Subunits Regulates Olfaction in Caenorhabditis elegans - NCBI
Upload
independentCategory
view
1download
0
The Role of Body Wall Muscles in C. elegans Locomotion
Jordan H. Boyle and Netta CohenSchool of Computing, University of Leeds, Leeds LS2 9JT, UK
Abstract
Over the past four decades, one of the simplest nervous systems across the animal kingdom, that of the nematodeworm C. elegans, has drawn increasing attention. This system is the subject of an intensive concerted effort tounderstand the behaviour of an entire living animal, from the bottom up and the top down. C. elegans locomotion,in particular, has been the subject of a number of models, but there is as yet no general agreement about the key(rhythm generating) elements. In this paper we investigate the role of one component of the locomotion subsystem,namely the body wall muscles, with a focus on the role of inter-muscular gap junctions. We construct a detailedelectrophysiological model which suggests that these muscles function, to a first approximation, as mere actuatorsand have no obvious rhythm generating role. Furthermore, we show that within our model inter-muscular couplingis too weak to have a significant electrical effect. These results rule out muscles as key generators of locomotion,pointing instead to neural activity patterns. More specifically, the results imply that the reduced locomotion velocityobserved in unc-9 mutants is likely to be due to reduced neuronal rather than inter-muscular coupling.
Key words: Caenorhabditis elegans locomotion, body wall muscles, gap junctions, conductance based models
1. Introduction
Over more than a century, huge advances havebeen made in understanding the operation of neu-rons at a cellular and sub-cellular level, as well as inabstractly understanding the operation of large scaleneural networks, such as those found in mammaliancortex. However, despite these impressive advances,we have yet to succeed in fully understanding theneural basis of all but the simplest behaviours. Oneorganism, Caenorhabditis elegans, provides us withthe first tangible possibility of understanding com-plex behaviours of an organism from the genetic andmolecular level, through the cellular level, right upto the system level (and back down).
C. elegans is a small (about 1mm long) nematodeworm [1], which has been the subject of much scien-tific research over the years. It is extensively used as
Email address: [email protected] (Netta Cohen).
a model organism for genetic research, due in partto its short life cycle and fully sequenced genome [2].In addition, it is an attractive subject for researchon the neural basis of behaviour due to an invariantnervous system consisting of a mere 302 neurons inthe adult hermaphrodite. While the connectivity ofits nervous system is known to an unprecedented de-gree of accuracy, far less information is available onthe electrical properties of these neurons. Thus thequestion of the neural basis of C. elegans behaviouris still an open and attractive one.
1.1. C. elegans locomotion
C. elegans movement typically consists of periodsof forward motion interspersed with short periodsof backward motion and turns. Forwards and back-wards locomotion are achieved by propagating sinu-soidal undulations along the body from head to tail(or tail to head) respectively. The worm’s locomo-
Preprint submitted to Elsevier 12 July 2007

tion is controlled by a small, known subset of its ner-vous system [3], and manipulations at the genetic orneuronal level allow insight into its inner workings.The apparent modularity of this subsystem, alongwith its easily observable output, make the locomo-tion nervous system a particularly appealing subjectfor modelling work.
Various models of C. elegans locomotion have al-ready been developed [4–9]. Nonetheless the systemis still only partially understood, and remains of sig-nificant interest to modellers and experimentalists.
One fundamental unresolved question is whetherthe rhythmic motor control of forward locomotionrelies on a central pattern generator (CPG) circuitor on sensory feedback. The conventional wisdom(supported by all evidence to date) is that all rhyth-mic motor behaviours across the animal kingdom areproduced by CPGs, but such a CPG has been elu-sive in the C. elegans locomotion circuit [7,8]. Thus,some models [4,7] propose a mechanism wherebyundulations are activated by stretch receptors thatprovide sensory feedback from body posture, withno intrinsic neuronal oscillator. One possible issuewith regard to any locomotion model in the wormis the relatively sparse modelling work on the bodywall muscles.
In particular, it is not known which componentsof the locomotion subsystem are actively involvedin generating and shaping locomotion. The candi-dates are the interneurons, ventral cord motorneu-rons, body wall muscles and the C. elegans body it-self. Putting aside the possible contribution of thebody (but see [4,5,7,8]), the two alternatives are ei-ther that the patterned activity of the motor neu-rons activates the muscles which then act as actua-tors to deliver the mechanical contractions, or, per-haps more interestingly, that in addition to neuronalactivity, the muscles themselves are capable of gen-erating oscillatory dynamics and/or of propagatingsuch signals down the length of the worm. The for-mer holds in most studied motor systems: the neuralciruit generates a patterned output, and the mus-cles serve as actuators of that output. Interestingly,this does not appear to be the case in Ascaris lum-bricoides [10,4] – a much larger but closely relatednematode whose nervous system is structurally verysimilar to that of C. elegans.
In Ascaris, the body wall muscles are electricallycoupled by gap junctions 1 and appear to form a
1 Gap junctions, or electrical synapses, are non-selectivechannels that form between two adjacent cells, allowing ions
functional syncytium which produces spontaneousmyogenic activity: graded spikes superimposed onslow depolarisations, which propagate indepen-dently of the nervous system [10]. From this per-spective, it may not be surprising if C. elegans mus-cles had a similar pattern generating (or patternmodulating) role in locomotion.
In the absence of a direct answer to this ques-tion, one may turn to behavioural evidence fromlocomotion-defective (or so called uncoordinated)mutant strains of the worm. Particularly instruc-tive are mutations that might disrupt electrical sig-nal flow between muscles. There are two known gapjunction genes in C. elegans namely unc-7 and unc-9, mutations of which result in virtually identicalphenotypes [11] where locomotion is severely im-paired. Both are widely expressed, but only unc-9is expressed in muscles. In Ref. [12] it is shown thatworms treated with unc-9 RNA interference (RNAi)to suppress unc-9 gene expression exhibit substan-tially reduced locomotion velocities, and the authorssuggest that this effect should be attributed specif-ically to the reduction in gap junction coupling be-tween body wall muscles.
In this paper, we rely on electrophysiological datarecorded from body wall muscles in vitro to con-struct a model of individual and coupled musclecells. We then use this model to determine whatpossible active role may be attributed to individ-ual C. elegans body wall muscles and, furthermore,to determine the consistency of such a model withthe observed unc-9 phenotype. More specifically, weattempt to address the following questions: Do themuscles exhibit action potentials? What is their con-tribution to the generation of rhythmic behaviour?And finally, how strong is the inter-muscular cou-pling, and to what extent does it affect locomotion?
1.2. Typical effects of diffusive coupling
Gap junctions are found in a range of organsand cell types in vertebrates as well as invertebratespecies and are very common in excitable tissue(e.g., heart muscle, pancreas, and the nervous sys-tem). Many gap junctions have fixed conductancesand act as resistive elements. The current flowingthrough such a resistor would be proportional tothe voltage drop across it, or the potential differ-ence between the two coupled cells, j and k: Ij,k =
and small molecules to pass between the them.
2

G (Vk − Vj) where G denotes a constant conduc-tance (or inverse resistance). In C. elegans, it ap-pears that the conductance of some gap junctions isitself a function of the potential difference betweenthe coupled cells. Nonetheless, for the purposes ofthis introduction, suffice it to consider whether thecoupling is relatively ‘weak’ or ‘strong’, neglectingany functional dependence of the conductance.
If coupling is sufficiently strong, the coupled ele-ments will fully synchronise. Therefore the more in-teresting cases are those of weak and intermediatecoupling. Weakly coupled limit cycle oscillators havebeen the subject of much theoretical investigation[13–15]. Typically, the frequencies of the oscillatorsare pulled towards each other, and full or partial en-trainment may result, depending on the difference innatural frequency and the strength of the coupling.In certain situations unexpected behaviour can re-sult, such as antiphase oscillation [16], bursting [17]and even quenching [18].
Coupling between non-oscillating elements has re-ceived less attention, but here one would expect twomain effects. First, gap junctions will allow diffusivecurrents to diminish potential differences betweencoupled cells. In addition, the input impedance ofthe cells will be affected altering their frequency re-sponse [19].
2. Methods
The body wall muscles of C. elegans are dividedinto four quadrants, two ventral and two dorsal, eachquadrant consisting of 23 or 24 trapezoidal cells, ar-ranged in two staggered rows. When it locomotes,the worm lies on its side, with the pairs of ventraland dorsal muscle quadrants contracting in unison.Gap junctions couple cells within each quadrant, aswell as between quadrants, on either the ventral ordorsal sides. Within a quadrant, gap junctions arefound between each muscle cell and the two over-lapping cells from the other row [12]. In the workthat follows, we have reduced each muscle quadrantto a chain of identical cells with nearest neighbourcoupling.
Nematode muscles are unusual in that they ex-tend thin, non-contractile processes to the nervecord, where they receive their neuromuscular in-put. Gap junctions are found between these musclearms, coupling cells from the two ventral (or dor-sal) quadrants. Each muscle typically has three tofive arms [20]. We developed a compartmental con-
ductance based model, with one compartment forthe cell body and ten compartments for each mus-cle arm. All active currents are included in the maincompartment, while the arms are modelled as pas-sive cables.
2.1. Electrical properties of the muscle body
We began by developing a conductance basedmodel of the muscle body. The model contains threeactive currents [21,22]: fast and slow potassium cur-rents (IKf and IKs) and a calcium current (ICa)that exhibits inactivation on both fast and slowtimescales, mediated by Ca2+ and voltage respec-tively. We also include a standard leak conductanceIL. A circuit diagram of the complete model isshown in Figure 1. The membrane potential for theith muscle in the chain is therefore given by
CdVi
dt= −ΣIion + Iin + Ii−1,i + Ii+1,i
ΣIion = IKs + IKf + ICa + IL ,
where Iin is the input current from the muscle arms(see Section 2.2) and Ij,k are gap junction currents(see Section 1.2). The membrane currents are givenby
IKs = gKsn(V − VKs)
IKf = gKfp4q(V − VKf )
ICa = gCae2f(1 + (h− 1)αCa)(V − VCa)IL = gL(V − VL) ,
with activation variables e, n, p, and inactivationvariables f , h, q. Gating kinetics are given in termsof a generic variable x
dx
dt=
x∞(V, Vhalf(x), k(x))− x
τx
with a steady state given by
x∞(X, Xhalf, k) =1
1 + exp[Xhalf−X
k
] ,
and the calcium mediated inactivation is given by
h = x∞([Ca2+]i, Cahalf(h), k(h)).
In order to find values for the 29 parameters ofthis model we simulated voltage clamp and currentclamp traces of the model, fitting the parametersto corresponding whole cell recordings of body wallmuscle cells in vitro [22]. We similarly fit the steadystate I-V curves for IK and ICa [21,22]. Parametersfor which values were reported in the literature were
3

Fig. 1. Equivalent circuit diagram of the muscle model showing currents, voltages and parameters of the muscle body (bottomright of figure), muscle arms (with two of ten compartments of one arm shown and remaining arms appearing in parallel) andcoupling. All labels correspond to those in the text.
limited to a range close to the reported values. Fitswere obtained by an evolutionary algorithm (differ-ential evolution [23]). The parameters obtained aregiven in Appendix A. Note that the value of C =30 pF was not evolved, but was taken from [12] andused for compatibility with coupling parameters. Allsimulations were run with a 4th order Runge-Kuttamethod with a time step of 0.1 ms.
2.2. Muscle arms
The muscles arms are modelled as passive cables,each characterised by membrane capacitance (cm),membrane resistance (rm) and longitudinal resis-tance (rl) (see Figure 1). We used N = 10 discretecompartments for each arm, with five arms per mus-cle. The membrane potential of each arm compart-ment evolves according to
cmdvn
dt= Iinn
− Ioutn− Imn for n = 1 : N,
where Iinnis the current flowing into the nth com-
partment, Ioutnis the current flowing out of the nth
compartment and Imn is the current leaking throughthe cell membrane, according to
Imn=
vn − VL
rmfor n = 1 : N.
The current flowing out of compartment n into com-partment n + 1 is
Ioutn=
vn − vn+1
rlfor n = 1 : N − 1
IoutN=
vN − V
rl,
where V is the membrane potential of the musclebody. With the exception of the first compartment,the current that flows out of one compartment mustflow into the next, so
Iinn= −Ioutn−1 for n = 2 : N.
For two coupled muscles (whether on the dorsal orventral side), the first compartment of arms on theright (left) muscle quadrant have
Iin1= INMJ ± IR,L,
where INMJ is the neuromuscular junction (NMJ)input, and IR,L is the inter-quadrant gap junctioncurrent described in Section 2.3. Finally, the totalcurrent flowing into the muscle body is the sum ofcurrents flowing out of each of the five arms
Iin = −ΣIoutN.
Since no data on the electrical properties of thearms could be found, the cable parameters werebased on estimates of the specific capacitance (Cm)and resistance (Rm) of the membrane, and the spe-cific resistance of the cytoplasm (Rl), scaled by thearm dimensions. This is described in more detail,along with a table of the parameters, in AppendixA.
4

2.3. Coupling
The inter-muscular gap junctions are described in[12], in which the intra-quadrant gap junction con-ductance was found to be a function of the poten-tial difference across the junction, while coupling be-tween quadrants was reported to have no voltage de-pendence (and a significantly smaller conductance).The intra-quadrant gap junction current introducedin Section 2.1 is given by
Ij,k = Gss(Vj − Vk)(Vj − Vk),
where
Gss(∆V ) = gintra
[1− Γmin
1 + exp(A(|∆V | − V0))+ Γmin
].
The inter-quadrant coupling from Section 2.2 is sim-ply
IR,L = ginter(v1(R)− v1(L)),
where v1(R) is the potential of the first compartmentof a right muscle arm and v1(L) is the potential ofthe first compartment of a left muscle arm.
2.4. Current stimuli
Inter-quadrant coupling occurs between cells thatshould be coactive (either on the ventral or dorsalside). To investigate whether these gap junctionscould contribute to equalising input to left and rightmuscles, we stimulate the arms of the right musclewith a constant current step and monitor the result-ing potential change in both left and right musclebodies.
Intra-quadrant coupling is different, as it occursbetween cells which would be expected to haveslightly different input. The locomotion waveform isperiodic in time, with a frequency of about 0.5 Hz.It is also periodic in space, with a wavelength of
approximately23
of the body length on agar. There-fore the inputs to adjacent muscles should be phase
shifted by approximately2π2324
=π
8. It has been
shown that the related nematode Ascaris has non-spiking neurons and graded synaptic transmission[24]. We will make the common assumption that C.elegans motorneurons share these properties. Wetherefore expect the current input to each muscleto be some smoothly varying function. The resultsfrom two such functions are shown in Section 3.3.Similar results were obtained for all other wave-forms tested.
2.5. Signal-to-noise ratio
Another potential effect of coupling is on signal-to-noise ratio (SNR). Synaptic vesicle release is astochastic process, so neuromuscular currents arelikely to have a random component. Rather than ex-plicitly modelling vesicle release, we have used anapproximation where input to each muscle consistsof a periodic signal combined with additive whiteGaussian (zero-mean) noise. The method used forestimating the SNR of a signal is described in Ap-pendix B.
3. Results
3.1. Behaviour of the single cell model
Figure 2 shows the experimental voltage- andcurrent-clamp traces [22] along with correspond-ing traces produced by simulation of our modelwith matching inputs. Likewise Figure 3 shows I-Vcurves for IK and ICa (recorded and simulated).The model output is quantitatively very similar tothe experimental data.
In contrast to Ascaris body wall muscles [25], ourmodel muscles do not produce spikes, either spon-taneously or in response to current injection. Evenwhen the fitness function for the parameter optimi-sation was altered to specifically reward oscillatorybehaviour, spiking could not be achieved for realisticvalues of gCa. While the non-spiking nature of themuscles may be surprising, it is consistent with be-haviour of the muscles in vitro [26]. The steady stateI-V curve for an entire model muscle cell is shownin Figure 4. The voltage response diminishes as cur-rent increases, giving the muscles a wider dynamicrange.
3.2. Propagating activity
A key question is whether the combination ofactive, excitatory currents and electrical couplingmight allow regenerative propagation of activitydown the chain of muscles, as reported in Ascaris[10]. Not only does the absence of spikes makethis unlikely, but the coupling is also insufficientlystrong. Experiments with our model suggest thatan increase of four fold in gCa and about six foldin gintra would be required to allow regenerativepropagation (not shown).
5
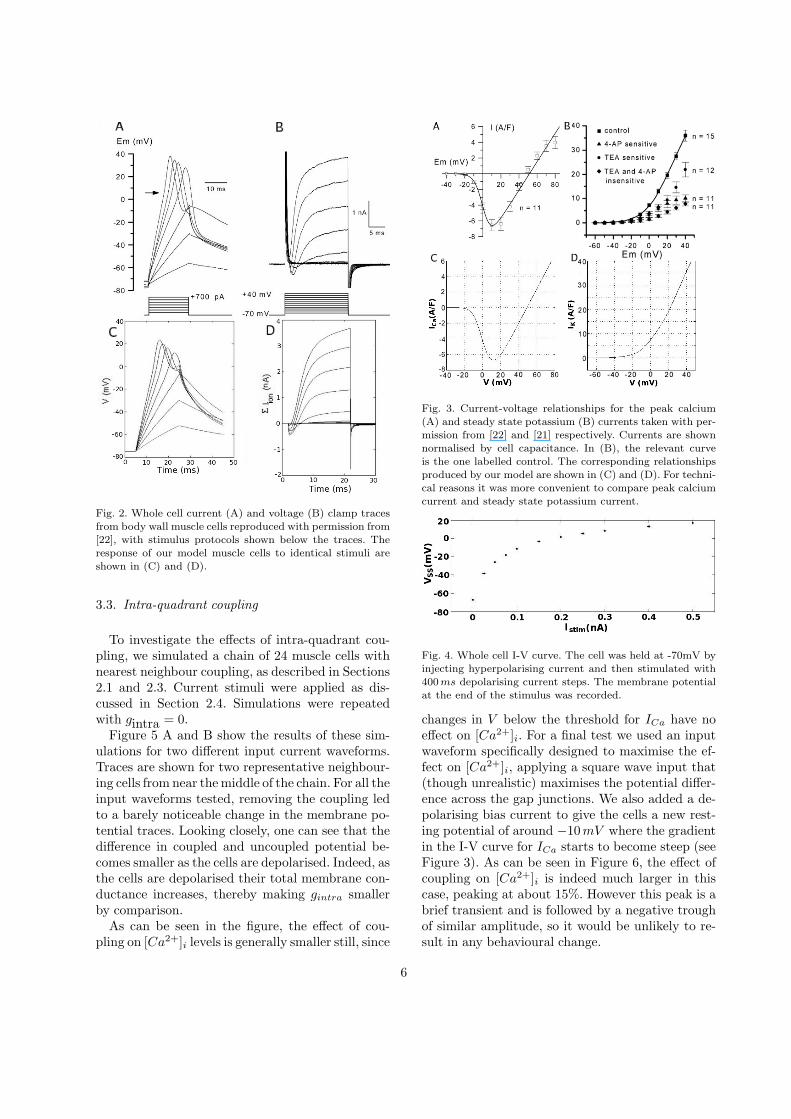
Fig. 2. Whole cell current (A) and voltage (B) clamp tracesfrom body wall muscle cells reproduced with permission from[22], with stimulus protocols shown below the traces. Theresponse of our model muscle cells to identical stimuli areshown in (C) and (D).
3.3. Intra-quadrant coupling
To investigate the effects of intra-quadrant cou-pling, we simulated a chain of 24 muscle cells withnearest neighbour coupling, as described in Sections2.1 and 2.3. Current stimuli were applied as dis-cussed in Section 2.4. Simulations were repeatedwith gintra = 0.
Figure 5 A and B show the results of these sim-ulations for two different input current waveforms.Traces are shown for two representative neighbour-ing cells from near the middle of the chain. For all theinput waveforms tested, removing the coupling ledto a barely noticeable change in the membrane po-tential traces. Looking closely, one can see that thedifference in coupled and uncoupled potential be-comes smaller as the cells are depolarised. Indeed, asthe cells are depolarised their total membrane con-ductance increases, thereby making gintra smallerby comparison.
As can be seen in the figure, the effect of cou-pling on [Ca2+]i levels is generally smaller still, since
Fig. 3. Current-voltage relationships for the peak calcium(A) and steady state potassium (B) currents taken with per-mission from [22] and [21] respectively. Currents are shownnormalised by cell capacitance. In (B), the relevant curveis the one labelled control. The corresponding relationshipsproduced by our model are shown in (C) and (D). For techni-cal reasons it was more convenient to compare peak calciumcurrent and steady state potassium current.
Fig. 4. Whole cell I-V curve. The cell was held at -70mV byinjecting hyperpolarising current and then stimulated with400 ms depolarising current steps. The membrane potentialat the end of the stimulus was recorded.
changes in V below the threshold for ICa have noeffect on [Ca2+]i. For a final test we used an inputwaveform specifically designed to maximise the ef-fect on [Ca2+]i, applying a square wave input that(though unrealistic) maximises the potential differ-ence across the gap junctions. We also added a de-polarising bias current to give the cells a new rest-ing potential of around −10 mV where the gradientin the I-V curve for ICa starts to become steep (seeFigure 3). As can be seen in Figure 6, the effect ofcoupling on [Ca2+]i is indeed much larger in thiscase, peaking at about 15%. However this peak is abrief transient and is followed by a negative troughof similar amplitude, so it would be unlikely to re-sult in any behavioural change.
6
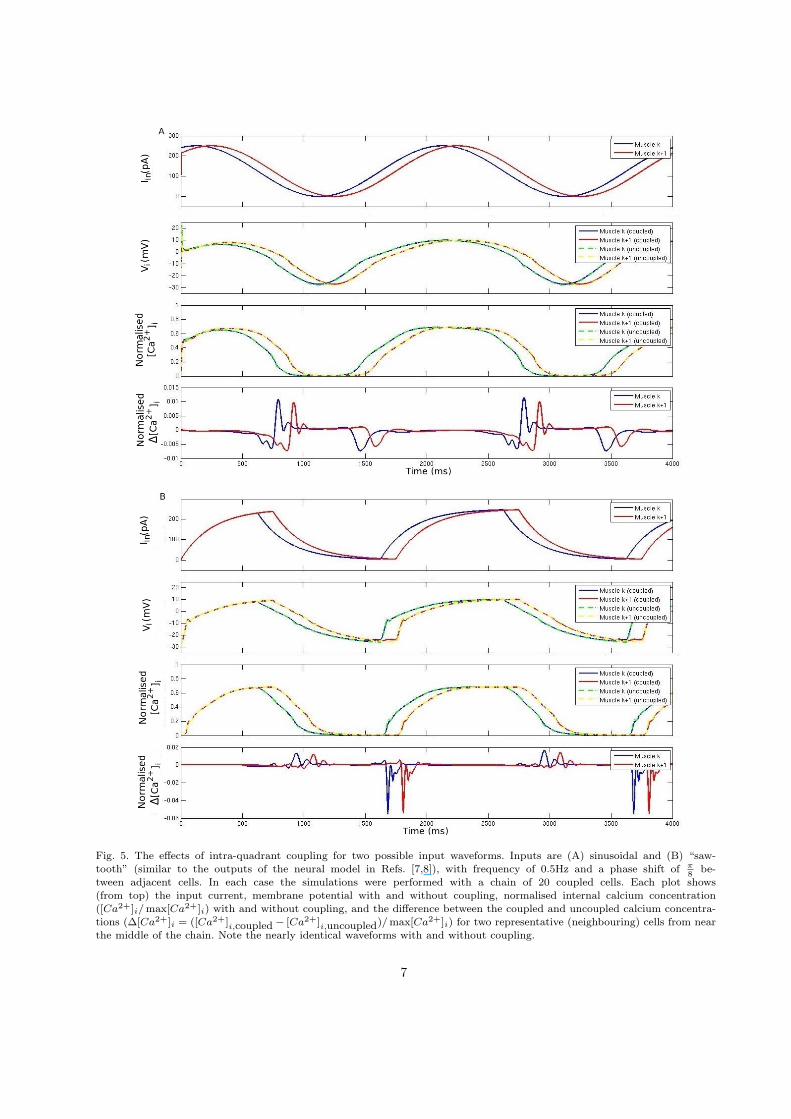
Fig. 5. The effects of intra-quadrant coupling for two possible input waveforms. Inputs are (A) sinusoidal and (B) “saw-tooth” (similar to the outputs of the neural model in Refs. [7,8]), with frequency of 0.5Hz and a phase shift of π
8be-
tween adjacent cells. In each case the simulations were performed with a chain of 20 coupled cells. Each plot shows(from top) the input current, membrane potential with and without coupling, normalised internal calcium concentration([Ca2+]i/ max[Ca2+]i) with and without coupling, and the difference between the coupled and uncoupled calcium concentra-tions (∆[Ca2+]i = ([Ca2+]i,coupled − [Ca2+]i,uncoupled)/ max[Ca2+]i) for two representative (neighbouring) cells from nearthe middle of the chain. Note the nearly identical waveforms with and without coupling.
7
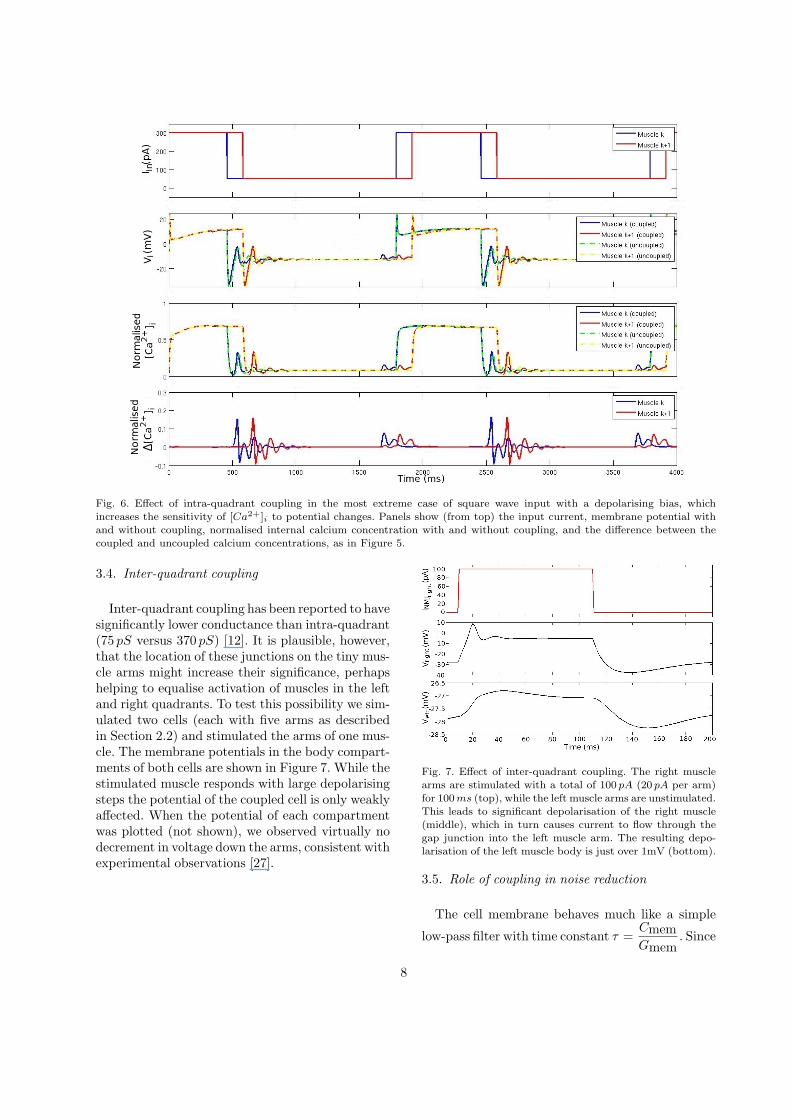
Fig. 6. Effect of intra-quadrant coupling in the most extreme case of square wave input with a depolarising bias, whichincreases the sensitivity of [Ca2+]i to potential changes. Panels show (from top) the input current, membrane potential withand without coupling, normalised internal calcium concentration with and without coupling, and the difference between thecoupled and uncoupled calcium concentrations, as in Figure 5.
3.4. Inter-quadrant coupling
Inter-quadrant coupling has been reported to havesignificantly lower conductance than intra-quadrant(75 pS versus 370 pS) [12]. It is plausible, however,that the location of these junctions on the tiny mus-cle arms might increase their significance, perhapshelping to equalise activation of muscles in the leftand right quadrants. To test this possibility we sim-ulated two cells (each with five arms as describedin Section 2.2) and stimulated the arms of one mus-cle. The membrane potentials in the body compart-ments of both cells are shown in Figure 7. While thestimulated muscle responds with large depolarisingsteps the potential of the coupled cell is only weaklyaffected. When the potential of each compartmentwas plotted (not shown), we observed virtually nodecrement in voltage down the arms, consistent withexperimental observations [27].
Fig. 7. Effect of inter-quadrant coupling. The right musclearms are stimulated with a total of 100 pA (20 pA per arm)for 100 ms (top), while the left muscle arms are unstimulated.This leads to significant depolarisation of the right muscle(middle), which in turn causes current to flow through thegap junction into the left muscle arm. The resulting depo-larisation of the left muscle body is just over 1mV (bottom).
3.5. Role of coupling in noise reduction
The cell membrane behaves much like a simple
low-pass filter with time constant τ =CmemGmem
. Since
8

Gmem is dynamic τ will also be. The propertiesof this filter will also be affected by coupling. Thecurrent stimulus used is a periodic signal with anadditive noise component (as described in Section2.5). Simulations were run both with and withoutcoupling, for various values of noise variance σ. Wethen estimated the signal-to-noise ratio (SNR) ofthe input current and of the resulting V and [Ca2+]iwaveforms (as described in Appendix B).
In the absence of coupling, the low-pass character-istics of the cell leads to a significant improvementin the SNR of the membrane potential over that ofthe input current. The additional low-pass effect of[Ca2+]i leads to an even better SNR in the calciumwaveform. When coupling is added, however, thereis only a very small further improvement in SNR, asillustrated in Figure 8.
Fig. 8. Two stages of low-pass filtering occur in the muscles,having a large effect on SNR. The first stage has its effecton the membrane potential, and the second on the internalcalcium concentration. Coupling results in a small furtherimprovement due to a small change in the effective inputimpedance.
4. Conclusions and discussion
We have presented a first detailed electrophysio-logical model of C. elegans body wall muscles. Weuse this model to test several hypotheses about thepossible role of these muscle cells in the locomotionof the worm. Model parameters were found such thatthe membrane properties of the cell bodies matchexperimental recordings. A possible caveat on thoseparameters is the lack of relevant in vivo data todate. Nonetheless, the model presented is likely toprovide a valid first approximation of muscle be-haviour in a behaving worm.
First, we rule out the possibility of calcium actionpotentials for parameters anywhere within the ball-park of the model we have presented. The fact thatconductances would have to increase many fold toyield any oscillations in the model cells suggests that
such behaviour is unlikely in vivo. This is an inter-esting result and contrasts with reported results forthe related nematode worm, Ascaris lumbricoides.That said, the absence of action potentials in bodywall muscles does not, in itself, preclude a possiblerole of these muscle cells in rhythmic pattern gen-eration. (Indeed, there are no known action poten-tials in the interneurons and motor neurons of thelocomotion system either.)
To determine the possible roles of muscle cells,first, in pattern generation, and second, in the prop-agation of signals along the worm, we present simu-lations of chains of coupled muscle cells (modeling asingle quadrant of body wall muscles in the worm)and pairs of coupled muscle cells (modeling inter-quadrant coupling). In these simulations, the limit-ing factor is the strength of the gap junctional cou-pling. In fact, it appears that the reported conduc-tance values are too low to support effective signaltransmission between adjacent muscle cells. Hence,all of the results presented strongly suggest that infact, C. elegans muscles are most likely to act onlyas actuators, and are not capable of communicatingsignals in a sufficiently effective manner, either toparticipate in pattern generation, or to propagateelectrical oscillations.
It has been suggested that these body wall mus-cle cells also possess so called stretch receptor chan-nels, that depolarise the cell in response to bend-ing or stretching of the body [28]. If so, in a rhyth-mically bending or undulating worm, the musclesmay, in principle, have the capacity to respond tothe alternating body posture actively (but still withgraded potential changes) thus aiding and main-taining this oscillatory behaviour. Such a sensory-feedback mechanism mimics closely existing mod-els of sensory feedback driving neuronal activationin the ventral cord [5,7,8]. However, if this weretrue, such a mechanism would operate effectivelyindependently from any neuronally generated os-cillations, since the only points of contact betweenmuscles and the nervous system (the neuromuscu-lar junctions) only allow information flow in one di-rection – from the neurons to the muscles. Thus,to be interesting (i.e. to contribute significantly topattern generation), muscles distributed along thebody of the worm would need to coordinate theiroscillations. Here, we have demonstrated that theweakness of the gap junctional coupling precludesthe communication of such signals along a chain ofmuscles.
As a direct consequence, the model and simula-
9

tions presented here strongly suggest that the neu-ral circuit is the active component generating therhythmic patterns of locomotion (though we shedno light on the neural mechanism of generating suchpatterns – whether via a central pattern generatingcircuit, or via sensory feedback from stretch recep-tor channels).
Another question is what role the muscles mayhave in shaping locomotion. Figure 5 shows that thewaveform of activity in muscles closely follows thewaveform of the input. This holds true both with andwithout inter-muscular coupling. The muscles there-fore seem to effectively be actuators. The musclesexhibit extended dynamic range due to the concaveI-V relationship shown in Figure 4 and are capableof some (very limited) low-pass filtering. This rela-tively simple interpretation of muscle function hasimportant implications for any neural model. Themuscles’ transfer function suggests that it shouldbe possible to infer neural activity patterns frommuscle output. The extended dynamic range andlow-pass characteristics should, if anything, improverobustness to changes in levels of neural activityand stochasticity of neurotransmitter release respec-tively. In fact, in the absence of muscle output tracesfrom behaving locomoting worms, the developmentof an appropriate physical model of the worm [4,29]would make it possible to infer neural activity pat-terns from behavioural observations.
Finally we have shown that the phenotypes of unc-7 or unc-9 mutation, or of unc-9 RNAi, cannot beexplained by our model in terms of muscle gap junc-tions. One may therefore speculate as to which gapjunctions (elsewhere in the locomotion system) mayaccount for the reduced velocity observed in thesemutant and RNAi treated worms. A likely candi-date is the gap junctional coupling between the for-ward locomotion command interneurons AVB andforward locomotion motor neurons of classes VB andDB. Interestingly, models of the neural control offorward locomotion [7,8] have also suggested thatmodulation of command input into these motor neu-rons can modulate locomotion velocity [8].
Appendix A. Model parameters
A.1. Parameters for the muscle body
Due to the complexity of the fitness landscapefor these parameters, multiple iterations of theevolutionary algorithm produced similar, but non-
identical parameter sets. One such set (the one usedfor the simulations in this paper) is presented inTable A.1.
Param. Value Reported Val. Param. Value Reported Val.
CA 72.3pF ∼ 75pF [22] CB 30pF 29.6pF [12]
gKs 436S/F 399S/F [21] VKs −64.3mV −67.9mV [21]
gKf 400S/F 423S/F [21] VKf −55.0mV −47.0mV [21]
gCa 220S/F 199S/F [22] VCa 49.1mV 50.0mV [22]
gL 19.3S/F 22S/F [12] VL 10.0mV n/a
αCa 0.283 n/a φCa 2.39 · 10−6 n/a
V0.5n 19.9mV n/a kn 15.9mV n/a
V0.5p −8.1mV n/a kp 7.4mV n/a
V0.5q −15.6mV n/a kq −10.0mV n/a
V0.5e −3.4mV n/a ke 6.7mV n/a
V0.5f25.2mV n/a kf 5.0mV n/a
Ca0.5h64.1 · 10−9 n/a kh −10µM n/a
τn 25.0ms n/a τp 2.3ms n/a
τq 150ms n/a τe 0.10ms n/a
τf 151ms n/a τCa 11.6ms n/a
Table A.1Parameters for the muscle body obtained by fits to experi-mental traces (see text for detail). Reported values are givenwhere possible. The two values of CA and CB were used inour simulations for compatibility with voltage- and current-clamp data [21,22] and coupling parameters [12] respectively.
A.2. Muscle arm parameters
We began by estimating values for the specific ca-pacitance (Cm) and resistance (Rm) of the mem-brane, and the specific resistance of the cytoplasm(Rl). Using the standard value of Cm = 1 µF/cm2,a cell with C = 30 pF (as in [12]) should have asurface area of 3 × 10−3 mm2. The same cell wasreported to have Gin = 1/Rin = 666 ps, so Rm =45 × 103 Ω cm2. Finally a standard value of Rl =100 Ω cm was chosen. Approximating each musclearm as a cylinder with l = 10µm and r = 0.75µm(which we divide into ten compartments), we obtain:
cm = Cm2πrl
rm =Rm
2πrl
rl =Rll
πr2.
The resulting parameter values are given in TableA.2.
10

N cm rm rl
10 47 fF 950 GΩ 570 kΩ
Table A.2Muscle arm compartment parameters, obtained from esti-mates of the cell properties and dimensions (see text).
A.3. Coupling parameters
As a conservative choice of gintra, we used thepeak value reported in Ref. [12]. The value of ginteris the reported value of 75 pS divided evenly acrossthe five arms of each muscle cell. The remaining pa-rameters set the voltage dependence of the intra-quadrant coupling and were obtained by curve fit-ting to the experimental G/V curve in [12]. The pa-rameters are given in Table A.2.
gintra ginter Γmin A V0
370 pS 15pS 0.13 40 60 mV
Table A.3Intra- and inter-quadrant coupling parameters. gintra andginter were reported [12]. The remaining parameters wereobtained by curve fitting to Figure 2. B of Ref. [12].
Appendix B. Calculating signal-to-noiseratio
We use spectral methods to calculate the signal-to-noise ratio (SNR) in a model muscle cell, stimu-lated with a superposition of a sinusoidal wave andwhite Gaussian noise. The input signal we use has
a period T =2π
ωin= 2s. Simulations were run for a
total duration of D = 8s with a time step of ∆t =0.01ms.
The SNR is estimated from the power spectrumdensity (PSD) of the muscle output. For a signalx(t), the PSD is given by
PSD(ω) = X(ω)X∗(ω)/N ,
where X(ω) is the Fourier transform of the signal(calculated with a fast Fourier transform or FFT,with N = 219, and the asterisk denotes complexconjugation.
To estimate the SNR from the PSD, we mustfirst specify what frequency range will be considered“signal”. Ideally the input should appear in the fre-quency domain as a spike of zero width at 0.5 Hz.In reality the signals (particularly the “filtered” sig-nals Vi and [Ca2+]i) will be smeared to some extent,
leading to a peak of nonzero width. Based on visualinspection of the PSD, we have assigned all compo-nents on the range 0-5 Hz to the signal, and from5 Hz to 50 kHz to noise. While there will be somenoise in the 0- 5Hz range, this represents only 0.1%of the total bandwidth, and will not significantly af-fect the results. Finally we obtain values for the sig-nal power, Psignal, the noise power Pnoise and theratio SNR as follows
Psignal =1N
Σ5Hz−5HzPSD(ω)
Pnoise =1N
[Σ50kHz
5Hz PSD(ω) + Σ−5Hz−50kHzPSD(ω)
]SNR =
PsignalPnoise
.
The SNR is then calculated for different values ofthe input noise variance.
References
[1] D. L. Riddle, T. Blumenthal, B. J. Meyer, J. R. Priess(Eds.), C. elegans II, 2nd Edition, Cold Spring HarbourLaboratory Press, New York, 1997.
[2] C. elegans Sequencing Consortium, Genome sequence ofthe nematode C. elegans: A platform for investigatingbiology, Science 282 (1998) 2012–2018.
[3] J. G. White, E. Southgate, J. N. Thomson, S. Brenner,The structure of the nervous system of the nematodeCaenorhabditis elegans, Philosophical Transactions ofthe Royal Society of London, Series B 314 (1986) 1–340.
[4] P. Erdos, E. Niebur, The neural basis of the locomotionof nematodes, Lecture Notes in Physics 368 (1990) 253–267.
[5] E. Niebur, P. Erdos, Theory of the locomotion ofnematodes: Control of the somatic motor neurons byinterneurons, Mathematical Biosciences 118 (1993) 51–82.
[6] J. A. Bryden, A simulation model of the locomotioncontrollers for the nematode Caenorhabditis elegans,Master’s thesis, University of Leeds (2003).
[7] J. A. Bryden, N. Cohen, A simulation model of thelocomotion controllers for the nematode Caenorhabditiselegans., in: S. Schaal, A. J. Ijspeert, A. Billard,S. Vijayakumar, J. Hallam, J. A. Meyer (Eds.),Proceedings of the eighth international conference on thesimulation of adaptive behaviour, MIT Press / BradfordBooks, 2004, pp. 183–192.
[8] J. A. Bryden, N. Cohen, Neural control of C. elegansforward locomotion: The role of sensory feedback,submitted.
[9] M. Suzuki, T. Tsuji, H. Ohtake, A model of motorcontrol of the nematode C. elegans with neuronalcircuits, Artificial Intelligence in Medicine 35 (2005) 75–86.
[10] D. A. Weisblat, R. L. Russell, Propagation of electricalactivity in the nerve cord and muscle syncytium of the
11

nematode Ascaris lumbricoides, Journal of comparativePhysiology 107 (1976) 293–307.
[11] P. Phelan, T. A. Starich, Innexins get into the gap,BioEssays 23 (2001) 388–396.
[12] Q. Liu, B. Chen, E. Gaier, J. Joshi, Z. W. Wang, Lowconductance gap junctions mediate specific electricalcoupling in body-wall muscle cells of Caenorhabditiselegans, Journal of Biological Chemistry 281 (2006)7881–7889.
[13] G. B. Ermentrout, n:m phase-locking of weakly coupledoscillators, Journal of Mathematical Biology 12 (1981)327–342.
[14] B. D. Ermentrout, N. Kopell, Frequency plateaus in achain of weakly coupled oscillators, I., SIAM Journal ofMathematical Analysis 15 (1984) 215–237.
[15] N. Kopell, G. B. Ermentrout, Symmetry andphaselocking in chains of weakly coupled oscillator,Communications on Pure and Applied Mathematics 39(1986) 623–660.
[16] A. Sherman, J. Rinzel, Rhythmogenic effects of weakelectrotonic coupling in neuronal models, Neurobiology89 (1992) 2471–2474.
[17] A. K. Han, C. Kurrer, Y. Kuramoto, Dephasing andbursting in coupled neural oscillators, Physical ReviewLetters 75 (1995) 3190–3193.
[18] D. Ramana Reddy, A. Sen, G. L. Johnston, Time delayinduced death in coupled limit cycle oscillators, PhysicalReview Letters 80 (1998) 5109–5112.
[19] N. Cohen, The development of spontaneous beatingactivity in cultured heart cells: from cells to networks,Ph.D. thesis, Israel Institute of Technology, Kislev, 5761,Haifa (December 2000).
[20] S. J. Dixon, P. J. Roy, Muscle arm development inCaenorhabditis elegans, Development 132 (2005) 3079–3092.
[21] M. Jospin, M. C. Mariol, L. Segalat, B. Allard,Characterisation of K+ currents using an in situpatch clamp technique in body wall muscle cells fromCaenorhabditis elegans, Journal of Physiology 544.2(2002) 373–384.
[22] M. Jospin, V. Jacquemond, M. C. Mariol, L. Segalat,B. Allard, The l-type voltage-dependent Ca2+ channelEGL-19 controls body wall muscle function inCaenorhabditis elegans, Journal of Cell Biology 159(2002) 337–347.
[23] K. Price, R. Storn, Differential evolution, Dr. Dobb’sJournal April (1997) 18–24.
[24] R. E. Davis, A. O. W. Stretton, Signalling properties ofAscaris motorneurons: Graded active responses, gradedsynaptic transmission, and tonic transmitter release,Journal of Neuroscience 9 (1989) 415–425.
[25] D. A. Weisblat, L. Byerly, R. L. Russel, Ionicmechanisms of electrical activity in somatic muscle of thenematode Ascaris lumbricoides, Journal of comparativePhysiology 111 (1976) 93–113.
[26] B. Allard, Personal Communication (2006).[27] J. E. Richmond, E. M. Jorgensen, One GABA and
two acetylcholine receptors function at the C. elegansneuromuscular junction, Nature neuroscience 2 (1999)791–797.
[28] J. Liu, B. Schrank, R. H. Waterston, Interaction betweena putative mechanosensory membrane channel and acollagen, Science 273 (1996) 361–364.
[29] J. H. Boyle, J. Bryden, N. Cohen, An integrated neuro-mechanical model of C. elegans forward locomotion,submitted.
12
Copyright © 2022 FDOKUMEN