
A new classification of a preovulatory oocyte maturation stage suitable for the synchronization of...
16
Vol. 11, No. 3 194 ORIGINAL RESEARCH A new classification of a preovulatory oocyte maturation stage suitable for the synchronization of ovulation in controlled reproduction of Eurasian perch, Perca fluviatilis L Daniel Żarski 1,2 , Zoltán Bokor 3 , László Kotrik 3 , Béla Urbanyi 3 , Akos Horváth 3 , Katarzyna Targońska 2 , Sławomir Krejszeff 2 , Katarzyna Palińska 2, 4 , Dariusz Kucharczyk 2 2 Department of Lake and River Fisheries, University of Warmia and Mazury, Olsztyn, Poland; 3 Department of Aquaculture, Szent István University, Gödöllő, Hungary; 4 Department of Zoology, University of Warmia and Mazury, Olsztyn, Poland Received: 6 July 2011; accepted: 25 September 2011 SUMMARY To improve controlled reproduction of Eurasian perch, Perca fluviatilis, the criteria for the evaluation of final oocyte maturation stages were revised. The new classification covers six preovulatory maturational stages (I -VI) from the end of vitellogenesis to germinal vesicle breakdown (GVBD) and was based on macroscopic changes of preovulatory oocytes (position of the germinal vesicle, GVBD, oil droplets coalescence). The observation was performed during out-of-season artificial reproduction with the use of a single hCG injection (500 IU/kg). The classification was subsequently verified with the controlled reproduction of wild female perch with the use of hormonal stimulation (500 IU hCG/kg of body weight at 12°C). The females were at different maturational stages and constituted respective experimental groups (I-VI). During the experiment, 1 Corresponding author: Department of Lake and River Fisheries, University of Warmia and Mazury, Oczapowskiego 5; 10-957 Olsztyn, Poland; e-mail: [email protected]
Transcript of A new classification of a preovulatory oocyte maturation stage suitable for the synchronization of...

Vol. 11, No. 3 194
ORIGINAL RESEARCH
A new classification of a preovulatory oocyte maturation stage suitable for the
synchronization of ovulation in controlled reproduction of Eurasian perch,
Perca fluviatilis L Daniel Żarski1,2, Zoltán Bokor3, László Kotrik3, Béla Urbanyi3, Akos
Horváth3, Katarzyna Targońska2, Sławomir Krejszeff2, Katarzyna Palińska2, 4, Dariusz Kucharczyk2
2Department of Lake and River Fisheries, University of Warmia and Mazury, Olsztyn, Poland; 3Department of Aquaculture, Szent István
University, Gödöllő, Hungary; 4Department of Zoology, University of Warmia and Mazury, Olsztyn, Poland
Received: 6 July 2011; accepted: 25 September 2011
SUMMARY To improve controlled reproduction of Eurasian perch, Perca fluviatilis, the criteria for the evaluation of final oocyte maturation stages were revised. The new classification covers six preovulatory maturational stages (I -VI) from the end of vitellogenesis to germinal vesicle breakdown (GVBD) and was based on macroscopic changes of preovulatory oocytes (position of the germinal vesicle, GVBD, oil droplets coalescence). The observation was performed during out-of-season artificial reproduction with the use of a single hCG injection (500 IU/kg). The classification was subsequently verified with the controlled reproduction of wild female perch with the use of hormonal stimulation (500 IU hCG/kg of body weight at 12°C). The females were at different maturational stages and constituted respective experimental groups (I-VI). During the experiment,
1Corresponding author: Department of Lake and River Fisheries, University of Warmia and Mazury, Oczapowskiego 5; 10-957 Olsztyn, Poland; e-mail: [email protected]

195 Oocyte maturity in perch
ovulation appeared to be considerably synchronized within particular groups. Statistical differences in latency time (time between hormonal treatment and ovulation) were found between experimental groups (mean latency time: 110, 92, 68, 49, 29 and 18 h in groups representing VI, V, IV, III, II and I stage of the proposed classification, respectively). The proposed classification and the results presented in the study allowed for effective synchronisation of ovulation. The use of our new oocyte maturation classification may positively influence the effectiveness of Eurasian perch production. Reproductive Biology 2011 11 3: 194-209. Key words: controlled reproduction, Eurasian perch (Perca fluviatilis), oocyte maturation stage INTRODUCTON The Eurasian perch, Perca fluviatilis L., has been identified as a very important candidate species for freshwater aquaculture [23, 30], and the technology of its production is being constantly improved [9, 11, 21, 22, 25, 29, 31]. Controlled reproduction is an important part of the culture process that determines egg and larval quality and quantity [4, 15, 16, 23, 26, 36] with further implications in aquaculture [3]. Like other percids, the Eurasian perch has been reproduced under controlled conditions without using hormonal agents. However, if hormonal preparations were not administered, a huge asynchronous ovulation was observed [15, 16, 27, 29, 31]. Unpredictable latency time (time between hormonal treatment and ovulation) was the reason for the deposition of eggs in the tank without human control [16, 29]. Variability in the latency time results in size differentiation of the larvae during rearing and can thereby intensify cannibalism which is a significant limiting factor during perch larval rearing [1, 5]. It has been reported that the necessity and dosage of a hormonal preparation during the reproductive season in percids depends on the maturity stage of oocytes [15, 16, 29]. Thus, precise determination of the oocyte maturity stage before hormonal stimulation could allow for precise prediction of the moment of ovulation which is important for the control of fertilization process. It is especially necessary when “dry” eggs are needed for applications such as genome manipulation (androgenesis, gynogenesis, polyploidization) or fertilization with cryopreserved sperm.

196 Żarski et al
In artificial reproduction, prediction of ovulation is achieved only by the determination of oocyte maturity [12, 17, 33] i.e. determination of position of germinal vesicle (GV) in oocytes. The relationship between the maturation stage and latency time was also found in percids [29, 32]. The following classification, developed for cyprinids [2], has been applied in fish reproduction studies. It includes four stages: - stage I: GV is in the centre of oocyte, - stage II: early stage of a migrating GV, GV is less than half the
oocyte diameter, - stage III: late stage of a migrating GV, GV is more than half the
oocyte diameter, - stage IV: peripheral GV or germinal vesicle break down (GVBD). Such classification accurately describes the final oocyte maturation (FOM) in many cyprinid species, and its application gives excellent reproductive effects, taking into account the ovulation time [12, 17, 35]. However, for many fish species, such as Eurasian perch [16, 24], eel Anguilla anguilla [28], pikeperch Sander lucioperca (L.) [32] or some species of marine fish, e.g., sea bass Dicentrarchus labrax (L.) [6] this description is not sufficiently precise. In fact, it is extremely difficult to predict the time of ovulation with the use of this system in percids because of a large variability in latency time reported earlier [15, 16, 29, 31, 32, 34]. Thus, it is important to verify and specify the process of oocyte maturation in percids.
The aim of the study was to develop the classification of oocyte maturation stages in Eurasian perch for the improvement of controlled reproduction. In this study, changes which take place during FOM have been analyzed and classified into six pre-ovulatory stages. Induced reproduction has also been carried out in order to verify the system intended to evaluate the maturity of females.
MATERIALS AND METHODS Determination of maturation stages All experiments were performed in accordance with the principles and procedures of Animal Ethics Committee of the University of Warmia and Mazury in Olsztyn. The determination of perch oocytes’s maturation stages was conducted during induced out-of-season reproduction. This procedure was chosen because the maturation stage of females during a season is very variable among individuals, even

197 Oocyte maturity in perch
within one population. Only perch spawners captured in the late autumn and kept in strictly controlled conditions had a high probability of being in the same maturation stage before the FOM starts. Perch spawners (9 females and 4 males), with an average weight of 467±122 g, were caught in cage-traps in Lake Sasek Wielki (north-eastern Poland) on October 25, 2009. Immediately afterwards, the fish were transported to the hatchery of the Department of Lake and River Fisheries, Olsztyn, and placed into a 1 000 l tank with controllable environmental conditions [18]. The fish were acclimated for 10 days at 10ºC. Subsequently, water was gradually cooled down for 15 days (by less than 0.5°C a day) to 4°C. They were subsequently kept at that temperature for 60 days. The fish were kept in darkness (0L:24D) during this period. Then, the water temperature was gradually increased (by less than 1ºC a day) up to 12°C, and the photoperiod duration was gradually extended to 12L:12D. Next, the fish were marked and randomly divided into two groups. The females which were given an intraperitoneal injection of physiological saline (0.9% NaCl) were the control group (n=5), whereas the fish in the experimental group (n=4) were given human chorionic gonadotropin (hCG; Biomed, Poland) at a dose of 500 IU/kg. Following stimulation, the temperature of water in the tank was raised to 14°C (by 1°C a day) and the photoperiod was set to 14L:10D. This procedure, including thermal manipulations was previously reported to be effective in Eurasian perch out-off-season reproduction [31]. These conditions were maintained until the end of the experiment. During the entire period of this experiment females (n=9) were kept together with males (n=4).
Every day at the same time, starting with the day of hormonal stimulation, each female from both groups was weighed (±0.1 g), and oocytes were sampled with a catheter. The oocytes were put in Petri dishes and poured over with Serra’s solution (70% ethanol, 40% formaldehyde and 99.5% glacial acetic acid in the ratio of 6:3:1). Following cytoplasm clarification, the samples were photographed under a stereoscopic microscope (Leica MZ 12.5, Switzerland, Heerbrugg). The procedure was repeated every day until spontaneous spawning of hCG-treated females took place (i.e. the appearance of eggs in the form of ribbons). The spontaneous spawning was chosen to check the time when females were ready to spawn without additional manipulations. Since both females and males were present

198 Żarski et al
in the tank, the obtained egg ribbons were fertilized. Samples of oocytes from the control group were taken for subsequent five days after spawning of hCG-treated fish was completed. The documentation produced in this manner was used to analyze changes occurring in 30 randomly chosen oocytes from each sample and the results of measurements were taken within an accuracy of 0.01 mm with the use of software for acquisition and analysis of pictures taken under the stereoscopic microscope (ProgRes® Capture Pro 2.5, Jenoptic, Germany). The oocyte diameter at each maturation stage and the diameter of oil droplets at a proposed stage V were measured. In addition, three egg samples (200-300 eggs each) were taken from each ribbon and subsequently incubated in separate 1.5 l tanks. The incubation was carried out in a closed water system at 14°C [14]. Embryo survival was determined at the pigmented-eyed stage.
Practical implementation of the proposed classification of oocyte maturation Perch spawners (47 females with an average weight 148±41 g and 14 males with an average weight 170±78 g) were obtained at the end of March from Lake Sasek Wielki. The maximum water temperature in the lake did not exceed 10°C by that time. Immediately after the fish were caught, they were transported to the hatchery and placed in the 1000 l tanks with controlled temperature and photoperiod [18]. After a three day acclimation period, oocyte samples from each female were taken with a catheter and were immersed in the Serra’s solution. After clarification of the cytoplasm maturation, the stage of the oocytes was recognized according to the developed classification. Afterwards, the females were tagged and divided into six groups according to their maturation stage. The decisive criterion was the majority of oocytes (over 50%) representing a specific stage. In this way, groups representing particular stages of oocyte maturation (I, II, III, IV, V and VI) were isolated. Next, all females were injected with hCG (500 IU/kg). After injection, the temperature was raised up to 12°C. The males were kept in separate tanks where the temperature was maintained at 12°C. Males were not stimulated hormonally.
The control of ovulation, performed by gentle pressure of the abdomen in order to determine if the eggs were able to be stripped, started 20, 30, 50, 70, 90 hours following hormonal treatment in groups representing V, IV, III, II and I stage, respectively. Next, all

199 Oocyte maturity in perch
females were checked for ovulation every 5-6 h until the first female in a respective group ovulated. Afterwards, the rest of the females from this group were checked every 3-4 h. Ovulation control in the group representing stage VI started 5 h after the hormonal stimulation and was carried out every 2-3 hours. If ovulation occurred, eggs from each female were stripped to a separate dry plastic container. After spawning, the females were transferred to a separate tank. The latency time was recorded throughout the experiment. When the females were found to deposit eggs in the tank, the spawned female was localized and removed from the tank. The latency time for females which ovulated in the tank was regarded as the moment when the female was localized. Female mortality rate was recorded during the examination in each group until day 5 after spawning. Freshly obtained and pooled (from three males) sperm was used to fertilize the egg-ribbons. Sperm was collected with syringes. For each 100 g of eggs, 1 ml of pooled sperm was used. Clean hatchery water was used for the activation of gametes. Whole ribbons were incubated in the floating plastic cages which allowed the circulation of well oxygenated water around the whole ribbon [36] in a closed recirculating system. The temperature of incubation was 14°C. The survival rate of embryos at the pigmented-eyed stage was recognized on the basis of 100 eggs randomly photographed under the stereoscopic microscope (in triplicates for each ribbon). All manipulations were done after the fish were anaesthetized in the MS-222 (Argent, Redmond, Washington, USA; 150 mg/l).
Statistical analysis Statistical analysis was performed with STATISTICA 9.0 (StatSoft Inc., Tulsa, Oklahoma, USA) software and MS Excel for Windows. All data were expressed as mean±SD. Before the statistical analysis, the data expressed in percentage were subjected to arcsin transformation. Regression analysis was performed to compare the females’ body weight and oocytes’ maturity stage. The determination of maturation stages (oocyte diameter, oil droplet diameter at the proposed stage V) was analyzed by Student t-test (at a 5% significance level). In experimental verification of the proposed classification of oocyte maturity stages, the data were subjected to one-way analysis of variance (ANOVA) followed by a post-hoc Duncan’s test (at a 5% significance level).

200 Żarski et al
RESULTS Determination of maturation stages No female mortality was observed during the experiment. During out-of -season spawning, 75% of hCG-treated females ovulated. Ovulation took place after an increase in the body weight by 11% relative to the initial weight. No ovulation or body weight growth was observed in the control, untreated group. The survival rate of embryos at the pigmented-eyed stage was 63±21%.
During the experiment, the migration of GV from the centre of the oocyte to its periphery was observed, and was accompanied by a coalescence of oil droplets into one large droplet. Next, GVBD was observed shortly before ovulation. These morphological changes were associated with an increase in the body weight of females (fig. 1). The changes provided grounds for the determination of the stages which describe consecutive changes of maturing oocytes (fig. 2). The changes were divided into seven stages (six preovulatory stages and ovulation regarded as a seventh stage): - stage I: the GV is situated in the oocyte centre, oil droplets are poorly
visible (fig. 2a); - stage II: beginning of GV migration, beginning of coagulation of clearly
visible oil droplets (fig. 2b); - stage III: migration of GV (above half of the oocyte diameter), oil
droplets are clearly visible (fig. 2c); - stage IV: the GV is at the oocyte periphery, a large forming oil droplet is
clearly visible; the droplet diameter is greater than the GV diameter and it reaches the size of about 1/3 of the oocyte diameter; smaller droplets are also visible (fig. 2d);
- stage V: the GV is situated at the oocyte edge, one large (size of about half the oocyte diameter) oil droplet is clearly visible (fig. 2e);
- stage VI: oocyte samples taken for analysis are macroscopically transparent; no visible GV after they were placed in Serra’s solution (following GVBD), oocytes at the pre-ovulation stage (fig. 2f);
- stage VII: ovulation. The experiment did not reveal any differences among individual
fish or among individual development stages in terms of oocyte diameter (t-test, p>0.05). The value remained within the range of 0.99-1.08 mm at all stages. No differences were recorded among oil droplet diameters in different individuals at the proposed stage V (t-test, p>0.05), and the diameter stayed within the range of 0.46-0.53 mm.
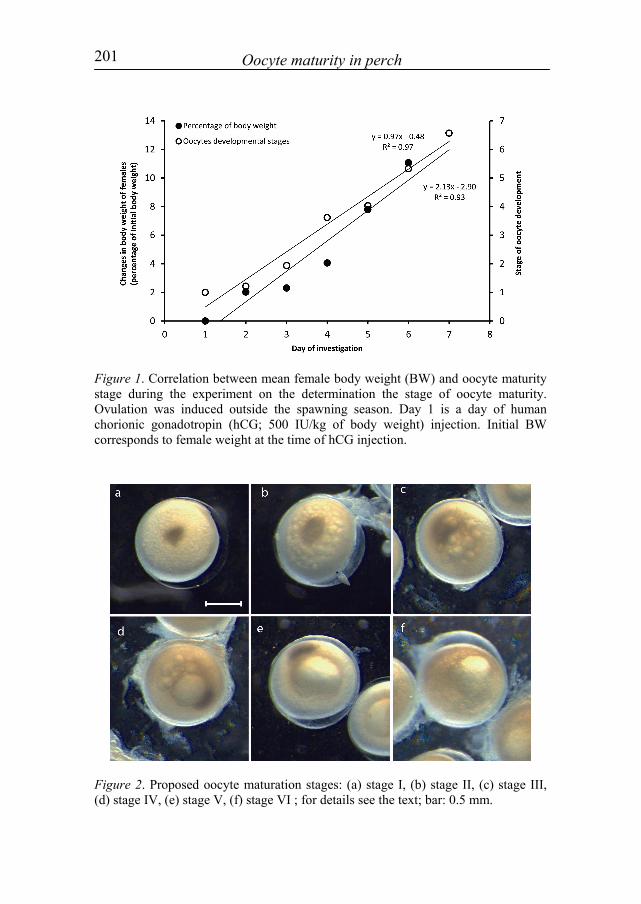
201 Oocyte maturity in perch
Figure 1. Correlation between mean female body weight (BW) and oocyte maturity stage during the experiment on the determination the stage of oocyte maturity. Ovulation was induced outside the spawning season. Day 1 is a day of human chorionic gonadotropin (hCG; 500 IU/kg of body weight) injection. Initial BW corresponds to female weight at the time of hCG injection.
Figure 2. Proposed oocyte maturation stages: (a) stage I, (b) stage II, (c) stage III, (d) stage IV, (e) stage V, (f) stage VI ; for details see the text; bar: 0.5 mm.

202 Żarski et al
Practical implementation of the proposed classification of oocyte maturation The highest percentage of ovulation (100% ovulation rate; p<0.05) was recorded in groups with the most advanced oocyte maturation (stages IV, V and VI) established at the time of hormonal stimulation. The ovulation rate in the remaining groups (stages I, II and III) ranged from 63 to 78%, and no differences were found among these groups (p>0.05; tab. 1). The survival of females was the lowest in groups representing stage I and II (50%), and the highest in group representing maturation stage VI (88%). However, no significant differences were found (p>0.05) in the female survival rate among the examined groups. The survival of embryos was related to the oocyte maturation stage at the time of hCG treatment. In female groups with less advanced maturation stage (stages I and II), embryo survival was the lowest (p<0.05). Embryo survival was similar (p>0.05) in the remaining groups (stages III, IV, V and VI). In contrast, latency times differed among all groups (p<0.05; tab. 1). Table 1. Results of controlled reproduction of Eurasian perch, Perca fluviatilis, obtained during practical implementation of the proposed classification of oocyte maturation
Each treatment group was injected with hCG (500 IU/kg of body weight) during I, II, III, IV, V or VI oocyte maturity stage; different superscript depict significant (p<0.05) differences among groups; n: number of fish; *time between hormonal stimulation and ovulation.

203 Oocyte maturity in perch
DISCUSSION If fish spawn once a year, a feature of fish production is seasonal fluctuation of the commercial-size fish supply. One of the measures used to increase production capacity is to carry out reproduction year round which entails the necessity to conduct out-of-season reproduction. It is then necessary to perform hormonal stimulation in order to obtain high quality gametes [10, 20, 31]. In percids, hCG stimulation is applied successfully and induces ovulation after a single injection [15]. In the present study, hCG administration caused synchronous ovulation in 75% of females during the out-of-season spawning. It is noteworthy that the variable quality of oocytes (expressed as embryo survival at the pigmented-eyed stage) is typical for off-season reproduction in percids [10, 25, 31]. Therefore, the results of out-of-season reproduction obtained in the study should be regarded as satisfactory. The observed variability in embryo survival could also result from variable effectiveness of spontaneous reproduction and other potentially stress-inducing manipulations applied (e.g. catheterization). Such variable results were obtained by Ronyai and Lengyel [29] during the reproductive season when spontaneous reproduction was examined. Thus, the FOM observed in the present study proceeded typically for the species. Moreover, none of the changes observed in hCG-treated fish were observed in control females examined at the end of the experiment. The lack of progression in maturation of females which were not hormonally stimulated confirms the necessity of hormonal stimulation in out-of-season reproduction in Eurasian perch [23, 31].
The proposed classification of oocyte maturity stages covers the entire FOM including stage I in which, in addition to the GV in the central position, yolk granularities at the oocyte edge were also visible [24, 30]. It is noteworthy that no oil droplets were visible at this stage (fig. 2a). Only the application of the out-of-season reproduction allowed oocytes to be observed at the beginning of FOM and simultaneous progression in maturation in all hCG-treated females.
The germinal vesicle position in non-ovulated perch oocytes was difficult to determine when the previous (developed for cyprinids) classification was used. The precise localization of the GV requires tilting or rolling individual oocyte for it be viewed from several angles. Variability in the intensity of cytoplasm clarification with the use of Serra’s solution might also have a negative effect on maturation

204 Żarski et al
stage determination. This issue especially refers to stages II and III in the previous classification.
It was previously reported that the coalescence rate of oil droplets could be a valuable and helpful additional morphological indicator of oocyte maturity [32]. Migaud et al. [24] conducted a histological analysis of FOM in perch and described nucleus migration and oil droplet coalescence. However, these authors assigned the process of GV migration and oil droplet coalescence to one maturational stage. In the present study, the same phenomena were described not in one but in three stages (III, IV, V). This move is justified by the relatively long period of the transformations (without hormonal stimulation even 3-4 weeks) during which the GV migrates and the oil droplets coalesce. For comparison, GVBD and ovulation last only few days [24]. Moreover, from stage III to V of the proposed classification, the female body weight increased rapidly (over 7%) which probably resulted from high gonadal hydration rate. A comparable tendency was reported for FOM perch by Sulistyo et al. [30]. These observations pointed to the significant and rapid changes occuring in the ovary between stage III and V of the proposed classification, reflected additionally in changes in oil droplet coalescence. Palstra et al. [28] also regarded oil droplet coagulation in oocytes of the European eel as one of the criteria of oocyte maturity. Migaud et al. [24] and Fontaine et al. [7] reported that in the Eurasian perch, the GVBD stage and ovulation stage (proposed stages VI and VII) were observed in relatively short time intervals, as compared to GV migration progression. These observations are of great importance for the practice of artificial reproduction since they allow planning and synchronisation of reproduction by hormonal stimulation.
Females were classified to the respective maturation stage according to the percentage of oocytes being at respective stages since usually oocytes of two stages were found in one female at the same time. This was observed earlier in perch, regardless of the applied classification [15, 16, 24]. In the second experiment, the range of latency time resulted from the fact that not all females in each group possessed 100% of oocytes at the same maturity stage. Such synchronic maturation stage is very difficult to achieve when wild spawners are reproduced [15, 16, 24, 29, 31]. It should be noted that fish caught at the same time from the same population are characterized by oocytes being at various maturation stages. Such a

205 Oocyte maturity in perch
phenomenon was reported earlier for pikeperch [36], and additionally confirms the necessity to determine the maturational stage of oocytes when wild fish are considered for reproduction. During the study, in which fish at each stage (I, II, III, IV, V and VI) were treated as separate experimental groups, the latency time differed among them. These results strongly supported the notion that a new classification of oocyte maturation is justified. The application of the “old” classification (for cyprinids) in percids has created many ambiguities. For example, on the basis of the descriptions given by Zakęś and Demska-Zakęś [34] or by Szkudlarek et al. [32] for pikeperch, groups representing I and II maturity stages of the proposed classification would be assigned to stage I of the “old” classification. Moreover, fish from groups representing III, IV and V maturation stage would be classified as one group. It can be concluded that the new classification system organizes in a simple way the recognition of maturation stage in perch, and determines more precisely the latency time following hormonal stimulation. It has to be pointed out that the proposed classification was already successfully applied in pikeperch [37] which makes it very useful also for other percids. This is particularly important when the reproduction of fish from different populations or with different degrees of domestication must be synchronized. The maturity of such fish varies [12, 13, 19], and synchronization of reproduction is frequently a condition of production success. For this purpose, it may be recommended to divide perch spawners according to the maturation stages and induce spawning at preferable times with proper hormonal and environmental manipulations. However, it should be noted that prediction of the moment of ovulation using this system should be verified under local conditions because of the inter-population differences. The type of hormonal treatment may also affect latency time significantly [8, 15, 16, 35].
ACKNOWLEDGEMENTS This study was partially financed by the project “Innovations in finfish aquaculture with special references to reproduction” (InnovaFish), Operational Programme Sustainable Development of the Fisheries Sector and Coastal Fishing Areas 2007-2013" (OR14-61724-OR1400003/09/10/11).

206 Żarski et al
REFERENCES 1. Baras E, Kestemont P, Melard C 2003 Effect of stocking density
on the dynamics of cannibalism in sibling larvae of Perca fluviatilis under controlled conditions. Aquaculture 219 241-255.
2. Brzuska E 1979 The in vivo method of estimating the stages of oocytes maturation in carp Cyprinus carpio L. Acta Hydrobiologica 21 423-433.
3. Bobe J, Labbe C 2010 Egg and sperm quality in fish. General and Comparative Endocrinology 165 535-548.
4. Cejko BI, Kucharczyk D, Targońska K, Kubiak D, Starosiek B, Glogowski J 2008 Quality parameters and selected biochemical markers of asp, Aspius aspius (L.), semen obtained after hormonal stimulation with Ovaprim or Ovopel. Archives of Polish Fisheries 16 179-188.
5. Fiogbé ED, Kestemont P 2003 Optimum daily ration for Eurasian perch Perca fluviatilis L. reared at its optimum growing temperature. Aquaculture 216 243-252.
6. Firat K, Saka S, Suzer C 2005 Gonadal oocyte development in LHRHa hormone treated European sea bass (Dicentrarchus labrax L., 1758) Broodstock. Turkish Journal of Veterinary and Animal Sciences 29 83-87.
7. Fontaine P, Migaud H, Mandiki R, Gardeur JN, Kestemont P, Fostier A 2003 Involvement of sex steroids in final stages of oogenesis in Eurasian perch, Perca fluviatilis. Fish Physiology and Biochemistry 28 331-332.
8. Horvath L, Szabo T, Burke J 1997 Hatchery testing of GnRH analogue-containing pellets on ovulation in four cyprinid species. Polish Archives of Hydrobiology 44 281-285.
9. Kestemont P, Vandeloise E, Mélard C, Fontaine P, Brown PB 2001 Growth and nutritional status of Eurasian perch Perca fluviatilis fed graded levels of dietary lipids with or without added ethoxyquin. Aquaculture 203 85-99.
10. Kolkovski S, Dąbrowski K 1998 Off-season spawning of yellow perch. Progressive Fish-Culturist 60 133-136.
11. Kouril J, Linhart O, Relot P 1997 Induced spawning of perch by means of a GnRH analogue. Aquaculture International 5 375-377.

207 Oocyte maturity in perch
12. Krejszeff S, Targońska K, Żarski D, Kucharczyk D 2009 Domestication affects spawning of the ide (Leuciscus idus)-preliminary study. Aquaculture 295 145-147.
13. Krejszeff S, Targońska K, Żarski D, Kucharczyk D 2010a Artificial reproduction of two different spawn-forms of the chub. Reproductive Biology 10 67-74.
14. Krejszeff S, Żarski D, Kucharczyk D, Kupren K, Targońska K, Mamcarz A 2010b An experimental device for eggs incubation and fish larvae rearing under laboratory conditions. Polish Journal of Natural Science 25 190-199.
15. Kucharczyk D, Kujawa R, Mamcarz A, Skrzypczak A, Wyszomirska E 1996 Induced spawning in perch, Perca fluviatilis L. using carp pituitary extract and HCG. Aquaculture Research 27 847-852.
16. Kucharczyk D, Kujawa R, Mamcarz A, Skrzypczak A, Wyszomirska E 1998 Induced spawning in perch, Perca fluviatilis L., using FSH + LH with pimozide or metoclopramide. Aquaculture Research 29 131-136.
17. Kucharczyk D, Kujawa R, Mamcarz A, Targońska-Dietrich K, Wyszomirska E, Glogowski J, Babiak I, Szabo T 2005 Induced spawning in bream (Abramis brama L.) using pellets containing GnRH. Czech Journal of Animal Science 50 89-95.
18. Kujawa R, Kucharczyk D, Mamcarz A 1999 A model system for keeping spawners of wild and domestic fish before artificial spawning. Aquacultural Engineering 20 85-89.
19. Kujawa R, Kucharczyk D, Mamcarz A, Żarski D, Targońska K 2011 Artificial spawning of common tench Tinca tinca (Linnaeus, 1758), obtained from wild and domestic stocks. Aquaculture International 19 513-521.
20. Malison JA, Procarione LS, Kayes TB, Hansen JF, Held JA 1998 Induction of out-of-season spawning in walleye (Stizostedion vitreum). Aquaculture 163 151-161.
21. Mandiki SNM, Blanchard G, Melard C, Koskela J, Kucharczyk D, Fontaine P, Kestemont P 2004 Effects of geographic origin on growth and food intake in Eurasian perch (Perca fluviatilis L.) juveniles under intensive culture conditions. Aquaculture 229 117-128.
22. Mélard C, Baras E, Mary L, Kestemont P 1996 Relationships between stocking density, growth, cannibalism and survival rate in

208 Żarski et al
intensively cultured larvae and juveniles of perch (Perca fluviatilis). Annales Zoologici Fennici 33 643-651.
23. Migaud H, Fontaine P, Sulistyo I, Kestemont P, Gardeur J-N 2002 Induction of out-of-season spawning in Eurasian perch Perca fluviatilis: effects of rates of cooling and cooling durations on female gametogenesis and spawning. Aquaculture 205 253-267.
24. Migaud H, Mandiki R, Gardeur J-N, Fostier A, Kestemont P, Fontaine P 2003 Synthesis of sex steroids in final oocyte maturation and induced ovulation in female Eurasian perch, Perca fluviatilis. Aquatic Living Resources 16 380-388.
25. Migaud H, Gardeur N-J, Kestemont P, Fontaine P 2004a Off-season spawning of Eurasian perch Perca fluviatilis. Aquaculture International 12 87-102.
26. Migaud H, Fontaine P, Kestemont P, Wang N, Brun-Bellut J 2004b Influence of photoperiod on the onset of gonadogenesis in Eurasian perch Perca fluviatilis. Aquaculture 241 561-574.
27. Migaud H, Wang N, Gardeur J-N, Fontaine P 2006 Influence of photoperiod on reproductive performances in Eurasian perch Perca fluviatilis. Aquaculture 252 385-393.
28. Palstra AP, Cohen EGH, Niemantsverdriet PRW, van Ginneken VJT, van den Thillart GEEJM 2005 Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture 249 533-547.
29. Ronyai A, Lengyel SA 2010 Effects of hormonal treatments on induced tank spawning of Eurasian perch (Perca fluviatilis L.). Aquaculture Research 41 e345-e347.
30. Sulistyo I, Rinchard J, Fontaine P, Gardeur JN, Capdeville B, Kestemont P 1998 Reproductive cycle and plasma levels of sex steroids in female Eurasian perch Perca fluviatilis. Aquatic Living Resources 11 101-110.
31. Szczerbowski A, Kucharczyk D, Mamcarz A, Łuczyński MJ, Targońska K, Kujawa R 2009 Artificial off-season spawning of Eurasian perch Perca fluviatilis L. Archives of Polish Fisheries 17 95-98.
32. Szkudlarek M, Kujawa R, Mamcarz A, Bieńkiewicz M, Kestemont P, Szczerbowski A, Łuczyński MJ, Krejszeff S, Targońska K, Kucharczyk D 2007 Checking maturation stages of females. In: Kucharczyk D, Kestemont P, Mamcarz A (eds)

209 Oocyte maturity in perch
Artificial reproduction of pikeperch, Mercurius Kaczmarek Andrzej, Olsztyn, Poland.
33. Yueh W, Chang C 2000 Morphological changes and competence of maturing oocytes in the protandrous black porgy, Acanthopagrus schlegeli. Zoological Studies 39 114-122.
34. Zakęś Z, Demska-Zakęś K 2009 Controlled reproduction of pikeperch Sander lucioperca (L.): a review. Archives of Polish Fisheries 17 153-170.
35. Żarski D, Kucharczyk D, Targońska K, Jamróz M, Krejszeff S, Mamcarz A 2009 Application of ovopel and ovaprim and their combinations in controlled reproduction of two reophilic cyprinid fish species. Polish Journal of Natural Science 24 235-244.
36. Żarski D, Palińska K, Targońska K, Bokor Z, Kotrik L, Krejszeff S, Kupren K, Horvath A, Urbanyi B, Kucharczyk D 2011a Oocyte quality indicators in Eurasian perch, Perca fluviatilis L., during reproduction under controlled conditions. Aquaculture 313 84-91.
37. Żarski D, Kucharczyk D, Targońska K, Palińska K, Kupren K, Fontaine P, Kestemont P 2011b A new classification of pre-ovulatory oocyte maturation stages in pikeperch, Sander lucioperca (L.), and its application during artificial reproduction. Aquaculture Research (in press).




















