
Early development of the retina and pineal complex in the sea lamprey: comparative...
16
Early Development of the Retina and Pineal Complex in the Sea Lamprey: Comparative Immunocytochemical Study MIGUEL MELE ´ NDEZ-FERRO, 1 BEGON ˜ A VILLAR-CHEDA, 1 XESU ´ S MANOEL ABALO, 1 EMMA PE ´ REZ-COSTAS, 1 ROLANDO RODRI ´ GUEZ-MUN ˜ OZ, 2 WILLEM J. DEGRIP, 3 JULIA ´ N YA ´ N ˜ EZ, 4 MARI ´ A CELINA RODICIO, 1 AND RAMO ´ N ANADO ´ N 1 * 1 Department of Fundamental Biology, Faculty of Biology, University of Santiago de Compostela, 15782 Santiago de Compostela, Spain 2 Department of Biology of Organisms and Systems, Faculty of Biology, University of Oviedo, 33006 Oviedo, Spain 3 Department of Biochemistry, Nijmegen Centre for Molecular Life Sciences, University of Nijmegen, 6500 HB Nijmegen, The Netherlands 4 Department of Cell and Molecular Biology, Faculty of Sciences, University of A Corun ˜a, 15071 A Corun ˜ a, Spain ABSTRACT Lampreys have a complex life cycle, with largely differentiated larval and adult periods. Despite the considerable interest of lampreys for understanding vertebrate evolution, knowledge of the early development of their eye and pineal complex is very scarce. Here, the early immu- nocytochemical organization of the pineal complex and retina of the sea lamprey was studied by use of antibodies against proliferating cell nuclear antigen (PCNA), opsin, serotonin, and -aminobutyric acid (GABA). Cell differentiation in the retina, pineal organ, and habenula begins in prolarvae, as shown by the appearance of PCNA-negative cells, whereas differentiation of the parapineal vesicle was delayed until the larval period. In medium-sized to large larvae, PCNA- immunoreactive (-ir) cells were numerous in regions of the lateral retina near the differentiated part of the larval retina (central retina). A late-proliferating region was observed in the right habenula. Opsin immunoreactivity appears in the pineal vesicle of early prolarvae and 3 or 4 days later in the retina. In the parapineal organ, opsin immunoreactivity was observed only in large larvae. In the pineal organ, serotonin immunoreactivity was first observed in late prolarvae in photoreceptive (photoneuroendocrine) cells, whereas only a few of these cells appeared in the parapineal organ of large larvae. No serotonin immunoreactivity was observed in the larval retina. GABA immunoreactivity appeared earlier in the retina than in the pineal complex. No GABA-ir perikaryon was observed in the retina of larval lampreys, although a few GABA-ir centrifugal fibers innervate the inner retina in late prolarvae. First GABA-ir ganglion cells occur in the pineal organ of 15–17 mm larvae, and their number increases during the larval period. The only GABA-ir structures observed in the parapineal ganglion of larvae were afferent fibers, which appeared rather late in development. The time sequence of development in these photoreceptive structures is rather different from that observed in teleosts and other vertebrates. This suggests that the unusual development of the three photoreceptive organs in lampreys reflects special- ization for their different functions during the larval and adult periods. J. Comp. Neurol. 442: 250 –265, 2002. © 2001 Wiley-Liss, Inc. Indexing terms: photoreceptors; opsin; GABA; cell proliferation; photoneuroendocrine cells; Agnathans; Petromyzon marinus Grant sponsor: Spanish Science and Technology Ministry; Grant num- ber: BMC2000-0283; Grant sponsor: Xunta de Galicia; Grant number: PGIDT99BIO20002; Grant number: PGIDT99PXI20003B. *Correspondence to: Ramo ´n Anado ´n, Department of Fundamental Biol- ogy, Faculty of Biology, University of Santiago de Compostela, 15782 Santiago de Compostela, Spain. E-mail: [email protected] Received 13 February 2001; Revised 12 July 2001; Accepted 20 October 2001 THE JOURNAL OF COMPARATIVE NEUROLOGY 442:250 –265 (2002) © 2001 WILEY-LISS, INC. DOI 10.1002/cne.10090
Transcript of Early development of the retina and pineal complex in the sea lamprey: comparative...

Early Development of the Retina andPineal Complex in the Sea Lamprey:
Comparative Immunocytochemical Study
MIGUEL MELENDEZ-FERRO,1 BEGONA VILLAR-CHEDA,1 XESUS MANOEL ABALO,1
EMMA PEREZ-COSTAS,1 ROLANDO RODRIGUEZ-MUNOZ,2 WILLEM J. DEGRIP,3
JULIAN YANEZ,4 MARIA CELINA RODICIO,1AND RAMON ANADON1*
1Department of Fundamental Biology, Faculty of Biology, University of Santiago deCompostela, 15782 Santiago de Compostela, Spain
2Department of Biology of Organisms and Systems, Faculty of Biology, University ofOviedo, 33006 Oviedo, Spain
3Department of Biochemistry, Nijmegen Centre for Molecular Life Sciences, University ofNijmegen, 6500 HB Nijmegen, The Netherlands
4Department of Cell and Molecular Biology, Faculty of Sciences, University of A Coruna,15071 A Coruna, Spain
ABSTRACTLampreys have a complex life cycle, with largely differentiated larval and adult periods.
Despite the considerable interest of lampreys for understanding vertebrate evolution, knowledgeof the early development of their eye and pineal complex is very scarce. Here, the early immu-nocytochemical organization of the pineal complex and retina of the sea lamprey was studied byuse of antibodies against proliferating cell nuclear antigen (PCNA), opsin, serotonin, and�-aminobutyric acid (GABA). Cell differentiation in the retina, pineal organ, and habenula beginsin prolarvae, as shown by the appearance of PCNA-negative cells, whereas differentiation of theparapineal vesicle was delayed until the larval period. In medium-sized to large larvae, PCNA-immunoreactive (-ir) cells were numerous in regions of the lateral retina near the differentiatedpart of the larval retina (central retina). A late-proliferating region was observed in the righthabenula. Opsin immunoreactivity appears in the pineal vesicle of early prolarvae and 3 or 4 dayslater in the retina. In the parapineal organ, opsin immunoreactivity was observed only in largelarvae. In the pineal organ, serotonin immunoreactivity was first observed in late prolarvae inphotoreceptive (photoneuroendocrine) cells, whereas only a few of these cells appeared in theparapineal organ of large larvae. No serotonin immunoreactivity was observed in the larvalretina. GABA immunoreactivity appeared earlier in the retina than in the pineal complex. NoGABA-ir perikaryon was observed in the retina of larval lampreys, although a few GABA-ircentrifugal fibers innervate the inner retina in late prolarvae. First GABA-ir ganglion cells occurin the pineal organ of 15–17 mm larvae, and their number increases during the larval period. Theonly GABA-ir structures observed in the parapineal ganglion of larvae were afferent fibers, whichappeared rather late in development. The time sequence of development in these photoreceptivestructures is rather different from that observed in teleosts and other vertebrates. This suggeststhat the unusual development of the three photoreceptive organs in lampreys reflects special-ization for their different functions during the larval and adult periods. J. Comp. Neurol. 442:250–265, 2002. © 2001 Wiley-Liss, Inc.
Indexing terms: photoreceptors; opsin; GABA; cell proliferation; photoneuroendocrine cells;
Agnathans; Petromyzon marinus
Grant sponsor: Spanish Science and Technology Ministry; Grant num-ber: BMC2000-0283; Grant sponsor: Xunta de Galicia; Grant number:PGIDT99BIO20002; Grant number: PGIDT99PXI20003B.
*Correspondence to: Ramon Anadon, Department of Fundamental Biol-ogy, Faculty of Biology, University of Santiago de Compostela, 15782Santiago de Compostela, Spain. E-mail: [email protected]
Received 13 February 2001; Revised 12 July 2001; Accepted 20 October2001
THE JOURNAL OF COMPARATIVE NEUROLOGY 442:250–265 (2002)
© 2001 WILEY-LISS, INC.DOI 10.1002/cne.10090

Lampreys belong to an ancient lineage of vertebratesthat flourished before the appearance of the gnathos-tomes. Sea lampreys have a complex life cycle that startswith egg laying in a river. The embryonic period is fol-lowed by a nonfeeding prolarval phase and a micropha-gous larval period (several years long) in which the ani-mals are blind and live burrowing in the bed of the river inwhich they were born. Through a complex metamorphosis,larvae (ammocoetes) transform into young adult lampreysthat descend to the sea to feed parasitically on marinefishes, growing rapidly until they finally ascend the riverto breed. Sea lampreys die after breeding (Hardisty andPotter, 1971).
Lampreys show a primitive pineal complex, with bothpineal and parapineal photoreceptive organs (Studnicka,1893, 1905; Retzius, 1895; Collin, 1969; Meiniel and Col-lin, 1971; Cole and Youson, 1982; Samejima et al., 1989;Tamotsu et al., 1990, 1994; Pombal et al., 1999; Yanez etal., 1999). The pineal organ of adult lampreys consists of apineal vesicle and a long pineal stalk containing photore-ceptors and ganglion neurons. Ganglion neurons and somephotoreceptor-like cells project in the pineal tract, enter-ing the brain at the level of the posterior commissure(Samejima et al., 1989; Pombal et al., 1999). The pinealorgan has been shown to be light sensitive both in adultand in larval lampreys (Tamotsu and Morita, 1986). Theparapineal organ consists of a parapineal vesicle withphotoreceptor cells and a ganglion that is thought to be aspecialized part of the left habenula (Studnicka, 1905;Meiniel and Collin, 1971; Yanez et al., 1999).
As in other vertebrates, the eye retina of adult lampreyscontains several types of neural cell, including one to threetypes of photoreceptors, bipolar cells, horizontal cells, am-acrine cells, and ganglion cells (see Dickson and Graves,1981; Rubinson and Cain, 1989; Collin and Pottert, 2000).The lateral eyes of larval lampreys exhibit dual retinaldevelopment, with a central part containing a single typeof photoreceptor that appears early in larval developmentand a lateral retina that begins to grow in medium-sizedlarvae, expanding considerably in large larvae and form-ing a layered structure in transforming larvae. In thislateral retina, which gives rise to practically the wholeadult retina, photoreceptors do not begin differentiationuntil advanced metamorphic stages (see Dickson andGraves, 1981; De Miguel, 1984, 1989; De Miguel and Ana-don, 1987). The lamprey retina receives centrifugal fibersfrom the brain (De Miguel et al., 1989; Vesselkin et al.,1989).
Here we report a comparative study of the photorecep-tive organs in the early stages of lamprey development.Because lampreys are among the most ancient extantvertebrates, knowledge of the early organization of thelamprey pineal complex and retina may be valuable forunderstanding the evolution of vertebrate photoreceptiveorgans in general. Specifically, we investigated proliferat-ing cell nuclear antigen (PCNA), opsin, serotonin, and�-aminobutyric acid (GABA) immunoreactivities in thedeveloping retina and pineal complex of the sea lampreyPetromyzon marinus. PCNA, the � subunit of DNA poly-merase, is a useful marker of cells that have not left themitotic cycle in the retina and brain of teleosts and othervertebrates (Negishi et al., 1990a,b; Negishi and Wagner,1995; Wullimann and Puelles, 1999). Opsins, the primaryphotoreceptive molecules of vertebrate photoreceptors, arevery useful markers of photoreceptor cell differentiation(van Veen et al., 1984; Saha and Grainger, 1993; Robinson
et al., 1995). Synthesis of opsins starts very early duringphotoreceptor cell differentiation, a few days before theexpression of other phototransduction cascade proteins(Sears et al., 2000). Some antiopsin antibodies cross-reactwith both lateral eye and pineal photoreceptors in larvallampreys (Garcıa-Fernandez et al., 1997), making themuseful tools for study of these organs. Serotonin is alsoexpressed in some lamprey pineal photoreceptors ( Ta-motsu et al., 1990, 1994, 1997; Pombal et al., 1999). Theseserotonin-immunoreactive (-ir) cells are thought to syn-thesize melatonin and are a key part of the photoneuroen-docrine system (Tamotsu et al., 1990, 1997). GABA is atransmitter in lamprey photoreceptive organs. The pres-ence of GABAergic ganglion cells in the pineal organ ofadult lampreys has already been noted (Pombal et al.,1999). These cells project in the pineal tract, entering thebrain through the posterior commissure. Although theacetylcholinesterase reaction has been widely used fordemonstration of pineal ganglion cells in teleosts (Wake,1973; Korf, 1974), this reaction and choline acetyltrans-ferase immunocytochemistry label mainly photoreceptorsin the lamprey pineal complex (Pombal et al., 1999; Yanezet al., 1999), making them useless for the study of pinealganglion cell development. The parapineal organ of theadult lamprey does not contain GABAergic cells but re-ceives abundant GABAergic innervation (Yanez et al.,1999). The retina of adult lampreys contains GABAergiccells (Rio et al., 1996), although these were not observed inthe larval retina (Anadon et al., 1998). The lamprey retinareceives GABAergic centrifugal fibers both in adults andlarvae (Rio et al., 1996; Vesselkin et al., 1996; Anadon etal., 1998). Because GABA is one of the earliest neurotrans-mitters expressed in both brain and retina (Ostholm et al.,1988; Ekstrom and Ohlin, 1995), knowledge of the ontog-eny of GABA immunoreactivity in lamprey photoreceptiveorgans may be useful for understanding the evolution ofearly neural circuits.
MATERIALS AND METHODS
Animals
Late embryos (E10–13), prolarvae (P0–23), and larvae(9–120 mm in length) of the sea lamprey (Petromyzonmarinus L.) were used. Embryos and prolarvae originatedfrom in-vitro-fertilized eggs obtained from sexually ma-ture adult lampreys caught in the River Sella (northernSpain). Fertilized eggs were reared in the laboratory un-der appropriate conditions of darkness and temperature(16°C). Larval lampreys were caught in the River Ulla(northwestern Spain) and kept in an aerated aquariumuntil processing. All experiments were conducted in accor-dance with European Community guidelines on animalcare and experimentation to minimize pain and discom-fort.
Stages of embryos, prolarvae, and early larvae are indi-cated by age (e.g., E12 indicates 12 days after fertilization;P3 indicates 3 days posthatching, and so on; in our broods,hatching occurred 11–13 days after fertilization). In addi-tion, to classify prolarvae, we used the stages defined forsea lamprey by Piavis (1971): hatching (P0–1), pigmenta-tion (P2–3), gill cleft (P4–7), and burrowing (P8–23)stages. Larval stages (with the exception of larvae frombroods in which age is known) are categorized simply bytotal body length.
251RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY

Immunohistochemistry
The specimens were fixed by immersion in fixative un-der deep anesthesia with 0.1% tricaine methane sulfonate(MS-222; Sigma, St. Louis, MO). For GABA immunocyto-chemistry, animals (late embryos, prolarvae, and 9–107mm larvae) were fixed in cold (4°C), freshly prepared 5%glutaraldehyde in 0.1 M phosphate buffer, pH 7.4 (PB),overnight. After rinsing in PB, samples were cryopro-tected with 30% sucrose in PB, embedded in Tissue Tek(Sakura, Torrance, CA), and frozen using liquid nitrogen-cooled isopentane. Transverse or horizontal sections werecut on a cryostat and mounted on gelatin-subbed slides.The sections were processed for immunocytochemistry asfollows: Nonspecific binding sites in the sections wereblocked by incubating with 10% normal goat serum (Vec-tor Laboratories, Burlingame, CA). Then, the sectionswere incubated in a humid chamber at room temperaturewith a polyclonal anti-GABA antibody (Affiniti, Mamhead,United Kingdom), diluted 1:1,000, overnight. After rinsingin Tris-buffered saline (TBS; pH 7.4), the sections wereincubated with a goat anti-rabbit immunoglobulin (Sig-ma), diluted 1:100, for 1 hour. The sections were developedby the peroxidase-antiperoxidase (PAP) procedure (Stern-berger et al., 1970; the PAP complex was purchased fromSigma and diluted 1:400) using 0.6 mg/ml 3-3�-diaminobenzidine (DAB; Sigma) and 0.003% H2O2. Allantibody dilutions were carried out in TBS containing0.2% Triton X-100 as detergent and 3% normal goat se-rum. After developing, the sections were rinsed in TBS,dehydrated, and coverslipped.
For PCNA, opsin, and serotonin immunocytochemistry,the animals (late embryos, prolarvae, and 9–75 mm lar-vae for PCNA; prolarvae and 9–38 mm larvae for opsin,and prolarvae and 9–120 mm larvae for serotonin) werefixed in 4% paraformaldehyde in PB (serotonin) or Bouin’sfluid (PCNA and opsins), embedded in paraffin, cut intransversal or sagittal planes on a rotary microtome (8–10�m thickness), and mounted on gelatin-subbed slides. De-waxed sections were then immunostained by a proceduresimilar to that used for GABA, using either a polyclonalantiserotonin antibody (Incstar, Stillwater, MN; diluted1:1,000) or a mouse monoclonal anti-PCNA antibody(clone PC10; Sigma; diluted 1:4,000), overnight, or twopolyclonal antibodies against visual pigment opsins(CERN-922, an anti-bovine rod opsin, and CERN-874, ananti-chicken cone opsin; diluted 1:1,000) and incubated for3 days. After rinsing in phosphate-buffered saline (PBS),sections were incubated with goat anti-rabbit (Sigma) oranti-mouse (Dako, Glostrup, Denmark) immunoglobulins,diluted 1:100 or 1:50, respectively, for 1 hour and devel-oped with DAB and H2O2 as described above.
The specificity of the antibodies against GABA, seroto-nin, and PCNA had previously been tested by the sup-pliers. Previous studies have shown that the antiopsinantibodies stain both pineal complex and retinal photore-ceptors in larval sea lampreys (Garcıa-Fernandez andFoster, 1994). As a further control, some sections wereprocessed without incubating with the primary antibody,or the primary antiserum was replaced by the correspond-ing preimmune serum. In these sections, no immunostain-ing was observed.
Photomicrographs were made with an Olympus DP-10color digital camera. The images were equalized, con-verted to gray scale, and adjusted for brightness and con-trast using Corel Photo-Paint 8. Some photographs were
slightly Nomarski-like enhanced with the relief 3-D effect.Photomontages were constructed and lettering was doneusing Corel Draw 8.
Hematoxylin-eosin staining andsemithin sections
Some prolarvae and small larvae were fixed in 1% para-formaldehyde and 1% glutaraldehyde in PB, rinsed in PB,postfixed in 1% OsO4 in 0.1 M cacodylate buffer, pH 7.4,dehydrated with acetone, and embedded in Spurr resin.Semithin transverse sections (1 �m thick) were cut on anultramicrotome and stained with 1% toluidine blue-1%borax solution in water. Series of hematoxylin-eosin-stained transverse sections of prolarvae and larvae fromour collection were also used for topographic purposes.
RESULTS
Late embryos (E10–13) showed highly elongated tubu-lar optic vesicles extending caudally from the terminalregion of the brain. In front of the posterior commissure,there was a small evagination of the roof of the forebrain,representing the pineal organ primordium. A parapinealorgan was not distinguished in embryos. No opsin-,serotonin-, or GABA-ir structures were noted in the pho-toreceptive organs, in accordance with their undifferenti-ated appearance. However, these embryos contained a fewsmall groups of GABA-ir neurons in the brain and spinalcord as well as fibers coursing longitudinally or crossing insome commissures.
Eye development and appearance of opsinand GABA immunoreactivities
In early prolarvae (hatchlings), the primordia of theeyes begin to thicken in a position caudal to the opticregion of the brain, just in front of the trigeminal ganglion,but their walls are still formed by a single epithelial layerof cells loaded with yolk platelets. The nuclei of the cells ofthe optic vesicle were PCNA-ir (Fig. 1A). A long optic stalkjoins the optic vesicle to the lateral optic recesses in thebrain (Fig. 1B). The epidermis close to the optic vesiclewas thickened, forming a prominent lens placode (Fig.1A). At the pigmentation stage, the terminal vesicularportion consisted of a flattened medial wall and a thick-ened lateral wall. All cells of the optic and lens vesicleshad PCNA-ir nuclei. At the gill cleft stage, several opsin-ircells were observed in the medial layer of the neural retina(Fig. 1C), indicating the start of differentiation of photo-receptors. Opsin immunoreactivity was observed in all cellregions, including the flattened apical region and a thinbasal process. The medial wall did not shown pigmenta-tion. A lens vesicle primordium could now be distin-guished. At the end of the gill cleft stage, some melaningranules appeared in the retinal epithelium, and the neu-ral retina consisted of two layers of nuclei, lateral (innerregion) and medial (outer region). No GABA-ir structureswere noted in the eyes at this stage or preceding stages. Inburrowing-stage prolarvae, light microscopy of the retinashowed a thin cleft between the pigmented epithelium andthe neural retina, and short but wide protrusions of pho-toreceptors were noted here (Fig. 1D). PCNA immunore-activity was very faint or absent in most cells of the opticvesicle, except in a few cells in the margin of the vesicleand occasional cells located among photoreceptors,whereas the cells of the optic stalk had PCNA-ir nuclei.
252 M. MELENDEZ-FERRO ET AL.
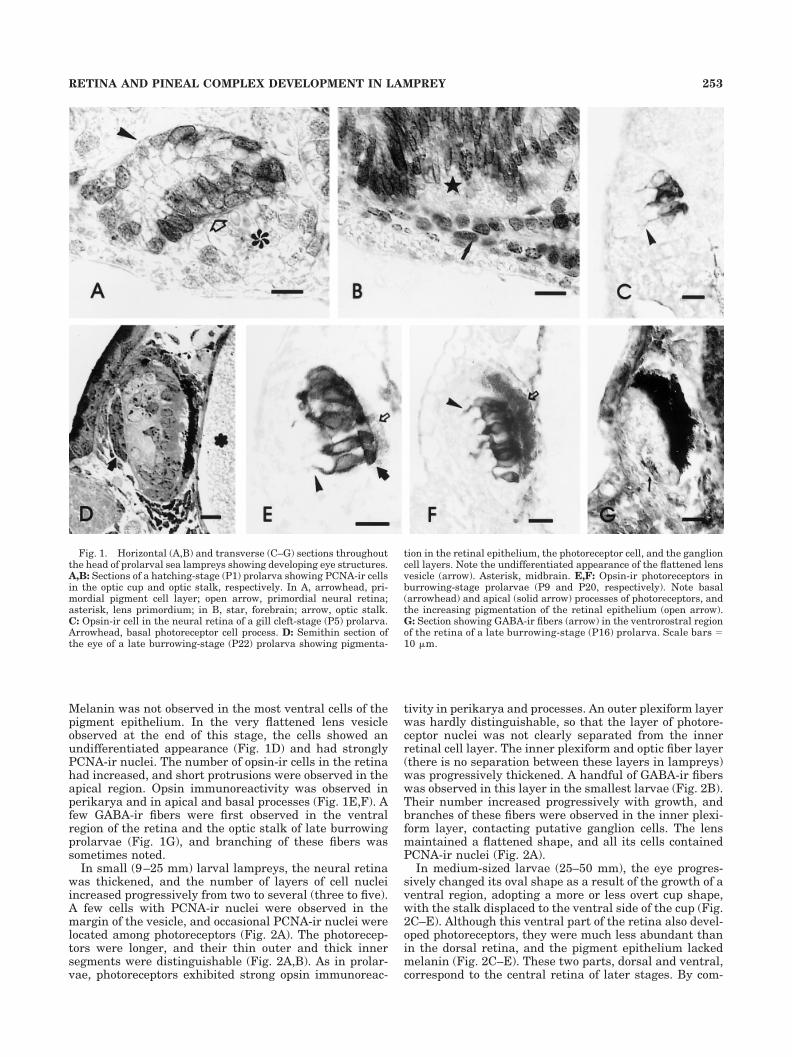
Melanin was not observed in the most ventral cells of thepigment epithelium. In the very flattened lens vesicleobserved at the end of this stage, the cells showed anundifferentiated appearance (Fig. 1D) and had stronglyPCNA-ir nuclei. The number of opsin-ir cells in the retinahad increased, and short protrusions were observed in theapical region. Opsin immunoreactivity was observed inperikarya and in apical and basal processes (Fig. 1E,F). Afew GABA-ir fibers were first observed in the ventralregion of the retina and the optic stalk of late burrowingprolarvae (Fig. 1G), and branching of these fibers wassometimes noted.
In small (9–25 mm) larval lampreys, the neural retinawas thickened, and the number of layers of cell nucleiincreased progressively from two to several (three to five).A few cells with PCNA-ir nuclei were observed in themargin of the vesicle, and occasional PCNA-ir nuclei werelocated among photoreceptors (Fig. 2A). The photorecep-tors were longer, and their thin outer and thick innersegments were distinguishable (Fig. 2A,B). As in prolar-vae, photoreceptors exhibited strong opsin immunoreac-
tivity in perikarya and processes. An outer plexiform layerwas hardly distinguishable, so that the layer of photore-ceptor nuclei was not clearly separated from the innerretinal cell layer. The inner plexiform and optic fiber layer(there is no separation between these layers in lampreys)was progressively thickened. A handful of GABA-ir fiberswas observed in this layer in the smallest larvae (Fig. 2B).Their number increased progressively with growth, andbranches of these fibers were observed in the inner plexi-form layer, contacting putative ganglion cells. The lensmaintained a flattened shape, and all its cells containedPCNA-ir nuclei (Fig. 2A).
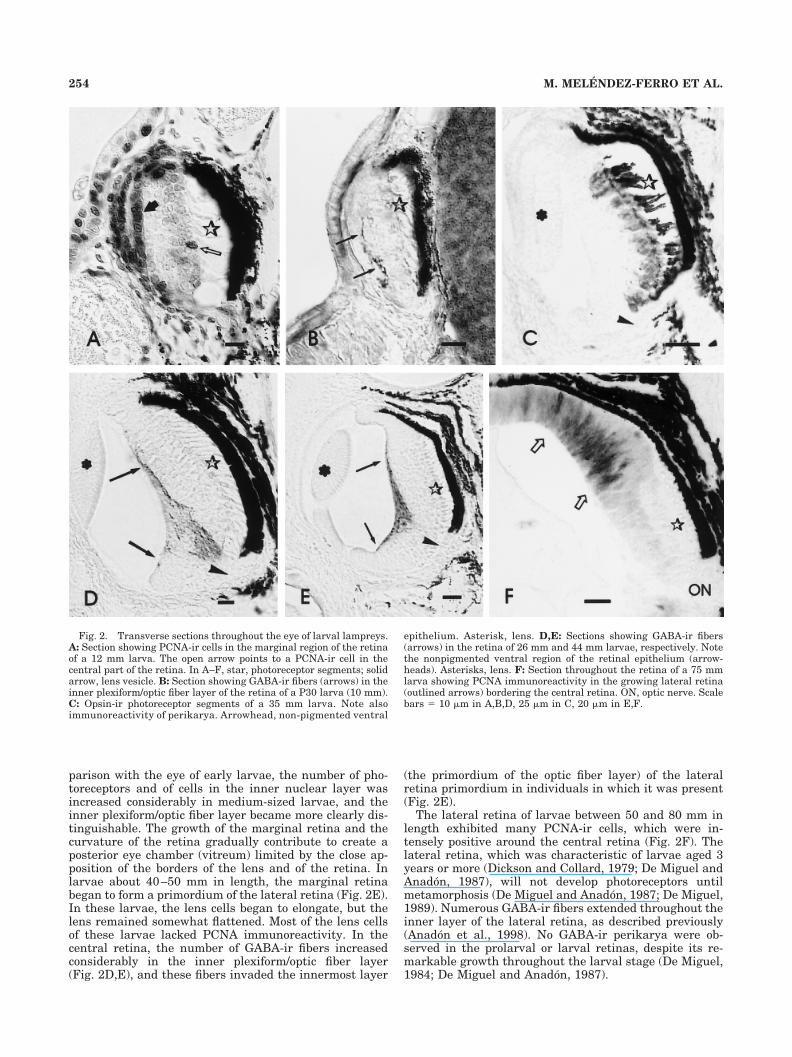
In medium-sized larvae (25–50 mm), the eye progres-sively changed its oval shape as a result of the growth of aventral region, adopting a more or less overt cup shape,with the stalk displaced to the ventral side of the cup (Fig.2C–E). Although this ventral part of the retina also devel-oped photoreceptors, they were much less abundant thanin the dorsal retina, and the pigment epithelium lackedmelanin (Fig. 2C–E). These two parts, dorsal and ventral,correspond to the central retina of later stages. By com-
Fig. 1. Horizontal (A,B) and transverse (C–G) sections throughoutthe head of prolarval sea lampreys showing developing eye structures.A,B: Sections of a hatching-stage (P1) prolarva showing PCNA-ir cellsin the optic cup and optic stalk, respectively. In A, arrowhead, pri-mordial pigment cell layer; open arrow, primordial neural retina;asterisk, lens primordium; in B, star, forebrain; arrow, optic stalk.C: Opsin-ir cell in the neural retina of a gill cleft-stage (P5) prolarva.Arrowhead, basal photoreceptor cell process. D: Semithin section ofthe eye of a late burrowing-stage (P22) prolarva showing pigmenta-
tion in the retinal epithelium, the photoreceptor cell, and the ganglioncell layers. Note the undifferentiated appearance of the flattened lensvesicle (arrow). Asterisk, midbrain. E,F: Opsin-ir photoreceptors inburrowing-stage prolarvae (P9 and P20, respectively). Note basal(arrowhead) and apical (solid arrow) processes of photoreceptors, andthe increasing pigmentation of the retinal epithelium (open arrow).G: Section showing GABA-ir fibers (arrow) in the ventrorostral regionof the retina of a late burrowing-stage (P16) prolarva. Scale bars �10 �m.
253RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY
parison with the eye of early larvae, the number of pho-toreceptors and of cells in the inner nuclear layer wasincreased considerably in medium-sized larvae, and theinner plexiform/optic fiber layer became more clearly dis-tinguishable. The growth of the marginal retina and thecurvature of the retina gradually contribute to create aposterior eye chamber (vitreum) limited by the close ap-position of the borders of the lens and of the retina. Inlarvae about 40–50 mm in length, the marginal retinabegan to form a primordium of the lateral retina (Fig. 2E).In these larvae, the lens cells began to elongate, but thelens remained somewhat flattened. Most of the lens cellsof these larvae lacked PCNA immunoreactivity. In thecentral retina, the number of GABA-ir fibers increasedconsiderably in the inner plexiform/optic fiber layer(Fig. 2D,E), and these fibers invaded the innermost layer
(the primordium of the optic fiber layer) of the lateralretina primordium in individuals in which it was present(Fig. 2E).
The lateral retina of larvae between 50 and 80 mm inlength exhibited many PCNA-ir cells, which were in-tensely positive around the central retina (Fig. 2F). Thelateral retina, which was characteristic of larvae aged 3years or more (Dickson and Collard, 1979; De Miguel andAnadon, 1987), will not develop photoreceptors untilmetamorphosis (De Miguel and Anadon, 1987; De Miguel,1989). Numerous GABA-ir fibers extended throughout theinner layer of the lateral retina, as described previously(Anadon et al., 1998). No GABA-ir perikarya were ob-served in the prolarval or larval retinas, despite its re-markable growth throughout the larval stage (De Miguel,1984; De Miguel and Anadon, 1987).
Fig. 2. Transverse sections throughout the eye of larval lampreys.A: Section showing PCNA-ir cells in the marginal region of the retinaof a 12 mm larva. The open arrow points to a PCNA-ir cell in thecentral part of the retina. In A–F, star, photoreceptor segments; solidarrow, lens vesicle. B: Section showing GABA-ir fibers (arrows) in theinner plexiform/optic fiber layer of the retina of a P30 larva (10 mm).C: Opsin-ir photoreceptor segments of a 35 mm larva. Note alsoimmunoreactivity of perikarya. Arrowhead, non-pigmented ventral
epithelium. Asterisk, lens. D,E: Sections showing GABA-ir fibers(arrows) in the retina of 26 mm and 44 mm larvae, respectively. Notethe nonpigmented ventral region of the retinal epithelium (arrow-heads). Asterisks, lens. F: Section throughout the retina of a 75 mmlarva showing PCNA immunoreactivity in the growing lateral retina(outlined arrows) bordering the central retina. ON, optic nerve. Scalebars � 10 �m in A,B,D, 25 �m in C, 20 �m in E,F.
254 M. MELENDEZ-FERRO ET AL.

PCNA and opsin immunoreactivities anddevelopment of the pineal complex
and habenula
Embryos and prolarvae. In late embryos and hatch-lings, the pineal organ primordium was a small, sphericaloutgrowth of the caudodorsal forebrain open to the ven-tricle just rostral to the posterior commissure (Fig. 3A).Most cells of this region showed PCNA-ir nuclei, and onlya few (one or two) cells showed PCNA-negative nuclei athatching. No opsin immunoreactivity could be distin-guished in the pineal vesicle at these stages. A parapinealprimordium could not be distinguished. A thickening ofthe dorsal wall of the forebrain marked the primordium ofthe habenulae, in which practically all cell nuclei werePCNA-ir.
In pigmentation- and gill cleft-stage prolarvae, the pi-neal organ progressively increased its size, and the cau-doventral wall became thickened (Fig. 3B–E). Some nucleiof the ventral wall were PCNA-negative, whereas those ofthe thin dorsal wall were PCNA-ir (Fig. 3C,D). In earlypigmentation-stage prolarvae (P2), a few cells in the cau-doventral region of the pineal vesicle exhibited moderateto intense opsin immunoreactivity (Fig. 3B). The numberof these cells increased by P3 (Fig. 3E), and, in gill cleft-stage prolarvae, many cell of the ventral wall wereopsin-ir cells. In pigmentation- and gill cleft-stage prolar-vae, a small cell mass with PCNA-ir nuclei was located inthe midline rostral to the pineal organ, representing theparapineal primordium (Fig. 3C). A cord of cells withelongated nuclei represents the communication with thethird ventricle (Fig. 3C). In the habenulae, in addition toproliferating cells (i.e., cells with PCNA-ir nuclei), someexternal cells were negative for PCNA.
In burrowing-stage prolarvae, the pineal organ was aclosed vesicle and had many photoreceptors, as indicatedby their PCNA-negative nuclei (Fig. 3F), their opsin im-munoreactivity (Fig. 3G), and the presence of recognizablephotoreceptor segments in the thin cleft between the outerand inner pineal walls (Fig. 3H). In semithin sections, theparapineal primordium appeared as a much flattened ves-icle (Fig. 3I). The cells of this vesicle were dense andshowed an undifferentiated appearance, as indicated bytheir PCNA immunoreactivity. The habenulae had in-creased in size since the hatchling stage, and PCNA-negative cells were rather numerous in the rostral regionof the right habenula, whereas the nuclei of most cells ofthe caudal region exhibited strong PCNA immunoreactiv-ity (Fig. 3F). A PCNA-negative layer of cells was alsoobserved in the left habenula. No opsin immunoreactivitywas observed in the parapineal organ or the habenula.
Larval pineal organ. In the pineal organ of earlylarvae (10–12 mm), the number of cells increased, and theorgan became clearly vesicular, with a thin dorsal walland a thickened ventral wall. Cells with PCNA-ir nucleiappeared more or less scattered in the periventricularregion of the vesicle, and PCNA-negative nuclei were nu-merous in basal regions of the ventral wall (Fig. 3H,I). Inthe pineal atrium (i.e., the ventrocaudal region of thevesicle joining the pineal stalk), most cells had PCNA-irnuclei. Distribution of opsin-ir cells was complementary tothe PCNA pattern: Most cells of the ventral wall exhibitedintense opsin immunoreactivity, whereas opsin-ir cellswere scarce in the atrium. Photoreceptors segments werenumerous in the slit-shaped ventricle. The pineal organ of20–30 mm larvae contained scattered cells with PCNA-ir
nuclei associated with the ventricular surface. In 35–38mm larvae, a large proportion (about half) of the cells ofthe ventral wall of the pineal vesicle exhibited opsin im-munoreactivity. In the atrium, however, most of the cellswere opsin negative. The thin dorsal wall of the vesiclelacked opsin-ir cells. In larger larvae, most pineal cellnuclei are PCNA negative (Fig. 4C).
Larval parapineal organ. In 12 mm larvae, mostcells of the parapineal vesicle contained PCNA-ir nuclei(Fig. 4A). The walls of this vesicle (mainly the ventralwall) were clearly thickened, and only a few PCNA-negative nuclei were observed in the dorsal wall (Fig. 4A).The caudal region of the vesicle was close to the small lefthabenula, which appeared as a poorly thickened regionwith some cells exhibiting PCNA-negative nuclei (Fig.4D). The ventral wall of the parapineal vesicle becameprogressively thickened in larger larvae, and more PCNA-negative nuclei could be observed. Fusion (i.e., disappear-ance of the intermediate basal laminae and connectivelayers) of the parapineal ganglion primordium (the rostralpart of the left habenula) and the parapineal vesicle wasfound in 23 mm larvae. In 33–35 mm larvae, the parapi-neal organ showed the organization found in large larvae;i.e., it consisted of a vesicle exhibiting a thickened innerwall that was joined to the external neuropil of the parapi-neal ganglion. Unlike the pineal organ and parapinealganglion, the parapineal vesicle of large larvae (50–75mm) contained numerous cells with PCNA-ir nuclei scat-tered among PCNA-negative cells (Fig. 4C). No opsin-ircells were observed in the parapineal organ of the larvalstages studied.
Larval habenula. In the habenula of 9–12 mm larvallampreys, a clear rostrocaudal gradient of proliferationwas observed with PCNA immunocytochemistry (Fig.4A,D,E). Most nuclei of the rostral regions of right and lefthabenulae were PCNA negative, PCNA-ir nuclei beinglimited to a few cells in ventral ventricular areas, whereasin the caudal region most cells exhibited strong PCNA-irnuclei (Fig. 4E). In 23 mm larvae, numerous cells withPCNA-ir nuclei were observed in a proliferative ventro-caudal region of the right habenula, whereas the left ha-benula showed only scattered PCNA-ir cell nuclei. Al-though most parts of the habenulae of larger larvaecontained only PCNA-negative cells, the ventrocaudal re-gion of the right habenula, close to the subcommissuralorgan, exhibited numerous PCNA-positive cells through-out the larval period studied (9–75 mm larvae; Fig. 4F). Inthe left habenula of the same larvae. some cells withPCNA-ir nuclei were scattered close to the ventricle incaudal regions. In larvae larger than 70 mm, the lefthabenula–parapineal ganglion became elongated, but aneat separation between these two regions was observedonly in adults. No opsin-ir cells were observed in thehabenula of the larval stages studied.
Serotonin immunoreactivity in thedeveloping pineal complex and habenula
A few serotonin-ir cells were observed in the pinealorgan of late burrowing-stage prolarvae (Fig. 5A). In lar-vae, the serotonin-ir cells were often bipolar, but they didnot exhibit well-developed photoreceptor segments pro-truding into the pineal lumen (Fig. 5B). The number ofserotonin-positive bipolar cells increased considerablyduring development. In larvae more than 35 mm inlength, the inner wall of the pineal vesicle becomes pro-gressively loaded with reflecting crystals, which makes
255RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY
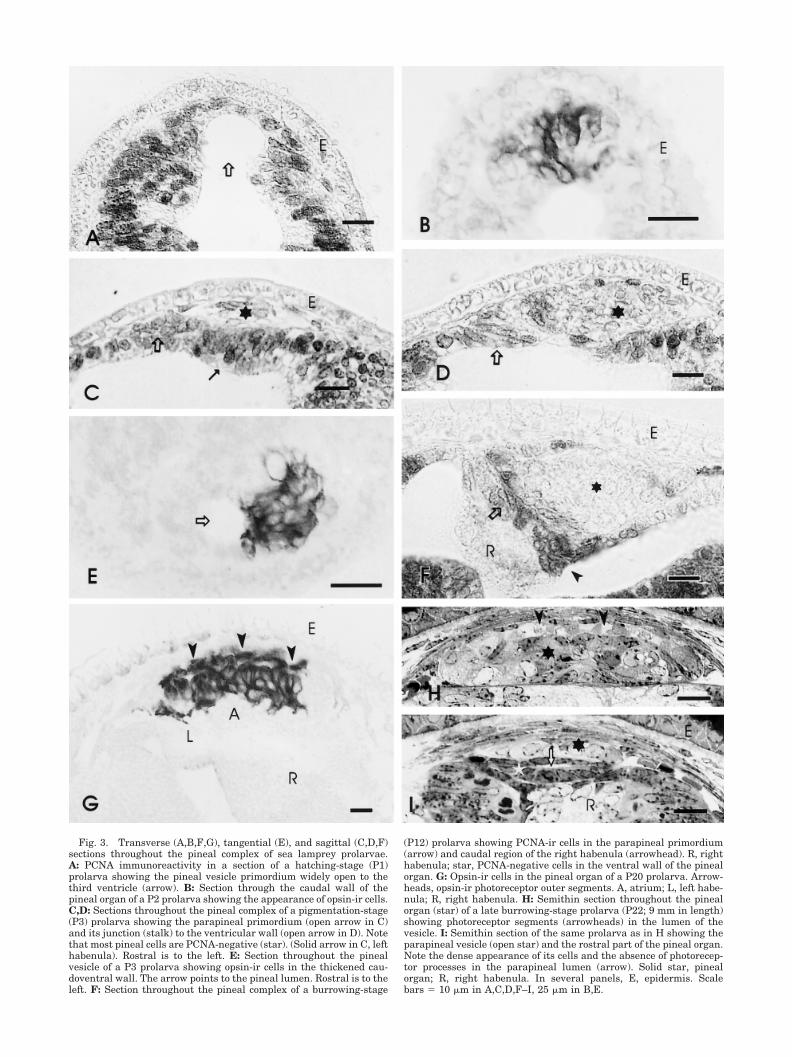
Fig. 3. Transverse (A,B,F,G), tangential (E), and sagittal (C,D,F)sections throughout the pineal complex of sea lamprey prolarvae.A: PCNA immunoreactivity in a section of a hatching-stage (P1)prolarva showing the pineal vesicle primordium widely open to thethird ventricle (arrow). B: Section through the caudal wall of thepineal organ of a P2 prolarva showing the appearance of opsin-ir cells.C,D: Sections throughout the pineal complex of a pigmentation-stage(P3) prolarva showing the parapineal primordium (open arrow in C)and its junction (stalk) to the ventricular wall (open arrow in D). Notethat most pineal cells are PCNA-negative (star). (Solid arrow in C, lefthabenula). Rostral is to the left. E: Section throughout the pinealvesicle of a P3 prolarva showing opsin-ir cells in the thickened cau-doventral wall. The arrow points to the pineal lumen. Rostral is to theleft. F: Section throughout the pineal complex of a burrowing-stage
(P12) prolarva showing PCNA-ir cells in the parapineal primordium(arrow) and caudal region of the right habenula (arrowhead). R, righthabenula; star, PCNA-negative cells in the ventral wall of the pinealorgan. G: Opsin-ir cells in the pineal organ of a P20 prolarva. Arrow-heads, opsin-ir photoreceptor outer segments. A, atrium; L, left habe-nula; R, right habenula. H: Semithin section throughout the pinealorgan (star) of a late burrowing-stage prolarva (P22; 9 mm in length)showing photoreceptor segments (arrowheads) in the lumen of thevesicle. I: Semithin section of the same prolarva as in H showing theparapineal vesicle (open star) and the rostral part of the pineal organ.Note the dense appearance of its cells and the absence of photorecep-tor processes in the parapineal lumen (arrow). Solid star, pinealorgan; R, right habenula. In several panels, E, epidermis. Scalebars � 10 �m in A,C,D,F–I, 25 �m in B,E.

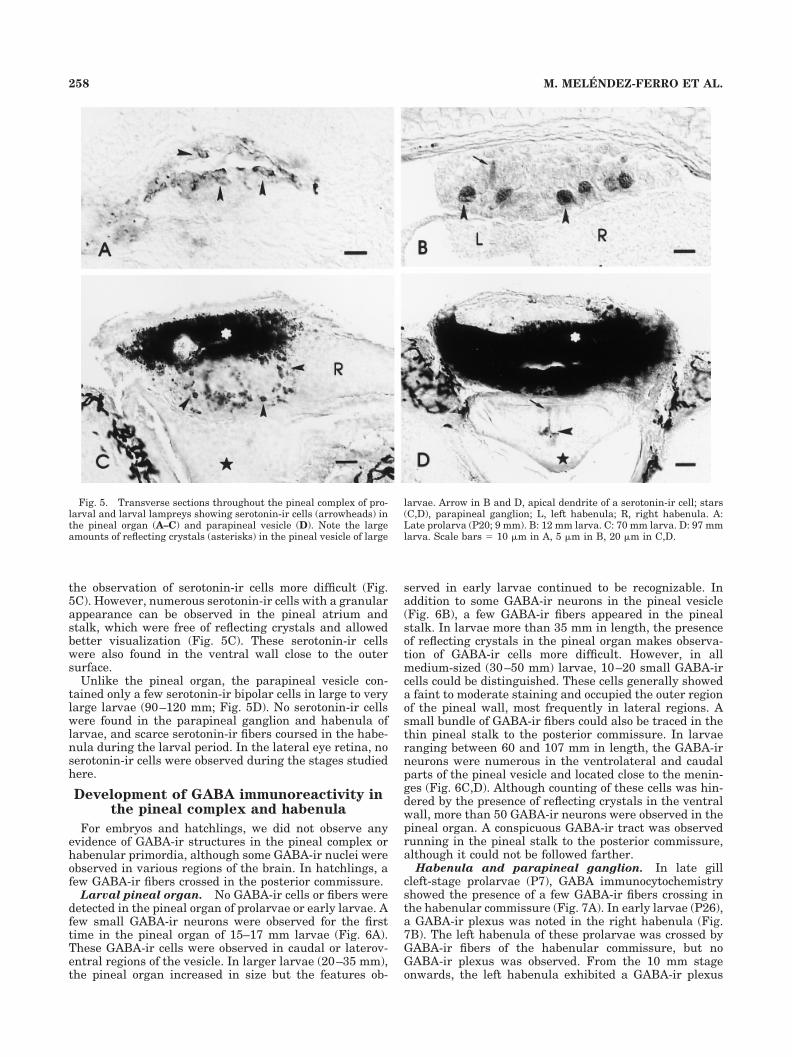
Fig. 4. Transverse sections throughout the pineal complex andhabenula of lamprey larvae showing the distribution of PCNA(A,C–F) and opsin (B) immunoreactivities. A: Section throughout thepineal complex of a 12 mm larva showing that most cell nuclei of theparapineal vesicle (open arrow) are PCNA-ir, with the exception of afew nuclei in the dorsal wall of the vesicle (solid arrow). Outlined star,pineal organ; arrowheads, PCNA-ir pineal cells; R, right habenula;white star, medial pallium (hippocampal) primordium. B: Sectionthrough the pineal complex of a 35 mm larva showing numerousopsin-ir cells in the pineal organ and the lack of immunoreactivity inthe parapineal organ (star). Arrowheads point to the photoreceptorsegments in the pineal lumen. C: Section showing numerous PCNA-ir
cells (arrows) in the parapineal vesicle of a 75 mm larva. Note that theparapineal ganglion (star) and pineal organ (asterisk) are almostdevoid of PCNA-ir cells. D,E: Sections throughout the rostral (D) andcaudal (E) regions of the right habenula (R) of a 12 mm larva. Note thePCNA-ir caudal region of the right habenula (arrow in E) and theoutstanding size difference between the two habenulae. PCNA-ir cellsare also found in the left habenula (L). F: Section throughout thecaudal region of the right habenula (R) showing the persistence of theventrocaudal proliferative region (arrow). The arrowhead points tothe corresponding region of the left habenula (L). HC, habenularcommissure. Scale bars � 10 �m in A, 25 �m in B, 20 �m in C–F.
257RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY
the observation of serotonin-ir cells more difficult (Fig.5C). However, numerous serotonin-ir cells with a granularappearance can be observed in the pineal atrium andstalk, which were free of reflecting crystals and allowedbetter visualization (Fig. 5C). These serotonin-ir cellswere also found in the ventral wall close to the outersurface.
Unlike the pineal organ, the parapineal vesicle con-tained only a few serotonin-ir bipolar cells in large to verylarge larvae (90–120 mm; Fig. 5D). No serotonin-ir cellswere found in the parapineal ganglion and habenula oflarvae, and scarce serotonin-ir fibers coursed in the habe-nula during the larval period. In the lateral eye retina, noserotonin-ir cells were observed during the stages studiedhere.
Development of GABA immunoreactivity inthe pineal complex and habenula
For embryos and hatchlings, we did not observe anyevidence of GABA-ir structures in the pineal complex orhabenular primordia, although some GABA-ir nuclei wereobserved in various regions of the brain. In hatchlings, afew GABA-ir fibers crossed in the posterior commissure.
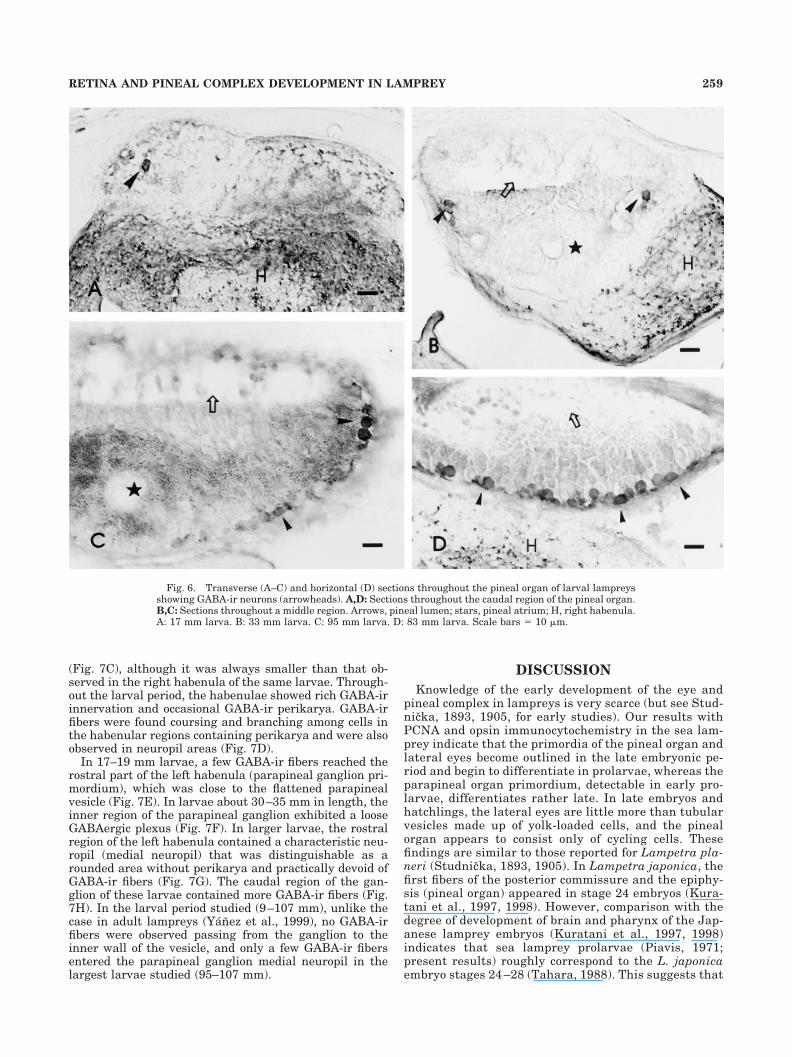
Larval pineal organ. No GABA-ir cells or fibers weredetected in the pineal organ of prolarvae or early larvae. Afew small GABA-ir neurons were observed for the firsttime in the pineal organ of 15–17 mm larvae (Fig. 6A).These GABA-ir cells were observed in caudal or laterov-entral regions of the vesicle. In larger larvae (20–35 mm),the pineal organ increased in size but the features ob-
served in early larvae continued to be recognizable. Inaddition to some GABA-ir neurons in the pineal vesicle(Fig. 6B), a few GABA-ir fibers appeared in the pinealstalk. In larvae more than 35 mm in length, the presenceof reflecting crystals in the pineal organ makes observa-tion of GABA-ir cells more difficult. However, in allmedium-sized (30–50 mm) larvae, 10–20 small GABA-ircells could be distinguished. These cells generally showeda faint to moderate staining and occupied the outer regionof the pineal wall, most frequently in lateral regions. Asmall bundle of GABA-ir fibers could also be traced in thethin pineal stalk to the posterior commissure. In larvaeranging between 60 and 107 mm in length, the GABA-irneurons were numerous in the ventrolateral and caudalparts of the pineal vesicle and located close to the menin-ges (Fig. 6C,D). Although counting of these cells was hin-dered by the presence of reflecting crystals in the ventralwall, more than 50 GABA-ir neurons were observed in thepineal organ. A conspicuous GABA-ir tract was observedrunning in the pineal stalk to the posterior commissure,although it could not be followed farther.
Habenula and parapineal ganglion. In late gillcleft-stage prolarvae (P7), GABA immunocytochemistryshowed the presence of a few GABA-ir fibers crossing inthe habenular commissure (Fig. 7A). In early larvae (P26),a GABA-ir plexus was noted in the right habenula (Fig.7B). The left habenula of these prolarvae was crossed byGABA-ir fibers of the habenular commissure, but noGABA-ir plexus was observed. From the 10 mm stageonwards, the left habenula exhibited a GABA-ir plexus
Fig. 5. Transverse sections throughout the pineal complex of pro-larval and larval lampreys showing serotonin-ir cells (arrowheads) inthe pineal organ (A–C) and parapineal vesicle (D). Note the largeamounts of reflecting crystals (asterisks) in the pineal vesicle of large
larvae. Arrow in B and D, apical dendrite of a serotonin-ir cell; stars(C,D), parapineal ganglion; L, left habenula; R, right habenula. A:Late prolarva (P20; 9 mm). B: 12 mm larva. C: 70 mm larva. D: 97 mmlarva. Scale bars � 10 �m in A, 5 �m in B, 20 �m in C,D.
258 M. MELENDEZ-FERRO ET AL.
(Fig. 7C), although it was always smaller than that ob-served in the right habenula of the same larvae. Through-out the larval period, the habenulae showed rich GABA-irinnervation and occasional GABA-ir perikarya. GABA-irfibers were found coursing and branching among cells inthe habenular regions containing perikarya and were alsoobserved in neuropil areas (Fig. 7D).
In 17–19 mm larvae, a few GABA-ir fibers reached therostral part of the left habenula (parapineal ganglion pri-mordium), which was close to the flattened parapinealvesicle (Fig. 7E). In larvae about 30–35 mm in length, theinner region of the parapineal ganglion exhibited a looseGABAergic plexus (Fig. 7F). In larger larvae, the rostralregion of the left habenula contained a characteristic neu-ropil (medial neuropil) that was distinguishable as arounded area without perikarya and practically devoid ofGABA-ir fibers (Fig. 7G). The caudal region of the gan-glion of these larvae contained more GABA-ir fibers (Fig.7H). In the larval period studied (9–107 mm), unlike thecase in adult lampreys (Yanez et al., 1999), no GABA-irfibers were observed passing from the ganglion to theinner wall of the vesicle, and only a few GABA-ir fibersentered the parapineal ganglion medial neuropil in thelargest larvae studied (95–107 mm).
DISCUSSION
Knowledge of the early development of the eye andpineal complex in lampreys is very scarce (but see Stud-nicka, 1893, 1905, for early studies). Our results withPCNA and opsin immunocytochemistry in the sea lam-prey indicate that the primordia of the pineal organ andlateral eyes become outlined in the late embryonic pe-riod and begin to differentiate in prolarvae, whereas theparapineal organ primordium, detectable in early pro-larvae, differentiates rather late. In late embryos andhatchlings, the lateral eyes are little more than tubularvesicles made up of yolk-loaded cells, and the pinealorgan appears to consist only of cycling cells. Thesefindings are similar to those reported for Lampetra pla-neri (Studnicka, 1893, 1905). In Lampetra japonica, thefirst fibers of the posterior commissure and the epiphy-sis (pineal organ) appeared in stage 24 embryos (Kura-tani et al., 1997, 1998). However, comparison with thedegree of development of brain and pharynx of the Jap-anese lamprey embryos (Kuratani et al., 1997, 1998)indicates that sea lamprey prolarvae (Piavis, 1971;present results) roughly correspond to the L. japonicaembryo stages 24 –28 (Tahara, 1988). This suggests that
Fig. 6. Transverse (A–C) and horizontal (D) sections throughout the pineal organ of larval lampreysshowing GABA-ir neurons (arrowheads). A,D: Sections throughout the caudal region of the pineal organ.B,C: Sections throughout a middle region. Arrows, pineal lumen; stars, pineal atrium; H, right habenula.A: 17 mm larva. B: 33 mm larva. C: 95 mm larva. D: 83 mm larva. Scale bars � 10 �m.
259RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY
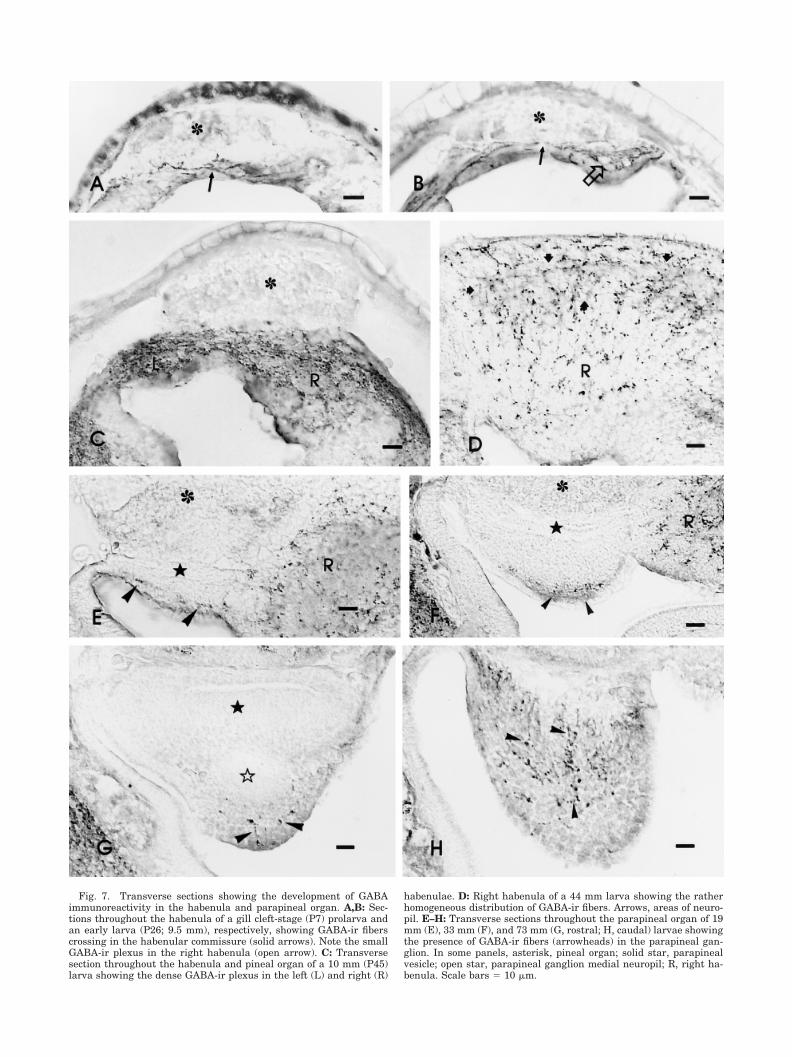
Fig. 7. Transverse sections showing the development of GABAimmunoreactivity in the habenula and parapineal organ. A,B: Sec-tions throughout the habenula of a gill cleft-stage (P7) prolarva andan early larva (P26; 9.5 mm), respectively, showing GABA-ir fiberscrossing in the habenular commissure (solid arrows). Note the smallGABA-ir plexus in the right habenula (open arrow). C: Transversesection throughout the habenula and pineal organ of a 10 mm (P45)larva showing the dense GABA-ir plexus in the left (L) and right (R)
habenulae. D: Right habenula of a 44 mm larva showing the ratherhomogeneous distribution of GABA-ir fibers. Arrows, areas of neuro-pil. E–H: Transverse sections throughout the parapineal organ of 19mm (E), 33 mm (F), and 73 mm (G, rostral; H, caudal) larvae showingthe presence of GABA-ir fibers (arrowheads) in the parapineal gan-glion. In some panels, asterisk, pineal organ; solid star, parapinealvesicle; open star, parapineal ganglion medial neuropil; R, right ha-benula. Scale bars � 10 �m.

relative time of appearance of pineal structures is sim-ilar in both lamprey species.
Cell proliferation in the retina andpineal complex
The present results in lampreys indicate that PCNAceases to be expressed in cell populations showing the firstsigns of differentiation, both in the eye and in the pinealcomplex. Our results with PCNA immunocytochemistryare comparable with those obtained in the developingretina of teleosts (Negishi et al., 1990a,b; Negishi andWagner, 1995). The parallelism between the expression ofPCNA in the retinas of lampreys and of teleosts is alsostrengthened by the marginal distribution of mostPCNA-ir cells in the retinas of late lamprey prolarvae andearly larvae and by the presence of occasional PCNA-ircells in the outer nuclear layer of the region with differ-entiated photoreceptors. A similar marginal zone of cellproliferation has also been reported for the chicken retina(Fisher and Reh, 2000). With the abundance of PCNA-ircells, proliferation is probably more active in the pinealorgan of late lamprey prolarvae and early larvae than inthe retina. In the parapineal organ, growth of the vesicleand cell differentiation appear to be much delayed. PCNAstudies have not been performed in the pineal complex ofteleosts, which precludes comparison with lampreys.
The late differentiation of the parapineal organ in lam-preys and the common origin of the parapineal ganglionand the left habenula have already been noted in riverlampreys ( Studnicka, 1893, 1905; Meiniel and Collin,1971). Our observations with PCNA immunocytochemis-try confirm that the beginning of cell differentiation in theparapineal organ is much delayed with respect to that inthe retina or the pineal organ. The projections of theparapineal vesicle to the medial neuropil of the ganglionrepresent the main parapineal afferent connection ob-served in adult lampreys (Yanez et al., 1999). The latefusion between the parapineal vesicle and the left-habenula-derived parapineal ganglion (in larvae about 23mm in length) indicates that the projections of the parapi-neal vesicle to the ganglion develop much later than theretinofugal and pinealofugal projections. In teleosts, theparapineal organ projects to a rostral portion of the lefthabenula (Yanez et al., 1996), which is similar to thatobserved in lampreys. However, the development of theprimordium of this organ and the parapineal tract inteleosts appears to occur almost as early as that of thepineal organ (Ekstrom et al., 1983; Corujo and Anadon,1986), in complete contrast to the pattern observed inlampreys. Moreover, the teleost parapineal organ growsvery little after the embryonic period (Holmgren, 1965;Corujo and Anadon, 1986), unlike the case in lampreys.
The habenulae and pineal complex of lampreys are no-table for their marked asymmetry (see Yanez and Anadon,1994b; Yanez et al., 1999). This asymmetry is evident assoon as the habenular primordia become recognizable. Onthe other hand, in the right habenula of lamprey larvaelarger than 20 mm, the PCNA-positive region is practi-cally limited to a caudoventral region close to the subcom-missural organ. The left habenula and the left-habenula-derived parapineal ganglion develop later and showconsiderably less cycling cell than the right habenula, asindicated by the scattered distribution and scarcity ofPCNA-ir cells. The marked difference in number of cyclingcells between the right and the left habenula, throughoutthe larval period, may explain the observed habenular
asymmetry. Asymmetrical cell proliferation in the habe-nulae has not been noted with PCNA immunocytochem-istry in zebrafish (Wullimann and Puelles, 1999), al-though recent studies have found that nodal-related geneexpression pathways underlie the anatomical asymmetryof the habenula and pineal complex in this species (Lianget al., 2000; Concha et al., 2000).
Development of opsin-ir cells in the pinealcomplex and retina
CERN-922 (antirod opsin) and CERN-874 (anticone op-sin) antisera give similar staining patterns in the pinealorgan and retina, as had been reported for adults of threespecies of lamprey (Garcıa-Fernandez et al., 1997). Simi-lar results with different antirod opsin antisera have beenobtained in river lamprey and some other vertebrates (vanVeen et al., 1984; Vigh-Teichmann et al., 1989, 1991). Thisindicates that these antisera cross-react with opsins ofsome pineal photoreceptors, suggesting close immunolog-ical properties between pineal and retinal opsins in lar-vae. However, Vigh-Teichmann et al. (1989) indicated thattheir antiopsin antiserum does not react with photorecep-tors of the pineal atrium, which also appears to be the casewith the two antisera used here.
Our results in sea lamprey reveal that opsin-ir photo-receptors first appear in the pineal organ of early prolar-vae and a few days later in the retina. The appearance ofthe pineal vesicle at that stage is of a single epithelium.An earlier expression of opsins in the pineal organ than inthe retina has been reported for some fishes (van Veen etal., 1984; Negishi and Wagner, 1995; Forsell et al., 1997),although not for zebrafish (Robinson et al., 1995) or forXenopus (Korf et al., 1989). Although it has been sug-gested that the appearance of photoreceptive molecules inthe pineal is related to hatching regulation in some fishes(Forsell et al., 1997), our results suggest that this is notthe case for lampreys. In lamprey, opsin antisera appearto label pineal and retinal photoreceptors in very earlystages of differentiation, as judged by the apparent lack ofphotoreceptor segments in the earliest stages at whichopsins were demonstrated. Similar early appearance ofopsins has been found in zebrafish and Xenopus (Saha andGrainger, 1993; Robinson et al., 1995).
Development of serotonin-ir cells in thepineal complex
In the pineal organ, serotonin-ir cells were first ob-served in late prolarvae, i.e., several days after the ap-pearance of opsin-ir photoreceptor. The parapineal organ,however, showed only a few serotonin-ir cells in largelarvae. These results indicate that the two organs alsofollow very different developmental patterns with respectto the appearance of serotonin immunoreactivity, which isin agreement with the notable difference in number ofserotonin-ir cells between these organs observed in adultlampreys (Tamotsu et al., 1990; Pombal et al., 1999; Yanezet al., 1999). Similar differences between the pineal andthe parapineal organs in the appearance of serotonin-ircells have been found in teleosts (van Veen et al., 1984). Indeveloping teleosts, the presence of serotonin-ir cells inthe parapineal organ has been observed only in sockeyesalmon juveniles (Ekstrom and Ebbesson, 1989).
Serotonin-containing cells of the lamprey pineal organare regarded as photoneuroendocrine cells synthesizingmelatonin (Meiniel and Hartwig, 1980; Guerlotte et al.,1986; Tamotsu et al., 1990, 1997). The first serotonin-ir
261RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY

cells observed in the developing lamprey pineal organhave the appearance of photoreceptors, as described foradults (Tamotsu et al., 1990, 1994). Their small photore-ceptor segments suggest that they correspond to type IIpineal photoreceptors (Meiniel, 1980), most of which bear-ing small rod opsin-ir outer segments (Tamotsu et al.,1997). The early appearance of serotonin in the pinealgland, though later than that of opsins, suggests thatphotoneuroendocrine action appears well before develop-ment of pinealofugal projections, as indicated by the lateappearance of GABA-ir fibers in the pineal stalk. In thepineal organ of teleosts, serotonin immunoreactivity alsoappears in the photoreceptor cell line early in develop-ment (van Veen et al., 1984). Serotonin and melatonin-synthesizing enzymes continue to be expressed in adultpineal organs (van Veen et al., 1980; Falcon et al., 1984;Ekstrom and Meissl, 1990; Begay et al., 1998). Unlike thecase in sockeye salmon fry (Ekstrom and Ebbesson, 1989),no serotonin-ir cells were observed in the habenulae of thelarval sea lamprey.
Development of GABAergic structures inthe retina and pineal complex
The present work is the first developmental study ofGABAergic structures in the photoreceptive epithalamusand the eyes of lampreys. Our findings indicate that thedevelopment of GABAergic structures in these organs ismuch delayed with respect to that in the brain, whichshows the first groups of GABAergic neurons in embryos(unpublished observations).
The first photoreceptors begin to differentiate in prolar-vae only a few days earlier in the pineal organ than in theretina (as indicated by the time of appearance of opsin-ircells). However, the development of GABA-ir structures inthese organs follows very different patterns. Thus, thefirst GABA-ir perikarya of the pineal organ appear infirst-year larvae after differentiation of photoreceptors,whereas GABA-ir cells are not observed in the retina oflarvae (Anadon et al., 1998; present results).
GABA immunoreactivity in the developing retina.
In late prolarvae and during the larval stage (a periodlasting for several years), the central retina containsstructurally and immunocytochemically identifiable pho-toreceptors (Dickson and Collard, 1979; De Miguel andAnadon, 1987; Rubinson and Cain, 1989; Garcıa-Fernandez and Foster, 1994; present results). In adultlampreys, some amacrine and horizontal cells areGABA-ir (Rio et al., 1993), as in most vertebrates (Agardhet al., 1987). Our results suggest that differentiation ofthese GABA-ir perikarya of the retina begins duringtransformation into young lampreys. A similar time se-quence of differentiation was observed for dopaminergiccells of the lamprey retina: They were absent throughoutthe larval period, even in the central retina, and were firstobserved in young postmetamorphic lampreys (De Migueland Wagner, 1990; Yanez and Anadon, 1994a). Our re-sults also indicate that no serotoninergic cells werepresent in the retinas of the prolarval and larval stagesstudied, despite the fact that adult lamprey retinas pos-sess serotonin-ir neurons, most of them amacrine cells(Negishi et al., 1986; Tamotsu et al., 1990; Versaux-Botteri et al., 1991).
In teleost retinas, as in those of amphibians and mam-mals, GABAergic, dopaminergic, and serotoninergic cellsappear around the time at which the first photoreceptorswith signs of morphological differentiation appear, in
early stages of retinogenesis (Ostholm et al., 1988; Negishiet al., 1990b; Becerra et al., 1994; Sandell et al., 1994;Negishi and Wagner, 1995; Hagedorn et al., 1998). Thisdoes not occur in sea lampreys, despite the fact that, in theeyes of late prolarvae and larvae, the retina (the “central”retina) possesses well-developed photoreceptors (Stud-nicka, 1912; De Miguel and Anadon, 1987; present re-sults). It was suggested that early GABA expression inhorizontal cells of the retina is related to rod (teleosts:Hagedorn et al., 1998) or cone (rabbit: Messersmith andRedburn, 1993; Mitchell and Redburn, 1996; Mitchell etal., 1999) differentiation, but this does not appear to be thecase in larval lampreys. One possible explanation forthese dramatic differences in retinal development be-tween lampreys and other vertebrates is that GABAergicand monoaminergic cells of the retina are involved in thecircuits for analysis of images formed on the retina. Theabsence of GABAergic and monoaminergic cells in thelarval lamprey eyes thus would suggest that they have notformed the image-analysis circuits yet. Unlike the eyes ofother developing vertebrates, the eyes of larval lampreyslie below a thickened skin and lack a transparent cornea(see Dickson and Graves, 1981; De Miguel and Anadon,1987). In larvae less than 50 mm in length, they also lacka lens with refractive properties, as judged by its flattenedshape and the PCNA positivity and undifferentiatedstructure of all cells of the lens primordium (present re-sults). Together, these results indicate that the larval eyesare only light detectors (ocellus).
The present results extend those of previous studies oflampreys showing the presence of GABA-ir centrifugalfibers in the retina of adults (Rio et al., 1993) and largelarvae (Anadon et al., 1998) to late (burrowing) prolarvaeand early larvae. Previous studies with tract-tracingmethods for the sea lamprey have suggested that thissystem appears later than was in fact observed here, giventhe absence or rarity of labeled M5 neurons after horse-radish peroxidase application to the eye in larvae between11 and 20 mm in length (Rodicio et al., 1995). The presentimmunocytochemical results revealed the presence ofGABAergic centrifugal fibers in prolarval eyes, which isearlier than previously thought and only a few days latterthan the start of differentiation of the first photoreceptors.The GABAergic fibers present in late prolarval and earlylarval eyes may in fact play developmental roles, as sug-gested for GABAergic fibers in various systems of othervertebrates. In all the GABAergic neurons of the develop-ing rat cerebellum, GABA-like immunoreactivity can beobserved long before synapse formation and the emer-gence of their electrophysiological activity (Aoki et al.,1989), and Aoki et al. suggest that GABA expressed indeveloping neurons may have not only neurotransmitterbut also developmental functions. In newborn mice andrats, GABA may similarly have both excitatory and tro-phic actions (Cherubini et al., 1991; Obata, 1997). In de-veloping rabbit retina, GABA released from horizontalcells is thought to intervene in the differentiation of pho-toreceptors (Messersmith and Redburn, 1993; Mitchelland Redburn, 1996; Hu et al., 1998; Sandell, 1998;Redburn-Johnson, 1998). Intracellular recordings of reti-nal cells responding to optic nerve stimulation in adultlamprey retinas have shown that, in most cells, nervestimulation produces excitatory postsynaptic potentials(Vesselkin et al., 1989); this raises the possibility thatcentrifugal fibers, and hence GABA, are excitatory even inthe adult lamprey retina.
262 M. MELENDEZ-FERRO ET AL.

The origin of the GABAergic retinopetal innervation ofthe larval lamprey retina can be traced to the M5 tegmen-tal mesencephalic nucleus (Rio et al., 1993; Anadon et al.,1998), the only retinopetal nucleus demonstrated withtract tracing methods in small larvae (Rodicio et al., 1995).In prolarval and early larval stages, the mesencephalictegmentum contains a rich GABAergic population in theregion of the M5 nucleus (unpublished observations), in-dicating that these GABAergic cells begin the differentia-tion early.
Development of GABAergic structures in the pineal
complex. Our results with the pineal organ of larvaeconfirm previous results in adult lampreys indicating thepresence of a population of GABA-ir cells with character-istics of ganglion cells (Pombal et al., 1999) and projectingthrough the pineal stalk and posterior commissure to thebrain. In the present study, no evidence of GABA immu-noreactivity in glial cells or pineal interneurons was ob-tained for the larval lamprey pineal gland, in agreementwith results in adults (Pombal et al., 1999). Our GABAresults confirm the close relationship between the pinealorgan and the posterior commissure observed by earlyauthors (Studnicka, 1893, 1905). The presence of GABAer-gic ganglion cells in the lamprey pineal organ (Pombal etal., 1999; present results) suggests that some of the pinealprojections are inhibitory. Note that GABA may have anexcitatory role in early stages of development (see above).The presence of a GABAergic system in the pineal organhas been also reported for other vertebrates. In the pinealorgan of a teleost, GABA immunoreactivity was reportedfor ganglion cells and glial cells and some interneuronsthat do not project to the brain (Ekstrom et al., 1987). Inthe pineal of teleosts, GABA participates in the modula-tion of light sensitivity (Meissl and Ekstrom, 1991). Un-like the case in lamprey and teleosts, interneurons werethe only GABA-ir cells observed in the pineal organ of frog(Ekstrom et al., 1990).
The pineal ganglion cells and some photoreceptor-likebipolar cells project to the brain in both lampreys (Same-jima et al., 1989; Pombal et al., 1999) and teleosts (Ek-strom, 1987). Although the two afferent systems (ganglioncells and photoreceptors) appear in early larval lampreys,differentiation of photoreceptors occurs sooner than thatof GABA-ir ganglion cells. However, whether brain-projecting photoreceptors develop before GABAergic pro-jections cannot be assessed. The appearance of GABAergicneurons in the pineal organ after the differentiation of thefirst pineal photoreceptors has likewise been found inteleosts (Ostholm et al., 1988; Ekstrom and Ohlin, 1995).
Finally, our results for the habenula of the sea lampreyindicate that GABAergic innervation appears shortly af-ter the first habenular neurons become recognizable bythe loss of PCNA positivity in their nuclei. This also ap-plies to the development of GABAergic innervation in theparapineal ganglion. Although in adult lampreys a fewGABA-ir fibers also extend from the parapineal ganglioninto the ventral wall of the parapineal vesicle (Yanez etal., 1999), such fibers were not observed during the devel-opmental period studied here. Though the origin of theGABA-ir fibers of the habenula/parapineal ganglion wasnot determined in the present study, the two main affer-ent neuronal populations found in adult lampreys (thesubhippocampal nucleus and the pretectal region; Yanezand Anadon, 1994b; Yanez et al., 1999) contained numer-ous GABA-ir cells in the larval stages studied here (un-published results).
CONCLUSIONS
Cell differentiation in the retina, pineal organ, and ha-benula of lampreys begins before cell differentiation of theparapineal organ, which was delayed until the larval pe-riod. However, proliferation was intense in regions of thelateral retina in the retina of medium-sized to large lar-vae, and a late-proliferating region was also observed inthe right habenula. Serotonin immunoreactivity in photo-receptive (photoneuroendocrine) cells first occurs in thepineal organ of prolarvae around the time of appearance ofphotoreceptors, whereas no serotonin immunoreactivitywas observed in the larval retina. The GABAergic struc-tures of the sea lamprey eyes develop before those of thepineal organ. The GABA-ir system of the pineal organ isafferent to different diencephalic and midbrain centers(see Yanez et al., 1993, 1999). On the other hand, the onlyGABA-ir structures observed in the parapineal ganglion oflarvae were afferent fibers, which appeared rather late indevelopment. Noticeably, the time sequence of develop-ment in these three photoreceptive structures is very dif-ferent from that observed in teleosts and other verte-brates. The GABAergic system observed in the larvallamprey retina is very different from that observed inearly retinas of other vertebrates; it might have evolved toaccomplish particular functions in a larva that lives bur-rowing in river sediment for many years. During most ofthis time, the eye does not form real visual images and isprobably little more than an ocellus-like light detector.Together, our results suggest that development of thethree photoreceptive organs in lampreys has been mark-edly specialized for the different functions they accom-plish during the larval and adult periods. Whether thischaracteristic of lamprey development is primitive or de-rived is not known, but it is intriguing that several typesof specialized photoreceptors were also found in am-phioxus (see Ruiz and Anadon, 1991a,b; Lacalli et al.,1994), a group of small burrowing prochordates (Cephalo-chordata).
ACKNOWLEDGMENTS
The University of Santiago de Compostela provided apredoctoral fellowship to M.M.-F.
LITERATURE CITED
Agardh E, Bruun A, Ehinger B, Ekstrom P, van Veen T, Wu JY. 1987.Gamma-aminobutyric acid- and glutamic acid decarboxylase-immunoreactive neurons in the retina of different vertebrates. J CompNeurol 258:622–630.
Anadon R, Melendez-Ferro M, Perez-Costas E, Pombal MA, Rodicio MC.1998. Centrifugal fibers are the only GABAergic structures of theretina of the larval sea lamprey: an immunocytochemical study. BrainRes 782:297–302.
Aoki E, Semba R, Kashiwamata S. 1989. When does GABA-like immuno-reactivity appear in the rat cerebellar GABAergic neurons? Brain Res502:245–251.
Becerra M, Manso MJ, Rodrıguez-Moldes MI, Anadon R. 1994. The struc-ture and development of dopaminergic interplexiform cells in the ret-ina of the brown trout, Salmo trutta fario: a tyrosine hydroxylaseimmunocytochemical study. J Anat 185:377–385.
Begay V, Falcon J, Cahill GM, Klein DC, Coon SL. 1998. Transcriptsencoding two melatonin synthesis enzymes in the teleost pineal organ:circadian regulation in pike and zebrafish, but not in trout. Endocri-nology 139:905–912.
Cherubini E, Gaiarsa JL, Ben-Ari Y. 1991. GABA: an excitatory transmit-ter in early postnatal life. Trends Neurosci 14:515–519.
Cole WC, Youson JH. 1982. Morphology of the pineal complex of the
263RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY

anadromous sea lamprey, Petromyzon marinus. Am J Anat 165:131–163.
Collin JP. 1969. La cupule sensorielle de l’organe pineal de la Lamproie dePlaner. L’ultrastructure des cellules sensorielles et ses implicationsfonctionelles. Arch Anat Microsc Morphol Exp 58:145–182.
Collin SP, Pottert IC. 2000. The ocular morphology of the southern hemi-sphere lamprey Mordacia mordax Richardson with special reference toa single class of photoreceptor and a retinal tapetum. Brain Behav Evol55:120–138.
Concha ML, Burdine RD, Russell C, Schier AF, Wilson SW. 2000. A nodalsignaling pathway regulates the laterality of neuroanatomical asym-metries in the zebrafish forebrain. Neuron 28:399–409.
Corujo A, Anadon R. 1986. Morfogenesis del diencefalo de la trucha ar-coiris. I. Epitalamo. Acta Microsc 9:55–66.
De Miguel E. 1984. Estudio comparado de la evolucion de la retina y techooptico de Petromyzon marinus L. durante el desarrollo y metamorfosis.Graduate Thesis, Facultad de Biologıa, Universidad Complutense, Ma-drid.
De Miguel E. 1989. Estudio del sistema visual y oculomotor de la lampreade mar Petromyzon marinus L. en estado larvario y transformacion dela retina durante la metamorfosis. PhD Thesis, Universidad de San-tiago de Compostela, Santiago de Compostela, Spain.
De Miguel E, Anadon R. 1987. Development of the retina and the optictectum of Petromyzon marinus L.: a light microscopic study. J Hirnfor-sch 28:445–456.
De Miguel E, Wagner HJ. 1990. Tyrosine hydroxylase immunoreactiveinterplexiform cells in the lamprey retina. Neurosci Lett 113:151–155.
De Miguel E, Rodicio MC, Anadon R. 1989. Ganglion cells and retinopetalfibers of the larval lamprey retina: an HRP ultrastructural study.Neurosci Lett 106:1–6.
Dickson HD, Collard TR. 1979. Retinal development in the lamprey (Petro-myzon marinus L.): premetamorphic ammocoete eye. Am J Anat 154:321–336.
Dickson HD, Graves DA. 1981. The ultrastructure and development of theeye. In: Potter IC, Hardisty MW, editors. The biology of lampreys, vol3. London: Academic Press. p 43–94.
Ekstrom P. 1987. Photoreceptors and CSF-contacting neurons in the pinealorgan of a teleost fish have direct axonal connections with the brain: anHRP-electron-microscopic study. J Neurosci 7:987–995.
Ekstrom P, Ebbesson SO. 1989. Distribution of serotonin-immunoreactiveneurons in the brain of sockeye salmon fry. J Chem Neuroanat 2:201–213.
Ekstrom P, Meissl H. 1990. Electron microscopic analysis of S-antigen- andserotonin-immunoreactive neural and sensory elements in the photo-sensory pineal organ of the salmon. J Comp Neurol 292:73–82.
Ekstrom P, Ohlin LM. 1995. Ontogeny of GABA-immunoreactive neuronsin the central nervous system in a teleost, Gasterosteus aculeatus L.J Chem Neuroanat 9:271–288.
Ekstrom P, Borg B, van Veen T. 1983. Ontogenic development of the pinealorgan, parapineal organ, and retina of the three-spined stickleback,Gasterosteus aculeatus L. (Teleostei). Cell Tissue Res 233:593–609.
Ekstrom P, van Veen T, Bruun A, Ehinger B. 1987. GABA-immunoreactiveneurons in the photosensory pineal organ of the rainbow trout: twodistinct neuronal populations. Cell Tissue Res 250:87–92.
Ekstrom P, Ostholm T, Meissl H, Bruun A, Richards JG, Møhler H. 1990.Neural elements in the pineal complex of the frog, Rana esculenta, II:GABA-immunoreactive neurons and FMRFamide-immunoreactive ef-ferent axons. Vis Neurosci 4:399–412.
Falcon J, Geffard M, Juillard MT, Steinbusch HW, Seguela P, Collin JP.1984. Immunocytochemical localization and circadian variations of se-rotonin and N-acetylserotonin in photoreceptor cells. Light and elec-tron microscopic study in the teleost pineal organ. J Histochem Cyto-chem 32:486–492.
Fischer AJ, Reh TA. 2000. Identification of a proliferating marginal zone ofretinal progenitors in postnatal chickens. Dev Biol 220:197–210.
Forsell J, Holmqvist B, Helvik JV, Ekstrom P. 1997. Role of the pinealorgan in the photoregulated hatching of the Atlantic halibut. Int J DevBiol 41:591–595.
Garcıa-Fernandez JM, Foster RG. 1994. Immunocytochemical identifica-tion of photoreceptor proteins in hypothalamic cerebrospinal fluid-contacting neurons of the larval lamprey (Petromyzon marinus). CellTissue Res 275:319–326.
Garcıa-Fernandez JM, Jimenez AJ, Gonzalez B, Pombal MA, Foster RG.1997. An immunocytochemical study of encephalic photoreceptors inthree species of lamprey. Cell Tissue Res 288:267–278.
Guerlotte J, Falcon J, Voisin P, Collin JP. 1986. Indoles in the photorecep-tor cells of the lamprey pineal complex. Ann Endocrinol 47:62–64.
Hagedorn M, Mack AF, Evans B, Fernald RD. 1998. The embryogenesis ofrod photoreceptors in the teleost fish retina, Haplochromis burtoni. DevBrain Res 108:217–227.
Hardisty MW, Potter IC. 1971. The general biology of adult lampreys. In:Hardisty MW, Potter IC, editors. The biology of lampreys, vol 1. Lon-don: Academic Press. p 127–206.
Holmgren U. 1965. On the ontogeny of the pineal and parapineal organs inteleost fishes. Progr Brain Res 10:172–182.
Hu M, Bruun A, Ehinger B. 1998. The expression of GABA(A) receptorsduring the development of the rabbit retina. Acta Ophthalmol Scand76:515–520.
Korf B. 1974. Acetylcholinesterase-positive neurons in the pineal andparapineal organs of the rainbow trout, Salmo gairdneri (with specialreference to the pineal tract). Cell Tissue Res 155:475–489.
Korf B, Rollag MD, Korf HW. 1989. Ontogenetic development of S-antigen-and rod-opsin immunoreactions in retinal and pineal photoreceptors ofXenopus laevis in relation to the onset of melatonin-dependent color-change mechanisms. Cell Tissue Res 258:319–329.
Kuratani S, Ueki T, Aizawa S, Hirano S. 1997. Peripheral development ofcranial nerves in a cyclostome, Lampetra japonica: morphological dis-tribution of nerve branches and the vertebrate body plan. J CompNeurol 384:483–500.
Kuratani S, Horigome N, Ueki T, Aizawa S, Hirano S. 1998. Stereotypedaxonal bundle formation and neuromeric patterns in embryos of acyclostome, Lampetra japonica. J Comp Neurol 391:99–114.
Lacalli TC, Holland ND, West JE. 1994. Landmarks in the anterior centralnervous system of amphioxus larvae. Phil Trans R Soc London [Biol]344:165–185.
Liang JO, Etheridge A, Hantsoo L, Rubinstein AL, Nowak SJ, IzpisuaBelmonte JC, Halpern ME. 2000. Asymmetric nodal signaling in thezebrafish diencephalon positions the pineal organ. Development 127:5101–5112.
Meiniel A. 1980. Ultrastructure of serotonin-containing cells in the pinealorgan of Lampetra planeri (Petromyzontidae). Cell Tissue Res 207:407–427.
Meiniel A, Collin JP. 1971. Le complexe pineal de l’ammocoete (Lampetraplaneri, Bl.). Identification du ganglion sous-jacent a l’organe parapi-neal et relations epithalamiques des organes pineal et parapineal. ZZellforsch 117:354–380.
Meiniel A, Hartwig HG. 1980. Indoleamines in the pineal complex ofLampetra planeri (Petromyzontidae). A fluorescence microscopic andmicrospectrofluorimetric study. J Neural Transm 48:65–83.
Meissl H, Ekstrom P. 1991. Action of gamma-aminobutyric acid (GABA) inthe isolated photosensory pineal organ. Brain Res 562:71–78.
Messersmith EK, Redburn DA. 1993. The role of GABA during develop-ment of the outer retina in the rabbit. Neurochem Res 18:463–470.
Mitchell CK, Redburn DA. 1996. GABA and GABA-A receptors are maxi-mally expressed in association with cone synaptogenesis in neonatalrabbit retina. Dev Brain Res 95:63–71.
Mitchell CK, Huang B, Redburn-Johnson DA. 1999. GABA(A) receptorimmunoreactivity is transiently expressed in the developing outer ret-ina. Vis Neurosci 16:1083–1088.
Negishi K, Wagner HJ. 1995. Differentiation of photoreceptors, glia, andneurons in the retina of the cichlid fish Aequidens pulcher: an immu-nocytochemical study. Dev Brain Res 89:87–102.
Negishi K, Kiyama H, Kato S, Teranishi T, Hatakenaka S, Katayama Y,Miki N, Tohyama M. 1986. An immunohistochemical study of the riverlamprey retina. Brain Res 362:389–393.
Negishi K, Stell WK, Takasaki Y. 1990a. Early histogenesis of the te-leostean retina: studies using a novel immunochemical marker, prolif-erating cell nuclear antigen (PCNA/cyclin). Dev Brain Res 55:121–125.
Negishi K, Teranishi T, Karkhanis A, Stell WK. 1990b. Emergence anddevelopment of immunoreactive cells in teleostean retinas during theperinatal period. Dev Brain Res 55:127–137.
Obata K. 1997. Excitatory and trophic action of GABA and related sub-stances in newborn mice and organotypic cerebellar culture. Dev Neu-rosci 19:117–119.
Ostholm T, Ekstrom P, Bruun A, van Veen T. 1988. Temporal disparity inpineal and retinal ontogeny. Brain Res 470:1–13.
Piavis GW. 1971. Embryology. In: Hardisty MW, Potter IC, editors. Thebiology of lampreys, vol 1. London: Academic Press. p 361–400.
Pombal MA, Yanez J, Marın O, Gonzalez A, Anadon R. 1999. Cholinergicand GABAergic neuronal elements in the pineal organ of lampreys, and
264 M. MELENDEZ-FERRO ET AL.

tract-tracing observations of differential connections of pinealofugalneurons. Cell Tissue Res 295:215–223
Redburn-Johnson D. 1998. GABA as a developmental neurotransmitter inthe outer plexiform layer of the vertebrate retina. Perspect Dev Neu-robiol 5:261–277.
Retzius G. 1895. Uber den Bau des sogennanten Parietalauges von Am-mocoetes. Biol Untersuch NF Bd VII.
Rio JP, Vesselkin NP, Kirpitchnikova E, Kenigfest NB, Versaux-Botteri C,Reperant J. 1993. Presumptive GABAergic centrifugal input to thelamprey retina: a double-labeling study with axonal tracing and GABAimmunocytochemistry. Brain Res 600:9–19.
Rio JP, Vesselkin NP, Reperant J, Kenigfest NB, Miceli D, Adanina VO.1996. Retinal and nonretinal inputs upon retinopetal RMA neurons inthe lamprey: a light and electron microscopic study combining HRPaxonal tracing and GABA immunocytochemistry. J Chem Neuroanat12:51–70.
Robinson J, Schmitt EA, Dowling JE. 1995. Temporal and spatial patternsof opsin gene expression in zebrafish (Danio rerio). Vis Neurosci 12:895–906.
Rodicio MC, Pombal MA, Anadon R. 1995. Early development and organi-zation of the retinopetal system in larval sea lamprey, Petromyzonmarinus L. An HRP study. Anat Embryol 192:517–526.
Rubinson K, Cain H. 1989. Neural differentiation in the retina of the larvalsea lamprey (Petromyzon marinus). Vis Neurosci 3:241–248.
Ruiz MS, Anadon R. 1991a. The fine structure of the lamellate cells in thebrain of amphioxus (Branchiostoma lanceolatum, Cephalochordata).Cell Tissue Res 263:597–600.
Ruiz MS, Anadon R. 1991b. Some considerations on the fine structure ofthe rhabdomeric photoreceptors in the amphioxus, Branchiostoma lan-ceolatum (Cephalochordata). J Hirnforsch 32:159–164.
Saha MS, Grainger RM. 1993. Early opsin expression in Xenopus embryosprecedes photoreceptor differentiation. Mol Brain Res 17:307–318.
Samejima M, Tamotsu S, Watanabe K, Morita Y. 1989. Photoreceptor cellsand neural elements with long axonal processes in the pineal organ ofthe lamprey, Lampetra japonica, identified by use of the horseradishperoxidase method. Cell Tissue Res 258:219–224.
Sandell JH. 1998. GABA as a developmental signal in the inner retina andoptic nerve. Perspect Dev Neurobiol 5:269–278.
Sandell JH, Martin SC, Heinrich G. 1994. The development of GABAimmunoreactivity in the retina of the zebrafish (Brachidanio rerio).J Comp Neurol 345:596–601.
Sears S, Erickson A, Hendrickson A. 2000. The spatial and temporalexpression of outer segment proteins during development of Macacamonkey cones. Invest Ophthalmol Vis Sci 41:971–979.
Sternberger LA, Hardy PH, Cuculis JJ, Meyer HG. 1970. The unlabeledantibody enzyme method of immunocytochemistry: preparation andproperties of soluble antigen-antibody complex (horseradishperoxidase-antiperoxidase) and its use in identification of spirochetes.J Histochem Cytochem 18:315–333.
Studnicka FC. 1893. Sur les organes parietaux de Petromyzon planeri.Sitzungs Gesell Wissensch (Prague). p 1–50.
Studnicka FK. 1905. Die Parietalorgane. In Oppel A, editor. Lehrbuch dervergleichenden mikroskopischen Anatomie der Wirbeltiere, pt 5. Jena:Gustav Fischer. p 1–254.
Studnicka FK. 1912. Ueber die Entwicklung und die Bedeutung derSeitenaugen von Ammocoetes. Anat Anz 41:561–578.
Tahara Y. 1988. Normal stages of development in the lamprey, Lampetrareissneri (Dybowski). Zool Sci 5:109–118.
Tamotsu S, Morita Y. 1986. Photoreception in pineal organs of larval andadult lampreys, Lampetra japonica. J Comp Physiol A159:1–5.
Tamotsu S, Korf H-W, Morita Y, Oksche A. 1990. Immunocytochemicallocalization of serotonin and photoreceptor-specific proteins (rod-opsin,S-antigen) in the pineal complex of the river lamprey, Lampetra ja-ponica, with special reference to photoneuroendocrine cells. Cell TissueRes 262:205–216.
Tamotsu S, Oishi T, Nakao K, Fukada Y, Shichida Y, Yoshizawa T, MoritaY. 1994. Localization of iodopsin and rod-opsin immunoreactivity in theretina and pineal complex of the river lamprey, Lampetra japonica.Cell Tissue Res 278:1–10.
Tamotsu S, Samejima M, Suzuki N, Morita Y. 1997. Three-dimensionalreconstruction of serotonin-immunoreactive photoreceptors in the pi-neal organ of the river lamprey, Lampetra japonica. Biol Signals6:184–190.
van Veen T, Ekstrom P, Borg B, Møller M. 1980. The pineal complex of thethree-spined stickleback, Gasterosteus aculeatus L.: a light-, electronmicroscopic and fluorescence histochemical investigation. Cell TissueRes 209:11–28.
van Veen T, Ekstrom P, Nyberg L, Borg B, Vigh-Teichmann I, Vigh B.1984. Serotonin and opsin immunoreactivities in the developing pinealorgan of the three-spined stickleback, Gasterosteus aculeatus L. CellTissue Res 237:559–564.
Versaux-Botteri C, Dalil N, Kenigfest N, Reperant J, Vesselkin N, Nguyen-Legros J. 1991. Immunohistochemical study of retinal serotonin cell inthe lamprey (Lampetra fluviatilis). Vis Neurosci 7:171–177.
Vesselkin NP, Reperant J, Kenigfest NB, Rio JP, Miceli D, Shupliakov OV.1989. Centrifugal innervation of the lamprey retina. Light- andelectron-microscopic and electrophysiological investigations. Brain Res493:51–65.
Vesselkin NP, Rio JP, Reperant J, Kenigfest NB, Adanina VO. 1996.Retinopetal projections in lampreys. Brain Behav Evol 48:277–286.
Vigh-Teichmann I, Vigh B, Wirtz GH. 1989. Immunoelectron microscopy ofrhodopsin and vitamin A in the pineal organ and lateral eye of lamprey.Exp Biol. 48:203–213.
Vigh-Teichmann I, Ali MA, Szel A, Vigh B. 1991. Ultrastructure and opsinimmunocytochemistry of the pineal complex of the larval Arctic charrSalvelinus alpinus: a comparison with the retina. J Pineal Res 10:196–209.
Wake k. 1973. Acetylcholinesterase-containing cells and their distributionin the pineal complex of the goldfish, Carassius auratus. Z Zellforsch145:287–298.
Wullimann MF, Puelles L. 1999. Postembryonic neural proliferation in thezebrafish forebrain and its relationship to prosomeric domains. AnatEmbryol 199:329–348.
Yanez J, Anadon R. 1994a. Are the dopaminergic cells of the lampreyretina interplexiform cells? Neurosci Lett 165:63–66.
Yanez J, Anadon R. 1994b. Afferent and efferent connections of the habe-nula in the larval sea lamprey (Petromyzon marinus L.). An experi-mental study. J Comp Neurol 345:148–160.
Yanez J, Anadon R, Holmqvist BI, Ekstrom P. 1993. Neural projections ofthe pineal organ in the larval lamprey (Petromyzon marinus L.) re-vealed by indocarbocyanine dye tracing. Neurosci Lett 164:213–216.
Yanez J, Meissl H, Anadon R. 1996. Central projections of the parapinealorgan of the adult rainbow trout (Oncorhynchus mykiss). Cell TissueRes 285:69–74.
Yanez J, Pombal MA, Anadon R. 1999. Afferent and efferent connections ofthe parapineal organ in lamprey: a tract tracing and immunocytochem-ical study. J Comp Neurol 403:171–189.
265RETINA AND PINEAL COMPLEX DEVELOPMENT IN LAMPREY




















