
Distribution of egg strands of perch (Perca fluviatilis L.) with respect to depth and spawning...
10
PRIMARY RESEARCH PAPER Distribution of egg strands of perch (Perca fluviatilis L.) with respect to depth and spawning substrate Martin C ˇ ech Æ Jir ˇı ´ Peterka Æ Milan R ˇ ı ´ha Æ Toma ´s ˇ Ju ˚za Æ Jan Kubec ˇka Received: 8 December 2008 / Revised: 5 April 2009 / Accepted: 6 April 2009 / Published online: 3 May 2009 Ó Springer Science+Business Media B.V. 2009 Abstract The distribution of egg strands of perch Perca fluviatilis was studied in relation to water depth and spawning substrate during late April and early May 2007 in Chabarˇovice Lake, Czech Repub- lic, using SCUBA divers and parallel transects. The depth distribution of egg strands differed significantly between the two sampling dates, being much deeper in early May compared to late April. Perch used at least seven different spawning substrates of which the most important were curly pondweed Potamog- eton crispus, worm weed Artemisia sp. and common reed Phragmites communis. However, while living submerged vegetation, although more abundant, was generally avoided, dead submerged vegetation was strongly preferred. It appears that dead vegetation is an ideal spawning substrate for perch since placement of the egg strands over those hard, complex three- dimensional structures ensures that the eggs remain well oxygenated 24 hours a day. Factors with significant influence on the distribution of egg strands were as follows (in order of decreasing influence): depth of deposition, type of spawning substrate, date of spawning and temperature at the depth of deposition. Keywords ChabarˇoviceLake SCUBA diving Aquatic vegetation Curly pondweed Potamogeton crispus Common reed Phragmites communis Eurasian water milfoil Myriophyllum spicatum Introduction European perch Perca fluviatilis L. is often assumed to spawn in the shallow water of rivers, lakes and reservoirs, where the majority of gelatinous egg strands are deposited to a maximum depth of 2 m (Holc ˇı ´k, 1965; Frank, 1967; Stehlı ´k, 1969; Viljanen & Holopainen, 1982; Treasurer, 1983; Urho, 1996; Smith et al., 2001). Lang (1987), however, reported perch spawning at 3–5 m and Craig (1987) from 0.5 up to a depth of 8 m. The results of Kubec ˇka (1992) have suggested that during the spawning period, warming of surface water could shift the depth of peak spawning activity from shallower towards the deeper layers, so that at the end of a prolonged spawning period, the peak activity could be found in a depth of 6–9 m. Similar results were found by Gillet & Dubois (1995, 2007). Perch spawn over a wide range of substrates including sand, boulders and gravels, submerged or emergent aquatic vegetation, roots and stumps of Handling editor: J. A. Cambray M. C ˇ ech (&) J. Peterka M. R ˇ ı ´ha T. Ju ˚za J. Kubec ˇka Biology Centre, Academy of Sciences of the Czech Republic, Institute of Hydrobiology, Na Sa ´dka ´ch 7, 370 05 C ˇ eske ´ Bude ˇjovice, Czech Republic e-mail: [email protected] 123 Hydrobiologia (2009) 630:105–114 DOI 10.1007/s10750-009-9783-z
Transcript of Distribution of egg strands of perch (Perca fluviatilis L.) with respect to depth and spawning...

PRIMARY RESEARCH PAPER
Distribution of egg strands of perch (Perca fluviatilis L.)with respect to depth and spawning substrate
Martin Cech Æ Jirı Peterka Æ Milan Rıha ÆTomas Juza Æ Jan Kubecka
Received: 8 December 2008 / Revised: 5 April 2009 / Accepted: 6 April 2009 / Published online: 3 May 2009
� Springer Science+Business Media B.V. 2009
Abstract The distribution of egg strands of perch
Perca fluviatilis was studied in relation to water
depth and spawning substrate during late April and
early May 2007 in Chabarovice Lake, Czech Repub-
lic, using SCUBA divers and parallel transects. The
depth distribution of egg strands differed significantly
between the two sampling dates, being much deeper
in early May compared to late April. Perch used at
least seven different spawning substrates of which the
most important were curly pondweed Potamog-
eton crispus, worm weed Artemisia sp. and common
reed Phragmites communis. However, while living
submerged vegetation, although more abundant, was
generally avoided, dead submerged vegetation was
strongly preferred. It appears that dead vegetation is
an ideal spawning substrate for perch since placement
of the egg strands over those hard, complex three-
dimensional structures ensures that the eggs remain
well oxygenated 24 hours a day. Factors with
significant influence on the distribution of egg strands
were as follows (in order of decreasing influence):
depth of deposition, type of spawning substrate, date
of spawning and temperature at the depth of
deposition.
Keywords Chabarovice Lake � SCUBA diving �Aquatic vegetation � Curly pondweed
Potamogeton crispus � Common reed Phragmites
communis � Eurasian water milfoil Myriophyllum
spicatum
Introduction
European perch Perca fluviatilis L. is often assumed
to spawn in the shallow water of rivers, lakes and
reservoirs, where the majority of gelatinous egg
strands are deposited to a maximum depth of 2 m
(Holcık, 1965; Frank, 1967; Stehlık, 1969; Viljanen
& Holopainen, 1982; Treasurer, 1983; Urho, 1996;
Smith et al., 2001). Lang (1987), however, reported
perch spawning at 3–5 m and Craig (1987) from 0.5
up to a depth of 8 m. The results of Kubecka (1992)
have suggested that during the spawning period,
warming of surface water could shift the depth of
peak spawning activity from shallower towards the
deeper layers, so that at the end of a prolonged
spawning period, the peak activity could be found in
a depth of 6–9 m. Similar results were found by
Gillet & Dubois (1995, 2007).
Perch spawn over a wide range of substrates
including sand, boulders and gravels, submerged or
emergent aquatic vegetation, roots and stumps of
Handling editor: J. A. Cambray
M. Cech (&) � J. Peterka � M. Rıha � T. Juza �J. Kubecka
Biology Centre, Academy of Sciences of the Czech
Republic, Institute of Hydrobiology, Na Sadkach 7, 370
05 Ceske Budejovice, Czech Republic
e-mail: [email protected]
123
Hydrobiologia (2009) 630:105–114
DOI 10.1007/s10750-009-9783-z

trees, dead branches dipping into the water, dislodged
trees lying on the bottom and other materials (e.g.
Zhakov, 1964; Frank, 1967; Pivnicka, 1979; Viljanen
& Holopainen, 1982; Treasurer, 1983; Craig, 1987;
Lang, 1987; Urho, 1996; Pedicillo et al., 2008).
Although the preference for submerged vegetation
sensus lato as a spawning substrate seems to be obvious
in perch, most studies do not provide any evaluation of
this preference (Frank, 1967; Holcık, 1969; Craig,
1987; Gillet & Dubois, 1995; Urho, 1996).
The present study focussed on the distribution of
egg strands of perch with respect to depth and
spawning substrate in a relatively deep lake, charac-
terized by large stands of submerged vegetation (both
dead and alive), i.e. the study area represents a water
body with no limitations to perch spawning activity
concerning depth and spawning substrate.
The main questions were (1) whether perch really
prefer spawning in very shallow areas up to a depth
of 2 m or whether the depth of spawning is more
likely controlled by the presence of appropriate
spawning substrate, or by the optimal temperature
for egg development and (2) whether perch use all
available spawning substrates (all types of submerged
vegetation in particular), in proportion to those
available in the lake, for deposition of their egg
strands. Thus, the main aim of this study was to find
and evaluate factors influencing the deposition of egg
strands in the system without any obvious limitation
to perch spawning and to provide recommendations
for management.
The study is part of a long-term project focussing
on succession processes in a restored opencast mine
lake where aquatic restoration has been applied.
Materials and methods
Study area
The study was carried out in the oligo- to mesotro-
phic Chabarovice Lake, Czech Republic (80 km
north-west of Prague), which has an area of ca.
200 ha, volume of 18 9 106 m3 and maximum depth
of 20 m (Fig. 1a). Secchi depth exceeded 7 m for
most of the year.
Chabarovice Lake is a newly restored, opencast
mine lake where aquatic restoration started in 2001.
The lake contains large stands of submerged
vegetation composed of curly pondweed Potamog-
eton crispus, Eurasian water milfoil Myriophyl-
lum spicatum, common stonewort Chara vulgaris
and Canadian waterweed Elodea canadensis. Due to
the seasonal succession of vegetation, the last two
species are much more abundant mainly later in the
year (July–October; J. Peterka, M. Rıha, M. Cech,
unpublished data).
During the filling of the lake, young trees and their
dislodged branches, bushes (especially black elder
Sambucus nigra), beds of common reed Phrag-
mites communis, worm weed Artemisia sp. and
common rush Juncus effusus became submerged in
large numbers (J. Peterka, M. Rıha, M. Cech, unpubl.
data). Today they have created a unique habitat of
dead vegetation with complex three-dimensional
structures (Fig. 1b).
The ichthyofauna of the lake consists of 12 fish
species, but only rudd Scardinius erythrophthalmus
(L.) (58% in abundance of fish [ 0?), perch
Perca fluviatilis L. (25%) and roach Rutilus rutilus
(L.) (14%) reach ecologically significant levels
(Kubecka et al., 2007). Despite the extensive stocking
programme focussing on larger individuals of pike
Esox lucius L., zander Sander lucioperca (L.) and
wels catfish Silurus glanis L. (since 2005; for
biomanipulation purposes), perch is still the most
abundant predatory fish in the lake, with a unimodal
cohort of older fish of standard lengths of 240–
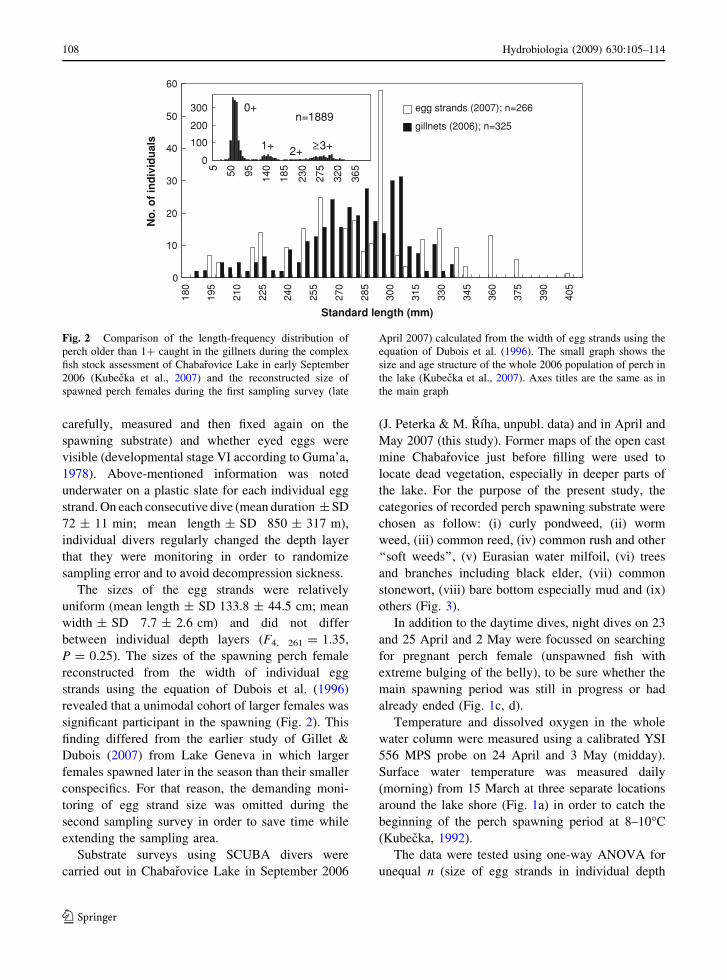
340 mm (Kubecka et al., 2007; Fig. 2).
Sampling
Three SCUBA divers monitored the depth distribution
of egg strands and spawning substrates of perch in
depth layers of 2–4, 4–6 and 6–8 m during 24–26 April
and 2–3 May 2007. On the same dates, additional dives
were made to depths [8 m (Fig. 1c, d). The divers
swam parallel to each other at minimum horizontal
distances of 5 m apart, each sampling the space of 3 m
to each side of him. Simultaneously to this SCUBA
research, the whole littoral zone of the lake (depth layer
0–2 m) was sampled visually from the boat using
polarized glasses.
During the first sampling survey, the divers
recorded the type of substrate on which individual
egg strands were deposited, the depth of their
deposition, the sizes of the egg strands (length, width;
using ruler, for which the egg strands were unravelled
106 Hydrobiologia (2009) 630:105–114
123
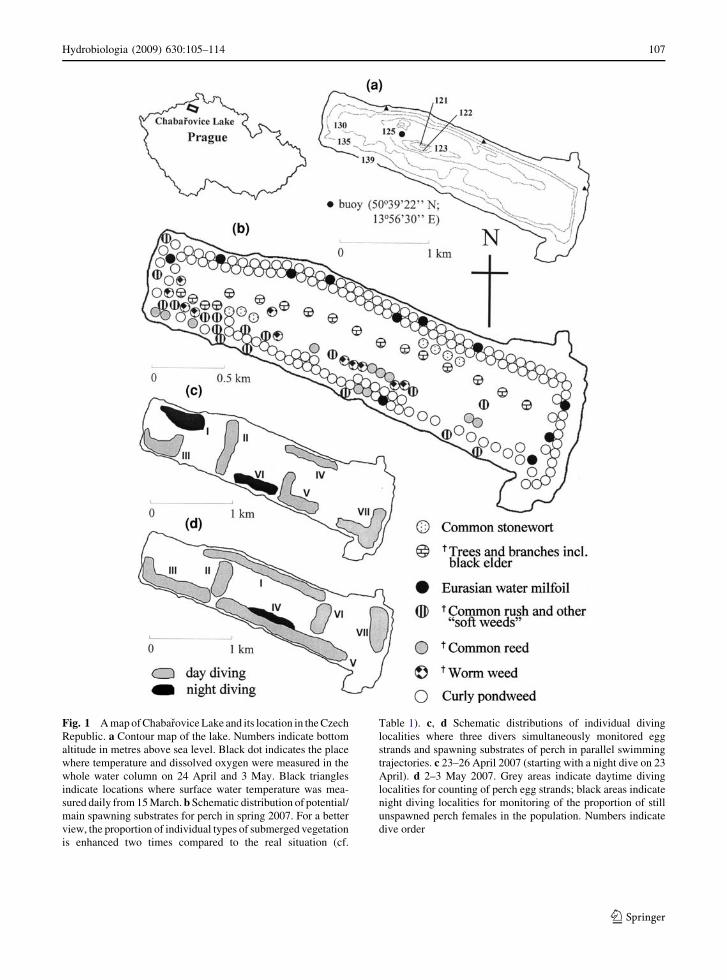
Fig. 1 A map of Chabarovice Lake and its location in the Czech
Republic. a Contour map of the lake. Numbers indicate bottom
altitude in metres above sea level. Black dot indicates the place
where temperature and dissolved oxygen were measured in the
whole water column on 24 April and 3 May. Black triangles
indicate locations where surface water temperature was mea-
sured daily from 15 March. b Schematic distribution of potential/
main spawning substrates for perch in spring 2007. For a better
view, the proportion of individual types of submerged vegetation
is enhanced two times compared to the real situation (cf.
Table 1). c, d Schematic distributions of individual diving
localities where three divers simultaneously monitored egg
strands and spawning substrates of perch in parallel swimming
trajectories. c 23–26 April 2007 (starting with a night dive on 23
April). d 2–3 May 2007. Grey areas indicate daytime diving
localities for counting of perch egg strands; black areas indicate
night diving localities for monitoring of the proportion of still
unspawned perch females in the population. Numbers indicate
dive order
Hydrobiologia (2009) 630:105–114 107
123
carefully, measured and then fixed again on the
spawning substrate) and whether eyed eggs were
visible (developmental stage VI according to Guma’a,
1978). Above-mentioned information was noted
underwater on a plastic slate for each individual egg
strand. On each consecutive dive (mean duration ± SD
72 ± 11 min; mean length ± SD 850 ± 317 m),
individual divers regularly changed the depth layer
that they were monitoring in order to randomize
sampling error and to avoid decompression sickness.
The sizes of the egg strands were relatively
uniform (mean length ± SD 133.8 ± 44.5 cm; mean
width ± SD 7.7 ± 2.6 cm) and did not differ
between individual depth layers (F4, 261 = 1.35,
P = 0.25). The sizes of the spawning perch female
reconstructed from the width of individual egg
strands using the equation of Dubois et al. (1996)
revealed that a unimodal cohort of larger females was
significant participant in the spawning (Fig. 2). This
finding differed from the earlier study of Gillet &
Dubois (2007) from Lake Geneva in which larger
females spawned later in the season than their smaller
conspecifics. For that reason, the demanding moni-
toring of egg strand size was omitted during the
second sampling survey in order to save time while
extending the sampling area.
Substrate surveys using SCUBA divers were
carried out in Chabarovice Lake in September 2006
(J. Peterka & M. Rıha, unpubl. data) and in April and
May 2007 (this study). Former maps of the open cast
mine Chabarovice just before filling were used to
locate dead vegetation, especially in deeper parts of
the lake. For the purpose of the present study, the
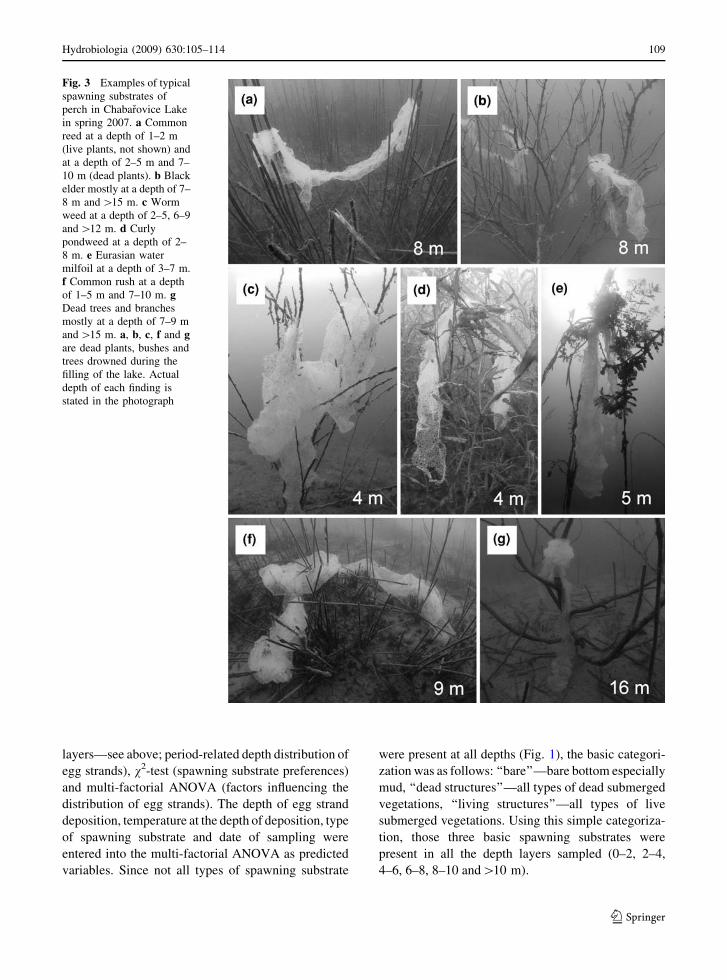
categories of recorded perch spawning substrate were
chosen as follow: (i) curly pondweed, (ii) worm
weed, (iii) common reed, (iv) common rush and other
‘‘soft weeds’’, (v) Eurasian water milfoil, (vi) trees
and branches including black elder, (vii) common
stonewort, (viii) bare bottom especially mud and (ix)
others (Fig. 3).
In addition to the daytime dives, night dives on 23
and 25 April and 2 May were focussed on searching
for pregnant perch female (unspawned fish with
extreme bulging of the belly), to be sure whether the
main spawning period was still in progress or had
already ended (Fig. 1c, d).
Temperature and dissolved oxygen in the whole
water column were measured using a calibrated YSI
556 MPS probe on 24 April and 3 May (midday).
Surface water temperature was measured daily
(morning) from 15 March at three separate locations
around the lake shore (Fig. 1a) in order to catch the
beginning of the perch spawning period at 8–10�C
(Kubecka, 1992).
The data were tested using one-way ANOVA for
unequal n (size of egg strands in individual depth
0
10
20
30
40
50
60
180
195
210
225
240
255
270
285
300
315
330
345
360
375
390
405
Standard length (mm)
No
. of
ind
ivid
ual
s
egg strands (2007); n=266
gillnets (2006); n=325
0
100
200
300
5 50 95 140
185
230
275
320
365
0+
1+ 2+ ≥3+
n=1889
Fig. 2 Comparison of the length-frequency distribution of
perch older than 1? caught in the gillnets during the complex
fish stock assessment of Chabarovice Lake in early September
2006 (Kubecka et al., 2007) and the reconstructed size of
spawned perch females during the first sampling survey (late
April 2007) calculated from the width of egg strands using the
equation of Dubois et al. (1996). The small graph shows the
size and age structure of the whole 2006 population of perch in
the lake (Kubecka et al., 2007). Axes titles are the same as in
the main graph
108 Hydrobiologia (2009) 630:105–114
123
layers—see above; period-related depth distribution of
egg strands), v2-test (spawning substrate preferences)
and multi-factorial ANOVA (factors influencing the
distribution of egg strands). The depth of egg strand
deposition, temperature at the depth of deposition, type
of spawning substrate and date of sampling were
entered into the multi-factorial ANOVA as predicted
variables. Since not all types of spawning substrate
were present at all depths (Fig. 1), the basic categori-
zation was as follows: ‘‘bare’’—bare bottom especially
mud, ‘‘dead structures’’—all types of dead submerged
vegetations, ‘‘living structures’’—all types of live
submerged vegetations. Using this simple categoriza-
tion, those three basic spawning substrates were
present in all the depth layers sampled (0–2, 2–4,
4–6, 6–8, 8–10 and[10 m).
Fig. 3 Examples of typical
spawning substrates of
perch in Chabarovice Lake
in spring 2007. a Common
reed at a depth of 1–2 m
(live plants, not shown) and
at a depth of 2–5 m and 7–
10 m (dead plants). b Black
elder mostly at a depth of 7–
8 m and [15 m. c Worm
weed at a depth of 2–5, 6–9
and [12 m. d Curly
pondweed at a depth of 2–
8 m. e Eurasian water
milfoil at a depth of 3–7 m.
f Common rush at a depth
of 1–5 m and 7–10 m. gDead trees and branches
mostly at a depth of 7–9 m
and [15 m. a, b, c, f and gare dead plants, bushes and
trees drowned during the
filling of the lake. Actual
depth of each finding is
stated in the photograph
Hydrobiologia (2009) 630:105–114 109
123
Results
During two sampling surveys of the lake, 14 dives
were carried out (3 night dives, 11 day dives,
comprising 50 h underwater) during which 896 indi-
vidual egg strands of perch were found (n24–26 April =
266; n2–3 May = 630). Night dives revealed that
during the first survey a significant proportion of the
females had not spawned (nobserved adult perch = 153;
nobserved pregnant female = 27). No egg strands were
found to be close to hatching, i.e. in an eyed eggs
stage. Throughout the daylight hours, males were
frequently ‘guarding’ the spawning substrate. They
were waiting for females in between stalks and
branches or they were slowly cruising in close
proximity to those structures. In contrast to females
occurring in shoals in deeper open water, males
showed very poor avoidance reaction and occasion-
ally attempted to approach the SCUBA diver.
During the second survey, night observations of resting
fish revealed that all females had spawned their eggs
(nobserved adult perch = 88; nobserved pregnant female = 0).
No males were observed ‘guarding’ the spawning
substrate at any depth. At that time, many egg strands
were close to hatching or in the process of hatching
(embryos leaving the egg envelope). In the depth
layers of 2–4 and 4–6 m, egg strands of 76.5% and
82.3%, respectively, were in the eyed eggs stage (at
least); however, in the 6–8 m depth layer it was only
3.9%, whereas in the depth [8 m, it was 0%.
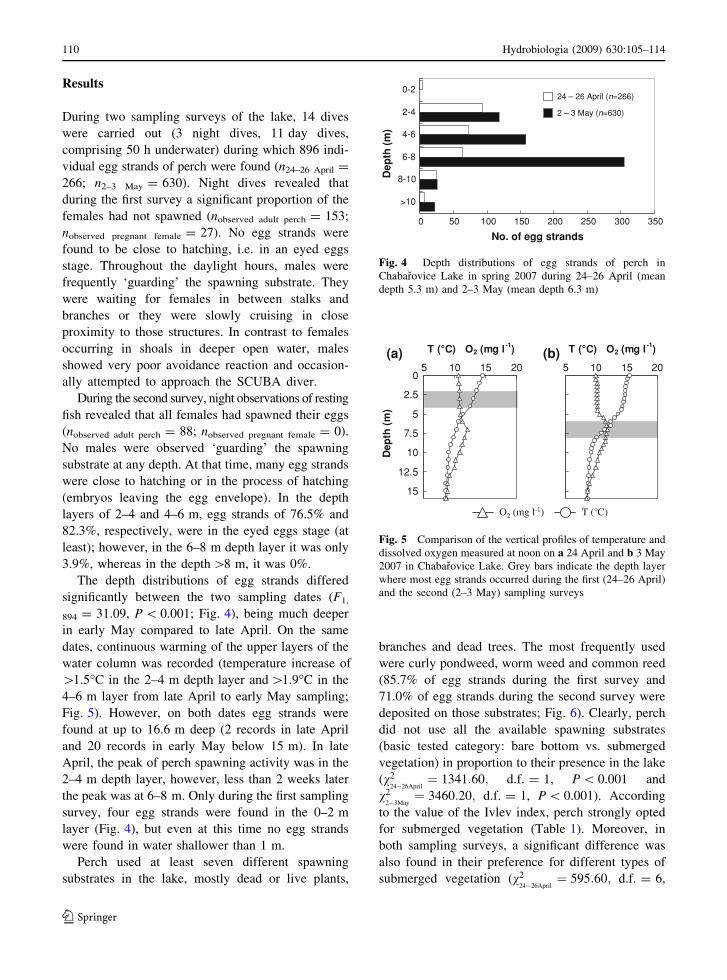
The depth distributions of egg strands differed
significantly between the two sampling dates (F1,
894 = 31.09, P \ 0.001; Fig. 4), being much deeper
in early May compared to late April. On the same
dates, continuous warming of the upper layers of the
water column was recorded (temperature increase of
[1.5�C in the 2–4 m depth layer and [1.9�C in the
4–6 m layer from late April to early May sampling;
Fig. 5). However, on both dates egg strands were
found at up to 16.6 m deep (2 records in late April
and 20 records in early May below 15 m). In late
April, the peak of perch spawning activity was in the
2–4 m depth layer, however, less than 2 weeks later
the peak was at 6–8 m. Only during the first sampling
survey, four egg strands were found in the 0–2 m
layer (Fig. 4), but even at this time no egg strands
were found in water shallower than 1 m.
Perch used at least seven different spawning
substrates in the lake, mostly dead or live plants,
branches and dead trees. The most frequently used
were curly pondweed, worm weed and common reed
(85.7% of egg strands during the first survey and
71.0% of egg strands during the second survey were
deposited on those substrates; Fig. 6). Clearly, perch
did not use all the available spawning substrates
(basic tested category: bare bottom vs. submerged
vegetation) in proportion to their presence in the lake
(v224�26April
¼ 1341:60; d.f. = 1, P \ 0.001 and
v22�3May
¼ 3460:20; d.f. = 1, P \ 0.001). According
to the value of the Ivlev index, perch strongly opted
for submerged vegetation (Table 1). Moreover, in
both sampling surveys, a significant difference was
also found in their preference for different types of
submerged vegetation (v224�26April
¼ 595:60; d.f. = 6,
0 50 100 150 200 250 300 350
0-2
2-4
4-6
6-8
8-10
>10
Dep
th (
m)
No. of egg strands
24 – 26 April (n=266)
2 – 3 May (n=630)
Fig. 4 Depth distributions of egg strands of perch in
Chabarovice Lake in spring 2007 during 24–26 April (mean
depth 5.3 m) and 2–3 May (mean depth 6.3 m)
5 10 15 20
T (°C) O2 (mg l-1)
0
2.5
5
7.5
10
12.5
15
5 10 15 20
T (°C) O2 (mg l-1)
Dep
th (
m)
T (°C)O2 (mg l-1)
(a) (b)
Fig. 5 Comparison of the vertical profiles of temperature and
dissolved oxygen measured at noon on a 24 April and b 3 May
2007 in Chabarovice Lake. Grey bars indicate the depth layer
where most egg strands occurred during the first (24–26 April)
and the second (2–3 May) sampling surveys
110 Hydrobiologia (2009) 630:105–114
123
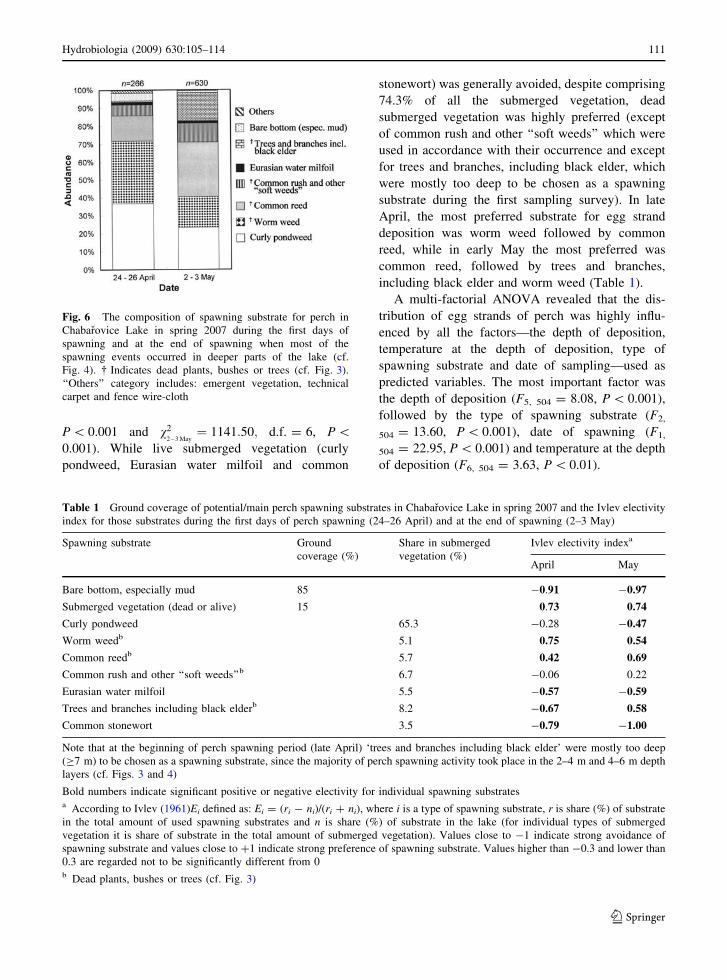
P \ 0.001 and v22�3 May
¼ 1141:50; d.f. = 6, P \0.001). While live submerged vegetation (curly
pondweed, Eurasian water milfoil and common
stonewort) was generally avoided, despite comprising
74.3% of all the submerged vegetation, dead
submerged vegetation was highly preferred (except
of common rush and other ‘‘soft weeds’’ which were
used in accordance with their occurrence and except
for trees and branches, including black elder, which
were mostly too deep to be chosen as a spawning
substrate during the first sampling survey). In late
April, the most preferred substrate for egg strand
deposition was worm weed followed by common
reed, while in early May the most preferred was
common reed, followed by trees and branches,
including black elder and worm weed (Table 1).
A multi-factorial ANOVA revealed that the dis-
tribution of egg strands of perch was highly influ-
enced by all the factors—the depth of deposition,
temperature at the depth of deposition, type of
spawning substrate and date of sampling—used as
predicted variables. The most important factor was
the depth of deposition (F5, 504 = 8.08, P \ 0.001),
followed by the type of spawning substrate (F2,
504 = 13.60, P \ 0.001), date of spawning (F1,
504 = 22.95, P \ 0.001) and temperature at the depth
of deposition (F6, 504 = 3.63, P \ 0.01).
Fig. 6 The composition of spawning substrate for perch in
Chabarovice Lake in spring 2007 during the first days of
spawning and at the end of spawning when most of the
spawning events occurred in deeper parts of the lake (cf.
Fig. 4). � Indicates dead plants, bushes or trees (cf. Fig. 3).
‘‘Others’’ category includes: emergent vegetation, technical
carpet and fence wire-cloth
Table 1 Ground coverage of potential/main perch spawning substrates in Chabarovice Lake in spring 2007 and the Ivlev electivity
index for those substrates during the first days of perch spawning (24–26 April) and at the end of spawning (2–3 May)
Spawning substrate Ground
coverage (%)
Share in submerged
vegetation (%)
Ivlev electivity indexa
April May
Bare bottom, especially mud 85 -0.91 -0.97
Submerged vegetation (dead or alive) 15 0.73 0.74
Curly pondweed 65.3 -0.28 -0.47
Worm weedb 5.1 0.75 0.54
Common reedb 5.7 0.42 0.69
Common rush and other ‘‘soft weeds’’b 6.7 -0.06 0.22
Eurasian water milfoil 5.5 -0.57 -0.59
Trees and branches including black elderb 8.2 -0.67 0.58
Common stonewort 3.5 -0.79 -1.00
Note that at the beginning of perch spawning period (late April) ‘trees and branches including black elder’ were mostly too deep
(C7 m) to be chosen as a spawning substrate, since the majority of perch spawning activity took place in the 2–4 m and 4–6 m depth
layers (cf. Figs. 3 and 4)
Bold numbers indicate significant positive or negative electivity for individual spawning substratesa According to Ivlev (1961)Ei defined as: Ei = (ri - ni)/(ri ? ni), where i is a type of spawning substrate, r is share (%) of substrate
in the total amount of used spawning substrates and n is share (%) of substrate in the lake (for individual types of submerged
vegetation it is share of substrate in the total amount of submerged vegetation). Values close to -1 indicate strong avoidance of
spawning substrate and values close to ?1 indicate strong preference of spawning substrate. Values higher than -0.3 and lower than
0.3 are regarded not to be significantly different from 0b Dead plants, bushes or trees (cf. Fig. 3)
Hydrobiologia (2009) 630:105–114 111
123

Discussion
According to the Czech Hydrometeorological Insti-
tute (unpubl. data), winter 2006/2007 was the warm-
est winter since 1922. For that reason, routine
SCUBA diving for monitoring of perch spawning
activity started in Chabarovice Lake very early, on 19
March with further attempts on 15 and 22 April. Up
to the latest of these dates, no egg strands were
observed.
The first bright white egg strands were recorded
during the night dive on 23rd April. The same night
diving, as well as night diving on 25th April, revealed
that a significant proportion of the perch females
remained unspawned. Hence, the first sampling
survey (24–26 April) could be considered as the
beginning of the perch spawning period (Craig,
1987).
Unusually, warm weather for the next 2 weeks
caused shortening of the spawning period (Gillet &
Dubois, 1995), since all the perch females recorded
during the night dive on 2nd May had already
released their eggs. Many egg strands, especially in
shallower depth layers (depth 2–6 m), were close to
hatching or in the middle of hatching. The difference
of 10–11 days corresponds well to the period of time
required for perch embryo development at tempera-
tures between 13 and 15�C (Guma’a, 1978). That was
why the second sampling survey (2–3 May) could be
referred to as the end of the main perch spawning
period.
During both sampling surveys, all but four egg
strands were found deeper than 2 m, which is in sharp
contrast to previous studies from various lakes and
reservoirs (Holcık, 1965; Frank, 1967; Stehlık, 1969;
Viljanen & Holopainen, 1982; Treasurer, 1983; Urho,
1996; Smith et al., 2001) but corresponds with the
findings of Lang (1987) and Gillet & Dubois (1995,
2007) from Lake Geneva, on the Swiss–French
border and with the findings of Probst et al. (2009)
from Lake Constance, on the Swiss–Austrian–Ger-
man border. The avoidance of the 0–2 m depth layer
as a spawning habitat is surprising since boulders,
beds of live common reed, bundles of common rush
and other ‘‘soft weeds,’’ while not as abundant as in
deeper layers (except of boulders), are generally
considered to be a suitable spawning substrate for
perch (Treasurer, 1983; Lang, 1987; Urho, 1996;
Smith et al., 2001).
It is most likely that, in Chabarovice Lake, the perch
avoided the shallowest littoral habitats due to relatively
high temperatures ([14�C), which may cause a
significant increase in egg mortality (Guma’a, 1978;
Sandstrom et al., 1997) and/or due to the supposed
damaging effect of waves (Probst et al., 2009).
Chabarovice is an opencast mine lake and the whole
surrounding landscape is deforested which makes the
lake vulnerable to winds from all directions. It has
already been described that strong winds provide a
potential mechanism for blowing perch egg strands
ashore (Clady & Hutchinson, 1975; Jones, 1982).
Similarly, there is evidence that ultraviolet radia-
tion can lead to a high mortality rate of perch eggs
incubated in the surface waters of lakes with low-
dissolved organic carbon (DOC) levels (Williamson
et al., 1997). Huff et al. (2004) have shown that 92%
of the eggs spawned in a low-DOC lake were located
at depths greater than 3 m, while 76% of eggs in the
higher-DOC lake were spawned in water less than
1 m deep. Unfortunately, the values of DOC were not
measured during the current study but during the first
sampling survey 87.2% of egg strands were deposited
at depths greater than 3 m and during the second
sampling survey this portion increased to 96.7%.
In Lake Geneva, perch have been reported to spawn
on both natural and artificial spawning substrate to a
maximum depth of 12–15 m (Gillet & Dubois, 1995;
Dubois et al., 1996; Gillet & Dubois, 2007). In
Lochaber Lake, Nova Scotia, females of the closely
related yellow perch Perca flavescens (Mitchill)
deposited their egg strands to a maximum depth of
12 m (Alto & Newsome, 1989). Similarly in Lake
Giles, Pennsylvania, spawned egg strands of yellow
perch were sporadically observed up to a depth of 15 m
(Huff et al., 2004). In the case of Chabarovice Lake,
however, egg strands were found up to a depth of
16.6 m during both sampling surveys. It seems that, at
least to this depth, their deposition and further devel-
opment was not limited by temperature (ca. 8.5�C),
dissolved oxygen (ca. 8.5 mg l-1) or suitable spawn-
ing substrate (worm weed, trees and branches includ-
ing black elder).
Previous studies from Rımov Reservoir and Lake
Geneva have shown that during the spawning period
warming of surface water could shift the depth of
perch spawning activity from shallower (depth 0–
4 m) towards deeper (depth 6–12 m) layers
(Kubecka, 1992; Gillet & Dubois, 1995, 2007), and
112 Hydrobiologia (2009) 630:105–114
123

these results are well supported by the data from
Chabarovice Lake. At the beginning of the spawning
period, the peak of perch spawning activity was
observed in the depth layer 2–4 m, however, less than
2 weeks later the peak of spawning activity was
observed in the deeper layer at 6–8 m.
The present study has shown that for egg strand
deposition perch do not use all the available spawn-
ing substrates in proportion to which they are present
in the lake. This is in agreement with previous studies
where perch generally preferred submerged vegeta-
tion and avoided bare bottom (e.g. Frank, 1967;
Holcık, 1969; Treasurer, 1983; Craig, 1987; Gillet &
Dubois, 1995; Urho, 1996; Nash et al., 1999; Smith
et al., 2001). Egg strands deposited directly on the
lake bottom may risk being smothered by silt or have
increased exposure to microorganisms (Smith et al.,
2001). They may also suffer from low levels of
dissolved oxygen, which is an important hazard for
developing embryos (Wootton, 1998).
The study of Chabarovice Lake, however, also
shows a significant difference in preference for
different types of submerged vegetation. While live
submerged vegetation (usually quite dense), although
more abundant, is generally avoided, dead submerged
vegetation (usually sparse) is highly preferred.
Perch egg strands deter predators (Newsome &
Tompkins, 1985); hence there is no apparent need to
hide them in dense vegetation. Moreover, dense, live
vegetation could deplete a reasonable amount of
oxygen from the surrounding water due to intensive
night respiration (Taiz & Zeiger, 2006). Since night
oxygen depletion, in the case of sparse dead
submerged vegetation (including periphyton), is of
much lower extent it seems reasonable that this type
of spawning substrate is highly preferred by perch.
The only exception is bundles of common rush and
other ‘‘soft weeds.’’ Due to its limited growth above
the bottom contour, this spawning substrate is
qualitatively somewhere between large, dead sub-
merged vegetation (common reed, worm weed, trees
and branches including black elder) and bare bottom.
Common reed, worm weed, trees and branches,
including black elder, are undoubtedly an ideal
spawning substrate for perch since placement of the
egg strands over those large dead structures, extend-
ing into the open water column, ensures that eggs
remain well oxygenated 24 hours a day (Reyes et al.,
1992). This finding is consistent with the results of
Nash et al. (1999) who used artificial spawning
substrates in Manchester Docks, UK, where perch
preferred branches of spruce Pilea abies, sycamore
Acer pseudoplatanus and willow Salix alba. It also
appears that hard, three-dimensional structures like
branches of spruce, sycamore and willow (Nash et al.,
1999) or stems of dead common reed, worm weed,
dislodged trees, bushes and their branches in general
(this study) are more appropriate substrates for egg
strand deposition by perch than the soft stems of any
live submerged vegetation.
Conclusion
The present study has shown that even in the system
without any ‘obvious limitation to perch spawning’
there are always some limitations. Perch did not use
for egg strands deposition shallowest littoral habitat
(high temperature causing increase of egg mortality,
damaging effect of waves) and when possible avoided
live submerged vegetation. Factors with significant
influence on the distribution of egg strands were as
follows (in order of decreasing influence): depth of
deposition, type of spawning substrate, date of
spawning and temperature at the depth of deposition.
In Chabarovice Lake, the most important substrates
for perch spawning—beds of dead common reed and
worm weed—reveal signs of progressive degradation
and this substrate will disappear from the lake within
several years. The spawning activity of perch in
shallower water (up to a depth of 7 m) will then have
to be concentrated almost exclusively on stands of live
vegetation whose presence in the lake is, however, less
predictable in time and depends on the weather
conditions during the year and the system of lake
filling. Moreover, the results have shown that live
vegetation is not much suitable spawning substrate for
perch. For that reason, installation of an artificial
spawning substrate (Gillet & Dubois, 1995; Dubois
et al., 1996; Nash et al., 1999; Pedicillo et al., 2008;
Probst et al., 2009) would, in the near future, be an
important means of both sustaining the stock of perch,
the main predatory fish in the lake, and for overall
management of the lake’s ichthyofauna.
Acknowledgements The authors thank B. Tlougan and M.
Burgis for careful reading and correcting the English and W.N.
Probst and one anonymous referee for valuable comments on
an earlier version of this manuscript. The study was supported
Hydrobiologia (2009) 630:105–114 113
123

by the Grant Agency of the Czech Republic (projects No. 206/
06/1371 and 206/09/P266), the Grant Agency of the Czech
Academy of Sciences (project No. 1QS600170504) and the
Palivovy kombinat Ustı nad Labem s.p.
References
Alto, S. K. & G. E. B. Newsome, 1989. Evidence of demic
structure for a population of yellow perch (Perca flaves-cens). Canadian Journal of Fisheries and Aquatic Sciences
46: 184–190.
Craig, J. F., 1987. The biology of perch and related fish. Croom
Helm Ltd, Kent.
Clady, M. D. & B. Hutchinson, 1975. Effect of high winds on
eggs of yellow perch, Perca flavescens, in Oneida Lake,
New York. Transactions of the American Fisheries Soci-
ety 104: 524–525.
Dubois, J. P., C. Gillet, S. Bonnet & Y. Chevalier-Weber,
1996. Correlation between the size of mature female perch
(Perca fluviatilis L.) and the width of their egg strands in
Lake Geneva. Annales Zoologici Fennici 33: 417–420.
Frank, S., 1967. The growth of perch, Perca fluviatilis, in the
course of the first year of life in two valley reservoirs in
Czechoslovakia. Ichthyologica (The Aquarium Journal)
39(3–4): 155–166.
Gillet, C. & J. P. Dubois, 1995. A survey of the spawning of
perch (Perca fluviatilis), pike (Esox lucius), and roach
(Rutilus rutilus), using artificial spawning substrate in
lakes. Hydrobiologia 300–301: 409–414.
Gillet, C. & J. P. Dubois, 2007. Effect of water temperature and
size of females on the timing of spawning of perch
Perca fluviatilis L. in Lake Geneva from 1984 to 2003.
Journal of Fish Biology 70: 1001–1014.
Guma’a, S. A., 1978. The effects of temperature on the
development and mortality of eggs of perch, Perca flu-viatilis. Freshwater Biology 8: 221–227.
Holcık, J., 1965. Dynamics of fish populations in Klıcava water
supply reservoir. PhD Thesis, Faculty of Science, Charles
University, Prague (in Slovak).
Holcık, J., 1969. The natural history of perch, Perca fluviatilis,
Linneaus 1758, in the Klıcava reservoir. Prace Laboratoria
Rybarstva 2: 269–305.
Huff, D. D., G. Grad & C. E. Williamson, 2004. Environmental
constraints on spawning depth of yellow perch: the roles of
low temperature and high solar ultraviolet radiation. Trans-
actions of the American Fisheries Society 133: 718–726.
Ivlev, V. S., 1961. Experimental ecology of the feeding of
fishes. Yale University Press, New Haven.
Jones, D. H., 1982. The spawning of perch (Perca fluviatilisL.) in Loch Leven, Kinross, Scotland. Fisheries Manage-
ment 13: 139–151.
Kubecka, J., 1992. Fluctuations in fyke-net catches during the
spawning period of the Eurasian perch (Perca fluviatilis)
in the Rımov Reservoir, Czechoslovakia. Fisheries
Research 15: 157–167.
Kubecka, J., J. Peterka, V. Drastık, T. Juza, M. Prchalova & M.
Rıha, 2007. Complex fish stock assessment of Chabarovice
Lake in year 2006. Report of the Biology Centre, Academy
of Sciences of the Czech Republic, Institute of Hydrobiol-
ogy (in Czech).
Lang, C., 1987. Mortality of perch, Perca fluviatilis L., esti-
mated from the size and abundance of egg strands. Journal
of Fish Biology 31: 715–720.
Nash, K. T., K. Hendry & D. Cragg-Hine, 1999. The use of
brushwood bundles as fish spawning media. Fisheries
Management and Ecology 6: 349–355.
Newsome, G. E. & J. Tompkins, 1985. Yellow perch egg
masses deter predators. Canadian Journal of Zoology 63:
2882–2884.
Pedicillo, G., F. Merulli, A. Carosi, P. Viali & M. Lorenzoni,
2008. The use of artificial spawning substrates as media to
support the reproduction of Eurasian perch in Lake
Piediluco. Hydrobiologia 609: 219–223.
Pivnicka, K., 1979. Production of perch and roach fry in the
Klıcava reservoir. Acta Societatis Zoologicae Bohemo-
slovacae 43(1): 43–47.
Probst, W. N., S. Stoll, H. Hofmann, P. Fischer & R. Eckmann,
2009. Spawning site selection by Eurasian perch
(Perca fluviatilis L.) in relation to temperature and wave
exposure. Ecology of Freshwater Fish 18: 1–7.
Reyes, M. R., H. H. Arzbach & E. Braum, 1992. In situ
development of perch eggs, Perca fluviatilis L. (Pisces,
Percidae) in a small eutrophic lake, Lake Plussee, Hol-
stein, Germany. Internationale Revue Ergebnisse Hydro-
biologie 77: 467–481.
Sandstrom, O., I. Abrahamsson, J. Andersson & M. Vetemaa,
1997. Temperature effects on spawning and egg devel-
opment in Eurasian perch. Journal of Fish Biology 51:
1015–1024.
Smith, C., A. Douglas & P. Jurajda, 2001. Oviposition site
selection and embryo mortality in perch. Journal of Fish
Biology 58: 880–882.
Stehlık, J., 1969. The fecundity of perch Perca fluviatilisLinneaus, 1758 in the Klıcava water reservoir. Acta So-
cietatis Zoologicae Bohemoslovacae 33(1): 88–95.
Taiz, L. & E. Zeiger, 2006. Plant Physiology. Sinauer Asso-
ciates Inc, Publishers, Sunderland.
Treasurer, J. W., 1983. Estimates of egg and viable embryo
production in a lacustrine perch, Perca fluviatilis. Envi-
ronmental Biology of Fishes 8/1: 3–16.
Urho, L., 1996. Habitat shifts of perch larvae as survival
strategy. Annales Zoologici Fennici 33: 329–340.
Viljanen, M. & I. J. Holopainen, 1982. Population density of
perch (Perca fluviatilis L.) at egg, larval and adult stages
in the dys-oligotrophic Lake Suomunjarvi, Finland. An-
nales Zoologici Fennici 19: 39–46.
Williamson, C. E., S. L. Metzgar, P. A. Lovera & R. E. Mo-
eller, 1997. Solar ultraviolet radiation and the spawning
habitat of yellow perch, Perca flavescens. Ecological
Applications 7: 1017–1023.
Wootton, R. J., 1998. The Ecology of Teleost Fishes. Kluwer
Academic Publishers, Dordrecht.
Zhakov, L. A., 1964. Method of assessment of absolute num-
bers of fish using census of perch egg strands. In Kitaev,
S. P. (ed.), Ozera Karelskogo peresejka. Izd. Nauka,
Moscow: 128–139 (in Russian).
114 Hydrobiologia (2009) 630:105–114
123




















