). The abilityof nitrite to activate GAL-ER suggests that the anion activates ERathrough the ligand-binding domain.Because helix H11 and helix H12 undergo conformational
changes when ERa is activated, amino acids on helices H10 andH11, in the loop structure between helix H11 and helix H12, and
on helix H12, that can potentially interact with nitrite, wereexamined. The ability of the anions to activate the mutant H516A(helix H10), mutants N519A and K520A (helix H11), mutantsK529A, C530A, and N532D (loop 11-12), and mutant H547A(helix H12) were tested. As negative controls, E523Q (helix H11)and D538N (loops 11–12) were tested. In addition to therepositioning of helices H11 and H12, several other movementsare thought to occur upon activation of the ligand-bindingdomain. To identify amino acids that can potentially facilitatethese movements, cysteine mutants C381A, C417A, and C447Alocated on helix H4, helix H6/H7, and helix H8, respectively, weretested due to the ability of nitrite to interact with thiol groups. Asshown in Fig. 4C all of the mutants were activated by estradiol.Treatment with the hormone resulted in a 2.5-fold to 4-foldincrease in CAT activity. Treatment with nitrite and nitrate alsoactivated the mutants C417A, C447A, N519A, E523Q, C530A, andD538N; there was f3-fold increase in CAT activity. However,treatment with the anions failed to activate the mutants C381A,
Figure 4. Effects of nitrite and nitrate on the activation of ERa mutants. COS-1 cells were transiently cotransfected with ERa mutants and a CAT reporterconstruct. Cells were treated for 24 h with 1 nmol/L estradiol, 1 Amol/L sodium nitrite, or 1 Amol/L sodium nitrate in the presence or absence of 500 nmol/L ICI 182,780.CAT activity was measured, normalized to the amount of h-galactosidase activity, and expressed as percentage control (mean F SD; n = 3; *, P < 0.05). A,diagram of ERa expression vectors. B, ERa ligand-binding domain chimera (GAL-ERa). C, ERa mutants C381A, C417A, C447A, H516A, N519A, K520A, E523Q,K529A, C530A, N532D, D538N, and H547A.
Cancer Research
Cancer Res 2008; 68: (10). May 15, 2008 3954 www.aacrjournals.org
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

H516A, K520A, K529A, N532D, and H547A suggesting that theseamino acids are involved in nitrite activation of the receptor.
Effects of nitrite on estradiol binding to ERA. To determinewhether nitrite blocks the binding of estradiol to ERa, purifiedhuman recombinant ERa was incubated with various concen-trations of nitrite (10�12 to 10�6 mol/L) for 1 hour. [3H]Estradiol(10 nmol/L) was then added in the presence or absence of a200-fold molar excess of diethylstilbestrol for 2 hours at 37jC.Specific binding of [3H]estradiol was determined and expressed aspercentage control. As shown in Fig. 5, increasing concentrations ofnitrite blocked the binding of estradiol to ERa suggests that theanion interacts directly with the ligand-binding domain.
Molecular modeling of the nitrite ion into potential bindingsites of ERA. The mutational analysis described above identifiedcys381 on helix H4, his516 on helix H10, lys520 and lys529 onhelix H11, asn532 in the loop between H11 and H12, and his547on helix H12 as potential interaction sites of nitrite with ERa.Based on these data, the crystal structure of the ligand-bindingdomain of ERa (PDB: 3ERD; ref. 25), and the chemical propertiesof nitrite, potential binding sites of the anion with the receptorwere identified using molecular modeling. For the construction,visualization, and analysis of the model, Insight II and Sybyl7.2 were used. Three potential binding sites were identified fornitrite on the solvent accessible surface of the ligand-bindingdomain (Fig. 6A–C). Site 1 was formed by a direct interaction ofnitrite with Lys529 on helix H11 and Asn532 in the loop betweenhelices H11 and H12 and by an indirect interaction through waterwith Val534 also in the loop between helices H11 and H12. Site2 was formed by a direct interaction of the anion with His516 onhelix H10 and lys520 on helix H11 and an indirect interactionthrough water with Asn519 at the interface of helices H10 andH11 and Glu523 on helix H11. The third site, site 3, was formedby the direct interaction of nitrite with Cys381 on helix H4,Arg515 on helix H10, and His547 on helix H12. The identificationof three potential binding sites formed by amino acids ondifferent helices suggests that nitrite promotes a conformationalchange in the ligand-binding domain, thereby activating thereceptor.
Discussion
This study shows the ability of physiologically and environmen-tally relevant doses of nitrite and nitrate to mimic the effects ofestradiol in human breast cancer cells. The ability of low doses ofnitrite and nitrate to mimic estradiol suggests that the anions mayplay a role in the etiology and progression of breast cancer due, inpart, to their ability to activate ERa. Human exposure to theseanions occurs through both exogenous and endogenous sources asa result of the environment, diet, and medicinal drugs and as thebreakdown product of nitric oxide. In surface and drinking water,nitrate is generally low but has increased over the last few decades(27) due to the use of nitrogen-based fertilizers, contaminationfrom refuse dumps, oxidation of ammonia from human and animalwaste, and treatment of drinking water with chloramines (28). Highamounts of nitrate are also present in some fruits and vegetables asa result of cultivation in greenhouses and in cured and processedmeats due to their addition as preservatives and color enhancers(29). Medications, including antimalarials, antidiarrheals, diuretics,and vasodilators, also contribute to exposure. Our total estimateddaily exposure to exogenous nitrites and nitrates ranges from 1.2 to3 mg and from 39 to 268 mg, respectively (30). Exposure to theanions also occurs as a result of the endogenous oxidation of nitricoxide to nitrites and nitrates. Nitric oxide is synthesized by a familyof nitric oxide synthases (NOS). In serum, the normal concen-tration of nitrite is f18.0 to 43.5 Amol/L (31–34) whereas, inbreast cancer patients, serum concentrations range from 246 to363 Amol/L (35). The amount of nitrite is also elevated in breasttissue from breast cancer patients (745 nmol/g wet weight)compared with the amount in benign breast disease (442 nmol/gwet weight; ref. 36). In breast tumors, the expression of NOS isassociated with increased proliferation and tumor grade and theexpression of PgR and nitric oxide is associated with invasivedisease (37). Similarly, in animal models, nitric oxide promotesthe progression of mammary tumors (38–40). Interestingly, epide-miologic studies suggest a link between postmenopausal breastcancer and farming (41, 42). There is also evidence that nitric oxideand/or its metabolites have other endocrine-like effects. In olderwomen, intermittent use of nitrates is associated with increasedbone mineral density (43), and in the neuronal NOS knockoutmouse, more aggressive and inappropriate sexual behavior is ob-served (44). Taken together, these observations suggest a possibleendocrine-like role for nitrite and nitrates in breast cancer.The ability of nitrite and nitrate to mimic the effects of estradiol
is due to an interaction of nitrite with the ligand-binding domain ofERa. Similar to other nuclear receptors (25, 45–47), the ligand-binding domain of ERa contains 11 a helices (H1–H12 minus H2)folded into a three-layered antiparallel a helical sandwich. Thecentral core layer contains three a-helices (H5/6, H9, and H10)sandwiched between two additional layers of helices composed ofH1-H4, H7, H8, and H11 and flanked by helix H12 (46, 47). Uponbinding, the ligand induces a conformational change that results ina transcriptionally active receptor. Based upon the crystal structureof apo-RXR-a and holo-RXR-a (45, 48), several major structuralchanges are thought to occur as a result of ligand binding; helixH12 is repositioned over the ligand-binding pocket; helix H11rotates f180j around its helical axis and tilts away from thepocket; the NH2 terminal end of helix H3 rotates f90j around itsaxis and bends toward helix H4 and the core of the pocket; theadjacent N loop relocates under helix H6; and the h turns, locatedbetween helices H4/H5 and H6, shift away from the core of thepocket (45, 48). To account for these structural changes, it has been
Figure 5. Effects of nitrite on estradiol binding to ERa. Purified humanrecombinant ERa was incubated with various concentrations of sodium nitrite(10�12 to 10�6 mol/L) for 1 h. [3H]Estradiol (10 nmol/L) was added in thepresence or absence of a 200-fold molar excess of diethylstilbestrol for 2 h at37jC. The amount of specific binding of [3H]estradiol was determined andexpressed as percentage control. Columns, mean; bars, SD (n = 3).
Nitrite Activates ERa
www.aacrjournals.org 3955 Cancer Res 2008; 68: (10). May 15, 2008
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

proposed (49, 50) that the retinoid is initially attracted into theligand-binding pocket by electrostatic interactions. Once inside thepocket, the retinoid induces the rearrangement of helix H3 which,in turn, causes helix H11 to rotate and to form a continuous benthelix with helix H10. The repositioning of helix H11 pulls helix H12under helix H4 freeing the N loop from its interaction with helixH12. The repositioning of helices H3 and H12 results in theformation of a shallow hydrophobic groove that constitutes theAF-2 domain, the binding site for steroid receptor coactivators. Inthe case of ERa, it has been suggested that, in addition to inducinga conformational change, estradiol functions inside the pocket as ascaffold for the structure of the ligand-binding domain (47). Thehydroxyl group on the A ring of the steroid hydrogen bonds withGlu353 on helix H3, Arg394 on helix H5, and a water molecule,whereas the hydroxyl group on the D ring hydrogen bonds withHis524 on helix H11. The steroid backbone forms van der Waals
interactions with hydrophobic amino acids on helices H3, H5, H7,and H11 and in the h-turn (46, 47). In contrast to estradiol, nitriteseems to interact with amino acids on the solvent accessiblesurfaces of the ligand-binding domain. Mutational analysisidentified Cys381 on helix H4, His516 on helix H10, Lys520 andLys529 on helix H11, Asn532 in the loop between H11 and H12, andHis547 on helix H12 as potential sites of interaction between nitriteand the receptor. Molecular modeling suggests that these aminoacids form three potential binding sites on the surface of theactivated receptor. Site 1 is formed by the interaction of nitrite withLys529 on helix H11 and Asn532 in the loop between helices H11and H12; site 2 results from the interaction of the anion withHis516 on helix H10 and Lys520 on helix H11, whereas site 3 is dueto an interaction of nitrite with Cys381 on helix H4 and His547 onhelix H12. Molecular modeling also identified Val534 in the loopbetween helices H11 and H12 as a potential amino acid in site
Figure 6. Molecular model of the interaction of nitrite with ERa. Three potential nitrite-binding sites on the solvent accessible surface of the ligand-binding domainof ERa were identified by computational analysis as described in Materials and Methods. A shows site 1 which is formed by Lys529 on helix H11 and Asn532 andVal534 in the loop between helices H11 and H12. B displays site 2 which is formed by His516 on helix H10 and Asn519, Lys520, and Glu523 on helix H11. C showssite 3 which is composed of Cys381 on helix H4, Arg515 on helix H10, Asn519 on helix H11, and His547 on helix H12. D shows the proposed model of ERaactivation by nitrite. In step 1, nitrite interacts with site 1 on helix H11 and in the loop between helices H11 and H12, resulting in the movement of helix H12 toward theligand-binding pocket. In step 2, the anion interacts with site 2 on helix H10 and on helix H11 resulting in the formation of a continuous bent helix between helicesH10 and H11. In step 3, nitrite interacts with site 3 on helix H4 and helix H12 pulling helix H12 under helix H4 resulting in the displacement of the N loop and therepositioning of helix H3 to form the coactivator binding site. Water molecules are represented by balls (red); nitrite and amino acid side chains are presented as ball andstick model.
Cancer Research
Cancer Res 2008; 68: (10). May 15, 2008 3956 www.aacrjournals.org
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

1; Asn519 and Glu523 on helix H11 as potential amino acids in site2; and Arg515 on helix H10 as a potential amino acid in site 3. Theinteractions of nitrite with Val534, Asn519, and Glu523 seem to beindirect interactions through water and may explain the ability ofnitrite to activate the N519A mutant. Although the precisemechanism by which nitrite interacts with these sites to activateERa remains to be defined, it is proposed that the interaction ofthe anion with these sites induces structural changes that mimicthe structural changes induced upon the binding of estradiol(Fig. 6D). In step 1, the interaction of nitrite with Lys529 on helixH11 and Asn532 in the loop between helices H11 and H12 (site 1)may result in the movement of helix H12 over the ligand-bindingpocket; in step 2, the interaction with His516 on helix H10 andLys520 on helix H11 (site 2) would result in the formation of acontinuous bent helix between helices H10 and H11; and in step3, the interaction of nitrite with Cys381 on helix H4 and His547 onhelix H12 (site 3) would pull helix H12 under helix H4 resulting inthe displacement of the N loop that causes the repositioning ofhelix H3 to form the coactivator binding site. It is not knownwhether nitrite interacts with these sites in a sequential manner orwhether the anion occupies all three sites simultaneously. In asimultaneous model, three molecules of nitrite bind to the ligand-binding domain resulting in the formation of the activeconformation of the receptor. In a sequential model (Fig. 6D), site1 is postulated as the initial entrant site located in the flexibleregion near the COOH terminal end of the ligand-binding domain.
As a consequence of the flexibility of the ligand-binding domainand the conformational changes that occur upon binding of nitrite,the anion is displaced from site 1 and moves into the secondbinding site and subsequently into the third binding site. Becausethe molecular modeling is based on the static, activated receptor,not all of the amino acids in transient interaction sites will beidentified. For example, if sites 1 and 2 are transient binding sites, itis possible that additional amino acids may be involved in theinteraction of the anion with these sites and between the sites.These models remain to be tested. The amino acids involved innitrite activation of ERa are conserved in ERh, suggesting that theanion may also activate other steroid receptors.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Received 7/20/2007; revised 2/8/2008; accepted 3/17/2008.Grant support: Komen Foundation, AICR, NIH ES11745, NIH P30-CA51008, P50-
CA58185, and U54 CA0100970.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.We thank Prof. P. Chambon and Dr. B. Katzenellenbogen for estrogen receptor
mutants, Dr. S. Byers, Dr. B. Katzenellenbogen, and Dr. A.T. Reigel for helpfuldiscussion, National Cancer Institute for allocation of computing time, and staffsupport at the Advanced Biomedical Computing Center.
Nitrite Activates ERa
www.aacrjournals.org 3957 Cancer Res 2008; 68: (10). May 15, 2008
References
1. Richie RC, Swanson JO. Breast cancer: a review of theliterature. J Insur Med 2003;35:85–101.
2. Althuis MD, Dozier JM, Anderson WF, Devesa SS,Brinton LA. Global trends in breast cancer incidenceand mortality 1973–1997. Int J Epidemiol 2005;34:405–12.
3. Feigelson HS, Jonas CR, Teras LR, Thun MJ, Calle EE.Weight gain, body mass index, hormone replacementtherapy, and postmenopausal breast cancer in a largeprospective study. Cancer Epidemiol Biomarkers Prev2004;13:220–4.
4. Miki Y, Swensen J, Shattuck-Eidens D, et al. A strongcandidate for the breast and ovarian cancer suscepti-bility gene BRCA1. Science 1994;266:66–71.
5. Kampert JB, Whittemore AS, Paffenbarger RS, Jr.Combined effect of child-bearing, menstrual events,and body size on age-specific breast cancer risk. Am JEpidemiol 1988;128:962–79.
6. MacMahon B, Cole P, Brown J. Etiology of humanbreast cancer: a review. J Natl Cancer Inst 1973;50:21–42.
7. Steinberg KK, Thacker SB, Smith SJ. A meta-analysis ofthe effect of estrogen replacement therapy on the risk ofbreast cancer. JAMA 1991;265:1985–90.
8. Madigan MP, Ziegler RG, Benichou J, Byrne C, HooverRN. Proportion of breast cancer cases in the UnitedStates explained by well-established risk factors. J NatlCancer Inst 1995;87:1681–5.
9. Tokunaga M, Land CE, Yamamoto T, et al. Incidence offemale breast cancer among atomic bomb survivors,Hiroshima and Nagasaki, 1950–1980. Radiat Res 1987;112:243–72.
10. Dorgan JF, Baer DJ, Albert PS, et al. Serum hormonesand the alcohol-breast cancer association in postmen-opausal women. J Natl Cancer Inst 2001;93:710–5.
11. Peeters PH, Keinan-Boker L, van der Schouw YT,Grobbee DE. Phytoestrogens and breast cancer risk.Review of the epidemiological evidence. Breast CancerRes Treat 2003;77:171–83.
12. Colborn T, von Saal FS, Soto AM. Developmentaleffects of endocrine-disrupting chemicals in wildlife andhumans. Environ Health Perspect 1993;101:378–84.
13. Safe S. Endocrine disruptors and human health: isthere a problem? Toxicology 2004;205:3–10.
14. Garcia-Morales P, Saceda M, Kenney N, et al. Effect ofcadmium on estrogen receptor levels and estrogen-induced responses in human breast cancer cells. J BiolChem 1994;269:16896–901.
15. Stoica A, Katzenellenbogen BS, Martin MB. Activa-tion of estrogen receptor-a by the heavy metalcadmium. Mol Endocrinol 2000;14:545–53.
16. Stoica A, Pentecost E, Martin MB. Effect of arseniteon estrogen receptor-a expression and activity inMCF-7 breast cancer cells. Endocrinology 2000;141:3595–602.
17. Stoica A, Pentecost E, Martin MB. Effects of seleniteon estrogen receptor-a expression and activity in MCF-7breast cancer cells. J Cell Biochem 2000;79:282–92.
18. Martin MB, Reiter R, Pham T, et al. Estrogen likeactivity of metals in MCF-7 breast cancer cells.Endocrinology 2003;144:2425–36.
19. Shao W, Halachmi S, Brown M. ERAP140, a conservedtissue-specific nuclear receptor coactivator. Mol CellBiol 2002;22:3358–72.
20. Webster NJ, Green S, Jin JR, Chambon P. Thehormone-binding domains of the estrogen and gluco-corticoid receptors contain an inducible transcriptionalactivation function. Cell 1988;54:199–207.
21. Tora L, White J, Brou C, et al. The human estrogenreceptor has two independent nonacidic transcriptionalactivation functions. Cell 1989;59:477–87.
22. Reese JC, Katzenellenbogen BS. Mutagensis ofcysteines in the hormone binding domain of the humanestrogen receptor. J Biol Chem 1991;266:10880–7.
23. Pakdel F, Katzenellenbogen BS. Human estrogenreceptor mutants with altered estrogen and anties-trogen ligand discrimination. J Biol Chem 1992;267:3429–37.
24. Wrenn CK, Katzenellenbogen BS. Structure-functionanalysis of the hormone binding domain of the humanestrogen receptor by region-specific mutagenesis andphenotypic screening in yeast. J Biol Chem 1993;268:24089–98.
25. Shiau AK, Barstad D, Loria PM, et al. The structuralbasis of estrogen receptor/coactivator recognition and
the antagonism of this interaction by tamoxifen. Cell1998;95:927–37.
26. Millar TM, Stevens CR, Benjamin N, Eisenthal R,Harrison R, Blake DR. Xanthine oxidoreductase cataly-ses the reduction of nitrates and nitrite to nitric oxideunder hypoxic conditions. FEBS Lett 1998;427:225–8.
27. WHO. Guidelines for drinking water quality. Healthcriteria and other supporting information. 2nd ed.Geneva: WHO; 1998.
28. Nitrification occurrence and control in chlorami-nated water systems. Denver (CO): American WaterWorks Association Research Foundation; 1995.
29. WHO. Evaluation of certain food additives andcontaminants Geneva: World Health Organization, JointFAO/WHO Expert Committee on Food Additives WHOTechnical Report Series No.859, 1995.
30. ECETOC. Nitrate and drinking water Brussels:European Chemical Industry Ecology and ToxicologyCentre Technical Report No. 27, 1988.
31. Kleinbongard P, Dejam A, Lauer T, et al. Plasmanitrite reflects constitutive nitric oxide synthase activityin mammals. Free Radic Biol Med 2003;35:790–6.
32. Hirokawa T, Yoshioka M, Okamoto H, Timerbaev AR,Blaschke G. High-sensitivity capillary electrophoresisdetermination of inorganic anions in serum and urineusing on-line preconcentration by transient isotacho-phoresis. J Chromatogr B Analyt Technol Biomed LifeSci 2004;811:165–70.
33. Hwang J, Mack WJ, Xiang M, Sevanian A, Lobo RA,Hodis HN. Long-term effect of estrogen replacement onplasma nitric oxide levels: results from the estrogen inthe prevention of atherosclerosis trial (EPAT). Athero-sclerosis 2005;181:375–80.
34. Mansoor MA, Kristensen O, Hervig T, et al.Relationship between serum folate and plasma nitrateconcentrations: possible clinical implications. ClinChem 2005;51:1266–8.
35. Gunel N, Coskun U, Sancak B, Gunel U, Hasdemir O,Bozkurt S. Clinical importance of serum interleukin-18and nitric oxide activities in breast cancer patients.Cancer 2002;95:663–7.
36. Gonenc A, Erten D, Aslan S, Akinci M, Simsek B,Torun M. Lipid peroxidation and antioxidant status in
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from

Cancer Research
Cancer Res 2008; 68: (10). May 15, 2008 3958 www.aacrjournals.org
blood and tissue of malignant breast tumor and benignbreast disease. Cell Biol Int 2006;30:376–80.
37. Reveneau S, Arnould L, Jolimoy G, et al. Nitric oxidesynthase in human breast cancer is associated withtumor grade, proliferation rate, and expression ofprogesterone receptors. Lab Invest 1999;79:1215–25.
38. Jadeski LC, Charkraborty C, Lala PK. Nitric oxidemediated promotion of mammary tumour cell migra-tion requires sequential activation of nitric oxidesynthase, guanylate cyclase and mitogen activatedprotein kinase. Int J Cancer 2003;106:496–504.
39. Jang TJ, Kim DK. Inducible nitric oxide synthaseexpression to tumor and stromal cells is associatedwith the progression of 7,12-dimetylbenz(a)anthraceneinduced rat mammary tumors. Cancer Lett 2003;182:121–6.
40. Ellies LG, Fishman M, Hardison J, et al. Mammarytumor latency is increased in mice lacking the induciblenitric oxide synthase. Int J Cancer 2003;106:1–7.
41. Gardner KM, Ou SX, Jin F, et al. Occupations andbreast cancer risk among Chinese women in urbanShanghai. Am J Ind Med 2002;42:296–308.
42. Band PR, Le ND, Fang R, Deschamps M,Gallagher RP, Yang P. Identification of occupationalcancer risks in British Columbia. A population-based case-control study of 995 incident breastcancer cases by menopausal status, controlling forconfounding factors. J Occup Environ Med 2000;42:284–310.
43. Jamal SA, Browner WS, Bauer DC, Cummings SR.Intermittent use of nitrates increases bone mineraldensity: the study of osteoporotic fractures. J BoneMiner Res 1998;13:1755–9.
44. Nelson RJ, Demas GE, Huang PL, et al. Behaviouralabnormalities in male mice lacking neuronal nitricoxide synthase. Nature 1995;378:383–6.
45. Bourguet W, Ruff M, Chambon P, Gronemeyer H,Moras D. Crystal structure of the ligand-binding domain
of the human nuclear receptor RXR-a. Nature 1995;375:377–82.
46. Brzozowski AM, Pike ACW, Dauter Z, et al. Molecularbasis of agonism and antagonism in the estrogenreceptor. Nature 1997;389:753–8.
47. Tanenbaum DM, Wang Y, Williams SP, Sigler PB.Crystallographic comparison of the estrogen andprogesterone receptor’s ligand binding domain. ProcNatl Acad Sci U S A 1998;95:5998–6003.
48. Egea PF, Mitschler A, Rochel N, Ruff M, Chambon P,Moras D. Crystal structure of the human RXRa ligand-binding domain bound to its natural ligand: 9-cisretinoic acid. EMBO J 2000;19:2592–601.
49. Wurtz JM, Bourguet W, Renaud JP, et al. A canonicalstructure for the ligand-binding domain of nuclearreceptors. Nat Struct Biol 1996;3:87–94.
50. Egea PF, Klaholz BP, Moras D. Ligand-proteininteractions in nuclear receptors of hormones. FEBSLett 2000;476:62–7.
Research. on July 9, 2014. © 2008 American Association for Cancercancerres.aacrjournals.org Downloaded from




















