
Synaptic circuitry in the retinorecipient layers of the optic tectum of the lamprey...
28
ORIGINAL ARTICLE Synaptic circuitry in the retinorecipient layers of the optic tectum of the lamprey (Lampetra fluviatilis). A combined hodological, GABA and glutamate immunocytochemical study Jacques Repe ´rant Roger Ward Monique Me ´dina Natalia B. Kenigfest Jean-Paul Rio Dom Miceli Bruno Jay Received: 5 September 2008 / Accepted: 2 February 2009 Ó Springer-Verlag 2009 Abstract The ultrastructure of the retinorecipient layers of the lamprey optic tectum was analysed using tract tracing techniques combined with GABA and glutamate immunocytochemistry. Two types of neurons were identi- fied; a population of large GABA-immunonegative cells, and a population of smaller, highly GABA-immunoreac- tive interneurons, some of whose dendrites contain synaptic vesicles (DCSV). Five types of axon terminals were identified and divided into two major categories. The first of these are GABA-immunonegative, highly gluta- mate-immunoreactive, contain round synaptic vesicles, make asymmetrical synaptic contacts, and can in turn be divided into AT1 and AT2 terminals. The AT1 terminals are those of the retinotectal projection. The origin of the nonretinal AT2 terminals could not be determined. AT1 and AT2 terminals establish synaptic contacts with DCSV, with dendrites of the retinopetal neurons (DRN), and with conventional dendritic (D) profiles. The terminals of the second category are GABA-immunoreactive and can sim- ilarly be divided into AT3 and AT4 terminals. The AT3 terminals contain pleiomorphic synaptic vesicles and make symmetrical synaptic contacts for the most part with glu- tamate-immunoreactive D profiles. The AT4 terminals contain rounded synaptic vesicles and make asymmetrical synaptic contacts with DRN, with DCSV, and with D profiles. A fifth, rarely observed category of terminals (AT5) contain both clear synaptic vesicles and a large number of dense-core vesicles. Synaptic triads involving AT1, AT2 or AT4 terminals are rare. Our findings are compared to these of previous studies of the fine structure and immunochemical properties of the retinorecipient layers of the optic tectum or superior colliculus of Gnathostomes. Keywords Lamprey Optic tectum Synaptic connections Immunocytochemistry Abbreviations AOA Area optica accessoria AT1 Type 1 axon terminals AT2 Type 2 axon terminals AT3 Type 3 axon terminals AT4 Type 4 axon terminals AT5 Type 5 axon terminals AX2 Axon of type 2 terminal AXg GABA-immunoreactive axon BA Background immunolabelling CGL Corpus geniculatum laterale J. Repe ´rant R. Ward M. Me ´dina N. B. Kenigfest D. Miceli B. Jay De ´partement Re ´gulations, De ´veloppement et Diversite ´ Mole ´culaire, USM 501, CNRS UMR 7221, Muse ´um National d’Histoire Naturelle, Paris, France J. Repe ´rant R. Ward M. Me ´dina D. Miceli Laboratoire de Neuropsychologie Expe ´rimentale et Compare ´e, Universite ´ du Que ´bec, Trois-Rivie `res, Canada N. B. Kenigfest Laboratory of Evolution of Neuronal Interactions, Sechenov Institute, Russian Academy of Sciences, St. Petersburg, Russia J.-P. Rio INSERM U-616, Ho ˆpital de la Salpe ˆtrie `re, Paris, France M. Me ´dina (&) De ´partement RDDM, USM 501, CNRS UMR 7221, Muse ´um National d’Histoire Naturelle, CP 55, Ba ˆtiment d’Anatomie Compare ´e, 57 rue Cuvier, 75231 Paris Cedex 05, France e-mail: [email protected]; [email protected] 123 Brain Struct Funct DOI 10.1007/s00429-009-0205-9
-
Upload
acanthoweb -
Category
Documents
-
view
0 -
download
0
Transcript of Synaptic circuitry in the retinorecipient layers of the optic tectum of the lamprey...

ORIGINAL ARTICLE
Synaptic circuitry in the retinorecipient layers of the optic tectumof the lamprey (Lampetra fluviatilis). A combined hodological,GABA and glutamate immunocytochemical study
Jacques Reperant Æ Roger Ward Æ Monique Medina ÆNatalia B. Kenigfest Æ Jean-Paul Rio Æ Dom Miceli ÆBruno Jay
Received: 5 September 2008 / Accepted: 2 February 2009
� Springer-Verlag 2009
Abstract The ultrastructure of the retinorecipient layers
of the lamprey optic tectum was analysed using tract
tracing techniques combined with GABA and glutamate
immunocytochemistry. Two types of neurons were identi-
fied; a population of large GABA-immunonegative cells,
and a population of smaller, highly GABA-immunoreac-
tive interneurons, some of whose dendrites contain
synaptic vesicles (DCSV). Five types of axon terminals
were identified and divided into two major categories. The
first of these are GABA-immunonegative, highly gluta-
mate-immunoreactive, contain round synaptic vesicles,
make asymmetrical synaptic contacts, and can in turn be
divided into AT1 and AT2 terminals. The AT1 terminals
are those of the retinotectal projection. The origin of the
nonretinal AT2 terminals could not be determined. AT1
and AT2 terminals establish synaptic contacts with DCSV,
with dendrites of the retinopetal neurons (DRN), and with
conventional dendritic (D) profiles. The terminals of the
second category are GABA-immunoreactive and can sim-
ilarly be divided into AT3 and AT4 terminals. The AT3
terminals contain pleiomorphic synaptic vesicles and make
symmetrical synaptic contacts for the most part with glu-
tamate-immunoreactive D profiles. The AT4 terminals
contain rounded synaptic vesicles and make asymmetrical
synaptic contacts with DRN, with DCSV, and with D
profiles. A fifth, rarely observed category of terminals
(AT5) contain both clear synaptic vesicles and a large
number of dense-core vesicles. Synaptic triads involving
AT1, AT2 or AT4 terminals are rare. Our findings are
compared to these of previous studies of the fine structure
and immunochemical properties of the retinorecipient
layers of the optic tectum or superior colliculus of
Gnathostomes.
Keywords Lamprey � Optic tectum �Synaptic connections � Immunocytochemistry
Abbreviations
AOA Area optica accessoria
AT1 Type 1 axon terminals
AT2 Type 2 axon terminals
AT3 Type 3 axon terminals
AT4 Type 4 axon terminals
AT5 Type 5 axon terminals
AX2 Axon of type 2 terminal
AXg GABA-immunoreactive axon
BA Background immunolabelling
CGL Corpus geniculatum laterale
J. Reperant � R. Ward � M. Medina � N. B. Kenigfest �D. Miceli � B. Jay
Departement Regulations, Developpement et Diversite
Moleculaire, USM 501, CNRS UMR 7221,
Museum National d’Histoire Naturelle, Paris, France
J. Reperant � R. Ward � M. Medina � D. Miceli
Laboratoire de Neuropsychologie Experimentale et Comparee,
Universite du Quebec, Trois-Rivieres, Canada
N. B. Kenigfest
Laboratory of Evolution of Neuronal Interactions, Sechenov
Institute, Russian Academy of Sciences, St. Petersburg, Russia
J.-P. Rio
INSERM U-616, Hopital de la Salpetriere, Paris, France
M. Medina (&)
Departement RDDM, USM 501, CNRS UMR 7221,
Museum National d’Histoire Naturelle,
CP 55, Batiment d’Anatomie Comparee,
57 rue Cuvier, 75231 Paris Cedex 05, France
e-mail: [email protected]; [email protected]
123
Brain Struct Funct
DOI 10.1007/s00429-009-0205-9

CH Cerebral hemisphere
CV Centrifugal visual pathway
D Conventional dendrite
DCSV Dendrite of interneuron containing synaptic
vesicles
DCSV1 Dendrite of interneuron containing synaptic
vesicles of the first type
DCSV2 Dendrite of interneuron containing synaptic
vesicles of the second type
Dg? Conventional GABA-immunoreactive dendrite
of interneuron
Dg- Conventional GABA-immunonegative dendrite
of efferent neuron
DG? Conventional glutamate-immunoreactive
dendrite of efferent neuron
DG- Conventional glutamate-immunonegative
dendrite of interneuron
DRN HRP-labelled dendrite of the centrifugal
neurons of RMA
DRNg? HRP-labelled GABA-immunoreactive dendrite
of the centrifugal neurons of RMA
GABA c-Aminobutyric acid
HI Horizontal interneurons
HRP Horseradish peroxidase
HY Hypothalamus
INS Soma of interneuron
ir Immunoreactive
M5 Centrifugal visual neurons of the nucleus M5 of
Schober
MT Mesencephalic tegmentum
NP Nucleus preopticus
NPO Nucleus of the postoptic commissure
NPR Nucleus pretectalis
OB Olfactory bulb
OC Optic chiasma
OF Optic fibre
ON Optic nerve
PCSV Profiles containing synaptic vesicles
PNS Soma of projecting neuron
RI Radial interneurons
RITC Rhodamine b-isothiocyanate
RMA Centrifugal visual neurons of the reticular
mesencephalic area
RP Retinal projections
TO Tectum opticum
SCE Stratum cellulare ependimale
SCFC Stratum cellulare et fibrosum centrale
SCFE Stratum cellulare et fibrosum externum
SCFI Stratum cellulare et fibrosum internum
SCP Stratum cellulare periventriculare
SFP Stratum fibrosum periventriculare
SM Stratum marginale
Introduction
The Agnatha, or jawless vertebrates, belong to the most
ancient extant vertebrate group, which appeared over
500 million years ago (Forey and Janvier 1993; Janvier
1996). The living representatives of these ancestral verte-
brates fall into one of two groups, the Myxinidae (hagfish)
and Petromyzontidae (lampreys). In spite of their superfi-
cial resemblances, the hagfish and lampreys diverged early
in vertebrate phylogeny, and lampreys are now considered
to be more closely related to the Gnathostomes, or jawed
vertebrates, than to the hagfish (Forey and Janvier 1993;
Janvier 1996). In this light, authors are in general agree-
ment that the central nervous system of the lamprey can be
considered as the prototype of that of the Gnathostomes
(Heier 1948; Nieuwenhuys 1977; Karamian et al. 1984;
Nieuwenhuys and Nicholson 1998; Robertson et al. 2006,
2007). The numerous studies of the functional anatomical
organisation of the primary visual system of the lamprey,
carried out over several decades (Northcutt and Przybylski
1973; Kennedy and Rubinson 1977; Kosareva et al. 1977;
Rubinson and Kennedy 1979; Reperant et al. 1980, 1990;
Kosareva 1980; Vesselkin et al. 1980, 1996; Karamian
et al. 1984; de Miguel et al. 1990; Rubinson, 1990; Pierre
et al. 1992; Rio et al. 1993) are in agreement with this
hypothesis. In the lamprey, as in all Gnathostomes (cf.
Ebbesson 1970, 1972; Reperant et al. 1990), the retina
projects contralaterally to five distinct regions of the brain
(Fig. 1): the postoptic region (nucleus of the postoptic
commissure); the thalamus (corpus geniculatum laterale);
the pretectum (nucleus pretectalis); the tegmentum mes-
encephali (accessory optic area); and the superficial layers
of the tectum opticum. Of these, the retinotectal projection
is by far the most important, as it is in all nonmammalian
vertebrates, in particular the anamniotes (cf. Ebbesson
1970, 1972; Reperant et al. 1990). The organisation of this
projection in the lamprey has been studied by electro-
physiological methods (Karamian et al. 1984) and by a
variety of hodological techniques at the level of resolution
of the light microscope (Northcutt and Przybylski 1973;
Kennedy and Rubinson 1977; Kosareva et al. 1977;
Kosareva 1980; Reperant et al. 1980, 1990; Vesselkin et al.
1980, 1984, 1996; de Miguel et al. 1990; Rubinson 1990).
While the synaptic circuitry of the retinorecipient layers
of the optic tectum, or its mammalian equivalent, the
superior colliculus, has been extensively studied in all
major groups of Gnathostomes (for reviews: teleosts,
Vanegas et al. 1984; Peyrichoux et al. 1986; Kageyama and
Meyer 1989; amphibians, Lazar 1984; Antal 1991; Gabriel
and Straznicky 1995; reptiles, Reperant et al. 1997; Rio
et al. 1995; birds, Morino et al. 1991; Tombol 1998;
Tombol and Nemeth 1999; Tombol et al. 2003; mammals,
Brain Struct Funct
123

Mize 1992; Mize and Butler 1995; Mize et al. 1994; Ortega
et al. 1995) no comparable ultrastructural studies have been
carried out in the lamprey, with the exception of a pre-
liminary study by Rio et al. (1996).
The immediate aim of the present investigation was to
carry out a detailed analysis of the synaptic circuitry of the
retinorecipient layers of the optic tectum of the lamprey,
using a variety of morphological techniques: tract tracing,
immunocytochemistry, double and triple labelling, mor-
phometry, and stereology. These were intended to specify
the cytological, neurochemical and synaptological charac-
teristics of the profiles containing synaptic vesicles (PCSV)
and other neuronal profiles. We pay particular attention to
the following points: (1) the characterisation of retinal
terminals and their postsynaptic targets; (2) the same
properties of nonretinal terminals; (3) the possible exis-
tence of a projection of the centrifugal visual neurons onto
this structure, a hypothesis recently proposed by Robertson
et al. (2006); and (4) the identification of interneurons and
their dendrites containing synaptic vesicles (DCSV), of
their afferent supply and the targets of these profiles, and a
concomitant analysis of the c-aminobutyric acid (GABA)-
immunoreactive system. In an attempt to determine the
extent to which the details of the organisation of the reti-
norecipient tectal layers of this ancestral vertebrate form
are conserved in more recent Gnathostomes, we compare
our findings to those reported in the literature.
Materials and methods
A total of 64 adult specimens of Lampetra fluviatilis were
used. The animals were caught in October in the Neva
River (St. Petersburg, Russia) during their migration
upstream from the Baltic Sea. The animals, approximately
20 cm in length, were kept in aerated aquaria at a tem-
perature of 5–7�C; the experimental manipulation of these
animals followed European, Canadian and Russian guide-
lines for the treatment of experimental animals.
Degeneration experiments (n = 12)
Animals were anaesthetised in a 0.4% solution of tricaine
methane sulfonate (MS 222, Sandoz), and subsequently
received an injection of 20 ll of a 40% aqueous solution of
calcium chloride into the posterior chamber of the eye.
They were allowed to survive for 12–30 days and then,
under deep anaesthesia (MS 222), were perfused transcar-
dially with 20–30 ml of 0.7% saline followed by 100–
150 ml of a fixative composed of 1% paraformaldehyde
and 2.5% glutaraldehyde in 0.12 M phosphate buffer at
pH 7.4. The brains were dissected out and stored for 3.5 h
at room temperature in fresh fixative. Small blocks of tectal
tissue, cut transversely, were then post-fixed for 3–4 h in
2% buffered osmium tetroxide, stained en bloc in 2.5%
buffered uranyl acetate, dehydrated in a graded series of
ethanol and embedded in Araldite. Semithin (1 lm) sec-
tions were cut and stained with toluidine blue. Ultrathin
(70–80 nm) sections were cut on a Reichert OM U2
ultramicrotome, mounted on copper grids, double-stained
with ethanolic uranyl acetate and lead citrate, and exam-
ined under a Philips EM 400 electron microscope. The
same procedure was used in preparing tissue for electron
microscopy in other experiments.
Axonal tracing experiments (n = 20)
Animals (n = 16) were anaesthetised by flooding the gills
with a 0.3% solution of urethane. After ablation of one or
two eyes, each severed optic nerve was gently exposed and
NPR
SM +SCFE
TO
NP
CGL
AOA
ON
M5RMA
OC
RP
CV
MT
OBCH
HY
DRN
DRN
DRN
NPO
Fig. 1 A schematic drawing of
a longitudinal section of the
brain of Lampetra fluviatilisindicating the retinal projection
(RP) and the centrifugal visual
pathway (CV) visualised after
either iontophoretic deposit of
HRP into the contralateral optic
nerve or intraocular injection of
RITC into the contralateral eye
Brain Struct Funct
123

isolated from the oculomotor nerves which were covered
with paraffin oil. A glass micropipette with a tip diameter
of 20–50 lm, filled with a 4% solution of horseradish
peroxidase (HRP; either Sigma Type VI or Boehringer
Grade I) in 0.1 M PB at pH 8.0, was placed in contact with
the sectioned right optic nerve and a positive current of
10 lA was passed for 20–30 min at a rate of 7 s on/7 s off.
In some cases the same procedure was then carried out on
the left optic nerve. The animals were reanimated in oxy-
gen-saturated water, and allowed to survive for 4–16 days.
They were then anaesthetised with MS 222 and perfused
transcardially, using the procedure described above. Thick
(50–80 lm) transverse sections of the optic tectum were
cut on an Oxford Vibratome and collected in 0.1 M Tris
buffer at pH 7.6. HRP was visualised either by the method
of Adams (1981), using cobalt-nickel intensified 3,30
diaminobenzidine (DAB) or by Mesulam’s (1982) tetra-
methylbenzidine (TMB) procedure. After the reaction,
some sections were mounted directly on glass slides for
examination by light microscopy, while others were pre-
pared for electron microscopy by the methods described
above. Under anaesthesia four other animals received a
unilateral intraocular injection, into either the left or right
eye, of 10 ll (w/v) of rhodamine b-isothiocyanate (RITC,
Sigma-Aldrich) containing 2% dimethyl sulfoxide. The
injections were performed with the lamprey maintained in
a stereotaxic apparatus, using a Hamilton syringe with a
maximum capacity of 20 ll. The solution was injected
slowly into the posterior chamber of the eye, as close to the
retina as possible but avoiding direct contact of the needle
with this structure. Survival time ranged from 10 to
18 days. Following perfusion the brains were cryostat
sectioned in the transverse plane at a thickness of 20 lm.
The sections were immediately mounted in slides,
coverslipped and viewed using a Leitz Ploemopack fluo-
rescence microscope fitted with an N2 filter (barrier filter
580 nm).
Combined degeneration and HRP tracing experiments
(n = 12)
Six to 12 days after the injection of calcium chloride into
the posterior chambers of each eye, the eyeballs were
removed under urethane anaesthesia and HRP was applied
to each optic nerve by the iontophoretic procedure descri-
bed above. The lampreys were allowed to survive for 6–
25 days and then perfused; the methods described above
were used both for fixation and revelation of HRP activity
in sections of the optic tectum, and also for the preparation
of sections for electron microscopy.
In addition, four intact lampreys were prepared for
examination by light microscopy, and a further six for
electron microscopy. The brains prepared for light
microscopy were embedded in paraffin, serially sectioned
at 10 lm and stained with cresyl violet.
Immunocytochemical methods
GABA immunofluorescence procedures
(normal material, n = 6)
The animals were perfused transcardially with 20–30 ml of
0.7% saline followed by 150 ml of a fixative composed of
4% paraformaldehyde and 0.5% glutaraldehyde in 0.1 M
phosphate buffer at pH 7.4. The brains were subsequently
removed and stored in fresh fixative for 2 h at 4�C, then
transferred to a 30% solution of sucrose in phosphate
buffer, in which they remained overnight at the same
temperature. Transverse sections of the mesencephalon
were cut at 40–50 lm in a freezing microtome. They were
incubated for 48 h at 4�C in a primary rabbit anti-GABA
antibody (Immunotech, Marseille, France) diluted 1:3,000
in phosphate buffer saline (PBS). The sections were sub-
sequently incubated for 2 h in a fluorescein-conjugated
(FITC) goat anti-rabbit IgG diluted 1:50 in PBS, rinsed in
PB, mounted on slides and coverslipped with Vectashield
(Vector Laboratories, Temecula CA, USA). Sections were
observed under a Leitz Ploemopack fluorescence micro-
scope fitted with I3 (448 nm) filter mirror systems to detect
the FITC fluorescence of GABAergic neurons. Labelled
profiles in serial sections were counted either directly under
the microscope or on a monitor screen driven by a Pana-
sonic WV1900 video camera.
GABA or glutamate pre-embedding procedure
(normal material, n = 4)
After perfusion under urethane anaesthesia with the same
dialdehyde fixative as that used in the HRP experiments,
slabs of tectal tissue from normal animals were serially
sectioned at 40–50 lm in the transverse plane in a
Vibratome and processed for GABA or glutamate-immu-
nocytochemistry. Sections were collected in cold 0.1 M
PBS, and preincubated for 30 min in 0.1 M PBS con-
taining 0.2% gelatine, 0.1% Triton-X 100 and 0.1 M
lysine. After several rinses in PBS the sections were
incubated overnight in a cold chamber with a humid
atmosphere in a 1:2,000 dilution of an rabbit anti-GABA
(Immunotech, France) or a 1:300 dilution of rabbit anti-
glutamate (Chemicon Temecula CA, USA) antibodies
containing 0.2% gelatin and 0.1% Triton-X 100; sections
were then rinsed in PBS and incubated for 1 h in a 1:100
dilution of a goat anti-rabbit immunoglobulin (Chemicon,
USA), rinsed again in PBS and incubated for 1 h in a
1:200 dilution of a rabbit avidin–biotin–HRP complex
Brain Struct Funct
123

(Vectastain Elite, Vector Laboratories, USA). Peroxidase
activity was visualised with 0.03% DAB (Sigma, USA)
and 0.005% hydrogen peroxide in 0.01 M Tris–HCl buffer
at pH 7.6. Sections were then embedded in Araldite.
The GABA and glutamate-immunoreactive elements of
the tissue were then examined by optical microscopy in
semithin (B1 lm) sections counterstained with toluidine
blue, or under the electron microscope in thin (70–80 nm)
sections.
GABA or glutamate post-embedding procedure
(normal, n = 6 and experimental material:
HRP, n = 20; HRP ? degeneration, n = 12)
Light microscopy
Semithin sections throughout the optic tectum were
mounted on gelatin-coated slides, etched in saturated
sodium ethoxide, treated with 10% hydrogen peroxide, and
preincubated for 30 min in 10% bovine serum albumin
(BSA) in PBS containing 0.2% gelatin, 0.25% Triton-X
100, and 0.1 M lysine prior to being incubated overnight at
4�C, in a humid atmosphere, in a 1:5,000 dilution of rabbit
anti-GABA antiserum (Immunotech, Marseille, France)
diluted in PBS containing 0.2% gelatin, 0.25% Triton-X
100, 0.1% lysine, and 1% BSA. Sections were subse-
quently rinsed in PBS and incubated for 1 h in a 1:100
dilution of a goat anti-rabbit antiserum, rinsed again in
PBS, and then incubated for 1 h in a 1:200 dilution of a
rabbit peroxidase–antiperoxidase complex, following the
technique of Sternberger et al. (1970). After several rinses
in PBS the peroxidase activity was visualized with 0.03%
DAB and 0.005% hydrogen peroxide in 0.1 M Tris–HCl
buffer. Sections were coverslipped, and examined under
the microscope.
Electron microscopy
Thin sections were gathered onto nickel grids and immersed
in the following solutions: (1) 1% aqueous periodic acid,
7 min; (2) rinse in distilled water; (3) 1% sodium metape-
riodate, 7 min; (4) rinse in distilled water; (5) 5% BSA in
0.05 M Tris-buffered saline (TBS) at pH 7.6, 30 min; and
(6) rinse in TBS. Sections were then incubated overnight at
4�C in one of two primary antibodies. These were either
rabbit anti-glutamate (Chemicon, Temecula CA, USA)
diluted 1/300 in TBS at pH 7.6, or rabbit anti-GABA
(Immunotech, France) diluted 1/2,000 in TBS at pH 7.6.
After several rinses in TBS, the sections were treated with a
goat anti-rabbit immunoglobulin (Chemicon) conjugated
with 10, 15 or 30 nm colloidal gold particles (Janssen,
Belgium), diluted 1/75 in TBS at pH 8.2, for 1 h at room
temperature. The sections were then rinsed in TBS and
distilled water, and counterstained with uranyl acetate and
lead citrate prior to examination under the electron micro-
scope. Some sections were also processed for double
immunolabelling; after incubation in the rabbit anti-gluta-
mate antibody and the second antibody conjugated with 30
or 10 nm gold particles, the sections were exposed to the
vapour of 4% paraformaldehyde at 80�C for 1 h (Wang and
Larsson 1985) thoroughly rinsed in PBS and then incubated
in the anti-GABA antibody followed by the secondary
antibody conjugated with 10 or 20 nm particles.
Specificity of antibodies and control procedures
The two polyclonal antibodies used in this study have
already been characterised in previous investigations;
the anti-glutamate antibody (Chemicon) by Sherry and
Ulshafer (1992) and Kalloniatis and Fletcher (1993), and
the anti-GABA antibody (Immunotech) by Seguela et al.
(1984) and Miceli et al. (2008); they were generated in
rabbit against glutamate or GABA coupled to BSA with
glutaraldehyde. The cross-reactivities were determined
using ELISA or RIA techniques. It has been shown that
each antibody shows a high specificity for the particular
haptene (glutaraldehyde-fixed glutamate, or glutaralde-
hyde-fixed GABA) and negligible cross-reactivity with
the other haptene of the pair, or with other amino acids
(L-aspartate, L-glutamate, taurine and glycine for the anti-
GABA antibody, L-aspartate, taurine, glycine and GABA
for the anti-glutamate antibody).
In the present study, in each immunolabelling session,
control grids were tested for any cross-reactivity with
glutamate and GABA. Lamprey brain macromolecules
were treated with glutaraldehyde either alone or in the
presence of one of these amino acids. The conjugates thus
formed were embedded and processed according to tech-
nique developed by Ottersen (1987, 1989), Ottersen and
Storm-Mathisen (1984) and Ottersen (1990); sections of
conjugates then underwent the same immunogold proce-
dure as sections of tectal tissue. Electron micrographs were
printed at the same magnification as that used for tissue
sections, and the density of gold particles determined (see
below).
The results obtained with the conjugated material
revealed a high degree of selectivity of each antiserum.
Thus, for the sections of glutamate conjugate, a mean
density of 3,172 ± 283 (n = 12) particles/lm2 was
observed when they were treated with the anti-glutamate
antibody, and only 108 ± 10 (n = 14) particles/lm2 when
treated with the anti-GABA antibody. Conversely, the
immunoprocessing of sections of the GABA conjugate led
to a mean density of 2,892 ± 304 (n = 12) particles/lm2
for the anti-GABA antibody, a value to be compared to the
mean density of 2.9 ± 1.2 particles/lm2 obtained with the
Brain Struct Funct
123

anti-glutamate antibody. Additional controls were made by
either omitting the primary antibody or replacing it with
non-immune normal serum, or by immunoadsorbtion of
free glutamate (for the anti-glutamate antibody) or of free
GABA (for the anti-GABA antibody) on polyacrylamide
beads with glutaraldehyde as a linking agent. Under none
of these conditions was any labelling ever observed. The
specificity of the second antibody was demonstrated by the
complete absence of labelling observed after omission of
the primary antibody.
Quantitative methods
Light microscopy
Video images of GABA-immunoreactive cells in sections
treated either for immunofluorescence or by pre and post-
embedding immunocytochemistry were observed and
mapped directly on the monitor screen; cell counts were
performed using NIH Image software. The GABA-reac-
tive cells of the dorsolateral region of the retinorecipient
layers of the optic tectum were counted within a
263 9 263 lm2 (i.e. an area of 69,169 lm2); cells tran-
sected by either the bottom or the right-hand side of this
square were not included in the count (Gundersen 1978;
Williams and Rakic 1988). The sizes of the somata were
estimated by measuring the longest and shortest axes of
the cell body and converting these to mean circular
diameters. The relative proportions of GABA-immunore-
active and GABA-immunonegative cells were estimated
in semithin sections of pre and post-embedded material by
counting the numbers in each category. Simple counting
methods yield biased estimates (cf. Williams and Rakic
1988; Guillery 2002); in our material the larger size of
GABA-negative cells led to an overestimate of their
number in comparison to that of the smaller GABA-
reactive neurons. However, since the mean cell body
diameters were known, as well as the thickness of the
tectal sections, we applied Abercrombie’s (1946) correc-
tion to the raw data in order to obtain unbiased estimates
of cell densities, expressed as cells/mm3, and the relative
proportions of each cell type.
Electron microscopy
The quantitative analysis was carried out on a total area of
6,500 lm2 of sections derived from the dorsolateral
superficial layers (SM and SCFE) of the central portion of
the optic tectum (Fig. 2c).
For each category of the profiles (axons, terminals, con-
ventional dendrites, DCSV), a sample of 40 randomly chosen
profiles was analysed to estimate various morphometric
parameters; the area of the profile and that of its synaptic
vesicles, the length of the synaptic differentiation, and the
density of immunogold particles lying over the profile.
The cross-sectional areas of dendritic and axon terminal
profiles, and the lengths of the synaptic zones, were
measured in electron micrographs printed at a final magni-
fication of 28,0009 or 46,0009, by means of the NIH Image
software package. Areas (a) and perimeters (p) of synaptic
vesicles were measured in micrographs printed at a final
magnification of 160,0009 by means of the Optilab image
analysis system. From these a dimensionless shape factor
was calculated as:
S ¼ 4pa=p2
A value of 1.0 for this factor indicates a perfect circle,
lower values indicating progressively elongated ellipsoids.
The density of immunogold particles (either GABA-ir or
Glu-ir) over different profiles was measured in micrographs
printed at a final magnification of 46,0009; the area of
each profile (axon terminals, dendrites containing synaptic
vesicles or DCSV, conventional dendrites, cell bodies) was
measured as above, and the density expressed as particles/
lm2 uncorrected for background. The same technique was
used to estimate the background intensity of labelling,
counts in this case being made over 100 lm2 areas of
empty resin or the lumina of blood vessels. Estimates of the
background intensity were made in randomly selected
micrographs of 36 sections of tissue.
All the numerical results are presented as mean val-
ues ± standard deviation; statistical analyses were made
by Student’s test or analyses of variance.
In order to estimate the relative proportions of each of
the different types of profile observed in the retinorecipient
layers of the dorsolateral tectum, we carried out a stere-
ological analysis using the point counting method
(Mayhew 1979; Underwood 1970; Weibel 1979). A 6 cm2
grid was randomly positioned on each of 30 electron
micrographs printed at a final magnification of 28,0009;
the profile underlying each of the 36 points of the grid was
noted as a conventional dendrite (noting the presence or
absence of HRP-labelling, GABA- or glutamate-immuno-
reactivity, or a combination of these), an axon (similarly
annotated) a PCSV (AT1, AT2, AT3, AT4, AT5 or DCSV),
these being defined by cytological, hodological and
immunocytochemical criteria; see below), a neuronal cell
body or a glial cell; a total of 1,080 points was counted.
Results
Light microscopy: general morphology
The optic tectum is a prominent external feature of the
adult lamprey brain. The two halves of the tectum bulge
Brain Struct Funct
123
laterally, forming a deep lateral tectal recess. The resulting
tectal ventricle is roofed over by the choroidal lamina. The
structure of the tectum has been extensively described
(Johnston 1902; Ariens-Kappers et al. 1936; Heier 1948;
Leghissa 1962; Schober 1964; Pfister 1971; Nieuwenhuys
1977; Kennedy and Rubinson 1977, 1984; Rubinson and
Kennedy 1979; Nieuwenhuys and Nicholson 1998; Iwahori
et al. 1999). These authors all agree in recognising a well-
developed tectal lamination, but differences of nomencla-
ture and the use of different histological techniques have
led them to disagree over the number, from six to nine, of
tectal laminae. In this investigation we have chosen the
nomenclature proposed by Kennedy and Rubinson (1984),
since it has the advantage of being based on the study of
the development of the tectal lamination. In this termi-
nology, the following seven layers are described from the
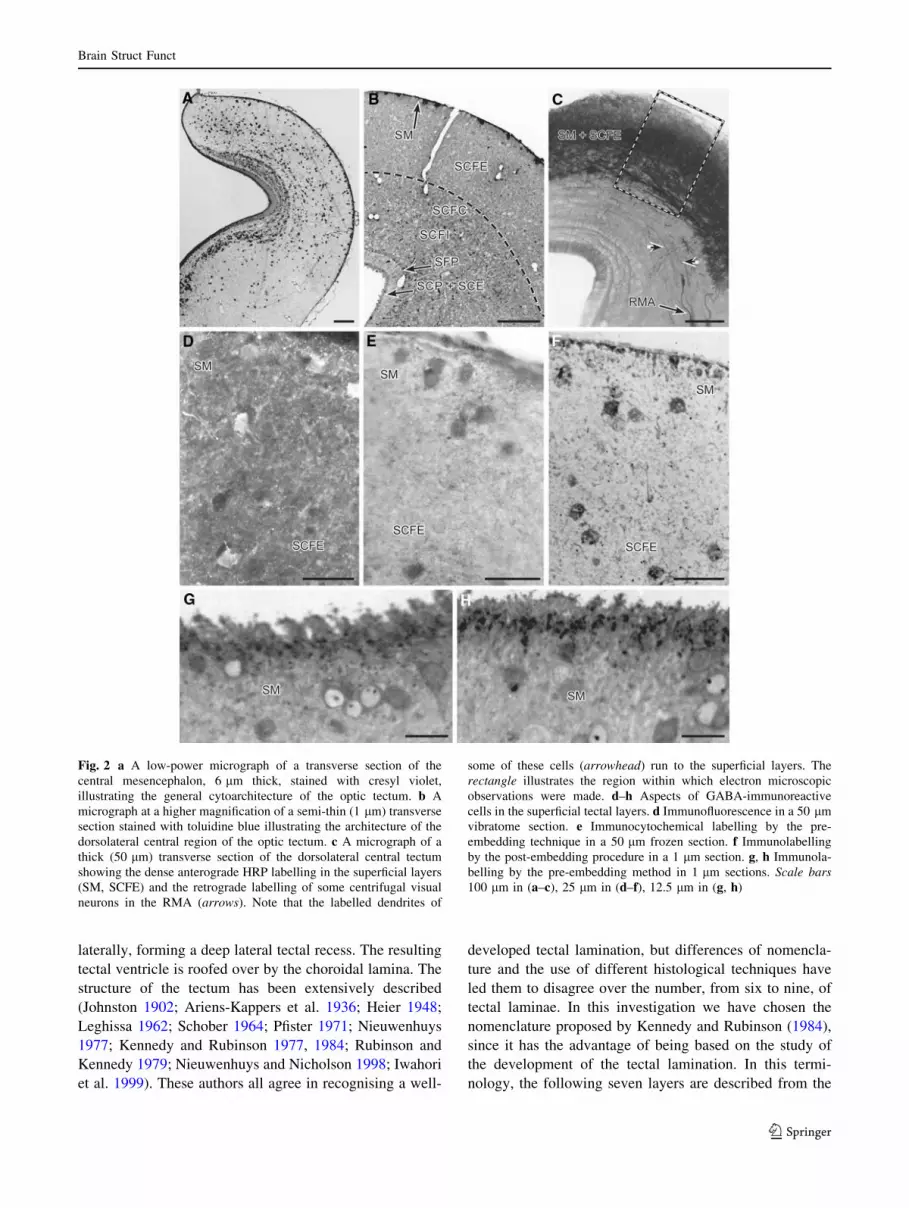
Fig. 2 a A low-power micrograph of a transverse section of the
central mesencephalon, 6 lm thick, stained with cresyl violet,
illustrating the general cytoarchitecture of the optic tectum. b A
micrograph at a higher magnification of a semi-thin (1 lm) transverse
section stained with toluidine blue illustrating the architecture of the
dorsolateral central region of the optic tectum. c A micrograph of a
thick (50 lm) transverse section of the dorsolateral central tectum
showing the dense anterograde HRP labelling in the superficial layers
(SM, SCFE) and the retrograde labelling of some centrifugal visual
neurons in the RMA (arrows). Note that the labelled dendrites of
some of these cells (arrowhead) run to the superficial layers. The
rectangle illustrates the region within which electron microscopic
observations were made. d–h Aspects of GABA-immunoreactive
cells in the superficial tectal layers. d Immunofluorescence in a 50 lm
vibratome section. e Immunocytochemical labelling by the pre-
embedding technique in a 50 lm frozen section. f Immunolabelling
by the post-embedding procedure in a 1 lm section. g, h Immunola-
belling by the pre-embedding method in 1 lm sections. Scale bars100 lm in (a–c), 25 lm in (d–f), 12.5 lm in (g, h)
Brain Struct Funct
123

external surface to the ventricle: (1) the stratum marginale
(SM); (2) the stratum cellulare et fibrosum externum
(SCFE); (3) the stratum cellulare et fibrosum centrale
(SCFC); (4) the stratum cellulare et fibrosum internum
(SCFI); (5) the stratum fibrosum periventriculare (SFP); (6)
the stratum cellulare periventriculare (SCP); and (7) the
stratum cellulare ependimale (SCE) (Fig. 2a, b).
Simple inspection of 10 lm paraffin sections stained
with cresyl violet (Fig. 2a) or of semithin (1 lm) sections
counterstained with toluidine blue (Fig. 2b), of the normal
optic tectum shows that the majority of tectal cell bodies is
concentrated in the deeper tectal layers (SCFI and SCP in
particular), whereas these cells are considerably rarer in the
superficial layers wherein they are widely dispersed.
HRP and RITC experiments
After application of HRP to the central stump of the optic
nerve, examination of vibratome sections revealed an
extremely dense anterograde labelling of the entire extent
of the primary visual system, in particular the superficial
layers (SM, SCFE) of the optic tectum (Fig. 2c), covering
approximately half of the tectal thickness. Optimal results
were obtained after a survival time of 4–6 days following
the iontophoretic application of the tracer and 10 days after
intraocular injection of RITC. The retinotectal fibres are
organised as the lateral optic tract, arising from the mar-
ginal optic tract; these densely packed fibres penetrate the
lateral aspect of the medial part of the SCFE in a fan-like
fashion. Collateral branches of these axons arborise within
the SM and throughout the entire thickness of the SCFE;
the retinotectal projection is predominantly contralateral,
but with a modest ipsilateral component which principally
concerns the lateroventral and mediodorsal regions of the
SM and SCFE. Comparable results were obtained after
intraocular injection of RITC.
The iontophoretic application of HRP to the stump or
intraocular injection of RITC of the optic nerve also led to
a bilateral, but predominantly contralateral, retrograde
labelling of the centrifugal visual neurons situated in the
nucleus M5 of Schober and in the reticular mesencephalic
area (RMA). The retinopetal neurons in the RMA were
mainly located in the dorsal tegmental region of the mid-
brain, but some were located in the SCFI of the
lateroventral region of the optic tectum (Fig. 2c). The
labelled cell bodies in M5 were pyramidal in shape, and
larger than those observed in RMA, which generally
appeared fusiform, or rarely multipolar. The dendrites of
the RMA retinopetal neurons are mainly oriented towards
the optic tectum; these labelled dendrites follow a latero-
medial path, reaching the deepest layers of the tectum
through its ventrolateral flank. They then cross the inter-
mediate layers (SCFI, SFC) extending finally into the
dorsolateral zone of optic terminal labelling (SCFE, SM;
Fig. 2c). These observations are entirely comparable to
those made after intraocular injection of RITC (Fig. 3). In
those specimens which received HRP after an intraocular
injection of calcium chloride, no orthograde labelling of
the primary visual system, including the retinotectal path-
way, was observed. However, the retrograde labelling of
the neurons of the centrifugal visual system in M5 and
RMA corresponded to that described above.
GABA immunofluorescence and immunocytochemistry
The general distribution of GABA-immunoreactive somata
in the SM and SCFE of the optic tectum was clearly visible
in frozen immunofluorescence and semithin pre-embedded
or post-embedded sections (Fig. 2d–h). The two layers
contained small GABA-ir cell bodies with a mean equiva-
lent diameter of 8.9 ± 2.32 lm (n = 66); a few of these
had fusiform somata oriented parallel to the tectal surface,
while the majority had vertically oriented pear-shaped
somata. The GABA-immunonegative cell bodies, observed
in semi-thin sections, were larger in size with a mean
diameter of 11.8 ± 3.44 lm (n = 57). In the neuropil
numerous GABA-ir puncta were also observed, represent-
ing axon terminals and dendritic shafts. Cell counts of
GABA-ir somata in pre- and post-embedded semithin sec-
tions of the dorsolateral part of the superficial layers of the
central region of the optic tectum, corrected by Aber-
crombie’s (1946) method, indicated their mean packing
density to be 684 ± 203 cells/mm3, and that they accounted
for 50.3% of the total population of cells in this region.
Electron microscopy
Examination of the ultrathin sections revealed that the SM
and SCFE of the dorsolateral part of the central region of
Fig. 3 Micrograph of a transverse frozen section, 25 lm thick, of
the midbrain of the lamprey showing the retrograde labelling of
centrifugal visual neurons in M5 and RMA after contralateral
intraocular injection of RITC. The double arrow indicates the
mediolateral axis and the single arrow the dorsoventral axis. Scalebar 200 lm
Brain Struct Funct
123

the tectum contained several types of profile containing
synaptic vesicles (PCSV), dendritic profiles, unmyelinated
axons, and occasional profiles of glial cells and neurons
(Fig. 4a).
General criteria used in the identification of profiles
Several criteria were used to classify the profiles observed in
the superficial layers of the central optic tectum: the presence
or absence of tracer labelling or signs of degenerative
change, and the presence or absence of immunoreactivity to
glutamate or GABA. In addition, other criteria were used to
classify the axonal profiles and DCSV; the size, shape, ori-
entation and position (presynaptic or postsynaptic) of the
profiles; the relative density of the hyaloplasm; the presence
of microtubules, microfilaments and ribosomes; the number
of mitochondria; and the shape of the vesicles (rounded or
flattened), together with their size, distribution and density.
The final criterion was the type of synaptic junction: on
the one hand, those described as Type I (Gray 1959) or
asymmetrical (Colonnier 1968), with a synaptic cleft
approximately 30 nm wide, with a well-developed synaptic
grid and a slight degree of postsynaptic differentiation; and
on the other hand those described as Type II (Gray 1959) or
Fig. 4 a A low-power
electromicrograph showing
HRP-labelled (DAB method)
optic fibres (OF) and their AT1terminals in the central region of
the SCFE. b An optic fibre (OF)
making synaptic contact enpassant (arrow) with a dendrite
profile (D). c An unmyelinated
HRP-labelled (TMB method)
optic fibre (OF) showing a high
degree of glutamate-
immunoreactivity (post-
embedding technique, 10 nm
gold particles). d A triple-
labelled preparation showing an
HRP-labelled (TMB method),
glutamate-immunoreactivity,
OF (post-embedding technique,
10 nm gold particles) and
GABA-immunoreactive AT3
terminals (30 nm gold
particles). Note the presence of
many 10 nm gold particles,
indicating glutamate-
immunoreactivity, over the
mitochondria of the AT3
terminals. Scale bars 2 lm in
(a), 1 lm in (c), 0.5 lm in
(b, d)
Brain Struct Funct
123

symmetrical (Colonnier 1968), in which the synaptic cleft is
approximately 18 nm wide, and with a considerably poorer
degree of postsynaptic differentiation.
General considerations of GABA- and glutamate-
immunoreactivity
The levels of background immunogold labelling were
0.94 ± 0.20 particles/lm2 for the anti-GABA antibody
(Fig. 13), and 1.38 ± 0.62 particles/lm2 for the anti-gluta-
mate antibody (Fig. 12). We do not subtract these values
from those presented below concerning the immunoreac-
tivity of the PCSV, conventional dendrites and neuronal
somata.
The density of immunolabelling of the two categories
of dendritic profiles, not containing synaptic vesicles,
(conventional dendrites: D), postsynaptic to the various
categories of terminal that we describe below was, for the
first, 23.2 ± 4.5 particles/lm2 for the anti-glutamate anti-
body (Fig. 12) and for the second 17.4 ± 2.2 particles/lm2
for the anti-GABA antibody (Fig. 13) (n = 40 for each).
The analysis of double immuno-labelled preparations
(Figs. 4d, 7a) treated with anti-glutamate and anti-GABA,
showed that the GABA-ir terminals (AT3, AT4, see below)
and GABA-ir (DCSV, see below) were also moderately
glutamate-immunoreactive (8.03 ± 0.7 particles/lm2 for
AT3 and AT4, n = 37, and 8.43 ± 1.2 particles/lm2 for
DCSV, n = 21) (Fig. 12). A more detailed analysis
revealed that these particles were generally situated over
mitochondria (Fig. 7a), while in GABA-negative, strongly
glutamate-ir profiles (AT1, AT2, see below) the gold par-
ticles were more concentrated over the synaptic vesicles
(Figs. 7, 8b, e).
Profiles containing synaptic vesicles (PCSV)
Six categories of PCSV could be distinguished on the
basis of several cytological; and immunochemical criteria.
Five of these correspond to axon terminals (AT) and the
sixth to DCSV. Of a total of 430 PCSV seen to make
synaptic contact, we were able to estimate that 91% of
these were axo-dendritic contacts with spines or den-
dritic shafts which sometimes contained synaptic vesicles,
8.9% were dendro-dendritic contacts, and 1.1% were axo-
somatic. We also point out the total absence of myelin-
ated axons.
Axon terminals
AT1. These correspond to retinal terminals, since they are
consistently labelled with tracer after iontophoretic
deposit of HRP into the optic nerve, or show degenerative
changes after intraocular injection of calcium chloride.
All the other types of PCSV, which we describe below,
were unlabelled and showed no signs of degenerative
change.
Many HRP-labelled (Figs. 4, 5d–g, 6, 7) or degenerating
profiles (Fig. 5a–c) were observed in the superficial reti-
norecipient layers of the tectum. These were unmyelinated
visual fibres and their terminals. The visual fibres were
irregularly shaped (Fig. 4a), displaying numerous neuro-
filaments, few microtubules, and occasional agranular
synaptic vesicles (Fig. 4b). These highly glutamate-ir fibres
(42.6 ± 3.9 particles/lm2, Fig. 4d) occasionally made
synaptic contact en passant with small, glutamate-ir or
GABA-ir dendrites. The diameter of these fibres varied
from 0.54 to 4.2 lm, about 60% of them having diameters
between 1 and 3 lm. In lampreys having received intra-
ocular calcium chloride, the retinal terminals showed a
variety of degenerative changes (Fig. 5a–c). The first sign
of degeneration was a swelling of the synaptic vesicles,
followed by a progressive darkening of the axoplasm in
which the organelles were more and more disrupted. In
those cases involving both degeneration and HRP tracing
techniques, we never observed HRP-labelled terminals in
the superficial layers of the optic tectum, but exclusively
labelled dendrites arising from the centrifugal visual neu-
rons of the RMA.
The AT1 terminals, with a mean cross-sectional area of
0.96 ± 0.09 lm2 and an irregular and sometimes scalloped
outline, were densely packed with round agranular synaptic
vesicles with a mean size of 1,340 ± 74 nm2 and a shape
index of 0.90 ± 0.10 (n = 420) (Fig. 11). The terminals
were often gathered into clusters (Fig. 4a), and were
GABA-immunonegative (Fig. 7a) their density of labelling
(0.98 ± 0.15 particles/lm2) (Fig. 12) being close to that of
the background; on the other hand, they showed strong
glutamate-immunoreactivity (43.8 ± 2.10 particles/lm2,
compared to a background of 1.38 ± 2.3 particles/lm2,
n = 33) (Figs. 7, 13). Their active zone is small (0.23 ±
0.04 lm). These terminals make asymmetrical synaptic
contact, their postsynaptic targets being either DCSV,
(19% of cases) (Figs. 5b–d, f, g, 6a–c, 7b), the distal
dendritic segments of HRP-labelled retinopetal neurons
of the RMA (7% of cases) (Fig. 5e), or conventional
dendrites unlabelled by HRP (74% of cases) (Fig. 5d).
About one-third of these latter profiles are GABA-immu-
noreactive (17.4 ± 2.2 particles/lm2), the remainder being
glutamate-positive (23.2 ± 4.5 particles/lm2, Fig. 5d).
The HRP-labelled dendrites of the centrifugal visual neurons,
which never contain synaptic vesicles, are either glutamate-
immunoreactive or GABA-immunoreactive (Fig. 5e). The
DCSV, which are of two types, are consistently GABA-ir
(see below). The AT1 retinal terminals represented 31.3% of
the total population of PCSV in the dorsolateral retinore-
cipient layers of the contralateral optic tectum.
Brain Struct Funct
123
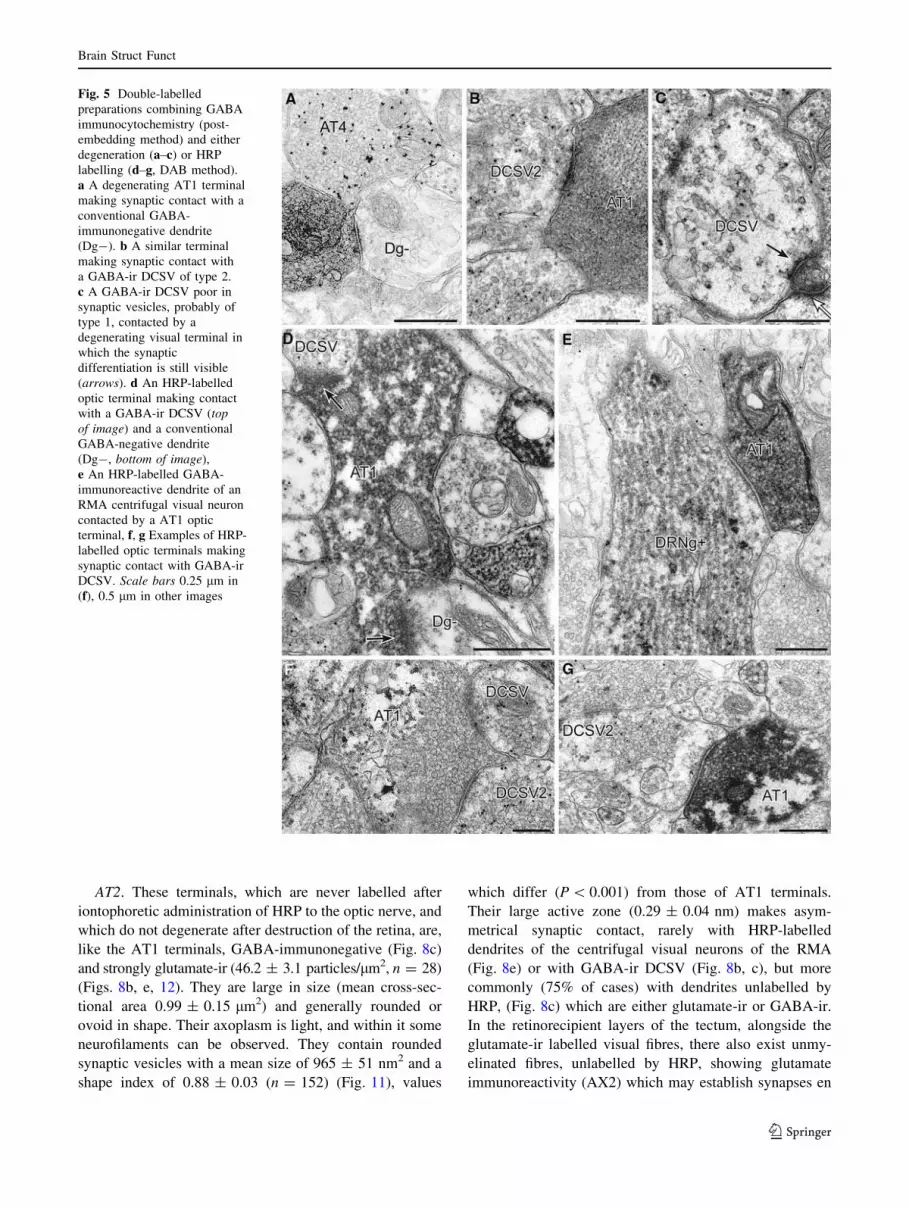
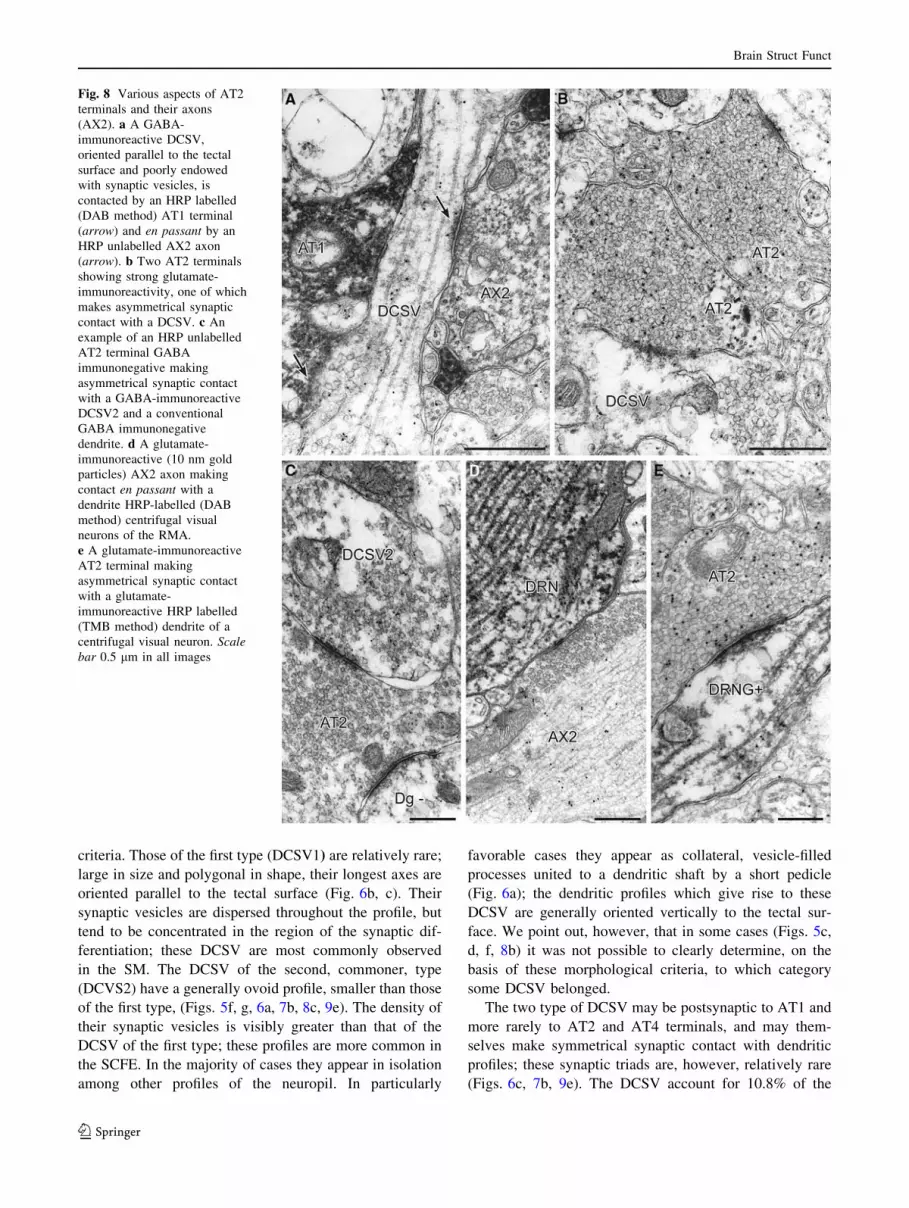
AT2. These terminals, which are never labelled after
iontophoretic administration of HRP to the optic nerve, and
which do not degenerate after destruction of the retina, are,
like the AT1 terminals, GABA-immunonegative (Fig. 8c)
and strongly glutamate-ir (46.2 ± 3.1 particles/lm2, n = 28)
(Figs. 8b, e, 12). They are large in size (mean cross-sec-
tional area 0.99 ± 0.15 lm2) and generally rounded or
ovoid in shape. Their axoplasm is light, and within it some
neurofilaments can be observed. They contain rounded
synaptic vesicles with a mean size of 965 ± 51 nm2 and a
shape index of 0.88 ± 0.03 (n = 152) (Fig. 11), values
which differ (P \ 0.001) from those of AT1 terminals.
Their large active zone (0.29 ± 0.04 nm) makes asym-
metrical synaptic contact, rarely with HRP-labelled
dendrites of the centrifugal visual neurons of the RMA
(Fig. 8e) or with GABA-ir DCSV (Fig. 8b, c), but more
commonly (75% of cases) with dendrites unlabelled by
HRP, (Fig. 8c) which are either glutamate-ir or GABA-ir.
In the retinorecipient layers of the tectum, alongside the
glutamate-ir labelled visual fibres, there also exist unmy-
elinated fibres, unlabelled by HRP, showing glutamate
immunoreactivity (AX2) which may establish synapses en
Fig. 5 Double-labelled
preparations combining GABA
immunocytochemistry (post-
embedding method) and either
degeneration (a–c) or HRP
labelling (d–g, DAB method).
a A degenerating AT1 terminal
making synaptic contact with a
conventional GABA-
immunonegative dendrite
(Dg-). b A similar terminal
making synaptic contact with
a GABA-ir DCSV of type 2.
c A GABA-ir DCSV poor in
synaptic vesicles, probably of
type 1, contacted by a
degenerating visual terminal in
which the synaptic
differentiation is still visible
(arrows). d An HRP-labelled
optic terminal making contact
with a GABA-ir DCSV (topof image) and a conventional
GABA-negative dendrite
(Dg-, bottom of image),
e An HRP-labelled GABA-
immunoreactive dendrite of an
RMA centrifugal visual neuron
contacted by a AT1 optic
terminal, f, g Examples of HRP-
labelled optic terminals making
synaptic contact with GABA-ir
DCSV. Scale bars 0.25 lm in
(f), 0.5 lm in other images
Brain Struct Funct
123

passant with DCSV (Fig. 8a) or with HRP-labelled den-
drite of the centrifugal visual neurons (Fig. 8d). While we
have no tangible proof, it is quite possible that the AT2
terminals, which account for 18.8% of the total population
of PCSV, arise from these unlabelled fibres.
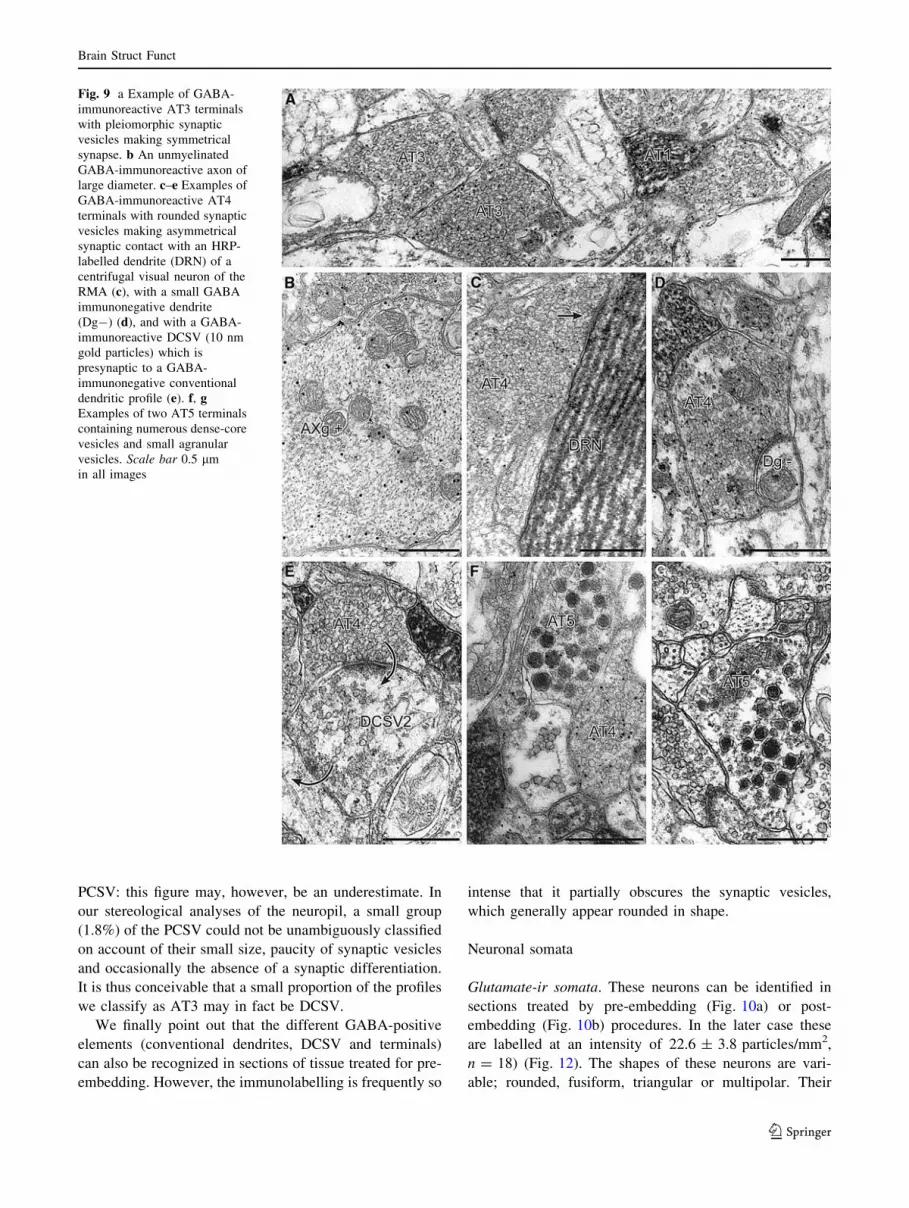
AT3. These GABA-ir (20.8 ± 2.4 particles/mm2,
n = 26) (Fig. 13) terminals are medium-sized (0.95 ±
0.204 lm2) and display a medium to dense matrix with
densely packed flattened vesicles (535 ± 65 nm2 in size
with a shape index of 0.42 ± 0.06, n = 132) (Figs. 4d, 9a,
11). They make symmetrical synaptic contact, for the most
part with small glutamate-ir profiles. They account for
22.6% of the total population of PCSV.
AT4. These terminals are also GABA-ir (Figs. 5a, 9c–e),
but more so (P \ 0.001) than the AT3 terminals
(24.2 ± 3.1 particles/mm2, n = 28) (Fig. 13), with a mean
cross-sectional area of 0.94 ± 0.27 lm2, their light to
moderately dense axoplasm contains numerous small,
rounded to oval, synaptic vesicles (802 ± 70 nm2 in size,
with a mean shape index of 0.82 ± 0.16, n = 143), these
Fig. 6 Examples of the two
types of DCSV observed in the
superficial tectal layers of the
lamprey. a A GABA-
immunoreactive dendrite,
oriented vertically with respect
to the tectal surface, gives off a
collateral branch containing
synaptic vesicles (arrows)
which ends in a swelling packed
with synaptic vesicles (DCSV2)
which is contacted by a HRP-
labelled AT1 terminal (DAB
method). b and c The type 1
DCSV are less frequently
observed than the former; they
generally appear as large
GABA-immunoreactive profiles
oriented parallel to the tectal
surface and contain
pleiomorphic vesicles at a lower
density than the DCSV2. The
DCSV1 are contacted by HRP-
labelled AT1 terminals (DAB
method). In c, note the synaptic
triad (arrows) involving an
HRP-labelled AT1 terminal,
a GABA-immunoreactive
DCSV1 and a small GABA
immunonegative dendrite
profile (Dg-). Scale bar 0.5 lm
in all images
Brain Struct Funct
123

values differing (P \ 0.001) from those of AT3 terminals
(Fig. 11). They make asymmetrical synaptic contact with
HRP-labelled dendrites of RMA neurons (Fig. 9c), with
GABA-ir DCSV (Fig. 9e), and small dendritic profiles
(Fig. 9d) which may be either GABA- or glutamate-ir.
They account for 14.8% of the total population of PCSV.
We point out the existence within the retinorecipient layers
of the tectum of large (C2 lm) strongly GABA-ir (23 ±
2.6 particles/mm2) unmyelinated axons (Fig. 9b), from
which these terminals may possibly arise.
AT5. These terminals (Fig. 9f, g), which are rarely
observed, accounting for 1.7% of the total population of
PCSV, are characterised by the fact that they contain both
clear synaptic vesicles and a large number of dense core
vesicles. Some of them show an accumulation of GABA-
immunoreactive particles over their synaptic vesicles.
Dendrites containing synaptic vesicles (DCSV)
These profiles are GABA-ir (23.04 ± 3.9 particles/mm2,
n = 38) (Fig. 13); their sizes vary considerably as a
function of the plane of section. Within their clear axo-
plasm microtubules, ribosomes and mitochondria may be
observed; finger-like blobs of smooth endoplasmic reticu-
lum can also be observed. Their synaptic vesicles are large
[1,270 ± 76 nm2) and ovoid to flattened in shape (shape
index 0.67 ± 0.11, n = 143) (Fig. 11)]. In general, two
types of DCSV can be distinguished by morphological
Fig. 7 a An example of triple
labelling, showing an HRP-
labelled AT1 terminal (TMB
method) expressing glutamate
immunoreactivity (10 nm gold
particles) and two GABA-ir
AT3 (30 nm gold particles);
note that in these, the smaller
particles indicating glutamate
reactivity are generally
concentrated over mitochondria.
b An example of a synaptic
triad, in which a glutamate-
immunoreactive HRP-labelled
AT1 terminal (DAB method)
makes synaptic contact with a
glutamate-immunonegative
DCSV2, which in turn contacts
a conventional glutamate-
immunoreactive dendrite
(DG?). Scale bars 0.25 lm in
(a), 0.5 lm in (b)
Brain Struct Funct
123
criteria. Those of the first type (DCSV1) are relatively rare;
large in size and polygonal in shape, their longest axes are
oriented parallel to the tectal surface (Fig. 6b, c). Their
synaptic vesicles are dispersed throughout the profile, but
tend to be concentrated in the region of the synaptic dif-
ferentiation; these DCSV are most commonly observed
in the SM. The DCSV of the second, commoner, type
(DCVS2) have a generally ovoid profile, smaller than those
of the first type, (Figs. 5f, g, 6a, 7b, 8c, 9e). The density of
their synaptic vesicles is visibly greater than that of the
DCSV of the first type; these profiles are more common in
the SCFE. In the majority of cases they appear in isolation
among other profiles of the neuropil. In particularly
favorable cases they appear as collateral, vesicle-filled
processes united to a dendritic shaft by a short pedicle
(Fig. 6a); the dendritic profiles which give rise to these
DCSV are generally oriented vertically to the tectal sur-
face. We point out, however, that in some cases (Figs. 5c,
d, f, 8b) it was not possible to clearly determine, on the
basis of these morphological criteria, to which category
some DCSV belonged.
The two type of DCSV may be postsynaptic to AT1 and
more rarely to AT2 and AT4 terminals, and may them-
selves make symmetrical synaptic contact with dendritic
profiles; these synaptic triads are, however, relatively rare
(Figs. 6c, 7b, 9e). The DCSV account for 10.8% of the
Fig. 8 Various aspects of AT2
terminals and their axons
(AX2). a A GABA-
immunoreactive DCSV,
oriented parallel to the tectal
surface and poorly endowed
with synaptic vesicles, is
contacted by an HRP labelled
(DAB method) AT1 terminal
(arrow) and en passant by an
HRP unlabelled AX2 axon
(arrow). b Two AT2 terminals
showing strong glutamate-
immunoreactivity, one of which
makes asymmetrical synaptic
contact with a DCSV. c An
example of an HRP unlabelled
AT2 terminal GABA
immunonegative making
asymmetrical synaptic contact
with a GABA-immunoreactive
DCSV2 and a conventional
GABA immunonegative
dendrite. d A glutamate-
immunoreactive (10 nm gold
particles) AX2 axon making
contact en passant with a
dendrite HRP-labelled (DAB
method) centrifugal visual
neurons of the RMA.
e A glutamate-immunoreactive
AT2 terminal making
asymmetrical synaptic contact
with a glutamate-
immunoreactive HRP labelled
(TMB method) dendrite of a
centrifugal visual neuron. Scalebar 0.5 lm in all images
Brain Struct Funct
123
PCSV: this figure may, however, be an underestimate. In
our stereological analyses of the neuropil, a small group
(1.8%) of the PCSV could not be unambiguously classified
on account of their small size, paucity of synaptic vesicles
and occasionally the absence of a synaptic differentiation.
It is thus conceivable that a small proportion of the profiles
we classify as AT3 may in fact be DCSV.
We finally point out that the different GABA-positive
elements (conventional dendrites, DCSV and terminals)
can also be recognized in sections of tissue treated for pre-
embedding. However, the immunolabelling is frequently so
intense that it partially obscures the synaptic vesicles,
which generally appear rounded in shape.
Neuronal somata
Glutamate-ir somata. These neurons can be identified in
sections treated by pre-embedding (Fig. 10a) or post-
embedding (Fig. 10b) procedures. In the later case these
are labelled at an intensity of 22.6 ± 3.8 particles/mm2,
n = 18) (Fig. 12). The shapes of these neurons are vari-
able; rounded, fusiform, triangular or multipolar. Their
Fig. 9 a Example of GABA-
immunoreactive AT3 terminals
with pleiomorphic synaptic
vesicles making symmetrical
synapse. b An unmyelinated
GABA-immunoreactive axon of
large diameter. c–e Examples of
GABA-immunoreactive AT4
terminals with rounded synaptic
vesicles making asymmetrical
synaptic contact with an HRP-
labelled dendrite (DRN) of a
centrifugal visual neuron of the
RMA (c), with a small GABA
immunonegative dendrite
(Dg-) (d), and with a GABA-
immunoreactive DCSV (10 nm
gold particles) which is
presynaptic to a GABA-
immunonegative conventional
dendritic profile (e). f, gExamples of two AT5 terminals
containing numerous dense-core
vesicles and small agranular
vesicles. Scale bar 0.5 lm
in all images
Brain Struct Funct
123
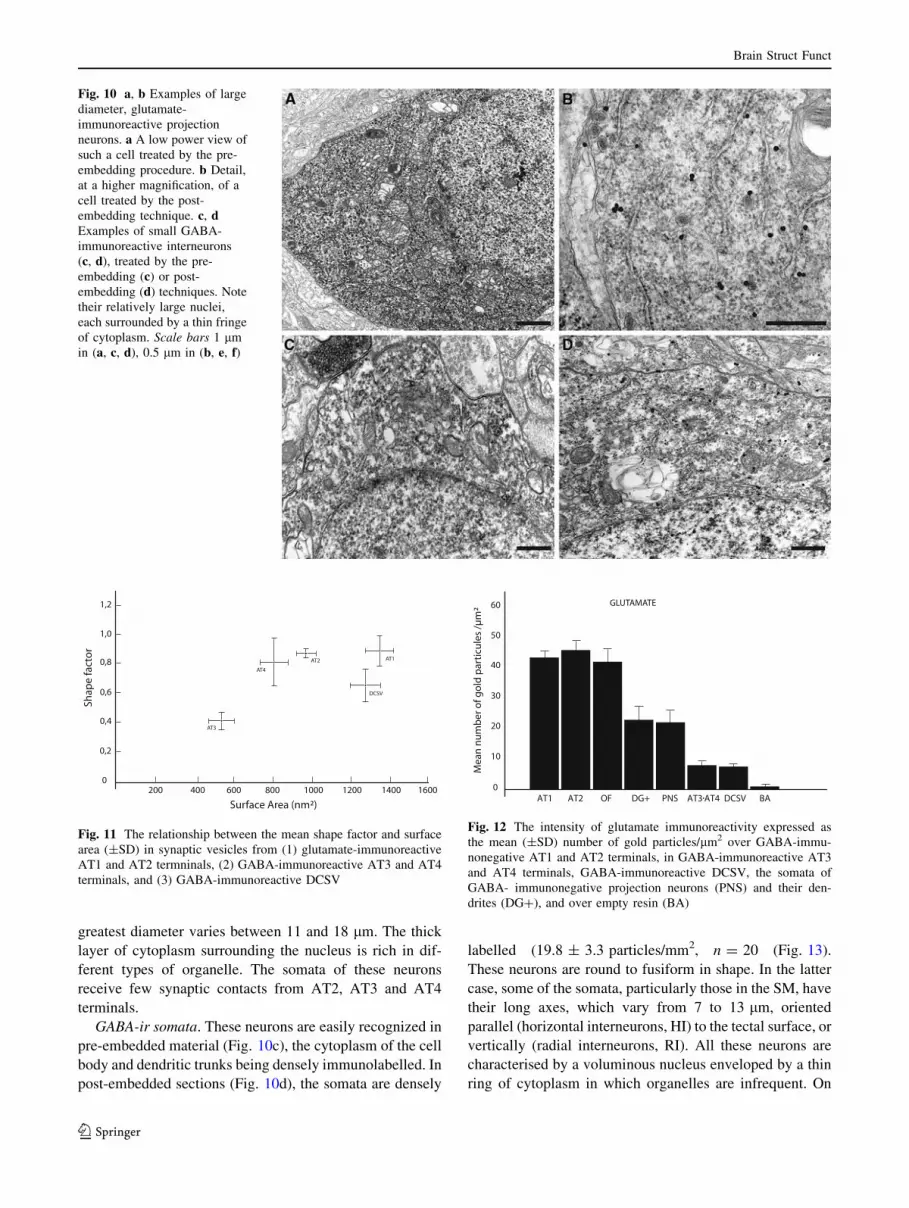
greatest diameter varies between 11 and 18 lm. The thick
layer of cytoplasm surrounding the nucleus is rich in dif-
ferent types of organelle. The somata of these neurons
receive few synaptic contacts from AT2, AT3 and AT4
terminals.
GABA-ir somata. These neurons are easily recognized in
pre-embedded material (Fig. 10c), the cytoplasm of the cell
body and dendritic trunks being densely immunolabelled. In
post-embedded sections (Fig. 10d), the somata are densely
labelled (19.8 ± 3.3 particles/mm2, n = 20 (Fig. 13).
These neurons are round to fusiform in shape. In the latter
case, some of the somata, particularly those in the SM, have
their long axes, which vary from 7 to 13 lm, oriented
parallel (horizontal interneurons, HI) to the tectal surface, or
vertically (radial interneurons, RI). All these neurons are
characterised by a voluminous nucleus enveloped by a thin
ring of cytoplasm in which organelles are infrequent. On
Fig. 10 a, b Examples of large
diameter, glutamate-
immunoreactive projection
neurons. a A low power view of
such a cell treated by the pre-
embedding procedure. b Detail,
at a higher magnification, of a
cell treated by the post-
embedding technique. c, dExamples of small GABA-
immunoreactive interneurons
(c, d), treated by the pre-
embedding (c) or post-
embedding (d) techniques. Note
their relatively large nuclei,
each surrounded by a thin fringe
of cytoplasm. Scale bars 1 lm
in (a, c, d), 0.5 lm in (b, e, f)
0
Shap
e fa
cto
r
200 400 600 800 1000 1200 1400 1600
AT3
AT4
AT2 AT1
DCSV
Surface Area (nm²)
0,2
0,4
0,6
0,8
1,0
1,2
Fig. 11 The relationship between the mean shape factor and surface
area (±SD) in synaptic vesicles from (1) glutamate-immunoreactive
AT1 and AT2 termninals, (2) GABA-immunoreactive AT3 and AT4
terminals, and (3) GABA-immunoreactive DCSV
GLUTAMATE
0
10
20
30
40
50
60
Mea
nn
um
ber
ofg
old
par
ticu
les
/µm
²
AT1 AT2 OF DG+ PNS AT3 AT4 DCSV BA
Fig. 12 The intensity of glutamate immunoreactivity expressed as
the mean (±SD) number of gold particles/lm2 over GABA-immu-
nonegative AT1 and AT2 terminals, in GABA-immunoreactive AT3
and AT4 terminals, GABA-immunoreactive DCSV, the somata of
GABA- immunonegative projection neurons (PNS) and their den-
drites (DG?), and over empty resin (BA)
Brain Struct Funct
123

occasion, cilia issuing from the somatic cytoplasm can be
observed. The somata of these neurons receive some syn-
aptic contact from AT2 and AT4 terminals.
The synaptic circuitry that we propose is summarised in
a schematic diagram (Fig. 14).
Discussion
Comparison with previous findings in the lamprey
Light microscopy
Cyto/chemoarchitecture of the superficial layers of the
optic tectum. The fact that the superficial layers of the optic
tectum of the lamprey are generally poorly furnished with
neurons has been mentioned in many studies of Lampetra
and other lamprey species (Ariens-Kappers et al. 1936;
Heier 1948; Leghissa 1962; Schober 1964; Pfister 1971;
Nieuwenhuys 1977; Kennedy and Rubinson 1984; Rio
et al. 1996; Nieuwenhuys and Nicholson 1998; Iwahori
et al. 1999). We are able to support this finding with some
quantitative data, concerning in particular the GABA-ir
neurons of this region. These cells, which account for half
the neurons of the superficial layers of the dorsolateral
region of the central portion of the tectum, are sparsely
distributed at a density of 684 ± 203 cells/mm3. In a
recent study of the distribution of GABA in the central
nervous system of L. fluviatilis, Robertson et al. (2007)
note that GABA-ir neurons are particularly numerous in
the deeper tectal layers, the stratum griseum periventricu-
lare in particular, whereas in the superficial layers they are
widely dispersed and weakly labelled. They note, as we do,
that in the latter region the neurons are for the most part
small or medium sized. They also describe several large
GABA-containing cells, which we did not observe in our
material. The fact that the small cells are GABAergic
indicates that they are most likely local circuit neurons or
interneurons (see below). The nature of the larger, GABA-
immunonegative neurons, which are highly glutamate-
immunoreactive, is more difficult to determine, but it is
quite likely that they are intra- or extratectal projection
neurons.
Hodological data. The distribution of retinal projec-
tions to the optic tectum, and that of the retinopetal
neurons, that we describe above after intraocular injection
AT1AT2AT4
AT1AT2AT4
DCVS1
HI
Dg+
AT2AT4
DG+
AT1AT2AT4
DCVS1
AXg(?)
Dg+
RI
AT1AT2AT4
AT1AT2AT4
DCSV2DG+
AT1AT2AT4
AT2AT4
DCSV2
AXg(?)
DCF, G+, g+
AT1AT2AT4
DG+
AT1AT2
AT4AT3
Fig. 14 A schematic
representation of the proposed
synaptic circuitry in the
retinorecipient layers of the
optic tectum of the lamprey
GABA
0
5
10
15
20
25
30
Mea
nn
um
ber
ofg
old
par
ticu
les
/µm
²
AT3 AT4 INS BADg+DCSV
Fig. 13 The intensity of GABA-immunoreactivity expressed as the
mean (±SD) number of gold particles/lm2 over AT3 and AT4
terminals, DCSV, the somata of interneurons (INS) and their
dendrites (Dg?), and over empty resin (BA)
Brain Struct Funct
123

of RITC or iontophoretic deposit of HRP into the optic
nerve of L. fluviatilis are entirely comparable to those
obtained in previous studies of this and other species
of lamprey using differents tracers: [3H] proline, [3H]
adenosine, [3H] glycine, HRP (Kosareva et al. 1977;
Kosareva 1980; Reperant et al. 1980, 1982, 1985; Ves-
selkin et al. 1980, 1984, 1996; de Miguel et al. 1990;
Pierre et al. 1992; Rio et al. 1992, 1996; Rodicio et al.
1995). We nevertheless point out that in our experiments
involving the degeneration of optic fibres by intraocular
injection of calcium chloride, followed several days later
by the iontophoretic deposit of HRP into the optic nerve,
anterograde labelling was totally absent from the primary
visual centres and in particular from the superficial tectal
layers. In these latter regions the only labelled structures
were a few dendrites of the centrifugal visual cells of the
RMA, labelled by retrograde transport of the enzyme. No
labelled prolongations, possibly corresponding to axons of
the centrifugal neurons of M5, were observed to pass in
their direction of the tectum. These observations indicate
(see also below) that none of the centrifugal visual neu-
rons project by way of collaterals in the superficial tectal
layers, a hypothesis recently formulated by Robertson
et al. (2006).
Electron microscopy
A single study of the fine structure of the optic tectum of
the lamprey (Rio et al. 1996) has been carried out; it was
primarily concerned with the synaptic relationships of the
intratectal dendrites of the centrifugal visual neurons of the
RMA with retinal terminals and other categories of ter-
minal. Our results are, on the whole, in general agreement
with several of those reported by Rio et al. (1996), par-
ticularly those concerning the cytological, morphometric
and synaptological properties of the retinal terminals. Rio
et al. (1996) also describe seven other types of PCSV,
including DCSV, of which the majority were situated in the
central and deep layers of the optic tectum. In the present
study devoted to the retinorecipient layers, in addition to
the retinal terminals (AT1) and the DCSV, we were able to
identify only four categories of terminal, of which one
(AT5) was not described by Rio et al. (1996). The
remaining three (AT2, AT3 and AT4) are difficult to
compare to the nonvisual terminals contacting the RMA
dendrites described by Rio et al. (1996), since these latter
PCSV were mainly observed in the nonretinorecipient
tectal layers.
Comparison with Gnathostomes
We propose to compare our ultrastructural data concerning
the retinorecipient layers of the optic tectum of the lamprey
with those of the literature concerning the same layers of
the optic tectum or superior colliculus of jawed vertebrates.
Retinal terminals (AT1)
The cytological features of these terminals (scalloped
outline, electron-lucent axoplasm, round synaptic vesicles
and asymmetrical synaptic contacts) in the lamprey are
totally comparable to those of retinotectal or retinocollic-
ular terminals described in Gnathostomes (for reviews:
teleosts: Peyrichoux et al. 1986; Vanegas et al. 1984;
Kageyama and Meyer 1989; amphibians, Lazar 1984;
Antal 1991; Gabriel and Straznicky 1995; reptiles, Repe-
rant et al. 1981, 1997; birds, Reperant and Angaut 1977;
Acheson et al. 1980; Morino et al. 1991; Tombol 1998;
Tombol and Nemeth 1999; Tombol et al. 2003. mammals:
Mize 1983, 1988; Huerta and Harting 1984; Schonitzer and
Hollander 1984; Ortega et al. 1995; Mize and Butler 1996).
The proportion of the total population of PCSV represented
by the retinal terminals of the lamprey (31.3%, present
results) lies within the range of values of this proportion
described in Gnathostomes (Teleosts: Carassius, 37% for
Murray and Edwards (1982) but 27% for Airhart and
Kriebel (1984); Rutilus, 23.5% (Peyrichoux et al. 1986);
Holocentrus, 16% (Ito et al. 1980); Reptiles: Vipera aspis,
25% (Reperant et al. 1981); Birds: Columba livia, 23%
(Angaut and Reperant 1976); Mammals: rabbit, 20%
(Vrensen and de Groot 1977); cat, 24% (Sterling 1971); the
primate Galago, 45% (Tigges et al. 1973). This variation
may reflect interspecific differences as well as different
methods of estimation.
The retinal terminals (AT1) of the lamprey tectum are
GABA-immunonegative and strongly glutamate-immuno-
reactive. It has been shown that the same is true not only of
retinotectal or retinocollicular terminals in all major groups
of Gnathostomes (fish, Kageyama and Meyer 1989;
amphibians, Gabriel and Straznicky 1995; reptiles, Repe-
rant et al. 1997; birds, Morino et al. 1996; mammals,
Ortega et al. 1995; Mize and Butler 1996) but also of all
retinal terminals in general (Reperant et al. 1997; Kenigfest
et al. 1998 for review), with the exception of a small
population of GABAergic visual terminals (Kenigfest et al.
1998; Miceli et al. 2008 for review) of which we found not
the slightest trace in the lamprey.
The fact that the retinal terminals are highly glutamate-
reactive, and that this immunoreactivity is preferentially
expressed over synaptic vesicles, strongly suggests that
these terminals use glutamate as a neurotransmitter (cf.
Montero 1990, 1994; Mize and Butler 1996; Reperant et al.
1997; Kenigfest et al. 1998). Other data support this
hypothesis. In the optic tectum and other primary visual
centres (dorsal lateral geniculate nucleus, nucleus of the
optic tract, suprachiasmatic nucleus) of a wide variety of
Brain Struct Funct
123

species (goldfish, frog, pigeon, chicken, mouse, rat, rabbit,
cat) extra- and intracellular recordings in vivo or in vitro
have shown that antagonists of excitatory amino acids such
as glutamate block the excitatory postsynaptic potentials
induced by stimulation of the retina. In addition, the
iontophoretic administration of glutamate produces an
excitatory response of visually sensitive neurons (Felix and
Frangi 1977; Langsdon and Freeman 1986; Shibata et al.
1986; Debski et al. 1987; van Deusen and Meyer 1988;
Cahill and Menaker 1989; Murphy et al. 1989; Hartveit and
Heggelund 1990; Heggelund and Hartveit 1990; Sillito
et al. 1990a, b; Funke et al. 1991; Kim and Dudek 1991;
Kwon et al. 1991; Roberts et al. 1991; Schmidt 1991;
Hestrin 1992; Binns and Salt 1994; Dye and Karten 1996).
In the pigeon optic tectum (Canzek et al. 1981) and the
rabbit superior colliculus (Sandberg et al. 1982), electrical
stimulation of the optic nerve leads to a release of endog-
enous glutamate and aspartate in vivo. In vitro, a calcium-
dependent potassium-induced release of glutamate has
been demonstrated in pigeon brain slices (Reubi 1980;
Toggenburger et al. 1982). The high-affinity uptake of
glutamate is greatly reduced in synaptosomes prepared
from the retinorecipient layers of enucleated chickens
(Henke et al. 1976) or pigeons (Bondy and Purdy 1977);
retinal ablation also leads to a significant reduction of
glutamate and aspartate levels in the optic nerve and reti-
norecipient tectal layers in the pigeon (Fonnum and Henke
1982; Morino et al. 1991) and the corresponding collicular
layers in the rat (Sakurai and Okada 1992; Sakurai et al.
1990). Mize and Butler (1995) have shown that, in the
superficial layers of the cat superior colliculus, antibody
labelling of NMDA R1 receptor subtypes is present at the
postsynaptic densities opposite to retinal terminals. Finally,
NMDA and nonNMDA agonists have been shown to excite
the retinorecipient layers of the feline superior colliculus in
vitro (Grantyn et al. 1987; Perouansky and Grantyn 1990).
Our data show that the retinotectal terminals in Lam-
petra make synaptic contact essentially with dendritic
profiles, of which the majority are devoid of synaptic
vesicles (conventional dendritic profiles, D), and with a
minority of DCSV. The D profiles are either GABA-ir
(Dg?) or glutamate-ir (DG?). It has been shown that
the same applies to all major groups of Gnathostomes
(fish, Kageyama and Meyer 1989; amphibians, Antal 1991,
Gabriel and Straznicky 1995; reptiles, Rio et al. 1995,
Reperant et al. 1997; birds, Morino et al. 1991; mammals,
Houser et al. 1983; Mize 1988; Mize and Butler 1996).
However, the relative proportions of the two types of
postsynaptic target vary somewhat. In Lampetra, 19% of
the retinal terminals contact DCSV (present results), a
value to be compared with 4% in the amphibian Ambys-
toma mexicanum (Ingham and Guldner 1981), 10% in the
teleost Rutilus (Peyrichoux et al. 1986), 11% in the viper
(Reperant et al. 1981), 33–50% in the cat (Sterling 1971;
but 20–34% in the same species for Behan 1981) and 57%
in the cane toad Bufo marinus (Gabriel and Straznicky
1995). The precise cellular origins of these dendritic targets
are for the most part unknown, but it is highly probable that
in all taxonomic groups the GABA-ir DCSV arise from
interneurons (see below), as do a proportion of GABA-ir D
profiles. However, in the lamprey, we have seen that some
of these latter profiles arise from the retinopetal neurons of
the RMA, and a small proportion of GABA-negative,
glutamate-positive dendritic profiles have the same origin
(present results).
With the exception of a relatively modest proportion of
synaptic contacts between retinal terminals and the den-
drites of the glutamate-ir retinopetal neurons of the RMA
(present results), we have no data concerning the intratectal
origin of the glutamate-ir neurons whose conventional
dendrites are postsynaptic to retinal terminals. They may
possibly arise from the GABA-immunonegative neurons of
the superficial tectal layers, of which the somata are
strongly glutamate-immunoreactive. It is also possible that
these dendrites arise from efferent neurons situated in the
deep tectal layers (Heier 1948; Kennedy and Rubinson
1984; Iwahori et al. 1999), whose long vertical dendrites
penetrate the retinorecipient layers (Iwahori et al. 1999).
Such an organisation has been clearly demonstrated in the
superior colliculus of the cat (Jeon et al. 1997) and the
optic tectum of the chicken (Tombol and Nemeth 1999;
Tombol et al. 2003).
We point out that one of the synaptological peculiarities
of the retinorecipient layers of the lamprey tectum, in
comparison with those of Gnathostomes, is that the visual
fibres make direct synaptic contact with the dendrites of
retinopetal neurons, thus forming a fast visual feedback
loop (retina ? tectum ? centrifugal neurons ? retina;
cf. Rio et al. 1996; Vesselkin et al. 1996; Reperant et al.
2006). While a similar feedback loop exists in birds and
crocodiles, in these organisms the dendrites of the cen-
trifugal neurons do not penetrate the optic tectum, and the
feedback loop contains an additional mono- or polysyn-
aptic pathway (cf. Reperant et al. 1989, 2006, 2007;
Uchiyama 1989; Ward et al. 1991; Medina et al. 2004).
Nonretinal terminals
In contrast to the retinal terminals, we have no experi-
mental hodological data concerning the origins of the
nonretinal GABA-ir and glutamate-ir terminals. However,
the recent light microscopic studies by Robertson et al.
(2006, 2007) which combine axonal tracing techniques and
immunohistochemistry in an attempt to identify the origins
of tectal afferent supplies and their neurochemical prop-
erties provide us with the means to advance several
Brain Struct Funct
123

hypotheses concerning the origin of these terminals. In the
same light, the comparison of our ultrastructural findings
with those in the literature concerning the superficial tectal
or collicular layers of Gnathostomes enables us, on the
basis of cytological, neurochemical and synaptological
criteria, to establish some evident points of comparison.
GABA-negative, glutamate-positive, nonvisual AT2
terminals
A relatively important proportion (18.8% of PCSV) of this
category of terminals has been recognised in the retinore-
cipient layers of the tectum of Lampetra (present results).
Given the relatively large proportion of AT2 terminals, it is
quite likely that they represent several subgroups arising in
different structures. In this context, Robertson et al. (2006)
have identified several structures containing GABA-im-
munonegative neurons (possibly glutamate-ir) which
provide afferent supplies to the tectum, for example those
situated in the caudo-ventral telencephalic pallium, the
pretectal region, the isthmic region and the octavolateral
area.
In the same tectal or collicular regions of Gnathostomes,
profiles with the same cytological characteristics as the
AT2 terminals of the lamprey (size and shape of the
synaptic vesicles, asymmetrical synaptic contact and con-
ventional dendrites or DCSV as targets) have been
identified (teleosts, Ito et al. 1980; Meek 1981; Peyrichoux
et al. 1986; amphibians, Antal 1991; reptiles, Reperant
et al. 1981; birds, Angaut and Reperant 1976; Tombol et al.
2003; mammals, Lund 1969, 1972; Sterling 1971; Tigges
et al. 1973; Tigges and Tigges 1975; Mize and Butler
1996), and some of these studies (Antal 1991; Mize and
Butler 1996) have shown that, like the AT2 terminals of the
lamprey, they are GABA-negative and glutamate-ir. The
proportion of these terminals varies considerably between
groups: our value of 18.8% is to be compared with 1% in
the pigeon (Angaut and Reperant 1976), 4% in the viper
(Reperant et al. 1981), 11% in the teleost Rutilus (Peyric-
houx et al. 1986), and 30% in the primate Galago (Tigges
et al. 1973). Despite these cytological, synaptological
and neurochemical similarities between these terminals
observed in lampreys and Gnathostomes, it is, at present,
impossible to affirm that they are homologous between the
two groups, since their origins remain largely unknown.
Thus, for example, in mammals it has been shown that
these glutamate-ir terminals (Mize and Butler 1996) are the
endings of the cortico-collicular pathway (Lund 1969,
1972; Sterling 1971; Tigges et al. 1973; Behan 1980;
Graham and Casagrande 1980; Mathers 1977; Valverde
1973; Mize and Butler 1996). Ito et al. (1980) have shown
that in the retinorecipient layers of the tectum of the teleost
Holocentrus rufus, there exists a category of nonvisual
terminals with round synaptic vesicles and asymmetrical
synaptic contact (S4), which degenerate after ablation of
the ipsilateral telencephalon. In the lamprey, the existence
of a telencephalo-tectal pathway has been demonstrated by
a variety of tracing techniques (Polenova and Vesselkin
1993; Northcutt and Wicht 1997; Robertson et al. 2006).
This pathway arises in the primordium hippocampi (Pole-
nova and Vesselkin 1993; Northcutt and Wicht 1997), a
structure also described as the medial pallium (Robertson
et al. 2006). Using a combination of tracing and GABA-
immunocytochemical techniques, Robertson et al. (2006)
conclude that some of these cells at the origin of this
projection, situated in the caudo-ventral pallium, are
GABA-immunoreactive, whereas the majority of these
neurons are GABA-immunonegative and probably gluta-
mate-ir.
GABAergic terminals (AT3, AT4)
In the superficial layers of the lamprey tectum, 37.4% of
PCSV are GABAergic terminals, and fall into one of two
categories; those with pleiomorphic synaptic vesicles and
symmetrical synaptic contacts (22.6%), and those with
round vesicles and asymmetrical synaptic contacts (14.8%)
(present results).
In the lamprey, it is highly likely that the GABAergic
terminals of the superficial tectal layers have several
origins. Some are almost certainly those of interneurons,
while others may be part of the afferent supplies to these
layers. In this context, Robertson et al. (2006) have shown
that in L. fluviatilis many sources of the GABAergic
afferent supply to the tectum arise in the caudo-ventral
pallium, the ventral and dorsal thalamus, the torus semi-
circularis and the nucleus M5 of Schober, within which
lie some of the centrifugal visual neurons. These authors
therefore advance two hypotheses: (1) the tectum is
innervated by collateral branches of the axons of the
centrifugal visual neurons of M5, a large number of
which are GABAergic (Rio et al. 1993); (2) the tectum
receives a GABAergic projection from M5 arising in
exclusively tectopetal neurons. This is quite plausible,
since the nucleus M5 is not composed exclusively of
centrifugal visual neurons (cf. Rio et al. 1993). Our
observations lead us to support the second of these
hypotheses (see also above). As we have pointed out,
after chemical destruction of the retina by calcium chlo-
ride, and subsequent administration of HRP to the optic
nerve several days later, only the elements of the cen-
trifugal visual neurons (axons, somata and dendrites) are
labelled with tracer. Examination of the retinorecipient
tectal layers reveals only the presence of dendrites of the
neurons of the RMA, with not the slightest trace of
labelled axon terminals.
Brain Struct Funct
123

In double-labelled preparations we have noticed that
these terminals, like other GABAergic elements in the
superficial tectal layers of the lamprey (cell bodies, den-
drites, DCSV) also display glutamate-immunoreactivity.
However, the density of particles from the anti-glutamate
antibody is consistently lower than those of the anti-GABA
antibody (see above), and that the latter are seen predom-
inantly over synaptic vesicles, whereas the former are
concentrated over mitochondria. Similar observations have
been made in other contexts (cf. Montero and Wenthold
1989; Montero 1990, 1994; Reperant et al. 1997; Kenigfest
et al. 1998). In contrast to the neurochemical identification
of other substances such as GABA, for which positive
immunoreactivity appears to be a specific marker of
GABAergic neurons, immunoreactivity to glutamate
appears to be more difficult to interpret. This amino acid is
present in most cellular compartments, and is involved in a
number of metabolic pathways. Several authors (Ottersen
and Storm-Mathisen 1984; Ottersen 1987, 1989; Storm-
Mathisen and Ottersen 1990) have suggested that gluta-
mate immunoreactivity reveals both the ‘‘metabolic’’ and
‘‘transmitter’’ pools of this amino acid in neural processes.
However, a high concentration of particles indicating
glutamate-immunoreactivity over the vesicles of AT1 and
AT2 terminals suggests that this substance is used as a
neurotransmitter (see above), while in the GABAergic
PCSV these particles are concentrated over mitochondria
and very likely reveal the metabolic pool of glutamate,
which serves as a precursor of GABA by the action of
glutamate decarboxylase (see Ottersen and Storm-Mathisen
1984).
A comparison of our findings with the data presented in
the literature concerning the superficial tectal or collicular
layers of Gnathostomes shows that in the two cases, while
the proportions of the two types of GABAergic terminal
vary from one species to another, they show a number of
cytological and synaptological features in common.
For example, Antal (1991) recognises, as in the lamprey,
two types of GABA-immunoreactive terminals in the ret-
inorecipient tectal layers of Rana esculenta; those with
pleiomorphic vesicles and symmetrical synaptic contact,
and those with rounded vesicles and asymmetrical contact;
in the same way, Tombol et al. (2003) recognize two types
of GABA-ir terminals in these regions of the chicken
tectum, one with pleiomorphic synaptic vesicles and
symmetrical contact, the other with round vesicles but with
symmetric contact. On the other hand, Rio et al. (1996)
distinguish four types of GABAergic terminal in the reti-
norecipient layers of the tectum of Vipera. Three of these
contain pleiomorphic synaptic vesicles and make sym-
metrical synaptic contact, while the fourth contains
rounded vesicles and makes asymmetrical contact. In
mammals, the majority of the GABAergic terminals of the
retinorecipient layers of the superior colliculus contain
pleiomorphic vesicles and make symmetrical synaptic
contact (cf. Lund 1969, 1972; Tigges et al. 1973; Valverde
1973; Houser et al. 1983; Mize 1992). However, Houser
et al. (1983) consider that in the retinorecipient layers of
the rat superior colliculus 4% of the GABAergic terminals
make assymmetrical synaptic contacts. In the lamprey, as
in Gnathostomes, the GABAergic terminals make synaptic
contacts with the dendrites of efferent neurons, and also
with the GABAergic DCSV of interneurons which may in
turn be presynaptic to the dendrites of efferent neurons.
Despite these similarities, it is difficult to establish
homologies between different taxonomic groups for these
different GABAergic terminals, since in the majority of
cases their origins have not been determined with
precision.
AT5 terminals
These terminals, with their many dense-core vesicles, are
rare in the retinorecipient tectal layers of the lamprey. They
have been described, without comment, in the corre-
sponding tectal layers of the viper (Rio et al. 1995), in the
isthmo-optic nucleus of the pigeon (Miceli et al. 2002; Rio
et al. 2002), the nucleus dorsolateralis anterior of the same
species (Miceli et al. 2008), and in the nucleus geniculatus
lateralis dorsalis of mammals (Papadopoulos and Parnav-
elas 1990; Pasik et al. 1988). In the latter cases it has been
shown that they correspond to serotoninergic terminals. At
the level of resolution of the light microscope, serotonin-
immunoreactive fibres have been described in the superfi-
cial tectal layers of the lamprey (Pierre et al. 1992), and it
is thus possible that the AT5 terminals correspond to the
terminals of these fibres.
Interneurons and DCSV
It has been shown many times that in Gnathostomes, as in
the lamprey (Robertson et al. 2007), GABA-ir neurons
exist in the various layers of the optic tectum or superior
colliculus In anamniotes (Franzoni and Morino 1989; Antal
1991; Medina et al. 1994), as in the lamprey (Robertson
et al. 2007; present results) these neurons are considerably
more numerous in the deeper tectal layers, and scattered at
a lower density in the superficial layers. In amniotes, par-
ticularly mammals, on the other hand, the situation is
inversed; the GABA-ir neurons are much commoner in the
superficial collicular layers than in the deeper ones (cf.
Mugnaini and Oertel 1985; Mize 1988, 1992).
The GABA-ir neurons of the superficial tectal layers
described in the lamprey (Robertson et al. 2007; present
results) resemble those described in the corresponding
tectal or collicular regions of Gnathostomes (teleosts:
Brain Struct Funct
123

Medina et al. 1994; amphibians, Franzoni and Morino
1989; Antal 1991; reptiles, Bennis et al. 1991; Rio et al.
1995; birds, Domenici et al. 1988; Granda and Crossland
1989; Veenman and Reiner 1994; mammals, Mugnaini and
Oertel 1985; Mize 1988, 1992; Mize et al. 1991,
1994).They are generally described as small in size, of
variable shape, and oriented either vertically or horizon-
tally with respect to the tectal or collicular surface. In
Lampetra they account for 50.1% of the neurons of the
superficial tectal layers, a figure close to the value of 50.3%
obtained by Rio et al. (1995) for the corresponding tectal
layers of Vipera aspis. However, different proportions of
these cells have been described in amphibians (61%; Antal
1991), the cat (45%; Mize 1988) and the rhesus monkey
(32.5%; Mize et al. 1991).
The GABA-immunoreactive profiles containing synap-
tic vesicles which we have observed in the superficial tectal
layers of the lamprey are undoubtedly of dendritic origin.
In contrast to axonal profiles, they contain polyribosomes
and some form of granular reticulum. In addition, micro-
tubules are very common, and neurofilaments are absent.
For want of serial sections, we were unable to determine
the origin of these profiles, but their high degree of
immunoreactivity strongly suggests that they arise from the
small GABAergic neurons, which show a high degree of
immunoreactivity, while the larger ones are GABA-im-
munonegative and highly glutamate-ir. We were not able to
show whether these small neurons were devoid of axons or
possessed very short axons, but in other respects they show
the cytological features of interneurons. In contrast to
projection neurons, these show a voluminous nucleus,
surrounded by a thin fringe of cytoplasm, which is poorly
endowed with organelles (cf. Peters et al. 1970; Tigges and
Tigges 1975; Mize 1992; Mize et al. 1994), as do the small
GABAergic neurons observed in our material.
In Lampetra, the DCSV are postsynaptic to glutamate-ir
retinal AT1 terminals and AT2 terminals, to GABAergic
AT4 terminals, and presynaptic to conventional dendritic
profiles (present results). In many Gnathostomes, the
DCSV of the superficial tectal or collicular layers are also
postsynaptic in most cases to retinal terminals, but also
to terminals with pleiomorphic synaptic vesicles (fish,
Peyrichoux et al. 1986; amphibians, Szekely et al. 1973;
Ingham and Guldner 1981; Antal 1991; reptiles, Reperant
et al. 1981; Rio et al. 1995; birds, Angaut and Reperant
1976; mammals, Lund 1969, 1972; Tigges et al. 1973;
Valverde 1973; Tigges and Tigges 1975; Graham and
Casagrande 1980; Vrensen and de Groot 1977; Houser
et al. 1983; Mize et al. 1991; Pinard et al. 1991).
Two types of DCSV exist in the superficial tectal layers
of the lamprey (present results). Those of the first type are
dendritic profiles generally oriented in the horizontal plane
that contain discrete clusters of pleiomorphic synaptic
vesicles located at sites of synaptic contact; those of the
second type are more richly endowed with synaptic vesi-
cles. They appear either in isolation or as varicosities
arising from collateral dendritic branches. These two types
of DCSV have been observed in Gnathostomes, most
systematically in the superficial layers of the mammalian
superior colliculus. The DCSV of the first type have been
described in the rat (Lund 1969), the mouse (Valverde
1973), the rabbit (Mize et al. 1994), the cat (Mize et al.
1982; Mize 1988), the rhesus monkey (Mize et al. 1991)
and the chimpanzee (Tigges and Tigges 1975), and it has
been shown that these DCSV arise from the horizontal cells
(cf. Mize 1992). It is likely that the same is true in the
lamprey, given that the horizontal interneurons are present
in the superficial tectal layers. These DCSV have also been
described in the superficial tectal layers of birds (Streit
et al. 1978) and reptiles (Reperant et al. 1981; Rio et al.
1995). In mammals, these profiles receive few synaptic
contacts, particularly from retinal terminals (cf. Mize
1992); in reptiles the inverse is true. In these latter forms
the DCSV are richly innervated, particularly by visual
terminals, and are frequently involved in synaptic triads
(Reperant et al. 1981; Rio et al. 1995), which does not
appear to be the case in mammals (cf. Mize 1992). In the
lamprey, these DCSV receive different types of synaptic
contact, including retinal terminals, and may be involved in
synaptic triads.
The DCSV of the second type have been described in
the superficial tectal layers of reptiles (Reperant et al.
1981), birds (Angaut and Reperant 1976) and on several
occasions in the superficial collicular layers of mammals
(Lund 1969; Sterling 1971; Valverde 1973; Tigges and
Tigges 1975; Mize 1988, 1992; Mize et al. 1982, 1991,
1994).In the lamprey, as in Gnathostomes, these DCSV
may be involved in synaptic triads of which the presynaptic
elements are generally retinal terminals. In mammals the
DCSV have been described in a variety of terms; balloon-
like dilatations (Sterling 1971), vesicle-filled spines (Lund
1969), spine-like appendages or spine-bearing dendrites
(Mize 1988, 1992), gemmule-like dendritic excrescences
(Houser et al. 1983), or as varicose dendritic enlargements
(Mize et al. 1991). It follows from this that the profiles are
interpreted as arising from dendritic spines, dendritic var-
icosities, or both of these. In the lamprey it is highly likely
that the DCSV2 arise from radial interneurons since the
collateral dendrites which give rise to them are generally
oriented perpendicular to the lateral surface. However, it is
also possible that some arise from dendritic spines. Iwahori
et al. (1999) find, in Golgi preparations of Lampetra
japonica, that the majority of neurons of the superficial
tectal layers, particularly those of small size, bear numer-
ous dendritic spines; this in marked contrast to Kennedy
and Rubinson (1984), whose Golgi preparations of
Brain Struct Funct
123

Petromyzon marinus indicate that the superficial tectal
neurons are devoid of dendritic spines. Our own findings
under the electron microscope fall between these positions,
tending towards that of Kennedy and Rubinson (1984). The
different dendritic branches of GABA-ir neurons which
appear in many of the sections we examined are very
poorly endowed with dendritic spines, and in no case did
we observe synaptic vesicles in any of these latter
structures.
Conclusions
A comparison of our data with other observations leads us
to conclude that the dorsolateral retinorecipient layers of
the optic tectum of the lamprey show many synaptological
and neurochemical similarities with the retinorecipient
layers of the tectum or superior colliculus of Gnathosto-
mes, even though it has some idiosyncrasities such as the
direct relationship of the retinal terminals to the dendrites
of retinopetal neurons and the total absence of myelinated
fibres.
These similarities are of several types. For example,
the nonvisual glutamatergic and GABAergic terminals of
lampreys and many Gnathostome species share many
common cytological, synaptological and neurochemical
characteristics; although, as we have pointed out, while
the origins of these terminals remain in many cases
unknown it is difficult to claim that the circuitry in which
they are involved is comparable from one taxonomic
group to another. On the other hand, the functional
anatomical data concerning the retinotectal system are
sufficiently extensive to reveal many similarities between
Gnathostomes and Agnathans. Thus, with the exception of
a small number of GABAergic retinal terminals (Kenigfest
et al. 1998; Miceli et al. 2008 for reviews) which are
absent in lampreys (present results), the retinotectal ter-
minals of all vertebrates are glutamatergic, have rounded
synaptic vesicles, and make asymmetrical synaptic contact
with three types of profile. The first of these are the
dendrites of glutamatergic efferent neurons, which are
activated by the retinal terminals. In the remaining two
cases, which are less numerous, the retinal terminals make
contact either with conventional GABAergic dendrites of
interneurons, or with GABAergic DCSV which may in
turn be presynaptic to the glutamatergic dendrites of
efferent neurons. In this latter triadic configuration, the
retinal terminals activate the DCSV which in turn inhibit
the efferent neurons.
It thus appears that the retinotectal system of verte-
brates, including the Petromyzontiform Agnatha, seems to
be characterised by a fundamental consistency of synaptic
organisation. This leads us to suppose that its underlying
plan probably existed in the ancestral Petromyzontiforms,
from which the Gnathostomes diverged some 450 million
years ago, and also that the consistency of synaptic cir-
cuitry probably implies a similarity of function in visual
information processing.
Numerous correlations between anatomy and function
suggest that interneurons play a powerful inhibitory role, in
particular by way of the dendro-dendritic synapses which
appear to act as a neuron-saving device (cf. Miceli et al.
2008). In addition, the microcircuits represented by the
synaptic triads involving retinal terminals (optic termi-
nal ? DCSV ? conventional dendrite) appear to play a
fundamental role in the local treatment of visual informa-
tion. Sherman (2004), for example, has suggested that they
are involved in the gain control of the visual relay cells to
maintain their modulation transfer functions within a linear
region.
In the superficial tectal layers of the lamprey this
GABAergic interneuronal network appears poorly devel-
oped in comparison to that described in the equivalent
region of Gnathostomes and particularly of Amniotes. In
the superficial tectal layers of the lamprey, the DCSV
account for 10.8% of the total population of PCSV (present
results); this proportion is considerably greater in amniotes
(21% in the viper, Rio et al. 1995; 23.5% in the rabbit,
Vrensen and de Groot 1977; 60% in the pigeon, Angaut
and Reperant 1976). In addition, in the lamprey the pro-
portion of dendro-dendritic synapses in these layers (8.9%
of synaptic contacts, present results) rises to 20% in the
viper (Rio et al. 1995) and cat (Sterling 1971) and to 55.2%
in the pigeon (Angaut and Reperant 1976). Finally, the
axo-dendro-dendritic synaptic triads involving retinal
terminals are extremely rare in the lamprey tectum but
commoner in Amniotes (reptiles; Rio et al. 1995; birds;
Angaut and Reperant 1976; mammals; Lund 1972; Houser
et al. 1983). In the light of these findings it is entirely
reasonable to suppose that the degree of information
processing is considerably less elaborate in the tectal
retinorecipient layers of Lampetra than in those of Gnat-
hostomes and in particular amniotes.
Acknowledgments We thank Denis LeCren for his extremely
skilful photographic assistance. This work was supported by MNHN,
CNRS (France), Academy of Sciences (Russia) NSERC (Canada).
References
Abercrombie M (1946) Estimation of nuclear population from
microtome sections. Anat Rec 94:239–247. doi:10.1002/ar.
1090940210
Acheson DW, Kemplay SK, Webster KE (1980) Quantitative analysis
of optic terminal profile distribution within the pigeon optic
tectum. Neuroscience 5:1067–1084. doi:10.1016/0306-4522
(80)90187-6
Brain Struct Funct
123

Adams JC (1981) Heavy metal intensification of DAB-based reaction
product. J Histochem Cytochem 29:775
Airhart MJ, Kriebel RM (1984) Retinal terminals in the goldfish optic
tectum: identification and characterization. J Comp Neurol
226:377–390. doi:10.1002/cne.902260307
Angaut P, Reperant J (1976) Fine structure of the optic fibre
termination layers in the pigeon optic tectum: a Golgi and
electron microscope study. Neuroscience 1:93–105. doi:10.1016/
0306-4522(76)90003-8
Antal M (1991) Distribution of GABA immunoreactivity in the optic
tectum of the frog: a light and electron microscopic study.
Neuroscience 42:879–891. doi:10.1016/0306-4522(91)90051-O
Ariens-Kappers CV, Huber GC, Crosby EC (1936) The comparative
anatomy of the nervous system including man. Hafner, New York
Behan M (1980) An EM-autoradiographic analysis of the corticocol-
licular terminals in the cat. Soc Neurosci Abstr 6:480
Behan M (1981) Identification and distribution of retinocollicular
terminals in the cat: an electron microscopic autoradiographic
analysis. J Comp Neurol 199:1–15. doi:10.1002/cne.901990102
Bennis M, Calas A, Geffard M, Gamrani H (1991) Distribution of
GABA immunoreactive system in the forebrain and midbrain
of the chameleon. Brain Res Bull 26:891–898. doi:10.1016/
0361-9230(91)90254-H
Binns KE, Salt TE (1994) Excitatory amino-acid receptors participate
in synaptic transmission of visual responses in the superficial
layers of the cat superior colliculus. Eur J Neurosci 6:161–169.
doi:10.1111/j.1460-9568.1994.tb00257.x
Bondy SC, Purdy JL (1977) Putative neurotransmitters of the avian
visual pathway. Brain Res 119:417–426. doi:10.1016/0006-
8993(77)90320-1
Cahill GM, Menaker M (1989) Effects of excitatory amino acid
receptor antagonists and agonists on suprachiasmaticus nucleus
responses to retinohypothalamic tract volleys. Brain Res 479:76–
82. doi:10.1016/0006-8993(89)91337-1
Canzek V, Wolfensberger M, Amsler V, Cuenod M (1981) In vivo
release of glutamate and aspartate following optic nerve
stimulation. Nature 293:572–574. doi:10.1038/293572a0
Colonnier M (1968) Synaptic pattern on different cell types in the
different laminae of the cat visual cortex. An electron-microscopic
study. Brain Res 9:268–287. doi:10.1016/0006-8993(68)90234-5
de Miguel E, Rodicio MC, Anadon R (1990) Organization of the
visual system in larval lampreys: an HRP study. J Comp Neurol
302:529–542. doi:10.1002/cne.903020309
Debski EA, Cline HT, Constantine-Paton M (1987) Kynurenic acid
blocks retinal-tectal transmission in Rana pipiens. Soc Neurosci
Abstr 13:1691
Domenici L, Waldvogel HJ, Matute C, Streit P (1988) Distribution of
GABA-like immunoreactivity in the pigeon brain. Neuroscience
25:931–950. doi:10.1016/0306-4522(88)90047-4
Dye TC, Karten HJ (1996) An in vitro study of retinotectal
transmission in the chick: role of glutamate and GABA in
evoked field potentials. Vis Neurosci 13:747–775
Ebbesson SOE (1970) On the organization of central visual pathways in
vertebrates. Brain Behav Evol 3:178–194. doi:10.1159/000125470
Ebbesson SOE (1972) A proposal for a common nomenclature for
some optic nuclei in vertebrates and the evidence for a common
origin of two such cell groups. Brain Behav Evol 6:75–91. doi:
10.1159/000123698
Felix D, Frangi V (1977) Dimethoxyaporphine as an antagonist of
chemical excitation in the pigeon optic tectum. Neurosci Lett
4:347–350. doi:10.1016/0304-3940(77)90182-3
Fonnum F, Henke H (1982) The topographical distribution of alanine,
aspartate, gamma-aminobutyric acid, glutamate, glutamine and
glycine in the pigeon optic tectum and the effect of retinal
ablation. J Neurochem 38:1130–1134. doi:10.1111/j.1471-4159.
1982.tb05358.x
Forey PL, Janvier P (1993) Agnathans and the origin of jawed
vertebrates. Nature 361:129–134. doi:10.1038/361129a0
Franzoni MF, Morino P (1989) The distribution of GABA-like-
immunoreactive neurons in the brain of the newt, Trituruscristatus carnifex and the green frog, Rana esculenta. Cell
Tissue Res 255:155–166. doi:10.1007/BF00229077
Funke K, Eysel UT, Fitzgibbon T (1991) Retinogeniculate transmis-
sion by NMDA and non-NMDA receptors in the cat. Brain Res
547:229–238. doi:10.1016/0006-8993(91)90966-Y
Gabriel R, Straznicky C (1995) Synapses of optic axons with GABA-
and Glutamate- containing elements in the optic tectum of Bufomarinus. J Hirnforsch 36:329–340
Graham J, Casagrande VA (1980) A light and electron microscopic
study of the superficial layers of the superior colliculus of the
tree shrew (Tupaia glis). J Comp Neurol 191:133–151. doi:
10.1002/cne.901910108
Granda R, Crossland WJ (1989) GABA-like immunoreactivity of
neurons in the chicken diencephalon and mesencephalon.
J Comp Neurol 287:455–469. doi:10.1002/cne.902870405
Grantyn R, Perouansky M, Lin HD, Hablitz JJ (1987) Glutamate-
induced ionic currents in cultured neurons from the rat superior
colliculus. Brain Res 420:182–187. doi:10.1016/0006-8993(87)
90258-7
Gray EG (1959) Axo-somatic and axo-dendritic synapses of the
cerebral cortex. an electron-microscopy study. J Anat Lond
93:420–433
Guillery RW (2002) On counting and counting errors. J Comp Neurol
447:1–7. doi:10.1002/cne.10221
Gundersen HJ (1978) Estimators of the number of objects per area
unbiased by edge effects. Microsc Acta 81:107–117
Hartveit E, Heggelund P (1990) Neurotransmitter receptors mediating
excitatory input to cells in the cat lateral geniculate nucleus. II.
Nonlagged cells. J Neurophysiol 63:1361–1372
Heggelund P, Hartveit E (1990) Neurotransmitter receptors mediating
excitatory input to cells in the cat lateral geniculate nucleus I.
Lagged cells. J Neurophysiol 63:1347–1360
Heier P (1948) Fundamental principles in the structure of the brain.
A study of the brain of Petromyzon fluviatilis. Acta Anat Suppl
(Basel) 8:1–213
Henke H, Schenker TM, Cuenod M (1976) Effects of retinal ablation
on uptake of glutamate, glycine, GABA, proline and choline in
pigeon tectum. J Neurochem 26:131–134
Hestrin S (1992) Developmental regulation of NMDA receptor
mediated synaptic currents at a central synapse. Nature 357:686–
689. doi:10.1038/357686a0
Houser CR, Lee M, Vaughn JE (1983) Immunocytochemical
localisation of glutamic acid decarboxylase in normal and
deafferented colliculus evidence for reorganization of c-amino-
butyric acid synapses. J Neurosci 3:2030–2042
Huerta MF, Harting JK (1984) The mammalian superior colliculus:
studies of its morphology and connections. In: Vanagas H (ed)
Comparative neurology of the optic tectum. Plenum Press, New
York, pp 687–773
Ingham CA, Guldner FH (1981) Identification and morphometric
evaluation of the synapses of optic nerve afferents in the optic
tectum of the axolotl (Ambystoma mexicanum). Cell Tissue Res
214:593–611. doi:10.1007/BF00233499
Ito H, Butler AB, Ebbesson SOE (1980) An ultrastructural study
of the normal synaptic organization of the optic tectum and
degenerating tectal afferents from retina, telencephalon and
contralateral tectum in a teleost, Holocentrus rufus. J Comp
Neurol 191:639–659. doi:10.1002/cne.901910409
Iwahori N, Kawasaki T, Baba J (1999) Neuronal organization of the
optic tectum in the river lamprey, Lampetra japonica: A Golgi
study. J Hirnforsch 39:409–424
Janvier P (1996) Early Vertebrates. Oxford University Press, Oxford
Brain Struct Funct
123

Jeon CJ, Gurski MR, Mize RR (1997) Glutamate containing neurons
in the cat superior colliculus revealed by immunocytochemistry.
Vis Neurosci 14:387–393
Johnston JB (1902) The brain of petromyzon. J Comp Neurol 12:1–
86. doi:10.1002/cne.910120102
Kageyama GH, Meyer RL (1989) Glutamate-immunoreactivity in the
retina and optic tectum of goldfish. Brain Res 503:118–127. doi:
10.1016/0006-8993(89)91711-3
Kalloniatis M, Fletcher EL (1993) Immunocytochemical localisation
of the amino-acid neurotransmitters in the chicken retina.
J Comp Neurol 336:174–193. doi:10.1002/cne.903360203
Karamian AI, Vesselkin NP, Agayan AL (1984) Electrophysiological
and behavioural studies of the optic tectum in Cyclostomes. In:
Vanegas H (ed) Comparative neurology of the optic tectum.
Plenum Press, New York, pp 15–32
Kenigfest NB, Reperant J, Rio JP, Belelkova MG, Ward R, Vesselkin
NP, Miceli D, Herbin M (1998) Retinal and cortical afferents
to the dorsal lateral geniculate nucleus of the turtle, Emysorbicularis: a combined axonal tracing glutamate and GABA
immunocytochemical electron microscopic study. J Comp
Neurol 391:470–490. doi:10.1002/(SICI)1096-9861(19980222)
391:4\470::AID-CNE5[3.0.CO;2-Y
Kennedy MC, Rubinson K (1977) Retinal projections in larval,
transforming and adult sea lamprey, Petromyzon marinus.
J Comp Neurol 171:465–468. doi:10.1002/cne.901710404
Kennedy MC, Rubinson K (1984) Development and structure of the
lamprey optic tectum. In: Vanegas H (ed) Comparative neurol-
ogy of the optic tectum. Plenum Press, New York, pp 1–13
Kim YI, Dudek FE (1991) Intracellular electrophysiological study of
the suprachiasmaticus nucleus neurons in rodents: excitatory
synaptic mechanisms. J Physiol 444:269–287
Kosareva AA (1980) Retinal projections in lamprey (Lampetrafluviatilis). J Hirnforsch 21:243–256
Kosareva AA, Vesselkin NP, Ermakova TV (1977) Retinal projection
in the lamprey Lampetra fluviatilis, as revealed by horseradish
peroxidase transport along the optic nerve. Zh Evol Biokhim
Fisiol Leningrad 13:405–407 In Russian
Kwon YH, Esguerra M, Sur M (1991) NMDA and non NMDA
receptors mediated visual geniculate nucleus. J Neurophysiol
66:414–428
Langsdon RB, Freeman JA (1986) Antagonists of glutamatergic
neurotransmission block retinotectal transmission in goldfish.
Brain Res 398:169–174. doi:10.1016/0006-8993(86)91263-1
Lazar GY (1984) Structure and connections of the frog optic tectum.
In: Vanegas H (ed) Comparative neurology of the optic tectum.
Plenum Press, New York, pp 185–210
Leghissa S (1962) La structure della corteccia mesencefalica dei
ciclostomi selaci ed urodeli. Att Accad Sci Ist Bologna Cl Sci Fis
Ser II 9:123–152
Lund R (1969) Synaptic patters of the superficial layers of the
superior colliculus of the rat. J Comp Neurol 135:179–208. doi:
10.1002/cne.901350205
Lund R (1972) Synaptic patters of the superficial layers of the
superior colliculus of the monkey, Macaca mulatta. Exp Brain
Res 15:194–211. doi:10.1007/BF00235582
Mathers LH Jr (1977) Retinal and visual cortical projections to the
superior colliculus of the rabbit. Exp Neurol 57:698–712. doi:
10.1016/0014-4886(77)90103-0
Mayhew TM (1979) Sterological approach to the study of synapse
morphometry with particular regard to estimating number in a
volume and on the surface. J Neurocytol 8:121–138. doi:
10.1007/BF01175556
Medina M, Reperant J, Dufour S, Le Belle N, Miceli D (1994) The
distribution of GABA immunoreactive neurons in the brain of
the silver eel (Anguilla anguilla). Anat Embryol (Berl) 189:25–
39. doi:10.1007/BF00193127
Medina M, Reperant J, Ward R, Miceli D (2004) The centrifugal
visual system of Crocodylus niloticus: a hodological, histochem-
ical and immunocytochemical study. J Comp Neurol 468:65–85.
doi:10.1002/cne.10959
Meek J (1981) A Golgi-electron microscopic study of goldfish optic
tectum. I. Description of afferents, cell types and synapses.
J Comp Neurol 199:149–173. doi:10.1002/cne.901990202
Mesulam MM (1982) Principles of horseradish peroxydase neurohis-
tochemistry and their applications for tracing pathways–axonal
transport, enzyme histochemistry and light microscopic analysis.
In: Mesulam MM (ed) Tracing Neuronal Connections with
Horseradish Peroxydase. John Wiley & Sons, New York, pp 1–
151
Miceli D, Reperant J, Rio JP, Hains P, Medina M (2002) Serotonin
immunoreactivity in the retinal projecting isthmo-optic nucleus
and evidence of brainstem raphe connections in the pigeon.
Brain Res 958:122–129. doi:10.1016/S0006-8993(02)03515-1
Miceli D, Reperant J, Ward R, Rio JP, Jay B, Medina M, Kenigfest
NB (2008) Fine structure of the visual dorsolateral anterior
thalamic nucleus of the pigeon (Columba livia): a hodological
and GABA-immunochemical study. J Comp Neurol 507:1351–
1378. doi:10.1002/cne.21635
Mize RR (1983) Patterns of convergence and divergence of retinal
and cortical synaptic terminals in the cat superior colliculus. Exp
Brain Res 51:88–96. doi:10.1007/BF00236806
Mize RR (1988) Immunocytochemical localization of gamma-
aminobutyric acid (GABA) in the cat superior colliculus.
J Comp Neurol 276:169–187. doi:10.1002/cne.902760203
Mize RR (1992) The organization of GABAergic neurons in the
mammalian superior colliculus. Prog Brain Res 90:219–248. doi:
10.1016/S0079-6123(08)63616-X
Mize RR, Butler GD (1995) N-methyl-D-aspartate receptor localiza-
tion identified using antibody immunocytochemistry, is primarily
postsynaptic in the cat superior colliculus. Soc Neurosci Abstr
21:1618
Mize RR, Butler GD (1996) Postembedding immunocytochemistry
demonstrates directly that both retinal and cortical terminals in
the cat superior colliculus are glutamate immunoreactive.
J Comp Neurol 371:633–648. doi:10.1002/(SICI)1096-9861
(19960805)371:4\633::AID-CNE11[3.0.CO;2-K
Mize RR, Spencer RF, Sterling P (1982) Two types of GABA-
accumulating neurons in the superficial gray layer of the cat
superior colliculus. J Comp Neurol 206:180–192. doi:10.1002/
cne.902060207
Mize RR, Jeon CJ, Hamada OL, Spencer RF (1991) Organization of
neurons labelled by antibodies to gamma-aminobutyric acid
(GABA) in the superior colliculus of the Rhesus monkey. Vis
Neurosci 6:75–92
Mize RR, Whitworth RH, Nunes-Cardozo B, van der Want (1994)
Ultrastructural organization of GABA in the rabbit superior
colliculus revealed by quantitative postembedding immunocy-
tochemistry. J Comp Neurol 341:273–287. doi:10.1002/cne.
903410211
Montero VM (1990) Quantitative immunogold analysis reveals high
glutamate levels in synaptic terminals of retino-geniculate
cortico-geniculate and geniculo-cortical axons in the cat. Vis
Neurosci 4:437–443
Montero VM (1994) Quantitative immunogold evidence for enrich-
ment of glutamate but not aspartate in synaptic terminals of
retino-geniculate, geniculo-cortical and cortico-geniculate axons
in the cat. Vis Neurosci 11:675–681
Montero VM, Wenthold RJ (1989) Quantitative immunogold
analysis reveals high glutamate levels in retinal and cortical
synaptic terminals in the lateral geniculate nucleus of the
macaque. Neuroscience 31:639–647. doi:10.1016/0306-4522
(89)90429-6
Brain Struct Funct
123

Morino P, Bahro M, Cuenod M, Streit P (1991) Glutamate like
immunoreactivity in the pigeon optic tectum and effect of retinal
ablation. Eur J NeuroSci 3:366–378. doi:10.1111/j.1460-9568.
1991.tb00824.x
Mugnaini E, Oertel WH (1985) An atlas of the distribution of
Gabaergic neurons and terminals in the rat CNS as revealed by
GAD immunhistochemistry. In: Bjorklund A, Hokfeld T (eds)
Handbook of Chemical Neuroanatomy. GABA and Neuropep-
tides in the CNS, vol. 4, part I. Elsevier, Amsterdam, New York,
Oxford, pp 436–622
Murphy PC, Salt TE, Sillito AM (1989) Contribution of both NMDA
and non NMDA receptors to transmission of the visual input to X
and Y cells in the anesthetized feline dorsal lateral geniculate
nucleus. Proc Physiol Soc J Physiol 415:46
Murray M, Edwards MA (1982) A quantitative study of the
reinnervation of the godfish optic tectum following optic nerve
crush. J Comp Neurol 209:363–373. doi:10.1002/cne.902090406
Nieuwenhuys R (1977) The brain of the lamprey in a comparative
perspective. Ann N Y Acad Sci 299:97–145. doi:10.1111/j.1749-
6632.1977.tb41902.x
Nieuwenhuys R, Nicholson C (1998) Lampreys, Petromyzontidae. In:
Nieuwenhuys R, Ten Donkelaar HJ, Nicholson C (eds) The
central nervous system of vertebrates, vol 1, chap 10. Springer,
New York, pp 397–495
Northcutt RG, Przybylski RJ (1973) Retinal projection in the lamprey
Petromyzon marinus L. Anat Rec 175:400
Northcutt RG, Wicht H (1997) Afferent and efferent connections of
the lateral and medial pallia of the silver lamprey. Brain Behav
Evol 49:1–19. doi:10.1159/000112978
Ortega F, Hennequet L, Sarria R, Streit P, Grandes P (1995) Changes
in the pattern of glutamate-like immunoreactivity in rat superior
colliculus following retinal and visual cortical lesions. Neuro-
science 67:125–134. doi:10.1016/0306-4522(95)00057-P
Ottersen OP (1987) Post-embedding light- and electron microscopic
immunocytochemistry of amino acids: description of a new
model allowing identical conditions for specificity testing and
tissue processing. Exp Brain Res 69:167–174. doi:10.1007/
BF00247039
Ottersen OP (1989) Quantitative electron microscopic immunocyto-
chemistry of neuroactive amino acids. Anat Embryol (Berl)
180:1–15. doi:10.1007/BF00321895
Ottersen OP, Storm-Mathisen J (1984) Glutamate and GABA-
containing neurons in the mouse and the rat brain as demon-
strated with a new immunocytochemistry technique. J Comp
Neurol 229:374–392. doi:10.1002/cne.902290308
Papadopoulos GC, Parnavelas JG (1990) Distribution and synaptic
organization of serotoninergic and noradrenergic axons in the
lateral geniculate nucleus of the rat. J Comp Neurol 294:345–
355. doi:10.1002/cne.902940304
Pasik P, Pasik T, Holstein GR (1988) Serotonin immunoreactivity in
the monkey lateral geniculate nucleus. Exp Brain Res 69:662–
666. doi:10.1007/BF00247318
Perouansky M, Grantyn R (1990) Is GABA release modulated by
presynaptic excitatory amino-acid receptors? Neurosci Lett
113:292–297. doi:10.1016/0304-3940(90)90600-E
Peters A, Palay SL, de Webster HFF (1970) The fine structure of the
nervous system. The cells and their processes. Harper and Row,
New York
Peyrichoux J, Pierre J, Reperant J, Rio JP (1986) Fine structure of the
optic fiber termination layer in the tectum of the teleost Rutilus:
a stereological and morphometric study. J Comp Neurol
246:364–381. doi:10.1002/cne.902460306
Pfister C (1971) Die Matrixentwicklung in Mes- und Rhombenceph-
alon von Lampetra planeri (Bloch) (Cyclostomata) im Verlaufe
des Individualzyklus. J Hirnforsch 13:377–383
Pierre J, Reperant J, Ward R, Vesselkin NP, Rio JP, Miceli D,
Kratskin I (1992) The serotoninergic system of the brain of the
lamprey, Lampetra fluviatilis: an evolutionary perspective.
J Chem Neuroanat 5:195–219. doi:10.1016/0891-0618(92)
90046-S
Pinard R, Benfares J, Lanoir J (1991) Electron microscopic study of
GABA-immunoreactive neural process in the superficial gray
layer of the rat superior colliculus their relationship with
degenerating retinal nerve endings. J Neurocytol 20:262–276.
doi:10.1007/BF01235544
Polenova OA, Vesselkin NP (1993) Olfactory and no olfactory
projections in the river lamprey (Lampetra fluviatilis) telenceph-
alon. J Hirnforsch 34:261–279
Reperant J, Angaut P (1977) The retinotectal projections in the
pigeon. An experimental optical and electron microscope study.
Neuroscience 2:119–140. doi:10.1016/0306-4522(77)90073-2
Reperant J, Vesselkin NP, Ermakova TV, Kenigfest NB, Kosareva
AA (1980) Radioautographic evidence for both orthograde and
retrograde axonal transport of label compounds after intraocular
injection of [3H] proline in the lamprey (Lampetra fluviatilis).
Brain Res 200:179–183. doi:10.1016/0006-8993(80)91105-1
Reperant J, Peyrichoux J, Rio JP (1981) Fine structure of the
superficial layers of the viper optic tectum. A Golgi and electron
microscopic study. J Comp Neurol 199:393–417. doi:10.1002/
cne.901990306
Reperant J, Vesselkin NP, Kenigfest NB, Miceli D, Rio JP (1982)
Bidirectional axonal and transcellular transport of [3H] adeno-
sine from the lamprey retina. Brain Res 246:305–310. doi:
10.1016/0006-8993(82)91181-7
Reperant J, Vesselkin NP, Miceli D, Kenigfest NB, Rio JP (1985) A
comparative radioautographic study of the bidirectional axonal
and transcellular transport of different amino acids and sugars
in the lamprey visual system. Brain Res 348:348–354. doi:
10.1016/0006-8993(85)90455-X
Reperant J, Miceli D, Vesselkin NP, Molotchnikoff S (1989) The
centrifugal visual system of Vertebrates. A century-old, research
reviewed. Int Rev Cytol 118:115–171. doi:10.1016/S0074-7696
(08)60874-8
Reperant J, Vesselkin NP, Kenigfest NB, Rio JP (1990) Modern
concepts of structural evolution of the nervous system exempli-
fied by studies on the visual system of vertebrates. Zh Evol
Biokhim Fisiol Leningrad 26:514–535 In Russian
Reperant J, Rio JP, Ward R, Wasowicz M, Miceli D, Medina M,
Pierre J (1997) Enrichment of glutamate-like immunoreactivity
in the retinotectal terminals of the viper Vipera aspis: an electron
microscope quantitative immunogold study. J Chem Neuroanat
12:267–280. doi:10.1016/S0891-0618(97)00018-5
Reperant J, Ward R, Miceli D, Rio JP, Medina M, Kenigfest NB,
Vesselkin NP (2006) The centrifugal visual system of verte-
brates: a comparative analysis of its functional anatomical
organization. Brain Res Brain Res Rev 52:1–57. doi:10.1016/j.
brainresrev.2005.11.008
Reperant J, Medina M, Ward R, Miceli D, Kenigfest NB, Rio JP,
Vesselkin NP (2007) The evolution of the centrifugal visual
system of vertebrates. A cladistic analysis and new hypotheses.
Brain Res Brain Res Rev 53:161–197. doi:10.1016/j.brainresrev.
2006.08.004
Reubi JC (1980) Comparative study of the release of glutamate and
GABA newly synthetized from glutamate in various regions
of the central nervous system. Neuroscience 5:2145–2150. doi:
10.1016/0306-4522(80)90130-X
Rio JP, Dalil N, Kirpitchnikova E, Vesselkin NP, Versaux-Botteri C,
Reperant J (1992) Recherche immunocytochimique sur les
mediateurs des neurones visuels centrifuges de la lamproie
(Lampetra fluviatilis). CR Acad Sci Paris 315:501–511
Brain Struct Funct
123

Rio JP, Vesselkin NP, Kirpitchnikova E, Kenigfest NB, Versaux-
Botteri C, Reperant J (1993) Presumptive GABAergic centrif-
ugal input to the lamprey retina: a double-labeling study with
axonal tracing and GABA-immunocytochemistry. Brain Res
600:9–19. doi:10.1016/0006-8993(93)90395-4
Rio JP, Reperant J, Herbin M, Miceli D (1995) Distribution of GABA
immunoreactivity in the retino-recipient layer of the viper optic
tectum. A light and electronic microscope quantitative study.
Anat Embryol (Berl) 191:251–265. doi:10.1007/BF00187824
Rio JP, Vesselkin NP, Reperant J, Kenigfest NB, Miceli D, Adanina
V (1996) Retinal and non-retinal inputs upon retinopetal RMA
neurons in the lamprey: a light and electron microscopic study
combining HRP axonal tracing and GABA immunocytochem-
istry. J Chem Neuroanat 12:51–57. doi:10.1016/S0891-0618
(96)00177-9
Rio JP, Reperant J, Miceli D, Medina M, Kenigfest-Rio NB (2002)
Serotoninergic innervation of the isthmo-optic nucleus of the
pigeon centrifugal visual system. An immunocytochemical elec-
tron microscopic study. Brain Res 924:127–131. doi:10.1016/
S0006-8993(01)03262-0
Roberts WA, Eaton SA, Salt TE (1991) Excitatory amino acid
receptors mediate synaptic responses to visual stimuli in superior
colliculus neurons of the rat. Neurosci Lett 129:161–164. doi:
10.1016/0304-3940(91)90451-X
Robertson B, Saitoh K, Menard A, Grillner S (2006) Afferents of the
lamprey optic tectum with special reference to the GABA input:
combined tracing and immunohistochemical study. J Comp
Neurol 499:106–119. doi:10.1002/cne.21078
Robertson B, Auclair F, Menard A, Grillner S, Dubuc R (2007)
GABA distribution in lamprey is phylogenetically conserved.
J Comp Neurol 503:47–63. doi:10.1002/cne.21348
Rodicio MC, Pombal MA, Anadon R (1995) Early development and
organization of the retinopetal system in larval sea lamprey,
Petromyzon marinus L. An HRP study. Anat Embryol (Berl)
192:517–526. doi:10.1007/BF00187182
Rubinson K (1990) The developing visual system and metamorphoses
in the lamprey. J Neurobiol 21:1123–1135. doi:10.1002/neu.
480210715
Rubinson K, Kennedy MC (1979) The organization of the optic
tectum in larval, transforming and adult sea lamprey, Petromy-zon marinus. In: Freeman RD (ed) Developmental neurobiology
of vision. Plenum Press, New York, pp 359–369
Sakurai T, Okada Y (1992) Selective reduction of glutamate in the rat
superior colliculus and dorsal lateral geniculate nucleus after
contralateral enucleation. Brain Res 573:197–203. doi:10.1016/
0006-8993(92)90763-Y
Sakurai T, Miyamoto T, Okada Y (1990) Reduction of glutamate content
in rat superior colliculus after retino-tectal denervation. Neurosci
Lett 109:299–303. doi:10.1016/0304-3940(90)90011-W
Sandberg M, Jacobson I, Hamberger A (1982) Release of endogenous
amino-acids from superior colliculus of the rabbit. In vitro
studies after retinal ablation. J Neurochem 40:917–921. doi:
10.1111/j.1471-4159.1983.tb08074.x
Schmidt M (1991) Mediation of visual responses in the nucleus of the
optic tract in cats and rats by excitatory amino-acid receptors.
Neurosci Res 12:111–121. doi:10.1016/0168-0102(91)90104-7
Schober W (1964) Vergleichend-anatomische untersuchungen am
Gehirn der larven und adulten Tiere von Lampetra fluviatilis(Linne 1758) und Lampetra planeri (Bloch, 1784). J Hirnforsch
7:107–209
Schonitzer K, Hollander H (1984) Retinotectal terminals in the
superior colliculus of the rabbit: A light and electron micro-
scopic analysis. J Comp Neurol 223:153–162. doi:10.1002/
cne.902230202
Seguela P, Geffard M, Buijs RM, Le Moal ML (1984) Antibodies
against gamma-aminobutyric acid: specificity studies and
immunocytochemical results. Proc Natl Acad Sci USA
81:3888–3892. doi:10.1073/pnas.81.12.3888
Sherman SM (2004) Interneurons and triadic circuitry of the
thalamus. Trends Neurosci 27:670–675. doi:10.1016/j.tins.2004.
08.003
Sherry DM, Ulshafer RJ (1992) Neurotransmitter-specific identifica-
tion and characterization of neurons in the all-cone retina of
Anolis carolinensis. II. glutamate and aspartate. Vis Neurosci
9:313–323
Shibata S, Liou SY, Ueki S (1986) Influence of excitatory amino-acid
receptor antagonists and baclofen on synaptic transmission in
optic nerve of the suprachiasmatic nucleus in slices of rat
hypothalamus. Neuropharmacology 25:403–409. doi:10.1016/
0028-3908(86)90235-2
Sillito AM, Murphy PC, Salt TE (1990a) The contribution of the non-
NMDA group of excitatory amino-acid receptors to retinogen-
iculate transmission in the cat. Neuroscience 34:273–280. doi:
10.1016/0306-4522(90)90137-S
Sillito AM, Murphy PC, Salt TE, Moady CI (1990b) The dependence
of retinogeniculate transmission in the cat on NMDA receptors.
J Neurophysiol 63:347–355
Sterling P (1971) Receptive fields and synaptic organization of the
superficial gray layer of the cat superior colliculus. Vision Res
Suppl 3:309–328. doi:10.1016/0042-6989(71)90048-4
Sternberger LA, Hardy PH Jr, Cuculis J, Meyer HG (1970) The
unlabeled antibody enzyme method of immunochemistry: prep-
aration and properties of soluble antigen-antibody complex
(horseradish peroxydase-antiperoxydase) and its use in identifi-
cation of spirochetes. J Histochem Cytochem 18:315–334
Storm-Mathisen J, Ottersen OP (1990) Immunocytochemistry of
glutamate at the synaptic level. J Histochem Cytochem
38(1):733–743
Streit P, Knecht E, Reubi JC, Hunt SP, Cuenod M (1978) GABA
specific presynaptic dendrites in the pigeon optic tectum: a high-
resolution autoradiographic study. Brain Res 149:204–210. doi:
10.1016/0006-8993(78)90599-1
Szekely G, Setalo G, Gy Lazar (1973) Fine structure of the frog’s
optic tectum: optic fibre termination layers. J Hirnforsch 14:189–
225
Tigges M, Tigges J (1975) Presynaptic dendrite cells and two other
classes of neurons in the superficial layers of the superior
colliculus of the chimpanzee. Cell Tissue Res 162:279–295. doi:
10.1007/BF00220175
Tigges M, Tigges J, Luttrell GL, Frazier CM (1973) Ultrastructural
changes in the superficial layers of the superior colliculus in
Galago crassicaudatus (Primates) after eye enucleation. Z
Zellforsch 140:291–307. doi:10.1007/BF00307019
Toggenburger G, Felix D, Cuenod M, Henke H (1982) In vivo release
of endogenous b-alanine, GABA and glutamate and electrophys-
iological effect of b-alanine in pigeon optic tectum. J Neurochem
39:176–183. doi:10.1111/j.1471-4159.1982.tb04716.x
Tombol T (1998) Golgi and electron-microscopic Golgi-GABA
immunostaining study of the avian optic tectum. Acta Anat
(Basel) 162:209–225. doi:10.1159/000046436
Tombol T, Nemeth A (1999) Direct connections between dendrite
terminals of tectal ganglion cells and glutamate-positive termi-
nals of presumed optic fibres in layers 4–5 of the optic tectum of
Gallus domesticus. A light-and electron microscopic study.
Neurobiology 7:45–67
Tombol T, Eyre M, Zayats N, Nemeth A (2003) The ramifications and
terminals of optic fibres in layers 2 and 3 of the avian optic
tectum: a Golgi and light and electron microscopic anterograde
tracer study. Cells Tissues Organs 175:202–222. doi:10.1159/
000074942Uchiyama H (1989) Centrifugal pathways to the retina: influence of
the tectum. Vis Neurosci 3:183–206
Brain Struct Funct
123

Underwood EE (1970) Quantitative stereology. Addison-Wesly,
Reading
Valverde F (1973) The neuropil in superficial layers of the superior
colliculus of the mouse. Z Anat Entwickl Gesch 142:117–147.
doi:10.1007/BF00519719
van Deusen EB, Meyer RL (1988) Differential effects of KA/QA,
NMDA and APB receptor antagonists on optic and toral afferent
stimulated activity on goldfish tectal neurons in vitro. Soc
Neurosci Abstr 14:96
Vanegas H, Ebbesson SOE, Laufer M (1984) Morphological aspects
of the teleostean optic tectum. In: Vanegas H (ed) Comparative
neurology of the optic tectum. Plenum Press, New York, pp 93–
120
Veenman CL, Reiner A (1994) The distribution of GABA-containing
perikarya, fibres and terminals in the forebrain and midbrain of
the pigeon, with particular reference to the basal ganglion and its
projection targets. J Comp Neurol 339:209–250
Vesselkin NP, Ermakova TV, Reperant J, Kosareva AA, Kenigfest
NB (1980) The retinofugal and retinopetal system in Lampetrafluviatilis. An experimental study using radioautographic and
HRP methods. Brain Res 195:453–460
Vesselkin NP, Reperant J, Kenigfest NB, Miceli D, Ermakova TV,
Rio JP (1984) An anatomical and electrophysiological study of
the centrifugal visual system in the lamprey (Lampetra fluvia-tilis). Brain Res 292:41–56
Vesselkin NP, Rio JP, Reperant J, Kenigfest NB, Adanina VO (1996)
Retinopetal projections in lampreys. Brain Behav Evol 48:277–
286
Vrensen G, de Groot D (1977) Quantitative aspects of the synaptic
organization of the superior colliculus in control and dark-reared
rabbits. Brain Res 134:417–428
Wang BL, Larsson LI (1985) Simultaneous demonstration of multiple
antigens by indirect fluorescence or immunogold staining: novel
light and electron microscopical double and triple staining
method employing primary antibodies from the same species.
Histochem 83:47–56
Ward R, Reperant J, Miceli D (1991) The centrifugal visual system:
what can comparative studies tell us about evolution and
possible function? In: Bagnoli P, Hodos W (eds) The changing
visual system. Plenum Press, New York, pp 61–76
Weibel ER (1979) Stereological methods. I. Practical methods for
biological morphometry. Academic Press, London
Williams RW, Rakic P (1988) Three-dimensionnal counting an
accurate and direct method to estimate numbers of cells in
sectioned material. J Comp Neurol 278:344–352
Brain Struct Funct
123




















