
The mysterious Spotted Green Pigeon and its relation to the Dodo and its kindred

Ž .Molecular Brain Research 50 1997 190–196
Research report
Immunocytochemical identification of pinopsin in pineal glands ofchicken and pigeon
Toshiyuki Okano a, Yoko Takanaka a, Atsushi Nakamura a, Kanjun Hirunagi b,Akihito Adachi c, Shizufumi Ebihara c, Yoshitaka Fukada a,)
a Department of Biophysics and Biochemistry, Graduate School of Science, The UniÕersity of Tokyo, Tokyo 113, Japanb Department of Animal Morphology and Function, School of Agricultural Sciences, Nagoya UniÕersity, Nagoya 464-01, Japan
c Department of Animal Physiology, School of Agricultural Sciences, Nagoya UniÕersity, Nagoya 464-01, Japan
Accepted 6 May 1997
Abstract
Pinopsin is a blue-sensitive photoreceptive molecule possibly involved in photic entrainment of the circadian pacemaker in the chickenpineal gland. To characterize pinopsin as a circadian photoreceptor, antibodies were raised against the C-terminal portion of pinopsin. Asexpected from the divergence of the amino acid sequence of this region, the resultant antibody cross-reacted with neither chicken
Ž .rhodopsin nor red-sensitive cone pigment chicken red . In Western blot analysis, the antibody stained a single band of 42-kDa protein inŽ .a detergent-extract of chicken pineal membranes, suggesting that pinopsin calculated molecular weight, 38 187 might be glycosylated
andror palmitoylated. Immunocytochemical examination of pineal sections of the chicken and the pigeon with this antibody revealedstrong positive images for most of the membrane structures in the lumen of the follicles. This antibody also stained string- andbulb-shaped structures of the chicken parafollicular cells, the morphology of which resembles those of retinal photoreceptor cells. Incontrast to the predominant distribution of pinopsin, a monoclonal antibody specific for chicken red stained a smaller number ofmembrane structures in the lumen of chicken pineal follicles. These results strongly suggest that the chicken pineal gland contains at least
Ž . Ž .two types of photoreceptive molecules, pinopsin major and chicken red minor . We show that the former molecule is localized inparafollicular pinealocytes and in the outer segments of pinealocytes that make contact with the follicular lumen. q 1997 Elsevier ScienceB.V.
Keywords: Pinopsin; Immunocytochemistry; Circadian clock; Chicken red; Pineal gland; Chicken
1. Introduction
Besides retinal photopigments, such as rhodopsin andcone pigments, many vertebrates have extra-retinal pho-
w xtoreceptive molecules 4–8,10,23,24 . In the case of birds,a pineal photoreceptive molecule has been implicated inthe photic entrainment of the circadian pacemaker which
w xcontrols the rhythmic production of melatonin 4,5 .
w Ž . xAbbreviations: CHAPS, 3- 3- cholamidopropyl dimethyl ammonio -1-propanesulfonate; ELISA, enzyme-linked immunosorbent assay; IPTG,isopropyl-b-D-thiogalactopyranoside; MBP, maltose-binding protein;
Ž .MBP-p7 or p9 , a fusion protein composed of maltose-binding proteinŽ .and pinopsin p7 or p9 fragment; MBP-bgal, a fusion protein composed
of maltose-binding protein and the amino-terminal region of b-galacto-sidase
) Corresponding author. Fax: q81 3 5802 8871; E-mail:[email protected]
Ž .Hydroxyindole-O-methyltransferase HIOMT and sero-Ž .tonin N-acetyltransferase NAT are enzymes involved in
melatonin biosynthesis. By using an antibody to HIOMTw x w x3 and a cRNA probe for NAT 2 , two types of pinealo-cytes have been detected in the chicken pineal gland: one
Žis the follicular pinealocytes termed ‘modified photore-w x.ceptor cells’ in 3 and the other is distributed at the
Žperiphery of the follicles termed ‘parafollicular pinealo-w x.cytes’ in 3 . Apical portions of the modified photorecep-
tor cells face the follicular lumina, into which modifiedw xcilia and cytoplasmic processes are projected 15 . Some of
these cells were immunocytochemically stained by eitherw xanti-rhodopsin or anti-chicken red antibodies 1,13 . On the
other hand, the parafollicular pinealocytes were stained bynone of the anti-opsin antibodies. Thus, the parafollicularpinealocytes were postulated to respond to neurotrans-mitters to synthesize and secrete melatonin. It has been
0169-328Xr97r$17.00 q 1997 Elsevier Science B.V. All rights reserved.

( )T. Okano et al.rMolecular Brain Research 50 1997 190–196 191
Žunclear whether the opsin-negative pinealocytes para-follicular pinealocytes and some of the modified photore-
.ceptors have any photoreceptive molecule.To investigate the molecular mechanism of the photic
input pathway to the circadian pacemaker in the chickenpineal gland, we previously cloned a blue-light sensitivemolecule belonging to a novel class of opsins from a
w xchicken pineal cDNA library 19 . The pineal opsin, whichw xwe named ‘pinopsin’ after pineal opsin 19 , is transcribed
in the pineal gland, but not in other tissues. In the presentstudy, we prepared antibodies highly specific for pinopsinin order to allow immunocytochemical localization. Theantibody strongly reacted with membranes in the parafol-licular zone in addition to the luminal surface of thefollicles, suggesting the presence of pinopsin in modifiedphotoreceptor cells and in the parafollicular pinealocytes.
2. Materials and methods
2.1. Preparation of fusion proteins
ŽBsrFI–Hind III and BbsI–Hind III fragments 456 and396 bp, respectively, corresponding to the C-terminal re-
.gion of the protein of the pinopsin cDNA were blunt-endedand subcloned into the XmnI site of a prokaryote expres-
Ž .sion vector pMAL-c2 New England Biolabs to obtainrecombinant plasmids termed pMAL-p7 and pMAL-p9,respectively. Since the XmnI site of pMAL-c2 is locatedjust downstream of the mal E gene for maltose-binding
Ž .protein MBP , pMAL-p7 and pMAL-p9 encode MBPsŽ . Ž .fused with 75 p7 region and 55 p9 region amino acid
Ž .residues of pinopsin, respectively Fig. 1 . The BL21 strainof Escherichia coli was transformed either by the recom-
Žbinant plasmids or by a control plasmid pMAL-c2 without.insert which encodes MBP fused with the N-terminal
Žregion of b-galactosidase. The fusion proteins MBP-p7,.MBP-p9 and MBP-bgal expressed in BL21 cells after 2 h
Ž .of IPTG 0.3 mM induction were extracted from the cellsand then purified by amylose affinity column chromatogra-phy according to a protocol recommended by the manufac-
Ž .turer New England Biolabs . The MBP-fusion proteinspurified to near homogeneity were dialyzed several times
Žagainst phosphate-buffered saline PBS; 10 mM Na-phos-.phate, 140 mM NaCl, and 10 mM KCl pH 7.4 just before
immunization.
2.2. Immunization of mice and purification of antisera
Four 6-week-old female Balbrc mice were injectedintraperitoneally with 50 mg of the purified fusion protein,MBP-p7 or MBP-p9 in 10 mM PBS emulsified in com-
Ž .plete Freund’s adjuvant Difco . Each mouse was boostedintraperitoneally with 50 mg of the same antigen in 10 mMPBS at day 21, and bled at day 28.
A control fusion protein, MBP-bgal was used to re-move anti-MBP antibodies from the resultant sera. Briefly,
Ž .purified MBP-bgal 1.67 mg was immobilized to HiTrapŽ .NHS-activated Sepharose 1 ml, Pharmacia according to a
protocol supplied by the manufacturer. Sera obtained fromthe immunized mice were applied to the column and theflow-through fractions were collected. The column wassuccessively washed with 100 mM sodium-citrate bufferpH 3.0 and 50 mM Tris-HCl buffer pH 8.6 to wash outanti-MBP antibodies bound to the column. Then, the col-umn was re-equilibrated with 10 mM PBS and the firstflow-through fraction was loaded again to collect a secondflow-through fraction. These procedures were repeateduntil the flow-through fraction did not react with MBP in
Ž .ELISA see below . The final flow-through fraction ob-tained from anti-MBP-p9 antiserum was applied to a Hi-
Ž .Trap Protein G-Sepharose column Pharmacia pre-equi-librated with 10 mM PBS. The antibodies bound to thecolumn were eluted with 100 mM sodium-citrate bufferpH 3.0, and neutralized immediately by adding 2 M Tris-HCl pH 9.0, followed by dialysis against 10 mM PBS.
Ž .In an enzyme-linked immunosorbent assay ELISA ,Ž0.1 mg of the purified fusion proteins MBP-p7, MBP-p9,
.or MBP-bgal in 100 ml of 10 mM PBS was adsorbed toŽ .each well of a microtitre plate MaxiSorp; Nunc at 48C
overnight. Subsequent procedures were performed accord-w xing to the method described previously 21 .
2.3. Western blot analysis
Chicken red and rhodopsin were purified from freshlyw xdissected chicken retinas 17 . Proteins separated by
Ž . w xSDS–polyacrylamide 10% gel electrophoresis 12 wereelectrophoretically transferred to a polyvinylidene difluo-
Ž .ride membrane Immobilon transfer membrane, Millipore .After the transfer, the blotted membrane was incubated in
Ž .1% skim milk Difco dissolved in Tris-buffered salineŽTBS; 50 mM Tris-HCl, 200 mM NaCl, and 1 mM MgCl2
.pH 7.4 for 1 h at 378C to block the remaining bindingsites, followed by incubation with a test antibody solutionin 1% skim milkrTBS at 48C overnight or at 378C for 1.5h. The membrane was then washed with 1% skimmilkrTBS, and incubated with 0.5 mgrml of peroxidase-
Žlabelled goat anti-mouse IgG Kirkegaard and Perry Labo-.ratories in 1% skim milkrTBS for 1–2 h at 378C. After
successive washing of the membrane with 0.05% Tween20rTBS and TBS, positive bands were detected using a
Ž .chemiluminescence reagent Renaissance, DuPont NEN .Alternatively, alkaline phosphatase-labelled horse anti-
Ž .mouse IgG 1 : 1000 dilution, New England Biolabs wasused as the second antibody. In this case, the positivebands were detected using CDP-Star chemiluminescence
Ž .substrate New England Biolabs .
2.4. Immunocytochemistry
2.4.1. Staining of pinopsinTwo-month-old chickens and adult pigeons were used
for the experiments. Anaesthetized animals were perfused
( )T. Okano et al.rMolecular Brain Research 50 1997 190–196192
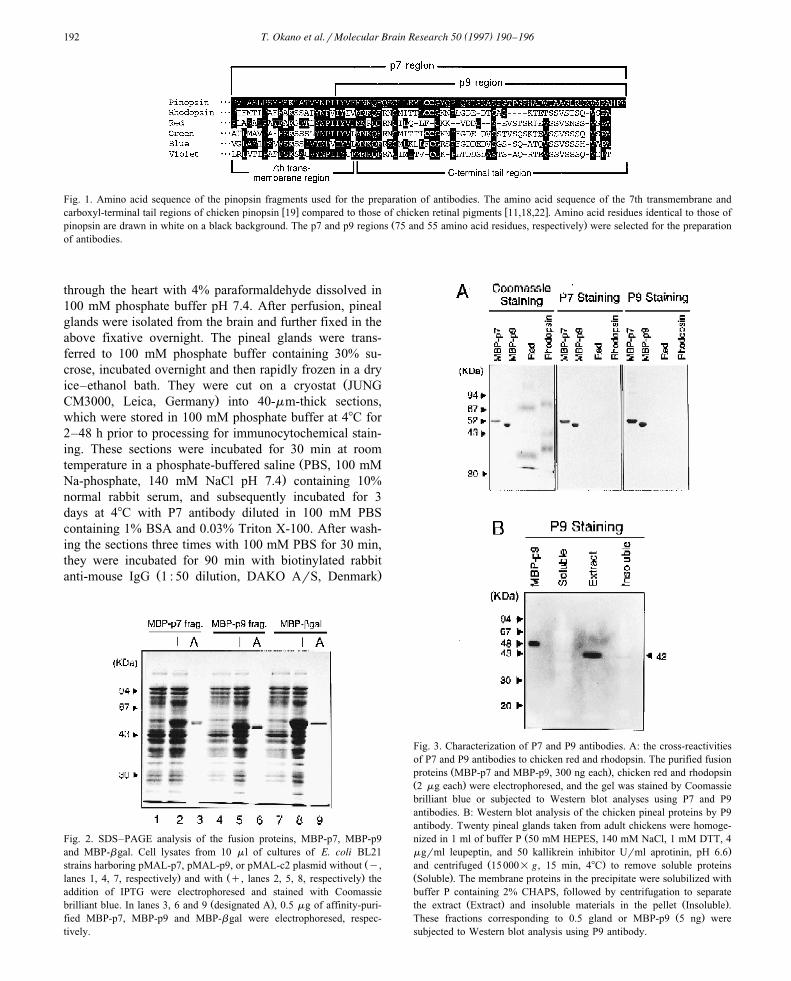
Fig. 1. Amino acid sequence of the pinopsin fragments used for the preparation of antibodies. The amino acid sequence of the 7th transmembrane andw x w xcarboxyl-terminal tail regions of chicken pinopsin 19 compared to those of chicken retinal pigments 11,18,22 . Amino acid residues identical to those of
Ž .pinopsin are drawn in white on a black background. The p7 and p9 regions 75 and 55 amino acid residues, respectively were selected for the preparationof antibodies.
through the heart with 4% paraformaldehyde dissolved in100 mM phosphate buffer pH 7.4. After perfusion, pinealglands were isolated from the brain and further fixed in theabove fixative overnight. The pineal glands were trans-ferred to 100 mM phosphate buffer containing 30% su-crose, incubated overnight and then rapidly frozen in a dry
Žice–ethanol bath. They were cut on a cryostat JUNG.CM3000, Leica, Germany into 40-mm-thick sections,
which were stored in 100 mM phosphate buffer at 48C for2–48 h prior to processing for immunocytochemical stain-ing. These sections were incubated for 30 min at room
Žtemperature in a phosphate-buffered saline PBS, 100 mM.Na-phosphate, 140 mM NaCl pH 7.4 containing 10%
normal rabbit serum, and subsequently incubated for 3days at 48C with P7 antibody diluted in 100 mM PBScontaining 1% BSA and 0.03% Triton X-100. After wash-ing the sections three times with 100 mM PBS for 30 min,they were incubated for 90 min with biotinylated rabbit
Ž .anti-mouse IgG 1 : 50 dilution, DAKO ArS, Denmark
Fig. 2. SDS–PAGE analysis of the fusion proteins, MBP-p7, MBP-p9and MBP-bgal. Cell lysates from 10 ml of cultures of E. coli BL21
Žstrains harboring pMAL-p7, pMAL-p9, or pMAL-c2 plasmid without y,. Ž .lanes 1, 4, 7, respectively and with q, lanes 2, 5, 8, respectively the
addition of IPTG were electrophoresed and stained with CoomassieŽ .brilliant blue. In lanes 3, 6 and 9 designated A , 0.5 mg of affinity-puri-
fied MBP-p7, MBP-p9 and MBP-bgal were electrophoresed, respec-tively.
Fig. 3. Characterization of P7 and P9 antibodies. A: the cross-reactivitiesof P7 and P9 antibodies to chicken red and rhodopsin. The purified fusion
Ž .proteins MBP-p7 and MBP-p9, 300 ng each , chicken red and rhodopsinŽ .2 mg each were electrophoresed, and the gel was stained by Coomassiebrilliant blue or subjected to Western blot analyses using P7 and P9antibodies. B: Western blot analysis of the chicken pineal proteins by P9antibody. Twenty pineal glands taken from adult chickens were homoge-
Žnized in 1 ml of buffer P 50 mM HEPES, 140 mM NaCl, 1 mM DTT, 4.mgrml leupeptin, and 50 kallikrein inhibitor Urml aprotinin, pH 6.6
Ž .and centrifuged 15000= g, 15 min, 48C to remove soluble proteinsŽ .Soluble . The membrane proteins in the precipitate were solubilized withbuffer P containing 2% CHAPS, followed by centrifugation to separate
Ž . Ž .the extract Extract and insoluble materials in the pellet Insoluble .Ž .These fractions corresponding to 0.5 gland or MBP-p9 5 ng were
subjected to Western blot analysis using P9 antibody.

( )T. Okano et al.rMolecular Brain Research 50 1997 190–196 193
and then rinsed in 100 mM PBS. They were furtherincubated for 90 min in 100 mM PBS containing peroxi-
Ždase-conjugated streptavidin 1 : 80 dilution, DAKO ArS,.Denmark , and rinsed twice by 100 mM PBS and 100 mM
Tris-HCl buffer pH 7.6 for 30 min. Positive signals wererevealed by incubation of the section for 5 min in Tris
Ž .buffer containing 0.05% diaminobenzidine DAB and0.01% H O . After rinsing, the sections were mounted on2 2
poly-L-lysine-coated slides, dehydrated, and coverslippedŽ .with an Entellan Merck, Germany .
The procedures for immunoelectron microscopy arew xdescribed elsewhere 9 . Briefly, the fixed pineal glands
were cut into 50-mm-thick sections with a VibratomeŽ .Technical Products International, USA . The sections vi-sualized by DAB were further processed with 1% OsO in4
100 mM phosphate buffer pH 7.4 for 90 min, dehydrated,and flat-embedded in Araldite for ultra-thin sectioning.The ultrathin sections stained with uranyl acetate and lead
Žcitrate were examined with an electron microscope JEM-.1210, Jeol .
( )2.4.2. Staining of chicken red iodopsinOne-day-old chickens were used for experiments. After
the procedures of tissue fixation described above, theŽpineal glands were sectioned with a Vibratome Technical
.Products International Inc., USA . The 20-mm-thick sec-tions were processed for immunocytochemical staining
Ž .using a monoclonal antibody 2 mgrml against chickenŽ w x.red R2; 20 according to the procedures described above.
Fig. 4. Immunoreactivities of anti-pinopsin and anti-chicken red antibodies in pineal glands of the chicken and pigeon. Frozen thin sections of pineal glandsŽ . Ž .from 2-month-old male chickens A,B and adult pigeons C were subjected to immunostaining by P7 antibody. Strong immunoreactivity was found in the
Ž . Ž . Ž .pineal lumina L of follicles A–C . Anti-chicken red monoclonal antibody R2-stained processes D, arrows in the follicular lumina of the pineal glandfrom 1-day-old chickens. In addition to the follicular lumen, pinopsin-positive cellular elements with a bulb- or string-shape were observed at the
Ž .parafollicular zone in the chicken pineal gland B, arrowhead . All the pinopsin-positive reactions were completely suppressed by preincubation of P7Ž .antibody with 20 mgrml of MBP-p7 not shown . Scale bar in A corresponds to: A, 500; and B,C,D, 50 mm.

( )T. Okano et al.rMolecular Brain Research 50 1997 190–196194
3. Results
3.1. Preparation of fusion proteins and antibodies
Ž .Pinopsin 351 amino acids is overall 43–48% identicalin amino acid sequence to known vertebrate visual pig-
w xments 14,19 , but identity is extremely low in the 40Ž .amino acid C-terminal region Fig. 1 . This led us to select
the C-terminal region for the preparation of pinopsin-specific antibodies. The cDNA fragment encoding the
ŽC-terminal 75 or 55 amino acid region p7 or p9 fragment,.respectively; Fig. 1 was subcloned into pMAL-c2 to yield
two recombinant plasmids, with which the E. coli BL21strain was transformed. Expression of the fusion protein
Žwas clearly detected 2 h after the addition of IPTG Fig..2 .
The fusion proteins, MBP-p7 and MBP-p9, were puri-fied by amylose affinity column chromatography, and thenused for immunization of mice. The anti-MBP antibodieswere eliminated from the antisera by passing them throughthe MBP-bgal-immobilized Sepharose column.
3.2. Characterization of the antibodies
Ž .In ELISA, the anti-MBP-p7 termed P7 antibody andŽ .anti-MBP-p9 termed P9 antibody prepared as described
above reacted strongly with the immunized antigensŽ .MBP-p7 and MBP-p9, respectively , but showed a very
Ž .weak reaction with MBP data not shown . The ELISA
also showed that the immunoreactions of both P7 and P9antibodies were almost completely and equally suppressedby pre-incubation with either MBP-p9 or MBP-p7 pro-teins. This indicated that both antibodies recognized theC-terminal 55 amino acid sequence, though the p7 regionencompasses an additional 20 amino acid residues of the
Ž .7th membrane spanning helix Fig. 1 . Since retinalrhodopsin and chicken red may be expressed in the pineal
w xgland in addition to pinopsin 1,19 , we examined thecross-reactivities of the P7 and P9 antibodies to the twopigments. As expected from the low homology between
Ž .the C-terminal sequence of pinopsin p9 region and thoseŽ .of the retinal pigments Fig. 1 , neither P7 nor P9 antibod-
Ž .ies cross-reacted with the two retinal pigments Fig. 3A .Taken together, these results indicate that both P7 and P9antibodies share similar affinity and high specificity for theC-terminal tail of pinopsin.
The immunoreactivity of P9 antibody to chicken pinealŽ .proteins was examined by Western blot analysis Fig. 3B .
The pineal membrane fraction was isolated from chickenpineal glands, and the membrane proteins were extractedwith 2% CHAPS. As shown in Fig. 3B, P9 antibody
Ž . Ž .stained a single band 42 000 Da in the extract Extractand a faint band with similar mobility in the insoluble
Ž .materials Insoluble . These reactions were completelysuppressed by pre-incubation of P9 antibody with 20
Ž .mgrml of MBP-p9 not shown . The results indicate thatpinopsin can be extracted effectively by 2% CHAPS justlike retinal pigments.
Fig. 5. Immunoelectron microscopic observation of sections of the parafollicular zone of the chicken pineal gland treated with anti-pinopsin antibody.Ultrathin sections were cut out from the parafollicular zone of the chicken pineal sections which had been stained with P7 antibody. A: a strong
Ž . Ž .immunoreactive element having a ciliary structure connected with the basal body b of the inner segment i . B: pinopsin-negative cellular elementsŽ .having whorl-like structures w were observed inside the pinopsin-positive lamellar complex. Scale bars: 1 mm.

( )T. Okano et al.rMolecular Brain Research 50 1997 190–196 195
3.3. Immunocytochemical localization of pinopsin in thepineal glands of chicken and pigeon
Cells expressing pinopsin were detected in the chickenand pigeon pineal glands by P7 antibody. As shown in Fig.4A,B, strong immunoreactivity was found in the lumina ofalmost all follicles of the chicken pineal gland. Similarly,immunoreactive pear-shaped processes were observed in
Ž .the pineal follicular lumen of the pigeon Fig. 4C . In spiteof the slight morphological difference between the folliclesof the two species, the localization of the immunoreactivi-ties are consistent with the presence of pinopsin at theregressed outer segments of modified photoreceptor cellsin the pineal gland. In addition to the follicular lumen,pinopsin-positive cellular elements with string- or bulb-shapes were observed at the periphery of the folliclesŽ . Žparafollicular zone in the chicken pineal gland Fig. 4B,
.arrowheads .These pinopsin-positive cellular elements in the parafol-
licular zone were further characterized by means of immu-w xnoelectron microscopy 9 . Fig. 5A shows that a pinopsin-
Ž .positive cilium-like process arose from the basal body bŽ .of the inner segment i . This process probably corre-
sponds to the string-shaped cellular element observed byŽ .the light-microscopy Fig. 4B . On the other hand, a
Ž .bulb-shaped pinopsin-positive element Fig. 4B seems toŽ .represent a whorl-like structure as shown in Fig. 5B w .
In contrast, immunoreactivity to a monoclonal antibodyŽ . w xR2 specific for chicken red 20 was found only in thefollicular lumina as shown in Fig. 4D. The parafollicularpinealocytes were immunonegative as we observed previ-
w xously for the quail pineal gland 13 . The immunoreactiveapical portions of the follicular cells protruded into the
Ž .lumina Fig. 4D , indicating the presence of chicken red atthe regressed outer segments of chicken modified photore-ceptor cells.
4. Discussion
The isolation of the cDNA for the pineal photoreceptivew xmolecule, pinopsin 19 , allowed us to prepare a fusion
protein consisting of MBP and pinopsin. Western blotŽ .analysis using the resultant antibody Fig. 3B showed the
apparent molecular weight of pinopsin to be 42 000. Thisis slightly higher than the value calculated from the de-
Ž w x.duced primary structure of pinopsin 38 187; 14,19 andmay indicate post-translational modification of pinopsin,
Ž 4 11.such as glycosylation potential sites, Asn and AsnŽ 314 315.andror palmitoylation possible sites, Cys and Cys
which are known to occur in the case of rhodopsin.However, the apparent molecular weights of chicken redŽ . Ž .35 000; Fig. 3A and chicken rhodopsin 37 000; Fig. 3A
w xare lower than the calculated values 40 279 11 and 39 281w x22 , respectively. Pinopsin might be modified by unknown
Ž .group s of high molecular weight.
Light- and electron microscopic observation of thechicken pineal gland revealed the localization of pinopsin
Žat both follicular and parafollicular pinealocytes Figs. 4.and 5 . Interestingly, the pinopsin-positive cellular ele-
ments in the parafollicular zone had string- or bulb-shapesŽ .Fig. 4B, arrowhead . The connection of the string-shaped
Ž .ciliary structure with the basal body Fig. 5A stronglysuggests that the immunoreactive elements represent outersegments of the parafollicular pinealocytes. Recently, wehave studied the ultrastructures of these elements in more
w xdetail by immunoelectron microscopy 9 , and shown thatsome of the whorl-like structures at the parafollicular zoneŽ .Fig. 5B arise from basal bodies located in the innersegments. Such bulbous protrusions or ciliary structures ofparafollicular pinealocytes have not been identified in the
w xpineal gland, although Ohshima and Matsuo 16 notedmorphological similarities between the follicular andparafollicular pinealocytes using conventional transmissionelectron microscopy. Taking these observations into ac-
Žcount, we suggest that not only the follicular cells mod-.ified photoreceptor cells but also the parafollicular
pinealocytes are photosensitive.Immunocytochemistry has shown that chicken red-posi-
Ž .tive cells Fig. 4D are fewer in number compared toŽ .pinopsin-positive cells Fig. 4B . This is consistent with
the previous result of Northern blot analysis showing thatmRNA for pinopsin is more abundant than that for chicken
w xred 19 . On the other hand, the distribution of thepinopsin-immunoreactivities is reminiscent of the localiza-
w xtion of HIOMT in both types of chicken pineal cells 3 .Based on a pre-supposition that photoreceptive moleculesare localized only in modified photoreceptor cells, theparafollicular pinealocytes were postulated to respond toneurotransmitters to synthesize and secrete melatonin whilethe modified photoreceptor cells were attributed the role of
w xmediating the action of light 3 . Although this is stillpossible, the present results rather suggest that both typesof pineal cells have the ability to respond to light andsynthesize melatonin.
Acknowledgements
This work was supported in part by grants-in-aid fromthe Japanese Ministry of Education, Science, Sports andCulture, and by research grants of SUNBOR, the Sumit-omo Foundation, and CREST.
References
w x1 M. Araki, Y. Fukada, Y. Shichida, T. Yoshizawa, F. Tokunaga,Differentiation of both rod and cone types of photoreceptors in thein vivo and in vitro developing pineal glands of the quail, Dev.
Ž .Brain Res. 65 1992 85–92.w x2 M. Bernard, M. Iuvone, V.M. Cassone, P.H. Roseboom, S.L. Coon,

( )T. Okano et al.rMolecular Brain Research 50 1997 190–196196
D.C. Klein, Avian melatonin synthesis: photic and circadian regula-tion of serotonin N-acetyltransferase mRNA in the chicken pineal
Ž .gland and retina, J. Neurochem. 68 1997 213–224.w x3 M. Bernard, P. Voisin, J. Guerlotte, J.P. Collin, Molecular and´
cellular aspects of hydroxyindole-O-methyltransferase expression inŽ .the developing chick pineal gland, Dev. Brain Res. 59 1991
75–81.w x4 T. Deguchi, Circadian rhythm of serotonin N-acetyltransferase activ-
Ž .ity in organ culture of chicken pineal gland, Science 203 19791245–1247.
w x5 T. Deguchi, Rhodopsin-like photosensitivity of isolated chickenŽ .pineal gland, Nature 290 1981 706–707.
w x6 R.G. Foster, J.J. Schalken, A.M. Timmers, W.J. DeGrip, A compari-son of some photoreceptor characteristics in the pineal and retina
Ž .I: the Japanese quail Coturnix coturnix , J. Comp. Physiol. A 165Ž .1989 553–563.
w x7 R.G. Foster, A.M. Timmers, J.J. Schalken, W.J. DeGrip, A compari-son of some photoreceptor characteristics in the pineal and retina
Ž .II: the Djungarian hamster Phodopus sungorus , J. Comp. Physiol.Ž .A 165 1989 565–572.
w x8 K. Goto, N. Miki, H. Kondo, An immunohistochemical study ofpinealocytes of chicks and some other lower vertebrates by means of
Ž .visinin retinal cone-specific protein -immunoreactivity, Arch. His-Ž .tol. Cytol. 52 1989 451–458.
w x9 K. Hirunagi, S. Ebihara, T. Okano, Y. Takanaka, Y. Fukada,Immunoelectron-microscopic investigation for the subcellular local-ization of pinopsin in the pineal gland of the chicken, Cell Tissue
Ž .Res. 289 1997 235–241.w x10 H.-W. Korf, R.G. Foster, P. Ekstrom, J.J. Schalken, Opsin-like¨
immunoreaction in the retinae and pineal organs of four mammalianŽ .species, Cell Tissue Res. 242 1985 645–648.
w x11 O. Kuwata, Y. Imamoto, T. Okano, K. Kokame, D. Kojima, H.Matsumoto, A. Morodome, Y. Fukada, Y. Shichida, K. Yasuda, Y.Shimura, T. Yoshizawa, The primary structure of iodopsin, a chicken
Ž .red-sensitive cone pigment, FEBS Lett. 272 1990 128–132.w x12 U.K. Laemmli, Cleavage of structural proteins during the assembly
Ž .of the head of bacteriophage T4, Nature 227 1970 680–685.
w x13 H. Masuda, T. Oishi, M. Ohtani, M. Michinomae, Y. Fukada, Y.Shichida, T. Yoshizawa, Visual pigments in the pineal complex ofthe Japanese quail, Japanese grass lizard and bullfrog: immunocyto-
Ž .chemistry and HPLC analysis, Tissue Cell 26 1994 101–113.w x14 M. Max, P.J. McKinnon, K.J. Seidenman, R.K. Barrett, M.L. Apple-
bury, J.S. Takahashi, R.F. Margolskee, Pineal opsin: a nonvisualŽ .opsin expressed in chick pineal, Science 267 1995 1502–1506.
w x15 K. Ohshima, S. Matsuo, Functional morphology of the pineal glandŽ .in young chickens, Anat. Anz. Jena 156 1984 407–418.
w x16 K. Ohshima, S. Matsuo, Cytodifferentiation of the chick pinealgland, with special reference to the photosensory and secretory
Ž .elements, J. Pineal Res. 5 1988 397–410.w x17 T. Okano, Y. Fukada, I.D. Artamonov, T. Yoshizawa, Purification
Ž .of cone visual pigments from chicken retina, Biochemistry 28 19898848–8856.
w x18 T. Okano, D. Kojima, Y. Fukada, Y. Shichida, T. Yoshizawa,Primary structures of chicken cone visual pigments: vertebraterhodopsins have evolved out of cone visual pigments, Proc. Natl.
Ž .Acad. Sci. USA 89 1992 5932–5936.w x19 T. Okano, T. Yoshizawa, Y. Fukada, Pinopsin is a chicken pineal
Ž .photoreceptive molecule, Nature 372 1994 94–97.w x20 Y. Shichida, Y. Taniguchi, O. Kuwata, Y. Fukada, T. Yoshizawa, S.
Horiuchi, M. Takeichi, Monoclonal antibodies to chicken iodopsin,Ž .Exp. Eye Res. 48 1989 281–293.
w x21 O. Shouno, K. Kokame, M. Araki, T. Takao, Y. Shimonishi, M.Murata, T. Yoshizawa, Y. Fukada, Preparation and characterizationof monoclonal antibodies specific for lauroylated isoform of bovinetransducin a-subunit: immunohistochemical analysis of bovine reti-
Ž .nas, J. Neurochem. 66 1996 2188–2196.w x22 M. Takao, A. Yasui, F. Tokunaga, Isolation and sequence determina-
Ž .tion of the chicken rhodopsin gene, Vision Res. 28 1988 471–479.w x23 B. Vigh, I. Vigh-Teichmann, Light- and electron-microscopic
demonstration of immunoreactive opsin in the pinealocytes of vari-Ž .ous vertebrates, Cell Tissue Res. 221 1981 451–463.
w x24 B. Vigh, I. Vigh-Teichmann, Three types of photoreceptors in thepineal and frontal organs of frogs: ultrastructure and opsin immuno-
Ž .reactivity, Arch. Histol. Jpn. 49 1989 495–518.
Copyright © 2022 FDOKUMEN




















