
Chronic exposure to 60Hz electric fields: Effects on pineal function in the rat
10
Bioelectromagnetics 2371-380 (1981) Chronic Exposure to 60-Hz Electric Fields: Effects on Pineal Function in the Rat Bary W. Wilson, Larry E. Anderson, D. Ian Hilton, and Richard D. Phillips Physical Sciences Department (8. W W) and Biology Department (L. E.A., D.I. H., R. D.P), Battelle Memorial Institute, Pacific Northwest Laboratory, Richland, Mshington As a component of studies to search for effects of 60-Hz electric field exposure on mamma- lian endocrine function, concentrations of melatonin, 5-methoxytryptophol, and serotonin-N- acetyl transferase activity were measured in the pineal glands of rats exposed or sham-exposed at 65 kV/m for 30 days. In two replicate experiments there were statistically significant differences between exposed and control rats in that the normal nocturnal increase in pineal melatonin content was depressed in the exposed animals. Concentnitions of 5-methoxytryp- tophol were increased in the pineal glands of the exposed groups when compared to sham- exposed controls. An alteration was also observed in serotonin-N-acetyl transferase activity, with lower levels measured in pineal glands from exposed animals. Key words: electric fields, 60 Hz, pineal gland, circadian rhythm, melatonin, 5 methoxytryptophol, acetyl transferase INTRODUCTION In the last decade, a number of studies have been conducted to determine if exposure to extremely low frequency (ELF) electric fields can give rise to measurable biologic effects. From such studies several lines of evidence have resulted that indicate that ELF exposure may affect the central nervous system (CNS). One of the earlier reports of a possible interaction between ELF and the CNS came from a Russian study in which a variety of neurologic symptoms including headache, excitability, and fa- tigue were reported in workers in ultrahigh-voltage (? 345 kV) switchyards [Asanova and Rakov, 1966; Korobkova et al, 19721. While much of this information must be considered anecdotal because of the lack of an appropriate control group, it has prompted further investigation of ELF exposure using animal models. pigeons are able to detect the presence of an electric field at strengths as low as 32 Behavioral studies by Graves et a1 [1978], for example, have demonstrated that Received for review April 1, 1981; revision received August 31, 1981. Address reprint requests to B. W. Wilson, Physical Sciences Department, Battelle, Pacific Northwest Labo- ratory. P.O. Box 999, Richland, WA 99352. 0197-846218110204-0371$03.00 0 1981 Alan R. Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Chronic exposure to 60Hz electric fields: Effects on pineal function in the rat

Bioelectromagnetics 2371 -380 (1981)
Chronic Exposure to 60-Hz Electric Fields: Effects on Pineal Function in the Rat Bary W. Wilson, Larry E. Anderson, D. Ian Hilton, and Richard D. Phillips Physical Sciences Department (8. W W) and Biology Department (L. E.A., D.I. H., R. D.P), Battelle Memorial Institute, Pacific Northwest Laboratory, Richland, Mshington
As a component of studies to search for effects of 60-Hz electric field exposure on mamma- lian endocrine function, concentrations of melatonin, 5-methoxytryptophol, and serotonin-N- acetyl transferase activity were measured in the pineal glands of rats exposed or sham-exposed at 65 kV/m for 30 days. In two replicate experiments there were statistically significant differences between exposed and control rats in that the normal nocturnal increase in pineal melatonin content was depressed in the exposed animals. Concentnitions of 5-methoxytryp- tophol were increased in the pineal glands of the exposed groups when compared to sham- exposed controls. An alteration was also observed in serotonin-N-acetyl transferase activity, with lower levels measured in pineal glands from exposed animals.
Key words: electric fields, 60 Hz, pineal gland, circadian rhythm, melatonin, 5 methoxytryptophol, acetyl transferase
INTRODUCTION
In the last decade, a number of studies have been conducted to determine if exposure to extremely low frequency (ELF) electric fields can give rise to measurable biologic effects. From such studies several lines of evidence have resulted that indicate that ELF exposure may affect the central nervous system (CNS). One of the earlier reports of a possible interaction between ELF and the CNS came from a Russian study in which a variety of neurologic symptoms including headache, excitability, and fa- tigue were reported in workers in ultrahigh-voltage (? 345 kV) switchyards [Asanova and Rakov, 1966; Korobkova et al, 19721. While much of this information must be considered anecdotal because of the lack of an appropriate control group, it has prompted further investigation of ELF exposure using animal models.
pigeons are able to detect the presence of an electric field at strengths as low as 32 Behavioral studies by Graves et a1 [1978], for example, have demonstrated that
Received for review April 1, 1981; revision received August 31, 1981.
Address reprint requests to B. W. Wilson, Physical Sciences Department, Battelle, Pacific Northwest Labo- ratory. P.O. Box 999, Richland, WA 99352.
0197-846218110204-0371$03.00 0 1981 Alan R. Liss, Inc.

372 Wilson et al
kV/m. Additional evidence of CNS interaction with ELF fields has been reported by Bayer et a1 [1977]; rats exposed to 50-Hz electric fields were able to perceive and avoid the field at 100 kV/m. Extending these observations, Hjeresen et a1 [1980], in this laboratory, have shown that rats exposed to 60-Hz electric fields at field strengths of 25-50 kV/m, spent most of their daily inactive period in the field; those exposed at field strengths greater than 75 kV/m spent most of the inactive period out of the field. Stern et al [1980] have reported data from preliminary studies that indicate that rats may be able to detect fields as low as 5 kV/m.
The ability of monkeys to judge accurately the passage of time has been shown to be affected by exposure to ELF fields of 7-75 Hz [Gavalas-Medici and Magada- leno, 19751. During and following the exposure, the animals, which had been trained to press a bar at 5- to 7.5-s intervals, showed a decrease of up to 60% in interresponse time.
As a measure of brain activity, electroencephalograms (EEGs) have been re- corded during and following exposure to ELF electric fields. At a variety of frequen- cies and field strengths, exposure appears to cause significant changes in EEG patterns of monkeys [Gavalas et al, 19701, cats [Bawin et al, 19731, mice and guinea pigs [Blanchi et al, 19731, and rats [Lott and McCain, 19731. Jaffe et al [1980] have re- ported additional neurophysiologic evidence that exposure to 60-Hz electric fields significantly increases the synaptic excitability of the superior cervical ganglia in rats.
and neuroendocrinologic effects of electric field exposure. Carmaciu et a1 [ 19771 have reported that rats exposed to 50-Hz, 100-kVIM electric fields for 2 h had significantly higher levels of plasma antidiuretic hormone compared to controls. Later work from the same laboratory [Groza et al, 19781 showed that rats exposed to 50-Hz, 100-kV/M fields for 6 h or 12 days had altered levels of urinary and blood catecholamines. Free et a1 [1981] have reported some subtle changes in endocrine levels in rats exposed to 60-Hz fields. However, in another laboratory, investigators thus far have failed to identify any effects of 60-Hz electric field exposure on various endocrine parameters [Quinlan et al, 19801.
may affect neuronally regulated biologic rhythms. Wever [1973] has shown that when humans are deprived of external time cues, the internal desynchronization and phase shifts that occur in circadian periodicity were arrested with the imposition of an ELF field.
The involvement of the pineal gland in mammalian circadian rhythms suggests that it might play a role in the interaction of the CNS with the ELF fields. The pineal gland of the rat weighs about 1 mg and is located outside the blood-brain bamer in the epithalamic region of the forebrain. There are enzyme systems contained within the pinealocytes of the gland that metabolize serotonin to other indoleamines (Fig. 1) that exhibit circadian rhythmicity in the pineal gland, such as melatonin [Wurtman and Axelrod, 1965al and 5-methoxytryptophol (5-MTOL) [Wilson et al, 19781. The organ is well vascularized and is innervated by adrenergic and some cholinergic autonomic fibers. The gland responds to stimulation via neuronal inputs (specifically, the release of norepinephrine from adrenergic fibers originating in the superior cervical ganglion) to increase the synthesis and release of melatonin into the systemic blood circulation. Hence, the pineal gland has been designated a ‘ ‘neuroendocrine transducer” [Wurtman and Axelrod, 1965bI.
Recent reports in the literature indicate that there may be some endocrinologic
Results from other experiments have suggested that exposure to electric fields

ELF Effects on the Pineal Gland 373
NOR EPlN EPHRINE (NEURONAL INPUT)
TRYPTOPHAN I BETA ADRENERGIC RECEPTOR 1 + TRYPTOPHAN
ATP CAMP I
5-OH TRYPTOPHAN I + +
RNA SYNTHESIS I + SEROTONIN
1 s N A i + PROTEIN SYNTHESIS
5-OH N ACETYL SEROTONIN TRYPTOPHOL
PINEAL GLAND + HIOMT -1 HlOMT
5 -M ETHOXY- TRY PTO PH 0 L
M ELATON IN
1 ME LATON IN
-1 5- M ETH OXY - TRYPTOPHOL
Fig. 1 . Production of melatonin and 5-MTOL in the rat pineal gland modulated by neumnal input from sympathetic autonomic ganglia. Abbreviations: MAO, monoamine oxidase; SNAT, serotonin N-acetyl transferase; HIOMT, hydmxyindole-0-methyl transferase; AR, acetaldehyde reductase.
We postulated that the pineal, in its role of modulating internal rhythms with external stimuli, might be used as a model for studying the neural effects of ELF exposure. Effects of electric field exposure on pineal function were investigated by comparing indole hormone production in exposed and control populations. We pursued the metabolic pathway in which serotonin is acetylated by serotonin-N-acetyl trans- ferase (SNAT) and subsequently methylated to melatonin. In two separate experiments we exposed rats to 60-Hz electric fields at 65 kV/m for 30 days (20 h/day), and mea- sured pineal hormone concentrations of melatonin as well as SNAT activity. We also measured 5-MTOL (Fig. l), a product of serotonin metabolism ,from an alternate path- way.
METHODS Animals
Male Sprague-Dawley-derived rats (Charles River SPF, Portage, Minnesota) were carefully examined on delivery to verify that they were healthy before being used in experiments. A sample population from each shipment was sacrificed for virologic screening and histopathologic examination. The remaining animals were maintained in standard cages in an isolation room for 2 weeks, ear-coded, and then acclimated for 14

374 Wilson et al
days in cages identical to those used to house animals in the electric field exposure systems. Animals were acclimated to a daily 14-h light : 10-h dark photoperiod, and were provided water and food ad libitum, Temperatures in the isolation and acclima- tion rooms were maintained at 21 “C (20-23 “C range). Relative humidity was 20-40 % .
After acclimation, the animals for each experiment were randomly subdivided into two groups (exposed and sham-exposed) for subsequent treatment. The animals were 56 days of age at the start of exposure. In the first study, melatonin and 5- MTOL levels were measured in 20 exposed and 20 sham-exposed animals. In the second experiment, both melatonin and SNAT levels were assayed in 20 exposed and 20 sham-exposed animals.
for exposed and sham-exposed animals, except for the presence or absence of the electric field. At the end of 30 days of exposure, animals were killed in groups of 10 (5 exposed, 5 sham-exposed) at four different times during the light/dark cycle. The electric field was turned off, and remained off, throughout the sacrifice times, which were at 1400, 2200, 0200 and 0800h. After animals were decapitated, the pineal gland was removed and quickly frozen on dry ice, usually within 2 min of death.
Electric Field Exposure The system used to expose rats to a uniform, vertical 60-Hz electric field has
been described by Jaffe et a1 [1980], and by Hilton and Phillips [1980]. Briefly, the rats were exposed in a parallel-plate exposure system, with the rat in electrical contact with the reference ground electrode. In both experiments, animals were exposed at an unperturbed field strength of 65 kV/m, for 20 h/day over 30 days. Because animals in adjacent cages shield one another, unperturbed field strength was reduced by approxi- mately 35% [Kaune, 19811; therefore, the “effective” field strength in this study was approximately 39 kV/m.
Gas ChromatographiclMass Spectrometric Assay for Melatonin and 5-Methoxytryptophol
melatonin and 5-MTOL according to a modification of the method of Wilson et a1 [ 19771. A gas chromatography/mass spectrometry (GC/MS) data system, (Hewlett- Packard 5985A) fitted with a 2-m X 4-mm ID glass column packed with OV-17 liquid phase, was used for low-resolution measurements. Analyses at intermediate and high mass spectral resolution were performed with fused silica capillary columns coated with SE52 stationary phase, on a double-focusing mass spectrometer (VG Micromass ZAB-IF) interfaced with a GC (HP5840A) operated in the direct splitless injection mode. The fused silica column was inserted directly into the ion source, resulting in superior gas chromatographic fidelity and minimal sample loss in the G U M S inter- face. Both melatonin and 5-MTOL were assayed in the same GUMS runs by selected ion monitoring using an internal standard [Wilson et al, 19781. Standard solutions of both melatonin and 5-methoxytryptophol containing internal standard were prepared and analyzed. The intensities (peak height) of the molecular ions from the two com- pounds were normalized to that of the internal standard generating standard curves. The assay was run in duplicate and samples were picked at random for multiple injec- tion. Variation in the ratios for multiple injections (three or more) did not exceed 15%
During each experiment, environment, housing, and husbandry were identical
Pineal glands were sonicated in 0.5 ml of H,O, and the sonicate was assayed for

ELF Effects on the Pineal Gland 375
and were typically 3-5 % . Solvent and extracted water blanks were analyzed periodi- cally throughout the assay. Melatonin and 5-MTOL were generally not detected in the blanks, and when present, they did not exceed a ratio corresponding to 10 pg per pineal.
Enzyme Assay
nate was adjusted to pH 6.8 with 0.1 ml, 0.2 M, 2-(N-morpholino)-ethanesulfonic acid (MES). The homogenates were centrifuged 5 min at 10,OOOg and the supernatant was subsequently analyzed for SNAT activity by means of a modification of the method described by Deguchi and Axelrod [1972]. The MES buffer (25 pl, 0.2 M) was added to 50 or 100 pl of pineal supernatant, 1 pmol tryptamine hydrochloride, and 17.5 pmol l-(I4C)-acetyl CoA (specific activity, 58 pCi/pmol). This mixture was incubated for 15 min at 37 "C, and the reaction was stopped by addition of 0.5 ml of 0.45 M borate buffer. The reaction mixture was subsequently transferred to a glass tube containing 6 ml toluene-isoamyl alcohol (97:3) and vortexed for 30 s. Following separation of the phases, an aliquot of the organic phase was transferred to 10 ml scintillation fluid (Liquiflor, New England Nuclear) and radioactivity was counted in a liquid scintillation spectrometer (Beckmann LS 9000).
Statistical Analysis
Data on pineal indole hormone concentrations were analyzed by means of a one-way analysis of variance, assuming a completely randomized experimental design, and a 2 X 4 factorial treatment design for each of the two hormones, melatonin and 5-MTOL. Prior to analysis, the original data were transformed using natural loga- rithms to stabilize the variance among treatments. SNAT activity data were analyzed by the unequal variance t test.
Each pineal gland was sonicated in 0.5 ml of H,O, and 0.1 ml of the homoge-
RESULTS
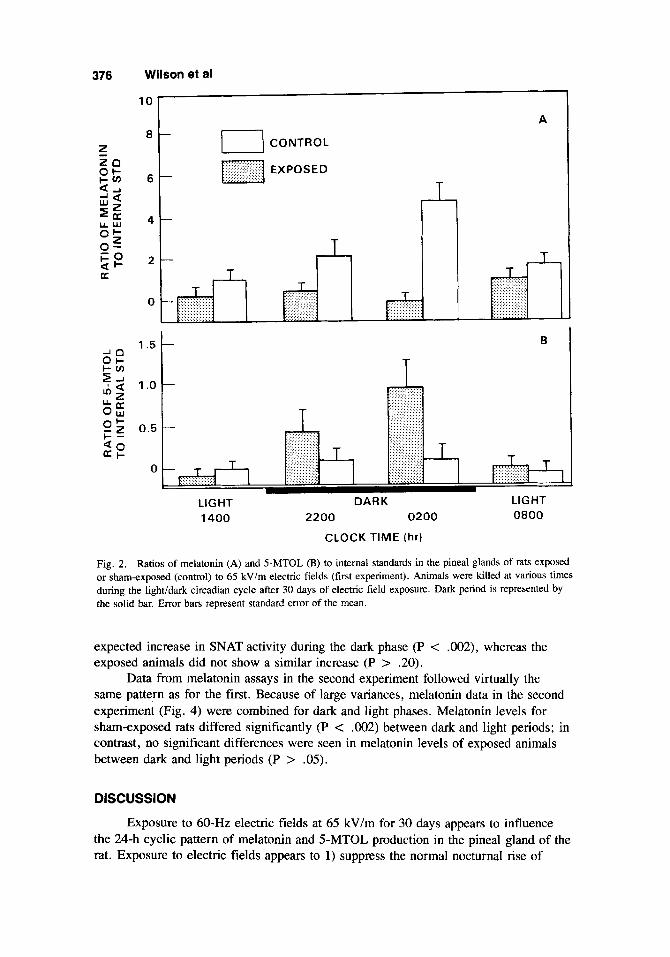
Data presented in Figure 2A show the ratio of pineal melatonin to internal stan- dard for control and exposed animals sacrificed at various times during the light/dark cycle after 30 days of exposure. Melatonin levels from exposed animals were sup- pressed during the dark phase when compared to sham-exposed controls. Exposed rats, sacrificed at 0200 h, showed a significant (P < .05) reduction in melatonin levels compared to controls. During the dark phase, there was a significant increase (P c . O l ) in pineal melatonin content for sham-exposed animals; however, there was no increase in melatonin in the exposed animals (P < .20).
During the dark phase, pineal 5-MTOL levels (Fig. 2B) in the exposed group tended to be higher than in the controls. However, the differences between the two groups at the four sampling periods were not statistically significant (P > .05).
during the lightldark cycle. The apparent reduction in SNAT values in exposed ani- mals was not significantly different from values of sham-exposed rats at any one of the four sampling times. Because of the large variance in the data, the two sampling times in the dark period (2200 and 0200 h) were combined and compared to the combined light-period sampling times (1400 and 0800 h). The sham-exposed animals showed the
In the second experiment, SNAT activity (Fig. 3) was measured at various times
376 Wilson et al
'k T I CONTROL
1 T I I I I f
A
B B
LIGHT DARK LIGHT 1400 2200 0200 0800
CLOCK TIME (hr)
LIGHT DARK LIGHT 1400 2200 0200 0800
CLOCK TIME (hr)
Fig. 2. Ratios of melatonin (A) and 5-MTOL (B) to internal standards in the pineal glands of rats exposed or shamexposed (control) to 65 kVlm electric fields (first experiment). Animals were killed at various times during the lightidark circadian cycle after 30 days of electric field exposure. Dark period is represented by the solid bar. Error bars represent standard error of the mean.
expected increase in SNAT activity during the dark phase (P < .002), whereas the exposed animals did not show a similar increase (P > .20).
Data from melatonin assays in the second experiment followed virtually the same pattern as for the first. Because of large variances, melatonin data in the second experiment (Fig. 4) were combined for dark and light phases. Melatonin levels for sham-exposed rats differed significantly (P < .002) between dark and light periods; in contrast, no significant differences were seen in melatonin levels of exposed animals between dark and light periods (P > .05).
DISCUSSION
Exposure to 60-Hz electric fields at 65 kV/m for 30 days appears to influence the 24-h cyclic pattern of melatonin and 5-MTOL production in the pineal gland of the rat. Exposure to electric fields appears to 1) suppress the normal nocturnal rise of

, 7 CON
TR
OL
....
....
....
. ..
....
....
...
EX
PO
SE
D
....
....
....
. ..
....
....
..
....
....
..
....
....
...
....
....
....
. ..
....
....
..
......
......
.....
l2
F
....
....
....
. ..
....
....
...
T
4 ..
...
....
. ..
...
....
. ...
.. ...
.. ..
...
.....
....
. ...
.. -r ..... ..... ., ,
, ..
...
....
. ..
...
....
. ..
...
....
. ...
.. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ..
....
....
. ...
......
..
T
d-1
..........
..........
. ..
....
....
..
....
....
. ..
....
....
..
....
....
. ....
... I ...
........
.. ..
....
....
LIG
HT
D
AR
K
LIG
HT
1
40
0
22
00
0
20
0
08
00
C
LO
CK
TIM
E (
hr)
Fig.
3.
Est
imat
ed p
inea
l SN
AT
act
ivity
in r
ats e
xpos
ed o
r sh
am-e
xpos
ed (c
ontr
ol) t
o el
ectr
ic fi
elds
at
65
kV/m
(se
cond
exp
erim
ent)
. Ani
mal
s w
ere
kille
d at
var
ious
tim
es d
urin
g th
e lig
hvda
rk c
irca
dian
cyc
le a
fter
30
day
s of
ele
ctri
c fie
ld e
xpos
ure.
Dar
k pe
riod
is r
epre
sent
ed b
y th
e so
lid b
ar. E
rror
bar
s re
pres
ent s
tand
ard
erro
r of
the
mea
n.
0 3
al 9
0
-4
-4

378 Wilson et al
Is) 0 - $ w 200 z a K W 0 z
4 g 100 I-
W 5
0 DARK LIGHT DARK LIGHT
SHAM-EXPOSED EXPOSED
Fig. 4. Pineal melatonin levels in rats exposed or sham-exposed to electric fields at 65 kV/m for 30 days (second experiment). Levels shown are those obtained during the dark or light phases of the diurnal cycle, immediately following electric field exposure. Error bars represent standard e m r of the mean.
pineal melatonin, 2) cause a delay and/or suppress the expected nocturnal increase in pineal SNAT activity, and 3) leads to an increase in pineal 5-MTOL content during the nocturnal period of the circadian cycle.
The reduced levels of pineal melatonin in the exposed animals during the daily cycle, especially during the dark phase, is probably due to an overall decrease in the production of pineal melatonin. A comparison of the patterns in SNAT activity at each of the four time points for the exposed and control groups suggests a possible delay or shift in the expected rhythm of SNAT; however, data from additional sampling times would be required to substantiate this hypothesis.
The increase in pineal 5-MTOL in the exposed group during the dark period suggests that exposure to electric fields may influence the biochemistry of the SNAT pathway for serotonin metabolism (Fig. 1). The final 5-MTOL concentration is dcpen- dent not only upon enzyme activity, but also upon substrate availability. It has been suggested that the release of “bound” serotonin may control the rate at which 5- MTOL and melatonin are produced [Wurtman and Ozaki, 19781. The increase in sub- strate availability for conversion into 5-MTOL, in this instance, may result from inter- ference with the SNAT pathway, which normally metabolizes much of the available serotonin in the pineal to melatonin. Experiments are in progress to measure serotonin levels as well as 5-MTOL and melatonin.
The response of the pineal gland to electric fields may be mediated by various humoral agents (ie, hormones, neurotransmitters), or by changes in the firing rate or pattern of neuronal input. These factors, in turn, may influence the sensitivity of the 0-adrenergic receptors, which are regulated at multiple sites [Zatz, 19781. We are aware of no experimental evidence suggesting that electric fields affect the levels of any humoral factor that might influence the production of pineal indoleamines. How- ever, there is some evidence that neuronal excitability may be altered in animals ex- posed to electric fields [Jaffe et al, 19801.
A number of factors can suppress the rhythmicity of pineal melatonin produc-

ELF Effects on the Pineal Gland 379
tion, including constant light [wurtman et al, 19641, bilateral superior ganglionectomy purtman et al, 19641, and treatment with various agents that interact with 6- adrenergic receptors [Deguchi and Axelrod, 19731. Biologic effects such as those ob- served in this and other studies [Hjeresen et al, 1980; Graves et al, 1979; Jaffe et al, 19801 may be the result of chronic low-level stimulation of the nervous system by the electric fields. This chronic stimulation may mimic constant photic input or may, in some way, alter normal neuronal activity.
ACKNOWLEDGMENTS
The authors thank Sally Hewitt, Laura Winn, and Jim Chandon for expert tech-
This work was supported by the U.S. Department of Energy under Contract nical assistance.
DEIAC06-76RLO 1830.
REFERENCES Asanova TP, Rakov A1 (1966): The state of health of persons working in electric fields of outdoor 400 and
500 kV switchyards. In “Hygiene of Labor Professional Diseases” Vol. 5.” Knickerbocker G (trans). Special Publication No. 10, 1975. Piscataway: IEEE Power Engineering Society.
Bawin SM, Gavalas-Medici RJ, Adey WR (1973): Effects of modulated very high frequency fields on specific brain rhythms in cats. Brain Res 58:365-384.
Bayer A, Brinkman J , Wittke G (1977): Experimentelle untersuchungen an ratten zur frage der wirkung electrischer wechselfelder auf lebewesen. [Experimental research on rats for determining the effect of electrical AC fields on living beings]. Elektrizitaetswirtschaft 4:77-81.
fields. 2. Effect on heart’s and brain’s electrical activity. Arch Fisiol 70:33-34.
diuretic hormone in rats. Physiologie 14:79-83.
Biochem 50: 174-179.
regulated by catecholamine transmitters. Pmc Natl Acad Sci USA 70:2411-2414.
Bioelectromagnetics 2 : 105- 122.
75-Hz Electric Fields on Neurophysiology and Behavior of Monkeys. Phase 1: Continuous Wave,” Technical Report, April 1976, No. AD-A008404/6ST. Springfield: NTIS.
Gavalas-Medici RJ, Watter DO, Hamer J , Adey WR (1970): Effect of low-level, low-frequency electric fields on EEG and behavior in Mucacu nemestrim. Brain Res 18:491-501.
Graves HB, Carter JH, Kellmel D, Cooper L (1978): Perceptibility and electrophysiological response of small birds to intense 60-Hz electric fields. IEEE Trans Power Appar Syst 97:1070-1073.
Graves HB, Long PD, Poznaniak D (1979): Biological effects of 60-Hz alternating-current fields: A chesh- ire cat phenomenon, pp 184-197. In “Biological Effects of Extremely Low-Frequency Electrnrnag- netic Fields.” Proceedings of the 18th Annual Hanford Life Sciences Symposium, October 16-18, Richland, Washington. CONF-781016. Springfield: NTIS.
high voltage electric field. Physiologie 15: 139-144.
magnetics 1:55-64.
behavior and activity of rats. Bioelectrumagnetics 1999-3 12.
on synaptic transmission and peripheral nerve function in the rat. Bioelectromagnetics 1 : 131-148.
Blanchi D, Cedrini L, Ceria F, Meda E, Re GG (1973): Exposure of mammalians to strong 50-Hz electric
Carmaciu R, Groza P, Daneluic E (1977): Effects of a high-tension electric field on the secretion of anti-
Deguchi T, Axelrod J (1972): Sensitive assay for serotonin N-acetyltransferase activity in rat pineal. Anal
Deguchi T, Axelrod J (1973): Supersensitivity and subsensitivity of the P-adrenergic receptor in pineal gland
Free MJ, Kaune WT, Phillips RD (1981): Endocrinological effects of strong 60-Hz electric fields on rats.
Gavalas-Medici RJ, Magadaleno SR (1975): “An Evaluation of Possible Effects of 45-Hz, 60-Hz and
Groza P, Carmaciu R, Bubuianu E (1978): Blood and urinary catecholamine variations under the action of a
Hilton DI. Phillips RD (1980): Cardiovascular response of rats exposed to 60-Hz electric fields. Bioelectro-
Hjeresen DL, Kaune WT, Decker JR, Phillips RD (1980): Effects of 60-Hz electric fields on avoidance
Jaffe RA, Laszewski BL, Carr DB, Phillips RD (1980): Chronic exposure to a 60-Hz electric field: Effects

380 Wilson et al
Kaune WT (1981): Interactive effects of 60-Hz electric field exposure systems. Bioelectromagnetics
Korobkova VP, Morozov UA, Stolarov MS, Yakub YA (1972): Influence of electric field in 500 and 750-kV 2 133-50.
switchyards on maintenance staff and means for its protection. CIGRE 2396. International Confer- ence on Large High Tension Electric Systems (28 August 1972).
in rats. Int J Biometeorol 17:221-225.
to 60-Hz power frequency. (Abstract). Bioelectromagnetics 1:243.
inary results. (Abstract.) Bioelectromagnetics 1:243.
aspects of these studies. Int J Biometeorol 17:227-232.
spectrometry method for the quantitative analysis of melatonin in plasma and cerebrospinal fluid. Anal Biochem 81:283-291.
tion, and evidence for daily rhythmicity. Life Sci 23:1019-1024.
ated by the sympathetic nervous system. Science 143:1328-1330.
mals. Prog Brain Res 10:520.
Lott JR, McCain HB (1973): Some effects of continuous and pulsating electric fields on brain wave activity
Quinlan WJ, Lebda N, Michaelson SM, Pettit S, Power S (1980): Neuroendocrine response of rats exposcd
Stern S, Laties VG, Stancampiano C, deLorge JO (1980): Detection of 60-Hz electric fields by rats: Prelim-
Wever R (1973): Human circadian rhythms under the influence of weak electric fields and the different
Wilson BW, Snedden W, Mullen PE, Sillman RE, Smith I, Landon J (1977): A gas chromatography-mass
Wilson, BW, Lynch HJ, Oraki Y (1978): 5-Methoxytryptophol in rat serum and pineal: Detection, quantita-
Wurtman RJ, Axelrod J, Fischer JE (1964): Melatonin synthesis in the pineal gland: Effect of light medi-
Wurtman RJ, Axelrod J (1965a): The formation, metabolism, and physiologic effects of melatonin in matn-
Wurtman RJ, Axelrod J (1965b): The pineal gland. Sci Am 213:50-60. Wurtman RJ, Ozaki Y (1978): Physiological control of melatonin synthesis and secretion: Mechanisms
generating rhythms in melatonin, methoxytryptophol, and arginine vasotocin levels and effects on the pineal of endogenous catecholamines, the estrous cycle, and environmental lighting. J Neurotrans (SUPPI) 13~59-70.
Zatz M (1978): Sensitivity and cyclic nucleotides in the rat pineal gland. J Neural Trans (Suppl) 13:97-114.




















