
Phylogeography and species discrimination in the Paracalliope fluviatilis species complex...
10
Phylogeography and species discrimination in the Paracalliope fluviatilis species complex (Crustacea: Amphipoda): can morphologically similar heterospecifics identify compatible mates? DARIN L. SUTHERLAND†, IAN D. HOGG* and JOSEPH R. WAAS Department of Biological Sciences, University of Waikato, Private Bag 3105, Hamilton, New Zealand Received 18 April 2009; accepted for publication 24 July 2009We examined the phylogeographic patterns of Paracalliope fluviatilis (Amphipoda) over its entire range and the prevalence of mate discrimination in laboratory mate choice tests using genetically distinct populations. We predicted that mate discrimination would increase as the level of genetic divergence increased between popula- tions. Thirty different haplotypes were found with pairwise sequence divergences in the range 1–23.5% between locations. Individuals were selected from seven genetically distinct populations and males were presented with ‘local’ (same population) or ‘foreign’ (genetically divergent) females. Males were more likely to pair with local than foreign females but there was no evidence of a gradual increase in discrimination. Discrimination became most prominent (approximately 5 : 1 local : foreign) when genetic divergences exceeded approximately 20%; matings between divergent individuals also resulted in significantly fewer females producing eggs. We suggest that: (1) this abrupt shift in discrimination occurs because individuals from different, but similarly divergent, clades rely on different recognition cues (e.g. moulting pheromones with disparate chemical signatures) that trigger recognition and subsequent discrimination of incompatible mates; (2) geological history associated with sea level changes and a series of isolation events may be responsible for the patterns of discrimination that we observed; and (3) amphipods may be more genetically variable relative to other invertebrate taxa. © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205. ADDITIONAL KEYWORDS: Arthropoda – biodiversity – cryptic – mate choice – mate guarding – outbreeding – recognition. INTRODUCTION The ability of individuals to recognize, and hence mate with, members of the same species, is an impor- tant component of reproductive success and, ulti- mately, speciation processes (Mayr, 1982; Colgan, 1983). Deviation from random mating between differ- ent populations may indicate the existence of differ- ent species or at least processes initiating speciation. Accordingly, studies of species recognition would help to identify or validate cryptic species, which cannot be elucidated using traditional morphological taxonomy, but may be indicated using molecular phylogenetic approaches (Adamowicz et al., 2007; Cook, Bunn & Hughes, 2007; Thum & Harrison, 2009). Additionally, experiments involving multiple, inter-population comparisons may improve our understanding of how genetic distances correspond with species-level boundaries. For example, is there a level of diver- gence above which species recognition occurs? The order Amphipoda is one of the most speciose aquatic groups (Ruppert & Barnes, 1994), and studies have indicated that species diversity in this group may actually be underestimated, primarily because of the existence of morphologically similar, but geneti- cally distinct (cryptic), species (Hogg et al., 1998; Witt *Corresponding author. E-mail: [email protected] †Present address: New Zealand Department of Conservation, Private Bag 701, Hokitika, New Zealand. Biological Journal of the Linnean Society, 2010, 99, 196–205. With 5 figures © 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205 196
Transcript of Phylogeography and species discrimination in the Paracalliope fluviatilis species complex...

Phylogeography and species discrimination inthe Paracalliope fluviatilis species complex(Crustacea: Amphipoda): can morphologically similarheterospecifics identify compatible mates?
DARIN L. SUTHERLAND†, IAN D. HOGG* and JOSEPH R. WAAS
Department of Biological Sciences, University of Waikato, Private Bag 3105, Hamilton, New Zealand
Received 18 April 2009; accepted for publication 24 July 2009bij_1344 196..205
We examined the phylogeographic patterns of Paracalliope fluviatilis (Amphipoda) over its entire range and theprevalence of mate discrimination in laboratory mate choice tests using genetically distinct populations. Wepredicted that mate discrimination would increase as the level of genetic divergence increased between popula-tions. Thirty different haplotypes were found with pairwise sequence divergences in the range 1–23.5% betweenlocations. Individuals were selected from seven genetically distinct populations and males were presented with‘local’ (same population) or ‘foreign’ (genetically divergent) females. Males were more likely to pair with local thanforeign females but there was no evidence of a gradual increase in discrimination. Discrimination became mostprominent (approximately 5 : 1 local : foreign) when genetic divergences exceeded approximately 20%; matingsbetween divergent individuals also resulted in significantly fewer females producing eggs. We suggest that: (1) thisabrupt shift in discrimination occurs because individuals from different, but similarly divergent, clades rely ondifferent recognition cues (e.g. moulting pheromones with disparate chemical signatures) that trigger recognitionand subsequent discrimination of incompatible mates; (2) geological history associated with sea level changes anda series of isolation events may be responsible for the patterns of discrimination that we observed; and (3)amphipods may be more genetically variable relative to other invertebrate taxa. © 2010 The Linnean Society ofLondon, Biological Journal of the Linnean Society, 2010, 99, 196–205.
ADDITIONAL KEYWORDS: Arthropoda – biodiversity – cryptic – mate choice – mate guarding – outbreeding– recognition.
INTRODUCTION
The ability of individuals to recognize, and hencemate with, members of the same species, is an impor-tant component of reproductive success and, ulti-mately, speciation processes (Mayr, 1982; Colgan,1983). Deviation from random mating between differ-ent populations may indicate the existence of differ-ent species or at least processes initiating speciation.Accordingly, studies of species recognition would helpto identify or validate cryptic species, which cannot be
elucidated using traditional morphological taxonomy,but may be indicated using molecular phylogeneticapproaches (Adamowicz et al., 2007; Cook, Bunn &Hughes, 2007; Thum & Harrison, 2009). Additionally,experiments involving multiple, inter-populationcomparisons may improve our understanding ofhow genetic distances correspond with species-levelboundaries. For example, is there a level of diver-gence above which species recognition occurs?
The order Amphipoda is one of the most specioseaquatic groups (Ruppert & Barnes, 1994), and studieshave indicated that species diversity in this groupmay actually be underestimated, primarily because ofthe existence of morphologically similar, but geneti-cally distinct (cryptic), species (Hogg et al., 1998; Witt
*Corresponding author. E-mail: [email protected]†Present address: New Zealand Department of Conservation,Private Bag 701, Hokitika, New Zealand.
Biological Journal of the Linnean Society, 2010, 99, 196–205. With 5 figures
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205196

& Hebert, 2000; Witt, Threloff & Hebert, 2006). Manyamphipod species are also known to display pre-copulatory mate guarding, whereby males guardfemales by remaining in close proximity, or physicallyattaching themselves, to females (Conlan, 1991). Thisbehaviour has been previously used for investigatingmate selection (Dick & Elwood, 1992; Jormalainen,1998; Sutherland, Hogg & Waas, 2007) and could beused as a model for investigations of cryptic species,as well as factors that may promote behavioural andgenetic differentiation in the absence of morphologicaldifferences.
The amphipod Paracalliope fluviatilis (FamilyParacalliopiidae) is endemic to New Zealand whereit is the most abundant and widespread freshwateramphipod species. It is commonly found amongstmacrophytes in the slow-flowing margins of riversand on the edges of lakes. It appears morphologi-cally uniform throughout its range but has mito-chondrial sequence divergences as high as 24%,suggesting the presence of one or more crypticspecies (Hogg et al., 2006). Congruent patterns ofdivergence have also been observed with nuclear(allozyme) markers (Hogg et al., 2006). Hence, popu-lations can be identified on the basis of mitochon-drial DNA data alone. Furthermore, the species isthe only New Zealand freshwater amphipod todisplay pre-copulatory (physical attachment) mateguarding (Chapman & Lewis, 1976).
To assess mate recognition in P. fluviatilis and howit relates to genetic divergence, we first assessedphylogeographic patterns to determine existing levelsof genetic divergence among populations and thetiming of divergence events. Second, we conductedlaboratory mate choice experiments to determinewhether males would discriminate againstgenetically-divergent females and, if so, at what levelof divergence discrimination would occur.
MATERIAL AND METHODSFIELD SAMPLING
Between July 2000 and April 2003, we sampled 419sites in a variety of freshwater habitats throughoutNew Zealand and, of these, P. fluviatilis were found at31 sites (Fig. 1). Animals were collected using 1-mmmesh size sieves, small hand nets or large nets withlong handled poles. Samples were live sorted on sitein trays, and specimens preserved in either 95%ethanol or in liquid nitrogen. Laboratory samplespreserved in ethanol were kept at -20 °C, whereas allothers were kept at -76 °C until needed for DNAanalysis. Samples were also obtained from GenBank(accession numbers DQ285299–DQ285343; Hogget al., 2006).
DNA SEQUENCE ANALYSIS
On the basis of previous data showing limited varia-tion within sites (Hogg et al., 2006), DNA wasextracted from one or two individuals per site usingthe DNeasy Tissue kit (Qiagen Inc.). A 710-bp frag-ment of the mitochondrial gene cytochrome c oxidaseI (COI) gene was amplified using the universalprimers LCO1490 and HCO2198 (Folmer et al., 1994).The forward direction was always sequenced,whereas the reverse was also sequenced in approxi-mately 10% of samples to verify sequence accuracy.Further details on polymerase chain reaction ampli-fication and thermocycling conditions are provided byHogg et al. (2006).
Sequences were aligned using SEQUENCHER,version 4.1.2 for Macintosh (Gene Codes) sequenceeditor and verified as being derived from amphipodDNA using the GenBank BLAST algorithm. A Manteltest was performed on pairwise sequence divergence
0 200 km
N
A
B
E
D
C
North Island
South Island
1
3
6
5
4
29
810
11 7
12
13
14
15
17
18
16
19
20
30
31
22
25
2421
23
26
28
29
27
Hamilton
Waihou
Napier
Wellington
Christchurch
Invercargill
Pirongia
Figure 1. Map of New Zealand showing collection sites.Sites with squares and place names indicate populationsused in the mate choice experiment; numbers next tocollection sites and letters identifying grouped populations(A, B, C, D, E), correspond to those used in the MaximumLikelihood Tree (Fig. 3).
MATE DISCRIMINATION IN P. FLUVIATILIS 197
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

and geographic distances using GENALEX, version 5(Peakall & Smouse, 2001) to determine whether therewas a significant relationship between genetic versusgeographic distance. The degree of mutational satu-ration was estimated by examining the correlationbetween the ts/tv ratio and the pairwise sequencedivergence. We used c2 tests, as implemented inPAUP* 4.0b10 (Swofford, 2002), to determine whetherthe assumption of equal base frequencies amongsequences was violated on: (1) all sites; (2) parsimony-informative sites only; and (3) the third codon posi-tion only. We then constructed phylogenetic treesusing PAUP* 4.0b10 (Swofford, 2002). A Neighbour-joining (NJ) phylogram was constructed using theTrN+G model (selected using MODELTEST, version3.7; Posada & Crandall, 1998). The estimated param-eters under this model were: ts/tv ratio = 5.056; andgamma distribution shape parameter = 0.1972. Amaximum likelihood (ML) search was conductedusing the above model parameters. A maximum par-simony (MP) analysis using the heuristic searchoption with unweighted characters was also imple-mented. Clades were selected on the basis of diver-gence values > 11%, following an analysis of 13320invertebrate congeneric species pairs found to havemean COI divergence values of 11% (Hebert, Ratnas-ingham & deWaard, 2003). Two amphipod sequenceswere obtained from GenBank for use as outgroups.These were Epimeria georgiana and Eusirus perden-tatus, and were selected on the basis of an analysis(MP tree, data not shown) of a range of amphipodspecies suggesting that, of amphipod sequences heldby GenBank, they were the most closely related to P.fluviatilis. We used three tree construction methodsaiming to minimize the potential for error that mayarise from assumptions inherent in phylogeneticreconstruction methods. Confidence in the cladisticanalyses was assessed by estimation of the g1 skew-ness statistic from 100 000 random tree length dis-tributions (Hillis & Huelsenbeck, 1992) and bybootstrap analysis with 1000 pseudoreplicates for theNJ tree (Felsenstein, 1976). Node support for the NJand ML trees was assessed using decay indices cal-culated in PAUP* 4.0b10 using command lines com-puted in MACCLADE, version 4.03 (Maddison &Maddison, 2004). A Templeton (Wilcoxon signedranks) test was used to determine whether significantdifferences existed between the trees. To examinewhether lineages within trees were evolving at asimilar rate, a two-cluster test was employed(Takezaki, Rzhetsky & Nei, 1995). To test whethersequences were diverging in a clock-like manner, alog-likelihood ratio test was carried out in PAUP*4.0b10 that compared ML trees generated with themolecular clock option enforced with unconstrainedtrees (Felsenstein, 1988). Divergence times were then
estimated by using a molecular clock approximationfor COI of 1.4–2.6% nucleotide sequence divergenceper million years (Knowlton, 1993; Knowlton &Weight, 1998).
MATE CHOICE EXPERIMENTS
We collected live P. fluviatilis from seven locationsthroughout New Zealand: (1) Hamilton (37.80°S,175.30°E); (2) Pirongia (37.82°S, 174.90°E); (3) TeAroha (37.55°S, 175.71°E); (4) Napier (39.39°S,176.87°E); (5) Wellington (41.12°S, 175.06°E); (6)Christchurch (43.69°S, 172.57°E); and (7) Invercargill(46.39°S, 168.38°E) (Fig. 1). Animals from each popu-lation were kept in separate 2-L holding tanks con-taining macrophytes (Elodea canadensis) to enrichthe environment and to provide oxygen. Fish foodflakes (Nutrafin Staplefood) were ground to a pasteand pipetted to supplement grazing on epiphyticalgae. Water was changed weekly and excess foodpipetted off every second day. Containers were main-tained at 18 °C under a 15 : 9 h light/dark cycle tosimulate summer conditions to enhance pairing. Allanimals were held at least 3 days in holding contain-ers before experiments began to acclimatize animalsto laboratory conditions.
Choice tests were conducted in clear plastic con-tainers (62 ¥ 62 ¥ 110 mm high) filled with 200 mL ofwater, an eight cm piece of E. canadensis and a dropof fish food. Choice tests consisted of placing threesingle individuals, a male and two females, together.For half of the tests, a Hamilton male was presentedwith a Hamilton female and a female from one of the‘foreign’ populations, whereas, for the other tests, amale from the same foreign population was usedinstead of the Hamilton male. Visual inspection of theoverall data revealed that the patterns of male choice(with respect to whether males selected local versusforeign females) was consistent for both Hamilton andforeign males. Accordingly, the data were pooledresulting in six interpopulation comparisons and 64individual choice tests. Female size (determined usingan ocular micrometer at ¥2.5 magnification) was keptconstant (e.g. a small female was used with anothersmall female) because female size has been shown toinfluence male choice in this and other amphipodspecies (Jormalainen, 1998; Sutherland et al., 2007,respectively). Because individuals from differentpopulations were morphologically indistinguishable,females were identified in choice tests by the place-ment of a single white spot of fast drying correctionfluid (Bic Wite Out) on a unique position on each oftheir backs. Subjects were checked every 24 h for upto 8 days, by which time the majority of animals hadpaired.
When pairing was first observed (i.e. a male wasphysically attached to the female consistent with
198 D. L. SUTHERLAND ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205
mate guarding), pairs were observed for at least 30 sto ensure that the male had a maintained grasp onthe female. The time to the first detected pairing wasrecorded, as well as the population the paired femalecame from. Pairs were left undisturbed until theyseparated (usually 1–4 days in this species); the dura-tion of pairing was then recorded and the animalsplaced in 70% ethanol. At the end of the experiment,the number of eggs produced by each female wasrecorded. Chi-square tests were used to determinewhether males preferred pairing with females fromtheir own population, a different population or pairedrandomly. One-way analysis of variance was per-formed on: (1) the length of time taken for pairing tobe detected; (2) the duration of pairing (to determinewhether males would invest more time guarding localfemales); and (3) female egg number (to determinewhether post-zygotic isolating barriers could limit eggproduction).
RESULTSGENETIC DIVERSITY AND PHYLOGEOGRAPHY
A 578-bp fragment of the COI mitochondrial gene wasused to analyse 48 P. fluviatilis from 31 sites. Con-sensus sequences of individuals from the same popu-lation that shared the same genotype were made.There were 239 informative sites, 32 variable butparsimony uninformative sites and 307 constantsites; no insertions or deletions were detected. A totalof 30 different haplotypes was found with pairwisesequence differences in the range 1–23.5%. Each sitewas represented by a single haplotype. The nucleotidecomposition across all sequences was: A = 26%,T = 28%, C = 27% and G = 19%. The assumption ofhomogeneity of base frequencies was supported using
all codon positions (c2 = 77.27, d.f. = 90, P = 0.828),and using the third codon position only (c2
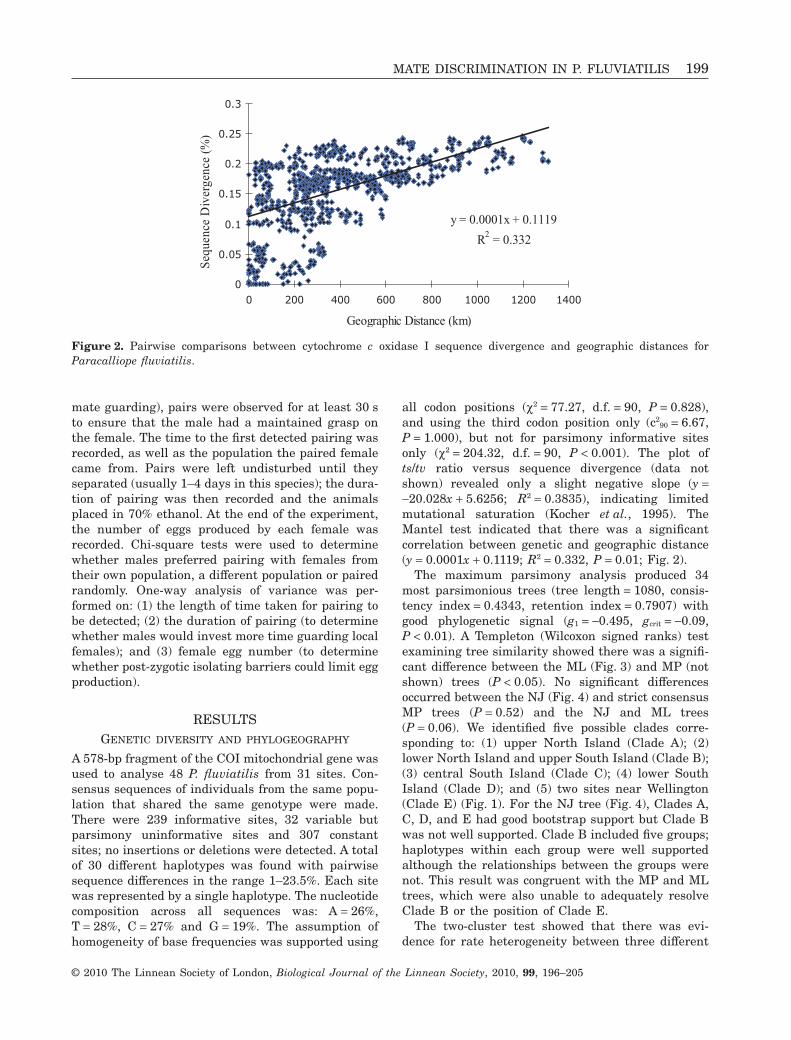
90 = 6.67,P = 1.000), but not for parsimony informative sitesonly (c2 = 204.32, d.f. = 90, P < 0.001). The plot ofts/tv ratio versus sequence divergence (data notshown) revealed only a slight negative slope (y =-20.028x + 5.6256; R2 = 0.3835), indicating limitedmutational saturation (Kocher et al., 1995). TheMantel test indicated that there was a significantcorrelation between genetic and geographic distance(y = 0.0001x + 0.1119; R2 = 0.332, P = 0.01; Fig. 2).
The maximum parsimony analysis produced 34most parsimonious trees (tree length = 1080, consis-tency index = 0.4343, retention index = 0.7907) withgood phylogenetic signal (g1 = -0.495, gcrit = -0.09,P < 0.01). A Templeton (Wilcoxon signed ranks) testexamining tree similarity showed there was a signifi-cant difference between the ML (Fig. 3) and MP (notshown) trees (P < 0.05). No significant differencesoccurred between the NJ (Fig. 4) and strict consensusMP trees (P = 0.52) and the NJ and ML trees(P = 0.06). We identified five possible clades corre-sponding to: (1) upper North Island (Clade A); (2)lower North Island and upper South Island (Clade B);(3) central South Island (Clade C); (4) lower SouthIsland (Clade D); and (5) two sites near Wellington(Clade E) (Fig. 1). For the NJ tree (Fig. 4), Clades A,C, D, and E had good bootstrap support but Clade Bwas not well supported. Clade B included five groups;haplotypes within each group were well supportedalthough the relationships between the groups werenot. This result was congruent with the MP and MLtrees, which were also unable to adequately resolveClade B or the position of Clade E.
The two-cluster test showed that there was evi-dence for rate heterogeneity between three different
y = 0.0001x + 0.1119
R2 = 0.332
0
0.05
0.1
0.15
0.2
0.25
0.3
0 200 400 600 800 1000 1200 1400
Geographic Distance (km)
Seq
uenc
e D
iver
genc
e (%
)
Figure 2. Pairwise comparisons between cytochrome c oxidase I sequence divergence and geographic distances forParacalliope fluviatilis.
MATE DISCRIMINATION IN P. FLUVIATILIS 199
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

clades (P < 0.05) indicating differential rates of evolu-tion. These differences were between: (1) Oamaru–Invercargill (sites 28 and 29) versus Christchurch–Lake Waihola (sites 26 and 27); (2) Clade C versusClades A and B; and (3) the Mangamuka–HokiangaHarbour clade (sites 4 and 5) versus the Pirongia,Hamilton, and Mangaonua Stream clade (sites 1–3)(Figs 1, 3). A log-likelihood ratio test supported thehypothesis that lineages were evolving according to aclock-like model of evolution (–ln L = 5161.34 withoutmolecular clock enforced and -ln L = 5170.33 withmolecular clock enforced [difference = 8.99]; c2 =15.655, d.f = 31, P > 0.99). Using a 2% midpoint for amolecular clock rate of 1.4–2.6% nucleotide sequencedivergence per million years (Knowlton, 1993; Knowl-ton & Weight, 1998), Clade E diverged approximately
13 Mya and Clades C and D diverged approximately12 Mya during the Miocene. The differences betweenClades A and B were smaller and occurred approxi-mately 7–9 Mya, during the late Miocene. Within-clade ranges for Clades A and B were between 0 and6 Mya (i.e. from the Pliocene onwards).
MATE CHOICE EXPERIMENTS
Male preference for females is shown in Table 1. Forthe first three population comparisons within 243 km(Pirongia, Waihou, Napier), males showed no prefer-ence for either local or foreign females (Table 1).However, for the remaining three populationcomparisons > 416 km (Wellington, Christchurch,Invercargill), males were more likely to choose local
1. Hamilton 2. Mangaonua 3. Pirongia4. Mangamuka 5. Hokianga Harbour6. Port Waikato 7. Lake Waikare 8. Waikato River9. Kawhia 10. Waitoa 11. Waihou12. Kaihere 13. Whangarai 14. Palmerston North15. Konini 16. Hawera 17.Wanganui 18. Otaki 19. Fox 20. Okato 21. Granity 22. Greymouth 23. Kaikoura 24. Blenheim 25. Napier 26. Christchurch 27. Lake Waihola28. Oamaru 29. Invercargill 30. Hutt River31. Wellington Eusirus perdentatus Epimeria georgiana
0.05 substitutions/site
A
B
C
D
E
98
78
98
90
98
99
96
100
76
78
94
60
99
97
78
88 54
95
51
10
3
2
320
10
10
11
15
17
162
10
5
69
18
11
2
2
Figure 3. Maximum likelihood tree for Paracalliope fluviatilis showing sites used in the mate choice experimenthighlighted in bold with bootstrap values (> 50) above nodes and decay indices (> 0) below nodes. Five clades(A, B, C, D, E) are highlighted on the tree based on 11% sequence divergence (Hebert et al., 2003).
200 D. L. SUTHERLAND ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

1. Hamilton2. Mangaonua
3. Pirongia
4. Mangamuka 5. Hokianga Harbour
6. Port Waikato 7. Lake Waikare
8. Waikato River
9. Kawhia10. Waitoa
11. Waihou
12. Kaihere13. Whangarai
14. Palmerston North15. Konini16. Hawera20. Okato
17. Wanganui19. Fox
18. Otaki 25. Napier
21. Granity 22. Greymouth
23. Kaikoura24. Blenheim
26. Christchurch
27. Lake Waihola 28. Oamaru 29. Invercargill
30. Hutt River31. Wellington
Epimeria georgiana Eusirus perdentatus
0.01 substitutions/site
74
98
100 100
100
100 60
69
98
99
56
99 100
100
97
100
82 86
52
85
100
100
100
100
100
A
D
E
B
C
16
2
11
10
18
2
17
10
11
15
2
10
96
3
5
2
20
3
10
Figure 4. Neighbour-joining tree for Paracalliope fluviatilis with bootstrap values (> 50) above nodes and decay indices(> 0) below nodes. Five putative clades (A, B, C, D, E) are highlighted on the tree.
Table 1. The ratio of local : foreign pairs formed from six intra- and inter-population crosses with significance values andtheir geographic distances (km) and pairwise genetic distances (uncorrected P)
Hamilton Local : foreign c2 PGeographicdistance
Geneticdivergence
Pirongia 25 : 24 (1.04 : 1) 0.083 0.773 15 0.019Waihou 25 : 23 (1.09 : 1) 0.191 0.662 25 0.140Napier 22 : 20 (1.10 : 1) 0.095 0.758 243 0.195Wellington 34 : 7 (4.86 : 1) 17.780 < 0.001 416 0.215Christchurch 20 : 4 (5.00 : 1) 10.667 0.001 689 0.235Invercargill 26 : 2 (13.00 : 1) 20.571 < 0.001 1120 0.215
MATE DISCRIMINATION IN P. FLUVIATILIS 201
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

females (P < 0.01 in all cases). There was no differ-ence between the six populations in the time taken toform pairs (P = 0.09–0.90) or in how long pairinglasted (P = 0.30–0.77; Table 2). There was also nodifference in mean egg number per female or theproportion of females producing eggs when thenumber of pairings was taken into account (P = 0.22–0.87; Table 2). However, there was a significant dif-ference for total number of females producing eggswhen the Hamilton population was crossed with Well-ington (local : foreign 11 : 2; F = 8.63, P = 0.005),Christchurch (6 : 0; F = 4.14, P = 0.046), and Inver-cargill populations (5 : 0; F = 5.74, P = 0.020; Table 2).
DISCUSSIONGENETIC DIVERSITY AND PHYLOGEOGRAPHY
There were considerable genetic divergences amongpopulations of P. fluviatilis with values ranging upto 23.5% suggesting that cryptic species may exist(Hebert et al., 2003). The greatest sequence diver-gences occurred between some of the South Islandpopulations (e.g. Invercargill and Lake Waihola;Fig. 1), whereas generally smaller differences occurredbetween North Island populations. Despite the largegenetic divergences among some lineages, no morpho-logical differences have yet been found (Hogg et al.,2006; M. A. Chapman, unpubl. data). This pattern ofmorphological conservatism coupled with large geneticdivergences has been previously reported among othercrustaceans (Lee, 2000; Witt & Hebert, 2000; Page &Hughes, 2007; Thum & Harrison, 2009).
The genus Paracalliope was likely introduced toNew Zealand waters from Australia via the Tasmancurrent (Barnard, 1972). This current has beenpresent subsequent to the Miocene (Field et al., 2002)and is considered to be responsible for the dispersal ofother aquatic species into New Zealand (e.g. mudfishgenus Neochanna: Waters & White, 1997; the galaxiidfish, Galaxias maculatus: Waters, Dijkstra & Wallis,2000). On the basis of the divergences that wedetected in P. fluviatilis (� 23.5%), dispersal to NewZealand may have occurred during the Miocene (upto 17 Mya; Fig. 5A). The ancestral form may haveundergone an estuarine phase before becoming fullyadapted to the freshwater environment. Geneticdivergences among populations would then haveresulted from a reduction in dispersal between rivercatchments as populations became more restricted tofreshwater habitats.
The beginning of the Pliocene (5.2 Mya) was char-acterized by rising sea levels that resulted in marineinundation in New Zealand creating two majorand several minor islands (Stevens & Ridge, 1995;Fig. 5B). Patterns of genetic divergence seen in ortho-pterans (e.g. Hemideina thoracica; Morgan-Richards,Trewick & Wallis, 2001), as well as speciation eventsin beetles (e.g. Lissotes; Fleming, 1979), have beenattributed to the resulting fragmentation of the NewZealand landmass. The divisions occurring withinand between Clades A and B suggest that individualsmay have become isolated on different islands duringthe Pliocene (Fig. 5B). At this time, individuals froman ancestral population on the east coast of the NorthIsland could have dispersed via prevailing ocean cur-rents (Fig. 5B) and colonized the upper South Islandand newly available habitat on the lower NorthIsland.
Lower sea levels during the Pleistocene (Fig. 5C)resulted in continuous land and would have allowedthe re-colonization of previously inundated habitat(e.g. sites 14–19; Figs 1, 3), and also prevented dis-persal between some sites (e.g. sites 21 and 22 andsites 23 and 24). Furthermore, it appears that indi-viduals from within Clade A (e.g. sites 1–9; Fig. 3)may have dispersed southward from the northernpart of the North Island to at least the central NorthIsland. Dispersal would have been further facilitatedby the Taupo eruption (approximately 27 000 yearsago), which changed the course of New Zealand’slongest river (the Waikato) and its associated tribu-taries from east to west flowing (Wilson & Walker,1985). Upstream dispersal into previously unavail-able habitat would then have resulted in the present-day east and west branches within Clade A (Fig. 3)(Hogg et al., 2006). This suggests that P. fluviatiliswere capable of at least some dispersal via coastalcurrents but not overland (e.g. via bird-mediated
Table 2. Comparisons of time (days) taken to pair (TTP),time paired (TP), the number of females who producedeggs, and the mean egg number per female for six intra-and inter-population crosses
HamiltonLocal :foreign TTP TP
Femaleswitheggs
Meaneggs(N)
Pirongia 25 1.84 1.84 4 3.7524 1.88 2.36 6 4.17
Waihou 25 1.68 1.00 7 3.8623 2.17 1.04 6 3.17
Napier 20 2.27 1.64 3 3.3320 1.50 1.50 3 2.00
Wellington 34 2.41 2.11 11 3.367 2.29 1.63 2 3.50
Christchurch 20 2.60 2.25 6 4.834 3.25 1.00 0 NA
Invercargill 26 4.08 1.75 5 3.202 5.00 1.00 0 NA
NA, not applicable.
202 D. L. SUTHERLAND ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

transport). However, the present-day Cook Strait(Fig. 5D) appears to be a significant barrier to dis-persal between the North and South Island, main-taining observed divergences between clades as wellas within clades (e.g. sites 21 and 22 versus 23 and24) (Figs 1, 3, 4).
MATE CHOICE EXPERIMENTS
There was evidence for nonrandom (assortative)mating between the Hamilton population and thethree populations that were the most geneticallydivergent (i.e. Wellington, Christchurch, and Inver-cargill). This finding, in conjunction with large geneticdivergences (� 21.5%; Table 1) and fewer or no eggsbeing produced from foreign/local pairings, suggeststhat individuals from the three populations areindeed a separate species from the Hamilton popula-tion. During Hamilton/Napier tests, 19.5% geneticdivergence, males showed no preference for localversus foreign females. However, discriminationoccurred between Hamilton and Wellington individu-als at 21.5% divergence. Because these levels weresimilar, we suggest that ancestry (i.e. being in differ-ent clades) and not divergence per se may explain thepatterns of discrimination that we observed. Withinclades, individuals may lack the ability to discrimi-nate sister taxa even when genetic divergences arehigh.
No obvious behavioural differences (e.g. orientation,postures, mating sequences) were observed amongthe genetically divergent populations of P. fluviatilis.When no obvious visual cues are available to distin-guish potential mates, pheromones may be used; con-specific recognition via these cues is common withinmany taxa (Colgan, 1983; Johnson, 2000; Wrens,
2000). The rapid evolution of pheromones (e.g. chang-ing ratios of chemical components, emission rates)could therefore facilitate the sudden shifts that weobserved in mate discrimination (Andersson, 1994).For example, in some amphipod species, femalesrelease hormones when their exoskeleton is shed; thiscan attract males because it indicates that the femaleis ready to be fertilized (Jormalainen, 1998). Wehypothesize that differences in pre-moulting hor-mones are used to identify conspecific mates (sensuCorotto et al., 1999). Although a strong bias in mateselection existed for the three most divergent crosses,some local : foreign pairings resulted in eggs beingproduced; hence, it is possible that viable hybridscould form between the putative sibling species.This could be particularly problematic for allopatricspecies when isolating barriers are removed becauselower levels of pre-zygotic reinforcement may exist(Coyne & Orr, 1997).
The large genetic divergences that we found in thepresent study suggest that P. fluviatilis is a speciescomplex. Significant discrimination was only appar-ent when genetic divergences were � 21.5%, which isalmost double the mean divergence of 11.3% for mostcongeneric pairs found by Hebert et al. (2003). Thissuggests that P. fluviatilis, and perhaps species in theAmphipoda in general, may be more genetically diver-gent relative to other animal taxa. There appears tobe limited gene flow among river catchments, sug-gesting that geographically isolated populations maybe undergoing speciation. Mate choice tests suggestthat discrimination is occurring among the mostdivergent populations. However, the changes are notgradual but abrupt, perhaps reflecting the ability ofindividuals to discriminate mates of disparate ances-try (i.e. between clades). Further work examining
1.7 m.y.a.PleistocenePliocene
5.2 m.y.a.Miocene12 m.y.a.
Present
A B C D
Figure 5. Time series showing the New Zealand land mass from the Upper Miocene (A) to the present (D) and showingland above sea-level (dark grey) superimposed on the current New Zealand coastline. Arrows indicate prevailing currents.Adapted with permission from Fleming (1979).
MATE DISCRIMINATION IN P. FLUVIATILIS 203
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

mate discrimination between other divergent (e.g.South Island) clades is needed to further test thishypothesis. We conclude that a combination ofmolecular, behavioural and other techniques (e.g. bio-chemical or neurosensory) may be required to accu-rately identify species which show no distinctivemorphological differences.
ACKNOWLEDGEMENTS
We are grateful to anonymous reviewers for theirvaluable comments on the manuscript. We also thankPeter Sutherland, Robert Torrens, Mike Winterbourn,Leanne O’Brien, and Paul Leisnham for providingsamples; Mark Stevens for advice on the geneticanalysis; and Graham Wallis, Pete Convey, andNaomi Crawford for their comments on an earlierversion of the manuscript. Members of the Centre forBiodiversity and Ecology Research group providedhelpful discussion. I.H. received financial supportfrom the New Zealand Foundation for ResearchScience and Technology (UOWX0505).
REFERENCES
Adamowicz SJ, Menu-Marque S, Hebert PDN, Purvis A.2007. Molecular systematics and patterns of morphologicalevolution in the Centropagidae (Copepoda: Calanoida) ofArgentina. Biological Journal of the Linnean Society 90:279–292.
Andersson M. 1994. Sexual selection. Princeton, NJ: Princ-eton University Press.
Barnard JL. 1972. Gammaridean Amphipoda of Australia,Part 1. Smithsonian Contributions in Zoology 103: 1–333.
Chapman MA, Lewis MH. 1976. An introduction to thefreshwater Crustacea of New Zealand. Auckland: Collins.
Colgan P. 1983. Comparative social recognition. New York,NY: Wiley-Interscience Publication.
Conlan KE. 1991. Pre-copulatory mating behaviour andsexual dimorphism in the amphipod Crustacea. Hydrobio-logia 223: 255–282.
Cook BD, Bunn SE, Hughes JM. 2007. A comparativeanalysis of population structuring and genetic diversity insympatric lineages of freshwater shrimp (Atyidae: Paratya):concerted or independent responses to hydrographic factors?Freshwater Biology 52: 2156–2171.
Corotto FS, Bonenberger DM, Bounkeo JM, Dukas CC.1999. Antennule ablation, sex discrimination, and matingbehavior in the crayfish Procambarus clarkii. Journal ofCrustacean Biology 19: 708–712.
Coyne JA, Orr HA. 1997. ‘Patterns of speciation in Droso-phila’ revisited. Evolution 51: 295–303.
Dick JTA, Elwood RW. 1992. Coexistence and exclusionamong Gammarus species: behavioural avoidance of inter-specific precopulation by male G. pulex (Amphipoda). Oikos64: 541–547.
Felsenstein J. 1976. The theoretical population genetics ofvariable selection and migration. Annual Review of Genetics10: 253–280.
Felsenstein J. 1988. Phylogenies from molecular sequences:inference and reliability. Annual Review of Genetics 22:521–565.
Field BD, Crundwell MP, Kennett JP, King PR, JonesCM, Scott GH. 2002. The early Middle Miocene paleoen-vironmental setting of New Zealand. Revista Mexicana deCiencias Geologica 3: 242–251.
Fleming CA. 1979. The geological history of New Zealandand its life. Auckland: Auckland University Press.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. 1994.DNA primers for the amplification of mitochondrial cyto-chrome c oxidase subunit 1 from diverse metazoan inverte-brates. Molecular Marine Biology and Biotechnology 3: 294–299.
Hebert PDN, Ratnasingham S, deWaard JR. 2003. Bar-coding animal life: cytochrome c oxidase subunit 1 diver-gences among closely related species. Proceedings of theRoyal Society of London Series B, Biological Sciences 270:S96–S99.
Hillis DM, Huelsenbeck JP. 1992. Signal, noise, and reli-ability in molecular phylogenetic analyses. Journal ofHeredity 83: 189–195.
Hogg ID, Larose C, Lafontaine Y, Doe KG. 1998. Geneticevidence for a Hyalella species complex within the GreatLakes–St Lawrence River: implications for ecotoxicologyand conservation biology. Canadian Journal of Zoology 76:1134–1140.
Hogg ID, Stevens MI, Schnabel KE, Chapman MA. 2006.Deeply divergent lineages among the widespread NewZealand amphipod Paracalliope fluviatilis revealed usingallozyme and mitochondrial DNA analyses. FreshwaterBiology 51: 236–248.
Johnson KP. 2000. The evolution of courtship displayrepertoire size in the dabbling ducks (Anatini). Journal ofEvolutionary Biology 13: 634–644.
Jormalainen V. 1998. Pre-copulatory mate guarding in crus-taceans, male competitive strategy and intersexual conflict.Quarterly Review of Biology 73: 275–304.
Knowlton N. 1993. Sibling species in the sea. Annual Reviewof Ecology and Systematics 24: 189–216.
Knowlton N, Weight LA. 1998. New dates and new rates fordivergence across the Isthmus of Panama. Proceedings ofthe Royal Society of London Series B, Biological Sciences265: 2257–2263.
Kocher TD, Conroy JA, McKaye KR, Stauffer JR, Lock-wood SF. 1995. Evolution of NADH dehydrogenase subunit2 in East African cichlid fish. Molecular Phylogenetics andEvolution 4: 420–432.
Lee CE. 2000. Global phylogeography of a cryptic copepodspecies complex and reproductive isolation betweengenetically proximate ‘populations’. Evolution 54: 2014–2027.
Maddison DR, Maddison WP. 2004. MacClade 4: analysisof phylogeny and character evolution, Release 4.03. Sunder-land, MA: Sinauer Associates .
204 D. L. SUTHERLAND ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205

Mayr E. 1982. The growth of biological thought. Cambridge,MA: Harvard University Press.
Morgan-Richards M, Trewick SA, Wallis GP. 2001. Chro-mosome races with Pliocene origins: evidence from mtDNA.Heredity 86: 303–312.
Page TJ, Hughes JM. 2007. Radically different scales ofphylogeographic structuring within cryptic species of fresh-water shrimp (Atyidae: Caridina). Limnology and Oceanog-raphy 52: 1055–1066.
Peakall R, Smouse PE. 2001. GenAlEx V5: genetic analysisin excel. Population genetic software for teaching andresearch. Australian National University, Canberra, Austra-lia. Available at: http://www.anu.edu.au/BoZo/GenA1Ex/.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14: 817–818.
Ruppert EE, Barnes RD. 1994. Invertebrate zoology, 6thedn. Philadelphia, PA: Saunders College Publishing.
Stevens G, Ridge P. 1995. Prehistoric New Zealand.Auckland: Reed Books.
Sutherland DL, Hogg ID, Waas JR. 2007. Mate choice,intraspecific competition and conflict in Paracalliopefluviatilis (Crustacea; Amphipoda). Biological Journal of theLinnean Society 92: 173–181.
Swofford DL. 2002. PAUP*. Phylogenetic analysis usingparsimony, version 4.0b10 for Macintosh. Sunderland MA:Sinauer Associates Inc.
Takezaki N, Rzhetsky A, Nei M. 1995. Phylogenetic test ofthe molecular clock and linearized trees. Molecular Biologyand Evolution 12: 823–833.
Thum RA, Harrison RG. 2009. Deep genetic divergencesamong morphologically similar and parapatric Skistodiap-tomus (Copepoda: Calanoida: Diaptomidae) challenge thehypothesis of Pleistocene speciation. Biological Journal ofthe Linnean Society 96: 150–165.
Waters JM, Dijkstra LH, Wallis GP. 2000. Biogeo-graphy of a southern hemisphere freshwater fish: Howimportant is marine dispersal? Molecular Ecology 9: 1815–1821.
Waters JM, White RWG. 1997. Molecular phylogeny andbiogeography of the Tasmanian and New Zealand mudfishes(Salmoniformes: Galaxiidae). Australian Journal of Zoology45: 39–48.
Wilson CS, Walker GPL. 1985. The Taupo eruption,New Zealand. I. General aspects. Philosophical Trans-actions of the Royal Society of London Series A 314: 199–228.
Witt JDS, Hebert PDN. 2000. Cryptic species diversity andevolution in the amphipod genus Hyalella within centralglaciated North America: a molecular phylogeneticapproach. Canadian Journal of Fisheries and Aquatic Sci-ences 57: 687–698.
Witt JDS, Threloff DL, Hebert PDN. 2006. DNA barcodingreveals extraordinary cryptic diversity in an amphipodgenus: implications for desert spring conservation. Molecu-lar Ecology 15: 3073–3082.
Wrens JJ. 2000. Decoupled evolution of display morphologyand behaviour in pharynosomatid lizards. BiologicalJournal of the Linnean Society 70: 597–612.
MATE DISCRIMINATION IN P. FLUVIATILIS 205
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 99, 196–205




















