
Trigeminal ganglion innervation of the cochlea—a retrograde transport study
Upload
independentCategory
view
0download
0
"HE JOURNAL OF COMPARATIVE NEUROLOGY 241:E-33 (1985)
Catecholaminergic Innervation of the Septa1 Area in Man: Immunocytochemical
Study Using TH and DBH Antibodies
P. GASPAR, B. BERGER, C. ALVAREZ, A. VIGNY, AND J.P. HENRY INSERM U 134-Laboratoire de Neuropathologie Charles Foix, Hijpital SalpBtriere 75651 Paris Cedex 13 (P.G., B.B., C.A.) and Institut de Biologie Physico-Chimique,
75005 Paris (A.V., J.P.H.)
ABSTRACT The catecholaminergic innervation of the human septal area and closely
related structures has been visualized by using tyrosine hydroxylase (TH) and dopamine-P-hydroxylase (DBH) as immunocytochemical markers. TH- like immunoreactivity with no corresponding DBH labelling was considered to be indicative of dopaminergic fibers.
Catecholaminergic innervation offered the following similarities to that of rodents: (1) moderate innervation in the medial septal division, with predominant DBH immunolabelling; (2) dense dopaminergic innervation in the lateral septal nuclei, organized in a laminar pattern; (3) presence of dopaminergic pericellular arrangements in the dorsal septum and bed n. of the stria terminalis; (4) clustering of dopaminergic terminals in n. accum- bens associated with a medioventral zone of DBH-like immunoreactive fi- bers; (5) close overlap between dopaminergic fields and acetylcholinesterase- reactive zones in both the lateral septum and the n. of the stria terminalis.
Differences with the catecholaminergic septal innervation of rodents consisted of (1) general caudal extension of the dopaminergic fields, possibly accounted for by the vertical stretching and caudal displacement of the septal nuclei in man; (2) complementary lateromedial topography of dopa- minergic and DBH-immunoreactive inputs in the n. of the stria terminalis as opposed to their dorsoventral organization in rodents; (3) presence of TH- immunolabelled cell group in the anterior olfactory nucleus and parolfactory cortex, which seems specific for primates.
Precise topographical mapping of the catecholaminergic structures in this central region of the limbic forebrain seems to be a prerequisite for accurate tissue sampling in the biochemical investigations of pathoIogical cases and should help in the interpretation of aminergic dysfunction in a variety of human diseases.
Key words: basal forebrain, accumbens, olfactory tubercle, acetylcholinesterase
Contrary to what had been assumed previously, the sep- ta] nuclei appear to be well developed in man. Indeed, although they occupy a relatively small volume in the primate telencephalon, both their absolute size and ratio to body weight increase regularly in the ascent of the phylo- genetic scale (Andy and Stephan, '68, '76). Furthermore, they seem to be important in human pathophysiology since neuropsychiatric disturbances have been reported in pa- tients with vascular or tumoral lesions involving this area (Taren, '65; Lindquist and NorlBn, '66; Eliott, '69; Laine and Blond, '80) and since early damage of the cholinergic septohippocampal pathway has been demonstrated in Alz-
0 1985 ALAN R. LISS. INC.
heimer's disease (Henke and Lang, '83; Arendt et al., '83). Nevertheless, studies of the septal nuclei in man have been scarce and almost exclusively limited to cytoarchitectonic analyses.
This stands in contrast to the numerous experimental studies, particularly in rodents, which have established the central position of the septal nuclei in the limbic system. The physiological importance of this area is further sug- gested by the precise and complex topographical organiza- tion of the afferent and efferent systems demonstrated in
Accepted June 17,1985.

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 13
rodents by both hodological data (Meibach and Siegel, '77; Swanson and Cowan, '79) and the distribution of aminergic and peptidergic inputs (Gall and Moore, '84; Kohler and Erikson, '84). It would appear of interest to investigate in man whether the different neurochemically defined inputs can also be identxied, and whether they respond to similar patterns of organization. Investigation of the catecholami- nergic innervation was studied first since it has been well characterized in the rodent septum as concerns the topog- raphy of both its noradrenergic (NA) and dopaminergic (DA) components, which predominate in the medial and lateral septum, respectively. This DA innervation is further characterized by the constitution of basketlike pericellular arrangements (Lindvall and Stenevi, '78; Moore, '78), form- ing direct synaptic contacts with perikarya and dendrites (Berger et al., '83; Onteniente et al., '84) and suggesting a peculiar action of DA on specific neuronal targets.
Finally, the DA septal innervation appears to be of partic- ular importance as it is one of the components of the meso- limbocortical DA system (Lindvall and Bjorklund '83) which has been implicated in the pathogenesis of schizophrenia (Stevens, '79) and of some symptoms observed in Parkin- son's disease (Price et al., '78; Javoy-Agid et al., '83).
In man, precise knowledge of the organization of the DA and NA innervations in the forebrain is generally lacking, since only limited biochemical data (McGeer and McGeer, '76; Farley and Hornykiewicz, '77; Nyberg et al., '82) or histochemical observations in fetuses (Nobin and Bjork- lund, '73) are available. On the other hand, direct compari- son with data obtained in rodents is not feasible because the configuration of this area is modified in the phyloge- netic scale and systematic reports on catecholaminergic innervation in primates are lacking.
In the present study, we have analyzed the catecholami- nergic innervation of the adult human septal nuclei, by using the synthesizing enzymes, tyrosine hydroxylase (TH) and dopamine-0-hydroxylase (DBH), as immunocytoehemi- cal markers. The distribution of the fiber tracts and axon terminals was established in cytoarchitectonically and his- tochemically defined nuclear areas. Comparison with the comprehensive results obtained in rodents disclosed both similarities and discrepancies in the organization of this aminergic innervation.
MATERIALS AND METHODS
The brains used in this study were obtained at autopsy from 15 patients deceased in general hospitals, with no known neurological or psychiatric illness, except in one case with familial myotonic dystonia but without evidence of central cerebral impairment. No neuropathological le- sions were observed, either at the gross macroscopic level or on histological examination of the forebrain.
Cyto-myelo-architectonic study was performed on four cases (one male, three females, age range 47-72 mean 63 years). Following formalin fmation, the whole septal area, between the mamillary bodies and the olfactory crus, was sampled as one large block and embedded in paraffin. Se- rial frontal and horizontal sections (10 or 20 pm thick ac- cording to the series) were retained at 500-pm intervals, stained with thionine and lux01 fast blue.
Prominent cyto-myelo-architectonic boundaries were charted (x-y plotter) in order to obtain a complete set of topographical maps of the septal area in the coronal plane. The diagrams presented in this work were selected from one representative case (male patient 47 years, paraffin block sections 20 pm thick).
Immunocytochemical and histochemical study was done on 11 other cases (five females; six males; age range 51-83 years, mean 68 years) with short postmortem delays: inter- val from death to fxation or freezing ranged from 4 to 14 hours (mean 8 hours).
Samples were transported and dissected on crushed ice. Brains were halved sagittally and sampling was done on the hemisphere in which the septal pellucidal membrane was intact (left hemisphere in five cases and right hemi- sphere in six). The unfixed tissue was sliced in the vertico- frontal plane, using the anterior commissure, the optic chiasm, and the lamina terminalis as landmarks (Fig. 1). In spite of slight variations in the axis of the frontal sec- tions, three comparable samples were usually obtained, two of the slices being located rostrally to the anterior commis- sure and one caudal to it.
The 3-4-mm-thick blocks, trimmed to the area of interest, were either directly frozen in liquid nitrogen-cooled isopen- thane (six cases) or immersed for 6 hours in ice-cold 4% paraformaldehyde (in 0.1 M phosphate bufTer pH 7.4). Fixed
AC Acc ACHE aon AHR aPS Cd . ~~
DB DBH DBH-IR F1 Fx GP ic ~~
IC Lat V Ld Lde Ldi Li
anterior commissure nucleus accumbens acetylcholinesterase anterior olfactory nucleus anterior hippocampal rudiment anterior perforated space caudate nucleus diagonal band dopamine-0-hydrox y lase dopamine-0-hydroxylase immunoreactivity fornus longus fornix columns globus pallidus islands of Calleja internal capsule lateral ventricle laterodorsal septal nucleus laterodorsal septal nucleus pars externa laterodorsal septal nucleus pars interna latero-intermediate septal nucleus
Abbreuiations
Lv mm mos MS NBM NDB NST OLT OT PG PSa Pt PV Sf so SP ST TH TH-IR 3v
lateroventral septal nucleus medial forebrain bundle medial olfactory stria medial septal nucleus nucleus basalis of Meynert nucleus of the diagonal band of Broca nucleus of the stria terminalis olfactory tubercle optic tract parolfactory gyrus anterior parolfactory sulcus putamen paraventricular nucleus Septofimbrial nucleus supraoptic nucleus Septum pellucidum stria terminalis tyrosine hydroxylase tyrosine hydroxylase immunoreactivity third ventricle
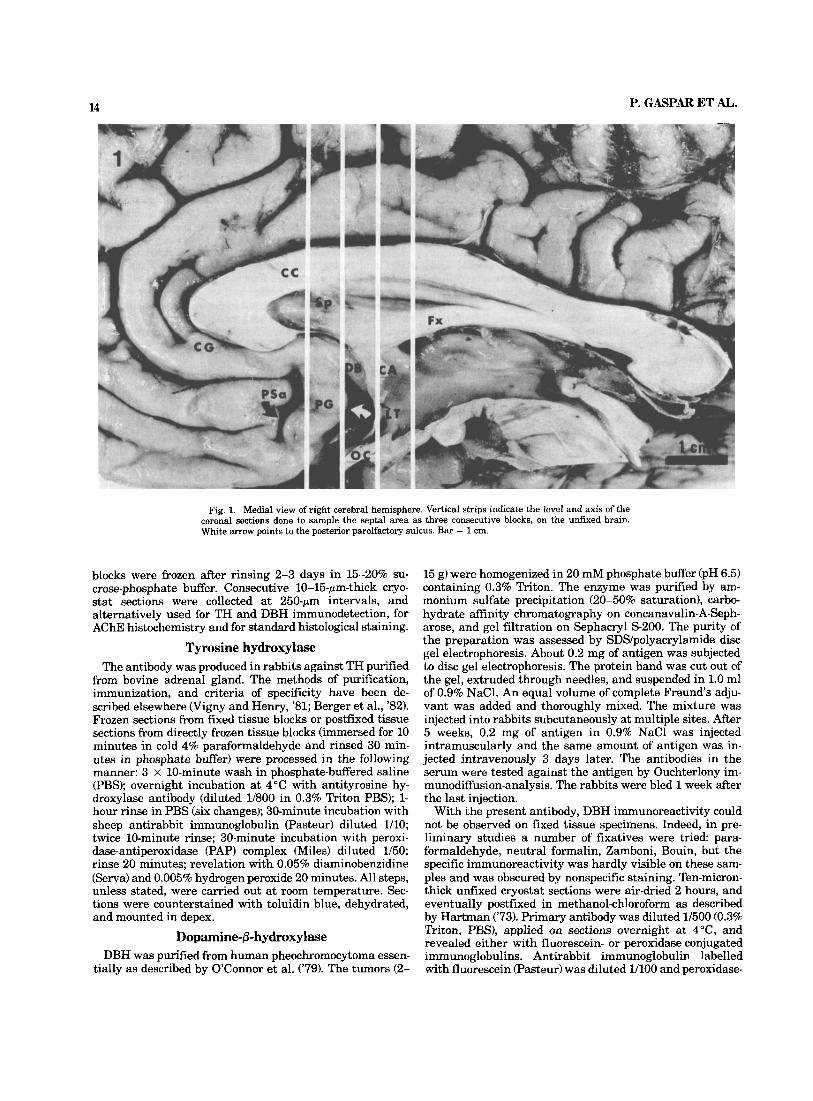
14 P. GASPAR ET AL. ~ -
Fig. 1. Medial view of right cerebral hemisphere. Vertical strips indicate the level and axis of the coronal sections done to sample the septa1 area as three consecutive blocks, on the unfixed brain. White arrow points to the posterior parolfactory sulcus. Bar = 1 cm.
blocks were frozen after rinsing 2-3 days in 15-20% su- crose-phosphate buffer. Consecutive 10-15-pm-thick cryo- stat sections were collected at 250-pm intervals, and alternatively used for TH and DBH immunodetection, for AChE histochemistry and for standard histological staining.
Tyrosine hydroxylase The antibody was produced in rabbits against TH purified
from bovine adrenal gIand. The methods of purification, immunization, and criteria of specificity have been de- scribed elsewhere (Vigny and Henry, '81; Berger et al., '82). Frozen sections from fixed tissue blocks or postfixed tissue sections from directly frozen tissue blocks (immersed for 10 minutes in cold 4% paraformaldehyde and rinsed 30 min- utes in phosphate buffer) were processed in the following manner: 3 x 10-minute wash in phosphate-buffered saline (PBS); overnight incubation at 4°C with antityrosine hy- droxylase antibody (diluted 1/800 in 0.3% Triton PBS); 1- hour rinse in PBS (six changes); 30-minute incubation with sheep antirabbit immunoglobulin (Pasteur) diluted 1/10; twice 10-minute rinse; 30-minute incubation with peroxi- dase-antiperoxidase (PAP) complex (Miles) diluted 1/50; rinse 20 minutes; revelation with 0.05% diaminobenzidine (Serva) and 0.005% hydrogen peroxide 20 minutes. All steps, unless stated, were carried out at room temperature. Sec- tions were counterstained with toluidin blue, dehydrated, and mounted in depex.
Dopamine-6-h yclrox ylase DBH was purified from human pheochromocytoma essen-
tially as described by O'Connor et al. ('79). The tumors (2-
15 g) were homogenized in 20 mM phosphate buffer (pH 6.5) containing 0.3% Triton. The enzyme was purified by am- monium sulfate precipitation (20-50% saturation), carbo- hydrate affinity chromatography on concanavalin-A-Seph- arose, and gel filtration on Sephacryl 5-200. The purity of the preparation was assessed by SDS/polyacrylamide disc gel electrophoresis. About 0.2 mg of antigen was subjected to disc gel electrophoresis. The protein band was cut out of the gel, extruded through needles, and suspended in 1.0 ml of 0.9% NaC1. An equal volume of complete Freund's adju- vant was added and thoroughly mixed. The mixture was injected into rabbits subcutaneously at multiple sites. After 5 weeks, 0.2 mg of antigen in 0.9% NaCl was injected intramuscularly and the same amount of antigen was in- jected intravenously 3 days later. The antibodies in the serum were tested against the antigen by Ouchterlony im- munodiffusion-analysis. The rabbits were bled 1 week after the last injection.
With the present antibody, DBH immunoreactivity could not be observed on fixed tissue specimens. Indeed, in pre- liminary studies a number of fixatives were tried para- formaldehyde, neutral formalin, Zamboni, Bouin, but the specific immunoreactivity was hardly visible on these sam- ples and was obscured by nonspecific staining. Ten-micron- thick unfixed cryostat sections were air-dried 2 hours, and eventually postfixed in methanol-chloroform as described by Hartman ('73). Primary antibody was diluted 1/500 (0.3% Triton, PBS), applied on sections overnight at 4"C, and revealed either with fluorescein- or peroxidase-conjugated immunoglobulins. Antirabbit immunoglobulin labelled with fluorescein (Pasteur) was diluted 1/100 and peroxidase-

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 15
linked donkey antirabbit globulin (Amersham) was diluted 1/50 and incubated 2 hours at room temperature. Peroxi- dase was revealed as aforementionned.
Acetylcholinesterase (AChE) activity was revealed ac- cording to the procedure of Karnovsky and Roots ('64). Pseudocholinesterase inhibitor ethopropazine (0.3 mM) was systematically added to the incubation medium. Control reactions included (1) omission of the substrate; (2) addition of cholinesterase inhibitor eserine sulfate (0.1 mM); (3) re- placement of acetylthiocholine by butyrylthiocholine. In the first two cases no staining was observed. When buty- rylthiocholine was used, the reaction product was scanty and essentially localized to blood vessels and to glial cells.
Analysis Bright- and darKield illumination were used to examine
the pattern of TH and DBH immunostaining. Charts of TH- and DBH-labelled structures (traced with an x-y plotter) were compared with cytoarchitectonic and histochemical data obtained from the Nissl- and AChE-stained sections of the same levels.
TH and DBH could be studied on alternate sections of the unfixed tissue blocks. Raw topographical data were trans- posed and schematized onto the series of line drawings obtained by the preliminary anatomical study.
RESULTS Cytoarchitecture and terminology
Because the delimitation, nuclear parcellation, and ter- minology of the human septal area have often differed in previous anatomical studies (Brockhaus, '42a,b; Crosby, '62; Andy and Stephan, '68; Nauta and Haymaker, '691, a pre- liminary reconsideration of the principal septal nuclei ap- peared to be a prerequisite to the topographical study of the catecholaminergic innervation. This was approached by both classical cyto-myelo-architectonic methods and histo- chemical detection of acetylcholinesterase (AChE) (Fig. 2). This enzymatic reaction, although not neurotransmitter- specific, provided information for further anatomical and functional subdivisions which were complementary to the cytoarchitectonic ones.
For the cytoarchitectonic compartmentation, we have re- lied on previous detailed works in man (Brockhaus '42a,b; Andy and Stephan '68) although we have identified a smaller number of nuclei. We have chosen to use a nomen- clature which would be more coherent with that used in the numerous anatomical, physiological, and biochemical studies in experimental animals and which would thus facilitate the comparison of immunocytochemical data ac- quired in man and rodents. The nomenclature employed was thus largely based on that established by Swanson and Cowan ('79) in the rat. Furthermore, it is in agreement with that used in other human anatomical studies (Crosby et al., '62; Nauta and Haymaker, '69). Terminological cor- respondences with other nomenclatural systems in man will be given as we proceed to the description of the differ- ent nuclear components.
General situation and limits. The human septal area occupies a portion of the medial telencephalic wall both in front and above the anterior commissure. Stretched be- tween the corpus callosum, the hemispheric base, and the fornix (Fig. l), it is composed of a dorsal part, the septum pellucidum, mainly formed of fibroglial elements and a ventral part, containing the septal nuclei.
The total extent of the septal nuclei in man was estimated to be of about 10 mm in the rostrocaudal axis and of 13 mm along the vertical axis.
They are limited rostrally and mediany by the anterior hippocampal rudiment and by the parolfactory gyrus Won Economo, '25; Nauta and Haymaker, '69) or area 25 of Brodmann. On a medial hemispheric view (Fig. l), the slight prominence of the medial precommissural septum, or diagonal band of Broca, is visible just below the beak of the corpus callosum and is separated rostrally from the parol- factory gyrus by the posterior parolfactory sulcus.
Caudally, the limits are formed by the anterior preoptic and hypothalamic nuclei, the anterior commissure, and the fornix columns. The laterodorsal limit is formed by the lateral ventricle. The ventrolateral limits consist of the nucleus accumbens, the islands of Calleja, and the anterior perforated space, which has been considered as more or less homologous to the olfactory tubercle of macrosmatics (Crosby and Humphrey, '41).
Nuclear components: cytoarchitectonic and histochemi- cal divisions (Fig. 2). Four main divisions were consid- ered: (1) the medial septal division consisting of the medial septal nucleus and the interstitial nucleus of the diagonal band of Broca; (2) the lateral septal division, subdivided into three main nuclei: dorsolateral, intermediolateral, and ventrolateral; (3) the posterior septal division, consisting of the atrophied and barely distinguishable triangular and septofimbrial nuclei; and (4) the bed nucleus of the stria terminalis.
(1) The medial septal division is located in an oblique band, along the interhemispheric fissure, and is character- ized by large-sized neurons with hyperchromic cytoplasm, irregular NissI staining, and frequent lipofuscin accumula- tion. The AChE-reactive neurons were observed only in this division among abundant AChE-stained fiber bundles. Two main nuclei could be individualized:
The medial septal nucleus (MS) corresponds to the nu- cleus diagonalis septalis of Brockhaus ('42b) or to the septal eomponent of the nucleus of the diagonal band of Andy and Stephan, ('68). This group formed a narrow strip of scat- tered neurons which extended for some distance above the interhemispheric fissure, along the fornix longus. Its ros- tralmost extension overlay the parolfactory cortical gyrus and the rostra1 hippocampal rudiment (Fig. 2a). Scattered neurons observed within the columns of the fornix (Fig. 2e), appeared to be a caudal extension of the MS. Neurons were medium to large-sized (18-35 pm large diameter) and only 30-60% of them (45% as a mean) contained intense AChE reactivity as judged from AChE-counterstained sections or from alternate Nissl- and AChE-stained sections.
The nucleus of the diagonal band (NDB), composed of a vertical and a horizontal portion, corresponds to the nu- cleus diagonalis angularis of Brockhaus ('42b) and to the tubercular portion of the diagonal band nucleus in Andy and Stephan's ('68) description.
Rostrally (Fig. 2b,c), the broad oblique mass of the verti- cal portion of the NDB lay in the ventral continuity of the MS, with which it had no sharp limits although it could be arbitrarily set at the level of a horizontal plane passing through the ventral tip of the lateral ventricle.
Caudally (Fig. 2d,e), the NDB arched ventrally and lat- erally below the ventral globus pallidus and anterior com- missure, forming the horizontal portion of the nucleus.
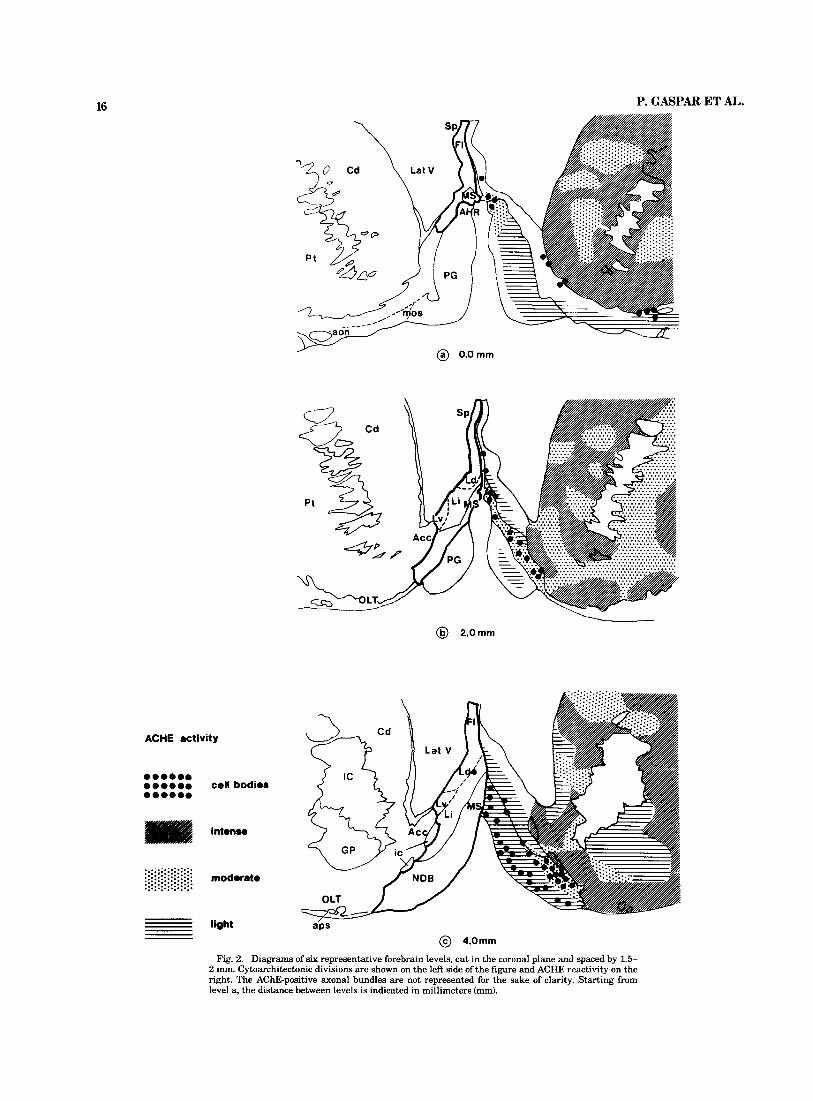
16 P. GASPAR ET AL.
ACHE activity
eeeeee eeeeee eeeeee
cell bodies
intense
moderato
light a'ps @ 4.0rnrn
Fig. 2. Diagrams of six representative forebrain levels, cut in the coronal plane and spaced by 1.5- 2 mm. Cytoarchitectonic divisions are shown on the left side of the figure and ACHE reactivity on the right. The AChE-positive axonal bundles are not represented for the sake of clarity. Starting from level a, the distance between levels is indicated in millimeters (mm).
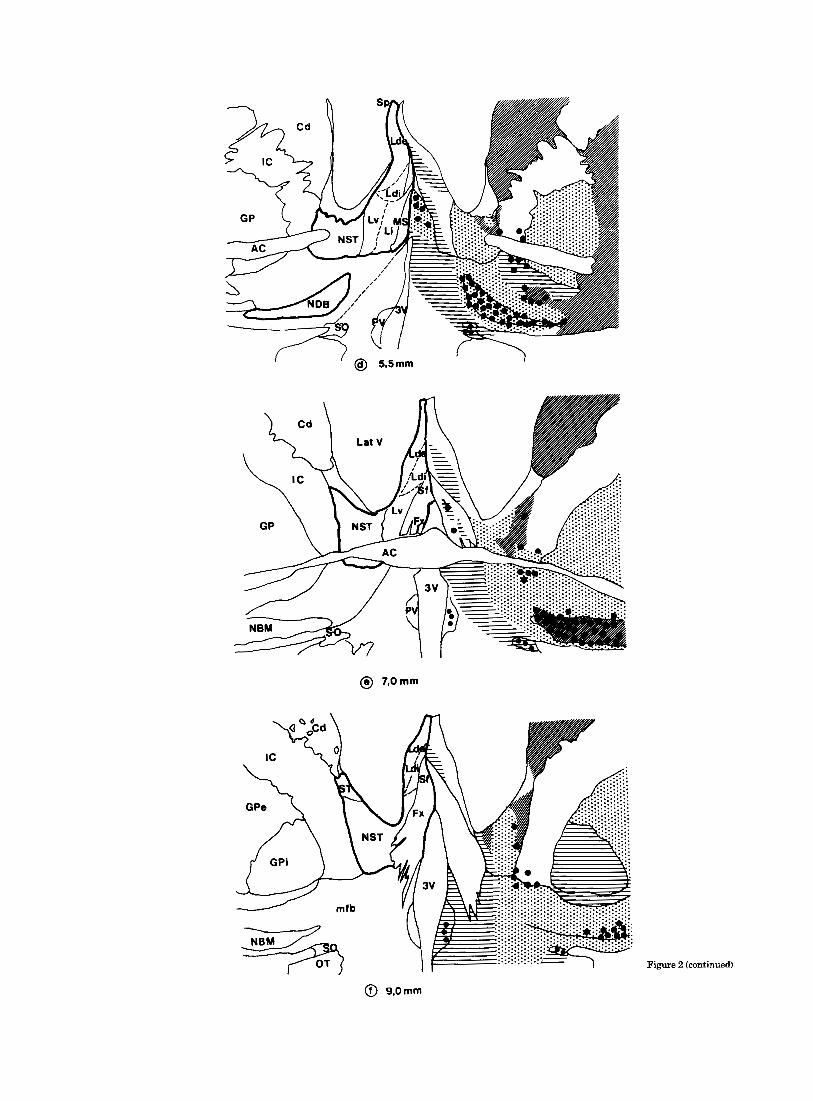
@ 7,Omm
Figure 2 (continued)

P. GASPAR ET AL. 18
Dorsally it became separated from the MS by the interven- ing cell masses of the anterior preoptic and hypothalamic nuclei. Laterally, it merged with the different cell groups of the nucleus basalis of Meynert (NBM) in the area of the substantia innominata.
The NDB neurons were more densely aggregated than in the MS and appeared to have larger cell sizes in the verti- cofrontal plane (20-55-pm broad diameter). The proportion of intensely AChE-reactive neurons with prominent den- dritic arborizations could be estimated to be 70% (60-80%) in the vertical portion of the NDB and 90% in the horizon- tal portion. However, very intense staining of the neuropil of the latter area rendered cell counts of the whole structure difficult.
Both the NDB and the MS, have been considered as the rostralmost continuation of the basal magnocellular com- plex (reviewed in Hedreen, '84). The link between the nu- cleus basalis of Meynert and the large neurons of the medial septal division is further supported by recent evidence in- dicating that this neuronal system provides the main source of cholinergic input to the telencephalic pallium (see Mesu- lam et al., '83a,b; McKinney et al., '83, for references). The delineation of the cholinergic neurons, on the basis of their immunolabelling with choline acetyltransferase (CAT) an- tibodies and their connections, both in rats and in primates, has led Mesulam et al. ('83a,b, '84) to propose an alternative nomenclature for the forebrain cholinergic neurons (Chl- Ch4). In this nomenclature, the MS corresponds to group Chl, the vertical portion of the NDB can be assimilated to Ch2, and the horizontal portion of NDB would correspond closest to what he has coined as the "antero-ventral part of Ch4," while no definite septal nucleus could be related to his Ch3 group. However, since a variable proportion of the neurons within these nuclei appears not to be cholinergic (Mesulam et al., '84), we have thought it preferable, in dealing with the present description of the septal area, to maintain the more classic cytoarchitectonic individualiza- tion of the MS and NDB.
(2) The lateral septal division was easily distinguished from the medial one by its smaller neurons, its small amount of myelinated fiber tracts, and its generally lower AChE reactivity of the neuropil and absence of AChE-posi- tive perikarya. It was further characterized by a laterome- dial increasing gradient of cell size and a dorsoventral increasing gradient of cell density. This allowed the distinc- tion of three main lateral septal nuclei.
The dorsolateral septal nucleus (Ld) corresponds to the n. septalis dorsalis of Andy and Stephan and to both the dorsal portion of n. septoventricularis and n. septofornicatus of Brockhaus ('42a). It stretched vertically between the mid- line and the lateral ventricle and extended caudally along the columns of the fornix. The pars externa (Lde) of this nucleus was the most extensive, characterized by a very low density of small, rounded neurons (12-20 pm broad diameter). The pars interna (Ldi) of the nucleus was a very conspicuous cell group visible at middle and caudal septal levels (Fig. 2d-D with densely packed fusiform perikarya. In AChE-stained preparations, the moderate staining in the neuropil of the pars interna formed a crescent which con- trasted with the absence of staining in pars externa.
The ventrolateral septal nucleus (Lv) corresponds to the n. septalis lateralis of Andy and Stephan, and to n. parvo- cellularis of Brockhaus. It formed a rounded ceIl mass which
became more conspicuous at midseptal levels (Fig. 2d). Neu- rons in the Lv were the smallest and most densely packed of the whole lateral septal division (large diameter 9-15 pm). Absence of AChE staining in the neuropil contrasted with the intense staining of the neighbouring n. accumbens and bed n. of the stria terminalis and facilitated its delimitation.
The intermediolateral septal nucleus (Li) was the least well-defined nucleus of the lateral septal division. Lying between the MS and the laterodorsal and lateroventral septal nuclei, the limits we have tentatively traced for it correspond to those of the medial septal nucleus of Andy and Stephan and to that of the ventral part of nucleus septoventricularis of Brockhaus. However, the terminology used by Andy and Stephan for this nuclear group appears to be confusing and inadequate, as both its cytological and histological properties are more similar to those of the other lateral septal nuclei. Indeed, it contains small to medium- sized neurons (maximal diameters ranging from 13 to 27 pm) which are not AChE reactive.
(3) As underlined by Andy and Stephan ('68, '76)' the two components of the caudal septal division are difficult to identify.
Nucleus septofimbrialis: Several authors (Lauer, '45; Crosby, '62; Nauta and Haymaker, '69) have designated as nucleus septofimbrialis the cell group which we considered to be the caudal continuation of the laterodorsal nucleus along the columns of the fornix. In the present account the scattered neuronal clusters in-between the columns of the fornix were considered to represent the nucleus septofim- brialis, following Andy and Stephan's description. The mod- erately dense array of AChE-reactive fibers running in the fornix expanded through the weakly reactive neuropil of this nuclear group.
The triangular septal nucleus does not figure on our dia- grams since there is no consensus as to its location. Indeed, Andy and Stephan located it between the ventral hippo- campal commissure and the fornix, while other authors (Crosby et al., '62; Nauta and Haymaker, '69) describe as the triangular nucleus a more conspicuous cell group lo- cated above the anterior commissure between the columns of the fornix.
(4) Description of the bed nucleus of the stria terminalis (NST) cell group is usually linked to that of the septal nuclei, although some authors considered it to belong to the amygdaloid complex on the basis of its cytological and neu- rochemical features as well as its connections (Krettek and Price, '78; Holstege et al., '84). Others have described it as a part of the striatum ("fundus subventricularis" for Brock- haus, '42a) or as a part of a wider complex which included also the anterior preoptic nuclei (Lauer, '45).
This nucleus lay caudal to nucleus accumbens and dorsal to the anterior commissure, wedged between the internal capsule and the lateral ventricle (Fig. 2d-D. It contained a heterogenous cell population with small (10-14 pm large diameter) and medium-sized neurons (15-20 pm large di- ameter) and could be further subdivided into a medial and external portion on the basis of cellular aggregation. The medial and rostra1 nuclear boundaries were not distinct on the Nissl-stained sections, but could be easily traced with the AChE reaction. Indeed, the moderate staining of the neuropil in the NST contrasted with both the intense stain- ing of the nucIeus accumbens rostrally and with the pale staining of the ventrolateral septal nucleus medially. AChE
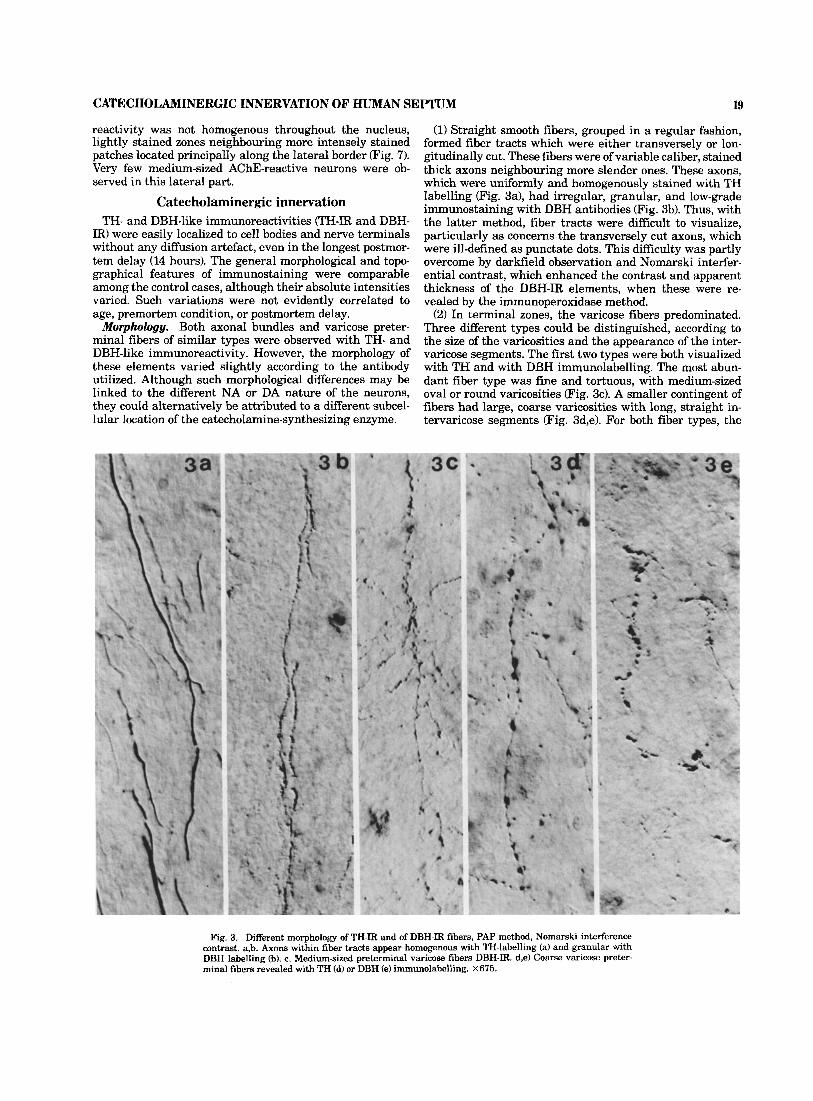
CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 19
reactivity was not homogenous throughout the nucleus, lightly stained zones neighbouring more intensely stained patches located principally along the lateral border (Fig. 7). Very few medium-sized AChE-reactive neurons were ob- served in this lateral part.
Catecholaminergic innervation TH- and DBH-like immunoreactivities (TH-IR and DBH-
IR) were easily localized to cell bodies and nerve terminals without any diffusion artefact, even in the longest postmor- tem delay (14 hours). The general morphological and topo- graphical features of immunostaining were comparable among the control cases, although their absolute intensities varied. Such variations were not evidently correlated to age, premortem condition, or postmortem delay.
Morphology. Both axonal bundles and varicose preter- minal fibers of similar types were observed with TH- and DBH-like immunoreactivity. However, the morphology of these elements varied slightly according to the antibody utilized. Although such morphological differences may be linked to the different NA or DA nature of the neurons, they could alternatively be attributed to a different subcel- lular location of the catecholamine-synthesizing enzyme.
(1) Straight smooth fibers, grouped in a regular fashion, formed fiber tracts which were either transversely or lon- gitudinally cut. These fibers were of variable caliber, stained thick axons neighbouring more slender ones. These axons, which were uniformly and homogenously stained with TH labelling (Fig. 3a), had irregular, granular, and low-grade immunostaining with DBH antibodies (Fig. 3b). Thus, with the latter method, fiber tracts were difficult to visualize, particularly as concerns the transversely cut axons, which were ill-defined as punctate dots. This difficulty was partly overcome by darkfield observation and Nomarski interfer- ential contrast, which enhanced the contrast and apparent thickness of the DBH-IR elements, when these were re- vealed by the immunoperoxidase method.
(2) In terminal zones, the varicose fibers predominated. Three different types could be distinguished, according to the size of the varicosities and the appearance of the inter- varicose segments. The first two types were both visualized with TH and with DBH immunolabelling. The most abun- dant fiber type was fine and tortuous, with medium-sized oval or round varicosities (Fig. 3c). A smaller contingent of fibers had large, coarse varicosities with long, straight in- tervaricose segments (Fig. 3d,e). For both fiber types, the
Fig. 3. Different morphology of TH-IR and of DBH-IR fibers, PAP method, Nomarski interference contrast. a,b. Axons within fiber tracts appear homogenous with TH-labelling (a) and granular with DBH labelling (b). c. Medium-sized preterminal varicose fibers DBH-IR. d,e) Coarse varicose preter- minal fibers revealed with TH (d) or DBH (e) immunolabelling. X675.
20 T H - l i ke immunoreactivity
P. GASP& ET AL. DBH- l ike immunoreactivity
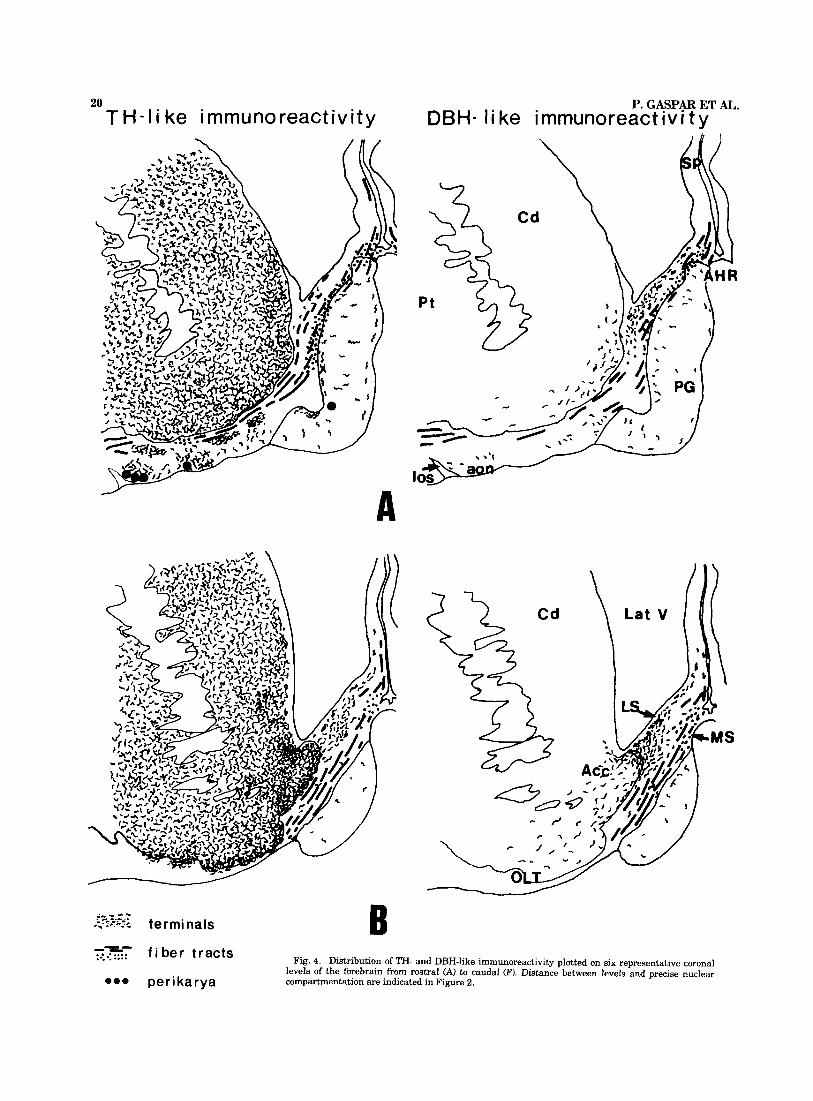
Fig. 4. Distribution of TH- and DBH-like immunoreactivity plotted on six representative coronal levels of the forebrain from rostra1 (A) to caudal (F). Distance between levels and precise nuclear compartmentation are indicated in Figure 2.

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM
T H - I R A\ ,-
21
DBH- I R
C
Figure 4 (continued)

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 23
intervaricose segments, well visualized with TH-IR, were fine or hardly visible with DBH-IR.
The last fiber type, consisting of a succession of very fine, dotlike varicosities, was only visible with TH-IR and could thus reflect specific DA-containing terminals. This latter type of fiber frequently surrounded neuronal perikarya and occasionally formed typical pericellular basketlike arrange- ments (Fig. 5c).
Ilbpography. The distribution of TH-IR and DBH-IR is represented diagrammatically at six representative verti- cofrontal levels of the forebrain (Fig. 4A-F) selected in the previous study.
Fiber tracts. The prominent catecholaminergic fiber tracts entering the septal area, or traversing it, had a roughly similar topography with DBH and TH immu- nostaining.
Incoming fibers to the septal nuclei could be followed essentially from the medial zone of the medial forebrain bundle and from the dense TH-IR fiber bundle surrounding the anterior commissure (Fig. 4D-F). Thereafter, fibers followed two main courses within the septal nuclei: a cau- dal contingent of catecholaminergic fibers ran dorsally to- ward the stria terminalis, forming a vertically oriented axonal bundle which followed the lateral border of the bed nucleus of stria terminalis medially to the internal capsule (Fig. 4E,F). This bundle provided fibers to the bed nucleus of stria terminalis, the caudal portion of the lateral septal nuclei, and the stria terminalis.
The largest contingent of catecholaminergic fibers ap- peared to be the rostral continuation of the medial fore- brain bundle, within the diagonal band. Two fascicles, running ventrodorsally in the lateral part of the diagonal band and just below the pial surface, respectively, provided fibers to the medial and lateral septal divisions. Dorsally, a fascicle continued into the septum pellucidum, along the precommissural fornix-fiber bundle, most of it appearing transversely cut in the coronal sections. These fibers could be followed up to the level of the corpus callosum but were not seen to penetrate into it. Rostrally, the TH-IR and DBH-IR fibers of the diagonal band appeared to be contin- uous with a transversely cut bundle oriented horizontally toward the cortex of the frontal lobe. This tract was located in the dorsal part of the medial septal nucleus and in- creased in bulk rostrally. It could be followed within the anterior hippocampal rudiment (Fig. 4Af and in the white matter underlying the medial cortex of the frontal lobe. At this level (Fig. 4A) labelled axons ran very close to the deepest cortical strata and were joined by oblique medi- odorsal fibers arising from below the striatum.
Terminal catecholaminergic fields. These fields had a characteristic pattern of distribution in the different septal nuclei, which differed significantly for DBH and TH-IR.
The medial septal division received only moderate ca- techolaminergic innervation with predominant DBH im- munolabelling. The DBH-IR terminal or preterminal fibers were essentially of the medium-sized varicose type, al- though a few fibers with coarse varicosities were present. Labelled fibers were observed in the close vicinity of the large hyperchromic neurons of the medial septal and diag- onal band nuclei but did not form close perikaryal contacts or pericellular arrangements.
The lateral septal division contained the major part of the catecholaminergic terminal innervation. The DBH-IR fi-
bers and terminals occupied a large area in the lateral septum, which was more extensive at rostral than at caudal levels. Rostrally (Fig. 4B,C) a moderately dense terminal network occupied the whole width of the ventrolateral and dorsolateral septal nucleus. This terminal field was contig- uous with that of nucleus accumbens but had a higher density of terminals. Caudally (Fig. 4D,E), DBH-IR fibers gradually became less numerous and tended to cluster ven- trally into two bands: laterally, close to the ventricular wall and medioventrally, overlapping the ventrolateral and in- termediolateral septal nuclei. DBH-IR fibers in the lateral septum were mainly of the medium-sized varicose type and did not form pericellular arrangements. Some coarse vari- cose fibers were also observed below the ventricular wall, coming into close contact with the subependymal cell layer and with the adventitia of blood vessels. No labelled termi- nals could be seen in direct contact with the ependymal cell layer.
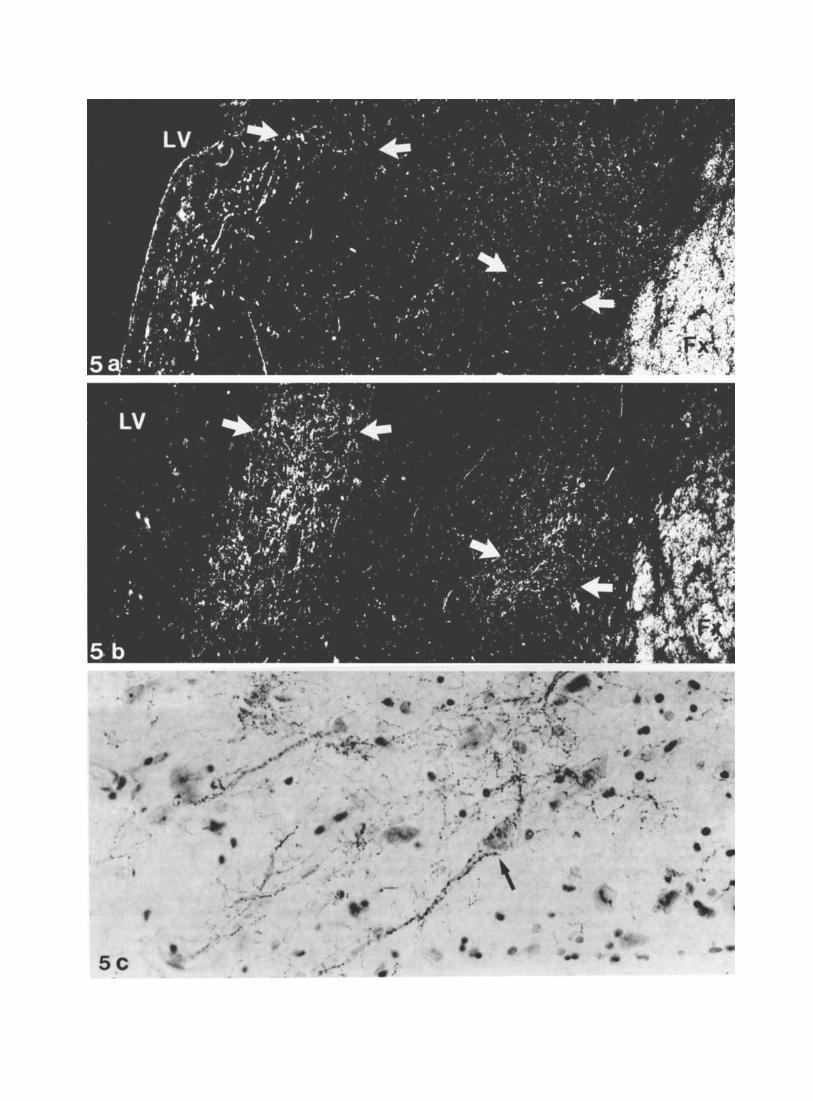
The TH-IR fibers had a slightly different pattern. Ros- trally, the TH-IR varicose network was less dense than that revealed with DBH, offering a sharp contrast to the densely innervated neighbouring structures such as the islands of Calleja and nucleus accumbens. Caudally, the septal TH-IR fibers increased in number and were clustered into three longitudinal strips. The lateral and medioventral bands partly overlapped those observed with DBH immunostain- ing but contained a much greater number of fibers (Fig. 5a,b). An additional terminal field was present dorsally in the pars interna and pars externa of the dorsolateral septal nucleus. Besides medium and coarse varicose fibers, similar to those visualized with DBH-IR, dotlike varicose TH-IR axons were observed in fairly large number either dis- persed, or forming dense perineuronal arrangements, with varicosities heavily surrounding the perikarya and their proximal dendrites (Fig. 5c). These formations, characteris- tic of the dorsolateral septal nucleus, caudally, were also occasionally observed in the medioventral field.
The nucleus septofimbrialis contained only a few DBH-IR varicose fibers but numerous TH-IR axons predominantly of the dotlike varicose type forming clusters interspaced between the unstained fornix bundles. TH-IR perineuronal arrangements were also a characteristic of this area.
In the space between the anterior pillars of the fornix, a moderate amount of DBH and TH-IR varicose fibers ex- tended in the anterior preoptic nuclei.
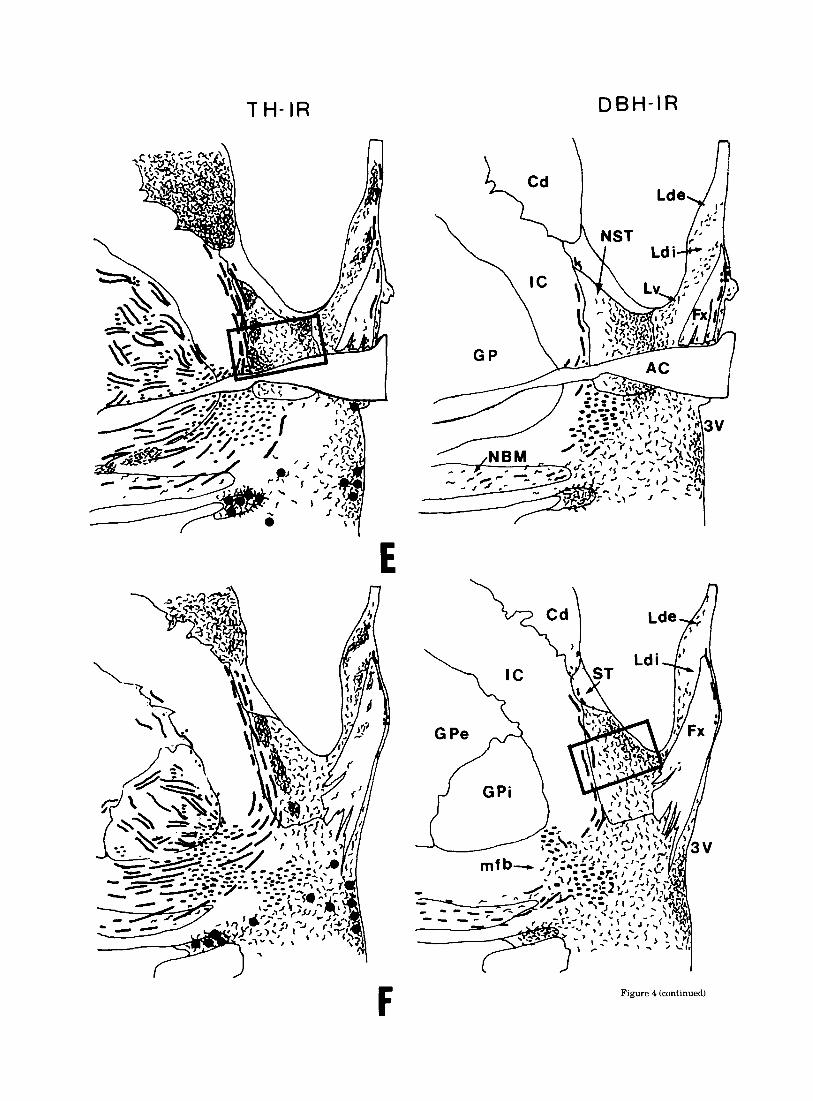
The bed nucleus of stria terminalis (NST) contained a massive catecholaminergic innervation. Abundant DBH-IR fibers were concentrated in the medial portion of the nu- cleus, the lateral portion being scarcely innervated. The terminal field in the NST was continuous ventrally with that of the medial preoptic area and medially with that of the ventrolateral septal nucleus. It consisted of about equal amounts of coarse and medium-varicose fibers.
Topography of the TH-IR terminal field was complemen- tary to that of DBH (Fig. 6a,b). Indeed, the varicose TH-IR fibers were unhomogenously distributed in a patchy man- ner, densely innervated areas neighbouring more lightly labelled zones. The dense TH-IR clusters were frequently lined along the the lateral edge of the NST or surrounded the dorsal and ventral aspect of the anterior commissure. Within these clusters, fine dotlike varicose fibers were most abundant and frequently formed basketlike pericellular ar- rangements. A rostrocaudal decreasing gradient of TH-IR terminals was also noted in the NST, particularly caudally

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 25
to the decussation of the anterior commissure where la- belled fibers became scarce.
Relationships of the catecholaminergic innemation to AChE reactiuity. Comparison of AChE activity with TH immunolabelling on consecutive sections revealed some striking topographical similarities especially in terminal fields containing densely packed dotlike varicose TH-IR fibers. Thus, a close correspondence was observed between the dense clusters of TH-IR terminals and the intensely AChE-reactive patches in the nucleus of the stria termin- alis (Fig. 7a,b). Similarly, in the lateral septum, the cres- cent-shaped area with moderate AChE reactivity cor- responded fairly well with the dorsal TH-IR terminal field.
Structures closely related to the septum (1) The anterior hippocampal rudiment, located at the
rostromedial edge of the septa1 nuclei, overlying the parol- factory gyrus, contained a transversely cut DBH and TH- IR fiber tract (Fig. 4A). In addition, moderate amounts of varicose DBH-IR fibers were visible between the pyramidal neurons, indicating a moderate terminal innervation. (2) The parolfactory cortex (Brodmann area 25) was distin-
guishable from the more rostra1 medial frontal cortex by its agranular structure. It contained only scarce catecholamin- ergic terminal innervation which appeared similar to that of more rostrally situated cortical fields (Brodmann areas 11 and 12). This consisted of a loose array of medium-sized varicose fibers oriented in tangential and radial planes, which seemed comparably stained with TH and DBH anti- bodies. A peculiar feature of this cortex was the presence of TH-immunolabelled neuronal perikarya. Although only one or two were generally observed on a section among numer- ous unstained neurons, they could be observed in almost all the cases studied and had a similar location in the deep cortical layers (V, VI). These small, fusiform, bipolar neu- rons (15-20 pm diameter, 140 pm2 cell area) had some resemblance to the TH-IR neurons of the anterior olfactory nucleus (vide infra), although anatomical continuity be- tween both was not clear. Precise estimation of their distri- bution and frequency would require further studies. Such perikarya were never DBH-immunolabelled, which strongly suggests their DA nature.
(3) The anterior olfactory nucleus and olfactory tubercle area has a complex structural organization consisting of several cytoarchitectonic divisions, for which we have fol- lowed the nomenclature of Crosby and Humphrey ('41). Rostrally, at the level of the anterior olfactory nucleus, a zone with moderately dense TH-IR terminals was visible between the medial and lateral olfactory stria, which con- trasted with the weak labelling of the neighbouring cortex. At this level, a group of TH-immunoreactive perikarya was present, lying at the periphery of the anterior olfactory nucleus, just below the pial surface, in the plexiform, cell- poor layer. These neurons were small with scant cytoplasm
Fig. 5. Catecholaminergic innervation in the caudal part of the lateral septum. a,b. Terminal fields are visualized as two bands: medially, next to the fornix (Fx) and laterally, below the lateral ventricle GV). A dense TH- IR network (pointed to by arrows in b) compared with Scarce DBH labelling (arrows point to the corresponding area in a) argue for the predominantly dopaminergic nature of the labelled fibers. Darkfield illumination. X68. c. Basketlike arrangement of dotlike varicose TH-IR fibers (arrow) around a perikaryon of the lateral septum and its proximal dendrites PAP method, toluidin blue counterstain. ~500.
(10-17 pm large diameter and 100 pm2 mean cell surface on coronal sections) and had round or fusiform shapes. In horizontal sections, including the olfactory stalk and pe- duncle, these TH-stained neurons appeared to be the caudal continuation of a fairly important cell group which followed the olfactory peduncle and the different components of the anterior olfatory nucleus. These neurons lay within the outer circumference of the peduncle. They presented very conspicuous dendritic arborizations (Fig. 8,b,c) which were oriented in the horizontal plane and extended for distances over 200-300 pm. In tangential sections, a cluster of TH-IR perikarya could be seen at the level of the attachment of the olfactory peduncle to the cortex (Fig. 8,a). These peri- karya were not labelled with DBH antibodies.
Caudally, at the level of the anterior perforated space or olfactory tubercle, as defined by Crosby and Humphrey ('41), an intricate arrangement of TH-IR axonal bundles and terminal fields was observed. Two types of TH-IR in- nervation could be distinguished one consisted of dense clustering of fine varicose fibers and terminals forming a network which resembled that visible in the striatum; the other TH-IR terminals were arranged according to a pat- tern which was specific for the Calleja islands (Fallon et al., '83). In the latter, a dense aggregation of TH-IR varicose fibers surrounded the granular cells, in a ringlike arrange- ment, with only few fibers penetrating between the granu- lar cells and into the central core of the Calleja island complex. These Calleja islands were disseminated not only within the olfactory tubercle but also along the medial border of nucleus accumbens.
Contrary to the distribution of TH-IR, DBH-IR fiber tracts and terminals were very scarce over the whole extent of the anterior olfactory nucleus and olfactory tubercle. In the islands of Calleja, they were evenly distributed within the core and the rim of the complex. (4) The pattern of catecholaminergic innervation in nu-
cleus accumbens distinguished it from neighbouring struc- tures. Indeed, the TH-IR network, instead of being homogenously distributed as in the caudate and putarnen, tended to form patches of more densely aggregated termi- nals. Also, the presence of DBH-IR fibers restricted to the medial and ventral zones of the nucleus differentiated it from the rest of the striatum and from the olfactory tuber- cle. Nevertheless, DBH-IR fibers were in far lower amounts in nucleus accumbens than in the bed nucleus of stria terminalis.
No clear relationship could be established between the dense TH-IR patches and the clusters of smaller neurons also present in nucleus accumbens, nor with the islands of intense AChE staining in the neuropil.
DISCUSSION Methodological considerations
The validity of immunocytochemical detection of TH and DBH for mapping catecholaminergic neurons has been am- ply demonstrated in animals ever since the pioneering stud- ies of Hartman ('73), Hokfelt et al. ('73, '771, and Pickel et al. ('75). The applicability of these techniques to human postmortem material has also been shown (Hartman, '74; Pearson et al., '79, '83; Gaspar et al., '83). The specificity of the antisera used in the present study was established by immunochemical tests (Vigny and Henry, '81) as well as by numerous immunocytochemical investigations (Berger et al. '82; Verney et al., '82, '85) with the TH antiserum.

Fig. 6. Micrograph of the bed n. of the stria terminalis, corresponding to the mirror image of the boxed area in Figure 4F. Midline is here to the left of the figure. The complementary pattern of DBH-IR and TH-IR is shown on consecutive sections with darkfield illumination, the star indicates the location of the same blood vessel a. DBH-IR terminal network predomi-
nates in the medial portion of the nucleus, beneath the lateral ventricle (LV). b. TH-IR forms dense clusters of terminals, laterally along the internal capsule (IC). Patches of TH-ZR terminals, pointed to by arrows in b, contain only scarce DBH-labelling in a. DBH is revealed with immunoperoxidase and TH with the PAP method. x44.
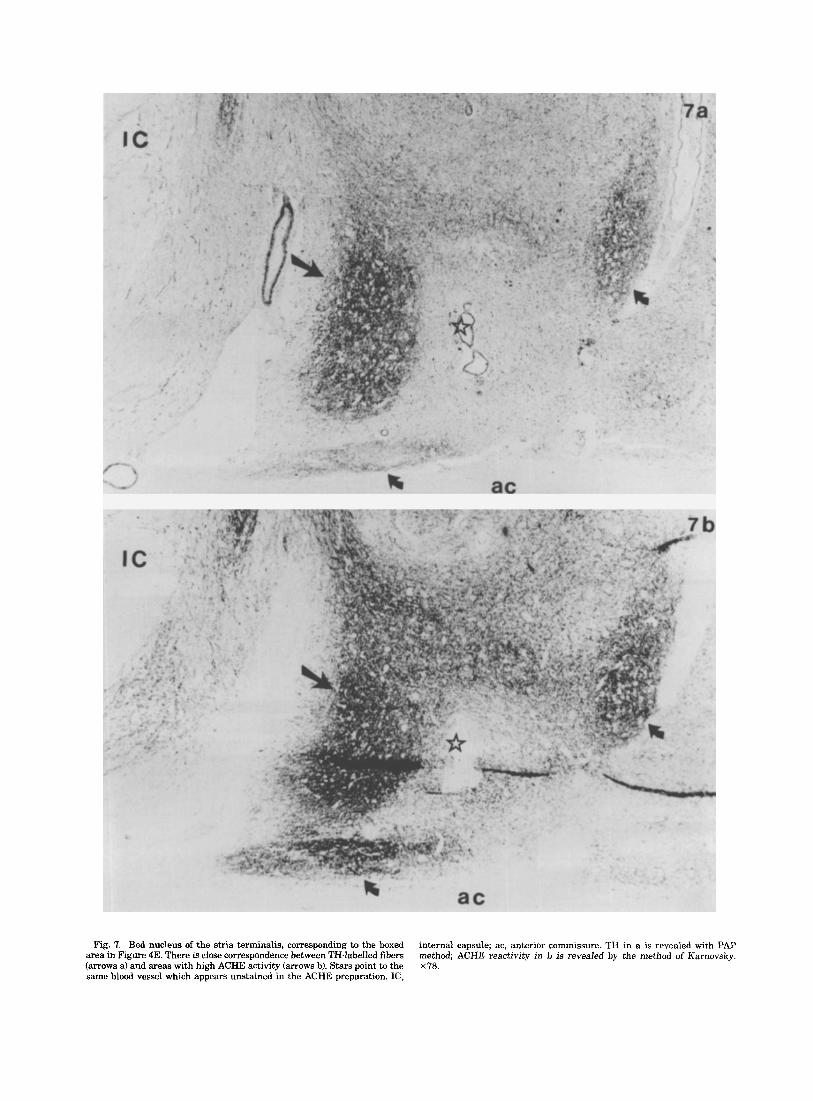
Fig. 7. Bed nucleus of the stria terminalis, corresponding to the boxed area in Figure 4E. There is close correspondence between TH-labelled fibers (arrows a) and areas with high ACHE activity (arrows b). Stars point to the same blood vessel which appears unstained in the ACHE preparation. IC,
internal capsule; ac, anterior commissure. TH in a is revealed with PAP method; ACHE reactivity in b is revealed by the method of Karnovsky. x78.

28 P. GASPAR ET AL.
Fig. 8. TH-labelled neurons, at the level of the anterior olfactory nucleus. PAP method, brightfield illuminaton. a. Horizontal section of the olfactory tract at the level of its attachment to the forebrain, revealing a cluster of these neurons (arrows). Rostra1 here is to the right of the figure. X78.
Higher magnification of neurons present in the boxed area in a. Toluidin blue counterstain. ~600. c. Extensive, Golgi-like dendritic arborizations of these small neurons can be seen in the horizontal plane. x500.

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 29
A limitation of our method resided in possible false-nega- tive reactions, since, with the human-specific DBH anti- body used in this study, specific immunoreactivity was substantially reduced by various aldehydic fixatives. How- ever, on directly frozen brain samples, extensive DBH im- munolabelling was observed. Since on these unfixed specimens the TH labelling was identical to that revealed on the paraformaldehyde-fixed samples, it may be assumed that a sufficient immobilization of both catecholamine-syn- thesizing enzymes was achieved with the method employed.
When using postmortem brain material, one must also consider possible degradation of the antigens andor diffu- sion out of the cellular processes. However, relative stabil- ity of both TH (Vogel et al., '69; Grote et al., '74; Puymirat et al., '79) and DBH (Vogel et al., '69) has been demon- strated in the early postmortem period (up to 16 hours). On the other hand, no diffusion artefacts could be observed on our tissue sections. The reliability of these methods in the human postmortem brain has also been demonstrated by concordant results obtained by different workers using dif- ferent antibody sources to TH (Gaspar et al., '83; Pearson et al., '83; German et al., '84).
The present study is the first attempt to identify and visualize the dopaminergic (DA) and noradrenergic (NA) innervation of the human septal area. This distinction was rendered difficult since TH, the first enzyme in catechol- amine biosynthesis, is present in the three types of cate- cholaminergic neurons (DA, NA, and adrenergic), whereas DBH, the noradrenergic-synthesizing enzyme, is contained in both NA and adrenergic neurons. However, this attempt was partly successful inasmuch as TH with no correspond- ing DBH labelling revealed DA neurons, whereas DBH labelling could be considered, in first approximation, to reflect essentially noradrenergic neurons.
Identification of the DA innervation was facilitated by the greater amount of TH in dopaminergic than in norad- renergic fibers, proved both biochemically (Schmidt and Bhatnagar, '79) and immunocytochemically in rodents (Pickel et al., '75; Hokfelt et al. '77; Gall and Moore, '84) and also observed in the present work.
The topography of DBH-IR corresponded closely to the NA innervation described in the rodent septal area. How- ever, the possibility that some of the DBH-IR fibers might be adrenergic cannot be excluded. Indeed, there have been contradictory biochemical reports on the presence of mod- erate amounts of adrenaline in the rodent septal nuclei (Van der Guten et al., '76) although immunocytochemical methods failed to detect terminals containing the adrener- gic synthesizing enzyme phenylethanolamine-N-methyl- transferase (PNMT) in this area (Hokfelt et al., '74). In man, the presence of relatively high levels of PNMT in the sep- tum (Lew et al., '77; Trocexicz et al., '82) could indicate that a subpopulation of the DBH-IR fibers might be adrenergic. On the other hand, the description of hypothalamic neurons which contained PNMT but not TH (Ross et al., '84) sug- gested that this enzyme might be present in noncatecho- laminergic neurons and subserve the methylation of other substrates than noradrenaline.
Catecholaminergic innervation of the septal area The catecholaminergic innervation of the human septal
area had a characteristic pattern of distribution-moderate or scarce in the medial septal division and denser in the lateral septal division and in the bed nucleus of stria ter- minalis. This organization was concordant with the well-
known topography of catecholaminergic fibers in the rat septal area (Fuxe, '65; Hokfelt et al., '77; Lindvall and Stenevi, '78; Moore, '78) and with the data obtained in human embryos using histofluorescence techniques (Nobin and Bjorklund, '73). The present immunocytochemical re- sults in adult human brain are also in agreement with the few biochemical measurements that have been made in postmortem samples (Farley et al., '77; Price et al., '78; Nyberg et al., '82).
Puthwags. Since TH-IR and DBH-IR fiber tracts always coexisted in superimposable topographical locations, no re- liable identification of the DA axons could be achieved. This distinction is also lacking in the few studies carried on in primates (Ishikawa and Tanaka, '77; Schofieid and Dixson, '82) or human fetuses (Nobin and Bjorklund, '73). However, when catecholaminergic pathways are considered globally, some correspondences can be found with our data. The caudal bundle, providing fibers to the stria terminalis and its bed nucleus, was in a location similar to an efferent axonal bundle from the locus coeruleus, traced in the ma- caque (Bowden et al., '78). The rostral catecholaminergic bundle, providing fibers to the diagonal band, has also been observed in human fetuses (Nobin and Bjorklund, '73) and its rostral continuation as a medial subcortical fiber tract has been described in the macaque (Bowden et al., '78, Levitt et al., '84).
DBH-positive terminal innervation. Although immuno- cytochemistry did not allow a morphological delineation as exquisite as the glyoxylic acid method, the two distinct types of DBH-IR terminals identified in the human septum were reminiscent of the two types of noradrenergic fibers described in the rodent septal nuclei (Lindvall and Stenevi, '78; Moore '78). In rodents, they could be related to different origins in the brainstem. Locus coeruleus NA fibers were identified as fine, branched fibers with medium-sized vari- cosities, whereas fibers with coarse varicosities were shown to arise from NA cell groups in the medulla oblongata. It could be hypothesized that in man, too, the specific mor- phology of DBH-labelled fibers in the septal nuclei was related to the origin of the NA septal afYerents, in both the locus coeruleus, as has been demonstrated in primates (Krayniak et al., '81) and the lower brainstem.
The topographical organization of the DBH-IR innerva- tion roughly paralleled that of the septal NA innervation in rodents (Swanson and Hartman, '75; Lindvall and Ste- nevi, '78; Moore, '78) with three distinctive features. The lateral septal division had a denser terminal network than the medial division (medial septal nucleus and diagonal band of Broca), which was confined to its ventral and inter- mediate portions with a decreasing rostrocaudal gradient in fiber density. Another discrepancy concerned the distri- bution of DBH-IR in the bed nucleus of stria terminalis (NST). In man, it was concentrated in the medial portion of the nucleus, whereas in the rat it has been described mainly in a ventral location below the anterior commissure. This difference could reflect the completion of a mediolateral differentiation within the nucleus in higher species. In fact, a functional mediolateral subdivision in the NST is already present in rat and cat as indicated by hodological studies (Krettek and Price, '78; Weller, and Smith, '82) and neuro- peptide mapping (Woodhams et al., '83), although it is not followed by a corresponding segregation of the DA and NA inputs (at least in rats), contrary to what was observed in man. Alternatively, the absence of a massive DBH-IR input to the ventral portion of NST in man could reflect the

30
decrease, or topographical displacement, of a particular type of NA afferents. Indeed, in rodents, the NA innervation of this area has been shown to be essentially provided by the medulla oblongata NA neurons (Lindvall and Stenevi, '78). Finally, the presumed nucleus septofimbrialis contained very rare DBH-IR terminal fibers, seemingly in much lower numbers than the NA innervation to this nucleus in rodents.
The dopaminergic innervation. This innervation was identified among the TH-labelled terminals by the absence of concomitant DBH-IR. This was particularly clear for the dotlike varicose fibers and the basketlike pericellular ar- rangements. As in the rodent (Lindvall and Stenevi, '78; Moore, '78; Gall and Moore, '84; Onteniente et al., '84), the DA innervation appeared to be restricted to the lateral septal division and to the bed nucleus of stria terminalis, with some differences concerning its intranuclear dis- tribution.
The terminal DA network in the lateral septum of rodents has generally been described in three subfields: rostral, medial, and subventricular, roughly organized in longitu- dinal strips. It was further characterized by the presence of numerous basketlike pericellular arrangements. In man, three subfields were also identified a dorsal one without any DBH-IR, and lateral and medioventral ones partially overlapping the DBH-IR terminal zones but with greater numbers of TH-IR fibers. No DA fibers could be detected in the rostral portion of the lateral division. Characteristic basketlike pericellular arrangements of the DA terminals around some lateral septal neurons were also conspicuous in human specimens. However, they appeared to be more restricted than in rodents, both in number and topography, being observed only at caudal levels, and principally in the dorsal DA field. This raises the question of a possible phy- logenetic regression of this particular mode of DA innerva- tion or of the specific neuronal targets to which it is destined. On the other hand, a prominent DA innervation and pericellular disposition of terminals was observed in the presumed nucleus septofimbrialis, contrasting with the scarce DA projection in rodents. This general caudal exten- sion of the DA fields could be explained by the structural modification of the septal area in man, as it seemed to follow the vertical stretching and caudal displacement of the septal nuclei.
In the bed nucleus of stria terminalis, the DA projection, including basketlike pericellular formations, was clustered in the outer portion of the nucleus instead of being homog- enously distributed in the dorsal part as in rodents. This lateral disposition of the DA input, complementary to the medial location of the NA projection, confirmed the medi- olateral organization of the NST in man.
A good overlap was observed between the DA terminal fields and the AChE-reactive neuropil in both the lateral septum and the NST. A similar correspondence has also been noted in the lateral septum of rodents (Lindvall, '77) and probably represents an example of a close association of AChE to DA neurons which has been described both at the level of cell bodies and of terminals (reviewed in Green- field, '84).
Catecholaminergic innervation in neighbouring structures
Nucleus accumbens. Two features characterized the ca- techolaminergic innervation of this nucleus. The first was
P. GASPAR ET AT,.
the clustering of the DA terminals, contrary to their more homogenous distribution in the caudate. These clusters only partly overlapped the patches of intense AChE reaction. Similar data have been reported in other species, and seem to reflect the complex organization of this area (reviewed in Graybiel and Ragsdale, '83; Herckenham et al., '84). See- ondly, the presence of DBH-IR innervation in this structure contrasted with the absence or scarcity of DBH-labelled fibers in the neighbouring caudate nucleus and olfactory tubercle. Our observation is concordant with biochemical studies in the human brain showing significant levels of NA in nucleus accumbens relative to the caudate nucleus (Farley et al., '78; Mackay et al., '78; Nyberg et al., '82). It is noteworthy that the DBH labelling corresponded to an area enriched in glutamic acid decarboxylase (GAD), as shown biochemically in microdissected human brains (Kwak et al. '84).
Finally, this medial area of DBH-IR afferents corre- sponded strikingly to the area defined as the hippocampal district (Kelley and Domesick, '82) in the rodent nucleus accumbens and corresponding to the specific terminal field of hippocampal input to the striatum. The nucleus accum- bens may thus be individualized by some specific neuro- chemical features in man, as well as in rodents (Herkenham et al., '84), although it is not well delimited cytoarchitecton- ically from the rest of the striatum.
Catecholaminergic cell group in the basal forebrain. The dopaminergic nature of the TH-labelled neurons ob- served at the level of the anterior olfactory nucleus and the parolfactory cortex is strongly suggested by their lack in DBH immunoreactivity. The presence of a similar group of neurons has not been reported in the brain of mature ro- dents. However, in the monkey, Kohler et al. ('83) have demonstrated a TH-immunolabelled neuronal group which extended from the orbitofrontal cortex through the olfac- tory and pyriform cortices. Although the latter cell group appears much more extensive than that observed in man, common morphological features and topographical loca- tions (around the olfactory tract) suggest that they may represent a similar catecholaminergic cell group particular t o the primate forebrain. In this respect, it was of interest t o note that these neurons were in continuity with the olfactory bulb dopaminergic neurons, as they had similar morphological aspects and were aligned all along the olfac- tory peduncIe. Incomplete migration of the olfactory DA neurons could account for the particular location of these neurons in the basal forebrain, since in man the elongated olfactory stalk is located below the forebrain instead of being rostral to it as in macrosmatics. This assumption is supported by the description of DA neurons in the most anteromedial part of the frontal cortex in the immature rodent brain (Specht et al., '81; Verney et al., '82).
As such, the "forebrain" DA neurons could be supposed to have only a local function, as was demonstrated for the periglomerular and bitufted DA cells in the rodent olfactory bulb (reviewed in Halasz and Sheperd, '83). It is noteworthy that in spite of the frequent atrophy of primary olfactory neurons in man, these olfactory DA cells do not appear to be atrophic and continue to express TH immunoreactivity. Involvement of these DA neurons in degenerative diseases would be of interest to investigate in view of the recently described specific neuronal alteration of olfactory nuclei in Alzheimer's disease (Esiri and Wilcock, '84).

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 31
Physiological considerations
The partition of the septum into two main anatomical and functional compartments, the medial septum providing the septohippocampal pathway and the lateral septum re- ceiving the bulk of the hippocampal formation input, has been well documented in rodents (Swanson and Cowan, '79) and primates (Siege1 et al., '75; Amaral and Cowan, '80). It corresponds, furthermore, to specific and highly organized aminergic, cholinergic, and peptidergic connections. From the numerous data accumulated, some are relevant to hu- man physiopathological problems.
Activation of the septohippocampal pathway has been repeatedly implied in learning processes (Galey et al., '83; Mitchell et al., '83). Several lines of evidence suggest that both DA and NA innervations regulate the cholinergic sep- tohippocampal neurons, the dopaminergic input exerting an inhibitory influence (Robinson et al., '79; Costa et al., '83) while noradrenergic innervation may have excitatory effects (Costa et al., '83). The presence of NA terminals in the medial septal and diagonal band nuclei supports a direct action of this amine on cholinergic neurons, whereas DA terminals in the lateral septum may influence the me- dial septal neurons indirectly by acting on inhibitory GABA interneurons (Costa et al., '83). Such functional interactions with aminergic systems could be relevant to the pathophys- iology of dementia. Indeed in demented patients with Alz- heimer's or Parkinson's disease, both cholinergic and noradrenergic deficits have been demonstrated (reviewed in Berger, '84). It may be hypothesized that loss of the NA input on the altered cholinergic neurons of the septohippo- campal pathway could impair normal function of the re- maining neurons.
In the lateral septal division, anatomical studies compar- ing the laminar distribution of aminergic and peptidergic neurons in the rodent have suggested that DA terminals may interact specifically with enkephalinergic neurons (Gall and Moore, '84). Relationships between enkephali- nergic and dopaminergic neural systems, at the septal level, are also suggested by pharmacological studies (Collu et al., '80; Megens and Cools, '83). Such interactions would be all the more interesting as the DA neurons of the ventral tegmentum, known to project to the lateral septum, are themselves under enkephalinergic control (Johnson et al., '80; Gaspar et al., '83). The existence of such anatomical relationships is presently under investigation in man.
The DA input would also be in a position to regulate the putative glutamatergic terminals of the hippocamposeptal pathway (Zazcek et al., '79; Joels and Urban, '841, although such interactions have not yet been reported to our knowl- edge. Interestingly, the topography of the DA terminals in the lateral septum of man overlaps with the zone of d e r - ents from the prefrontal cortex in rhesus monkey (Tanaka and Goldman, '76). The prefrontal cortex is also a site of DA projections not only in rodents but also in primates (Bjorklund et al., '78; Brown et al., '79) and in man (Scatton et al., '83). This convergence implies that both the origin and the terminal of the prefrontoseptal pathway might be modulated by DA inputs. This also seems to be the case for the hippocampo-accumbens pathway (Verney et al., '851, at least in rodents.
A last point of interest to consider is that of the origin of the septal DA innervation in man. It has been extensively analysed in the rat (see review in Lindvall and Bjorklund, '83); in studies where specific revelation of the amine could
be combined to anatomical tracing methods, the DA neu- rons projecting to the lateral septum were localized in the most medial and ventral portions of the ventral tegmental area. Collateralization of these DA neurons to other termi- nal fields such as the cortex or nucleus accumbens could only rarely be demonstrated directly (Deniau et al., '80). In primate, limited hodological data have demonstrated septal afferents from nucleus paranigralis and medial substantia nigra (Krayniak et al., '81). In man, biochemical analysis of pathological cases with degeneration of the DA systems may replace, to a certain extent, direct hodological data, by comparing the levels of DA and its metabolites in both the nuclei of origin and the presumed areas of terminal fields (Javoy-Agid et al., '83). In this respect, analysis of cases with supranuclear palsy might provide relevant informa- tion concerning the origin of the septal DA innervation, since in this disease, the DA nigrostriatal system is se- verely affected while the DA mesocortical system appears to be spared (Ruberg et al., '85).
ACKNOWLEDGMENTS We are indebted to Prof. Agid, Dr. Callard, Prof. Chom-
ette, Prof. Hauw and Dr. Josset for providing brain samples and to Mrs. C. Stone for improving the English of the manuscript.
LITERATURE CITED Amaral, D.G., and W.N. Cowan (1980) Subcortical afferents to the hippo-
campal formation in the monkey. J. Comp. Neurol. 189t573-591. Andy, O.J., and H. Stephan (1968) The septum in the human brain. J. Comp.
Neurol. 133:383-409. Andy, O.J., and H. Stephan (1976) Septum development in primates. In J.F.
De France (ed): The Septa1 Nuclei. New York Plenum Press, pp. 3-36. Arendt, T., V. Bigl, A. Arendt, and A. Tennstedt (1983) Loss of neurons in
the nucleus basalis of Meynert in Alzheimer's disease, Paralysis agitans and Korsakoffs disease. Acta Neuropathol. (Berl.) 61t101-108.
Berger, B., U. DiPorzio, M.C. Daguet, M. Gay, A. Vigny, J. Glowinski, and A. Prochiantz (1982) Long term development of mesencephalic neurons of mouse embryos in dissociated primary cultures: Morphological and histochemical characteristics. Neuroscience 7t193-205.
Berger, B., C. Verney, and C. Alvarez (1983) Postnatal development of catecholaminergic pericellulax baskets in the septal area of Sprague- Dawley rats. In: Abstracts of 5th CA Symposium, Giiteborg 1983. New York Pergamon Press, p. 79.
Berger, B. (1984) Anomalies des neurotransmetteurs dam la maladie d'Alz- heimer. Rev. Neurol. 140:539-552.
Bjorklund, A., I. Divac, and 0. Lindvall (1978) Regional distribution of catecholamines in monkey cerebral cortex, evidence for a dopaminergic innervation of the primate prefrontal cortex. Neurosci. Lett. 7t115-119.
Bowden, D.M., D.C. German, and W.D. Poynter (1978) An autoradiographic semistereotaxic mapping of major projections from locus coeruleus and adjacent nuclei in macaca mulatta. Brain Res. 145.257-276.
Brockhaus, H. (1942a) Zur feineren Anatomie des Septum und des Striatum. J. Psychol. Neurol. 51:l-56.
Brockhaus, H. (1942b) Vergleichend-anatomische Untersuchugen iiber den Basalkernkomplex. J. Psychol. Neurol. 51:57-95.
Brown, R.M., A.M. Crane, and P.S. Goldman (1979) Regional distribution of monoamines in the cerebral cortex and subcortical structures of the rhesus monkey: Concentrations and in vivo synthesis rates. Brain Res. 168t133-150.
Collu, R., E. Stefanini, F. Vernaleone, A.M. Marchisio, P. Devoto, and A. Argiolas (1980) Biochemical characterization of the dopaminergic inner- vation of the rat septum. Life Sci. 26t1665-1673.
Costa, E., P. Panula, H.K. Thompson, and D.L. Cheney (1983) The transsy- naptic regulation of the septal-hippocampal cholinergic neurons. Life Sci. 32165-179.
Crosby, E.C., and T. Humphrey (1941) Studies of the vertebrate telencepha- lon. 11. The nuclear pattern of the anterior olfactory nucleus, tubercu- lum olfactorium, and the amygdaloid complex in adult man. J. Comp. Neurol. 74:309-352.

32 P. GASPAR ET AL.
Crosby, E.C., T. Humphrey, and E.W. Lauer (1962) Correlative Anatomy of the Nervous System. New York MacMillan.
Deniau, J.M., A.M. Thierry, and J. Feger (1980) Electrophysiological iden- tification of mesencephalic ventro-medial tegmental (VMT) neurons pro- jecting to the frontal cortex, septum and nucleus accumbens. Brain Res. 189:315-326.
Eliott, F.A. (1969) The corpus callosum, cingulate gyrus, septum pelluci- dum, septal area and fornix. In P.J. Vinken and G.W. Bruyn (eds): Handbook of Clinical Neurology. Amsterdam: North Holland Publish- ing Co. Vol. 11, pp. 758-776.
Esiri, M., and G.K. Wilcock (1984) The olfactory bulbs in Alzheimer’s dis- ease. J. Neurol. Neurosurg. Psychiatry 47:56-60.
Fallon, J.H., S.E. Loughlin, and C.E. Ribak (1983) The islands of Calleja complex of ra t basal forebrain. III. Histochemical evidence for a striato- pallidal system. 3. Comp. Neurol. 218t91-120.
Farley, I.J., and 0. Hornykiewicz (1977) Noradrenaline distribution in sub- cortical areas ofshe human brain. Brain Res. 12653-62.
Farley, I.J., K.S. Price, and 0. Hornykiewicz (1977) Dopamine in the limbic regions of the human brain: Normal and abnormal. In E. Costa and G.L. Gesa (eds): Advences in Biochemical Psychopharmacology. New York: Raven Press, pp. 333-349.
Fuxe, K. (1965) Evidence for the existence of monoamine neurons in the central nervous system. IV. Distribution of monoamine nerve terminals in the central nervous system. Acta Physiol. Scand. 64:39-85.
Galey, D., Y. Jeantet, C. Destrade, and R. JafTard (1983) Facilitation of memory consolidation by post-training electrical stimulation of the me- dial septal nucleus: Is i t mediated by changes in rhythmic slow activity? Behav. Neural. Biol. 38240-250.
Gall, C., and R.Y. ‘Moore (1984) Distribution of enkephalin, substance P, tyrosine hydroxylase, and 5-hydroxytryptamine imrnunoreactivity in the septal region of the rat. J. Comp. Neurol. 255.212-227.
Gaspar, P., B. Berger, M. Hamon, F. Cesselin, A. Vigny, F. Javoy-Agid, and Y. Agid (1983) Tyrosine hydroxylase and methionine-enkephalin in the human mesencephalon. J. Neurol. Sci. 58247-267.
German, D.C., D.S. Schlussberg, and D.J. Woodward (1983) Three-dimen- sional reconstruction of midbrain dopaminergic neuronal populations: From mouse to man. J. Neural Transm. 57.243-254,
Graybiel, A.M., and C.W. Ragsdale (1983) Biochemical anatomy of the striatum. In P.C. Emson (ed): Chemical Neuroanatomy. New York Raven Press, pp. 427-504.
Greenfield, S. (1984) Acetylcholinesterase may have novel functions in the brain. TINS 7;364-368.
Grote, S.S., S.G. Moses, E. Robins, R.W. Hudgens, and A.B. Croninger (1974) A study of selected catecholamine metabolizing enzymes: A comparison of depressive suicides and alcoholic suicides with controls. J. Neuro- chem. 23:791-802.
Halasz, N., and G.M. Sheperd (1983) Neurochemistry of the vertebrate olfactory bulb. Neuroscience 10:579-619.
Hartman, B.K. (1973) Immunofluorescence of dopamine-0-hydroxylase, ap- plication of improved methodology to the localization of the peripheral and central noradrenergic nervous system. J. Histochem. Cytochem. 21:312-332.
Hartman, B.K. (1974) Localization of noradrenergic neuron systems in hu- man brain. J. Psychiatr. Res. 11:283-288.
Hedreen, J.C., R.G. Struble, P.J. Whitehouse, and D.L. Price (1984) Topog- raphy of the magnocellular basal forebrain in the human brain. J. Neuropathol. 4.31-21.
Henke, H., and W. Lang (1983) Cholinergic enzymes in neocortex, hippocam- pus and basal forebrain of non-neurological and senile dementia of Alzheimer-type patients. Brain Res. 2675281-291.
Holstege, G., L. Meiners, and K. Tan (1984) Efferent connections of the bed nucleus of the stria terminalis (BNST). An HRP and autoradiographic tracing study. Neurosci. Lett. [Suppl.]I8:S62.
Herkenham, M., S.M. Edley, and J. Stuart (1984) Cell clusters in the nucleus accumbens of the rat and the mosaic relationship of opiate receptors acetylcholinesterase and subcortical afferent terminations. Neurosci- ence 11:561-595.
Hokfelt, T., K. Fuxe, M. Goldstein, and T.H. Joh (1973) Irnmunohistochemi- cal localization of the three catecholamine synthesizing enzymes: As- pects on methodology. Histochemie 33.231-254.
Hokfelt, T., K. Fuxe, M. Goldstein, and 0. Johansson (1974) Immunohisto- chemical evidence for the existence of adrenaline neurons in the rat brain. Brain Res. 66235-251.
Hokfelt, T., 0. Johansson, K. Fuxe, M. Goldstein, and D. Par (1977) Immu- nohistochemical studies on the localization and distribution of mono-
amine neuron systems in the rat brain. 11. Tyrosine hydroxylase in the telencephalon. Med. Biol. 552-40.
Ishikawa, M., and C. Tanaka (1977) Morphological organization of catechol- amine terminals in the diencephalon of the rhesus monkey. Brain Res. 119:43-55.
Javoy-Agid, F., M. Ruberg, H. Taquet, B. Bokobza, Y. Agid, P. Gaspar, B. Berger, J. Nguyen-Legros, C. Alvarez, F. Gray, B. Scatton, and L. Rou- quier (1983) Biochemical neuropathology of Parkinson disease. Adv. Neurol. 40: 189-199.
Joels, M., and I.J.A. Urban (1984) Electrophysiological and pharmacological evidence in favor of amino acid neurotransmission in fimbria-fornix fibres innervating the lateral septal complex of rats. Exp. Brain Res. 54:455462.
Johnson, R.P., M. Sar, and W.E. Stumpf (1980) A topographic localization of enkephalin on dopamine neurons of the rat substantia nigra and ventral tegmental area demonstrated by combined histofluorescence immuno- cytochemistry. Brain Res. 194:566-571.
Karnovsky, M.J., and L. Roots (1964) A direct-coloring thiocholine method for cholinesterases. J. Histochem. Cytochem. 12:219-221.
Kelley, A.E., and V.B. Domesick (1982) The distribution of the projection from the hippocampal formation to the nucleus accumbens in the r a t An anterograde and retrograde-horseradish peroxidase study. Neurosci- ence 7.2321-2335.
Kohler, C., and L.G. Eriksson (1984) An immunohistochemical study of somatostatin and neurotensin positive neurons in the septal nuclei of the ra t brain. Anat. Embryol. 17Orl-10.
Kohler, C., B.J. Everitt, J. Pearson, and M. Goldstein (1983) Immunohisto- chemical evidence for a new group of catecholamine-containing neurons in the basal forebrain of the monkey. Neurosci. Lett. 37:161-166.
Krayniak, P.F., R.C. Meibach, and A. Siege1 (1981) Origin of brain stem and temporal cortical afferent fibers to the septal region in the squirrel monkey. Exp. Neurol. 72113-121.
Krettek, J.G., and J.L. Price (1978) Amygdaloid projections to subcortical structures within the basal forebrain and brain stem in the rat and cat. J. Comp. Neurol. 178:225-254.
Kwak, S., J. Kanazawa, H. Sugita, and Y. Toyokura (1984) Tridimensional distribution of markers of neurotransmitters within the “accumbens area” of normal human brains. Neuroscience, 13r717-733.
Laine, E., and S. Blond (1980) Tumeurs trigono-septales. Neurochirurgie 26947-278.
Lauer, E.W. (1945) The nuclear pattern and fiber connections of certain basal telencephalic centers in the macaque. J. Comp. Neurol. 82,215- 254.
Levitt, P., P. Rakic, and P. Goldman-Rakic (1984) Region-specific distribu- tion of catecholamine afferents in primate cerebral cortex: A fluores- cence histochemical analysis. J. Comp. Neurol. 227.23-36.
Lew, J.Y., Y. Matsumo, J. Pearson, M. Goldstein, T. Hokfelt, and K. Fuxe (1977) Localization and characterization of phenylethano1amine-N- methyl transferase in the brain of various mammalian species. Brain Res. I 19: 199-210.
Lindquist, G., and G. Norlen (1966) Korsakoffs syndrome after operation of the anterior communicating artery. Acta Psychiatr. Scand. 4224-34.
Lindvall, 0. (1977) Combined visualization of central catecholamine and acetylcholinesterase-containing neurons: Application of the glyoxylic acid and thiocholine histochemical methods to the same vibratome sec- tion. 50:191-196.
Lindvall, O., and U. Stenevi (1978) Dopamine and noradrenaline neurons projecting to the septal area in the rat. Cell Tissue Res. 190:383-407.
Lindvall, O., and A. Bjorklund (1983) Dopamine- and norepinephrinecon- taining neuron systems: Their anatomy in the rat brain. In P.C. Emson (ed): Chemical Neuroanatomy. New York: Raven Press, pp. 229-255.
Mackay, A.V.P., C.M. Yates, A. Wright, P. Hamilton, and P. Davies (1978) Regional distribution of monoamines and their distribution in the hu- man brain. J. Neurochem. 30:841-848.
McGeer, P.L., and E.G. McGeer (1976) Enzymes associated with the metab- olism of catecholamines, acetylcholine and GABA in human controls and patients with Parkinson’s disease and Huntington’s chorea. J. Neu- rochem. 26:65-76.
McKinney, M., J.T. Coyle, and J.C. Hedreen (1983) Topographic analysis of the innervation of ra t neocortex and hippocampus by the basal forebrain cholinergic system. J. Comp. Neurol. 217:103-121.
Mesulam, M.M., E.J. Mufson, A.I. Levey, and B.H. Wainer (1983a) Cholin- ergic innervation of cortex by the basal forebrain: Cytochemistry and cortical connections of the septal area, diagonal band nuclei, nucleus basalis (Sn and hypothalamus in the rhesus monkey. J. Comp. Neurol. 214:170-197.

CATECHOLAMINERGIC INNERVATION OF HUMAN SEPTUM 33
Mesulam, M.M., E.J. Mufson, B.H. Wainer, and A.I. Levey (1983b) Central cholinergic pathways in the rat: An overview based on an alternative nomenclature (Chl-Ch6). Neuroscience 1Or1185-1201.
Mesulam, M.M., E.J. Mufson, A.I. Levey, and B.H. Wainer (1984) Atlas of cholinergic neurons in the forebrain and upper brainstem of the ma- caque based on monoclonal choline acetyltransferase immunohisto- chemistry and acetylcholinesterase histochemistry. Neuroscience 121669-687.
Megens, A.P.A., and A.R. Cools (1983) Involvement of the septum in central dopamine-acetylcholine interactions in morphine treated rats. Pharma- col. Biochem. Behav. 181761-768.
Meibach, R., and A. Siegel (1977) Efferent connections of the hippocampal formation in the rat. Brain Res. 124t197-224.
Mitchell, S.J., J.N.P. Rawlins, 0. Steward, and D.S. Olton (1982) Medial septal area lesions disrupt 0 rhythm and cholinergic staining in medial entorhinal cortex and produce impaired radial arm maze behavior in rats. J. Neurosci. 2r292-302.
Moore, R.Y. (1978) Catecholamine innervation of the basal forebrain: I. The septal area. J. Comp. Neurol. 1771665-683.
Nauta, W.J.H., and W. Haymaker (1969) Hypothalamic nuclei and fiber connections. In W. Haymaker, E. Anderson, and W.J.H. Nauta (eds): The Hypothalamus. Springfield: C.C. Thomas, pp. 136-209.
Nobin, A., and A. Bjorklund (1973) Topography of the monoamine neuron systems in the human brain as revealed in fetuses. Acta Physiol. Scand. [Suppl.] 388:l-40.
Nyberg, P.R., Adolfsson, N.E. Anden, and B. Winblad (1982) Concentrations of dopamine and noradrenaline in some limbic and related regions of the human brain. Acta Neurol. Scand. 651267-273.
O'Connor, D.T., R.P. Frigon, and R.A. Stone (1979) Human pheochromocy- toma dopamine-0-hydroxylase: Purification and molecular parameters of the tetramer. Mol. Pharmacol. 16:529-538.
Onteniente, B., M. Geffard, and A. Calas (1984) Ultrastructural immunocy- tochemical study of the dopaminergic innervation of the rat lateral septum with anti-dopamine antibodies. Neuroscience 13~385-393.
Pearson, J., M. Goldstein, and L. Brandeis (1979) Tyrosine hydroxylase immunohistochemistry in human brain. Brain Res. 165333-337.
Pearson, J., M. Goldstein, K. Markey, and L. Brandeis (1983) Human brain- stem catecholamine neuronal anatomy as indicated by immunocyto- chemistry with antibodies to tyrosine hydroxylase. Neuroscience 813- 32.
Pickel, V.M., T.H. Joh, P.M. Field, C.G. Becker, and D.J. Reis (1975) Cellular localization of tyrosine hydroxylase by immunohistochemistry. J. His- tochem. Cytochem. 23:l-12.
Price, K.S., I.J. Farley, and 0. Hornyckiewicz (1978) Neurochemistry of Parkinson's disease: Relation between striatal and limbic dopamine. Adv. Biochem. Psychopharmacol. 19:293-300.
Puymirat, J., F. Javoy-Agid, P. Gaspar, A. Ploska, A. Prochiantz, and Y. Agid (1979) Post-mortem stability and storage in the cold of brain en- zymes. J. Neurochem. 32449454.
Robinson, S.E., D. Malthe-Sorenssen, P.L. Wood, and J. Commissiong (1979) Dopaminergic control of the septal-hippocampal pathway. J. Pharmacol. Exp. Ther. 208:476-479.
Ross, C.A., D.A. Ruggiero, M.P. Meeley, D.H. Park, T.H. Joh, and D.J. Reis (1984) A new group of neurons in hypothalamus containing phenyl- ethanolamine N-methyl transferase (PNMT) but not tyrosine hydroxyl- ase. Brain Res. 306349-353.
Ruberg, M., F. Javoy-Agid, N. Le Balc'h, E. Hirsch, B. Scatton, R. Lheureux, J.J. Hauw, C. Duyckaerts, F. Gray, A. Morel-Maroger, A. Rascol, M. Serdaru, and Y. Agid (1985) Dopaminergic and cholinergic lesions in
progressive supranuclear palsy. Ann. Neurol. (in Dress). Scatton, B., F. Javoy-Agid, L. Rouquier, B. Dubois, and Y. Agid (1983)
Reduction of cortical dopamine, noradrenaline, serotonin and their me- tabolite in Parkinson's disease. Brain Res. 275:321-328.
Schmidt, R.H., and R.K. Bhatnagar (1979) Assessment of the effects of neonatal subcutaneous 6-hydroxy-dopamine or noradrenergic and dopa- minergic innervation of the cerebral cortex. Brain Res. 166:309-320.
Schofield, S.P.M., and A.F. Dixson (1982) Distribution of catecholamine and indoleamine neurons in the brain of the common marmoset (Callithrix Jacchus). J. Anat. 1341315-338.
Siegel, A., S. Ohgami, and H. Edinger (1975) Projections of the hippocampus to the septal area in the squirrel monkey. Brain Res. 99:247-260.
Specht, L.A., V.M. Pickel, H.J. Tong, and D.J. Reis (1981) Light microscopic immunocytochemical localization of tyrosine hydroxylase in prenatal rat brain: II. Late ontogeny. J. Comp. Neurol. 199r255-276.
Stevens, J.R. (1979) Schizophrenia and dopamine regulation in the mesolim- bic system. TINS, 21102-105.
Swanson, L.W., and B.K. Hartman (1975) The central adrenergic system. An immunofluorescence study of the location of cell bodies and their efferent connections in the rat utilizing dopamine-6-hydroxylase as a marker. J. Comp. Neurol. 163r467-506.
Swanson, L.W., and W.M. Cowan (1979) The connections of the septal region in the rat. J. Comp. Neurol. 186r621-656.
Tanaka, D., and P.S. Goldman (1976) Silver and autoradiographic evidence for a projection from the principal sulcus to the septum in the rhesus monkey. Brain Res. 103r535-540.
Taren, J.A. (1965) Anatomical pathways related to the clinical findings in aneurysms of the anterior communicating artery. J. Neurol. Neurosurg. Psychiatry 281228-234.
Trocexicz, J., 0. Kazuhiro, and T. Nagatsu (1982) Distribution of noradren- aline-N-methyltransferase (phenylethanolamine N-methyltransferase) in rat and human brains: Application of a new assay with electrochemical detection. Neurosci. Lett. [Suppl.] 9:S80.
Van der Guten, J., M. Paklovits, H.L.M. Wijnen, and D.H.G. Versteeg (1976) Regional distribution of adrenaline in rat brain. Brain Res. 107r171- 175.
Verney, C., B. Berger, J. Adrien, A. Vigny, and M. Gay (1982) Development of the dopaminergic innervation of the rat cerebral cortex. A light microscopic immunocytochemical study using anti-tyrosine hydroxylase antibodies. Dev. Brain Res. 541-52.
Verney, C., M. Baulac, B. Berger, C. Alvarez, A. Vigny, and B. Helle (1985) Morphological evidence for a dopaminergic terminal field in the hippo- campal formation of young and adult rat. Neuroscience 4:1039-1052.
Vigny, A,, and J.P. Henry (1981) Bovine adrenal tyrosine hydroxylase: Comparative study of native and proteolyzed enzyme and their interac- tion with anions. J. Neurochem. 36r483-489.
Vogel, W.H., V. Orfei, and B. Century (1969) Activities of enzymes involved in the formation and destruction of biogenic amines in various areas of human brain. J. Pharmacol. Exp. Ther. 1651196-203.
Von Economo, C.V. (1925) L'architecture cellulaire normale de l'ecorce ce- rebrale. Paris: Masson.
Weller, K.L., and D.A. Smith (1982) Afferent connections to the bed nucleus of the stria terminalis. Brain Res. 232r255-270.
Woodhams, P.L., G.W. Roberts, J.M. Polak, and T.J. Crow (1983) Distribu- tion of neuropeptides in the limbic system of the r a t The bed nucleus of the stria terminalis, septum and preoptic area. Neuroscience 8:677-703.
Zazcek, R., J.C. Hedreen, and J.T. Coyle (19791 Evidence for a hippocampal septal glutamatergic pathway in the rat. Exp. Neurol. 65145-156.
Copyright © 2022 FDOKUMEN




















