
TAp73 and DNp73 activate the expression of the pro-survival caspase-2S
Upload
independentCategory
view
0download
0
Journal of Biotechnology 95 (2002) 205–214
The protection of hybridoma cells from apoptosis bycaspase inhibition allows culture recovery when exposed to
non-inducing conditions
Albert Tinto a, Carme Gabernet a, Joaquim Vives a, Eva Prats b,Jordi J. Cairo a, Lluıs Cornudella b, Francesc Godia a,*
a Dept. d’Enginyeria Quımica, Uni�ersitate Autonoma de Barcelona, Escola Tecnica Superior d’Enginyers, ETSE, 08193 Bellaterra,Barcelona, Spain
b Dept. de Biologia Molecular i Cel ·lular, CID-CSIC, C/ Jordi Girona 18–26, 08034 Barcelona, Spain
Received 1 August 2001; received in revised form 13 December 2001; accepted 20 December 2001
Abstract
Programmed cell death (PCD) or apoptosis process in a hybridoma cell line induced by the deprivation of one ofthe main nutrients, glutamine, has been studied. The use of caspase inhibitors has enabled maintenance of cellviability during a significant period of time, when glutamine depletion was maintained in the culture. Two caspaseinhibitors partially suppressed the progress of PCD under glutamine deprivation: Ac-DEVD-cho and z-VAD-fmk.Indeed, as a consequence of this protection, the number of viable cells decreased by 10% (for z-VAD-fmk) and by80% (for Ac-DEVD-cmk) after 36 h of culture, while it decreased by 90% for a control culture in the absence ofprotective compounds. However, when the culture was exposed to non-apoptotic conditions after this period of timeunder apoptosis protection conditions, a normal growth pattern was not recovered. Interestingly, the simultaneoususe of both inhibitors made the recovery of the cell culture possible even after a period of 36 h under glutaminedepletion, indicating that the inhibition of the effector caspases occurs upstream of the point in which hybridoma cellsenter into the commitment step of the death programme. © 2002 Elsevier Science B.V. All rights reserved.
Keywords: Hybridoma; Glutamine-induced apoptosis; Culture recovery; Caspases; Caspase inhibitors
www.elsevier.com/locate/jbiotec
1. Introduction
The potential for the large-scale use of mono-clonal antibodies in diagnosis, therapeutics,downstream processing, and other fields of bio-
technological interest has resulted in serious atten-tion being directed towards the improvement ofprocesses based on hybridoma cell cultures (Hesseand Wagner, 2000).
Glucose and glutamine are the main carbon,nitrogen and energy sources in batch cultures ofhybridoma. However, as a consequence of theirexhaustion the process of apoptosis is triggered(Mercille and Massie, 1994; Simpson et al., 1998).
* Corresponding author. Tel.: +34-3-581-1018; fax: +34-3-581-2013.
E-mail address: [email protected] (F. Godia).
0168-1656/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved.
PII: S0 168 -1656 (02 )00012 -3

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214206
This death process is an evolutionarily conserved,genetic form of cell death, also known as pro-grammed cell death (PCD). Once fully activated itleads to the complete loss of viability in normalculture conditions, due to the irreversibility of theprocess (Mastrangelo and Betenbaugh, 1994).
Apoptosis occurs when a cell activates an inter-nal suicide program, encoded in cellular genes,leading to important changes in cell morphology.Such changes are chromatin condensation, DNAfragmentation and cytoplasmatic collapse result-ing in disassembly of the cell (Thornberry andLabzebnik, 1998). The insults leading the cell tostart the apoptosis programme are of a widerange: such as depletion of main nutrients, glu-cose and glutamine, other aminoacids, lack ofoxygen, or high osmolarity (Al-Rubeai and Singh,1998).
When mammalian cells are cultured in bioreac-tors, cell death by apoptosis severely impairs theculture performance (Mercille and Massie, 1994).Therefore, the manipulation of the genetic cellmachinery of apoptosis has raised considerableinterest due to the possibility of preventing it andtherefore extending the life-span of cultures, im-proving both cell viability and productivity, asshown by Itoh et al. (1995).
In order to protect animal cells in culture fromapoptosis, by means of a genetic modification, itis important to examine two important aspects.First, the identification of the targets allowing forthis protection during a significant time window.Second, the identification of the possibility torecover normal cell growth when, after a certaintime under apoptosis inducing conditions and inthe presence of the protection mechanism, thecells are returned into normal culture conditions,non-apoptosis inducing. This second aspect is crit-ical for the practical application of apoptosis pro-tection strategies in cell culture in bioreactors,where the apoptosis process can be triggered bymany factors. It is therefore important to protectthe cells during a certain time while the anomalyin the process (for example the depletion of amedium component or a hardware failure) is be-ing corrected.
To achieve this goal, the present study is fo-cused on the aspartate-specific proteases termed
caspases which are critically involved in the apop-totic process in mammalian cells (Alnemri et al.,1996; Cohen, 1997; Nicholson, 1999) and have animportant role in the commitment, amplificationand demolition phases of the death programme(Slee et al., 1999). These enzymes exist as zy-mogens composed of a prodomain, and large andsmall catalytic subunits. Generation of active cas-pases requires accurate processing of internal as-partic residues to liberate the prodomain andproduce the two chain active enzymes (Stennickeand Salvesen, 1998). The use of specific peptidicinhibitors that bind to the catalytic site of cas-pases (Villa et al., 1997) should enable determina-tion of the possibility of recovering KB26.5hybridoma cell cultures from apoptosis. Particu-larly, the use of three specific peptide compoundsis investigated: z-VAD-fmk, Ac-DEVD-cho, andz-YAVD-cmk.
2. Material and methods
2.1. Cell line
The KB26.5 murine hybridoma was used in allthe experiments performed. This cell line producesan IgG3 monoclonal antibody against antigen A1
of the red blood cells as described previously(Damgaard et al., 1993).
2.2. Cell culture
The basal medium used was Dulbecco’smodified Eagle’s media (DMEM) from GIBCOsupplemented with 2% foetal calf serum (FCS,Biological Industries), 6 mM L-glutamine and 25mM glucose, both from Sigma-Aldrich, 3.7 g l−1
sodium bicarbonate (Probus), 15 mg l−1 phenolred, 0.391 mg l−1 �-mercaptoethanol (Sigma) and0.836 mg l−1 insulin (Novo). Cultures were car-ried out in 25 cm2 T-flasks with 10 ml of workingvolume, using 2×105 cells ml−1 as seeding den-sity, and grown at 37 °C in a humidified atmo-sphere of 5% CO2 air mixture (Forma ScientificCO2 incubator) as described previously (Paredeset al., 1999). Density and viability were assessedby the trypan blue exclusion method using a

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214 207
hemocytometer (Neubauer improved, Brand) andlight microscopy. This method has an error of 5%.
2.3. Glutamine measurement
Glutamine concentrations were measured byHPLC (Hewlett Packard 1090) using a reversephase column (aminoquant 200×2.1 mm) as pre-viously described (Sanfeliu et al., 1996). The errorin this analysis was less than 5%.
2.4. Caspase inhibitors
Ac-YVAD-cmk, Ac-DEVD-cho and z-VAD-fmk were obtained from Bachem. Ac-YVAD-cmkand Ac-DEVD-cho were reconstituted in 50%acetic acid and added to the cultures at differentfinal concentrations, and pH was adjusted withNaOH. z-VAD-fmk was dissolved in methanoland supplemented to the media at the same con-centrations as the former two. Control cultureswere performed with equivalent quantities ofmethanol and acetic acid than for the experimentsusing the peptide compounds. Previously, it waschecked that the quantities of methanol and aceticacid used had no effect on cell proliferation orviability.
2.5. Apoptosis induction
To induce apoptosis, 5×105 cells l−1 wereseeded into 25 cm2 T-flasks and cultured in glu-tamine deprived medium.
2.6. Quantification of apoptosis protection
In order to have a quantitative estimation ofthe apoptosis protection obtained by the use ofpeptide compounds, the decrease in viable cellconcentration was evaluated at a given time as apercentage of the initial viable cell concentration.
2.7. Reco�ery assay
Aliquots of cells cultured under conditions ofglutamine deprivation and at different concentra-tions of caspases inhibitors were harvested atdifferent times, centrifuged for 10 min at 500×gand grown in glutamine supplemented medium.
2.8. Analysis of apoptosis
2.8.1. Confocal microscopyCells were examined for apoptotic morphology
using a solution of 200 g l−1 acridine orange(Sigma) and 200 g l−1 ethidium bromide (Bio-probes) at a final concentration of 8 �g l−1.Phosphatidylserine translocation was examinedwith Annexine-V-fluos (Boehringer–Mannheim)specific staining and DNA fragmentation usingthe enzymatic assay TUNEL (Promega).
2.8.2. Flow cytometryAn aliquot of 3×105 cells was washed twice
with PBS and harvested by centrifugation, incu-bated for 15 min at room temperature with An-nexine-V-fluos (Roche), diluted 1:10 in 8.18 g l−1
NaCl (Panreac), 0.55 g l−1 CaCl2 (Panreac), 2.38g l−1 HEPES (Sigma), and light protected. Fi-nally the cells were analysed on a Becton–Dickin-son FCAScan.
2.8.3. DNA fragmentation assay by agarose gelelectrophoresis
Performed according to the methodology al-ready described (Sambrook et al., 1989).
2.8.4. Cytochrome c detectionCytochrome c release from mitochondria was
detected by Western blot analysis as described byMartin et al. (1996). Primary antibody againstcytochrome c (Ab-2) was obtained from Onco-gene (cPC333). Secondary goat polyclonal anti-body anti-rabbit IgG HRP-conjugate wasobtained from Upstate Biotechnology (c12-348).
3. Results and discussion
3.1. Induction of apoptosis in the hybridomaKB26.5 by glutamine depletion
When the cells of KB26.5 hybridoma are grownin normal conditions, using a complete mediumformulation with 5 mM glutamine, cell death isoccurring sharply at 72 h, when glutamine isexhausted, as shown in Fig. 1A. The mechanismof cell death was via apoptosis, and 52% of the
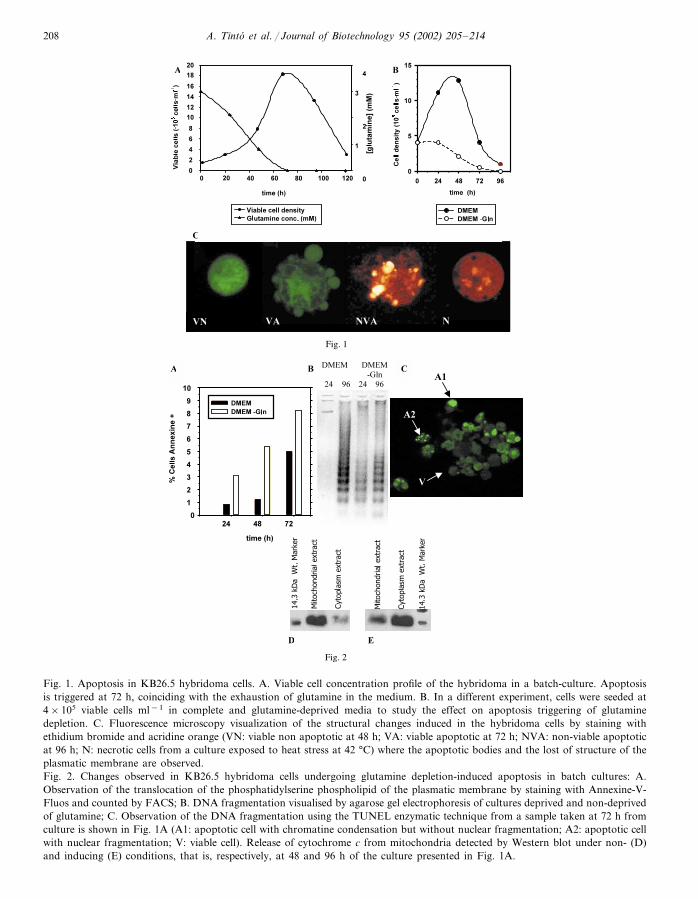
A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214208
Fig. 1. Apoptosis in KB26.5 hybridoma cells. A. Viable cell concentration profile of the hybridoma in a batch-culture. Apoptosisis triggered at 72 h, coinciding with the exhaustion of glutamine in the medium. B. In a different experiment, cells were seeded at4×105 viable cells ml−1 in complete and glutamine-deprived media to study the effect on apoptosis triggering of glutaminedepletion. C. Fluorescence microscopy visualization of the structural changes induced in the hybridoma cells by staining withethidium bromide and acridine orange (VN: viable non apoptotic at 48 h; VA: viable apoptotic at 72 h; NVA: non-viable apoptoticat 96 h; N: necrotic cells from a culture exposed to heat stress at 42 °C) where the apoptotic bodies and the lost of structure of theplasmatic membrane are observed.Fig. 2. Changes observed in KB26.5 hybridoma cells undergoing glutamine depletion-induced apoptosis in batch cultures: A.Observation of the translocation of the phosphatidylserine phospholipid of the plasmatic membrane by staining with Annexine-V-Fluos and counted by FACS; B. DNA fragmentation visualised by agarose gel electrophoresis of cultures deprived and non-deprivedof glutamine; C. Observation of the DNA fragmentation using the TUNEL enzymatic technique from a sample taken at 72 h fromculture is shown in Fig. 1A (A1: apoptotic cell with chromatine condensation but without nuclear fragmentation; A2: apoptotic cellwith nuclear fragmentation; V: viable cell). Release of cytochrome c from mitochondria detected by Western blot under non- (D)and inducing (E) conditions, that is, respectively, at 48 and 96 h of the culture presented in Fig. 1A.

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214 209
cells showed apoptotic morphology at 72 h (Fig.1C).
The effect of apoptosis triggering by glutamineis further evidenced when cells are cultured inmedium deprived of glutamine. In this case, celldeath is observed early in the experiment, whencompared to the control culture using completemedium formulation (Fig. 1B). In the glutaminedeprived culture, the phosphatidylserine external-ization (an early signal of the culture) is 56% at48 h and 82% at 72 h (Fig. 2A). Chromosomalcondensation followed by DNA degradation tooligonucleosomal fragments is also evidenced inFig. 2B and C. The release of cytochrome c fromthe mitochondria to the cytosol is also observedclearly in Fig. 2D and E, showing the key role ofthis event in the progress of the apoptotic path-way. Under conditions of glutamine starvation,the process of apoptosis starts due to the lack ofprecursors for aminoacids, nucleotides, and car-bon synthesis as well as energy supply.
3.2. Determination of the effect of chemicalinhibitors of caspases in�ol�ed in apoptosis in thehybridoma KB26.5 under glutamine depri�ation
Specific caspase inhibitors (Garcia-Calvo et al.,1998; Ekert et al., 1999) were added to the cellcultures in order to elucidate whether the actionof caspases in the apoptosis cascade is situatedup-stream or down-stream of the point whereirreversible commitment to death occurs in a glu-
tamine deprived culture. Three different moleculeswhere selected: the broad spectrum caspase in-hibitor z-VAD-fmk, the group II cysteineproteases inhibitor Ac-DEVD-cmk and the groupI specific inhibitor peptide z-YVAD-cmk. Theireffects were investigated at increasing concentra-tions up to 300 �M under conditions of glutaminedeprivation in the cell culture. As a control, anexperiment without any of the inhibitormolecules, also in conditions of glutamine depri-vation was carried out (Table 1).
3.3. Use of z-VAD-fmk
Incubation with the broad range caspase in-hibitor z-VAD-fmk at 50–100 �M substantiallydelayed the initiation of apoptosis and extendedthe viability of the cultures up to 36 h. Transloca-tion of phosphatidylserine was reduced by 75%and changes in cellular morphology were alsoobserved to a lower extent while the process ofdegradation of DNA was totally blocked (Fig.3A, B, C). However, at 60 h the protection is nolonger effective, and the viability in both experi-ments (that is, in presence of peptide and itscontrol with no peptide) is almost the same, aswell as the percentage of cells with phos-phatidylserine translocation. However, it shouldbe highlighted that at this time (60 h), DNAobtained from the cells cultured in the presence of100 �M z-VAD-fmk was still not fragmented.
Table 1Experiments performed with caspase inhibitors in batch cultures of hybridoma KB26.5
Culture z-VAD-fmk (�M) Ac-DEVD-cmk (�M) z-YVAD-cmk (�M)
Effect of z-VAD-fmk 10 – –50 – –
––10010 ––Effect of Ac-DEVD-cmk
– 50 –– 100 ––Effect of z-YVAD-cmk – 100– – 200
– 300–
For each series of experiments with a specific peptide molecule, a control experiment was run, exposing the cells to a glutaminedeprived medium, without addition of any peptide.
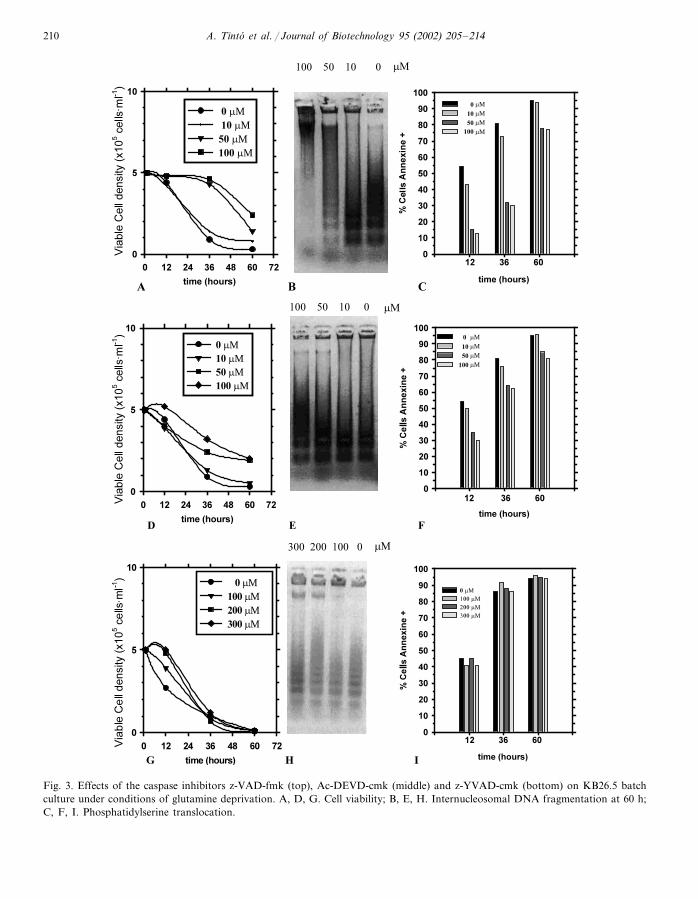
A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214210
Fig. 3. Effects of the caspase inhibitors z-VAD-fmk (top), Ac-DEVD-cmk (middle) and z-YVAD-cmk (bottom) on KB26.5 batchculture under conditions of glutamine deprivation. A, D, G. Cell viability; B, E, H. Internucleosomal DNA fragmentation at 60 h;C, F, I. Phosphatidylserine translocation.

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214 211
3.4. Use of Ac-DEVD-cho
When the peptide Ac-DEVD-cho was used, inconcentrations up to 100 �M, a certain delay ofapoptotis was also observed, although in a lesserextent than in the previous case, as can be ob-served in Fig. 3D, E, F. These effects could beobserved basically at the level of cell morphologyand to some degree by phosphatidylserine translo-cation, that was lower when the peptide waspresent, especially at the sample taken at 12 h, butthe samples taken at higher times showed norelevant differences among the use of this peptideand the control culture.
3.5. Use of z-YVAD-cmk
Concentrations of the z-YVAD-cmk tetrapep-tide up to 200 �M produced no effect on theprogress of apoptosis in KB26.5 hybridoma cells.When a higher concentration, 300 �M, was used,only a small effect could be observed and it wasperceptibly less effective than z-VAD-fmk andAc-DEVD-cho, already at the observation madeat 12 h. From this point cell viability decreaseddramatically and at 36 h the number of apoptoticcells was similar to the control culture, as ob-served in Fig. 3G, H, I.
Although caspase-inhibitors do not target indi-vidual caspases, considering the inhibitory con-stants reviewed by Ekert et al. (1999), and thebiochemical pathway of caspases (reviewed byBudihardjo et al., 1999, and Slee et al., 1999,among others) the results obtained with thesepeptidic inhibitors give some information aboutthe specific caspases involved in the glutamine-in-duced apoptosis in KB26.5 hybridoma cell line. Inthis sense, z-VAD-fmk has been reported as astrong inhibitor of caspases 1, 3, 5, 7, 8 and 9,with inhibition constants (Ki) values lower than0.1 �M. However, there is not any homologue ofcaspase 5 in murine cell lines (Nicholson, 1999)and caspase 8 can be most probably discarded inthis case, since it is mainly involved in apoptosisprocesses mediated through membrane receptorsand not by cellular stress (Varfolomeev et al.,1998). z-VAD-fmk showed a marked reduction ofdeath by apoptosis in hybridoma cultures but also
showed that even in this case, the death programwas activated after a certain time (60 h), thereforeindicating the possibility of other unknown cas-pases being active in the cells, although to a lowerextent.
z-YVAD-cmk, an inhibitor that has been de-scribed to affect in a specific way only caspase 1(Ekert et al., 1999), showed a low degree ofprotection, thus indicating that this particular cas-pase has a low implication in the apoptosis pro-gram in the cell line studied.
When Ac-DEVD-cho was used, an extra inhibi-tion of apoptosis was observed in z-VAD-fmktreated cultures. This effect could be done becauseits strong inhibition of caspase 3, but also cas-pases 7 and 8.
Finally, when Ac-DEVD-cho and z-VAD-fmkwere used simultaneously the strongest inhibitioneffect was obtained (Fig. 4A, B). The combinationof these results would indicate the principal roleof caspases 9 and 3 in the observed apoptosisprocess in this cell line, although they are notconclusive regarding a certain participation ofcaspase 7, and in a later stage of some unknowncaspases.
3.6. Reco�ery of culture �iability
After the study of the effect of different in-hibitors affecting different caspase families, thepossibility to recover cell growth after a period ofexposure to the specific inhibitors and glutaminedeprivation was further investigated, using thetwo peptides that had been more effective.
Medium depletion of glutamine was initiallysupplied with Ac-DEVD-cmk and z-VAD-fmk atoptimal concentrations of 100 �M. The experi-ments of cell recovery consisted in the expositionof cells to apoptosis inducing conditions and afterdifferent periods of time cells were harvested andresuspended in complete medium, with glutamine,that is normal growth conditions. These experi-ments where carried out using four different pos-sibilities: control (no peptide addition), onlyz-VAD-fmk, only Ac-DEVD-cho, and both pep-tides simultaneously. Cell recovery was studied at12, 24, 36 and 60 h. The results are summarized inTable 2.
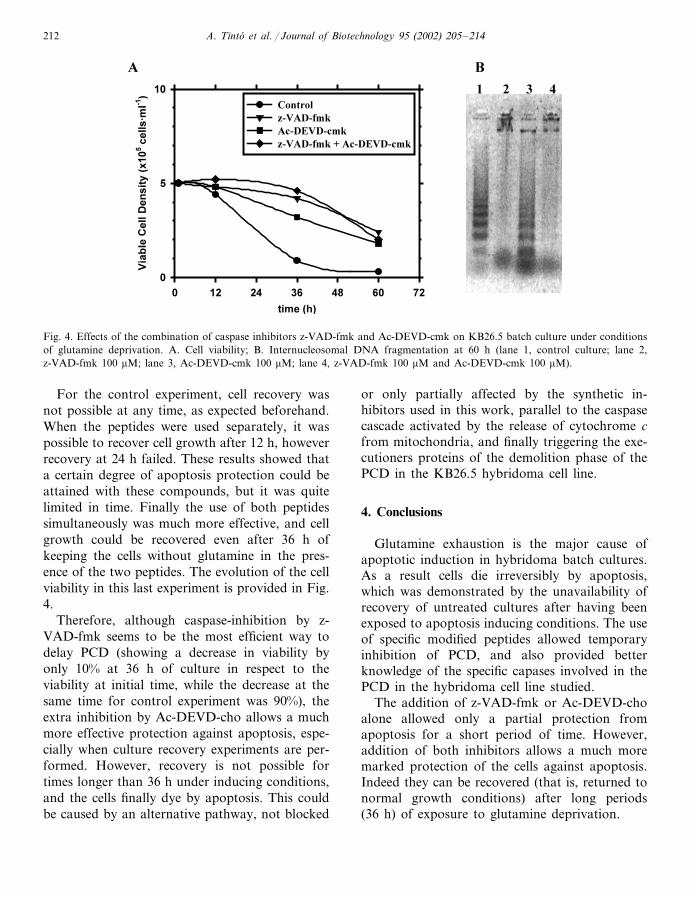
A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214212
Fig. 4. Effects of the combination of caspase inhibitors z-VAD-fmk and Ac-DEVD-cmk on KB26.5 batch culture under conditionsof glutamine deprivation. A. Cell viability; B. Internucleosomal DNA fragmentation at 60 h (lane 1, control culture; lane 2,z-VAD-fmk 100 �M; lane 3, Ac-DEVD-cmk 100 �M; lane 4, z-VAD-fmk 100 �M and Ac-DEVD-cmk 100 �M).
For the control experiment, cell recovery wasnot possible at any time, as expected beforehand.When the peptides were used separately, it waspossible to recover cell growth after 12 h, howeverrecovery at 24 h failed. These results showed thata certain degree of apoptosis protection could beattained with these compounds, but it was quitelimited in time. Finally the use of both peptidessimultaneously was much more effective, and cellgrowth could be recovered even after 36 h ofkeeping the cells without glutamine in the pres-ence of the two peptides. The evolution of the cellviability in this last experiment is provided in Fig.4.
Therefore, although caspase-inhibition by z-VAD-fmk seems to be the most efficient way todelay PCD (showing a decrease in viability byonly 10% at 36 h of culture in respect to theviability at initial time, while the decrease at thesame time for control experiment was 90%), theextra inhibition by Ac-DEVD-cho allows a muchmore effective protection against apoptosis, espe-cially when culture recovery experiments are per-formed. However, recovery is not possible fortimes longer than 36 h under inducing conditions,and the cells finally dye by apoptosis. This couldbe caused by an alternative pathway, not blocked
or only partially affected by the synthetic in-hibitors used in this work, parallel to the caspasecascade activated by the release of cytochrome cfrom mitochondria, and finally triggering the exe-cutioners proteins of the demolition phase of thePCD in the KB26.5 hybridoma cell line.
4. Conclusions
Glutamine exhaustion is the major cause ofapoptotic induction in hybridoma batch cultures.As a result cells die irreversibly by apoptosis,which was demonstrated by the unavailability ofrecovery of untreated cultures after having beenexposed to apoptosis inducing conditions. The useof specific modified peptides allowed temporaryinhibition of PCD, and also provided betterknowledge of the specific capases involved in thePCD in the hybridoma cell line studied.
The addition of z-VAD-fmk or Ac-DEVD-choalone allowed only a partial protection fromapoptosis for a short period of time. However,addition of both inhibitors allows a much moremarked protection of the cells against apoptosis.Indeed they can be recovered (that is, returned tonormal growth conditions) after long periods(36 h) of exposure to glutamine deprivation.

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214 213
Table 2Effect of caspase inhibitors on the reversibility of the apoptotic process in the hybridoma KB26.5
RecoveryInhibitor present
12 h 24 h 36 hAc-DEVD-cmk (�M) 60 hz-VAD-fmk (�M)
– – − − − −–100 + − − −
+ −100 −– −100100 + + + −
Cultures were deprived of glutamine and after the lapses of time indicated cells were centrifuged and cultured in fresh mediumcontaining glutamine. ‘+’ and ‘−’ symbols indicate whether the cultures were rescued or not, when re-exposed to non-inducingapoptosis conditions.
Caspases 3 and 9 were found to be the maineffectors of the apoptotic signal in the hybridomaKB26.5 and they are situated upstream from thepoint after which cells enter in the commitmentphase of apoptosis. Due to the economical unfea-sibility of the use of peptide caspase inhibitors inbioreactors, future work will be focused on thegenetic inhibition of these caspases by means ofthe overexpression of endogenous and viral in-hibitors of apoptosis or engineered proteins in-volved in delaying or blocking the apoptoticprocess. In fact, the protection observed by thecombination of different peptides indicate thatdifferent caspases are involved in channelling theapoptotic signal in the cells and that potentialgenetic modifications should consider more thanone single target.
Acknowledgements
The present work has been developed in theframework of the Centre de Referencia en Biotec-nologia (Generalitat de Catalunya) and supportedby the Plan Nacional de Biotecnologia (BIO97-0542). J.V. is a recipient of a fellowship fromCIRIT (1999FI00779) and C.G. from MEC(PN97 43709939).
References
Al-Rubeai, M., Singh, R.P., 1998. Apoptosis in cell culture.Curr. Op. Biotechnol. 9, 152–156.
Alnemri, E.S., Livingston, D.J., Nicholson, D.W., Salvesen,G., Thornberry, N.A., Wong, W.W., Yuan, J., 1996. Hu-man ICE/CED-3 protease nomenclature. Cell 87, 171.
Budihardjo, I., Olover, H., Lutter, M., Luo, X., Wang, X.,1999. Biochemical pathways of caspase activation duringapoptosis. Annu. Rev. Cell Dev. Biol. 15, 269–290.
Cohen, G.M., 1997. Caspases: the executioners of apoptosis.Biochem. J. 326, 1–16.
Damgaard, B., Sanfeliu, A., Cairo, J.J., Casas, C., Sola, C.,Godia, F., 1993. Substitution of transferrin by FeCl3 in thedevelopment of low foetal calf serum concentrations forKB-26.5 hybridoma cell line. Cytotechnology 13, 133–141.
Ekert, P.G., Silke, J., Vaux, D.L., 1999. Caspase inhibitors.Cell Death Differ. 6, 1081–1086.
Garcia-Calvo, M., Peterson, E.P., Leiting, B., Ruel, R.,Nicholson, D.W., Thornberry, N.A., 1998. Inhibition ofhuman caspases by peptide-based and macromolecular in-hibitors. J. Biol. Chem. 273, 32608–32613.
Hesse, F., Wagner, R., 2000. Development and improvementsin the manufacturing of human therapeutics with mam-malian cell cultures. TIBTECH 18, 173–180.
Itoh, Y., Veda, H., Suzuki, E., 1995. Overexpression of bcl-2,apoptosis suppressing gene: prolonged viable culture pe-riod of hybridoma and enhanced antibody production.Biotech. Bioeng. 48, 118–122.
Martin, S.J., Amarantes-Mendes, G.P., Shi, L., Chuang, T.H.,Casiano, C.A., O’Brien, G.A., Fitzgerald, P., Tan, E.M.,Bockoch, G.M., Greenberg, A.H., Green, D.R., 1996. Thecytotoxic cell protease granzyme B initiates apoptosis in acell-free system by proteolytic processing and activation ofthe ICE/CED-3 family protease, CPP32, via a novel two-step mechanism. EMBO J. 15, 2407–2416.
Mastrangelo, A.J., Betenbaugh, M.J., 1994. Overcoming apop-tosis: new methods for improving protein-expression sys-tems. TIBTECH 16, 88–95.
Mercille, S., Massie, B., 1994. Induction of apoptosis in nutri-ent-deprived cultures of hybridoma in myeloma cells. Bio-technol. Bioeng. 44, 1140–1154.
Nicholson, D.W., 1999. Caspase structure, proteolytic sub-strates, and function during apoptotic cell death. CellDeath Differ. 6, 1028–1042.

A. Tinto et al. / Journal of Biotechnology 95 (2002) 205–214214
Paredes, C., Prats, E., Cairo, J.J., Cornudella, L., Godia, F.,1999. Modification of glucose and glutamine metabolism inhybridoma cells through metabolic engineering. Cytotech-nology 30, 85–93.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. MolecularCloning: A Laboratory Manual, 2nd ed. Cold SpringHarbour Laboratory, Cold Spring Harbour, New York.
Simpson, N.H., Singh, R.P., Perani, A., Goldenzon, C., Al-Rubeai, M., 1998. In hybridoma cultures, deprivation ofany single amino acid leads to apoptotic death, which issuppressed by the expression of the bcl-2 gene. Biotech.Bioeng. 59, 90–98.
Sanfeliu, A., Cairo, J.J., Casas, C., Sola, C., Godia, F., 1996.Analysis of nutritional factors and physical conditionsaffecting growth and monoclonal antibody production ofthe hybridoma KB-26.5 cell line. Biotechnol. Prog. 12,209–216.
Slee, E.A., Adrain, C., Martin, S.J., 1999. Serial killers: order-ing caspase activation events in apoptosis. Cell DeathDiffer. 6, 1067–1074.
Stennicke, H.R., Salvesen, G.S., 1998. Properties of the cas-pases. Biochem. Biophys. Acta 1387, 17–31.
Thornberry, N.A., Labzebnik, Y., 1998. Caspases: enemieswithin. Science 281, 1312–1316.
Varfolomeev, E.E., Schuchmann, M., Luria, V., Chian-nilkulchai, N., Beckmann, J.S., Mett, I.L., Rebrikov, D.,Brodianski, V.M., Kemper, O.C., Kollet, O., Lapidot, T.,Soffer, D., Sobe, T., Avraham, K.B., Goncharov, T., Holt-mann, H., Lonai, P., Wallach, D., 1998. Targeted disrup-tion of the mouse Caspase 8 gene ablates cell deathinduction by the TNF receptors, Fas/Apo1, and DR3 andis lethal prenatally. Immunity 9, 267–276.
Villa, P., Kaufmann, S.H., Earnshaw, W.C., 1997. Caspasesand caspase inhibitors. TIBS 22, 388–393.
Copyright © 2022 FDOKUMEN




















