
Nitric oxide-evoked cGMP production in Purkinje cells in rat cerebellum: An immunocytochemical and...
8
Nitric oxide-evoked cGMP production in Purkinje cells in rat cerebellum: An immunocytochemical and pharmacological study Manuela Marcoli a , Guido Maura a,b , Chiara Cervetto a , Caterina Giacomini a , Diana Oliveri c , Simona Candiani c , Mario Pestarino c, * a Dipartimento di Medicina Sperimentale, Sezione di Farmacologia e Tossicologia, Universita ` di Genova, 16148 Genova, Italy b Centro di Eccellenza per la Ricerca Biomedica, 16132 Genova, Italy c Dipartimento di Biologia, Universita ` di Genova, Viale Benedetto XV 5, 16132 Genova, Italy Received 1 March 2006; received in revised form 18 May 2006; accepted 12 June 2006 Available online 9 August 2006 Abstract The cerebellar cells that account for glutamate-dependent cyclic GMP (cGMP) production, involving activation of the ionotropic glutamate receptors/nitric oxide synthase/soluble guanylyl cyclase pathway, are not fully established. In the present paper we have searched for the localisation of the cGMP response to the nitric oxide (NO) donor S-nitroso-penicillamine (SNAP 1 mM), expected to generate local NO concentrations in the low nanomolar physiological range and evoking a cGMP response dependent on glutamate release and on the consequent activation of ionotropic glutamate NMDA/non-NMDA receptors, in cerebellar slices from adult rat. We have found that low concentration of exogenous NO evoked cGMP accumulation in Purkinje cells in an ionotropic glutamate receptor-dependent and tetrodotoxin-sensitive manner. Such immunocytochemical localisation appears consistent with functional evidence for physiologically relevant glutamate-dependent cGMP production in Purkinje cells in rat cerebellar cortex. # 2006 Elsevier Ltd. All rights reserved. Keywords: Calbindin-D-28K; Cerebellar slices; 3 0 ,5 0 -Cyclic guanosine monophosphate; Immunocytochemistry; S-Nitroso-penicillamine; Tetrodotoxin 1. Introduction Cerebellar ionotropic glutamate receptors were recognised to be linked to cGMP production through nitric oxide synthase (NOS)/soluble guanylyl cyclase (sGC) activation (Bredt and Snyder, 1989) both in in vivo (Wood and Rao, 1991; Fedele and Raiteri, 1999) and in vitro models (Garthwaite, 1991; Maura et al., 1995). Through activation of NOS-linked NMDA/AMPA receptors and production of the diffusible messenger nitric oxide (NO), synaptic transmission onto glutamate ionotropic receptors is converted in nonsynaptic regulation of neuron communication (see Kiss and Vizi, 2001; Vizi et al., 2004): by this way, production of the unconventional messenger NO in the cerebellar cortex provides the anatomofunctional basis for glutamatergic transmission auto-regulation and for changes of synaptic communication efficiency. Although NO-mediated communication appears crucial to the glutamatergic transmis- sion regulation in the cerebellum, being NO involved in presynaptic facilitation (Jacoby et al., 2001) and postsynaptic depression (see Ito, 2001 and references therein) of glutama- tergic transmission during induction of synaptic plasticity, the sites for accumulation of cGMP dependent on activation of the cerebellar ionotropic glutamate receptors/NOS/sGC path- way(s) are not unequivocally established. Earlier functional evidence suggests that Purkinje cells might be responsible for cGMP production in the cerebellar cortex (Biggio and Guidotti, 1976; Rubin and Ferrendelli, 1977; De Vente et al., 1990), and NO-dependent cGMP production in Purkinje cells appears required for the long-term depression of excitatory transmission at parallel fibre-Purkinje cells synapses (Ito and Karachot, 1992; Daniel et al., 1993; Boxall and Garthwaite, 1996; Lev-Ram et al., 1997; Levenes et al., 1998; Hartell, 2001). Nevertheless, cGMP accumulation in Purkinje cells could not be easily detected. Purkinje cells were demonstrated to express sGC (Ariano et al., 1982; Nakane et al., 1983) and cGMP-dependent protein kinases and www.elsevier.com/locate/neuint Neurochemistry International 49 (2006) 683–690 * Corresponding author. Tel.: +39 010 353 8043; fax: +39 010 353 3062. E-mail address: [email protected] (M. Pestarino). 0197-0186/$ – see front matter # 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuint.2006.06.009
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Nitric oxide-evoked cGMP production in Purkinje cells in rat cerebellum: An immunocytochemical and...

www.elsevier.com/locate/neuint
Neurochemistry International 49 (2006) 683–690
Nitric oxide-evoked cGMP production in Purkinje cells in rat cerebellum:
An immunocytochemical and pharmacological study
Manuela Marcoli a, Guido Maura a,b, Chiara Cervetto a, Caterina Giacomini a, Diana Oliveri c,Simona Candiani c, Mario Pestarino c,*
a Dipartimento di Medicina Sperimentale, Sezione di Farmacologia e Tossicologia, Universita di Genova, 16148 Genova, Italyb Centro di Eccellenza per la Ricerca Biomedica, 16132 Genova, Italy
c Dipartimento di Biologia, Universita di Genova, Viale Benedetto XV 5, 16132 Genova, Italy
Received 1 March 2006; received in revised form 18 May 2006; accepted 12 June 2006
Available online 9 August 2006
Abstract
The cerebellar cells that account for glutamate-dependent cyclic GMP (cGMP) production, involving activation of the ionotropic glutamate
receptors/nitric oxide synthase/soluble guanylyl cyclase pathway, are not fully established. In the present paper we have searched for the
localisation of the cGMP response to the nitric oxide (NO) donor S-nitroso-penicillamine (SNAP 1 mM), expected to generate local NO
concentrations in the low nanomolar physiological range and evoking a cGMP response dependent on glutamate release and on the consequent
activation of ionotropic glutamate NMDA/non-NMDA receptors, in cerebellar slices from adult rat. We have found that low concentration of
exogenous NO evoked cGMP accumulation in Purkinje cells in an ionotropic glutamate receptor-dependent and tetrodotoxin-sensitive manner.
Such immunocytochemical localisation appears consistent with functional evidence for physiologically relevant glutamate-dependent cGMP
production in Purkinje cells in rat cerebellar cortex.
# 2006 Elsevier Ltd. All rights reserved.
Keywords: Calbindin-D-28K; Cerebellar slices; 30,50-Cyclic guanosine monophosphate; Immunocytochemistry; S-Nitroso-penicillamine; Tetrodotoxin
1. Introduction
Cerebellar ionotropic glutamate receptors were recognised
to be linked to cGMP production through nitric oxide synthase
(NOS)/soluble guanylyl cyclase (sGC) activation (Bredt and
Snyder, 1989) both in in vivo (Wood and Rao, 1991; Fedele and
Raiteri, 1999) and in vitro models (Garthwaite, 1991; Maura
et al., 1995). Through activation of NOS-linked NMDA/AMPA
receptors and production of the diffusible messenger nitric
oxide (NO), synaptic transmission onto glutamate ionotropic
receptors is converted in nonsynaptic regulation of neuron
communication (see Kiss and Vizi, 2001; Vizi et al., 2004): by
this way, production of the unconventional messenger NO in
the cerebellar cortex provides the anatomofunctional basis for
glutamatergic transmission auto-regulation and for changes of
synaptic communication efficiency. Although NO-mediated
* Corresponding author. Tel.: +39 010 353 8043; fax: +39 010 353 3062.
E-mail address: [email protected] (M. Pestarino).
0197-0186/$ – see front matter # 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.neuint.2006.06.009
communication appears crucial to the glutamatergic transmis-
sion regulation in the cerebellum, being NO involved in
presynaptic facilitation (Jacoby et al., 2001) and postsynaptic
depression (see Ito, 2001 and references therein) of glutama-
tergic transmission during induction of synaptic plasticity, the
sites for accumulation of cGMP dependent on activation of the
cerebellar ionotropic glutamate receptors/NOS/sGC path-
way(s) are not unequivocally established.
Earlier functional evidence suggests that Purkinje cells
might be responsible for cGMP production in the cerebellar
cortex (Biggio and Guidotti, 1976; Rubin and Ferrendelli,
1977; De Vente et al., 1990), and NO-dependent cGMP
production in Purkinje cells appears required for the long-term
depression of excitatory transmission at parallel fibre-Purkinje
cells synapses (Ito and Karachot, 1992; Daniel et al., 1993;
Boxall and Garthwaite, 1996; Lev-Ram et al., 1997; Levenes
et al., 1998; Hartell, 2001). Nevertheless, cGMP accumulation
in Purkinje cells could not be easily detected. Purkinje cells
were demonstrated to express sGC (Ariano et al., 1982;
Nakane et al., 1983) and cGMP-dependent protein kinases and

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690684
G-substrate (Aswad and Greengard, 1981; Nairn and Green-
gard, 1983; El-Husseini et al., 1999) as well as cGMP-specific
phosphodiesterases (Kotera et al., 1997; Giordano et al., 2001;
Okada and Shigeki, 2002). However, although it cannot be
excluded that contribution made by non-neuronal cells might
have been overemphasised over neuronal localisation
(Southam and Garthwaite, 1993), in image analysis studies
cGMP accumulation evoked by ionotropic glutamate receptor
activation or NO donors in neonatal or adult rat cerebellar
cortex was mainly observed in granules, Bergman glia or
astrocytes (De Vente et al., 1990, 1998). The failure to
demonstrate cGMP in adult cerebellar Purkinje cells is not
likely to depend on the detection limit of the antibody. In fact,
evidence on cGMP expression in tissue sections provided by
image analysis (cGMP-immunofluorescence) and comparison
with biochemical cGMP determination give comparable
results. Moreover, immunoreactivity was still visible even if
quantitation does not show significant increase (Van-Staveren
et al., 2001).
Identification of the cerebellar cells responsible for cGMP
accumulation dependent on activation of ionotropic gluta-
mate receptors/NOS/sGC pathway may allow a better
understanding of physiologically relevant glutamatergic
pathway(s) in the cerebellar cortex, as it can reflect the area
of influence of glutamate-dependent transmission extended to
nonsynaptic NO-mediated communication. We have obtained
evidence that low nanomolar exogenous NO generated by
1 mM of the NO donor S-nitroso-penicillamine (SNAP) could
substitute for endogenous NO acting as a physiological signal
in adult rat cerebellar slices, by evoking a cGMP response
dependent on release of endogenous glutamate at parallel
fibres/Purkinje cell synapses and on the consequent activation
of the cerebellar ionotropic glutamate receptors/NOS/sGC
pathway(s) (Marcoli et al., 2006). In the present paper
immunocytochemical localisation of the cGMP accumulation
in response to 1 mM SNAP was assessed in adult rat
cerebellar slices.
2. Experimental procedures
2.1. Animals
Adult male rats (Sprague–Dawley 200–250 g) were housed at constant
temperature (22 � 1 8C) and relative humidity (50%) under a regular light–dark
schedule (light 7 a.m. to 7 p.m.); food and water were freely available. The
animals were sacrificed by decapitation. Experimental procedures were
approved by the Ethical Committee of the Pharmacology and Toxicology
Section, Department of Experimental Medicine in accordance with the Eur-
opean legislation (European Communities Council Directive of 24 November
1986, 86/609/EEC). All efforts were made to minimize the number of animals
used and their suffering.
2.2. Cerebellar slices
The cerebellum was rapidly removed, placed in ice-cold medium and
chopped with a Mcllwain tissue chopper in 400 mm sagittal slices. Slices were
preincubated (90 min) in a physiological medium having the following com-
position (mmol/l): NaCl 125; KCl 3; MgSO4 1.2; CaCl2 1.2; NaH2PO4 1.0;
NaHCO3 22 and glucose 10 (gassed with 95% O2 and 5% CO2 at 37 8C); pH
7.2–7.4, with changes of the medium every 30 min. After preincubation slices
were transferred into tubes containing 2 ml standard medium or medium added
with tetrodotoxin (TTX) or glutamate receptor antagonists, saturated with 95%
O2 and 5% CO2, and incubated in a shaking water bath at 37 8C for 15 min; then,
the slices were added or not with SNAP (1 mM) with or without TTX or
glutamate receptor antagonists for 3 min, according to the protocol described by
Marcoli et al. (2006).
2.3. Imaging
Immunoreactive cGMP was localized according to De Vente et al. (1987)
and De Vente and Steinbusch (1993). After 3 min of incubation, the slices
were fixed in 4% paraformaldehyde in Tris buffered saline (pH 7.4) (TBS), at
4 8C for 120 min, then washed in TBS containing 10% sucrose at 4 8C for
90 min. After embedding in Tissue Freezing Medium (Electron Microscopy
Sciences, Fort Washington, PA), the frozen slices are cut in 20 mm-thick
sagittal sections with a cryostat Frigocut 2800E (Reichert-Jung, Germany).
The paraformaldehyde-fixed frozen sections were mounted on slides pre-
viously coated with 0.1% poly-L-lysine (Sigma, St. Louis, MO), and pro-
cessed by immunocytochemistry. Briefly, the sections were allowed to dry at
room temperature for 20–30 min, incubated overnight in a moist chamber at
4 8C with the following primary antisera: 1-anti-cGMP-formaldehyde-bovine
thyroglobulin obtained in sheep (kindly provided by De Vente, Maastricht,
The Netherlands), diluted 1:1000 in TBS containing 0.3% Triton X-100
(TBS-T); 2-anti-20O-succinyl cyclic GMP conjugated to BSA (Chemicon
International, Inc., Temecula, CA) diluted 1:1000 in TBS-T. Then, the
sections were rinsed several times in TBS and incubated for 1 h at room
temperature in a moist chamber with horseradish peroxidase-conjugated
rabbit anti-sheep IgG (Sigma, St. Louis, MO) diluted 1:200 in TBS-T. Finally,
the sections were washed in TBS, and treated for 30 min with a solution
containing 3,30-diaminobenzidine-HCl (40 mg/100 ml TBS) and 200 ml of
36% hydrogen peroxide in 100 ml of TBS. Afterwards, the sections were
dehydrated in ethanol, cleared in xylene and mounted in Eukit. Alternatively,
the secondary antiserum was FITC-conjugated rabbit anti-sheep IgG (Sigma,
St. Louis, MO), then the sections were mounted with PBS/glycerol and
observed by epifluorescence microscope.
In order to define the identity of cGMP positive cells we have performed a
double immunostaining. The sections were incubated overnight in a moist
chamber at 4 8C with both the following primary antisera diluted in TBS-T: 1-
anti-cGMP-formaldehyde-bovine thyroglobulin obtained in sheep, diluted
1:1000 (De Vente, Maastricht, The Netherlands); 2-anti-glial fibrillary acidic
protein (GFAP) obtained in rabbit diluted 1:100, or rabbit anti-calbindin-D-28K
diluted 1:500, or monoclonal antibody anti-calbindin-D-28K diluted 1:200
(Sigma, St. Louis, MO). After washing, the sections were incubated 1 h at
room temperature with both the following secondary antibodies diluted in TBS:
FITC-conjugate anti-sheep IgG developed in donkey diluted 1:200 (Sigma, St.
Louis, MO), Alexa Fluor1 546 goat anti-mouse IgG diluted 1:1000 (Molecular
Probes Europe BV, Leiden, The Netherlands) or tetramethylrhodamine iso-
thiocyanate-conjugate anti-rabbit IgG developed in goat diluted 1:200 (Sigma,
USA). After three washes in TBS, 15 min each, the slides were coverslipped
with glycerol:TBS (v/v, 1:9). Images were collected by using an Olympus IX71
microscope (Olympus Italia s.r.l., Italy) equipped with a Soft Imaging System
chilled colour digital camera ColorView II (Soft Imaging System GmbH,
Germany) and images were processed using the analySIS software package
(Soft Imaging System GmbH, Germany).
In order to check the specificity of the immunoreaction, the following
controls were performed: (1) omitting the primary or secondary antibodies, (2)
using preimmune serum from which we had obtained the primary antibody,
diluted 1:30 in TBS and (3) blocking of endogenous peroxidases by 3% H2O2.
2.4. cGMP immunoassay
After 3-min incubation the slices were transferred into 1 ml of Tris–HCl
(50 mM; pH 7.5 containing 4 mM EDTA, heated at 100 8C), then maintained at
100 8C for 10 min. The slices were homogenised by sonication and centrifuged
for 5 min at 5000 � g. The cGMP content was determined in l00-ml aliquots of
the supernatant using a commercially available radioimmunoassay kit (Amer-
sham Radiochemical Centre, Buckinghamshire, UK), as described in Marcoli
et al. (2006). The sensitivity of the assay was about 0.04 pmol. The levels of

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690 685
cGMP were expressed as pmol/mg protein. The cGMP response was calculated
by subtraction of the cGMP present in the controls from that present in the
sample containing the drugs tested. The effect of drugs was expressed as percent
variation with respect to the appropriate controls. Mean � S.E.M. of the
numbers of experiments (n) are indicated throughout. Significance of the
difference between means was analyzed by Anova followed by the Student’s
t-test; the level of significance was set at p < 0.05.
Protein determination was carried out according to Bradford (1976) using
bovine serum albumin as standard.
2.5. Materials
The following drugs were purchased: 6-cyano-7-nitroquinoxaline-2,3-dione
(CNQX); dizocilpine (MK 801); S-nitroso-penicillamine (SNAP) and tetrodo-
toxin (TTX) from Tocris Cookson (Bristol, UK). SNAP solutions were freshly
prepared 60 min before use and kept in dark tightly sealed at room temperature.
Specificity of the response to SNAP was checked by assessing lack of any
cGMP response to 1 mM SNAP after leaving 24-h opened the SNAP solution. In
these conditions SNAP is expected to be decomposed and to have lost its ability
to release NO.
Fig. 1. cGMP immunoreactivity in coronal frozen sections of rat cerebellum. Pu
treatment with SNAP (1 mM) (B). Additional treatment with TTX (0.5 mM) results
small blood vessels (C). Similarly, additional treatment with the NMDA receptor an
impaired the cGMP response to SNAP (D). Basal level of cGMP are shown in (A)
molecular layers. Scale bar = 100 mm.
3. Results
3.1. Immunocytochemistry: localisation of the cGMP
response to SNAP (1 mM), its TTX-sensitivity and
dependence on ionotropic glutamate receptor activation
Cyclic GMP immunostaining was obtained exclusively using
the antiserum against formaldehyde-fixed cGMP (De Vente
et al., 1987). In frozen cerebellar sections the basal cGMP
immunostaining intensity was low, even if few cGMP positive
elements were visible as well as positive blood vessel walls
(Fig. 1A). Intense cGMP immunostaining appeared in Purkinje
cell bodies and dendrites after 1 mM SNAP treatment (Figs. 1B,
2A and B). Inhibition of SNAP-evoked cGMP accumulation was
observed when ionotropic glutamate NMDA/AMPA receptors
were blocked by the NMDA receptor antagonist MK 801
(10 mM) plus the AMPA receptor antagonist CNQX (10 mM), or
rkinje cell bodies and dendrites show a strong cGMP immunopositivity after
in a loss of cGMP immunoreactivity, the only visible signals are associated to
tagonist MK801 (10 mM) plus the AMPA receptor antagonist CNQX (10 mM)
, where few positive Purkinje cells (arrowheads) are visible. g, granule and m,

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690686
Fig. 2. cGMP immunoreactivity in Purkinje cell bodies and dendrites after treatment with SNAP (1 mM). Using the indirect immunoperoxidase technique, cGMP
immunostaining is shown at level of bodies ((A) and (B) arrowheads) and dendrites ((B) arrows) of Purkinje cells. g, granule and m, molecular layers. Basal levels of
cGMP are shown in (C). Scale bar = 100 mm.
in the presence of TTX (0.5 mM) (Fig. 1C and D). The double
immunostaining showed that GFAP immunoreactivity was
localized in several glial cells of the granular layer, but not in
the cGMP-positive Purkinje cells. In particular, GFAP was found
in some large cellular bodies of the Bergman glia and in other
glial cell types widespread in the granule layer (Fig. 3A and B).
The cell processes of the Bergman glia projected right across the
molecular layer and reached the outer surface of the cerebellum
cortex, where they form conical end feet (Fig. 3A). Overlay of
cGMP and calbindin-D-28K (a Purkinje cell marker according to
Jande et al., 1981; Cesa et al., 2005) showed co-localisation of
cGMP and calbindin-D-28K in the Purkinje cells (Fig. 3C–E).
Decomposed SNAP (1 mM solution was left opened 24-h)
was unable to evoke any cGMP accumulation in Purkinje cells
(data not shown). No cGMP immunoreactivity was observed in
cerebellum frozen slices using the primary or secondary
antisera. The cGMP antiserum showed no cross-reactivity with
other nucleotides and nucleosides.
3.2. Immunoassay: TTX-sensitivity of the SNAP (1 mM)-
evoked cGMP production
In our conditions cGMP production in control slices
amounted to 8.70 � 0.81 pmol/mg protein/3 min (mean
� S.E.M., n = 5). SNAP (1 mM) increased cGMP level by
79.4 � 5.1% (mean � S.E.M., n = 5). TTX (0.5 mM) per se did
not modify basal cGMP (8.09 � 0.49 pmol/mg protein/3 min;
mean � S.E.M., n = 5) while it prevented the cGMP response
to 1 mM SNAP (Fig. 4).
Decomposed SNAP (1 mM solution was left opened 24-h)
did not evoke any cGMP response in rat cerebellar slices (data
not shown).
4. Discussion
The main result of the present study is that in adult rat
cerebellar slices the Purkinje cells appear responsible for cGMP
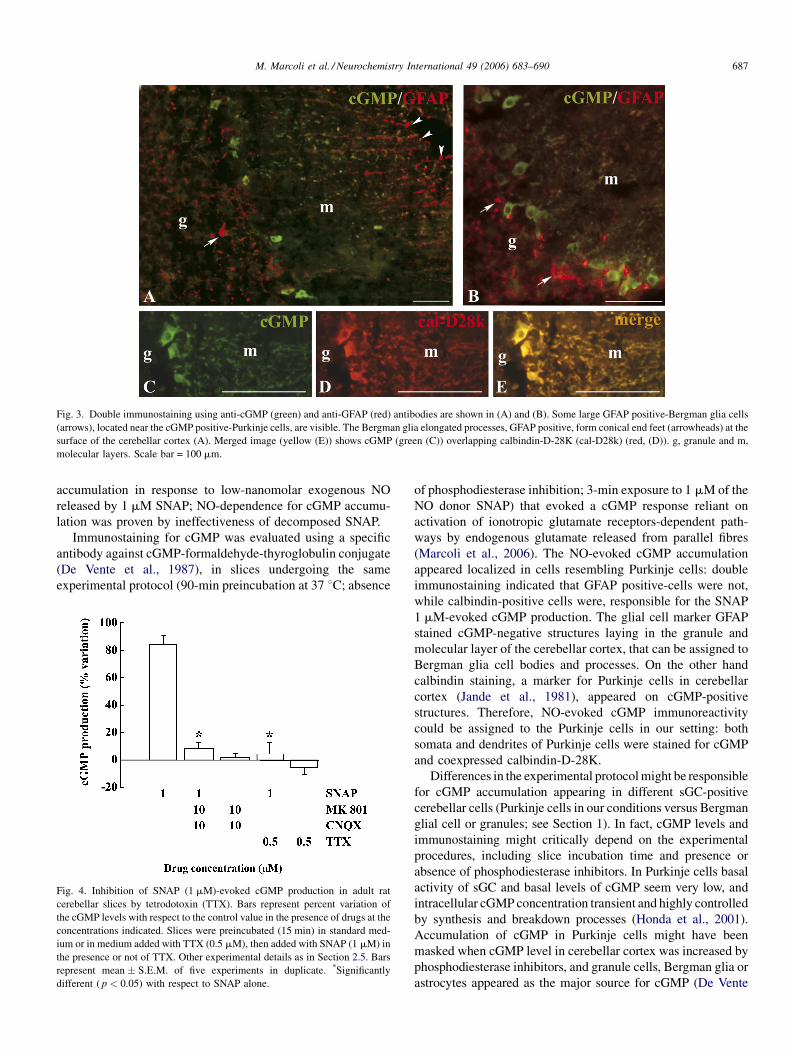
M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690 687
Fig. 3. Double immunostaining using anti-cGMP (green) and anti-GFAP (red) antibodies are shown in (A) and (B). Some large GFAP positive-Bergman glia cells
(arrows), located near the cGMP positive-Purkinje cells, are visible. The Bergman glia elongated processes, GFAP positive, form conical end feet (arrowheads) at the
surface of the cerebellar cortex (A). Merged image (yellow (E)) shows cGMP (green (C)) overlapping calbindin-D-28K (cal-D28k) (red, (D)). g, granule and m,
molecular layers. Scale bar = 100 mm.
accumulation in response to low-nanomolar exogenous NO
released by 1 mM SNAP; NO-dependence for cGMP accumu-
lation was proven by ineffectiveness of decomposed SNAP.
Immunostaining for cGMP was evaluated using a specific
antibody against cGMP-formaldehyde-thyroglobulin conjugate
(De Vente et al., 1987), in slices undergoing the same
experimental protocol (90-min preincubation at 37 8C; absence
Fig. 4. Inhibition of SNAP (1 mM)-evoked cGMP production in adult rat
cerebellar slices by tetrodotoxin (TTX). Bars represent percent variation of
the cGMP levels with respect to the control value in the presence of drugs at the
concentrations indicated. Slices were preincubated (15 min) in standard med-
ium or in medium added with TTX (0.5 mM), then added with SNAP (1 mM) in
the presence or not of TTX. Other experimental details as in Section 2.5. Bars
represent mean � S.E.M. of five experiments in duplicate. *Significantly
different ( p < 0.05) with respect to SNAP alone.
of phosphodiesterase inhibition; 3-min exposure to 1 mM of the
NO donor SNAP) that evoked a cGMP response reliant on
activation of ionotropic glutamate receptors-dependent path-
ways by endogenous glutamate released from parallel fibres
(Marcoli et al., 2006). The NO-evoked cGMP accumulation
appeared localized in cells resembling Purkinje cells: double
immunostaining indicated that GFAP positive-cells were not,
while calbindin-positive cells were, responsible for the SNAP
1 mM-evoked cGMP production. The glial cell marker GFAP
stained cGMP-negative structures laying in the granule and
molecular layer of the cerebellar cortex, that can be assigned to
Bergman glia cell bodies and processes. On the other hand
calbindin staining, a marker for Purkinje cells in cerebellar
cortex (Jande et al., 1981), appeared on cGMP-positive
structures. Therefore, NO-evoked cGMP immunoreactivity
could be assigned to the Purkinje cells in our setting: both
somata and dendrites of Purkinje cells were stained for cGMP
and coexpressed calbindin-D-28K.
Differences in the experimental protocol might be responsible
for cGMP accumulation appearing in different sGC-positive
cerebellar cells (Purkinje cells in our conditions versus Bergman
glial cell or granules; see Section 1). In fact, cGMP levels and
immunostaining might critically depend on the experimental
procedures, including slice incubation time and presence or
absence of phosphodiesterase inhibitors. In Purkinje cells basal
activity of sGC and basal levels of cGMP seem very low, and
intracellular cGMP concentration transient and highly controlled
by synthesis and breakdown processes (Honda et al., 2001).
Accumulation of cGMP in Purkinje cells might have been
masked when cGMP level in cerebellar cortex was increased by
phosphodiesterase inhibitors, and granule cells, Bergman glia or
astrocytes appeared as the major source for cGMP (De Vente

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690688
et al., 1990, 1998). Furthermore, especially in the presence of
phosphodiesterase inhibitors, cGMP accumulation might also
depend on sGC desensitization. Indeed, it appears that
distribution of sGC activity and cGMP production observed in
the presence of phosphodiesterase inhibitors, might poorly
describe the cGMP profile in the absence of phosphodiesterase
inhibitors (Southam and Garthwaite, 1993; Mullershausen et al.,
2001; Wykes et al., 2002). In any case, although cGMP
accumulation had never been directly demonstrated in adult
cerebellar Purkinje cells, indication that cGMP may be produced
in such cells was obtained using exogenous or endogenously
generated NO. NO-dependent activation of sGC in cultured
Purkinje cells was observed using an antibody sensing sGC
activation (Tsuyama et al., 1999). Furthermore, activation of
type-5 phosphodiesterase or phosphorylation of G-substrate by
cGMP-dependent protein kinase suggest that cGMP production
in Purkinje cells could be evoked by exogenous NO, or by
endogenous NO released by parallel fibre activation, in rat
cerebellar slices (Hartell et al., 2001; Endo et al., 2003) or in vivo
in the mouse (Shimizu-Albergine et al., 2003). Investigation on
temporal dynamics of cGMP with fluorescent indicator in single
Purkinje cells in organotypic cultures had shown that Purkinje
cells rapidly responded with cGMP transients to NO-donors or to
parallel fibres stimulation in conditions inducing long-term
potentiation of the parallel fibre-Purkinje cell synapse (Honda
et al., 2001).
In our experimental protocol, the low NO donor concentration
is likely to be a critical point accounting for cGMP localisation in
Purkinje cells. Indeed, it is conceivable that the cerebellar
powerful inactivation mechanism(s) shaping NO signals in the
low nanomolar range allow cerebellar cells to maintain NO
concentrations compatible with physiological roles, when
exposed to low NO donor concentrations (Griffiths and
Garthwaite, 2001). Low nanomolar exogenous NO concentra-
tions generated by 1 mM SNAP (Ishida et al., 1996; Megson
et al., 1997, 1999) indeed appeared to substitute for endogen-
ously produced NO acting as a physiological signal in facilitating
glutamate release onto ionotropic receptors linked to cGMP
production (Marcoli et al., 2006). In fact, the finding that 1 mM
SNAP released endogenous glutamate from parallel fibres
(Marcoli et al., 2006) in a glutamate receptor-dependent manner,
is consistent with the electrophysiological evidence for the NO
role in facilitating glutamate release at parallel/climbing fibre/
Purkinje cells synapses during induction of long-term potentia-
tion (Jacoby et al., 2001). As a matter of fact, NO direct releasing
effect at parallel fibre endings and activation of granules/parallel
fibres, related to NO-dependent potentiation of glutamate release
from mossy fibre endings (see Maffei et al., 2003) might co-
operate to evoke glutamate release from parallel fibres. We here
find that in the presence of TTX the cGMP production evoked by
1 mM SNAP was almost completely abolished and no cGMP
accumulated in Purkinje cells. TTX-sensitivity of the NO-
evoked cGMP accumulation in Purkinje cells, suggesting action
potential-dependent activation of the glutamatergic pathway
depending on neuronal firing, fits in well with mechanisms
operating during induction of synaptic plasticity. We cannot
however exclude that TTX-sensitive depolarization (see
Ohkuma et al., 1998), could also play a direct role in glutamate
release and consequent cGMP production by 1 mM SNAP, or
interfere with ionotropic glutamate receptor activation on
parallel fibres themselves and/or on intrinsic neurons involved
in the pathway.
The cGMP production in response to 1 mM SNAP, almost
matching the response to NMDA/AMPA receptor agonists
(Maura et al., 1995), appeared dependent on the activation of
cGMP-linked glutamate ionotropic receptors by endogenously
released glutamate (Marcoli et al., 2006). Fitting in with
quantitative determinations, we here obtain immunocytochem-
ical evidence for the inability of 1 mM SNAP to cause cGMP
accumulation in Purkinje cells when NMDA/AMPA receptors
were blocked by MK 801 plus CNQX. We can hypothesise that
the NO-depended glutamate release from parallel fibres might
activate endogenous NO production by acting upon NMDA/
AMPA receptors on NOS-positive inhibitory basket or stellate
cells (Garthwaite and Beaumont, 1989; Southam and
Garthwaite, 1993; Clark and Cull-Candy, 2002). Diffusion of
NO from basket/stellate cells might then evoke cGMP
production in neighbouring Purkinje cells somata or dendrites:
in fact, ionotropic glutamate receptor-dependent NO cascade
involved in cerebellar long term depression has been suggested
to be localized in interneurons (Shin and Linden, 2005). On the
other hand, glutamate released from parallel fibres might evoke
a cGMP response in Purkinje cells dendrites by activating
NMDA or AMPA-dependent NOS in parallel fibres themselves.
Indeed, frequency-dependent NO generation in parallel fibres
terminals during synaptic plasticity (Shibuki and Kimura, 1997;
Kimura et al., 1998; Casado et al., 2000), involving presynaptic
NMDA/AMPA receptor activation by released glutamate
(Gorbunov and Esposito, 1994; Casado et al., 2000, 2002;
Okada et al., 2004), might also be linked to postsynaptic cGMP
production in Purkinje cells.
A point often ignored when discussing the cerebellar NO
targets remains to be raised: visualisation of cGMP in parallel
fibres is difficult and depends on extreme section thinness and
on the fibre direction (as shown by De Vente et al., 1998).
Therefore, we cannot exclude that in our conditions cGMP
production in parallel fibre terminals might possibly contribute
to the immunopositivity in Purkinje cell dendrites.
In conclusion, in adult rat cerebellar slices 1 mM of the NO
donor SNAP induced cGMP production in Purkinje cells,
dependent on NMDA/AMPA receptor activation, and TTX-
sensitive. Our findings suggest that low-level exogenous NO in
cerebellar slices could activate cerebellar glutamatergic path-
way(s) onto Purkinje cells, by facilitating activity-dependent
glutamate release from parallel fibres.
Acknowledgements
The authors are grateful to Dr. J. De Vente (Department of
Psychiatry and Neuropsychology, European School of Neu-
roscience, Universiteit Maastricht, 6200 MD Maastricht, The
Netherlands) for generous gift of the antiserum anti-cGMP-
formaldehyde-bovine thyroglobulin. This work was supported
by an Italian MIUR Network grant.

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690 689
References
Ariano, M.A., Lewicki, J.A., Brandwein, H.J., 1982. Immunohistochemical
localization of guanylate cyclase within neurons of rat brain. Proc. Natl.
Acad. Sci. U.S.A. 79, 1216–1320.
Aswad, D.W., Greengard, P., 1981. A specific substrate from rabbit cerebellum
for guanosine 30, 50-monophosphate-dependent protein kinase. I. Purifica-
tion and characterization. J. Biol. Chem. 256, 3487–3493.
Biggio, G., Guidotti, A., 1976. Climbing fiber activation and 30,50-cyclic
guanosine monophosphate (cGMP) content in cortex and deep nucleai of
cerebellum. Brain Res. 107, 577–581.
Boxall, A.R., Garthwaite, J., 1996. Long-term depression in rat cerebellum
requires both NO synthase and NO-sensitive guanylyl cyclase. Eur. J.
Neurosci. 8, 2209–2212.
Bradford, M.M., 1976. Rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye
binding. Anal. Biochem. 72, 246.
Bredt, D.S., Snyder, S.H., 1989. Nitric oxide mediates glutamate-linked
enhancement of cGMP levels in the cerebellum. Proc. Natl. Acad. Sci.
U.S.A. 86, 9030–9033.
Casado, M., Dieudonne, S., Ascher, P., 2000. Presynaptic N-methyl-D-aspartate
receptors at the parallel fibre-Purkinje cell synapse. Proc. Natl. Acad. Sci.
U.S.A. 97, 11593–11597.
Casado, M., Isope, P., Ascher, P., 2002. Involvement of presynaptic N-methyl-D-
aspartate receptors in cerebellar long-term depression. Neuron 33, 123–
130.
Cesa, R., Morando, L., Strata, P., 2005. Purkinje cell spinogenesis during
architectural rewiring in the mature cerebellum. Eur. J. Neurosci. 22, 579–
586.
Clark, B.A., Cull-Candy, S.G., 2002. Activity-dependent recruitment of extra-
synaptic NMDA receptor activation at an AMPA receptor-only synapse. J.
Neurosci. 22, 4428–4436.
Daniel, H., Hemart, N., Jaillard, D., Crepel, F., 1993. Long-term depression
requires nitric oxide and guanosine 30-50 cyclic monophosphate production
in cerebellar Purkinje cells. Eur. J. Neurosci. 5, 1079–1082.
De Vente, J., Steinbusch, H.W.M., Schipper, J., 1987. A new approach to
immunocytochemistry of 30,50-cyclic guanosine monophosphate: prepara-
tion, specificity, and initial application of a new antiserum against for-
maldehyde-fixed 30, 50-cyclic guanosine monophosphate. Neuroscience 22,
361–373.
De Vente, J., Bol, J.G.J.M., Berkelmans, H.S., Schipper, J., Steinbusch, H.M.W.,
1990. Immunocytochemistry of cGMP in the cerebellum of the immature,
adult, and aged rat: the involvement of nitric oxide. A micropharmacolo-
gical study. Eur. J. Neurosci. 2, 845–862.
De Vente, J., Steinbusch, H.W.M., 1993. Immunocytochemistry of second
messenger molecules: the study of formaldehyde–fixed cyclic GMP. In:
Cuello, A.C. (Ed.), Immunohystochemistry II. John Wiley & Sons,
Chichester, pp. 409–427.
De Vente, J., Hopkins, D.A., Markerink-Van Ittersum, M., Emson, P.C.,
Schmidt, H.H.H.W., Steinbusch, H.W.M., 1998. Distribution of nitric oxide
synthase and nitric oxide-receptive, cyclic GMP-producing structures in the
rat brain. Neuroscience 87, 207–241.
El-Husseini, A.E.D., Williams, J., Reiner, P.B., Pelech, S., Vincent, S.R., 1999.
Localization of the cGMP-dependent protein kinases in relation to nitric
oxide synthase in the brain. J. Chem. Neuroanat. 17, 45–55.
Endo, S., Nairn, A.C., Greengard, P., Ito, M., 2003. Thr123 of rat G-substrate
contributes to its action as a protein phosphatase inhibitor. Neurosci. Res.
45, 79–89.
Fedele, E., Raiteri, M., 1999. In vivo studies of the cerebral glutamate receptor/
NO/cGMP pathway. Prog. Neurobiol. 58, 89–120.
Garthwaite, J., 1991. Glutamate, nitric oxide and cell–cell signalling in the
nervous system. Trends Neurosci. 14, 60–67.
Garthwaite, J., Beaumont, P.S., 1989. Excitatory amino acid receptors in the
parallel fibre pathway in rat cerebellar slices. Neurosi. Lett. 107, 151–156.
Giordano, D., De-Stefano, M.E., Citro, G., Modica, A., Giorni, M., 2001.
Expression of cGMP-binding cGMP-specific phosphodiesterase (PDE5) in
mouse tissues and cell lines using an antibody against the enzyme amino-
terminal domain. Biochim. Biophys. Acta 1539, 16–27.
Gorbunov, N., Esposito, E., 1994. Activation of glutamate receptors stimulates
the formation of nitrite in synaptosomes from rat cerebellum. J. Neurochem.
62, 2205–2211.
Griffiths, C., Garthwaite, J., 2001. The shaping of nitric oxide signals by a
cellular sink. J. Physiol. Lond. 536, 855–862.
Hartell, N.A., 2001. Receptors, second messengers and protein kinases required
for heterosynaptic cerebellar long-term depression. Neuropharmacology 40,
148–161.
Hartell, N.A., Furuka, S., Jacoby, S., Okada, D., 2001. Intercellular action of
nitric oxide increases cGMP in cerebellar Purkinje cells. Neuroreport 12,
25–28.
Honda, A., Adams, S.R., Sawyer, C.L., Lev-Ram, V., Tsien, R.Y., Dostmann,
W.R.G., 2001. Spatiotemporal dynamics of guanosine 30, 50-cyclic mono-
phosphate revealed by a genetically encoded, fluorescent indicator. Proc.
Natl. Acad. Sci. U.S.A. 98, 2437–2442.
Ishida, Y., Hashimoto, M., Fukushima, S., Masumura, S., Sasaki, T., Nakayama,
K., Tamura, K., Murakami, E., Isokawa, S., Momose, K., 1996. A nitric
oxide-sensitive electrode: requirement of lower oxygen concentration for
detecting nitric oxide from the tissue. J. Pharm. Tox. Meth. 35, 19–24.
Ito, M., 2001. Cerebellar long-term depression: characterization, signal trans-
duction, and functional roles. Physiol. Rev. 81, 1143–1195.
Ito, M., Karachot, L., 1992. Protein kinases and phospahatase inhibitors
mediating long-term desensitization of glutamate receptors in cerebellar
Purkinje cells. Neurosci. Res. 14, 27–38.
Jacoby, S., Sims, R.E., Hartell, N.A., 2001. Nitric oxide is required for the
induction and heterosynaptic spread of long-term potentiation in rat cer-
ebellar slices. J. Physiol. Lond. 535, 825–839.
Jande, S.S., Maler, L., Lawson, D.E.M., 1981. Immunohistochemical mapping
of vitamin D-dependent calcium-binding protein in brain. Nature 294, 765–
767.
Kimura, S., Uchiyama, S., Takahashi, H.E., Shibuki, K., 1998. cAMP-depen-
dent long-term potentiation of nitric oxide release from cerebellar parallel
fibres in rats. J. Neurosci. 18, 8551–8558.
Kiss, J.P., Vizi, E.S., 2001. Nitric oxide: a novel link between synaptic and
nonsynaptic transmission. Trends Neurosci. 24, 211–215.
Kotera, J., Yanaka, N., Fujishige, K., Imai, Y., Akatsuka, H., Ishizuka, T.,
Kawashima, K., Omori, K., 1997. Expression of rat cGMP-binding cGMP-
specific phosphodiesterase mRNA in Purkinje cell layers during postnatal
neural development. Eur. J. Biochem. 249, 434–442.
Lev-Ram, V., Jiang, T., Wood, J., Lawrence, D.S., Tsien, R.Y., 1997. Synergies
and coincidence requirements between NO, cGMP, and Ca2+ in the induc-
tion of cerebellar long-term depression. Neuron 18, 1025–1038.
Levenes, C., Daniel, H., Crepel, F., 1998. Long-term depression of synaptic
transmission in the cerebellum: cellular and molecular mechanisms revis-
ited. Prog. Neurobiol. 55, 79–91.
Maffei, A., Prestori, F., Shibuki, K., Rossi, P., Taglietti, V., D’Angelo, E., 2003.
NO enhances presynaptic currents during cerebellar mossy fiber-granule
cell LTP. J. Neurophysiol. 90, 2478–2483.
Marcoli, M., Cervetto, C., Paluzzi, P., Guarnieri, S., Raiteri, M., Maura, G.,
2006. Nitric oxide-evoked glutamate release and cGMP production in
cerebellar slices: control by presynaptic 5-HT1D receptors. Neurochem.
Int. 49, 12–19.
Maura, G., Guadagnin, A., Raiteri, M., 1995. Low nanomolar serotonin inhibits
the glutamate receptor/nitric oxide/cyclic GMP pathway in slices from adult
rat cerebellum. Neuroscience 68, 455–463.
Megson, I.L., Greig, I.R., Gray, G.A., Webb, D.J., Butler, A.R., 1997. Prolonged
effect of a novel S-nitrosated glyco-amino acid in endothelium-denuded rat
femoral arteries: potential as a low release nitric oxide donor drug. Br. J.
Pharmacol. 122, 1617–1624.
Megson, I.L., Morton, S., Greig, I.R., Mazzei, F.A., Field, R.A., Butler, A.R.,
Caron, G., Gasco, A., Fruttero, R., Webb, D.J., 1999. N-substituted analo-
gues of S-nitroso-N-acetyl-D, L-penicillamine: chemical stability and pro-
longed nitric oxide mediated vasodilatation in isolated rat femoral arteries.
Br. J. Pharmacol. 126, 639–648.
Mullershausen, F., Russwurm, M., Thompson, W.J., Liu, L., Koeslng, D.,
Friede, A., 2001. Rapid nitric oxide-induced desensitization of the cGMP
response is accused by increased activity of phopshodiesterase type 5
paralleled by phosphorilation of the enzyme. J. Cell. Biol. 155, 271–278.

M. Marcoli et al. / Neurochemistry International 49 (2006) 683–690690
Nakane, M., Ichikawa, M., Deguchi, T., 1983. Light and electron microscopic
demonstration of guanylate cyclase in rat brain. Brain Res. 273, 9–15.
Nairn, A., Greengard, P., 1983. Ciclyc GMP-dependent protein phosphorylation
in mammalian brain. Fed. Proc. 42, 3107–3113.
Ohkuma, S., Katsura, M., Hibino, Y., Xu, J., Shirotani, K., Kuriyama, K., 1998.
Multiple actions of nitric oxide on voltage-dependent Ca2+ channels in
mouse cerebral cortical neurons. Brain Res. Mol. Brain Res. 54, 133–140.
Okada, D., Shigeki, A., 2002. Allosteric activation of cGMP-specific, cGMP-
binding phosphodiesterase (PDE5) by cGMP. Biochemistry 41, 9672–9679.
Okada, D., Yap, C.C., Kojima, H., Kikuchi, K., Nagano, T., 2004. Distinct
glutamate receptors govern differential levels of nitric oxide production in
a layer-specific manner in the rat cerebellar cortex. Neuroscience 125,
461–472.
Rubin, E.H., Ferrendelli, J.A., 1977. Distribution and regulation of cyclic
nucleotide levels in cerebellum, in vivo. J. Neurochem. 29, 43–51.
Shibuki, K., Kimura, S., 1997. Dynamic properties of nitric oxide release from
parallel fibres in rat cerebellar slices. J. Physiol. Lond. 498, 443–852.
Shimizu-Albergine, M., Rybalkin, S.D., Rybalkina, I.G., Feil, R., Wolfsgruber,
W., Hofmann, F., Beavo, J.A., 2003. Individual cerebellar Purkinje cells
express different cGMP phosphodiesterases (PDEs): in vivo phosphoryla-
tion of cGMP-specific PDE (PDE5) as an indicator of cGMP-dependent
protein kinase (PKG) activation. J. Neurosci. 23, 6452–6459.
Shin, J.H., Linden, D.J., 2005. An NMDA receptor/nitric oxide cascade is
involved in cerebellar LTD but is not localized to the parallel fibre terminal.
J. Neurophysiol. 946, 4281–4289.
Southam, E., Garthwaite, J., 1993. The nitric oxide–cyclic GMP signalling
pathway in rat brain. Neuropharmacology 32, 1267–1277.
Tsuyama, S., Yamazaki, E., Tomita, T., Ihara, H., Takenaka, S., Kato, K.,
Kozaki, S., 1999. Characterization of a novel monoclonal antibody that
sense nitric oxide-dependent activation of soluble guanylate cyclase. FEBS
Lett. 455, 291–294.
Van-Staveren, W.C., Markerink-van-Ittersum, Steinbusch, H.W., de Vente, J.,
2001. The effects of phosphodiesterase inhibition on cyclic cGMP and
ciclyc AMP accumulation in the hyppocampus of the rat. Brain. Res. 12,
275–286.
Vizi, E.S., Kiss, J.P., Lendvai, B., 2004. Nonsynaptic communication in the
central nervous system. Neurochem. Int. 45, 443–451.
Wood, P.L., Rao, T.S., 1991. A review of in vivo modulation of cerebellar cGMP
levels by excitatory amino acid receptors: role of NMDA, quisqualate
and 1kainate subtypes. Prog. Neuropsychopharmacol. Biol. Psychiatry
15, 229–235.
Wykes, V., Bellamy, T.C., Garthwaite, J., 2002. Kinetics of nitric oxide-cyclic
GMP signalling in CNS cells and its possible regulation by cyclic GMP. J.
Neurochem. 83, 37–47.




















