
Trunk movements in human locomotion
14
Acta Physiol Scand 1984, 121: 9-22 Trunk movements in human locomotion ALF THORSTENSSON, JOHNNY NILSSON, HANS CARLSON and MICHAEL R. ZOMLEFER' Department of Physiology 111, Karolinska Institutet, Lidingoviigen 1, S-114 33 Stockholm, Sweden THORSTENSSON, A., NILSSON, J., CARLSON, H. & ZOMLEFER, M. R.: Trunk movements in human locomotion. Acta Physiol Scand 1984, 121:9-22. Received 21 Sept. Accepted 14 Nov. 1983. ISSN 0001-6772. Department of Physiology 111, Karolinska institutet, Stockholm, Sweden. Trunk movements in the frontal and sagittal planes were studied in 10 healthy males (18-35 yrs) during normal walking (1.0-2.5 ds) and running (2.0-6.0 m/s) on a treadmill. Move- ments were recorded with a Selspot optoelectronic system. Directions, amplitudes and phase relationships to the stride cycle (defined by the leg movements) were analyzed for both linear and angular displacements. During one stride cycle the trunk displayed two oscillations in the vertical (mean net amplitude 2.5-9.5 cm) and horizontal, forward-back- ward directions (mean net amplitude 0.5-3 cm) and one oscillation in the lateral, side to side direction (mean net amplitude 2-6 cm). The magnitude and timing of the various oscillations varied in a different way with speed and mode of progression. Differences in amplitudes and timing of the movements at separate levels along the spine gave rise to angular oscillations with a similar periodicity as the linear displacements in both planes studied. The net angular trunk tilting in the frontal plane increased with speed from 3-10". The net forward-backward trunk inclination showed a small increase with speed up to 5" in fast running. The mean forward inclination of the trunk increased from 6" to about 13" with speed. Peak inclination to one side occurred during the support phase of the leg on the same side. Peak forward inclination was reached at the initiation of the support phase in walking, whereas in running the peak inclination was in the opposite direction at this point. The adaptations of trunk movements to speed and mode of progression could be related to changing mechanical conditions and different demands on equilibrium control due to e.g. changes in support phase duration and leg movements. Key words: Human locomotion, kinematics, trunk movements, speed adaptations, balance control Compared to the considerable amount of informa- tion available on the human limb movements the knowledge about the control of the trunk is scarce (Grillner 1981, Thorstensson et al. 1982). In loco- motion the legs execute the primary propulsive movements, whereas the trunk motor system as- sists in the maintenance of body equilibrium and interacts with the limb movements to achieve eE- cient locomotion. The trunk represents more than 50% of the body mass and a detailed knowledge of its movements is a fust step towards understanding the physiological control system. Some aspects of trunk movements during loco- motion have been studied earlier (Braune & Fischer 1895, Fischer 1899-1904, Cavagna et al. 1963, 1964, Murray et al. 1964, 1966, Gregersen & Lucas 1967, Lamoreux 1971, Waters et al. 1973, Cappozzo et al. 1978, Inman et al. 1981, Cappozzo 1981, 1982, 1983). These studies have mainly been limited to walking over a narrow range of speeds and in run- ning to energetic considerations based on the dis- placement of the centre of gravity (Cavagna 1964, Ito et al. 1983). The changes occurring in the transi- tion between the two modes of human progression have not been described in detail in the literature. In recent studies of the control of the trunk dur- ing locomotion (Carlson et al. 1979, Carlson & Thorstensson 1981, Thorstensson et al. 1982) inter- est was focused on the activation pattern of lumbar back muscles in relation to the overall trunk move- ' Present address for M. R. Zomlefer: Rehabilitative Re- search and Development Center, Veterans Administration Hospital, Palo Alto, CA 94304, USA. Acta Physiol Scand 121
Transcript of Trunk movements in human locomotion

Acta Physiol Scand 1984, 121: 9-22
Trunk movements in human locomotion
ALF THORSTENSSON, JOHNNY NILSSON, HANS CARLSON and MICHAEL R. ZOMLEFER' Department of Physiology 111, Karolinska Institutet, Lidingoviigen 1, S-114 33 Stockholm, Sweden
THORSTENSSON, A., NILSSON, J., CARLSON, H. & ZOMLEFER, M. R.: Trunk movements in human locomotion. Acta Physiol Scand 1984, 121:9-22. Received 21 Sept. Accepted 14 Nov. 1983. ISSN 0001-6772. Department of Physiology 111, Karolinska institutet, Stockholm, Sweden.
Trunk movements in the frontal and sagittal planes were studied in 10 healthy males (18-35 yrs) during normal walking (1.0-2.5 d s ) and running (2.0-6.0 m/s) on a treadmill. Move- ments were recorded with a Selspot optoelectronic system. Directions, amplitudes and phase relationships to the stride cycle (defined by the leg movements) were analyzed for both linear and angular displacements. During one stride cycle the trunk displayed two oscillations in the vertical (mean net amplitude 2.5-9.5 cm) and horizontal, forward-back- ward directions (mean net amplitude 0.5-3 cm) and one oscillation in the lateral, side to side direction (mean net amplitude 2-6 cm). The magnitude and timing of the various oscillations varied in a different way with speed and mode of progression. Differences in amplitudes and timing of the movements at separate levels along the spine gave rise to angular oscillations with a similar periodicity as the linear displacements in both planes studied. The net angular trunk tilting in the frontal plane increased with speed from 3-10". The net forward-backward trunk inclination showed a small increase with speed up to 5" in fast running. The mean forward inclination of the trunk increased from 6" to about 13" with speed. Peak inclination to one side occurred during the support phase of the leg on the same side. Peak forward inclination was reached at the initiation of the support phase in walking, whereas in running the peak inclination was in the opposite direction at this point. The adaptations of trunk movements to speed and mode of progression could be related to changing mechanical conditions and different demands on equilibrium control due to e.g. changes in support phase duration and leg movements.
Key words: Human locomotion, kinematics, trunk movements, speed adaptations, balance control
Compared to the considerable amount of informa- tion available on the human limb movements the knowledge about the control of the trunk is scarce (Grillner 1981, Thorstensson et al. 1982). In loco- motion the legs execute the primary propulsive movements, whereas the trunk motor system as- sists in the maintenance of body equilibrium and interacts with the limb movements to achieve eE- cient locomotion. The trunk represents more than 50% of the body mass and a detailed knowledge of its movements is a fust step towards understanding the physiological control system.
Some aspects of trunk movements during loco- motion have been studied earlier (Braune & Fischer 1895, Fischer 1899-1904, Cavagna et al. 1963, 1964, Murray et al. 1964, 1966, Gregersen & Lucas 1967, Lamoreux 1971, Waters et al. 1973, Cappozzo et al.
1978, Inman et al. 1981, Cappozzo 1981, 1982, 1983). These studies have mainly been limited to walking over a narrow range of speeds and in run- ning to energetic considerations based on the dis- placement of the centre of gravity (Cavagna 1964, Ito et al. 1983). The changes occurring in the transi- tion between the two modes of human progression have not been described in detail in the literature. In recent studies of the control of the trunk dur-
ing locomotion (Carlson et al. 1979, Carlson & Thorstensson 1981, Thorstensson et al. 1982) inter- est was focused on the activation pattern of lumbar back muscles in relation to the overall trunk move-
' Present address for M. R. Zomlefer: Rehabilitative Re- search and Development Center, Veterans Administration Hospital, Palo Alto, CA 94304, USA.
Acta Physiol Scand 121

10 A . Thorstensson et al.
ments and on changes observed in the transition from walking t o running. In this paper we expand on the kinematical analysis. Data are presented on linear and angular displacements of the trunk in the frontal and sagittal planes during walking and run- ning on a treadmill over a wide velocity range. First a general description of the trunk movements in locomotion is given and then a detailed quantitative analysis of the speed adaptations in movement am- plitudes, angular excursions and phase relation- ships t o the stride cycle (defined by leg move- ments) is presented. Finally, an attempt is made to give a coherent functional view of the trunk move- ments in locomotion.
MATERIAL AND METHODS Subjects and procedure Ten males volunteered as subjects in the study. Their ages, heights and weights ranged 18-35 yrs (mean: 26 yrs), 1.79-1.89 m (mean: I .83 m) and 66-80 kg (mean: 73 kg), respectively. They had no history of back pain or any other disorders related to the locomotor apparatus. Dur- ing the experiments the subjects wore shorts and conven- tional sport shoes. They performed level walking and running on a large motor-driven treadmill (size of rubber belt: 2.7X0.8 m). The range of speeds investigated was 1.0-2.5 mis (1.0, 1.5, 2.0, 2.5 mls) for walking and 2 .M.O m / s (2.0, 2.5, 3.0, 4.0, 5.0, 6.0 d s ) for running. Several sessions were allowed before the actual recordings started in order to familiarize the subjects with the experimental procedure. Recordings were made during 15-30 s of “steady state” locomotion at each speed. A rest period of about I min was given between each recording session.
Movement recordings Selspot system. The movements of the trunk and left leg were recorded with an optoelectronic system (Selspot I, Selcom AB, Partille, Sweden). This system utilizes in- frared light emitting diodes (LEDs) as markers and the position of each LED is detected by a special detector surface in the focal plane of a camera. The horizontal and vertical resolution is 10 bits. The LEDs are pulsed se- quentially with a 50 ys pulse every 100 ps. The scan rate was set to 156 Hz (Halbertsma 1983). Two cameras, placed perpendicular to each other, were used, one from the rear (movements in the frontal plane) and one from the left side (movements in the sagittal plane). Both cameras were at a distance of 4.5 m from the subject (field of view: approximately 2x2 m). At this distance a certain position of a diode could be determined with a maximal error of +3 mm in both the horizontal and vertical direction. Meas- ures were taken to avoid reflections and to reduce inter- ference from other sources of infrared radiation (Hal- bertsma 1983).
LED positions. LEDs were glued onto the skin over the following anatomical landmarks: (1) ankle joint (left later- al malleolus), (2) knee joint meneath the centre of the lateral epicondylis of the left femur), (3) hip joint (on the
Acta Physiol Scand I21
superior part of the left trochanter major) and along the spine at L3 level (4), at C7 level (6), and at a level corresponding to 50% of the distance between L3 and C7 (diode 5) . LEDs 4-6 were recorded with the rear camera (frontal plane) and 1-3 from the side (sagittal plane). In addition, recordings in the sagittal plane were made from LEDs at L3 (7) and C7 level (8). To minimize obstruction by the arms and shoulders during locomotion LEDs 7 and 8 were placed on stiff light weight metal pins protruding approximately 7 cm from the spine. The pins were at- tached to sockets glued to the skin and further stabilized by rubber belts. The relative movements between mark- ers on the skin and the spine have been shown to be small (<2 mm) in extreme trunk positions (Thorstensson et al. 1982). However, during dynamic conditions, as in loco- motion, it cannot be excluded that there is a larger relative movement between markers and the underlying skeletal structures. This may affect the accuracy of the movement recordings. In walking the subjects were instructed to keep their arms semitlexed in order to reduce obstruction of the LEDs at the hip and L3 level.
Data processing. The analogue position signals were converted to digital form, fed on line to a computer (HP 21 17F) and stored on magnetic tape. Linear interpolation was used to substitute for missing data due to occasional obstruction of an LED. Corrections were made for lens and electronic distortion and parallax errors. After correc- tions the position data were smoothed with filters (33 point finite impulse response filters) with a pass band from 0 to 5 Hz (stop band 15 to 78 Hz, Halbertsma 1983). Net amplitudes and timing of different phases of joint angles and displacements were determined interactively with the aid of the graphic display. The angles chosen to represent the angular movements of the trunk and knee were calculated as shown in Fig. 1. The calculations of net linear and angular excursions and timing of different events were performed on averaged stride cycles obtained from 5-8 consecutive strides for each individual. The duration of a stride cycle (Tc) was defined as the time from touch down of the left foot (indicated by the start of the flexion phase of the left knee during support, cf. Thorstensson et al. 1982) to the next touch down of the same foot. Generally, timing values were obtained after normalization of the stride cycles and expressed in per cent of Tc.
Conventional statistical methods were used to calculate means and standard deviations (SD). Differences were tested for statistical significance with the Student’s t-test.
RESULTS
The movements of the trunk will first be briefly described in relation to the different phases of the stride cycle in walking and running. Then a quanti- tative analysis is made of changes in net linear and angular displacements and in timing of different events with speed and mode of locomotion. Finally, a more coherent analysis is performed of the trunk movements in space a t different phases of a loco- motor stride cycle in walking and running.

Trunk movements in human locomotion 1 1
A. WALK 1.5 rn/s RUN 6.0 rn/s
4 I 4 0 4 4 ITD
] .06 m
3.06 m
1.04 m
3.06 m
] .06 m
] 1 4 O
1 50
] 0 3 m
] 03m
1 4* f K 1 400
Bm) EXT
FLEX
REAR VIEW
SIDE VIEW
$7 ANGLE
Is c__
Fig. 1. Linear and angular displacements of the trunk during consecutive stride cycles in walking (1.5 m/s) and running (6.0 d s ) for one subject. Upper part (A) of the figure shows vertical and lateral movements, lower part (B) movements in the sagittal plane. For definitions of levels and angles, see insets to the right. Ipsilateral (left) support phases following ipsilateral touch down (iTD), are indicated by shaded areas; contrala- teral support phases following contralateral touch down (coTD), are marked by hori- zontal bars in the middle of the figure. In this experiment diodes were placed also on the right leg (cf. Methods) and touch down and toe off were determined as for the left leg, i.e. from the angular displacements of the knee ankle joints (cf. Nilsson et al. 1983).
I. General description In Fig. 1 typical displacement curves are shown for a few consecutive strides in walking and running. Fig. 1A shows the vertical movements of C7 and L3 (records 1-2) as well as the linear (records 3-5) and angular displacements (records 6-7) of the trunk from side to side (see inset for definitions). In Fig. 1 B the corresponding sagittal displacements are given for C7 and L3 and Ts, i.e. trunk inclina- tion (see inset). The lowermost record shows the concomitant angular excursions at the knee joint. The figure should serve as a general guide for the more detailed quantitative analysis that follows. To facilitate a comparison of the trunk movements and the different stride cycle phases, the support phases have been indicated: shaded area=ipsilateral (left)
and horizontal bar=contralateral (right) support phase. From this it is possible to define periods of double and single support in walking and periods both with and without ground contact in running.
I: 1. Vertical movements (Fig. 1 A, records 1-2) In walking the linear displacement of both L3 and C7 showed two full vertical oscillations per stride cycle. The lowest point was reached just after touch down, i.e. in approximately the middle of the double support phase, whereas the highest position occurred in the middle of the single support phase of each leg. During running the same general pat- tern was seen. However, there was a marked phase shift in relation to the stride cycle. In contrast to walking, a considerable downward movement oc-
Acta Physiol Scand 121

12 A . Thorstensson et al.
Table I . Mean values in seconds (fl SD and range) for stride cycle durations (Tc) at different velocities of walking and running (n=IO)
Walk Run
Velocity, rnis . . . 1.0 1.5 2.0 2.5 2.0 2.5 3.0 4.0 5.0 6.0
Tc, s + I SD Range
1.21 1.02 0.91 0.79 0.78 0.76 0.73 0.69 0.65 0.59 0.083 0.056 0.042 0.038 0.044 0.040 0.033 0.023 0.028 0.024
1.11- 0.93- 0.85- 0.73- 0.73- 0.71- 0.69- 0.66- 0.62- 0.56- 1.36 1.12 0.97 0.84 0.86 0.84 0.80 0.74 0.69 0.63
curred initially during the first half of the support period. The highest position coincided with the short phase in the air.
Linear displacements. In walking both L3 and C7 levels of the trunk were moving forward at touch down of each foot. This movement continued dur-
I: 2 . Lateral movements (Fig. 1 A, records 3-7) Linear displacements. In walking the linear dis- placement at all levels of the trunk showed one full oscillation from one side to the other and back during a stride cycle. At touch down of one foot, the trunk was moving towards the same side and reached an extreme position during the period of single support. In running the same general pattern was present. However, there was some phase shift, particularly at higher running velocities (see be- low). Thus, the extreme position of L3 to one side was reached late in the support phase of the corre- sponding limb or during the airborne phase.
Angular displacements. In walking also the inter- nal bending (BF) and the overall tilting (TF) of the trunk from side to side showed one full oscillation during a stride cycle (Fig. l A , records 6-7). At touch down of one foot both angular displacements were towards the same side, but the extreme posi- tion of BF was reached during double support and of TF somewhat later in the step cycle. During running the extreme position of BF to the side of touch down generally was reached before the foot was placed on the ground. The corresponding posi- tion for TF occurred early in the support phase. Thus, a phase difference between BF and TF was also observed in running.
I: 3. Forward-backward movements (Fig. 1 B, re- cords 1-3) The fore-aft movements of the trunk demonstrated a pattern of two oscillations per stride cycle. Occa- sionally, this pattern was somewhat irregular (asymmetric) and the oscillations hard to define due to the very small amplitudes, particularly at the C7 level.
ing the double support phase and attained the ex- treme forward position at about the transition from double to single support. Peak backward displace- ment was reached at the end of single support. In running the trunk movements at the L3 level in the sagittal plane showed a gradual phase shift with speed in relation to the stride cycle. At slow speeds touch down occurred when L3 was close to its most forward position, whereas at higher speeds (e.g. 6.0 m/s, Fig. 1B) L3 was moving backwards at touch down. At 6.0 m / s , peak backward displacement was present in the middle of the support phase and peak forward position during the airborne phase.
Angular displacements. At touch down in walk- ing the trunk inclination (Ts) was close to or had just reached its most forward position (Fig. IB). During double support the trunk was gradually tilt- ing in the backward direction until the beginning of single support. In running, on the other hand, touch down occurred when the trunk was in the most backward tilted position or had just started to move forwards (note that the trunk is not actually tilting backwards due to an increased mean forward incli- nation, cf. below and Fig. 7). Peak forward inclina- tion occurred in the middle of the support phase.
11. Adaptation of movement amplitudes and timing of different events to speed and made of locomotion 11: 1 . Stride cycle duration (Table 1) The recordings of walking and running were carried out over a wide velocity range. To facilitate a com- parison between different strides, speeds and sub- jects, the timing data will be given in a normalized form, i.e. the time between touch downs of the left foot is set to 100%. The absolute mean stride cycle durations are given in Table 1. As shown also earli-
Acta Physiol Scand I21

Trunk movements in human locomotion 13
.I0
.08
.06
04
.02
er by e.g. Grillner et al. (1979) the stride cycle duration decreases curvo-linearly with speed, i.e. a larger relative decrease at low speeds (35% de- crease from 1 to 2.5 m/s in walking) than at high speeds (about 25% decrease from 2 to 6 m/s in running). Over the whole range of speeds there was a 50% reduction of Tc.
-
-
-
-
-
11: 2. Vertical movements (Fig. 2 ) The net vertical excursions of both C7 and L3 in walking showed a continuous increase with increas- ing speed from 2.7 cm to 6.8 crn (L3, Fig. 2A). Three subjects tested also at 3 d s showed a de- crease in net amplitude at this velocity. An increase in net vertical displacement occurred in running (for L3 from 7.7 crn at 2 rn/s to 9.5 cm at 2.5 d s ) (Fig. 2 A), but at higher speeds it gradually declined again (to 4.8 cm at 6.0 d s ) . The maximum dis- placements occurred between 2.5-3 d s (in one case at 4.0 d s ) . Changing from walking to running at the same velocity (2-2.5 d s ) caused a signifcant increase in vertical displacement (from 6.0-6.8 cm to 7.7-9.5 cm for L3) (Fig. 2A). In general there were no or only small differences in net vertical displacement between L3 and C7 levels, but in running at 2.5-3 m / s all subjects showed lower val- ues at C7 than at L3-level.
As mentioned above there was a marked differ- ence in timing of the vertical trunk movements in relation to the stride cycle (leg movements) in walk- ing and running (cf. Fig. 1 A). In Fig. 2 B one in- stant of peak vertical displacement at the L3 level (downwards, cf. Fig. 1 A, upper 2 records) is shown. In walking the lowest position occurred just after touch down (7-10% of Tc), whereas in run- ning the corresponding position occurred later (15-19% of Tc after touch down). The timing of vertical displacement peaks was not a e c t e d by a change in velocity. No systematic difference could be detected between the L3 and C7 levels. There were very small differences between individuals.
11: 3. Lateral movements (Fig. 3 and 4) Linear displacements. Net lateral movement ampli- tudes (from side to side) at C7, 50% and L3 levels of the trunk are shown versus velocity in Fig. 3 A. In walking there was an initial decline in net ampli- tude at all levels from 1.0 to 2.0 d s . The largest decrease was seen at the 50% level (from 5.8 to 3.2 cm). During running there was a difference in speed adaptation between levels. At the C7 level there
A. VE RT I C A L D I S P L A C E M E N T, L 3, (PEAK TO PEAK), m r T
0 WALK 0 RUN
I - 20
I 2 3 4 5 6
VELOCITY, m/s
Fig. 2. (A) Net vertical displacement versus velocity for walking and running. In Figs. 2-7 values are given as means k 1 SD (vertical bars). (B) Time for first downward peak in vertical displacement in per cent of cycle duration (%Tc) at different velocities. In Fig. 2-6 positive numbers on the ordinate represent peaks occurring after and nega- tive numbers before ipsilateral (left) touch down (iTD); iTD is indicated by a solid horizontal line (at 0% of Tc) and contralateral touch down (coTD) by a broken line (at 50% of Tc).
was no change, whereas both the 50% and the L3 level showed an increase in net amplitude with speed. The interindividual differences were largest at high speeds both in walking and running. The net side to side displacement at the C7 level was similar
Acta Physiol Scand 121

14 A . Thorstensson et al.
A.
LATERAL DISPLACEMENT (PEAK TO PEAK), m
LLL 1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
B PEAK LATERAL DISPLACEMENT TO THE LEFT, % OF CYCLE DURATION
c7 0 WALK
RUN
40 t
I......; 1 2 3 4 5 6 1 2 3 4 5 6
VELOCITY, m k
Fig. 3. (A) Net lateral linear displacement and (B) time for left peak lateral displace- ment in percent of cycle duration (Tc) at levels C7, 50% and L3 versus velocity (cf. Fig. 1).
(about 4 cm) when the subject walked or ran at the same speed. There were no significant differences at the 50% or L3 levels either, even if there was a tendency towards smaller values at the L3 level in running. L3 showed the smallest lateral move- ments, except at the highest velocities of walking (2.5 d s ) and running (5-6 d s ) , where C7 displayed the smallest values (Fig. 3A). The largest lateral excursions were seen at the 50% level in fast run- ning (5-6 d s ) and in slow walking (1.0 d s ) (Fig. 3A).
Fig. 3B shows the timing of peaks in lateral displacements to the left at different velocities (ex- pressed in per cent of stride cycle duration, Tc). During walking a peak in lateral displacement of all three levels was seen at a time corresponding to 18-38% of Tc after touch down of the correspond- ing foot. The peaks tended to occur earlier in the stride cycle with increasing velocity. The interindi- vidual variation was higher at higher walking speeds. No significant phase differences occurred with increased running velocity at the C7 and 50% levels (2242% of Tc, Fig. 3B). At the L3 level, however, there was a marked phase shift at running
Acta Physiol Scand 121
velocities above 2.5 d s (Fig. 3B). The extreme position to one side actually occurred 3-13 % of Tc after touch down of the contralateral foot. This means that at touch down L3 was moving in the opposite direction to the upper part of the trunk at high running velocities. There were no differences in timing of lateral displacement peaks between walking and running at the same speeds, except for the L3 level at 2.5 d s , where the peaks occurred later in the stride cycle in running. The interindivi- dual variation was smal! for the C7 and 50% levels, especially in running.
Angular displacements (TF and BF) (Fig. 4). Net amplitudes. In walking the net side to side tilting of the trunk, TF, (cf. Fig. 1) was small, but it in- creased with velocity from about 2" to 5" (Fig. 4 A). The angular displacement of the "intrinsic" trunk angle in the frontal plane, BF (cf. Fig. l), showed a higher net amplitude than TF, approximately 7-9", but it remained unchanged with velocity (Fig. 4 A). In running both TF and BF increased markedly with velocity (from 5 to 11" and from 1 1 to 20", respectively, Fig. 4 A). The interindividual differ- ences increased with speed for both angles. Com-

Trunk movements in human locomotion 15
0 WAU. bMJN
TF
- - - - - - - -. - - - - - - - - - - -.
40 a: h - : L A + !
20 .
0
-20 . T . . . ' ' . " ' ' . . *
h LATERAL DISPLACEMENT (PEAK TO PEAK), deprws
TF 0 WALK
MJN
BF
. - - - - - - - - -. - - - - - - - - - - - - - - - - -.
------7
EF
I . . . . 1 2 3 4 5 6 1 2 3 4 5 6
R
paring walking and running at the same velocity, there were significantly larger BF excursions in running, whereas the TF angle showed similar val- ues (Fig. 4A).
There were differences in timing of displacement peaks in relation to the stride cycle between intrin- sic bending (BF) and overall tilting (TF) of the trunk (Fig. 4B). In walking peak angular displace- ment of BF to one side occurred 7-9% of Tc after touch down of the foot on the same side (Fig. 4 B). The corresponding peaks in TF were seen later in the stride cycle (at about 35 % of Tc) (Fig. 4 B). The difference between subjects was large for TF at low speeds. This was partly due to difficulties in defin- ing the peak because of the flat shape of the dis- placement curves in some individuals. The same relation between the timing of TF and BF peaks
was also observed in running. However, the TF peaks occurred at a time corresponding to 1626% of Tc, whereas the BF peak to the side of foot contact occurred 5-10% of Tc before touch down (Fig. 4B). Differences were seen in timing between walking and running at the same velocities (peaks occumng earlier in running) both for TF (in 4 sub- jects, average difference 10-14% of Tc) and for BF
(in 5 subjects, average difference 1620% of Tc). Changing velocities had no effect on the timing of TF or BF peaks in relation to the stride cycle (Fig. 4 B).
11: 4. Forward-backward movements (Fig. 5-6)
Linear displacements. The net horizontal rnove- ment amplitudes in the sagittal plane were small (t4 cm in all subjects). In all cases the values for
Acta Physiol Scand 121

16 A . Thorstensson et al.
04 - c7
0 3 . 0 WALK RUN
!
m : h / . 01
L3
:/
Fig. 5 . (A) Net forward-backward linear displacements and (B) time for peak forward displacement in percent of cycle duration at C7 and L3 levels versus velocity (cf. Fig. 1).
60
40'
2 0 .
0
20
C7 were smaller than for L3. In walking the mean values for C7 ranged approximately 1-1.5 cm and for L3 about 2-2.5 cm. The interindividual variation was large and there were no marked changes in mean amplitude with speed (Fig. 5A). In running there was a tendency of increasing net linear dis- placements with speed, both at C7 and L3, but the values were small and individual differences rela- tively large (Fig. 5A). There was no difference between walking and running at the same speeds at L3, whereas all subjects showed somewhat smaller values at C7 in running (Fig. 5A).
No or only minor changes were seen in the tim- ing of linear displacement peaks in relation to the stride cycle with increasing velocity, as shown in Fig. 5 B. A phase difference was observed between walking and running corresponding to approximate- ly 5-15% of Tc, the extreme forward position being closer to the instant of touch down in running.
. c7 L3
. - - - - - -. - - - - - - - - - - - - - - - - - - - . - - - - - - - - - - - - - - - - - - - - - - - -. - - 0 WALK . RUN - - 4 o - h 6
v-
. r
In running there was also a phase difference be- tween levels in such a way that the L3 level reached its extreme position about 10% of Tc earlier in the stride cycIe. The interindividual variation was rela- tively small.
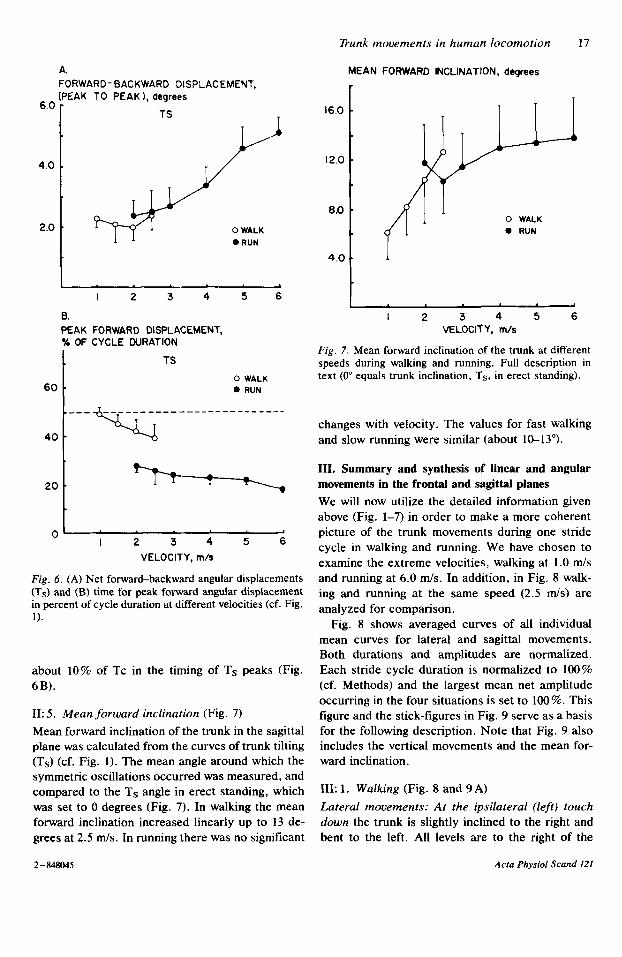
Angular di,splacement (Ts) (Fig. 6). The overall range in net angular displacements of the trunk in the sagittal plane for all subjects and velocities was 1.5-6". There was no change with speed in walking, whereas the values #in running increased (from, on average, 2.5 to 5') (Fig. 6A). There was no differ- ence in Ts between walking and running at the same velocity.
A gradual phase shift was observed in the timing of Ts peaks in relation to the stride cycle with in- creasing speed both in walking (10% of Tc, 1-2.5 d s ) and in running (10% of Tc, 2-6 m/s) (Fig. 6B). The transition from walking to running at the same speed (2-2.5 m l s ) resulted in a stepwise change of
r . . . . . .
Acra Physiol Scand 121
Trunk movements in human locomotion 17
16.0
12.0
8.0
4.0
6.0
4 .O
2.0
8
* I
A. FORWARD- BACKWARD DlSPLAC EMENT.
60
40
(PEAK
TS speeds during walking and running. Full description in text (0" equals trunk inclination, Ts, in erect standing). 0 WALK
* 0 RUN
_ _ _ _ _ _ _ _ _ _ - - _ _ _ _ - _ _ _ _ _ _ -- changes with velocity. The values for fast walking and slow running were similar (about 1G13"). :---\
t
TO PEAK 1, degrees
TS T
1 1 0 RUN
I 2 3 4 5 6
MEAN FORWARD INCLINATION, degrees r
0 WALK 0 RUN
0. PEAK FORWARD DISPLACEMENT.
I 2 3 4 5 6 VELOCITY. m/s
t 20
t 2 3 4 5 6
0' I
VELOCITY, m/r
Fig. 6. (A) Net forward-backward angular displacements (T,) and (B) time for peak forward angular displacement in percent of cycle duration at different velocities (cf. Fig. 1).
about 10% of Tc in the timing of Ts peaks (Fig. 6B).
11: 5 . Mean forward inclination (Fig. 7) Mean forward inclination of the trunk in the sagittal plane was calculated from the curves of trunk tilting (Ts) (cf. Fig. 1). The mean angle around which the symmetric oscillations occurred was measured, and compared to the Ts angle in erect standing, which w a s set to 0 degrees (Fig. 7). In walking the mean forward inclination increased linearly up to 13 de- grees at 2.5 m/s. In running there was no significant
111. Summary and synthesis of linear and angular movements in the frontal and sagittal planes We will now utilize the detailed information given above (Fig. 1-7) in order to make a more coherent picture of the trunk movements during one stride cycle in walking and running. We have chosen to examine the extreme velocities, walking at 1.0 d s and running at 6.0 m/s. In addition, in Fig. 8 walk- ing and running at the same speed (2.5 d s ) are analyzed for comparison.
Fig. 8 shows averaged curves of all individual mean curves for lateral and sagittal movements. Both durations and amplitudes are normalized. Each stride cycle duration is normalized to 100% (cf. Methods) and the largest mean net amplitude occurring in the four situations is set to 100%. This figure and the stick-figures in Fig. 9 serve as a basis for the following description. Note that Fig. 9 also includes the vertical movements and the mean for- ward inclination.
111: 1. Walking (Fig. 8 and 9 A) Lateral movements: At the ipsilateral (left) touch down the trunk is slightly inclined to the right and bent to the left. All levels are to the right of the
2-848045 Acta Physiol Scand I21

18 A . Thorstensson et al.
0 50 100 %Tc O 50 IOO Fig. 8. Averaged individual mean curves for linear and angular movements in the frontal (lateral movements) and sagittal planes (forward-backward movements) at different velocities (walking 1.0 and 2.5 m / s , running 2.5 and 6.0 m/s). For definitions of levels and angles, see Methods and Fig. 1. The curves are normalized with respect to time (stride cycIe=100%, cf. Fig. 2 and Methods) as well as to amplitude (the highest net amplitude in the four experimental situations is set to 100%). Solid vertical lines indicate iTD (at 0 and 100 % of Tc), broken vertical lines correspond to coTD (at 50 % of Tc). The duration of double leg support in walking and time in the air in running at the different velocities are given at the bottom of the figure.
midline and moving to the left. At higher walking (BF) is reached after &lo% of Tc. At this point the speeds the trunk is closer to the midline at touch C7 and L3 levels are close to the midline and the down. 50 % level is still to the right of the midline (walk 1 .O
The trunk continues to move towards the left side d s ) . At higher speeds the trunk has moved more to during the beginning of the ipsilateral support the left. At peak BF there is no inclination of the phase. Both inclination (TF) and bending (BF) are trunk (TF=O"). increasing to the left until the peak in feji bending The movement of all levels of the trunk continues
Acta Physiol Scand 121

Trunk movements in human locomotion 19
A. WALK 10 m/s 0. RUN 6.0 m/s
I TD co TD I TD coTD
0 10
MOVEMENTS IN THE FRONTAL PLANE MOVEMENTS IN THE FRONTAL PLANE LEFT - RIGHT L E F T - RIGHT
MOVEMENTS IN THE SAGITTAL PLANE MOVEMENTS IN THE SAGITTAL PLANE FMIWARO BAU(WR0 FOWbRO - BACKWARD
10 20 30 40 50 %Tc I I
coTD ? 0 10 20 30 40 50 %Tc
I TD coTD I TD
Fig. 9. Schematic two-dimensional representation of trunk movements in the frontal and sagittal planes during (A) walking (1.0 m / s ) and (B) running (6.0 d s ) . The figure is constructed from normalized mean values at every ten percent of Tc (0-50%). Symmetry in the stride cycle was found and therefore only 50% of the cycle is represented. The movement trajectories are successively built up at each level as the stride cycle proceeds from 0 to 50 %. At 50 % of Tc the trajectories in the frontal plane during a full stride cycle are completed with broken lines. In the sagittal plane the two corresponding trajectories per stride cycle are superimposed. The thin lines represent the mean inclination in the frontal (TF) and sagittal planes (Ts). The “intrinsic” lateral trunk bending, BF, is given by the thick broken lines between the C7, 50% and L3 markers. Definitions of levels and angles are given in Fig. 1. Vertical and horizontal amplitude scalings apply only to movement trajectories.
to the left during the single support phase of the left leg. At 1.0 m / s the peaks in left displacements occur at approximately 30% of Tc. Also at this point, the peak in left trunk inclination (TF), is attained. At higher walking speeds the extreme positions to the left are reached earlier in the stride cycle and all levels are moving to the right at peak left inclina- tion, which stiil occurs at about 30% of Tc after touch down. All levels are to the left of the midline. At peak inclination there is no bending of the trunk, B F = O O , i.e. a transition from left to right bending occurs.
At contralateral touch down the trunk is in a position corresponding to, but opposite to that at ipsilaterd touch down, i.e. all levels are to the left of the midline moving to the right. Then the move-
ments occur in analogy with the above but in the opposite direction until a “butterfly-like” trajec- tory is completed in one stride cycle (Fig. 9A).
Forward-backward movements. When discuss- ing the forward-backward movements of the trunk it should be remembered that they are measured in relation to the mean forward inclination, which in- creases linearly with walking speed (1-2.5 d s ) from 6 to 13 degrees (0’ at erect standing). This means that the trunk is never actually tilted back- wards, just moving in the backward direction.
At ipsilateral (left) touch down both the C7 and L3 levels are moving in the forward direction. The C7 level is close to (or somewhat in front of) the line of mean inclination, and the L3 level is behind (low speeds) or at this “midline” (high walking
Acta Physiol Scand I21

20 A. Thorstensson et al.
speeds). Peak forward inclination (Ts) occurs at touch down during walking at 1.0 m/s. At higher speeds there is a gradual shift of the forward peak to about 10% of Tc before touch down. Peak backward inclination, (Ts) of the trunk coin- cides approximately with the peak forward position of the C7 and L3 levels, particularly at higher speeds, and occurs in the beginning of the single support phase (15 to 25% of Tc). At contralateral touch down a similar trunk position is assumed as at ipsilateral touch down and two superimposed circular-shaped trajectories are completed during one stride cycle (Fig. 9A).
111: 2. Running (Fig. 8 and 9 B) Lateral movements. In slow running the relative movements of the three levels of the trunk are similar to that in walking at the corresponding speeds (cf. above). At running speeds above 2.5 d s , however, the situation is different: At ipsilater- a1 touch down the trunk is inclined to the left and bent to the left. The C7 level is to the left of the midline moving to the left, whereas both the 50% and the L3 levels are on the right side, the 50% level is moving to the left and the L3 level to the right. These movements continue until L3 reaches its peak right displacement at about 12% of Tc.
At peak left inclination (TF), (approximately 16% of Tc) all levels are moving to the left. The C7 level is close to its left peak (occurs at about 25 % of Tc) the 50% level to the midline and L3 to its right displacement peak. The trunk is almost straight (BF=OO), i.e. it is in a transition from left to right bending.
Peak trunk bending (BF) to the right is reached at approximately 40% of Tc, when the 50% level is close to its maximum left position, C7 and L3 are both close to the midline, C7 moving to the right and L3 to the left. The trunk is approximately vertical (TF=O”, i.e. in a transition from left to right inclination). At contralateral (right) touch down the trunk is inclined to the right and bent to the right. During the next half of the stride cycle similar trajectories as described above are formed in the opposite direction (Fig. 9B).
Forward-backward movements. In running the “midline” i.e. the mean forward inclination vanes between 10-14”. At ipsilateral touch down both the C7 and L3 levels are close to their most forward positions. Due to the larger movement amplitude of the L3 level there is a concomitant backward peak
in trunk inclination (Ts). The peaks in Ts coincide with the peaks in linear displacement (in the oppo- site direction) at the L3 level (Fig. 8).
Lateral versus forward-backward movements of the trunk (TF us. Ts): Walking. In slow walking the trunk is in its most forward position at touch down (Ts=7.3” at 1.0 d s ) . At this point the trunk is tilted (TF) towards the side opposite to that of touch down. When peaks in TF occur (at about 35 % and 85% of Tc) the trunk is close to the “midline” in the sagittal plane moving forward. With increasing speed the relation between TF and touch down is unchanged whereas Ts is closer to the “midline” (i.e. closer to the mean forward inclination). The peaks in lateral trunk inclination (TF) coincide with the forward peaks in Ts and occur at about 10% of Tc before touch down, whereas the backward peaks in Ts occur during a movement of TF to either side (Fig. 8 and 9A).
Running. Also in running the forward peak incli- nation (Ts) occurs at the same time as the peaks in lateral tilt (Tp). In slow running peak backward tilt occurs when TF is at the midline and coincides with touch down (Fig. 8).
DISCUSSION
This paper presents a comprehensive account of trunk movements in the lateral, forward-backward and vertical directions during human locomotion over a wide range of velocities, from slow walking (1.0 m/s) to relatively fast running (6.0 d s ) . Al- though the study is performed on a limited (n= 10) number of male subjects, it should give a general insight into the movement strategies used with changes in speed and mode of progression. The physical conditions during treadmill locomotion should not be different from overground, except for the absence of wind resistance, which might have an effect on the trunk movements, e.g. mean for- ward trunk inclination, at the highest velocities of running (Grillner 1981). Rotational movements of the trunk could not be analyzed with the present experimental design. It can be assumed that trunk rotation had minimal effects on the recorded dis- placements since the markers were placed axially along the spine (Gregersen & Lucas 1967, Thor- stensson et al. 1982, Cappozzo 1983).
The organization of the different phases of the stride cycle is different in walking and running (Grillner et al. 1979, Grillner 1981, Nilsson et al.
Acta Physiol Scand I21

Trunk movements in human locomotion 21
1983). In walking, the duration of the support phases is much longer and there are also phases of double leg support. In running at the normal speeds investigated, there are single support phases of short duration and periods of no support in between (Fig. 1). This means that the demands on equilibri- um control are markedly different during the two modes of locomotion. This is reflected in the move- ments of the trunk. In the frontal plane the trunk is essentially moved as a stiff segment from side to side in walking, but in running internal bending of the trunk (BF) is more pronounced. The turning points of BF occur mainly during the airborne phase in running in contrast to during the double support phase in walking. During the main part of the stride cycle in running the upper region of the trunk is on one side and the lower region on the other side of the midline. This is due to the fact that the L3 level is moving largely out of phase with the upper part of the trunk. At touch down of one leg the trunk is mainly on the opposite side of the midline, both in walking and running. However, the inclination of the trunk is towards the contralateral side in walking, with the lower level closest to the midline, whereas in run- ning the trunk inclination is towards the ipsilateral side with C7 at the midline. Also there are differ- ences in the amplitude and timing of the trunk movements in the sagittal plane. In walking, the trunk is inclined most forward at the end of single support or at the start of double support and least forward at the end of double support. In running, the trunk angular motion is essentially reversed as compared to walking with the least forward inclina- tion at touch down and the peak forward inclination coinciding with the termination of the support phase.
Another role of the trunk is to balance the head. It would seem advantageous if the trunk could act as a shock-absorber to minimize the movements (accelerations) of the head and thereby optimize the conditions for the visual and (perhaps) the vestibu- lar systems. This function is indicated when com- paring the movement amplitudes of the upper (C7) and lower part (L3) of the trunk (Fig. 1 , 3, 5). The movements at the C7 level are consistently smaller than at the L3 level in the forward-backward direc- tion at all speeds. The movements of C7 in this direction are not larger in fast running than in walk- ing (about 1.5 cm). There are more individual vari- ations in the lateral movements at different trunk
levels. Whereas the lateral movements of L3 in- crease with speed, C7-movements remain at the same amplitude or even decrease. This means that at running speeds of 4-6 m/s the smallest move- ments are seen at the C7-level. There is no marked difference between C7 and L3 in the vertical dis- placements. However, at low running speeds, where the vertical trunk movements are the largest (8-10 cm), there is a tendency towards smaller val- ues at the C7-level. A further damping of the move- ments may occur in the cervical spine.
In an earlier paper (Thorstensson et al. 1982)'we have discussed the function of the lumbar back muscles in relation to the overall angular move- ments (TF, Ts) of the trunk. It was concluded that the primary function of these muscles is to counter- act excessive trunk movements in the frontal plane during walking and in the sagittal plane during run- ning. The more detailed movement analysis in the present study further supports the previous conclu- sion. The coincidence of a peak in trunk lateral bending with a large peak of activity in the contrala- teral Erector Spinae muscle in walking could sug- gest that these muscles in addition act to induce the movement in the contralateral direction.
M. R. Zomlefer was supported by the Swedish Medical Research Council. The study was supported by grants from the Swedish Defence Research Organization (project no. H657), the Swedish Work Environment Fund (82- 0184) and the Swedish Medical Research Council (B83- O4X-06529-0 1 A).
REFERENCES BRAUNE, C. W. & FISCHER, 0. 1895. Der Gang des
Menschen. I. Abh Mat Phys C1 Kon Sachs Ges Wis- sensch 21: 151.
CAPPOZZO, A. 1981. Analysis of the linear displacement of the head and trunk during walking at different speeds. J Biomech 14: 41 1425.
CAPPOZZO, A. 1982. Low frequency self-generated vi- bration during ambulation in normal men. J Biomech 15: 599-609.
CAPPOZZO, A. 1983. The forces and couples in the human trunk during level walking. J Biomech 16: 599-609.
CAPPOZZO, A., FIGURA, F., LEO, T. & MAR- CHETTI, M. 1978. Movements and mechanical ener- gy changes in the upper part of the human body during walking. In: Biomechanics VI-A (ed. E. Asmussen and K. Jorgensen), pp. 272-279. University Park Press, Baltimore.
CARLSON, H. & THORSTENSSON, A. 1981. Control of the human trunk during locomotion. Acta Physiol Scand 114: 14A.
Acta Physiol Scand 121

22 A . Thorstensson et al.
CARLSON, H., NILSSON, J., THORSTENSSON, A. & ZOMLEFER, M. 1979. Control of the trunk in human treadmill walking. Commun. to the VIIth Int. Congr. of Biomechanics, Warzaw, Poland.
CAVAGNA, G. A., SAIBENE, F. P. & MARGARIA, R. 1963. External work in walking. J Appl Physiol 18: 1-9.
CAVAGNA, G. A., SAIBENE, F. P. & MARGARIA, R. 1964. Mechanical work in running. J Appl Physiol 19: 249-256.
FISCHER, 0. 1899-1904. Der Gang des Menschen. Abh Math Phys C1 Kon Sachs Ges Wissensch 11-25 (1900); 1-130. 111-26 (1901) 85-170, IV-26 (1901); 496-556. V- 28 (1904); 319418. VI-28 (1904); 531-617.
GREGERSEN, C. G. & LUCAS, D. B. 1967. An in vivo study of the axial rotation of the human thoracolumbar spine. J Bone Joint Surg 49A: 247-262.
GRILLNER, S. 1981. Control of locomotion in bipeds, tetrapods and fish. In: Handbook of Physiology-The nervous system I1 (ed. V. Brooks), pp. 1179-1236. Waverly Press, Baltimore, Maryland.
GRILLNER, S., HALBERTSMA, J., NILSSON, J. & THORSTENSSON, A. 1979. The adaptation to speed in human locomotion. Brain Research 165: 177-182
HALBERTSMA, J. 1983. The stride cycle of the cat: The modelling of locomotion by computerized analysis of automatic recordings. Acta Physiol Scand, Suppl. 521.
INMAN, V. T., RALSTON, H. J. & TODD, F. 1981. Human walking. Williams and Wilkins, Baltimore.
ITO, A., KOMI, P. V., SJODIN, B., BOSCO, C. & KARLSSON, J. 1983. Mechanical efficiency of posi- tive work in running at different speeds. Med Sci Sports 15: 299-308.
LAMOREUX, L. W. 1971. Kinematic measurements in the study of human walking. Bull Prosthetics Res 3: 10-15.
MURRAY, P., DROUGHT, B. & KORY, R. C. 1964. Walking patterns of normal men. J Bone Surg
MURRAY, M. P., KORY, R. C., CLARKSON, B. H. & SEPIC, S. B. 1966. Comparison between free and fast speed walking patterns of normal men. Am J Phys Med 45: 8-24.
SON, A. 1983. Speed adaptations in muscle activity and movement patterns of the lower limb in human locomotion (Manuscript).
THORSTENSSON, A., CARLSON, H., ZOMLEFER, M. R. & NILSSON, J. 1982. Lumbar back muscle activity in relation to trunk movements in man. Acta Physiol Scand 116: 13-20.
WATERS, R. L., MORRIS, J. & PERRY, J. 1973. Trans- lational motion of the head and trunk during normal walking. J Biomech 6: 167-172.
46A: 335-360.
NILSSON, J., HALBERTSMA, J. & THORSTENS-
Acta Physiol Scand 121




















