
Screening for Caspase-3 Inhibitors: A New Class of Potent Small-Molecule Inhibitors of Caspase-3
9
10.1177/1087057105285048 Okun et al. Novel Small-Molecule Caspase-3 Inhibitors Screening for Caspase-3 Inhibitors: A New Class of Potent Small-Molecule Inhibitors of Caspase-3 ILYA OKUN, 1,2 SERGEI MALARCHUK, 1 , ELENA DUBROVSKAYA, 1 ALEXANDER KHVAT, 1 SERGEY TKACHENKO, 1 VOLODYMYR KYSIL, 1 ALEXEY ILYIN, 2 DMITRY KRAVCHENKO, 2 ERIC R. PROSSNITZ, 3 LARRY SKLAR, 3 and ALEXANDRE IVACHTCHENKO, 1,2 From the authors’650,000 compound collection, they have selected approximately 15,000 potential small-molecule protease inhibitors, which were subjected to high-throughput screening against caspase-3. The screening yielded a series of hits that belong to 11 different scaffolds. Based on the structure of one of the hits, a new class of the small-molecule inhibitors with a double electrophilic warhead, 8-sulfonyl-pyrrolo[3,4-c]quinoline-1,3-diones (SPQ), was synthesized and tested in follow-up mechanistic and antiapoptosis assays. Mechanistic analysis of a representative compound of this class, CD-001-0011, showed that the compound exhibited a high potency (IC 50 = 130 nM), was reversible though noncompetitive, and had a broad selectivity profile to other caspases belonging to groups I to III. The compound was effective in preventing staurosporine- induced apoptosis in a few cell lines and retinoic acid–induced apoptosis in zebrafish. (Journal of Biomolecular Screening 2006:277-285) Key words: caspase-3, small molecules, inhibitors, apoptosis, structure-activity relationship INTRODUCTION A POPTOSIS, OR PROGRAMMED CELL DEATH, is a normal compo- nent of the development and health of organisms. Cells die in response to a variety of stimuli, and during apoptosis, they do so in a controlled, regulated fashion. Apoptosis is a process in which cell-signaling mechanisms play an active role in the cell’s own death without triggering inflammation. This makes apoptosis dis- tinct from other forms of cell death, such as necrosis, in which un- controlled cell death leads to cell lysis, release of intracellular con- tent, triggering of inflammatory responses, and, potentially, serious health problems. A major biochemical pathway involved in apoptosis includes a family of proteases, known as caspases, which act in a cascade to activate downstream caspases responsible for breakdown or cleav- age of key cellular substrates required for normal cellular function, including structural proteins in the cytoskeleton and nuclear pro- teins such as DNA repair enzymes. Caspases are found practically in all organisms from Caenorhabditis elegans to humans. 1 At least 12 of the caspases have been cloned (caspases 1 through 10, 13, and 14). 2,3 Caspases 11 and 12, 2 murine homologs, have no known human counterpart. Two major functionally distinct groups of the caspases are cur- rently recognized as participating in either inflammation or apoptosis. Caspases share similarities in amino acid sequence, structure, and substrate specificity 4 and are subdivided into 2 subfamilies based on their functionality: caspases involved in in- flammation (caspases 1, 4, 5, 11, 12, 13, and 14) and apoptosis- related caspases (caspases 2, 3, 6, 7, 8, 9, and 10). Based on their position in the apoptotic signal pathway, the subfamily of apoptosis-related caspases has also been subdivided into 2 groups: initiator caspases (caspases 2, 8, 9, and, probably, 10) and execu- tioner (or effector) caspases (caspases 3, 6, and 7). 5 Activation of caspase-3 is a key event integrating upstream sig- nals into final execution of cell death. This makes caspase-3 an in- teresting therapeutic target, and the search for caspase-3 inhibitors is an ongoing endeavor many pharmaceutical companies are em- bracing in the search for effective drugs. 6-9 As an alternative to the 2 extremes of drug discovery—purely rational drug design and blind screening campaigns—in silico computational docking has resulted in the design and synthesis of novel small-molecule inhibitors of cysteinyl proteases, including caspases. Different classes of the reversible and irreversible prote- ase inhibitors discovered through these efforts are described in Schirmeister and Kaeppler. 10 Screening of a protease-focused li- © 2006 Society for Biomolecular Sciences www.sbsonline.org 277 1 ChemDiv, Inc., 11885 Sorrento Valley Road, San Diego, CA 92121. 2 Chemical Diversity Research Institute, Khimki, Moscow Region, Russia 114401. 3 University of New Mexico, 915 Camino de Salud NE, Albuquerque, NM. Received May 31, 2005, and in revised form Nov 2, 2005. Accepted for publica- tion Nov 15, 2005. Journal of Biomolecular Screening 11(3); 2006 DOI: 10.1177/1087057105285048
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Screening for Caspase-3 Inhibitors: A New Class of Potent Small-Molecule Inhibitors of Caspase-3

10.1177/1087057105285048Okun et al.Novel Small-Molecule Caspase-3 Inhibitors
Screening for Caspase-3 Inhibitors:A New Class of Potent Small-Molecule Inhibitors of Caspase-3
ILYA OKUN,1,2 SERGEI MALARCHUK,1, ELENA DUBROVSKAYA,1 ALEXANDER KHVAT,1
SERGEY TKACHENKO,1 VOLODYMYR KYSIL,1 ALEXEY ILYIN,2 DMITRY KRAVCHENKO,2
ERIC R. PROSSNITZ,3 LARRY SKLAR,3 and ALEXANDRE IVACHTCHENKO,1,2
From the authors’650,000 compound collection, they have selected approximately 15,000 potential small-molecule proteaseinhibitors, which were subjected to high-throughput screening against caspase-3. The screening yielded a series of hits thatbelong to 11 different scaffolds. Based on the structure of one of the hits, a new class of the small-molecule inhibitors with adouble electrophilic warhead, 8-sulfonyl-pyrrolo[3,4-c]quinoline-1,3-diones (SPQ), was synthesized and tested in follow-upmechanistic and antiapoptosis assays. Mechanistic analysis of a representative compound of this class, CD-001-0011,showed that the compound exhibited a high potency (IC50 = 130 nM), was reversible though noncompetitive, and had a broadselectivity profile to other caspases belonging to groups I to III. The compound was effective in preventing staurosporine-induced apoptosis in a few cell lines and retinoic acid–induced apoptosis in zebrafish. (Journal of Biomolecular Screening2006:277-285)
Key words: caspase-3, small molecules, inhibitors, apoptosis, structure-activity relationship
INTRODUCTION
APOPTOSIS, OR PROGRAMMED CELL DEATH, is a normal compo-nent of the development and health of organisms. Cells die in
response to a variety of stimuli, and during apoptosis, they do so ina controlled, regulated fashion. Apoptosis is a process in whichcell-signaling mechanisms play an active role in the cell’s owndeath without triggering inflammation. This makes apoptosis dis-tinct from other forms of cell death, such as necrosis, in which un-controlled cell death leads to cell lysis, release of intracellular con-tent, triggering of inflammatory responses, and, potentially,serious health problems.
A major biochemical pathway involved in apoptosis includes afamily of proteases, known as caspases, which act in a cascade toactivate downstream caspases responsible for breakdown or cleav-age of key cellular substrates required for normal cellular function,including structural proteins in the cytoskeleton and nuclear pro-teins such as DNA repair enzymes.
Caspases are found practically in all organisms fromCaenorhabditis elegans to humans.1 At least 12 of the caspaseshave been cloned (caspases 1 through 10, 13, and 14).2,3 Caspases11 and 12, 2 murine homologs, have no known human counterpart.Two major functionally distinct groups of the caspases are cur-rently recognized as participating in either inflammation orapoptosis. Caspases share similarities in amino acid sequence,structure, and substrate specificity4 and are subdivided into 2subfamilies based on their functionality: caspases involved in in-flammation (caspases 1, 4, 5, 11, 12, 13, and 14) and apoptosis-related caspases (caspases 2, 3, 6, 7, 8, 9, and 10). Based on theirposition in the apoptotic signal pathway, the subfamily ofapoptosis-related caspases has also been subdivided into 2 groups:initiator caspases (caspases 2, 8, 9, and, probably, 10) and execu-tioner (or effector) caspases (caspases 3, 6, and 7).5
Activation of caspase-3 is a key event integrating upstream sig-nals into final execution of cell death. This makes caspase-3 an in-teresting therapeutic target, and the search for caspase-3 inhibitorsis an ongoing endeavor many pharmaceutical companies are em-bracing in the search for effective drugs.6-9
As an alternative to the 2 extremes of drug discovery—purelyrational drug design and blind screening campaigns—in silicocomputational docking has resulted in the design and synthesis ofnovel small-molecule inhibitors of cysteinyl proteases, includingcaspases. Different classes of the reversible and irreversible prote-ase inhibitors discovered through these efforts are described inSchirmeister and Kaeppler.10 Screening of a protease-focused li-
© 2006 Society for Biomolecular Sciences www.sbsonline.org 277
1ChemDiv, Inc., 11885 Sorrento Valley Road, San Diego, CA 92121.2Chemical Diversity Research Institute, Khimki, Moscow Region, Russia114401.3University of New Mexico, 915 Camino de Salud NE, Albuquerque, NM.
Received May 31, 2005, and in revised form Nov 2, 2005. Accepted for publica-tion Nov 15, 2005.
Journal of Biomolecular Screening 11(3); 2006DOI: 10.1177/1087057105285048

brary that we have assembled from our internal collection of morethan 650,000 compounds yielded several new classes of small-molecule caspase-3 inhibitors. In this report, we describe mecha-nistic and selectivity profiles of a new series of potent nonpeptideinhibitors, as well as their cell-based and zebrafish antiapoptoticaction.
MATERIALS AND METHODS
Reagents
Caspase-3 was purchased from Upstate USA (Charlottesville,VA). Caspases 1, 4 to 6, and 9, Ac-WEHD-AMC, Ac-WEHD-AMC, and Ac-WEHD-AMC were from Calbiochem (San Diego,CA). Caspases 2, 7, 8, and 10 were from R & D Systems (Minne-apolis, MN). Ac-DEHD-AMC, Ac-VEHD-AMC, Ac-IETD-AMC, Ac-VEHD-AMC, and Ac-IETD-AMC were from Sigma(St. Louis, MO), and Ac-DEVD-AMC and Ac-DEVD-AMC andAc-DEVD-CHO were from Bachem (King of Prussia, PA). Allother reagents were of a research grade.
Library selection
We used our “focused diversity” approach to select 15,338compounds from our 650,000 compound collection for screeningagainst caspase-3. In accordance with this approach, one of the se-lection criteria was based on the warhead concept,11-13 with a modi-fication to include only electrophilic compounds with reversibleformation of chemical bonds. As the active site of caspase-3 con-tains a sulfhydryl moiety necessary for its catalytic activity, we fo-cused the selection on molecules with the warheads capable of re-acting with the cysteinyl thiols. At the same time, the core of themolecules was represented by a diversity of heterocyclic structuressuch as quinoxalinones, pyridones, pyrimidinones,pyrimidinediones, qiunolones, and so forth.
Enzyme assay
The assay buffer contained 2 mM EDTA, 0.1% Chaps, 20 mMHEPES pH 7.4, and 5 mM DTT. Thirty microliters of the substrate,50 µM Ac-DEVD-AMC, and 30 µL of 0.7 U/µL caspase-3 wereadded to the microplates. The enzymatic activity of the caspase-3was measured at ambient temperature by registering the time-dependent accumulation of the fluorescent product, methyl-coumarine, which is formed during cleavage of the fluorogenicsubstrate, Ac-DEVD-AMC. Our data (not shown) indicated lin-earity of the Ac-DEVD-AMC cleavage for up to 10 h, indicatingabsence of a product inhibition or plate reader photomultiplier sat-uration. Caspase-3 was freshly prepared from a frozen stock by di-luting with the assay buffer. Activities of the other caspases weremeasured similarly using the corresponding fluorogenic sub-strates: Ac-WEHD-AMC for caspase-1, Ac-DEHD-AMC forcaspase-2, Ac-WEHD-AMC for caspase-4, Ac-WEHD-AMCfor caspase-5, Ac-VEID-AMC for caspase-6, Ac-DEVD-AMC
for caspase-7, Ac-IEPD-AMC for caspase-8, and Ac-LEHD-AMC for caspase-9.
Reversibility of inhibition assay
Thirty microliters of caspase-3 (14 U/µL) in assay buffer con-taining 5 mM DTT was mixed with 30 µL of 3 µM CD-001-0011(the final compound concentration of 1.5 µM is approximately 10-fold higher than its IC50). The mixture was incubated for 1 h atroom temperature and then diluted 10-fold with the assay buffer.The substrate was added as described above to the diluted aliquotstaken at the indicated time points. As a control, a mixture ofcaspase-3 and CD-001-0011 (0.7 U/µL and 0.15 µM, respectively)was prepared and incubated for the same time durations before ad-dition of the substrate.
Cell apoptosis assay
SH-S5Y5 (human neuroblastoma), Jurkat (human T-cell leuke-mia), and NIH-3T3 (mouse embryonic fibroblast) cell lines wereobtained from ATCC. SH-S5Y5 cells were cultured in DMEM/F-12 medium with L-glutamine, HEPES, and pyridoxine(Invitrogen, Carlsbad, CA). Jurkat and NIH-3T3 cells were cul-tured in RPMI-1640 medium with L-glutamine (Invitrogen). Allculture media were supplemented with 10% fetal bovine serum(NovaTech) and antibiotic/antimycotic solution (Sigma). Cellswere grown at 37° C in 5% CO2/95% air environment at 100%humidity.
SH-S5Y5 and HIH-3T3 cells were plated into half-area wells of96-well cell culture plates (Corning, Corning, NY) at 104 cell/wellin 45 µL of medium and grown overnight before compound addi-tion. Jurkat cells were plated in 45 µL/well of serum-freehybridoma medium (Invitrogen) at 2.5 × 105 cells/mL and incu-bated for 1 h before compound addition. A total of 10 mM DMSOstock compound solutions was diluted 1:10 with growth medium,and 5 µL of the resulting 1 mM (10% DMSO) compound solutionswas added to the cells. Ten percent DMSO was also used for con-trol (no compounds) wells. Cells were incubated for 30 min beforeaddition of 5 µL of the apoptosis inducer staurosporine (final con-centration of 100 nM). After 24 or 48 h of incubation, AlamarBlue® reagent (Biosource, Camarillo, CA) was added at 5.5 µL/well. Cells were further incubated for 90 min, and the fluorescenceof converted Alamar Blue® dye was measured using the WallacVictor2V plate reader (PerkinElmer, Wellesley, MA) at 531/589nm ex/em wavelengths. The percent protection from inducedapoptosis was calculated using the following equation:
% *protectionFl Fl
Fl Fli s
o s
=−
−
∆ ∆
∆ ∆100,
where ∆Fl denotes the difference in Alamar Blue® fluorescencemeasured at point 0 and after 90-min incubation with the cells;subscripts i, s, and o define, respectively, fluorescence in the pres-ence of a compound and staurosporine, staurosporine alone, andno staurosporine.
Okun et al.
278 www.sbsonline.org Journal of Biomolecular Screening 11(3); 2006

Zebrafish apoptosis assay
Embryo collection. Zebrafish experiments were performed byPhylonix Pharmaceuticals, Inc. Embryos were generated by natu-ral pairwise mating in accordance with the work of Westerfield.14
Embryos were maintained at 28° C in fish water (200 mg InstantOcean Salt; Aquarium Systems, Mentor, OH) per liter of deionizedwater, pH 6.6 to 7.0, maintained with 2.5 mg/L Jungle pH stabi-lizer (Jungle Laboratories Corporation, Cibolo, TX; conductivity670-760 µS). Embrions were used for the experiments 24 hpostfertilization (hpf).
Embryo treatment. Ten embryos were used for each regimen: 1)control (1% DMSO) 24-hpf embryos, 2) retinoic acid + 1%DMSO-treated 24-hpf embryos, and 3) retinoic acid pluscompound-treated 24-hpf embryos. Treatment duration was 24 h.Thus, apoptosis in the forebrain region was examined at 48 hpf.
Visualization of apoptotic cells in forebrain. Zebrafish were im-mersed in 1 µg/mL acridin orange in phosphate-buffered saline for1 h and rinsed twice in 10 mL of fresh fish water. Fish were anes-thetized in MESAB (0.5 mM 3-aminobenzoic acid ethyl ester, 2mM Na2HPO4) and mounted in methylcellulose in depressedslides for observation with a Zeiss M2Bio fluorescence micro-scope (Carl Zeiss Microimaging Inc., Thornwood, NY).
Compound screening
A library of small molecules (about 15,000 compounds) withpurity higher than 95% was screened in a 384-well plate format. ABiomekFX Assay Work Station (Beckman Coulter, Fullerton,CA) was used for plate formatting and reagent distribution. Fluo-rescence measurements were performed using a VICTOR2V 96/384 multilabel plate reader (PerkinElmer Life Sciences, Boston,MA) at λex = 360 nm and λem = 460 nm (top readout) in Corningblack 384-well plates.
A total of 195 µL of the assay buffer, as indicated above, wasadded to each well of the 96-well plates containing 5 µL of 10-mMcompounds dissolved in DMSO. The 384-well black assay plateswere prepared by distributing 2.5 µL of the diluted compoundsinto each well and sequentially adding the substrate (30 µL) andthe enzyme (30 µL). Thus, the final screening concentration of thecompounds was 10 µM (0.1% DMSO). The plates were thentransferred to the plate reader, and the fluorescence intensitieswere monitored for at least 1 h with 0.5-min reading intervals.
For IC50 determinations, the 10-mM compound solutions werefirst serially diluted in DMSO, and equal aliquots (5 µL) of each di-lution were added to 15 µL of the assay buffer. All other proce-dures were performed as described above. Thus, in these experi-ments, the highest compound concentration was 100 µM, withserial 3-fold dilutions at a constant DMSO concentration of 1%.
Data fitting and manipulations
All curve fittings were performed with Prism 4 software(GraphPad, San Diego, CA), using built-in equations describingcorresponding data models.
HTS quality was assessed using Z′ factor,15 which describes awindow between maximal and minimal signal compared to datavariability inside the screening experiment. Z′ factor calculationswere performed for each plate using data obtained from the platecolumns reserved for caspase-3 without (Max signal) and in thepresence of its tetrapeptide inhibitor, Ac-DEVD-CHO, (Min sig-nal). Calculations were performed using the following equation:
where σMax and σMin are, respectively, standard deviations of maxi-mal (Max) and minimal (Min) readings.
RESULTS
Primary screening
For assay quality control purposes and Z′ factor calculations,each 384-well plate contained 32 wells (columns A01-P01 andA02-P02) with the enzyme only (Max Enzyme Activity) and 32wells (columns A23-P23 and A24-P24) with the enzyme and a 60-nM peptide inhibitor, Ac-DEVD-CHO (Min Enzyme Activity).Thus, for each plate, its own screening window had been estab-lished between the caspase-3 Max Enzyme Activity and Min En-zyme Activity.
The quality of the caspase-3 assay was very good (Z′ factoracross all the plates tested was higher than 0.8). The primary screenof the selected compounds at a concentration of 10 µM revealed 299hits (higher than 50% enzyme inhibition threshold). Thus, the fo-cused library has produced a 1.9% hit rate, which is much better thantheusual rateof 0.01% to 0.1% when diverse libraries are screened.
The hits discovered during the primary HTS screening cam-paign represented at least 11 new chemotypes, which have notbeen previously described as inhibitors of cysteine proteases. Twochemotype libraries, which produced potent inhibitors, are de-scribed elsewhere.16,17
Here we describe in more detail 2 new chemotypes found amongthe hits belonging to 8-sulfonyl-pyrrolo[3,4-c]quinoline-1,3-diones(SPQ), which are based on the general template I (Table 1).
First, we tested compounds of the chemotype 1 series of SPQanalogs, 4-methyl-8-(4-methyl-piperidine-1-sulfonyl)-pyrrolo[3,4-c]quinoline-1,3-diones, by varying content of the Rgroup (Table 1). There was a clear trend in inhibition potency ofthe compounds. More bulky lipophilic groups such as cycloheptyland cyclohexyl substantially decreased the potency, and more po-lar and hydrophilic groups increased the potency. Next, we fixedthe R-group of the chemotype 1 to the ethoxy substitute found in
Novel Small-Molecule Caspase-3 Inhibitors
Journal of Biomolecular Screening 11(3); 2006 www.sbsonline.org 279

the most potent compound, CD-001-0011, and analyzed a series ofcompounds by substituting the piperidine moiety in position 8 (Ta-ble 1) with primary amines of benzene and benzene derivatives(chemotype 2). For all studied compounds of the chemotype 2, nosubstantial differences were evident, although all of them were lesspotent than CD-001-0011.
Unlike known caspase-3 inhibitors, the compounds of the SPQseries have 2 electrophilic carbonyls that might serve as sites ofnucleophilic attack by cysteine thiolates (Fig. 1).
The CD-001-0011 (Table 1), which exhibited fairly goodcaspase-3 inhibition potency (IC50 ≈ 130 nM), was further charac-
terized in the follow-up mechanistic, cell-based, and whole-organism (zebrafish) antiapoptosis studies.
Mechanistic analysis of caspase-3 inhibitionby CD-001-0011
Reversibility of the caspase-3 inhibition with CD-001-0011was determined in the dilution experiments as described in the Ma-terials and Methods section. From the data shown in Figure 2, onecan see that although control samples exhibited no time-dependentchange in caspase-3 activity, the inhibition after dilution decreased
Okun et al.
280 www.sbsonline.org Journal of Biomolecular Screening 11(3); 2006
Template I
N
O
OO
SO
N
NR
Chemotype 1
CH3
O
OO
SO
N
NN
H
OH
R1
Chemotype 2
Cpd ID R IC50, nM Cpd ID R1 IC50, nM
CD-004-0013 *
68,900 ± 7000 CD-004-0021 Cl
* 311 ± 28
CD-004-0025 * 40,600 ± 5000 CD-004-0023
O
CH3
* 310 ± 30
CD-004-0016 CH3 2130 ± 205 CD-004-0022 OH
* 268 ± 21
CD-004-0026 S
N
*
CO2C2H5
864 ± 57 CD-004-0024
CH3CH3
* 256 ± 24
CD-004-0014 CO2CH3
*
534 ± 46 CD-004-0019 CH3*
243 ± 32
CD-004-0015 CH2CO2tert
-Bu 378 ± 36 CD-004-0020 *
219 ± 18
CD-004-0017 CH2CO2H 321 ± 29
CD-004-0027 S
*
CO2CH3
306 ± 26
CD-001-0011 CH2CH2OH 130 ± 23
R3
O
O
O
SO
N
NR2
R1
Table 1. Structure-Activity Relationship Analysis of Hits Around Chemotypes 1 and 2 Derived From Compounds of Template I
Values of IC50 are shown for compounds with different R and R1
substitutes.

with time to the level characteristic of the control (1×) samples.This indicates reversible interaction of CD-001-0011 withcaspase-3. The dissociation half-life, estimated in accordance withan equation describing 1st-order dissociation kinetics, was ap-proximately equal to 50 min.
Competition studies were performed using 2 types of experi-ments. In one experimental setting, the CD-001-0011 inhibitionconcentration curves were registered at several substrate concen-trations (Fig. 3). In an orthologous setting, the substrateconcentration-dependent rates of enzyme activity were measuredin the presence of different concentrations of CD-001-0011 (Fig.4). Both experiments indicated noncompetitive inhibition. Inhibi-tion experiments such as illustrated in Figure 3 revealed no right-ward shift upon increasing the substrate concentration, whichwould be expected for competitive inhibition. In the classical Mi-chaelis-Menten experiments (Fig. 4), increases in inhibitor con-centration lead to diminishing of Vmax, with no statistically signifi-cant changes in the KM values.
The selectivity profile of CD-001-0011 was assessed using 9commercially available caspases, 1 through 9. Substrates and con-
ditions used for each of the kinases are described in the Materialsand Methods section. The results are presented in Figure 5.
The results were grouped for caspases in accordance with theirselectivity at the P4 site.18 CD-001-0011 nonselectively inhibitscaspases of group I with potencies in the range of 500 nM to 800nM. It is approximately an order of magnitude less potent in inhib-iting caspases of group III and shows some selectivity amongmembers of the group II, with caspase-3 having the highest sensi-tivity (pIC50 ≈ 7) among the caspases tested. The maximal differ-ence in potencies covers 2 orders of magnitude ranging from about10 µM (caspases 8 and 9, group III) to about 100 nM (caspase 3,group II).
Novel Small-Molecule Caspase-3 Inhibitors
Journal of Biomolecular Screening 11(3); 2006 www.sbsonline.org 281
Enz-SH
N
OH
O
N
R3
R 2
R 1
SEnz
R 2
R 1
N
O
O
N
R3
R 1
R 2
N
O
OH
N
R3
S
Enz
FIG. 1. Possible modes of caspase thiolate nucleophilic attack on the“phtalimide” carbonyls of 8-sulfonyl-pyrrolo[3,4-c]quinoline-1,3-diones.
0 30 60 90
50
60
70
80
90
100CD-001-0011 (10X)CD-001-0011 (1X)
Time after dilution, min
% In
hib
itio
n
FIG. 2. Reversibility of caspase-3 inhibition with CD-001-0011.Caspase-3 at a 10× concentration, as specified in the Materials and Meth-ods section, was preincubated with 1.5-µM CD-001-0011 for 1 h and thenwas diluted 10-fold with the assay buffer. At the indicated time points afterthe dilution, the caspase-3 substrate was added, and the reaction rate wasmeasured (closed circles). In control experiments (open circles), caspase-3 was preincubated with the compound at 1× concentrations, 0.7 U/µLand 0.15 µM, respectively, for 1 h, and the substrate was added at the indi-cated time points. The percentage inhibition was calculated relative to theactivity of caspase-3, which went through all the procedures for concen-trated or control samples but without the inhibitor. The inhibition reversaldata were fitted with the following equation: i = I0 × exp(–t/τ) + B, where I0
is a maximal inhibition at zero time point, τ is a lifetime constant, and B isa basal inhibition with CD-001-0011 at 150 nM.
10-9 10-8 10-7 10-6 10-5 10-4
0
25
50
75
100
2 mM6 mM20 mM60 mM
Ac-DEVD-AMC
[CD-001-0011], M
% In
hib
itio
n
FIG. 3. Concentration-dependent inhibition of caspase-3 with CD-001-0011 measured in the presence of different substrate, Ac-DEVD-AMC, concentrations. The percentage of inhibition was calculated rela-tive to the enzyme activity at each of the substrate concentrations with noinhibitor present.
FIG. 4. Substrate concentration-dependent caspase-3 enzymatic ratesmeasured in the presence of a different concentration of the inhibitor, CD-001-0011. The enzyme was preincubated for 15 min with the inhibitor atthe indicated concentrations, and the enzymatic rate was registered uponaddition of the substrate, Ac-DEVD-AMC. In the insert is a table with theenzymatic Vmax and KM values corresponding to the inhibitorconcentration.

Antiapoptotic effect of CD-001-0011
Antiapoptotic activity of CD-001-0011 was determined in sev-eral cell lines and in a whole animal model (zebrafish). The cellapoptosis was induced by staurosporine in cell cultures and byretinoic acid in zebrafish, as described in the Materials and Meth-ods section. Apoptosis in cells was measured as the suppression ofAlamar Blue® reduction rate after 24 to 48 h incubation withstaurosporine or with the mixture of staurosporine and CD-001-0011. Zebrafish experiments were performed at PhylonixPharmaceuticals, Inc. (Cambridge, MA) in accordance with theprotocol described in the Materials and Methods section.
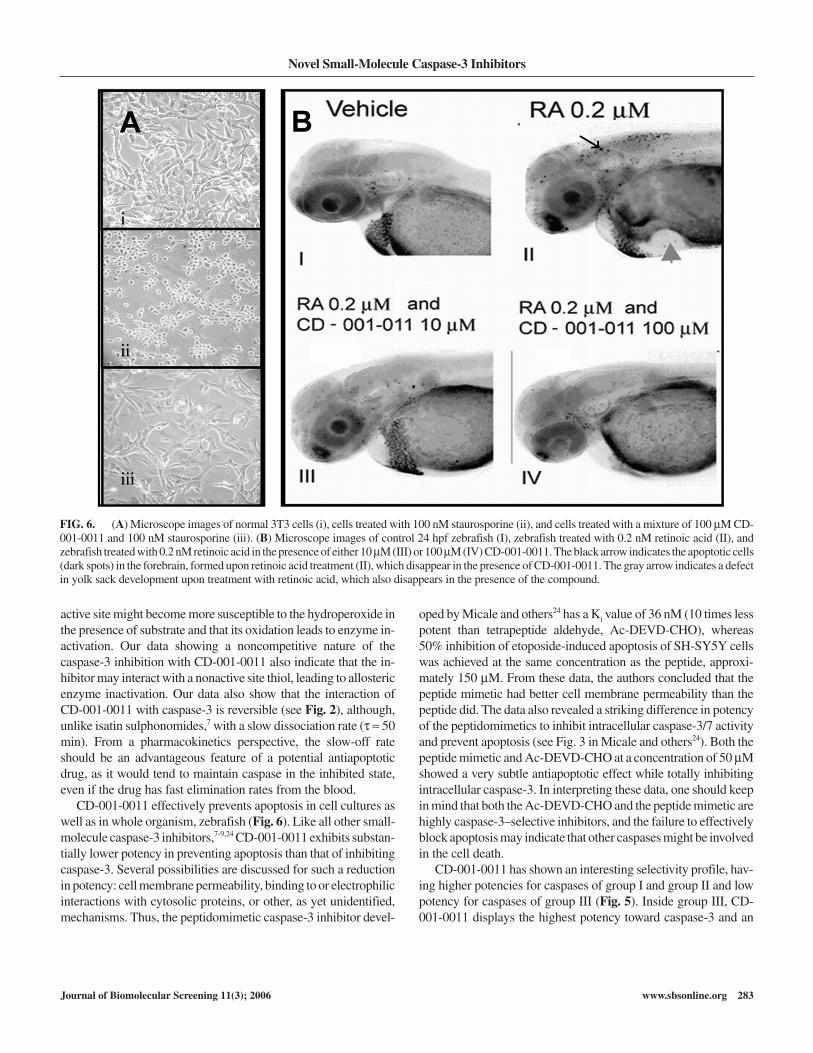
Both Table 2 and Figure 6A demonstrate the protective activityof CD-001-0011 against staurosporine-induced apoptosis in cellcultures. Apoptosis leads to drastic changes (rounding and shrink-age) in cell morphology (Fig. 6A, ii), increased intracellularcapase-3/7 activity (data not shown), and diminished rate ofAlamar Blue® reduction. CD-001-0011 effectively prevented thestaurosporine-induced cell morphology changes (compare Fig.6A, ii and iii) and staurosporine-induced cell death in several celllines (Table 2).
CD-001-0011 also effectively blocked retinoic acid–inducedapoptosis of neuronal cells in a whole animal (zebrafish). Theapoptotic cells appear as black dots in the forebrain (small arrow inFig. 6B, II). In the presence of CD-001-0011, retinoic acid failed toinduce neuronal cell apoptosis as evidenced by the absence of theblack dots in the forebrain (Fig. 6B, III and IV). The compoundalso effectively prevented retinoic acid–induced defects in devel-oping yolk sack (large arrow, Fig. 6B, II).
In the cell- and zebrafish-based assays, much higher concentra-tions of the inhibitor were needed to achieve protective effect as
compared with inhibition experiments using purified enzyme. Asimilar difference between caspase-3 inhibition and whole-cellantiapoptotic potency was shown for isatin sulfonamides7 andanilinoquinazolines.8 Such a difference could be due to many fac-tors, including poor cell membrane permeability, metabolic trans-formation inside the cells, protein binding, and so forth.7,8 At leastfor isatin sulfonamides, it was shown that cytosol binding is re-sponsible for lower potency in cell-based assay versus isolated en-zyme assay.7
DISCUSSION
Cysteine proteases play an important role in different patholo-gies and are expected to be valuable therapeutic and antiviral tar-gets. Efforts to identify small-molecule inhibitors are well underway.10 The SPQs are structurally distinct from other known classesof nonpeptide caspase-3 inhibitors, including isatin sulfonamides.Unlike previously reported inhibitors, they possess 2 potential sitesfor nucleophilic attack by cysteinyl thiolate (see Fig. 1). Our datashow that the interaction of the inhibitor with caspase-3 is revers-ible though noncompetitive, which raises the possibility that itmight act allosterically through interaction with sulfhydrylgroup(s) distinct from the active site. Indeed, it was shown thatcaspases can also be regulated by protein thiol modification at sitesother than the active site cysteine.19 Thus, Cys 244, located near theactive site of caspase-1, is believed to be essential for the caspase-1activity and is a target of the enzyme inhibition by thimerosal.20
The authors are not aware of any existing data that clarify the roleof other than active-site cysteinyls in caspase-3 activity. However,Hampton and others21 have shown that inactivation of caspase-3with hydrogen peroxide substantially increases when a substrate ispresent before hydrogen peroxide is added. It is difficult to explainsuch behavior by increased susceptibility of the active site thiol tothe peroxide, which is accessible to solvent in the groove of the ac-tive site formed by 4 loops when no substrate is present22 (Fig. 7A)and is inaccessible or partially accessible to a solvent in the boundstate23 (when a peptide is present in its active site; Fig. 7B).
Caspase-3 has total of 7 cysteinyls, although some of them areburied deep in the molecule. Still, at least 3 of them (excluding theone in active site) are readily accessible to water and hence to po-tential oxidation. One can speculate that cysteinyl thiol outside the
Okun et al.
282 www.sbsonline.org Journal of Biomolecular Screening 11(3); 2006
Caspas
e1
Caspas
e4
Caspas
e5
Caspas
e2
Caspas
e3
Caspas
e7
Caspas
e6
Caspas
e8
Caspas
e9
5
6
7
Group I Group II Group III
pIC
50
FIG. 5. Selectivity profile of CD-001-0011. Potencies of the com-pound to inhibit caspases tested were measured as described in the Materi-als and Methods section. Average potencies, pIC50 = –Log(IC50), withstandard deviations are shown. The caspases are grouped in accordancewith their substrate specificity at the P4 site.
Table 2. Antiapoptotic Activity of 100 µM CD-001-0011Measured by Prevention of Staurosporine-Induced Decrease in
Alamar Blue® Fluorescence
Cell Line % Protection from Apoptosisa
Jurkat 55 ± 7 (3)NIH-3T3 69 ± 8 (3)SH-SY5Y 52 ± 8 (3)
Calculation of percentage protection is described in the Materials and Methods section.Number of independent experiments, each in triplicate, is shown in parentheses.
active site might become more susceptible to the hydroperoxide inthe presence of substrate and that its oxidation leads to enzyme in-activation. Our data showing a noncompetitive nature of thecaspase-3 inhibition with CD-001-0011 also indicate that the in-hibitor may interact with a nonactive site thiol, leading to allostericenzyme inactivation. Our data also show that the interaction ofCD-001-0011 with caspase-3 is reversible (see Fig. 2), although,unlike isatin sulphonomides,7 with a slow dissociation rate (τ ≈ 50min). From a pharmacokinetics perspective, the slow-off rateshould be an advantageous feature of a potential antiapoptoticdrug, as it would tend to maintain caspase in the inhibited state,even if the drug has fast elimination rates from the blood.
CD-001-0011 effectively prevents apoptosis in cell cultures aswell as in whole organism, zebrafish (Fig. 6). Like all other small-molecule caspase-3 inhibitors,7-9,24 CD-001-0011 exhibits substan-tially lower potency in preventing apoptosis than that of inhibitingcaspase-3. Several possibilities are discussed for such a reductionin potency: cell membrane permeability, binding to or electrophilicinteractions with cytosolic proteins, or other, as yet unidentified,mechanisms. Thus, the peptidomimetic caspase-3 inhibitor devel-
oped by Micale and others24 has a Ki value of 36 nM (10 times lesspotent than tetrapeptide aldehyde, Ac-DEVD-CHO), whereas50% inhibition of etoposide-induced apoptosis of SH-SY5Y cellswas achieved at the same concentration as the peptide, approxi-mately 150 µM. From these data, the authors concluded that thepeptide mimetic had better cell membrane permeability than thepeptide did. The data also revealed a striking difference in potencyof the peptidomimetics to inhibit intracellular caspase-3/7 activityand prevent apoptosis (see Fig. 3 in Micale and others24). Both thepeptide mimetic and Ac-DEVD-CHO at a concentration of 50 µMshowed a very subtle antiapoptotic effect while totally inhibitingintracellular caspase-3. In interpreting these data, one should keepin mind that both the Ac-DEVD-CHO and the peptide mimetic arehighly caspase-3–selective inhibitors, and the failure to effectivelyblock apoptosis may indicate that other caspases might be involvedin the cell death.
CD-001-0011 has shown an interesting selectivity profile, hav-ing higher potencies for caspases of group I and group II and lowpotency for caspases of group III (Fig. 5). Inside group III, CD-001-0011 displays the highest potency toward caspase-3 and an
Novel Small-Molecule Caspase-3 Inhibitors
Journal of Biomolecular Screening 11(3); 2006 www.sbsonline.org 283
FIG. 6. (A) Microscope images of normal 3T3 cells (i), cells treated with 100 nM staurosporine (ii), and cells treated with a mixture of 100 µM CD-001-0011 and 100 nM staurosporine (iii). (B) Microscope images of control 24 hpf zebrafish (I), zebrafish treated with 0.2 nM retinoic acid (II), andzebrafish treated with 0.2 nM retinoic acid in the presence of either 10 µM (III) or 100 µM (IV) CD-001-0011. The black arrow indicates the apoptotic cells(dark spots) in the forebrain, formed upon retinoic acid treatment (II), which disappear in the presence of CD-001-0011. The gray arrow indicates a defectin yolk sack development upon treatment with retinoic acid, which also disappears in the presence of the compound.

order of magnitude lower potency toward caspase-2. At present,though a general quest is being directed to identifying and devel-oping highly selective caspase-3 inhibitors as potential drug candi-dates, there is not enough preclinical and clinical information tosuggest that the high selectivity would be of real advantage. Broad-spectrum inhibitors could be used for a multiplicity of therapeuticareas. Indeed, a role of caspase-1 inhibitors has been thought to bemainly useful in treatment of inflammation. However, there are in-dications that caspase-1 might also be a key enzyme inneurodegenerative disease-induced neuronal cell apoptosis.25
Other caspases, in particular caspase-3, can also be activated26 insuch diseases. Therefore, discovery of small-molecule caspase in-hibitors with different selectivity profiles could be important inunderstanding pathologies and could lead to a development ofhighly effective drugs.
ACKNOWLEDGMENTS
We gratefully acknowledge our collaborators from PhylonixPharmaceuticals, Inc. for determining efficacy of CD-001-0011 inthe zebrafish apoptosis model.
REFERENCES
1. Thornberry NA, Lazebnik Y: Caspases: enemies within. Science1998;281:1312-1316.
2. Earnshaw WC, Martins LM, Kaufmann SH: Mammalian caspases: structure,activation, substrates, and functions during apoptosis. Annu Rev Biochem1999;68:383-424.
3. Van de Craen M, Van Loo G, Pype S, Van Criekinge W, Van den Brande I,Molemans F, et al: Identification of a new caspase homologue: caspase-14.Cell Death Differ 1998;5:838-846.
4. Nicholson DW, Thornberry NA: Caspases: killer proteases. Trends BiochemSci 1997;22:299-306.
5. Slee EA, Adrain C, Martin SJ: Serial killers: ordering caspase activationevents in apoptosis. Cell Death Differ 1999;6:1067-1074.
6. Nuttall ME, Lee D, McLaughlin B, Erhardt JA: Selective inhibitors ofapoptotic caspases: implications for novel therapeutic strategies. Drug DiscovToday 2001;6:85-91.
7. Lee D, Long SA, Adams JL, Chan G, Vaidya KS, Francis TA, et al: Potent andselective nonpeptide inhibitors of caspase 3 and 7 inhibit apoptosis and main-tain cell functionality. J Biol Chem 2000;275:16007-16014.
8. Scott CW, Sobotka-Briner C, Wilkins DE: Novel small molecule inhibitors ofcaspase-3 block cellular and biochemical features of apoptosis. J PharmacolExp Ther 2003;304:433-440.
Okun et al.
284 www.sbsonline.org Journal of Biomolecular Screening 11(3); 2006
FIG. 7. (A) Three-dimensional structure of an unligated caspase-3 at 1.9 Å resolution.22 Both chain A and B are shown in magenta. (B) Three-dimen-sional structure of the caspase-3 in complex with AcDEVD-CHO peptide at 2.5 Å resolution.23 Chain A is shown in magenta, chain B is shown in blue, andpeptide is shown in brown. Active site and C-terminal cysteinyl moieties are shown in yellow in both panels.
A B

9. Lee D, Long SA, Murray JH, Adams JL, Nuttall ME, Nadeau DP, et al: Potentand selective nonpeptide inhibitors of caspase 3 and 7. J Med Chem
2001;44:2015-2026.
10. Schirmeister T, Kaeppler U: Non-peptidic inhibitors of cysteine proteases.Mini Rev Med Chem 2003;3:361-373.
11. Bamford M, Walkinshaw G, Brown R: Therapeutic applications of apoptosisresearch. Exp Cell Res 2000;256:1-11.
12. Ullman BR, Aja T, Deckwerth TL, Diaz JL, Herrmann J, Kalish VJ, et al:Structure-activity relationships within a series of caspase inhibitors: effect ofleaving group modifications. Bioorg Med Chem Lett 2003;13:3623-3626.
13. Perni RB, Pitlik J, Britt SD, Court JJ, Courtney LF, Deininger DD, et al: Inhib-itors of hepatitis C virus NS3.4A protease 2: warhead SAR and optimization.Bioorg Med Chem Lett 2004;14:1441-1446.
14. Westerfield M: The Zebrafish Book: A Guide for the Laboratory Use of
Zebrafish. Eugene, OR: University of Oregon Press; 1993.
15. Zhang JH, Chung TD, Oldenburgh KR: A simple statistical parameter for usein evaluation and validation of high throughput screening assays. J BiomolScreen 1999;4:67-73.
16. Kravchenko DV, Kysil VV, Ilyn AP, Tkachenko SE, Maliarchouk S, Okun I,et al: 1,3-dioxo-4-methyl-2,3-dihydro-1H-pyrrolo[3,4-c]quinolines as potentcaspase-3 inhibitors. Bioorg Med Chem Lett 2005;15/7:1841-1845.
17. Kravchenko DV, Kuzovkova YA, Kysil VM, Tkachenko SE, Maliarchouk S,Okun IM, et al: Synthesis and structure-activity relationship of 4-substituted2-(2-acetyloxyethyl)-8-(morpholine-1,3-diones as potent caspase-3 inhibi-tors. J Med Chem 2005;48:3680-3683.
18. Nicholson DW: Caspase structure, proteolytic substrates and function duringapoptotic cell death. Cell Death Diff 1999;6:1028-1042.
19. Nobel CSI, Kimland M, Nicholson DW, Orrenius S, Slater AFG: Disulfiramis a potent inhibitor of proteases of the caspase family. Chem Res Toxicol1997;10:1319-1324.
20. Wilson KP, Black J-AF, Thomson JA, Kim EE, Griffith JP, Navia MA, et al:Structure and mechanism of interleukin-1β converting enzyme. Nature
1994;370:270-275.
21. Hampton MB, Stamenkovic I, Winterbourn CC: Interaction with substratesensitizes caspase-3 to inactivation by hydrogen peroxide. FEBS Lett2002;517:229-232.
22. Ni C-Z, Li C, Wu JC, Spada AP, Ely KR: Conformational restrictions in theactive site of unliganded human caspase-3 [Online]. Pdb ID 1QX3. DepositedSeptember 04, 2003. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?CMD=search&DB=structure
23. Rotonda J, Becker JW: Crystal structure of the complex of apopain with thetetrapeptide aldehyde inhibitor Ac-Devd-Cho [Online]. Pdb ID 1PAU. De-posited June 06, 1996. Retrieved from http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?CMD=search&DB=structure
24. Micale N, Vairagoundar R, Yakovlev AG, Kozikowski AP: Design and syn-thesis of a potent and selective peptidomimetic inhibitor of caspase-3. J MedChem 2004;47:6455-6458.
25. Andreassen OA, Ferrante RJ, Hughes DB, Klivenyi P, Dedeoglu A, Ona VO,et al: Malonate and 3-nitropropionic acid neurotoxicity are reduced in trans-genic mice expressing a caspase-1 dominant-negative mutant. J Neurochem2000;75:847-852.
26. Miyashita T, Matsui J, Ohtsuka Y, Mami U, Fujishima S, Okamura-Ono Y, etal: Expression of extended polyglutamine sequentially activates initiator andeffector caspases. Biochem Biophys Res Commun 1999;257:724-730.
Address reprint requests to:Ilya Okun
11558 Sorrento Valley Rd.San Diego, CA 92121
E-mail: [email protected]
Novel Small-Molecule Caspase-3 Inhibitors
Journal of Biomolecular Screening 11(3); 2006 www.sbsonline.org 285




















