
Life-history traits of the endangered river blenny Salaria fluviatilis (Asso) and their implications...
21
Life-history traits of the endangered river blenny Salaria fluviatilis (Asso) and their implications for conservation D. VINYOLES* AND A. DE SOSTOA Departament de Biologia Animal (Vertebrats), Facultat de Biologia, Universitat de Barcelona, Avda. Diagonal 645, 08028 Barcelona, Spain (Received 31 July 2005, Accepted 9 November 2006) The life-history characteristics of the river blenny Salaria fluviatilis in a Mediterranean stream in north-east Spain have been studied. The maximum age observed was 4þ years in both sexes, although only age groups 0þ and 1þ years were well represented. For both sexes, growth was rapid between April and June (before the summer drought) and in October. The instantaneous rate of monthly growth (April to October) was positively correlated with water depth. All fish attained sexual maturity during their first year of life. Females spawned multiple times. Spawning occurred mainly from June to August, although older females (2þ years old) began in May. The relationship between fecundity and total length recorded for older females in the spring was not significantly different from that obtained for 1 year-old females in the summer. The mean diameter of ripe oocytes, however, was greater in older females. Larger eggs that were laid during a period of good food supply (spring) might have a higher probability of survival. The results suggest that female fecundity is affected by environmental conditions. The species is characterized by fast growth, early maturity, multiple spawning and limited longevity. This may be a good life-history strategy for small fish species in unstable environments, where adult mortality is high, variable or unpredictable. Some life-history traits of the river blenny suggest that this species might be vulnerable to more severe drought conditions than those associated with the Mediterranean climate. # 2007 The Fisheries Society of the British Isles Key words: age and growth; Blennius fluviatilis; fluctuating streams; fresh water; Mediterranean- type climate; reproduction. INTRODUCTION Many freshwater animal species are in sharp decline (Richter et al., 1997; Ricciardi & Rasmussen, 1999). Freshwater fishes, in particular, may be one of the most threatened groups of vertebrates (Bruton, 1995). The decline of freshwater fishes due to anthropogenic action is currently a matter of concern (Williams & Miller, 1990; Cambray & Bianco, 1998). In Europe, more than half of freshwater fish species are threatened, and actions to ensure their con- servation are required at all levels. Familiarity with the life-history patterns of *Author to whom correspondence should be addressed. Tel.: þ34 934021448; fax: þ34 934035740; email: [email protected] Journal of Fish Biology (2007) 70, 1088–1108 doi:10.1111/j.1095-8649.2007.01371.x, available online at http://www.blackwell-synergy.com 1088 # 2007 The Fisheries Society of the British Isles
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Life-history traits of the endangered river blenny Salaria fluviatilis (Asso) and their implications...

Life-history traits of the endangered river blennySalaria fluviatilis (Asso) and their implications
for conservation
D. VINYOLES* AND A. DE SOSTOA
Departament de Biologia Animal (Vertebrats), Facultat de Biologia, Universitat deBarcelona, Avda. Diagonal 645, 08028 Barcelona, Spain
(Received 31 July 2005, Accepted 9 November 2006)
The life-history characteristics of the river blenny Salaria fluviatilis in a Mediterranean stream in
north-east Spain have been studied. The maximum age observed was 4þ years in both sexes,
although only age groups 0þ and 1þ years were well represented. For both sexes, growth was
rapid between April and June (before the summer drought) and in October. The instantaneous
rate of monthly growth (April to October) was positively correlated with water depth. All fish
attained sexual maturity during their first year of life. Females spawned multiple times.
Spawning occurred mainly from June to August, although older females (2þ years old) began
in May. The relationship between fecundity and total length recorded for older females in the
spring was not significantly different from that obtained for 1 year-old females in the summer.
The mean diameter of ripe oocytes, however, was greater in older females. Larger eggs that were
laid during a period of good food supply (spring) might have a higher probability of survival.
The results suggest that female fecundity is affected by environmental conditions. The species is
characterized by fast growth, early maturity, multiple spawning and limited longevity. This may
be a good life-history strategy for small fish species in unstable environments, where adult
mortality is high, variable or unpredictable. Some life-history traits of the river blenny suggest
that this species might be vulnerable to more severe drought conditions than those associated
with the Mediterranean climate. # 2007 The Fisheries Society of the British Isles
Key words: age and growth; Blennius fluviatilis; fluctuating streams; fresh water; Mediterranean-
type climate; reproduction.
INTRODUCTION
Many freshwater animal species are in sharp decline (Richter et al., 1997;Ricciardi & Rasmussen, 1999). Freshwater fishes, in particular, may be oneof the most threatened groups of vertebrates (Bruton, 1995). The decline offreshwater fishes due to anthropogenic action is currently a matter of concern(Williams & Miller, 1990; Cambray & Bianco, 1998). In Europe, more thanhalf of freshwater fish species are threatened, and actions to ensure their con-servation are required at all levels. Familiarity with the life-history patterns of
*Author to whom correspondence should be addressed. Tel.: þ34 934021448; fax: þ34 934035740;
email: [email protected]
Journal of Fish Biology (2007) 70, 1088–1108
doi:10.1111/j.1095-8649.2007.01371.x, available online at http://www.blackwell-synergy.com
1088# 2007 The Fisheries Society of the British Isles

endangered species is essential for the design and implementation of effectivemanagement and conservation measures. Surprisingly, very little is knownabout the life history of many species in need of protection. Practical aspectsof conservation biology, such as captive breeding and recovery programmes,require information about natural populations, so as to ensure that endangeredspecies are efficiently maintained and enhanced (Caro, 1998). Life-history var-iables are also needed in behavioural and ecological studies to parameterizeconservation models (Goss-Custard & Sutherland, 1997). Indeed, the life his-tory of a species has important implications for predicting the way it willrespond to the timing and type of management applied (Sutherland, 1995).The river blenny Salaria fluviatilis (Asso) was long believed to be the only
freshwater member of a large marine family of small benthic fishes. Recently,another freshwater blenny species Salaria economidisi Kottelat, endemic toLake Trichonis (Greece), has been described (Kottelat, 2004). The river blennyappears in rivers and lakes in the Mediterranean Sea basin. During the breedingseason, males excavate a nest cavity under a stone, and several females layclutches of adhesive eggs in a monolayer under the stone (Wickler, 1957). Afterfertilization, males provide sole care to the eggs by fanning and defending themuntil they hatch. Specific habitat requirements in streams consist of moderateto fast-flowing sections with a mixture of gravel and stones, with males spawn-ing preferentially under large stones (Freeman et al., 1990). Cote et al. (1999)have reported the negative effects of stone and gravel extraction on the repro-ductive success of this species. Even though many studies have focused on theriver blenny (Vinyoles et al., 1999, 2002), little information about basic aspectsof its biology has been published. The only data available relate mainly to thegonadal cycle of the new species S. economidisi in Lake Trichonis (Psarraset al., 1997). Key biological traits of both species, however, remain unknown.The circum-Mediterranean distribution of the river blenny make it a subject
of interest from a conservational point of view. In five out of nine northernMediterranean countries where this species occurs, it has been classified as vul-nerable or endangered (Balik, 1995; Bianco, 1995; Changeux & Pont, 1995;Elvira, 1995a; Mrakovcic et al., 1995). Its conservation status has not been as-sessed in three other countries due to a lack of available information. Duringthe 1990s, the species disappeared from a large portion of freshwater environ-ments in eastern Sicily as a result of habitat destruction and the deteriorationof water quality (Ferrito & Tigano, 1996). Healthy populations of river blen-nies appear to occur only in Greece (Economidis, 1995). In the Iberian Penin-sula, it is restricted to six drainage basins (Elvira, 1995b; Doadrio, 2001) and isclassified as endangered (Blanco & Gonzalez, 1992). The decline of river blennypopulations was also noted during the last decade in freshwater habitats ofnorth-eastern Spain, leading to the isolation of small populations (Aparicioet al., 2000). Reasons for the decline and current threats in Spain and elsewhereare habitat loss, water pollution and the introduction of exotic fish predators(Elvira, 1995b; Hernandez et al., 2000), as well as excessive water extraction(Changeux & Pont, 1995; Collares-Pereira et al., 2000). Recently, laboratoryexperiments have shown that the river blenny is a suitable host for the glo-chidia of the freshwater mussel Margaritifera auricularia (Spengler) (Araujoet al., 2001), a species living in the lower section of the River Ebro that has been
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1089
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

classified as critically endangered by the IUCN, suggesting that the disappear-ance of river blennies could lead to secondary species losses.The river blenny inhabits mainly the intermediate and lower reaches of Med-
iterranean Sea rivers. Generally, these systems have strong seasonal fluctua-tions in water and flow levels, temperature and resource availability. Severalauthors have studied how freshwater fish species inhabiting Mediterranean Searivers cope with these harsh conditions (particularly summer droughts) (Encina &Granado-Lorencio, 1997; Pires et al., 2000; Ribeiro et al., 2000; Oliveira et al.,2002; Oliva-Paterna et al., 2003). In this context, the life history of river blenniesis of particular interest because, unlike most other sympatric species, it providesparental care. The aim of this study was to provide information for the first timeon the age, growth and reproduction of a population of river blenny inhabitinga Spanish stream under Mediterranean climate. The significance of life-historytraits for its future conservation will be also discussed.
MATERIALS AND METHODS
STUDY AREA
This study was part of a larger research programme on fish communities of the RiverMatarranya that began in 1984, 8 years before the designation of river blennies asa threatened species in Spain (Blanco & Gonzalez, 1992). Although river blenny pop-ulations are usually rare, this species is still abundant in the study area (Fayon site)today. The River Matarranya is a tributary of the River Ebro in north-eastern Spain.The river is c. 100 km long and is subject to a Mediterranean-type climate, with floodsand summer droughts. The sampling site was located c. 10 km from the river mouth, inan area where the river width was highly variable (from 4 to 15 m during the study)and where water depth was moderate (from 5 to 40 cm). The riverbed consisted mainlyof stones and gravel. The flow varied between 17 and 25 cm s�1 during most of thestudy period, except in the summer when some stretches of river became isolated pools,and during frequent floods in spring and autumn. During droughts, which occurred inJuly, August, September and even October, hydrological connectivity was disrupted.Such disruption varied from flow reduction to complete loss of surface water connec-tivity. Water depth during the summer of 1984 (August and September) was only 5cm. Water temperature in the summer reached 35° C (August), while in the winterwas as low as 8° C (January). Dissolved oxygen was usually close to saturation, withthe exception of isolated pools during the summer where minimum values of 6�3ppm were recorded. Dissolved oxygen concentration was undersaturated from July toSeptember. These harsh conditions are responsible for the sparse fish community pres-ent at this site, when compared with the upper reaches of the Matarranya (Sostoa et al.,1990). River blennies were the most abundant species in the study area both during thestudy period (Vinyoles, 1993) and at present (unpubl. data).
The cycles of rainfall andwater temperature shown inFig. 1 reflect the drought and rainyperiods that characterize the study area. Due to the large amount of rainfall during theautumn of 1983 and the spring rains in 1984, the river was relatively deep (30–40 cm) andflow was rapid (20–52 cm s�1) between January and June 1984. Thereafter, however, theamount of rainfall was not sufficient to re-establish winter flow after the severe summerdrought of 1984, despite two rainfall peaks in November 1984 and May 1985. Hence,although depth was 35 cm in November 1984, it was only 10 cm in April 1985 with littleor no flow. In May 1985, after the spring rains, the water column barely reached 20 cm,the same depth as during the first month of summer drought the previous year (July1984). In a river subject to a highly variable rainfall regime such as the Matarranya, waterdepth instead of monthly rainfall values as a good predictor of water flow has been chosen.
1090 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

DATA COLLECTION
A total of 1098 specimens (584 males, 463 females and 51 unsexed individuals) werecaught by electrofishing at monthly intervals from January 1984 to July 1985. Monthlysampling was always performed within the same river section (c. 100 m long). Afterbeing caught, specimens were preserved in dry ice and transported to the laboratorywhere they were measured (total length, LT, � 1 mm) and weighed (total mass, MT �0�01g), and gonads removed and weighed (gonad mass, MG � 0�01g). Sex was deter-mined by the examination of external characteristics (i.e. presence of anal glands inmales). Since young males of many blenniids do not have well developed anal glands,and could thus be erroneously classified as females, gonads were also inspected whenthe study specimens were dissected.
To determine age, a sub-sample of 765 opercular bones from 423 males and 342females were removed from the left side of the fish. The opercular bones were mounteddry between two slides for examination with a microfilm projector. Following Craig(1974) and Bagenal & Braum (1978), the use of the opercular bones for age determina-tion was validated by analysing the marginal growth of the opercular bones to testwhether increments on the bones were annual. Age validation was performed for eachsex using a length-frequency based method using the software Complete ELEFAN(Gayanillo et al., 1988). First, size categories representing age classes were identifiedusing the method of Bhattacharya (1967). A modal class progression analysis was thenapplied, in which an index >2 must be obtained to provide meaningful separations(Gayanillo et al., 1988). Due to the small sizes of the monthly samples, age validation
Rai
nfal
l (m
m)
Wat
er te
mpe
ratu
re (
º C
)
Jan.Mar.
MayJuly
Sep.Nov.
Jan.Mar.
MayJuly
Sep.Nov.
Jan.Mar.
MayJuly
Sep.Nov.
Jan.Mar.
MayJuly
Sep.Nov.
0
20
40
60
80
100
120
140
160
0
5
10
15
20
25
30
1982 1983 1984 1985
FIG. 1. Ombrothermal diagram of the lower course of the Matarrana, showing the water temperature
( ) and rainfall per month ( ) between 1982 and 1985. Periods of floods (>50 mm of
precipitation in 1 day) are marked ( ).
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1091
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

was performed for seasons. The seasonal growth pattern was described using the aver-age and monthly S.D. of LT for each cohort and sex. The instantaneous growth rate (G)was estimated as: G ¼ (ln LT2 � ln LT1) (t2 � t1)
�1, where LT1 and LT2 are the meantotal length (mm) at time t1 and t2 (i.e. 1 month).
Age and LT at first maturity were determined for 161 male individuals. Maturity wasestablished through testicle smear. Males were considered to be sexually mature whenthe proportion of spermatozoids was larger than that of germinal cells. In female indi-viduals, age and LT at first maturity were determined when ripe oocytes appeared in theovaries.
The gonadal cycle was described in terms of the gonado-somatic index (IG) with IG ¼100 MG MT
�1. Monthly mean � S.D. were calculated for each sex.Oocyte development and female fecundity were examined using the gravimetric
method (Bagenal & Braum, 1978). For this purpose, 207 females which include samplesfor all months of the study were considered. One ovary from each female was trans-ferred into Gilson’s fluid. Once the gonads had been separated, oocytes were pouredinto a column of 15 sieves with a range of mesh-sizes (between 1�0 and 0�1 mm) andflushed with running water to facilitate their descent. Based on oocyte diameter fre-quency distributions, developmental stages corresponding to previtellogenic (immature)and vitellogenic (ripening, or in maturation process, and ripe) oocytes were identified.Fecundity, defined here as the total number of ripening or vitellogenic oocytes in theovaries prior to spawning (Bagenal, 1978), was determined by counting all vitellogenicoocytes in gravid females (i.e. in females with ripe oocytes). In order to summarize theresults obtained, only the most representative months of the reproductive cycle wereconsidered. Fecundity (F) was measured as: ln F ¼ a þ b ln LT, LT is in mm. Femalesfor which the onset of spawning could not be confirmed were excluded from the anal-ysis. The fecundities and average diameter of eggs laid (ripe oocytes) of young (femalesthat were reproducing for the first time) and older (females that had reproduced at leastonce in previous years) females were compared.
Parametric statistics were used except when departure from normality or lack ofhomogeneity of variance required non-parametric tests. When multiple tests were per-formed, significance levels were corrected using the sequential Bonferroni method (Rice,1989). Real probability values are reported throughout, and Bonferroni correctionsnever overturned significant results.
RESULTS
AGE DETERMINATION
During the 2 years of study, annuli formation in river blennies occurredbetween the end of April (44% of individuals both in 1984 and 1985) andthe end of May (47 and 63% of individuals, respectively). The birth date ofnew cohorts was therefore considered to be 15 May. Most individuals showeda false check in autumn. False checks formed in previous years were observedin the opercular bones of old specimens, making the detection of annuli moredifficult. Autumn checks could be distinguished from true checks because theformer did not form a complete check around the opercular bones. Most speci-mens in the sub-sample (388 males and 323 females) were dated successfully.Individuals not showing clear checks (35 males and 19 females; 7% of thesub-sample) were omitted from age determination.Five different age groups (from 0þ to 4þ years) were observed in this pop-
ulation, although only three individuals (two males and one female) belongedto age group 4þ years (0�5%; Table I). The oldest age groups were poorly rep-resented (7% of individuals belonged to age group 2þ years and 1�5% to age
1092 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

group 3þ years). Age group 0þ years (53% of individuals) was the most abun-dant, followed by age group 1þ years (38%). Although the oldest females livedas long as the oldest males, a lower proportion of females was detected ingroups older than 1þ years (w2, d.f. ¼ 1, P < 0�001).
SEASONAL GROWTH PATTERN
River blenny males were longer than females (mean LT � S.D., males: 47�2 �8�3, females: 43�1 � 8; unpaired t-test, d.f. ¼ 1045, P < 0�001). The largest male
TABLE I. Estimated age groups from the length-frequency data of males and femalesof the river blenny using BHATTACHARYA’s method followed by modal class pro-gression analysis; the separation index (S.I.) must be >2 to provide meaningfully
separated groups. Values are means � S.D.; sample sizes are given in parentheses
Sample Group LT (mm) S.I. Estimated age (years)
MalesWinter 1984 1 33�3 � 4�3 (16) 0þ
2 47�9 � 3�0 (23) 4�04 1þ3 53�7 � 1�0 (19) 2�90 2þ4 62�5 � 2�1 (9) 5�64 3þ
Spring 1 41�0 � 3�3 (36) 0þ2 54�2 � 3�6 (19) 3�86 1þ
Summer 1 34�5 � 1�6 (13) 0þ2 51�8 � 2�8 (72) 7�82 1þ3 61�5 � 2�4 (14) 3�77 2þ
Autumn 1 39�9 � 4�5 (116) 0þ2 54�1 � 4�6 (22) 3�13 1þ
Winter 1985 1 41�7 � 3�3 (92) 0þ2 60�0 � 2�1 (8) 6�72 1þ
Spring 1 45�4 � 2�9 (57) 0þ2 57�2 � 4�2 (6) 3�31 1þ
Summer 1 52�8 � 2�1 (45) 1þ2 59�3 � 1�2 (6) 3�93 2þ
FemalesWinter 1984 1 30�0 � 2�1 (36) 0þ
2 43�9 � 3�9 (37) 4�65 1þ3 55�0 � 2�6 (4) 3�39 2þ
Spring 1 40�1 � 4�8 (47) 0þ2 50�4 � 2�2 (25) 2�95 1þ
Summer 1 31�5 � 1�6 (7) 0þ2 48�9 � 2�5 (59) 8�37 1þ
Autumn 1 38�3 � 2�8 (87) 0þ2 55�5 � 4�9 (8) 4�46 1þ
Winter 1985 1 40�4 � 2�8 (62) 0þ2 58�3 � 3�4 (4) 5�80 1þ
Spring 1 40�4 � 2�9 (51) 0þSummer 1 45�0 � 3�3 (16) 1þ
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1093
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108
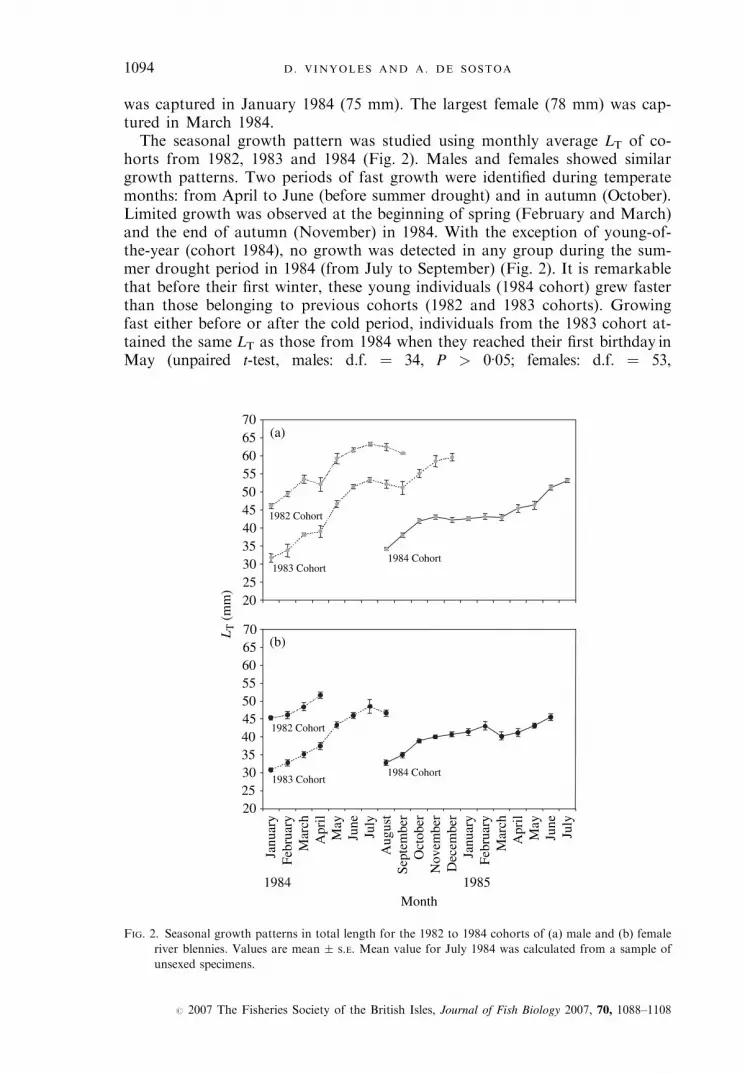
was captured in January 1984 (75 mm). The largest female (78 mm) was cap-tured in March 1984.The seasonal growth pattern was studied using monthly average LT of co-
horts from 1982, 1983 and 1984 (Fig. 2). Males and females showed similargrowth patterns. Two periods of fast growth were identified during temperatemonths: from April to June (before summer drought) and in autumn (October).Limited growth was observed at the beginning of spring (February and March)and the end of autumn (November) in 1984. With the exception of young-of-the-year (cohort 1984), no growth was detected in any group during the sum-mer drought period in 1984 (from July to September) (Fig. 2). It is remarkablethat before their first winter, these young individuals (1984 cohort) grew fasterthan those belonging to previous cohorts (1982 and 1983 cohorts). Growingfast either before or after the cold period, individuals from the 1983 cohort at-tained the same LT as those from 1984 when they reached their first birthday inMay (unpaired t-test, males: d.f. ¼ 34, P > 0�05; females: d.f. ¼ 53,
LT (
mm
)
(a)
1982 Cohort
1983 Cohort1984 Cohort
2025303540455055606570
2025303540455055606570
Mar
chA
pril
May
June
July
Aug
ust
Sept
embe
rO
ctob
erN
ovem
ber
Dec
embe
r
(b)
1984 Month
1985
1982 Cohort
1983 Cohort1984 Cohort
Janu
ary
Febr
uary
Mar
chA
pril
May
June
July
Janu
ary
Febr
uary
FIG. 2. Seasonal growth patterns in total length for the 1982 to 1984 cohorts of (a) male and (b) female
river blennies. Values are mean � S.E. Mean value for July 1984 was calculated from a sample of
unsexed specimens.
1094 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

P > 0�05). There was a positive correlation between monthly G (1983 and 1982cohorts) and water depth during the warm period (April to October 1984) (1983cohort: r2 ¼ 0�62, F1,5, P < 0�05; 1982 cohort: r2 ¼ 0�84, F1,4,P < 0�01; Fig. 3). No differences were detected between monthly G of malesand females during those months (paired t-test: d.f. ¼ 5, P > 0�05). Since waterdepth during the 1985 warm period (April to July) was not available, the rela-tionship between G and depth could not be examined for the 1984 cohort.
SEX RATIO
A small number of small specimens could not be sexed (n ¼ 51). The overallsex ratio (584 males:463 females) was significantly different from 1:1 (w2, d.f. ¼ 1,P < 0�001). Monthly differences in sex ratio were found in July (egg-layingperiod) in both years, with males outnumbering females (1984: w2, d.f. ¼ 1,P < 0�001; 1985: w2, d.f. ¼ 1, P < 0�001). Monthly biases were also found amongindividuals belonging to age group 0þ years (the dominant age group aftersummer 1984) in September 1984 (w2, d.f. ¼ 1, P < 0�001) and February 1985(w2, d.f. ¼ 1, P < 0�001), both with an over-representation of males.
AGE AND LENGTH AT FIRST MATURATION
During 1984, all males and females belonging to age group 0þ years weremature when they reached 1 year of age at the end of May. All older malesand females (age group �1þ years) were mature in April. During 1985, a delayin the onset of first sexual maturity was observed in females (all of thembelonging to age group 0þ years), which all became mature only in June.The smallest mature individuals for each gender were a 37 mm male anda 36 mm female.
Water depth (cm)
G
0 5 10 15 20 25 30 35 40 45–0·10
–0·06
–0·02
0·02
0·06
0·10
0·14
0·18
FIG. 3. The relationship between instantaneous growth rate (G) and water depth in the study area for
the 1983 ( ) and 1982 ( ) cohorts of river blennies during the warm months (from April
to October). The curves were fitted by y ¼ 0�79x � 0�03 (1983 cohort) and y ¼ 0�92x � 0�05 (1982
cohort).
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1095
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108
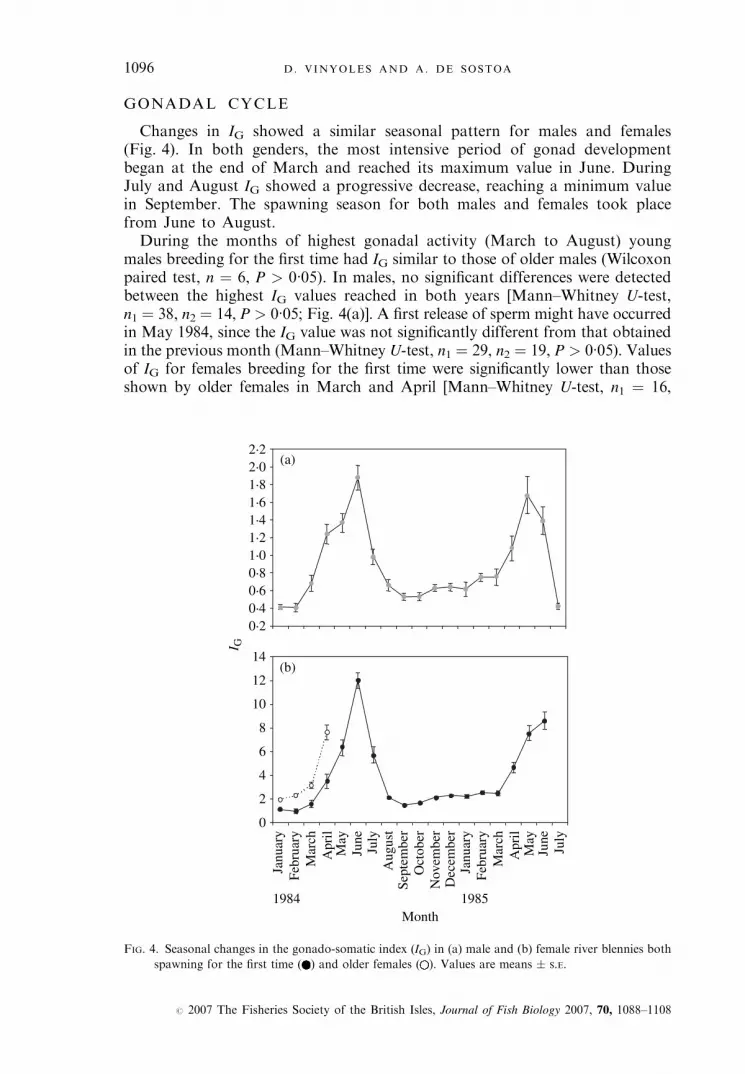
GONADAL CYCLE
Changes in IG showed a similar seasonal pattern for males and females(Fig. 4). In both genders, the most intensive period of gonad developmentbegan at the end of March and reached its maximum value in June. DuringJuly and August IG showed a progressive decrease, reaching a minimum valuein September. The spawning season for both males and females took placefrom June to August.During the months of highest gonadal activity (March to August) young
males breeding for the first time had IG similar to those of older males (Wilcoxonpaired test, n ¼ 6, P > 0�05). In males, no significant differences were detectedbetween the highest IG values reached in both years [Mann–Whitney U-test,n1 ¼ 38, n2 ¼ 14, P > 0�05; Fig. 4(a)]. A first release of sperm might have occurredin May 1984, since the IG value was not significantly different from that obtainedin the previous month (Mann–Whitney U-test, n1 ¼ 29, n2 ¼ 19, P > 0�05). Valuesof IG for females breeding for the first time were significantly lower than thoseshown by older females in March and April [Mann–Whitney U-test, n1 ¼ 16,
(a)
0·20·40·60·81·01·21·41·61·82·02·2
0
2
4
6
8
10
12
14
1984 Month
1985
(b)
I G
Mar
chA
pril
May
June
July
Aug
ust
Sept
embe
rO
ctob
erN
ovem
ber
Dec
embe
r
Janu
ary
Febr
uary
Mar
chA
pril
May
June
July
Janu
ary
Febr
uary
FIG. 4. Seasonal changes in the gonado-somatic index (IG) in (a) male and (b) female river blennies both
spawning for the first time ( ) and older females ( ). Values are means � S.E.
1096 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

n2 ¼ 26, P < 0�001; Fig. 4(b)]. A different pattern may have occurred in older fe-males, since they showed earlier ovarian development than younger females. FromApril 1984, however, very few of these older females were captured at the study site(n ¼ 12). Very few first-reproduction females (reproductive period of 1984 and1985) were captured after spawning (n ¼ 17 after August 1984, n ¼ 2 in July1985). Values of IG for first-reproduction females in June 1984 were significantlyhigher than those detected in June 1985 (Mann–Whitney U-test, n1 ¼ 33, n2 ¼19, P < 0�001). Such differences were not influenced by the size of the females(unpaired t-test, d.f. ¼ 50, P > 0�05).
FEMALE FECUNDITY AND EGG DIAMETER
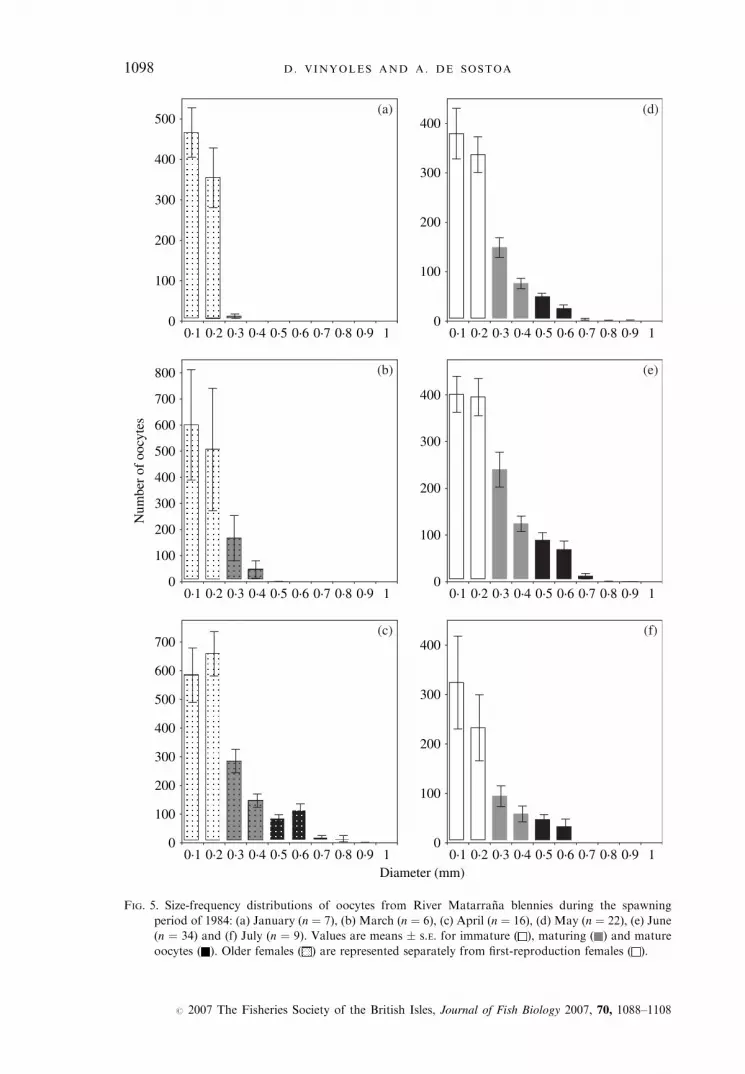
The highest numbers of oocytes were found between March and April inolder females (Fig. 5) and between May and June in females breeding forthe first time (Figs 5 and 6). From July (1984), the total number of oocytesin ovaries decreased until it reached a minimum value in September (mean �S.D. ¼ 80�2 � 49�5). Recruitment oocytes were observed from Septemberonwards. During the quiescent phase (September and October), ovaries onlycontained immature oocytes of translucent appearance (maximum diameter�0�2 mm). Larger opaque oocytes (0�25–0�4 mm diameter) in the process ofmaturation started to appear in November. They were not clearly differentiateduntil March (older females) or April (young females) and remained in theovaries for some time. By the following month (April or May, respectively)a new group of amber-coloured oocytes (0�5–1 mm diameter), or yolky eggs,was found, which remained until August. The fact that yolky eggs occurredin a low proportion throughout the reproductive period shows that riverblenny females are multiple spawners.Estimates of fecundity (i.e. the total number of vitellogenic oocytes � 0�25
mm in diameter prior to spawning) were obtained both for first-reproductionfemales (mature specimens in May and June) and older females (mature speci-mens in April and May). During the first reproductive cycle of the study(1984), the fecundity of older females in April and May was not significantlydifferent from that of first-reproduction females in May and June (ANCOVA,P > 0�05). The regression equations obtained for the two groups of femaleswere: first-reproduction females ln F ¼ �3�02 þ 2�4 ln LT (r2 ¼ 0�33, F1,36,P < 0�001) and older females ln F ¼ �7�42 þ 3�49 ln LT (r2 ¼ 0�75, F1,14,P < 0�001). The mean diameter of ripe oocytes (�0�5 mm) in the ovaries ofolder females in April and May 1984 (mean � S.D. ¼ 0�57 � 0�04 mm, n ¼ 16),however, was significantly larger than that observed later for first-reproductionfemales (0�55 � 0�03 mm, n ¼ 38; unpaired t-test, d.f. ¼ 52, P < 0�01). Duringthe following spawning period (1985) only a small sample of first-reproductionfemales could be analysed (in May and June). The regression equationobtained was: ln F ¼ �10�02 þ 4�12 ln LT (r2 ¼ 0�53, F1,6, P < 0�05).The fecundity estimate for first-reproduction females was significantly higher
in 1984 than in 1985 (ANCOVA, P < 0�01 for intercept values). Females didnot vary in LT between the 2 years (1984: mean � S.D. ¼ 46�2 � 4�8 mm;1985: mean � S.D. ¼ 46�1 � 4�9 mm; unpaired t-test, d.f. ¼ 44, P > 0�05).
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1097
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108
Diameter (mm)
Num
ber
of o
ocyt
es
0
100
200
300
400
500
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
0
100
200
300
400
500
600
700
800
0
100
200
300
400
500
600
700
0
100
200
300
400
0
100
200
300
400
0
100
200
300
400
(a)
(b)
(c)
(d)
(e)
(f)
FIG. 5. Size-frequency distributions of oocytes from River Matarrana blennies during the spawning
period of 1984: (a) January (n ¼ 7), (b) March (n ¼ 6), (c) April (n ¼ 16), (d) May (n ¼ 22), (e) June
(n ¼ 34) and (f) July (n ¼ 9). Values are means � S.E. for immature ( ), maturing ( ) and mature
oocytes ( ). Older females ( ) are represented separately from first-reproduction females ( ).
1098 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Diameter (mm)
Num
ber
of o
ocyt
es
0
50
100
150
200
250
300 (a)
(b)
(c)
(d)
(e)
(f)
0
100
200
300
400
500
600
0
100
200
300
400
0
100
200
300
400
500
0
100
200
300
400
500
600
700
800
0
100
200
300
400
500
600
700
800
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1 0·1 0·2 0·3 0·4 0·5 0·6 0·7 0·8 0·9 1
FIG. 6. Size-frequency distributions of oocytes from River Matarrana blennies during the spawning
period of 1984–1985: (a) November (n ¼ 6), (b) January (n ¼ 9), (c) March (n ¼ 7), (d) April (n ¼ 8),
(e) May (n ¼ 10) and (f) June (n ¼ 7). Values are means � S.E. for immature ( ), maturing ( ) and
mature oocytes ( ). All females are first-reproduction females (1984 cohort).
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1099
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

The mean diameter of ripe oocytes was also not significantly different betweenfirst-reproduction females of 1984 and those of 1985 (mean � S.D. ¼ 552�4 �16�1, n ¼ 8; unpaired t-test, d.f. ¼ 44, P > 0�05).
DISCUSSION
Streams in regions with Mediterranean-type climate are shaped by predict-able seasonal alternations of flooding and drying, but also show strong inter-annual flow variation. Precipitation has been described as the mostimportant of the many abiotic factors that regulate the dynamics of fish pop-ulations in Mediterranean Sea streams of the Iberian Peninsula (Bravo et al., 2001).The longevity of male and female river blennies in the highly variable RiverMatarranya is lower than in other Mediterranean rivers of the Ebro Riverbasin that show a more or less continuous water flow year round (unpubl.data). Severe drought conditions are associated with decreased longevity inother freshwater fish species (Lobon-Cervia et al., 1991; Oliveira et al., 2002).In the present study, both longevity and LT were greater in males than in fe-males. Sexual dimorphism in body size is typical of many species of marine blen-nies, all of them showing exclusive male parental care (Papaconstantinou, 1979).
GROWING IN A VARIABLE ENVIRONMENT
Fish growth is an indeterminate plastic process that can change considerablyin response to environmental factors (Weatherley & Gill, 1987). Intraspecificgrowth differences found in freshwater fish species living in MediterraneanSea rivers are strongly determined by environmental conditions (Lobon-Cerviaet al., 1991; Herrera & Fernandez-Delgado, 1994; Fernandez-Delgado & Herrera,1995; Geraldes & Collares-Pereira, 1995). Summer droughts in particular playa key role in the growth pattern of these fishes. In the River Matarranya, maleand female river blennies exhibited similar growth patterns. Both sexes showedtwo periods of fast growth during warm months (one in the spring, before thesummer drought, and another in autumn). No growth was observed duringsummer drought (July to September), with the exception of young individualsthat had been born a few months earlier. This might explain the appearanceof a false autumn check in the opercular bones of adult river blennies, corre-sponding with the resumption of fish growth. The cessation of growth duringthe dry summer coincides with an abrupt decline in macro-invertebrateabundance in the river, particularly of chironomid larvae and pupae (Vinyoleset al., 1999) which constitute the most common food item for river blennies(Vinyoles, 1993). Moreover, during this period the lower River Matarranyais reduced to shallow, isolated pools in which river blennies can be found atunusually high densities (up to 30 individuals m�2 during August; D. Vinyoles,pers. obs.) living under extreme environmental conditions (high temperatureand low dissolved oxygen concentration). Energy available for growth decreasesunder such conditions (Jobling, 1994; Barton & Taylor, 1996). Thus, the intermit-tent flow of the river results in reduced growth rates and higher mortalityamong large specimens due to physicochemical stressors (Oliveira et al.,2002). Fish populations living in environments shaped by predictable seasonal
1100 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

events, however, might have adapted to such conditions by growing fast duringfavourable conditions. The growth pattern of the river blenny is similar to thatof the gudgeon Gobio gobio (L.), both in this river and in other highly variablestreams (Lobon-Cervia et al., 1991). A phase of rapid growth when favourableconditions are restored after a period of growth depression (‘compensatorygrowth’) has been widely described in fishes (Ali et al., 2003). Periods of growthduring the spring have also been described in other freshwater fishes inhabitingthe Iberian Peninsula (Ribeiro et al., 2000). The River Matarranya is a produc-tive river, which harbours a high density of invertebrates in the spring andearly summer (Malo, 1993). These factors would explain why, in a variablestream such as the River Matarranya, fish growth rate during the warmmonths is positively correlated with water depth. The cumulative temperatureand the high productivity of this stream during the spring and early summerappear to be strong determinants of growth (Lobon-Cervia et al., 1991).River blennies from the River Matarranya born in different years under dif-
ferent environmental conditions all reached a similar size at reproduction,growing fast either before (spring and early summer) or after the dry period(during autumn). Thus, both males and females could attain sexual maturityat the end of their first year of life. Rapid growth during the first year and earlysexual maturity are a life-history tactic adopted by other species in fluctuatingstreams (Herrera & Fernandez-Delgado, 1994; Fernandez-Delgado & Herrera,1995; Aparicio & Sostoa, 1998; Pires et al., 2000; Ribeiro et al., 2000). Thesmallest mature female observed in this study was of a size similar to that re-ported for the marine blenny Aidablennius sphynx (Valenciennes) (Carrasson &Bau, 2003). Salaria economidisi in Lake Trichonis also starts reproducing ata similar small size (Psarras et al., 1997).
GONADAL CYCLE
River blennies do not migrate between spawning and feeding areas (Freemanet al., 1990). The male-biased sex ratios observed in the present study duringthe two reproductive periods can therefore be attributed to a greater mortalityof females during spawning.Reproduction in fishes depends on various environmental factors (De Vlaming,
1972; Joyeux et al., 1992), food availability (Reznick & Yang, 1993) and avail-ability of suitable nest sites (Breitburg, 1987). In the river blenny, gonad devel-opment started at the end of March and peaked in June. The spawningseason for first-reproduction females took place from June to August. Thegonadal cycle of river blennies is similar to that described by Patzner (1983)for the peacock blenny Salaria pavo (Risso) and by Carrasson & Bau (2003)for A. sphynx. In both of these marine species, however, few changes are observedin the ovaries during the winter months, while in river blennies, maturing oocytesstarted appearing in November. Salaria economidisi spawns from March toAugust, with a peak during April and May (Psarras et al., 1997). The differencesobserved between the patterns of gonad development of the two freshwaterblenny species could be related, at least in part, to the presence of larger femalesin the lake species. The present results suggest that in the River Matarranya,older river blenny females (i.e. female spawning for the second and third times)
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1101
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

could have started spawning earlier than the younger ones (during May 1984),which would explain the disappearance of the former from the populationfrom that month onwards after the reproductive effort. This would be consistentwith the assumption that a first period of sperm release occurred in May 1984.Moreover, such an early period of sperm release did not seem to occur inMay 1985, coinciding with a lack of older females in the population. Mostfirst-reproduction females disappeared in July and August, that is, after theiregg-laying period. Wootton (1979) described three patterns of gonad develop-ment for fish species from temperate climates. According to the present results,river blennies may be included in the group of fish species that spawn at theend of spring and during summer. In this group, gonad development is relatedto increases in the daylight period, water temperature and food supply. Gonaddevelopment of the river blenny in the River Matarranya occurs mainly duringthe spring, the period in which the river has plentiful food (Vinyoles, 1993).Somatic energy storage and intense gonad development at the time of the highestenvironmental food supply have been reported for fish species inhabiting a fluc-tuating river (Encina, 1991). The unusual drought after July 1984, however, mayhave caused a decrease in food availability during the second spring of the study.Clear evidence for this is that significantly fewer prey were found in the guts offemales in spring 1985 than in spring 1984 (Vinyoles, 1993). This low food intakecould have had a negative effect on the reproductive investment made by fe-males, whose IG index at the peak of gonad development (June) was significantlylower in the second year of study than in the previous one. Energy demands forgonad development can only be met when there is adequate food supply. Duringdry periods, when somatic energy declines, no additional gonad growth takesplace in adult fish, thus body energy storage can be used as an energy sourcefor other needs (e.g. increase in metabolic rate, decrease in diet content becauseof lower prey availability) (Encina & Granado-Lorencio, 1997).
EGG PRODUCTION PLASTICITY
Several peaks in oocyte size-frequency distributions during the spawningseason indicate that river blennies in the population are multiple spawners.Repeated spawnings by females over long periods have been described in othermembers of this family (Patzner, 1983; Robertson, 1990; Santos, 1995; Carrasson& Bau, 2003). The production of multiple batches of eggs, reaching maturationand being spawned at different times, present certain advantages (Cambray &Bruton, 1984; Burt et al., 1988; Kraak, 1996), especially for fish species livingin highly variable environments (Nikolsky, 1963). It has been suggested thatS. economidisi females are also multiple spawners (Psarras et al., 1997). The ad-vantages of dividing the spawn could moreover compensate, at least in certainfish species, a low fecundity value. Hence, the number of vitellogenic oocytesfound in river blennies in relation to female size (ranging from 416 at 44 mmLT to 1206 at 55 mm LT) are as low as that recorded for A. sphynx (from 432at 45 mm LT to 1682 at 53 mm LT; Carrasson & Bau, 2003). This low fecundity,linked to male parental care, suggests that phylogenetic constraints could shapesome life-history characteristics. In the River Matarranya, river blennieslengthen their reproductive period not only by dividing the spawn, but also
1102 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

through a different pattern of gonad development in females depending ontheir reproductive age. The advantages of this pattern are clear. In Mediterra-nean Sea rivers of the Iberian Peninsula, heavy floods are common, occurringmainly in autumn and occasionally in the spring (Martın, 1985). Hence, eggs laidearly by older females during the spring (April and May) have good chances ofdeveloping each year despite an occasional flood. On the contrary, eggs laid dur-ing the summer (from June onwards) by first-reproduction females have a lowerprobability of survival due to the lack of rainfall and the start of the summerdrought. In addition to affecting the physicochemical conditions of the water(temperature and oxygen content) and causing a decrease in food supply forfishes, summer droughts in the study area result in an important loss of micro-habitat availability for both young and adult river blennies (Freeman et al.,1990). The probability of survival of eggs laid by older females might also behigher because their eggs were larger. Ware (1975) showed that mortality rateof fish eggs is inversely proportional to egg diameter. The eggs of older femalesmay also survive better because the risk of filial cannibalism from male parents islow at the beginning of the reproductive period (Vinyoles et al., 1999). Overall,river blennies show a short life span and older females are relatively scarce, asmight be expected in a fluctuating environment, but a ‘pool’ of older femaleswhose eggs have a high likelihood of survival could ensure the persistence ofthe species in years with exceptionally severe summers. Egg production by youngfemales during the first year of study was larger than during the following cycle.These results are consistent with the differing IG observed and support the ideathat food shortage can be a limiting factor for egg production in some fish spe-cies. On the other hand, limiting egg production in an environment where a goodfood supply cannot be guaranteed could enhance adult survival and prevent in-vesting in offspring subject to a high risk of failure.
LIFE-HISTORY TRAITS AND CONSERVATION
The river blenny population living in the River Matarranya is characterizedby fast growth, early maturity and short life span. These life-history character-istics are typical of species inhabiting unstable environments, where adult mor-tality is high, variable or unpredictable (Stearns, 1976). Such life-history tacticsare frequently found in other freshwater fish species native to Mediterranean Seaenvironments (Fernandez-Delgado & Herrera, 1995; Encina & Granado-Lorencio,1997; Ribeiro et al., 2000), although the environmental thresholds affectinglife histories of freshwater fishes are species-specific (Lobon-Cervia et al.,1991). Early sexual maturity of fishes living in stressful environments is furthersupported by other reproductive life-history tactics. According to Ribeiro et al.(2000), such strategies are based on having a low proportion of individuals sur-viving beyond their second year of life (thus they only spawn once), femalesbeing batch spawners and the size of oocytes increasing with female size (withlarger eggs having a greater chance of survival). These characteristics yield clearadvantages in fluctuating environments (Ribeiro et al., 2000). Offspring productionis not limited to a single, potentially risky reproductive event where a climatecatastrophe (e.g. spring flood or summer drought) could preclude recruitment.Moreover, in a river subject to a rainfall regime such as that of the River
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1103
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Matarranya, both events will rarely happen within a particular year. Hence, anexceptional spring flood followed by a drought-free summer would favour thesurvival of the offspring of young females. Nonetheless, the biologicalcharacteristics of this species could be different in other rivers or in lakes, or evenin Mediterranean-type non-seasonal tributaries (i.e. those with a constant vol-ume of flow) in the same Ebro River basin where this study was carried out.Freshwater fishes are threatened by a wide array of factors, but anthropo-
genic disturbances underlie the rapid decline and extinction of many species(Cowx, 2000). In the Iberian Peninsula, habitat alterations, reduction of flowdue to excessive water extraction and river regulation, pollution and the intro-duction of exotic fish species have contributed to the demise of native speciesacross their natural range (Elvira, 1995b; Ribeiro et al., 2000; Elvira & Almodovar,2001). Demands on water resources are continuously increasing in arid andsemi-arid regions such as the Mediterranean Sea countries (Economidis, 1995;Ribeiro et al., 2000; Collares-Pereira & Cowx, 2004). Thus, natural droughts canbe exacerbated by excessive water extraction in these areas (Moyle, 1995). Unfor-tunately this is also the case with the River Matarranya. Extraction of water forirrigation has been occurring for some years and has already caused quantifiableimpacts on the indigenous fish community, although for the moment the riverblenny population appears to be relatively stable and healthy (unpubl. data).Habitat alterations have contributed to a decline of river blennies in recent years(Aparicio et al., 2000). Some features of the life-history characteristics of river blen-nies found in the RiverMatarranya (such as a short life span and low female fecun-dity that is even lower in unfavourable years) suggest that this species might bevulnerable to severe drought conditions, despite the advantages of other biologicalmechanisms (such as being a multiple spawner and having a long spawning period)or even behavioural mechanisms such as filial cannibalism during food shortageperiods (Vinyoles et al., 1999). The principle of prioritizing conservation areas tocreate nature reserves is well established in terrestrial systems (Cambray, 2002),but few reserves have been specifically designated for freshwater fishes (Crivelli,2002). The preservation of intact flow conditions is one of the key issues for theconservation of fish species inMediterranean Sea rivers (Fischer &Kummer, 2000).
We thank J. D. Rodrıguez-Teijeiro for comments on early drafts of the manuscript,and G. Grossman and M. A. Puig for the facilities provided. Thanks also to F. Casals,V. Fernandez, M. C. Freeman, J. Lobon, C. Lopez and F. de Sostoa for assistance inthe field. We would also like to thank V. C. Almada, I. M. Cote and an anonymousreferee for their comments, which have helped us to improve our work. Field workwas funded by the United States-Spain Joint Committee for Scientific and Technolog-ical Cooperation (CCA841065).
References
Ali, M., Nicieza, A. & Wootton, R. J. (2003). Compensatory growth in fishes: a responseto growth depression. Fish and Fisheries 4, 147–190. doi: 10.1046/j.1467-2979.2003.00120.x
Aparicio, E. & Sostoa, A. de (1998). Reproduction and growth of Barbus haasi in asmall stream in the N.E. of the Iberian peninsula. Archiv fur Hydrobiologie 142,95–110.
1104 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Aparicio, E., Vargas, M. J., Olmo, J. M. & Sostoa, A. de (2000). Decline of native fresh-water fishes in a Mediterranean watershed on the Iberian Peninsula: a quantitativeassessment. Environmental Biology of Fishes 59, 11–19. doi: 10.1023/A:1007618517557
Araujo, R., Bragado, D. & Ramos, M. A. (2001). Identification of the river blenny,Salaria fluviatilis, as a host to the glochidia of Margaritifera auricularia. Journal ofMolluscan Studies 67, 128–129. doi: 10.1093/mollus/67.1.128
Bagenal, T. B. (1978). Aspects of fish fecundity. In Ecology of Freshwater Fish Production(Gerking, S. D., ed.), pp. 75–101. Oxford: Blackwell Scientific Publications.
Bagenal, T. B. & Braum, E. (1978). Eggs and early life history. In Methods of Assessmentof Fish Production in Fresh Waters (Bagenal, T. B., ed.), pp. 165–201. Oxford &Edinburgh: Blackwell Scientific Publications.
Balik, S. (1995). Freshwater fish in Anatolia, Turkey. Biological Conservation 72, 213–223. doi: 10.1016/0006-3207(94)00084-4
Barton, B. A. & Taylor, B. R. (1996). Oxygen requirements of fishes in northern Albertarivers with a general review of the adverse effects of low dissolved oxygen. WaterQuality Research Journal of Canada 31, 361–409.
Bhattacharya, C. G. (1967). A simple method of resolution of a distribution intoGaussian components. Biometrics 23, 115–135.
Bianco, P. G. (1995). Mediterranean endemic fresh-water fishes of Italy. BiologicalConservation 72, 159–170. doi: 10.1016/0006-3207(94)00078-5
Blanco, J. C. & Gonzalez, J. L. (1992). Libro Rojo de los Vertebrados de Espana. Madrid:ICONA.
Bravo, R., Soriguer, M. C., Villar, N. & Hernando, J. A. (2001). The dynamics of fishpopulations in the Palancar stream, a small tributary of the River Guadalquivir,Spain. Acta Oecologica 22, 9–20.
Breitburg, D. (1987). Interspecific competition and the abundance of nest sites: factorsaffecting sexual selection. Ecology 68, 1844–1855.
Bruton, M. N. (1995). Have fishes had their chips? The dilemma of threatened fishes.Environmental Biology of Fishes 43, 1–27.
Burt, A., Kramer, D. L., Nakatsuru, K. & Spry, C. (1988). The tempo of reproduc-tion in Hyphessobrycon pulchripinnis (Characidae) with a discussion on thebiology of ‘‘multiple spawning’’ in fishes. Environmental Biology of Fishes 22,15–27.
Cambray, J. A. (2002). Conservation needs of Sandelia bainsii, an African anabantid. InConservation of Freshwater Fishes: Options for the Future (Collares-Pereira, M. J.,Cowx, I. G. & Coelho, M. M., eds), pp. 90–97. Oxford: Fishing News Books,Blackwell Science.
Cambray, J. A. & Bianco, P. G. (1998). Freshwater fish in crisis, a Blue Planet per-spective. Italian Journal of Zoology 65, 345–356.
Cambray, J. A. & Bruton, M. N. (1984). The reproductive strategy of a barb, Barbusanoplus (Pisces: Cyprinidae), colonizing a man-made lake in South Africa. Journalof Zoology 204, 143–168.
Caro, T. (1998). The significance of behavioral ecology for conservation biology. InBehavioral Ecology and Conservation Biology (Caro, T., ed.), pp. 3–26. New York:Oxford University Press.
Carrasson, M. & Bau, M. (2003). Reproduction and gonad histology of Aidablenniussphynx (Pisces: Blenniidae) of the Catalan Sea (northwestern Mediterranean).Scientia Marina 67, 461–469.
Changeux, T. & Pont, D. (1995). Current status of the riverine fishes of the FrenchMediterranean basin. Biological Conservation 72, 137–158.
Collares-Pereira, M. J., Cowx, I. G., Ribeiro, F., Rodrigues, J. A. & Rogado, L. (2000).Threats imposed by water resource development schemes on the conservation ofendangered fish species in the Guadiana River Basin in Portugal. FisheriesManagement and Ecology 7, 167–178. doi: 10.1046/j.1365-2400.2000.00202.x
Collares-Pereira, M. J. & Cowx, I. G. (2004). The role of catchment scale environmentalmanagement in freshwater fish conservation. Fisheries Management and Ecology11, 303–312. doi: 10.1111/j.1365-2400.2400.00392.x
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1105
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Cote, I. M., Vinyoles, D., Reynolds, J. D., Doadrio, I. & Perdices, A. (1999). Potentialimpacts of gravel extraction on Spanish populations of river blennies Salariafluviatilis (Pisces, Blenniidae). Biological Conservation 87, 359–367. doi: 10.1016/S0006-3207(98)00072-X
Cowx, I. G. (2000). Analysis of threats to freshwater fish conservation: past and presentchallenges. In Conservation of Freshwater Fishes: Options for the Future (Collares-Pereira, M. J., Cowx, I. G. & Coelho, M. M., eds), pp. 201–220. Oxford: FishingNews Books, Blackwell Science.
Craig, J. F. (1974). Population dynamics of perch, Perca fluviatilis L. in Slapton Ley,Devon. II. Age, growth, length-weight relationships and condition. FreshwaterBiology 4, 433–444.
Crivelli, A. J. (2002). The role of protected areas in freshwater fish conservation. InConservation of Freshwater Fishes: Options for the Future (Collares-Pereira, M. J.,Cowx, I. G. & Coelho, M. M., eds), pp. 373–388. Oxford: Fishing News Books,Blackwell Science.
De Vlaming, V. L. (1972). The effects of diurnal thermoperiod treatments on reproduc-tive function in the estuarine gobiid fish, Gillichthys mirabilis Cooper. Journal ofthe Experimental Marine Biology and Ecology 9, 155–163.
Doadrio, I. (2001). Atlas y Libro Rojo de los Peces Continentales de Espana. Madrid: CSIC-Ministerio de Medio Ambiente.
Economidis, P. S. (1995). Endangered freshwater fishes of Greece. Biological Conserva-tion 72, 201–211. doi: 10.1016/0006-3207(94)00083-3
Elvira, B. (1995a). Conservation status of endemic freshwater fish in Spain. BiologicalConservation 72, 129–136. doi: 10.1016/0006-3207(94)00076-3
Elvira, B. (1995b). Native and exotic freshwater fishes in Spanish river basins. FreshwaterBiology 33, 103–108.
Elvira, B. & Almodovar, A. (2001). Freshwater fish introductions in Spain: facts andfigures at the beginning of the 21st century. Journal of Fish Biology 59, 323–331.doi: 10.1006/jfbi.2001.1753
Encina, L. (1991). Ecologıa trofica y dinamica energetica de la comunidad ıctica del rıoGuadalete. PhD Dissertation, University of Sevilla, Spain.
Encina, L. & Granado-Lorencio, C. (1997). Seasonal variations in the physiologicalstatus and energy content of somatic and reproductive tissues of chub. Journal ofFish Biology 50, 511–522. doi: 10.1111/j.1095-8649.1997.tb01946.x
Fernandez-Delgado, C. & Herrera, M. (1995). Age structure, growth and reproductionof Rutilus lemmingii in an intermittent stream of the Guadalquivir River basin,southern Spain. Hydrobiologia 299, 207–213.
Ferrito, V. & Tigano, C. (1996). Decline of Aphanius fasciatus (Cyprinodontidae) andSalaria fluviatilis (Blenniidae) populations in freshwater of eastern Sicily. Ichthy-ological Exploration of Freshwaters 7, 181–184.
Fischer, S. & Kummer, H. (2000). Effects of residual flow and habitat fragmentation ondistribution and movement of bullhead (Cottus gobio L.) in an alpine stream.Hydrobiologia 422, 305–317. doi: 10.1023/A:1017083714513
Freeman, M. C., Vinyoles, D., Grossman, G. D. & Sostoa, A. de (1990). Microhabitatuse by Blennius fluviatilis in the Rıo Matarrana, Spain. Freshwater Biology 24,335–345.
Gayanillo, F. C., Soriano, M. & Pauly, D. (1988). A Draft Guide to the Complete ELEFAN.Manila: International Centre for Living Aquatic Resources Management.
Geraldes, A. M. & Collares-Pereira, M. J. (1995). Preliminary insight into age and growthof the Iberian chub (Leuciscus pyrenaicus) in the Sorraia system. Folia Zoologica44, 159–165.
Goss-Custard, J. D. & Sutherland, W. J. (1997). Individual behaviour, populations andconservation. In Behavioural Ecology: an Evolutionary Approach (Krebs, J. R. &Davies, N. B., eds), pp. 373–395. Oxford: Blackwell Science.
Hernandez, R., Lacomba, R. T., Uvinas, Y. N. & Oltra, R. (2000). Distribution patternof river blennies in the Jucar River basin (eastern Spain). Journal of Fish Biology57, 250–254. doi: 10.1111/j.1095-8649.2000.tb00790.x
1106 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Herrera, M. & Fernandez-Delgado, C. (1994). The age, growth and reproduction ofChondrostoma polylepis willkommi in a seasonal stream in the Guadalquivir Riverbasin (southern Spain). Journal of Fish Biology 44, 11–22. doi: 10.1111/j.1095-8649.1994.tb01581.x
Jobling, M. (1994). Fish Bioenergetics. London: Chapman & Hall.Joyeux, J. C., Tomasini, J. A. & Bouchereau, J. L. (1992). Modalites de la reproduction
de Gobius niger (Teleostei, Gobiidae) dans une lagune Mediterraneenne. Vie etMilieu 42, 1–13.
Kraak, S. B. M. (1996). A quantitative description of the reproductive biology of theMediterranean blenny Aidablennius sphynx (Teleostei, Blenniidae) in its naturalhabitat. Environmental Biology of Fishes 46, 329–342.
Kottelat, M. (2004). Salaria economidisi, a new species of freshwater fish from LakeTrichonis, Greece, with comments on variation in S. fluviatilis (Teleostei: Blennii-dae). Revue Suisse de Zoologie 111, 121–137.
Lobon-Cervia, J., Montanes, C. & Sostoa, A. de (1991). Influence of environment uponthe life history of gudgeon, Gobio gobio (L.): a recent and successful colonizer ofthe Iberian Peninsula. Journal of Fish Biology 39, 285–300.
Malo, J. (1993). Comunidades bentonicas de rıos mediterraneos. PhD Dissertation,University of Murcia, Spain.
Martın, X. (1985). Pluges i inundacions a la Mediterrania. Barcelona: Ketres Editora.Moyle, P. B. (1995). Conservation of native freshwater fishes in the Mediterranean-type
climate of California, USA: a review. Biological Conservation 72, 271–279. doi:10.1016/0006-3207(94)00083-3
Mrakovcic, M., Misetic, S. & Povz, M. (1995). Status of freshwater fish in CroatianAdriatic River systems. Biological Conservation 72, 179–185. doi: 10.1016/0006-3207(94)00080-A
Nikolsky, G. V. (1963). The Ecology of Fishes. London: Academic Press.Oliva-Paterna, F. J., Minano, P. A. & Torralva, M. (2003). Habitat quality affects the
condition of Barbus sclateri in Mediterranean semi-arid streams. EnvironmentalBiology of Fishes 67, 13–22. doi: 10.1023/A:1024481928161
Oliveira, J. M., Ferreira, A. P. & Ferreira, M. T. (2002). Intrabasin variations in age andgrowth of Barbus bocagei populations. Journal of Applied Ichthyology 18, 134–139.doi: 10.1046/j.1439-0426.2002.00333.x
Papaconstantinou, C. A. (1979). The secondary sex characteristics of the genus Blenniusof the Mediterranean Sea (Pisces: Blenniidae). Rapport Commitee Internationale dela Mer Mediterranee 25/26, 10.
Patzner, R. A. (1983). The reproduction of Blennius pavo (Teleostei, Blenniidae). I. Ovarialcycle, environmental factors and feeding. Helgolander Meeresunters 36, 105–114.
Pires, A. M., Cowx, I. G. & Coelho, M. M. (2000). Life history strategy of Leuciscuspyrenaicus (Cyprinidae) in intermittent streams of the Guadiana basin (Portugal).Cybium 24, 287–297.
Psarras, Th., Barbieri-Tseliki, R. & Economou, A. (1997). First data on the feeding andbiology of reproduction of Salaria fluviatilis. Proceedings 5th Hellenic Symposium onOceanography & Fisheries 2, 261–264.
Reznick, D. & Yang, A. P. (1993). The influence of fluctuating resources on lifehistory-patterns of allocation and plasticity in female guppies. Ecology 74, 2011–2019.
Ribeiro, F., Cowx, I. G. & Collares-Pereira, M. J. (2000). Life history traits of theendangered Iberian cyprinid Anaecypris hispanica and their implications forconservation. Archiv fur Hydrobiologie 149, 569–586.
Ricciardi, A. & Rasmussen, J. B. (1999). Extinction rates of North American fresh-water fauna. Conservation Biology 13, 1220–1222. doi: 10.1046/j.1523-1739.1999.98380.x
Rice, W. R. (1989). Analyzing tables of statistical tests. Evolution 43, 223–225.Richter, B. D., Braun, D. P., Mendelson, M. A. & Master, L. L. (1997). Threats to
imperiled freshwater fauna. Conservation Biology 11, 1081–1093. doi: 10.1046/j.1523-1739.1997.96236.x
LIFE-HISTORY TRAITS OF THE RIVER BLENNY 1107
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108

Robertson, D. R. (1990). Differences in the seasonalities of spawning and recruitment ofsome small neotropical reef fishes. Journal of Experimental Marine Biology andEcology 144, 49–62.
Santos, R. S. (1995). Anatomy and histology of secondary sexual characters, gonad andliver of the rock-pool blenny, Parablennius sanguinolentus parvicornis (Pisces:Blenniidae) of the Azores. Arquipe lago 13, 21–38.
Sostoa, A. de, Casals, F., Fernandez, J. V., Lobon-Cervia, J., Sostoa, F. J. & Vinyoles, D.(1990). Les comunitats de peixos continentals. In Historia Natural dels Paı sosCatalans, Vol. 11 (Sostoa, A. de, ed.), pp. 390–392. Barcelona: FundacioEnciclopedia Catalana.
Stearns, S. C. (1976). Life-history tactics: a review of the ideas. The Quarterly Review ofBiology 51, 3–47.
Sutherland, W. J. (1995). Introduction and principles of ecological management. InManaging Habitats for Conservation (Sutherland, W. J. & Hill, D. A., eds), pp. 1–21.Cambridge: Cambridge University Press.
Vinyoles, D. (1993). Biologia i ecologia de Blennius fluviatilis (Pisces: Blenniidae) al RiuMatarranya. PhD Dissertation, University of Barcelona, Spain.
Vinyoles, D., Cote, I. M. & Sostoa, A. de (1999). Egg cannibalism in river blennies: therole of natural prey availability. Journal of Fish Biology 55, 1223–1232. doi:10.1111/j.1095-8649.1999.tb02072.x
Vinyoles, D., Cote, I. M. & Sostoa, A. de (2002). Nest orientation patterns in Salariafluviatilis. Journal of Fish Biology 61, 405–416. doi: 10.1111/j.1095-8649.2002.tb01573.x
Ware, D. M. (1975). Relation between egg size, growth and natural mortality of larvalfish. Journal of the Fisheries Research Board of Canada 32, 2503–2512.
Weatherley, A. H. & Gill, H. S. (1987). The Biology of Fish Growth. London: AcademicPress Inc.
Wickler, W. (1957). Vergleichende Verhaltensstudien an Grundfischen. I. Beitrage zurBiologie, besonders zur Ethologie von Blennius fluviatilis Asso im Vergleich zueinigen anderen Bodenfischen. Zeitschrift fur Tierpsychologie 14, 393–428.
Williams, J. E. & Miller, R. R. (1990). Conservation status of the North American fishfauna in fresh water. Journal of Fish Biology 37, 79–85.
Wootton, R. J. (1979). Energy cost of egg production and environmental determinantsof fecundity in teleost fishes. Symposia of the Zoological Society of London 44,133–159.
1108 D. VINYOLES AND A. DE SOSTOA
# 2007 The Fisheries Society of the British Isles, Journal of Fish Biology 2007, 70, 1088–1108




















