
Building Capacity in Smaller Organizations: A Leadership Framework for Identity-Based Groups

Alternative splicing generates a smaller assortment of CaV2.1 transcripts incerebellar Purkinje cells than in the cerebellum
Srinivasan Kanumilli, Elizabeth W. Tringham, C. Elizabeth Payne, Jonathan R. B. Dupere,Kanamarlapudi Venkateswarlu, and Maria M. UsowiczDepartment of Pharmacology, University of Bristol, Bristol, United Kingdom
Submitted 27 June 2005; accepted in final form 7 November 2005
Kanumilli, Srinivasan, Elizabeth W. Tringham, C. ElizabethPayne, Jonathan R. B. Dupere, Kanamarlapudi Venkateswarlu,and Maria M. Usowicz. Alternative splicing generates a smallerassortment of CaV2.1 transcripts in cerebellar Purkinje cells than inthe cerebellum. Physiol Genomics 24: 86–96, 2006. First publishedNovember 8, 2005; doi:10.1152/physiolgenomics.00149.2005.—P/Q-type calcium channels control many calcium-driven functions in thebrain. The CACNA1A gene encoding the pore-forming CaV2.1 (�1A)subunit of P/Q-type channels undergoes alternative splicing at multi-ple loci. This results in channel variants with different phenotypes.However, the combinatorial patterns of alternative splice events at twoor more loci, and hence the diversity of CaV2.1 transcripts, areincompletely defined for specific brain regions and types of brainneurons. Using RT-PCR and splice variant-specific primers, we haveidentified multiple CaV2.1 transcript variants defined by differentpairs of splice events in the cerebellum of adult rat. We haveuncovered new splice variations between exons 28 and 34 (some ofwhich predict a premature stop codon) and a new variation in exon 47(which predicts a novel extended COOH-terminus). Single cell RT-PCR reveals that each individual cerebellar Purkinje neuron alsoexpresses multiple alternative CaV2.1 transcripts, but the assortment issmaller than in the cerebellum. Two of these variants encode differentextended COOH-termini which are not the same as those previouslyreported in Purkinje cells of the mouse. Our patch-clamp recordingsshow that calcium channel currents in the soma and dendrites ofPurkinje cells are largely inhibited by a concentration of �-agatoxinIVA selective for P-type over Q-type channels, suggesting that thedifferent transcripts may form phenotypic variants of P-type calciumchannels in Purkinje cells. These results expand the known diversityof CaV2.1 transcripts in cerebellar Purkinje cells, and propose theselective expression of distinct assortments of CaV2.1 transcripts indifferent brain neurons and species.
splice variants; P type; calcium channels; Purkinje neurons
0 VOLTAGE-GATED calcium (Ca2�) channels regulate numerouscellular functions, from neuronal electrical activity to intracel-lular signaling pathways. Diversity of Ca2� channels in thebrain is apparent from the many roles that Ca2� channels playin different cell types and subcellular compartments (3). Thediversity reflects, in part, the existence of nine classes of thepore-forming CaV (�1) subunit in the brain and alternativepre-mRNA splicing of the gene encoding each class of CaV
subunit (12, 17). P/Q-type Ca2� channels control many func-tions throughout the brain and contain a CaV2.1 subunit (alsoknown as �1A). However, information about splicing of theCACNA1A gene encoding CaV2.1 in different brain regions
and neurons is incomplete (2, 5, 16, 30, 33, 35, 37). Augmen-tation of what is currently known about CaV2.1 splicing in thecerebellum and cerebellar Purkinje cells is of particular inter-est, because P-type currents in Purkinje cells are the prototyp-ical P-type currents against which putative P-type and Q-typecurrents in other cells are compared, but it is not clear howmany CaV2.1 transcript variants there are in Purkinje cells, orwhich ones harbor the different CACNA1A mutations associ-ated with the loss of Purkinje cells in spinocerebellar ataxiatype 6 and familial hemiplegic migraine (27).
Alternative splicing of CACNA1A at multiple loci has beendemonstrated by the cloning of CaV2.1 cDNAs from varioustissues and cells of different species (2, 8, 9, 14, 23, 31, 35, 35,41) and by transcript mapping of CaV2.1 cDNAs from differentregions of human brain, including the cerebellum (30). Thecombinatorial patterns of alternate splice events at two or moreloci remain largely unidentified for CaV2.1 in the cerebellum,because each transcript analyzed in the transcript mappingstudy contained only a single splice locus. Furthermore, giventhe potential for species-specific splicing (26), it is unclear howaccurately splicing in human cerebellum predicts splicing inthe cerebellum of rodents, which are used for electrophysio-logical and pharmacological studies of native P/Q-type cur-rents and for manipulation of CACNA1A (18). Similarly, thetwo CaV2.1 splice variants cloned from mouse Purkinje cells(35) may not be representative of Purkinje cells in otherspecies, or they may not be the only variants in these cells.Here, we begin to address these issues by RT-PCR analysis ofpatterns of splicing in adult rat cerebellum and rat cerebellarPurkinje cells, and by searching for splice events known tooccur in Purkinje cells of the mouse.
MATERIALS AND METHODS
Isolation of cerebellar mRNA and PCR. Adult male Wistar rats[aged �40 postnatal days (P40) or 150–175 g in weight], young maleWistar rats (P8 and P20), and adult male C57BL/6J mice (P42) werekilled by cervical dislocation and decapitated in accordance with theAnimals (Scientific Procedures) Act 1986, and protocols were ap-proved by the University of Bristol Ethical Review Committee. cDNAwas obtained from the cerebellar vermis and amplified by PCR usingstandard procedures. (For further details, see Supplemental Methods;available at the Physiological Genomics web site.)1
Design and experimental verification of splice variant specificity ofprimers. Potential sites of splice variation were identified by aligningthe sequences of known rodent CaV2.1 cDNAs (Fig. 1; OMIGAsoftware or DS Gene software, Accelrys, Cambridge, UK). Primers(Table 1) were designed with the aid of Oligo 6.1 software (MolecularBiology Insights, Cascade, CO) and the basic local alignment searchArticle published online before print. See web site for date of publication
(http://physiolgenomics.physiology.org).Address for reprint requests and other correspondence: M. M. Usowicz,
Dept. of Pharmacology, Univ. of Bristol, Univ. Walk, Bristol BS8 1TD, UK(e-mail: [email protected]).
1 The Supplemental Material for this article is available online at http://physiolgenomics.physiology.org/cgi/content/full/00149.2005/DC1.
Physiol Genomics 24: 86–96, 2006.First published November 8, 2005; doi:10.1152/physiolgenomics.00149.2005.
1094-8341/06 $8.00 Copyright © 2006 the American Physiological Society86
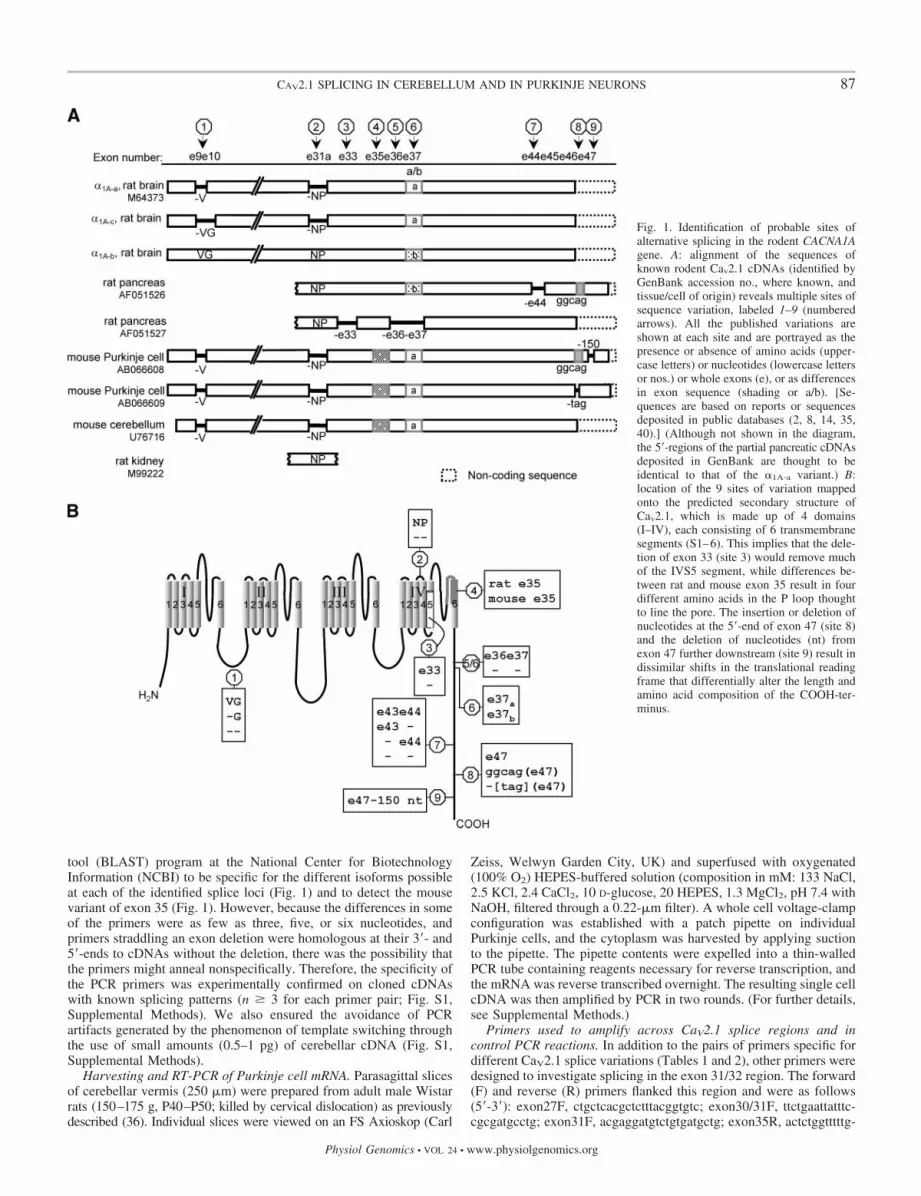
tool (BLAST) program at the National Center for BiotechnologyInformation (NCBI) to be specific for the different isoforms possibleat each of the identified splice loci (Fig. 1) and to detect the mousevariant of exon 35 (Fig. 1). However, because the differences in someof the primers were as few as three, five, or six nucleotides, andprimers straddling an exon deletion were homologous at their 3�- and5�-ends to cDNAs without the deletion, there was the possibility thatthe primers might anneal nonspecifically. Therefore, the specificity ofthe PCR primers was experimentally confirmed on cloned cDNAswith known splicing patterns (n � 3 for each primer pair; Fig. S1,Supplemental Methods). We also ensured the avoidance of PCRartifacts generated by the phenomenon of template switching throughthe use of small amounts (0.5–1 pg) of cerebellar cDNA (Fig. S1,Supplemental Methods).
Harvesting and RT-PCR of Purkinje cell mRNA. Parasagittal slicesof cerebellar vermis (250 �m) were prepared from adult male Wistarrats (150–175 g, P40–P50; killed by cervical dislocation) as previouslydescribed (36). Individual slices were viewed on an FS Axioskop (Carl
Zeiss, Welwyn Garden City, UK) and superfused with oxygenated(100% O2) HEPES-buffered solution (composition in mM: 133 NaCl,2.5 KCl, 2.4 CaCl2, 10 D-glucose, 20 HEPES, 1.3 MgCl2, pH 7.4 withNaOH, filtered through a 0.22-�m filter). A whole cell voltage-clampconfiguration was established with a patch pipette on individualPurkinje cells, and the cytoplasm was harvested by applying suctionto the pipette. The pipette contents were expelled into a thin-walledPCR tube containing reagents necessary for reverse transcription, andthe mRNA was reverse transcribed overnight. The resulting single cellcDNA was then amplified by PCR in two rounds. (For further details,see Supplemental Methods.)
Primers used to amplify across CaV2.1 splice regions and incontrol PCR reactions. In addition to the pairs of primers specific fordifferent CaV2.1 splice variations (Tables 1 and 2), other primers weredesigned to investigate splicing in the exon 31/32 region. The forward(F) and reverse (R) primers flanked this region and were as follows(5�-3�): exon27F, ctgctcacgctctttacggtgtc; exon30/31F, ttctgaattatttc-cgcgatgcctg; exon31F, acgaggatgtctgtgatgctg; exon35R, actctggtttttg-
Fig. 1. Identification of probable sites ofalternative splicing in the rodent CACNA1Agene. A: alignment of the sequences ofknown rodent Cav2.1 cDNAs (identified byGenBank accession no., where known, andtissue/cell of origin) reveals multiple sites ofsequence variation, labeled 1–9 (numberedarrows). All the published variations areshown at each site and are portrayed as thepresence or absence of amino acids (upper-case letters) or nucleotides (lowercase lettersor nos.) or whole exons (e), or as differencesin exon sequence (shading or a/b). [Se-quences are based on reports or sequencesdeposited in public databases (2, 8, 14, 35,40).] (Although not shown in the diagram,the 5�-regions of the partial pancreatic cDNAsdeposited in GenBank are thought to beidentical to that of the �1A-a variant.) B:location of the 9 sites of variation mappedonto the predicted secondary structure ofCav2.1, which is made up of 4 domains(I–IV), each consisting of 6 transmembranesegments (S1–6). This implies that the dele-tion of exon 33 (site 3) would remove muchof the IVS5 segment, while differences be-tween rat and mouse exon 35 result in fourdifferent amino acids in the P loop thoughtto line the pore. The insertion or deletion ofnucleotides at the 5�-end of exon 47 (site 8)and the deletion of nucleotides (nt) fromexon 47 further downstream (site 9) result indissimilar shifts in the translational readingframe that differentially alter the length andamino acid composition of the COOH-ter-minus.
87CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

gatcccgg. To confirm that the PCR reactions had worked, PCRreactions included pairs of primers targeted against mRNAs differen-tially expressed by different types of neurons in the cerebellum. Thesewere as follows (5�-3� sequences with predicted size of PCR productshown): calbindin D-28K (F, aggcacgaaagaaggctggat; R, tcccaca-cattttgattccctg; 432 bp) or the transcription factor zipro 1 (F,ggccctatgactgtaagtgtg; R, gtgtggactctctgatgcttg; 409 bp) or theGABAA�6 receptor subunit (F, atggactgatgagaggctga; R, tctgggac-ctctactgaataaagc; 342 bp) or GAD65 (F, tcttttctcctggtggtgcc; R,
ccccaagcagcatccacat; 390 bp) or the NR2A and NR2C N-methyl-D-aspartate (NMDA) receptor subunits (F, ggggttctgcatcgacatcc; R,gacagcaaagaaggcccacac; 546 bp).
Analysis of PCR products. PCR products were usually identifiedaccording to size by agarose gel electrophoresis. Some were se-quenced, either directly or after subcloning. (For further details, seeSupplemental Methods.)
Cell-attached recording of Ca2� channel currents. The surface ofthe soma of a Purkinje cell or of the first dendritic bifurcation of aPurkinje cell was cleaned of overlying debris by applying a stream ofextracellular solution from a pipette. Cell-attached recordings weremade from the cleaned membrane with pipettes (thick-walled boro-silicate glass capillaries; Harvard Apparatus, Kent, UK) filled with afiltered (0.2 �m) solution containing (in mM) 5 BaCl2, 10 CsCl, 10HEPES, 134 or 150 TEA-Cl, 0.1 EGTA, pH 7.4 with TEA-OH, plus1 �M TTX. For some recordings, the pipette solution also contained30 or 100 nM �-agatoxin IVA (Scientific Marketing Associates,Barnet, UK). Pipette resistances were 4–10 M� for somatic record-ings and 6–13 M� for dendritic recordings. A depolarizing voltageramp (170 mV, 0.53 mV/ms, starting 30–50 mV negative to theresting cell potential) followed by a repolarizing ramp was applied tothe pipette every 5 s using a Cambridge Electronic Design (CED)1401 plus A/D interface (Cambridge, UK). The evoked currents werelow-pass filtered at 2 kHz and acquired at 7 kHz. At the end ofcell-attached recording, a whole cell configuration was established tomeasure the cell resting potential. Patch potentials were calculated asthe resting cell potential measured at the end of cell-attached record-ing (e.g., approximately �60 mV) minus the pipette holding potential(e.g., �40 mV) and minus the applied voltage ramp (from 0 to �170mV). Thirty currents were recorded from each patch and averaged. Alinear leak current was then subtracted to give the mean Ca2� channelcurrent. Recordings with drug-containing and drug-free pipettes were
Table 1. PCR primers for analysis of splicing of the CACNA1A gene in the cerebellumand in cerebellar Purkinje cells of the rat
Name Sequence (5�-3�) Position Accession No.
Forward primers
e2 cctgatgatgacaagacacc 370–389 M64373�VG cttttgatgctcttcggaga 1,253 M64373*VG ccttttgatgttggagctctt 1,252 M64373*NP tcaccgagtttgggaataacttca 4,793–4,816 M64373NP accgagtttgggaatccgaata 94–115 AF051526�e33 ctttcgtgcagtctttcaaggtg 209–231 AF051527mouse e35 tgggatcctcacggcagac 5,237–5,255 AB066608e37a aggacatgtacagtttattgcgag 5,402–5,425 M64373e37b tcgcatgccttacccggac 693–711 AF051526e43 tggagatgagagaaatgggaac 6,050–6,071 M64373ggcag(e47) cgacacacagacagggcagtag 1,892–1,913 AF051526�tag(e47) acagacagttccgtcagtgga 6,686–6,706 AB066609
Reverse primers
�V tagctctccgaagagctcca 1,260–1,279 M64373e14 aacagcagacttatgatggatttc 1,839–1,862 M64373e35 actctggtttttggatcccgg 5,177–5,197 M64373�e36�e37 gtccatccgcaagagcctcag 593–513 AF051527e37ai cgatattactcgcaataaactgtacatg 5,406–5,433 M64373e37aii ccgagagggggcgatattac 5,425–5,444 M64373e37b ggacatgtgtctcagcatctg 718–738 AF051526�e43 ggccccttctcctctggga 6,040 M64373*�e44 gatggtctggttctctgctgg 1,450–1,470 AF051526e44 actgaggttatttccacgtgg 6,175–6,195 M64373�e43�e44 tggtatcagagatggtctggga 6,040 M64373*e47 tgccccgcgtagtaaccatt 2,281–2,300 AF051526
The position of each primer is given on a rat CaV2.1 cDNA (identified by accession no.); e, exon. In the cases denoted by an asterisk (*), the position of onlythe first nucleotide is indicated in the M64373 variant, since none of the CaV2.1 cDNAs in GenBank or EMBL-Bank contains the splice variation encoded bythe primer [either because the sequence to which the primer is targeted was described in a publication but not deposited in a public database, or because the primersequence (�e43, �e43�e44) is based on deletions in human CaV2.1 cDNA]. Two reverse exon 37a primers (i, ii) were used in the study.
Table 2. Pairs of primers used to detect CaV2.1 splicevariations in the cerebellum and in cerebellarPurkinje cells of the rat
Forward/ReversePrimer Pairs
Predicted ProductSize, bp
Forward/ReversePrimer Pairs
Predicted ProductSize, bp
e2/�V 910 e37a/�e43�e44 660�VG/e14 607 e37b/�e43–e44 674VG/e14 614 e43/e44 146�NP/e35 405 e37a/�e44 764NP/e35 409 e37b/�e44 778�NP/e37a(i) 641 e37a/e44 794�NP/e37b 647 e37b/e44 815NP/e37a(i) 645 NP/�e44 1,377NP/e37b 651 �NP/�e44 1,373mouse e35/e37a(ii) 267 �NP/e44 1,403�e33/�e36�e37 304 NP/e44 1,407e37a/�e43 658 ggcag(e47)/e47 409/257e37b/�e43 672 �tag(e47)/e47 394
The two product sizes shown for the ggcag (e47)/e47 pair of primers arepredicted for ggcag-containing transcripts with and without an 150-nucleo-tide deletion in exon 47 (site 9, Fig. 1A).
88 CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

interleaved throughout each day of recording to ensure that anydifferences in current size were due to an effect of �-agatoxin IVAand not due to variation arising from the use of different sets ofpipettes with and without drug, or from the use of different animals ondifferent days.
RESULTS
CaV2.1 cDNA analysis and location of splice event-specificprimers. To design primers that would enable an investigationof alternative splicing of the CACNA1A gene in rat cerebellumand in individual cerebellar Purkinje cells with RT-PCR, sitesof sequence variation in rodent CaV2.1 cDNAs were identifiedby aligning the sequences of cloned rodent CaV2.1 cDNAs.This comparison revealed nine sites of variation (Fig. 1A),many of which (1, 2, 6-8 in Fig. 1A) are now established as lociof alternative splicing by rat, mouse, and human genomic andcDNA analysis. Such analysis has determined that the �V,�VG, and VG variations (site 1) arise by the lengthening orshortening of the 5�-end of exon 10 during splicing (2, 30). The�NP and NP variations (site 2) arise by the exclusion orinclusion of a very short (6 nucleotides) exon 31a (2, 16, 30).Exons 37a and 37b (site 6) are alternate exons that are incor-porated into transcripts in a mutually exclusive manner (2, 30),and exon 44 (site 7) is a cassette exon (13, 14, 30). Extensionof the 5�-end of exon 47 by five nucleotides during splicing(ggcag, site 8) causes a shift in the reading frame that results inthe translation of exon 47 and an extended COOH-terminus (9,10, 13, 14, 28, 30, 33, 41).
The alignment also displays sites or patterns of variation inrodent Cav2.1 cDNAs not reported for other species. Theseinclude the coincident exclusion of exons 33, 36, and 37 (sites3, 5, and 6), removal of three nucleotides (the stop codon, tag)from the 5�-end of exon 47 (site 8), and deletion of 150nucleotides from exon 47 further downstream (site 9). Exclu-sion of the “tag” stop codon from the 5�-end of exon 47 duringsplicing results in the translation of exon 47. This encodes anextended COOH-terminus with no amino acid homology tothat of the extended tail generated by the ggcag insertion (35).The mechanisms by which 150 nucleotides are deleted fromexon 47 (site 9) remain to be elucidated. Finally, the differ-ences in the sequences of exon 35 (site 4, 17 nucleotides of151, 4 amino acids of 50) in mouse and rat Cav2.1 cDNAs arethought to be orthologous rather than splicing differences.
The various isoforms identified at the multiple locations aremapped on to the predicted membrane topology of the CaV2.1subunit in Fig. 1B. To determine which of these variants arepresent in rat cerebellum and rat cerebellar Purkinje cells, PCRprimers were designed to target each variation (Table 1). Inaddition, primers were designed to detect the inclusion orexclusion of exon 43 or both exons 43 and 44 (site 7, Fig. 1B),because transcripts differing in the presence or absence of theseexons have been reported in a human cerebellar CaV2.1 cDNAlibrary, and combinatorial inclusion and exclusion of exons 43and 44 affect current amplitude and Ca2�-dependent channelinactivation (30).
Multiple CaV2.1 transcripts in rat cerebellum. Twenty-sixpairs of primers were used to investigate the occurrence ofvarious splice events in rat cerebellar vermis (Table 2). Sevenof these consisted of a splice isoform-specific primer (�V or�VG or VG; �NP or NP; ggcag or �tag) and a primer locatedin a nonspliced region (in exon 2 or exon 14 or exon 35 or exon
47). These were used to explore single splice events in differ-ent transcripts. One pair (mouse e35/e37aii) contained a for-ward primer specific for the mouse version of exon 35. In theremaining 18 pairs, each primer was specific for one spliceisoform at one splice site. They were used to explore events attwo splice sites within individual transcripts. For example, thegel in Fig. 2A depicts an investigation into the pairing of exon37a or exon 37b with the presence or absence of exon 44(e37a/e37b with e44/�e44), and the pairing of the presence orabsence of the nucleotides encoding NP with the presence orabsence of exon 44 (�NP/NP with e44/�e44).
The results of the PCR experiments on adult rat cerebellarcDNA are summarized in Fig. 2, B and C. In total, 19 of the 26pairs of primers generated a PCR product in the majority ofreactions. In agreement with previous in situ hybridization,immunoblotting, and RT-PCR studies (2, 5, 28, 37), the histo-gram confirms the generation of three isoforms by alternativesplicing of nucleotides encoding valine and glycine (e2/�V,�VG/e14, and VG/e14 pairs of primers in Fig. 2B; site 1, Fig.2C), transcripts with or without the NP-encoding nucleotides(�NP/e35 and NP/e35 pairs of primers in Fig. 2B; site 2, Fig.2C), transcripts containing exon 37a or exon 37b (e.g., �NP/e37a, �NP/e37b, NP/e37a, NP/e37b pairs of primers in Fig.2B; site 6, Fig. 2C), transcripts with exon 44 (e.g., e43/e44 pairof primers in Fig. 2B; site 7, Fig. 2C), and ggcag-containingtranscripts (ggcag/e47 pair of primers in Fig. 2B; site 8, Fig.2C). Our results also indicate the existence of transcripts withor without exon 44 in rat cerebellum (Fig. 2B; site 7, Fig. 2C).CaV2.1 transcripts lacking exon 44 have not been observedpreviously in rat cerebellum but have been detected in ratpancreas (14) and in human cerebellum (10, 30, 33) and humanspinal cord (13).
The PCR experiments also uncovered the presence of mul-tiple transcripts in adult rat cerebellum generated by differentcombinations of known splice events. As shown in Fig. 2, Band C, transcripts were detected containing the following:NP/exon 37a, NP/exon 37b, �NP/exon 37a, and �NP/exon37b. Therefore, the two isoforms produced at each of these twosplice loci occur in all four possible combinations. In addition,PCR detected all four possible combinations of exon 37a or37b and exon 44 or �exon 44, and all four possible combina-tions of NP or �NP and exon 44 or �exon 44. Many of thesepairings have not been observed previously in rat cerebellum(crossed circles in Fig. 2B). Some have not been observed inbrain CaV2.1 transcripts of any species.
The mouse cerebellar Purkinje cell variant (Fig. 1A,AB066608) is characterized by the insertion of ggcag at the5�-end of exon 47 and the deletion of 150 nucleotides fromexon 47. Our experiments with the ggcag(e47)/e47 pair ofprimers did not provide evidence for such a transcript in ratcerebellum. Rather, these gave a PCR product that was of thesize predicted (Table 2) for the ggcag-containing exon 47 of arat pancreatic variant (Fig. 1A, AF051526) that does notcontain the deletion (14). In two of these reactions, an addi-tional smaller product was identified, but its size was margin-ally greater (300 bp) than the size predicted (Table 2) by the150-nucleotide deletion. Sequencing of these products con-firmed their lack of homology with the mouse cerebellarPurkinje cell variants (see below).
The absence of a PCR product in all or most reactions withthe remaining seven pairs of primers suggests an absence of
89CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

transcripts lacking exon 43; both exons 43 and 44; exons 33,36, and 37; or tag at the 5�-end of exon 47 and an absence oftranscripts containing the mouse version of exon 35 in combi-nation with exon 37a. The PCR reactions were repeated oncerebellar cDNA of rats of different age to determine whetherthe splicing profile changes during postnatal development (4,37) and whether age-related changes might explain why wewere unable to detect some splice events known to occur inhuman or mouse cerebellum [i.e., �e43 or �e43�e44 or�tag(e47)]. We found that the CaV2.1 splice variations absentfrom P40 rat cerebellum were also absent from the cerebellumof P8 and P21 rats. However, two transcript variants (�VG andNP/e37b) present at P40 did not occur at P8 (Fig. S2, Supple-mental Results), while some transcripts appeared to show adevelopmental increase in expression (ANOVA, P 0.05).These were transcripts lacking the nucleotides for valine (�V),transcripts containing nucleotides encoding the NP insert, andtranscripts with the combinations NP/exon 37a, NP/exon 37b,exon 37a/�exon 44, and exon 37b/�exon 44.
Further analysis of splicing in domain IV. The NP insert(exon 31a) in the IVS3-4 extracellular loop (Fig. 1B) is impor-tant in determining the sensitivity to block by �-agatoxin IVAand the electrophysiology of CaV2.1 channels (2, 9, 16, 33).We considered the possibility that splicing in this region mightgenerate cerebellar transcripts with the NP-encoding exonflanked by novel sequences or transcripts in which the NP-encoding exon is replaced by other nucleotides, all of whichwould not be detectable with our NP and �NP primers thatassume the simple inclusion and exclusion of a cassette exon.Therefore, PCR amplification was performed across this regionwith a reverse primer located in exon 35 and different forwardprimers (exon 27F, exon 30/31F, exon 31F). Subcloning andsequencing of 13 PCR products revealed 10 different splicingpatterns, illustrated in Fig. 3. (The inferred splicing mecha-nisms are shown in Fig. S3, Supplemental Results). Themajority (10/13) contained exon 31a, but only a minority ofthese (4/10) showed the simple incorporation of exon 31a (top2 entries in Fig. 3). In the others, exon 31a was accompaniedby alternative stretches of 24 or 168 nucleotides from the intronbetween exons 31 and 32 (EMBL-Bank accession nos.:AM040231, AM040232). One also included the intron be-tween exons 33 and 34 (EMBL-Bank, AM040234), whileanother also lacked exon 33 (EMBL-Bank, AM040233). Ad-ditionally, two transcripts lacked exons 29, 30, 31, and 31a(EMBL-Bank, AM040230). The simple exclusion of exon 31a,which has been described previously for rat and human CaV2.1(15, 30), was only observed in 1 of the 13 transcripts. Theinclusion of additional nucleotides from the intron betweenexons 31a and 32 predicts the introduction of a prematuretermination codon, located several amino acids after exon 31a(Fig. S3, Supplemental Results). Even when we alter thereading frame of these insertions, as could happen if thesesplice events are combined with upstream splice events, theinsertion of a stop codon is predicted, albeit in a differentlocation.
CaV2.1 alternative splicing in single cerebellar Purkinjecells. Having used RT-PCR to investigate splicing ofCACNA1A in rat cerebellum, we used the same primers toinvestigate CaV2.1 splice variants in individual cerebellar Pur-kinje neurons of adult rat with single cell RT-multiplex PCR.To confirm that the PCR products were generated from Pur-
Fig. 2. Identification of multiple Cav2.1 transcript variants with different splicingpatterns in adult rat cerebellum. A: agarose gel (negative image) illustrating theamplification of transcripts with the PCR primers indicated. The lane marked“calb.” depicts the amplification of calbindin D-28K, a positive control for PCR ofcerebellar cDNA. B: histogram summarizing the frequency with which PCRproducts of expected size (Table 2) were generated by the pairs of primersindicated. Nos. in bars indicate the no. of experiments carried out with each pairof primers. Asterisk (*) denotes that the forward ggcag primer and the reverse e47primer generated a single product (400 bp) in 7 reactions and 2 products (400bp, 300 bp) in an additional 2 reactions. Crossed circles denote splicing patternsthat have not been previously observed in rat cerebellar transcripts. C: schematicdiagram of the PCR products generated. Asterisk (*) emphasizes the generation oftwo ggcag-containing PCR products of distinct size. Numbered arrows refer to thesites of variation identified in Fig. 1.
90 CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

kinje cell mRNA, and that there was no contamination from thenumerous neighboring granule neurons, each reaction con-tained primers designed to amplify cDNAs for the transcriptionfactor zipro 1 or the GABAA �6-receptor subunit, both ofwhich are present only in granule cells (38, 39), and forcalbindin D-28K, which is found almost exclusively in Pur-kinje cells (1). Some reactions also included primers thatwould amplify GAD65, which is expressed in all GABAergicneurons but not in granule cells, or NR2A/C NMDA receptorsubunits, which are differentially expressed in adult rat cere-bellar granule (NR2A, NR2C) and Purkinje cells (NR2C) (32).The ability of these primers to distinguish between Purkinjeand granule neurons is demonstrated in Fig. 4.
Single cell RT-multiplex PCR experiments with the CaV2.1splice isoform-specific primers demonstrated the expression ofmultiple alternative CaV2.1 transcripts in single Purkinje neu-rons. This is exemplified by the simultaneous amplification ofPCR products containing alternative splice combinations (ex-emplar gel in Fig. 5A) and of multiple (2 or 3) ggcag(e47)-containing products of distinct size (asterisk in Fig. 5, B andC). However, only 8 of 26 pairs of primers (Fig. 5, B and C)amplified a product in a majority of cells (67–89%). Six pairsgenerated PCR products in a minority of cells (12–15%;dashed lines in Fig. 5C), which may represent rare transcriptsor false-positive results. This is in marked contrast with ourfinding that 19 pairs of primers generated a product fromcerebellar cDNA in 65–100% reactions (Fig. 2B). For instance,
of the three isoforms produced in the cerebellum by alternativesplicing at the 5�-end of exon 10 (Fig. 2), the �V isoform wasdetected in the majority of Purkinje cells, the �VG isoformwas undetected, and the VG isoform was detected in only 15%of cells. Likewise, of the NP and �NP isoforms produced inthe cerebellum by the inclusion or exclusion of exon 31a, onlythe �NP isoform was detected in the majority of Purkinjecells. These results suggest that, although each Purkinje cellcan expresses multiple transcript variants, regulation of alter-native splicing at the level of the individual Purkinje cellresults in a smaller range of CaV2.1 transcripts than in thewhole cerebellum.
An inconsistency in the single cell RT-PCR data is that itwas not possible to routinely amplify transcripts containing thesplice combination �NP/exon 37b, even though we readilydetected �NP transcripts (with �NP/e35 and �NP/e37aprimer pairs) and exon 37b transcripts (with e37b/e44 ore37b/�e44 primer pairs). This discrepancy could arise if novelsplicing events occur in the exon 30/32 region of transcriptscontaining exon 37b and are not detectable with our NP and�NP primers. In view of this, and because we found novelsplicing events between exons 28 and 34 in the cerebellum,splicing in Purkinje cells was further investigated with primerslocated in exon 27 (forward) and exon 35 (reverse). Agarosegel electrophoresis revealed only a single band (n � 6 cells).The sequences of these PCR products (not shown), obtained bydirect sequencing (n � 4 cells) or by subcloning and then
Fig. 3. Multiple CaV2.1 transcript variants in adult rat cerebellum generated by alternative splicing between exons 28 and 34. cDNAs were amplified with avariety of forward primers (F; located in exon 31a or at the exon 30/31 junction or in exon 27) and a reverse primer (R) in exon 35. The sequences of 13 clonedPCR products were compared with those of the known rodent CaV2.1 cDNAs and with rat CACNA1A using the Ensembl Genome Browser to deduce the 10splicing patterns shown. Empty boxes, exons; shaded boxes, introns. Exons and introns are drawn on different scales. The intron lengths shown are accordingto those in the public rat gene sequence, except for introns between exons 29 and 30 and between exons 30 and 31, which remain to be defined (unknown lengthsdenoted by breaks). The no. immediately to the right of each variant indicates the no. of products (1 or 2) with that particular splicing pattern. The pairs of primersused to amplify these products are indicated further to the right.
91CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

sequencing (n � 5 cells), were identical to the equivalentregions in the known NP-lacking Cav2.1 cDNAs (Fig. 1). Noneof the products contained exon 31a and none displayed any ofthe novel forms of splicing between exons 28 and 34 detectedin the cerebellum. Therefore, splicing in this region does notexplain our inability to amplify cDNAs from Purkinje cells
Fig. 4. Genotyping of cerebellar Purkinje cells. The PCR products shown inthe agarose gels (negative images) were amplified from cDNA of the cerebellarvermis, a single Purkinje cell, and a single granule cell. A: all cDNAs weredetected in the cerebellar vermis. B: as predicted for a Purkinje cell, there wasamplification of calbindin D-28K and GAD65 but no amplification of GABAA
receptor-�6 and zipro 1. C: in direct contrast with B, and as predicted for agranule cell, there was amplification of GABAA receptor-�6 and zipro 1 butnot of calbindin D-28K and GAD65. In A, B, and C, there was the generationof an NR2 product, which serves as a positive control.
Fig. 5. Individual cerebellar Purkinje neurons express a smaller assortment ofmultiple Cav2.1 variants than the cerebellum. A: representative gel (negativeimage) depicting the amplification of multiple transcripts from a single ratcerebellar Purkinje cell with pairs of primers designed against the splicevariations indicated. B: histogram summarizing the frequency with which PCRproducts of expected size (Table 2) were generated by the pairs of primersindicated. Nos. in bars denote the no. of cells used for reactions with each pairof primers. Compare with histogram in Fig. 2. Asterisk (*) denotes that, in allsuccessful reactions with the ggcag forward primer and the e47 reverse primer,multiple products of distinct size were amplified. C: schematic diagram of thetranscripts detected. Solid lines, transcripts observed in 67–89% of cells;dotted lines, transcripts detected in 12–15% of cells. Asterisk (*) emphasizesthe generation of 3 products containing the ggcag insert. The numbered arrowsrefer to the sites of variation identified in Fig. 1.
92 CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

with the �NP and exon 37b splice combination. An alternativeexplanation that requires future investigation is that the se-quence of exon 37b in these elusive transcripts is not the sameas that in the cDNAs described to date and contains thesequence for the exon 37b forward primer but not for the exon37b reverse primer, which is located downstream from theforward primer.
Our finding that the majority of CaV2.1 transcripts in Pur-kinje cells lack the NP-encoding exon is consistent with elec-trophysiological studies reporting almost complete block (90–95%) of Ca2� channel currents in the soma of rat Purkinje cells(22) by concentrations of �-agatoxin IVA considered to beselective for P-type (�100 nM) over Q-type channels (�100nM) (21, 29, 34). However, because these previous electro-physiological studies were carried out on cells isolated fromimmature rats (20, 22), whereas our study was on CaV2.1splicing in Purkinje cells in adult rat, we examined the sensi-tivity of Ca2� channels in mature Purkinje cells to 30 nM�-agatoxin IVA. Cell-attached recordings of Ca2� channelcurrents (Fig. 6A) from the soma or the first dendritic bifurca-tion with drug-free pipettes or drug-containing pipettes (6, 36)demonstrated inhibition of the majority of the somatic Ca2�
channel current (92%, Fig. 6B) and of the dendritic current(91%, Fig. 6C). Increasing the concentration 10-fold to 300 nMdid not increase block of somatic currents (n � 12), whichsuggests that other types of �-agatoxin IVA-sensitive chan-nels, such as Q-type channels, make no clear contribution tothe remaining somatic current. It remains possible, however,that a higher concentration of �-agatoxin IVA might haveblocked this current, but we did not investigate further thepharmacological identity of this current or of the dendriticcurrent resistant to block by 30 nM �-agatoxin IVA.
Identity of CaV2.1 COOH-termini. The largest of the PCRproducts containing the ggcag insert (exon 47) amplified fromindividual Purkinje cells (n � 5 cells) was identical in se-quence (EMBL-Bank, AM040229) to that of the ggcag-con-taining exon 47 of the rat pancreatic AF05126 variant (14)(Fig. S4, Supplemental Results). The smaller product repre-sents a novel transcript that contains a deletion of 123 nucle-otides (Fig. S4; EMBL-Bank, AM040228). We identified thesame exon 47 variants in cerebellar cDNA [and also confirmedthe cerebellar expression of a transcript variant with a stopcodon (tag) at the start of exon 47 (not shown), as in the ratbrain M64373 variant; Fig. 1], using PCR primers flanking the5�-end of exon 47 and the 123-nucleotide deletion (sequencenot shown). The 123-nucleotide deletion is downstream fromthe epitope for an exon 47-specific antibody used previously todetect the translation of exon 47 in rat cerebellar Purkinje cellsand in multiple rat cerebellar CaV2.1 protein isoforms (28). Itis in a similar position to the 150-nucleotide deletion in theAB066608 variant cloned from mouse Purkinje neurons (35).The similar location supports the notion that the deletions ariseby alternative splicing (35), but we could not compare thecDNA sequences with mouse or rat CACNA1A genes, becausethis region of the genes is incomplete in public genomicdatabases. The predicted amino acid sequences for the twoexon 47 variants (Fig. 7) imply that, like mouse cerebellarPurkinje cells (35), rat cerebellar Purkinje cells may express atleast two Cav2.1 variants with extended COOH-termini. How-ever, these have little amino acid homology with the extendedCOOH-termini in cerebellar Purkinje cells of the mouse (Fig.
7) or with extended COOH-termini of human or rabbit CaV2.1transcripts (9, 10, 13, 23, 28, 30, 33, 41).
DISCUSSION
This study identifies multiple transcript variants of theCaV2.1 subunit in rat cerebellum, which are distinguished bydifferent combinations of splice events and by novel splicingevents. Our detection of a variety of splice variations in ratcerebellum (�V or �VG or VG, exon 10; �NP or NP, exon
Fig. 6. Confirmation that the majority of Ca2� channel currents in mature ratcerebellar Purkinje cells is inhibited by P-type-selective concentrations of�-agatoxin IVA. A: example of cell-attached recording. The black trace is themean of 30 currents (superimposed gray traces) evoked by a voltage ramp (top)applied at 0.2 Hz to a single patch. The leak current (dotted line) was estimatedby fitting a straight line to the linear part of the mean current and extrapolatingit over the full range of the ramp. B: average current-voltage relationshipsrecorded with drug-free pipette solution from 24 somatic patches (each patchwas in a different cell) and with pipettes containing 30 nM �-agatoxin IVAfrom 26 somatic patches. These were obtained by subtracting the leak currentfrom the mean current obtained from each patch, plotting the current againstpatch potential, and averaging the resultant current-voltage relationships acrosspatches. Vertical bars represent SE and, for clarity, are shown every 10 mVrather than for every data point. C: average current-voltage relationshipsrecorded with drug-free pipette solution from 32 dendritic patches and withpipettes containing 30 nM �-agatoxin IVA from 9 dendritic patches.
93CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

31a; exon 37a or exon 37b; exon 44; ggcag or �ggcag, exon47) confirms previous studies of rat cerebellar CaV2.1 mRNA,cDNA, and protein (2, 5, 28, 37). We also detect the lack ofexon 44 in some transcripts, which was known to occur in ratpancreas (14) but not in rat cerebellum. CaV2.1 splice varia-tions not previously documented in rat cerebellum, or in anyother tissue or species, are the introduction of alternativepremature stop codons shortly after the NP insert via theincorporation of alternative sections of the intron betweenexons 31a and 32, the inclusion of the intron between exons 33and 34, the coincident exclusion of exons 29–31a, or thedeletion of 123 nucleotides from within exon 47. We find thattwo alternative splicing events at exon 31a (NP or �NP) andexon 37 (a or b) occur in all four possible combinations.Likewise, two alternative splicing events at exon 37 (a or b)and exon 44 (exon 44 or �exon 44) occur in all four possiblecombinations. Different patterns of splicing at these three sites(exon 31a, exon 37, exon 44) may therefore generate eightdistinct transcript variants. The actual number of variants in ratcerebellum may be higher, if each of the eight splicing patternscan take place within a transcript with any of the splice eventsidentified at other sites, as well as with splice events notidentified in this study.
Multiple CaV2.1 transcript variants are also present in indi-vidual mature Purkinje cells, but the range of splice variationsis smaller than in the cerebellum: �V (exon 10), �NP (exon31a), exon 37a or b, exon 44 or �exon 44, ggcag insert (exon47), deletion of 123 nucleotides (exon 47). All of the splicingevents and patterns identified in Purkinje cells occur in anindividual cell. There appears to be no mutual exclusion of thetwo extended versions of exon 47, or of exon 37a and exon37b, from individual neurons. Recent work (4) has suggestedthat exon 37a-containing transcripts predominate in Purkinjecells of young animals (P12), and in adult Purkinje cells, exon37a protein is restricted to the soma, whereas exon 37b proteinis in the soma and in dendrites. Our finding that the majority of
CaV2.1 transcripts in mature rat Purkinje cells lack the NP-encoding exon is in agreement with an earlier prediction (2)and with the isolation of transcripts lacking the NP-encodingexon from mouse Purkinje cells (33, 35). The scarcity in ratPurkinje cells of splicing events that introduce a premature stopcodon after the NP-encoding exon 31a in some cerebellartranscripts predicts a higher level of CaV2.1 protein expressionin Purkinje cells than in other cerebellar cells, if the truncatedCaV2.1 proteins encoded by these transcripts act in a dominantnegative manner to inhibit the expression of full-length CaV2.1proteins (12, 24, 25). A further difference between the cere-bellum and Purkinje cells is that, in Purkinje cells, the twosplice events at exon 37 (a or b) and the two splice events atexon 44 (inclusion or exclusion) do not occur in all fourpossible combinations. The exon 37a/�exon 44 combinationwas not detected.
Our finding that the exon 47 variants in cerebellar Purkinjecells of the rat are not the same as those in cerebellar Purkinjecells of the mouse (35) suggests species-specific alternativesplicing at the level of the individual type of cell. The expres-sion of the exon 47 variants found in rat or mouse Purkinjecells has not been intentionally explored in human cerebellarPurkinje cells, but none of the CaV2.1 cDNAs cloned fromhuman cerebellum contains these variations. Differences in theCOOH-terminus of CaV2.1 cDNAs obtained from varioustissues and species indicate that this is a region of functionalspecialization that may underly species-specific differences inCaV2.1 channel expression and function (12, 17). We do notyet know what influence the predicted extended COOH-terminihave on the expression or function of Ca2� channels in ratcerebellar Purkinje cells. However, the two extended COOH-termini found in mouse cerebellar Purkinje cells have no effecton the electrophysiological properties or the sensitivity toblock by �-agatoxin IVA (35). Alternative roles for differentCOOH-termini may be the differential subcellular localizationof the variants or differential interaction of the variants with
Fig. 7. Distinct amino acid sequences of predicted alternative extended CaV2.1 termini in rat cerebellar Purkinje cells. Predicted amino acid sequences for theggcag-containing cDNAs amplified from cerebellar Purkinje cells (i and ii) are compared with the equivalent regions in the extended COOH-termini in a ratpancreatic variant and in 2 mouse cerebellar Purkinje cell variants (protein accession nos. indicated). The vertical arrow denotes the exon 46/47 boundary. Grayshading highlights identical amino acids in 2 or more sequences. Asterisk (�) identifies a stop codon.
94 CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

intracellular proteins (19). Intriguingly, the position of the123-nucleotide deletion in rat cerebellar Purkinje cells and the150-nucleotide deletion in mouse cerebellar Purkinje cellscoincides with the nucleotides encoding a polyglutamine tractin some human CaV2.1 protein isoforms (11, 41). Expansion ofthis tract by a gene mutation is associated with the neurologicalcondition spinocerebellar ataxia 6 and selective degenerationof Purkinje cells (10, 41).
We have not yet investigated which of the transcript variantsidentified in rat cerebellum and rat Purkinje cells generatefunctional Ca2� channels, nor the influence of the differentpatterns of splicing on channel properties. However, previousstudies have investigated the functional impact of alternativesplicing at exon 10, exon 31a, exon 37, or exon 44. They haveshown that CaV2.1 channels without V in the I-II linkerinactivate more rapidly, have briefer openings, are morereadily blocked by G proteins, and are upregulated less byprotein kinase C than channels with VG in the I-II linker (2).The absence of NP (exon 31a) in the IVS3-4 linker enhancesthe affinity of the channel for �-agatoxin IVA, shifts thevoltage dependence of activation and inactivation to morenegative potentials, and speeds inactivation kinetics (2, 9, 16,33). Inclusion of exon 37, a or b, generates two variants of anEF hand-like domain in the COOH-terminus that alter calcium/calmodulin-dependent facilitation of the channels, without af-fecting calcium/calmodulin-dependent inhibition (5). Exclu-sion of exon 44 from the COOH-terminus results in more rapidinactivation kinetics (13). These previous studies suggest thatthe multiplicity of transcript variants in the cerebellum thatresults from different combinations of splice events at multipleloci may be manifest as a broad range of phenotypic variants ofP-type and Q-type Ca2� channels with different but overlap-ping functional profiles (2). The smaller transcript diversity inPurkinje cells, together with the relatively low abundance inPurkinje cells of NP-containing transcripts (Ref. 35 and thisstudy) and the block of 90% of Ca2� channel currents inimmature (20) and adult Purkinje cells (this study) by P-type-selective concentrations of �-agatoxin IVA, predicts the ex-pression of subtypes of P-type channels in cerebellar Purkinjecells with different functional properties. These may underliethe distinct Ca2� channel currents previously identified incell-attached recordings from cerebellar Purkinje cells (Refs. 6,7, and 36 and E. W. Tringham, C. E. Payne, and M. M.Usowicz, unpublished observations).
ACKNOWLEDGMENTS
M. M. Usowicz is particularly grateful to A. C. Dolphin and K. M. Page forpractical training in the early stages of this project. We are grateful fortechnical advice from L. Hall, J. Rossier, and B. Cauli. We thank B. Ligon, K.Dunlap, and T. P. Snutch for the gifts of CaV2.1 cDNA clones. A fewexperiments were carried out by R. Stewart or H. Gurney.
Present address of E. W. Tringham: NeuroMed Technologies Inc., Don RixBldg., 301-2389 Health Sciences Hall, Vancouver, British Columbia, V6T1Z4, Canada.
GRANTS
This work was supported by the United Kingdom Medical ResearchCouncil (MRC) and the Royal Society. E. W. Tringham was in receipt of aUniversity of Bristol PhD scholarship. C. E. Payne was in receipt of an MRCPhD scholarship. J. R. B. Dupere was in receipt of a Wellcome Trust PhDscholarship. K. Venkateswarlu was supported by a David Phillips ResearchFellowship from the United Kingdom Biotechnology and Biological SciencesResearch Council.
REFERENCES
1. Bastianelli E. Distribution of calcium-binding proteins in the cerebellum.Cerebellum 2: 242–262, 2003.
2. Bourinet E, Soong TW, Sutton K, Slaymaker S, Mathews E, MonteilA, Zamponi GW, Nargeot J, and Snutch TP. Splicing of �1A subunitgene generates phenotypic variants of P- and Q-type calcium channels.Nat Neurosci 2: 407–415, 1999.
3. Catterall WA. Structure and regulation of voltage-gated Ca2� channels.Annu Rev Cell Dev Biol 16: 521–555, 2000.
4. Chaudhuri D, Alseikhan BA, Chang SY, Soong TW, and Yue DT.Developmental activation of calmodulin-dependent facilitation of cerebel-lar P-type Ca2� current. J Neurosci 25: 8282–8294, 2005.
5. Chaudhuri D, Chang SY, DeMaria CD, Alvania RS, Soong TW, andYue DT. Alternative splicing as a molecular switch for Ca2�/calmodulin-dependent facilitation of P/Q-type Ca2� channels. J Neurosci 24: 6334–6342, 2004.
6. Dupere JR, Moya E, Blagbrough IS, and Usowicz MM. Differentialinhibition of Ca2� channels in mature rat cerebellar Purkinje cells bysFTX-3.3 and FTX-33. Neuropharmacology 35: 1–11, 1996.
7. Dupere JRB and Usowicz MM. Multiple types of Ca2� channels in thesoma and dendrites of adult rat cerebellar Purkinje cells. J Physiol 494P:84–85, 1996.
8. Fletcher CF, Lutz CM, O’Sullivan TN, Shaughnessy JD Jr, HawkesR, Frankel WN, Copeland NG and Jenkins NA. Absence epilepsy intottering mutant mice is associated with calcium channel defects. Cell 87:607–617, 1996.
9. Hans M, Urrutia A, Deal C, Brust PF, Stauderman K, Ellis SB,Harpold MM, Johnson EC, and Williams ME. Structural elements indomain IV that influence biophysical and pharmacological properties ofhuman �1A-containing high-voltage-activated calcium channels. Biophys J76: 1384–1400, 1999.
10. Ishikawa K, Fujigasaki H, Saegusa H, Ohwada K, Fujita T, IwamotoH, Komatsuzaki Y, Toru S, Toriyama H, Watanabe M, Ohkoshi N,Shoji S, Kanazawa I, Tanabe T, and Mizusawa H. Abundant expressionand cytoplasmic aggregations of �1A voltage-dependent calcium channelprotein associated with neurodegeneration in spinocerebellar ataxia type 6.Hum Mol Genet 8: 1185–1193, 1999.
11. Jodice C, Mantuano E, Veneziano L, Trettel F, Sabbadini G, Calan-driello L, Francia A, Spadaro M, Pierelli F, Salvi F, Ophoff RA,Frants RR, and Frontali M. Episodic ataxia type 2 (EA2) and spinocer-ebellar ataxia type 6 (SCA6) due to CAG repeat expansion in theCACNA1A gene on chromosome 19p. Hum Mol Genet 6: 1973–1978,1997.
12. Jurkat-Rott K and Lehmann-Horn F. The impact of splice isoforms onvoltage-gated calcium channel �1 subunits. J Physiol 554: 609–619, 2004.
13. Krovetz HS, Helton TD, Crews AL, and Horne WA. C-terminalalternative splicing changes the gating properties of a human spinal cordcalcium channel �1A subunit. J Neurosci 20: 7564–7570, 2000.
14. Ligon B, Boyd AE 3rd, and Dunlap K. Class A calcium channel variantsin pancreatic islets and their role in insulin secretion. J Biol Chem 273:13905–13911, 1998.
15. Lin Z, Haus S, Edgerton J, and Lipscombe D. Identification of func-tionally distinct isoforms of the N-type Ca2� channel in rat sympatheticganglia and brain. Neuron 18: 153–166, 1997.
16. Lin Z, Lin Y, Schorge S, Pan JQ, Beierlein M, and Lipscombe D.Alternative splicing of a short cassette exon in �1B generates functionallydistinct N-type calcium channels in central and peripheral neurons. J Neu-rosci 19: 5322–5331, 1999.
17. Lipscombe D, Pan JQ, and Gray AC. Functional diversity in neuronalvoltage-gated calcium channels by alternative splicing of Cav�1. MolNeurobiol 26: 21–44, 2002.
18. Liu L, Zwingman TA, and Fletcher CF. In vivo analysis of voltage-dependent calcium channels. J Bioenerg Biomembr 35: 671–685, 2003.
19. Maximov A, Sudhof TC, and Bezprozvanny I. Association of neuronalcalcium channels with modular adaptor proteins. J Biol Chem 274:24453–24456, 1999.
20. Mintz IM, Adams ME, and Bean BP. P-type calcium channels in ratcentral and peripheral neurons. Neuron 9: 85–95, 1992.
21. Mintz IM and Bean BP. Block of calcium channels in rat neurons bysynthetic omega-Aga-IVA. Neuropharmacology 32: 1161–1169, 1993.
22. Mintz IM, Venema VJ, Swiderek KM, Lee TD, Bean BP, and AdamsME. P-type calcium channels blocked by the spider toxin �-Aga-IVA.Nature 355: 827–829, 1992.
95CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org

23. Mori Y, Friedrich T, Kim MS, Mikami A, Nakai J, Ruth P, Bosse E,Hofmann F, Flockerzi V, and Furuichi T. Primary structure and func-tional expression from complementary DNA of a brain calcium channel.Nature 350: 398–402, 1991.
24. Okagaki R, Izumi H, Okada T, Nagahora H, Nakajo K, and OkamuraY. The maternal transcript for truncated voltage-dependent Ca2� channelsin the ascidian embryo: a potential suppressive role in Ca2� channelexpression. Dev Biol 230: 258–277, 2001.
25. Page KM, Heblich F, Davies A, Butcher AJ, Leroy J, Bertaso F, PrattWS, and Dolphin AC. Dominant-negative calcium channel suppressionby truncated constructs involves a kinase implicated in the unfoldedprotein response. J Neurosci 24: 5400–5409, 2004.
26. Pan Q, Bakowski MA, Morris Q, Zhang W, Frey BJ, Hughes TR, andBlencowe BJ. Alternative splicing of conserved exons is frequentlyspecies-specific in human and mouse. Trends Genet 21: 73–77, 2005.
27. Pietrobon D. Calcium channels and channelopathies of the central ner-vous system. Mol Neurobiol 25: 31–50, 2002.
28. Restituito S, Thompson RM, Eliet J, Raike RS, Riedl M, Charnet P,and Gomez CM. The polyglutamine expansion in spinocerebellar ataxiatype 6 causes a � subunit-specific enhanced activation of P/Q-type calciumchannels in Xenopus oocytes. J Neurosci 20: 6394–6403, 2000.
29. Sidach SS and Mintz IM. Low-affinity blockade of neuronal N-type Cachannels by the spider toxin �-agatoxin-IVA. J Neurosci 20: 7174–7182,2000.
30. Soong TW, DeMaria CD, Alvania RS, Zweifel LS, Liang MC, Mitt-man S, Agnew WS, and Yue DT. Systematic identification of splicevariants in human P/Q-type channel �12.1 subunits: implications forcurrent density and Ca2�-dependent inactivation. J Neurosci 22: 10142–10152, 2002.
31. Starr TV, Prystay W, and Snutch TP. Primary structure of a calciumchannel that is highly expressed in the rat cerebellum. Proc Natl Acad SciUSA 88: 5621–5625, 1991.
32. Thompson CL, Drewery DL, Atkins HD, Stephenson FA, and ChazotPL. Immunohistochemical localization of N-methyl-D-aspartate receptor
NR1, NR2A, NR2B and NR2C/D subunits in the adult mammaliancerebellum. Neurosci Lett 283: 85–88, 2000.
33. Toru S, Murakoshi T, Ishikawa K, Saegusa H, Fujigasaki H, UchiharaT, Nagayama S, Osanai M, Mizusawa H, and Tanabe T. Spinocere-bellar ataxia type 6 mutation alters P-type calcium channel function. J BiolChem 275: 10893–10898, 2000.
34. Tottene A, Moretti A, and Pietrobon D. Functional diversity of P-typeand R-type calcium channels in rat cerebellar neurons. J Neurosci 16:6353–6363, 1996.
35. Tsunemi T, Saegusa H, Ishikawa K, Nagayama S, Murakoshi T,Mizusawa H, and Tanabe T. Novel Cav2.1 splice variants isolated fromPurkinje cells do not generate P-type Ca2� current. J Biol Chem 277:7214–7221, 2002.
36. Usowicz MM, Sugimori M, Cherksey B, and Llinas R. P-type calciumchannels in the somata and dendrites of adult cerebellar Purkinje cells.Neuron 9: 1185–1199, 1992.
37. Vigues S, Gastaldi M, Massacrier A, Cau P, and Valmier J. The �1A
subunits of rat brain calcium channels are developmentally regulated byalternative RNA splicing. Neuroscience 113: 509–517, 2002.
38. Wisden W, Korpi ER, and Bahn S. The cerebellum: a model system forstudying GABAA receptor diversity. Neuropharmacology 35: 1139–1160, 1996.
39. Yang XW, Zhong R, and Heintz N. Granule cell specification in thedeveloping mouse brain as defined by expression of the zinc fingertranscription factor RU49. Development 122: 555–566, 1996.
40. Yu AS, Hebert SC, Brenner BM, and Lytton J. Molecular character-ization and nephron distribution of a family of transcripts encoding thepore-forming subunit of Ca2� channels in the kidney. Proc Natl Acad SciUSA 89: 10494–10498, 1992.
41. Zhuchenko O, Bailey J, Bonnen P, Ashizawa T, Stockton DW, AmosC, Dobyns WB, Subramony SH, Zoghbi HY, and Lee CC. Autosomaldominant cerebellar ataxia (SCA6) associated with small polyglutamineexpansions in the �1A-voltage-dependent calcium channel. Nat Genet 15:62–69, 1997.
96 CAV2.1 SPLICING IN CEREBELLUM AND IN PURKINJE NEURONS
Physiol Genomics • VOL 24 • www.physiolgenomics.org
Copyright © 2022 FDOKUMEN




















