
Remaining tree species from the indigenous vegetation of Ankara, Turkey
Upload
vogelbeschermingCategory
view
3download
0
- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 155
Applied Vegetation Science 8: 155-166, 2005© IAVS; Opulus Press Uppsala.
Effects of tree species composition on within-forest distribution ofunderstorey species
van Oijen, Danielle1*; Feijen, Markus1; Hommel, Patrick2; den Ouden, Jan1 & de Waal, Rein2
1Forest Ecology and Forest Management Group, Wageningen University and Research Centre, PO Box 342,6700 AH Wageningen, The Netherlands; 2Alterra, Wageningen University and Research Centre, PO Box 47,
6700 AA Wageningen, The Netherlands; *Corresponding author; E-mail [email protected]
AbstractQuestion: Do tree species, with different litter qualities,affect the within-forest distribution of forest understoreyspecies on intermediate to base-rich soils? Since habitat lossand fragmentation have caused ancient forest species todecline, those species are the main focus of this study.Location: Three ancient forests, along a soil gradient fromacidification-sensitive to base-rich, were studied: Limbrich-terbosch and Savelsbos in The Netherlands and Holtkrat inDenmark.Methods: Canopy and soil surveys along transects generateddata for Redundancy Analysis on tree - humus relationships.We analysed the distribution of forest plant species withCanonical Correspondence Analysis. The explanatory fac-tors were soil characteristics (pH, organic matter, loam con-tent and thickness of the humus layers), external crownprojection, groundwater and canopy data. We further ana-lysed the relationship between forest species and humuscharacteristics with Spearman correlations.Results: Tree species have a significant impact on humuscharacteristics through the nature of their litter. Humus char-acteristics significantly explain the distribution of forestunderstorey species. The pH of the first 25 cm mineral soiland the thickness of the F- (fermentation) layer are theprimary factors affecting the distribution of ancient forestspecies.Conclusion: This study indicates that the species composi-tion of the forest canopy affects the distribution of forestunderstorey species. Ancient forest species are more abun-dant and frequent underneath trees with base-rich litter. Onacidification-sensitive soils these relationships were strongerthan on more base-rich, loamy soils.
Keywords: Ancient forest species; Conservation; Fraxinus;Humus profile; Multi-variate analysis; Tilia; Quercus.
Abbreviations: Ah = Soil horizon consisting of mineral soilwith a high organic matter content; CEC = Cation exchangecapacity; F = Fermenting litter (layer); Ah = Humus-richmineral soil (layer); L = Litter (layer); RDA = Redundancyanalysis.
Introduction
Ancient forest species are species that are mostlyconfined to ‘ancient forests’ (Rackham 1980; Peterken1993), which have existed for a long period of time,since a certain threshold date which varies from coun-try to country (e.g. at least since 1600 in UK (Peterken1974), 1800 in Denmark (Petersen 1994) and 1850 inThe Netherlands (Grashof-Bokdam 1997; see over-view in Hermy et al. 1999). The term ‘ancient’ onlyrefers to the continuity as a forest and not to the age ofthe trees or to the naturalness of the stands (Wulf 1997).These forests are characterized by relatively undis-turbed soils, not altered by agriculture or other long-lasting soil perturbations (Koerner et al. 1997; Verheyenet al. 1999; Harmer et al. 2001). Habitat loss andfragmentation throughout history have caused ancientforest species to decline (Peterken 1977; Rackham 1980;Hermy et al. 1999). In recent decades forest manage-ment has taken a more ecological approach and strivesfor a higher diversity and naturalness in forests. Despitethese efforts the biodiversity of most broad-leaved for-ests in northwest European countries is still decreasing(Dirkse & Martakis 1998; Brunet et al. 2000; Peterken1993). Ecological knowledge of ancient forest speciesand their environment contributes to diversity conser-vation goals.
Lists of ancient forest species are regional becausethe affinity to ancient forests depends on plant geogra-phy and forest history. Within its natural distributionarea a certain species might be restricted to forests inone region while elsewhere it is a typical grasslandspecies (Hermy et al. 1999; Rackham 1980). Dispersallimitation is often seen as a very important, if not themost determining, factor in the distribution, coloniza-tion and establishment of ancient forest species (Grashof-Bokdam 1997; Bossuyt et al. 1999; Ehrlén & Eriksson2000; Jacquemyn et al. 2001; Graae et al. 2003). Otherauthors found that reduced establishment of ancientforest species is related to habitat quality such as base

156 VAN OIJEN, D. ET AL.
saturation, humus type, light climate and phosphatecontent (Koerner et al. 1997; Hermy et al. 1999; Honnayet al. 1999; Bijlsma et al. 2001; Hommel & de Waal2003b; De Keersmaeker et al. 2004). Verheyen et al.(2003) suggested that diaspore availability initially de-termines the colonization pattern of forest herbs infragmented landscapes whereas environmental sortingfollows at later stages.
The humus profile creates the chemical conditions(e.g. pH and nutrient availability) and the physicalconditions (e.g. water availability, obstructive litterlayer and rooting horizons) for forest plants. The hu-mus profile is the result of a broad range of biotic andabiotic site factors and can be seen as an indication offorest history and a source of information for futuredevelopment. Although development of the humus pro-file depends on parent material and other primary sitefactors, vegetation is the source of humus material(Jenny 1941).
The impact of trees on soil properties is tree speciesspecific (Whitney & Foster 1988; Nordén 1994 a-c).Trees influence soil properties through the quality oftheir litter as determined by nutrient contents, CEC(Cation exchange capacity) and base saturation. As aresult, different humus types (usually classified as mor,moder and mull, see Klinka et al. 1981) can developunderneath different tree species on the same soils(Birks 1986; Persson et al. 1987; Pigott 1989; Maddeleinet al. 1994; Neirynck et al. 2000). On acidification-sensitive soils in particular tree species with base-richlitter might direct the humus development towards mulltypes whereas a higher abundance of tree species withmore acid litter results in ectorganic humus types. Acidi-fication-sensitive soils are, for example, old river clays,sandy loess and Pleistocene sands with more than 15%loam. Their pH-KCL values are in a range of 3.5 - 4.0and their buffering capacity is intermediate (Hommel& de Waal 2003 a).
The impact of trees on soil and vegetation are mani-fold. Stem flow and throughfall of nutrients affectnutrient availability (Nordén 1994 a-c), large litter quan-tities and large leaf surface area inhibit sprouting ofcertain species (Sydes & Grime 1981; den Ouden &Vogels 1997) and the depth or density of tree rootsaffect below-ground competition for water and nutri-ents (Faliński & Falińska 1986). Tree species differ intheir phenology, and the quantity and quality of pen-etrating light depends on crown structure, leaf positionand texture (Rackham 1980).
Other authors have studied relationships betweentrees and soil, or soil and understorey, on several soiltypes (Nordén 1994 a,b,c; Koerner et al. 1997; Neiryncket al. 2000), but few studies include both interactions(Pigott 1989; Hommel & De Waal 2003 a). Here we
study the distribution of forest understorey species andtheir relationships with characteristics of the humusprofile and the composition of the canopy layer. Weconducted vegetation and soil surveys in three ancientforests situated on acidification-sensitive to more base-rich soils. Our main research question was whether treespecies affect the within-forest distribution of the forestunderstorey species. More specifically, we focus onancient forest species.
Methods
Study areas
We researched three ancient forests that were se-lected according to the following criteria. A wide rangeof understorey species should be present in varyingabundance and distribution. The forests should containdifferent tree species representing a range in litter qual-ity. We were particularly interested in Tilia cordata andT. platyphyllos as both species are valued for their soilimproving litter quality (Rackham 1980; Pigott 1989).The different forests should be on a gradient in parentmaterial from acidification-sensitive to base-rich. Theyshould be comparable in hydrology and climate. Theoccurrence of ancient Tilia stools, which are very rarein The Netherlands, was the deciding factor in alsoincluding a research area in Denmark despite problemscaused by geographical distance.
Limbrichterbosch and Savelsbos are both situatedin the southern part of The Netherlands (Table 1). Lim-brichterbosch was part of the ‘Graet’, a large more orless closed forest around 900 AD. It is mentioned in the13th century and on topographical maps dating from1842 AD (Schols 1986). The investigated Tilia coppicein Savelsbos has been forested for at least 200 years andprobably much longer as the steep slopes in this typicalgradient forest are not suitable for agricultural use(Schoonderwoerd & Nyssen 1999). Holtkrat is situatedin central Jutland, Denmark. In 1449 the name ‘Holt’was used and the forest was described in 1670 as acoppice (Kristiansen & Dalsgaard 2000). Climate, soiltype and vegetation are comparable to Dutch situations(Hommel & de Waal 2003 a). It must be stressed thatpopulations of ancient forest species may respond dif-ferently in the two countries.
Limbrichterbosch and Holtkrat were uniform, withinthe research area, with respect to primary site factorssuch as climate, parent material and hydrology.Savelsbos was heterogeneous in topography and parentmaterial. The hill sequence comprises heterogeneousparent material of sandy clay and gravel, loess (siltyclay loam) and marlstone. All three areas were coppice
- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 157
forests at some time in their history and for a number ofdecades have been managed for nature conservationonly. They can be arranged along a soil gradient fromacidification-sensitive (Limbrichterbosch) to morebase-rich soils (Holtkrat and parts of Savelsbos), cul-minating in calcareous soils in Savelsbos. Likewise,the vegetation follows a gradient from the Fago-Quercetum convallarietosum to the Stellario-Carpinetumoxalidetosum, typicum and allietosum (Hommel et al.1999; Stortelder et al. 1999).
Sampling
In the selected areas we positioned line transects,adjusted to height or groundwater variation accordingto maps and field visits. On these line transects square10 m × 10 m plots were marked covering the wholetransect (Table 1). In the centre, 1 m × 1 m plots werenested within the large plots, in which the abundance ofthe understorey ground flora was estimated using theBraun-Blanquet scale. Data gathered in the 10 m × 10 mplots were explanatory variables for the vegetation andcharacteristics of the humus profile of the nested plots.We used the European forest species list from Hermy etal. (1999) to select ancient forest species.
At three random points in the 1 m × 1 m plot area,the humus profile was described after Klinka et al.(1981), modified by Kemmers & de Waal (1999). Datasampling on humus profile and vegetation was con-ducted within one square metre, which benefits thestrength of relationships. Thickness and nature of hu-mus horizons were used to discern different humustypes. In this study, the ectorganic layer is divided intoL, the litter layer of fresh organic material withoutdecomposition and F, the more decomposed and frag-mented litter layer where some litter structure is stillvisible and mineral soil has not mixed. F/Ah is a transi-tion where mineral soil is mixed with F. The Ah horizonconsists of mineral soil with a high organic matter
content and the AC horizon is a transition from Ah to Clayer (parent material) (Klinka et al. 1981; Kemmers &de Waal 1999). In Savelsbos we described soil typesafter Vleeshouwer & Damoiseaux (1990) to analyse theheterogeneity in parent material and soil type.
Soil samples were taken at three depths after re-moval of the ectorganic layer, 0 - 5 cm, 5 - 25 cm and 80cm. With Merck Spezialindikator (pH 2.0-9.0) we deter-mined pH to an accuracy of 0.5 (Scheffer & Schacht-schabel 1982). Organic matter content and loam contentwere estimated (Anon. 1975) using references from digi-tized soil maps (www.bodemdata.nl) and Kristiansen &Dalsgaard (2000). We validated the estimated data byanalysing ca. five subsamples for each sampled site inthe laboratory (glow loss, texture analysis), 85% of theestimated data referred to the same soil class.Groundwater level was measured in a bore hole with ameasuring tape connected to a weight, the day afterboring. In Savelsbos, due to the hilly terrain andmarlstone parent material, we did not measuregroundwater level.
In the 10 m × 10 m plots the abundance of species inthe canopy and shrub layers were estimated using theBraun-Blanquet scale. As a measure of light availabil-ity we estimated external crown projection area using atube that restricted the angle of sight to ca. 22° toincrease the accuracy of the visual estimate. For eachplot, 20 random measurements (% cover) were takenand the mean value used for data analysis. All thevegetation data were gathered from 28.04. - 21.06. 2002,within a maximum span of three weeks per researcharea.
Data analysis
All statistical analyses were performed for eachresearch area separately. Braun-Blanquet data on treeand understorey species were converted into a 1 to 9scale (van der Maarel 1979).
Table 1. The main characteristics of the three studied ancient forests. Site age: first date at which a forest is mentioned in theliterature; No. species = total number of understorey species; No. afs = total number of ancient forest species
Area Site age Soil type/ Humus No./length (m) No. sampling No. species/(ha) (AD) Main tree species loam % types transects plots No. afs
Limbrichterbosch 91 900 Fraxinus excelsior Luvisol/ Moder 3 / 620 62 58 / 18Quercus robur 55-70 MullQuercus petraeaPopulus x canadensis
Holtkrat 10 1449 Quercus robur Sandy clay Mull 3 / 450 45 31 / 13Tilia cordata loam/18-25
Savelsbos 240 1800 Carpinus betulus Heterogenous Mull 5 / 450 45 34 / 14Quercus roburTilia cordata

158 VAN OIJEN, D. ET AL.
The relationship between tree species and humuscharacteristics was analysed with redundancy analysis(RDA) in Canoco for Windows version 4.0 (Jongmanet al. 1995; ter Braak 1996; ter Braak & Smilauer1998). All environmental factors were checked on nor-mality and transformed if necessary. Trees were di-vided into fractions indicating the quality of their litter(after Heath et al. 1966; Rackham 1980; Kuiters 1987;Maddelein et al. 1994). This resulted in a fraction oftrees with more base- and nutrient-rich litter (= fraction‘rich’), an intermediate or unknown fraction and afraction of tree species with acidifying litter (= fraction‘poor’) (Table 2).The explanatory variables in this multi-variate analysis were tree species and fractions, theresponse variables were the humus characteristics (hori-zon thickness (cm), pH and organic matter of soil sam-ples (%)).
We determined the relationship between the under-storey species and their environment with multivariateanalysis to obtain the direct ordination of species byvariance in environmental factors. Canonical Corre-spondence Analysis (CCA) provides a reliable analysisof spatial or temporal assessments of vegetation, evenif species distributions are skewed or not all of thefactors determining species composition are known(Palmer 1993). First, detrended correspondence analy-sis (DCA) of the understorey was carried out to deter-mine the variance in species data. If the resulting gradi-ent in species data was less than two standard devia-tions (SD), we used linear ordination techniques (RDA)in subsequent multivariate analysis and unimodal tech-niques (CCA) if this gradient was larger than three SD.The response variable was the abundance of allunderstorey species (including bryophytes). The ex-planatory variables were the abundance of the treespecies, tree fractions, pH, loam content and organicmatter content of soil samples at three depths, groundwater level, external crown projection and thickness ofthe humus horizons. The Monte Carlo permutation testwas used to determine the significance of the separateaxes and model. Earlier research on soil characteristicsin Holtkrat and Limbricht by Kristiansen & Dalsgaard(2000) and Hommel et al. (2002) respectively, wasused to forestall effects of covariation between treespecies and site conditions. The covariable soil typewas used to erase the variation in species explained bysoil heterogeneity in Savelsbos.
With non-parametric Spearman correlation analysisin SPSS (Anon. 2001) we determined the relationshipof separate species and their environment in addition toall species at once in multivariate analysis.
Results
Tree species - humus relationship
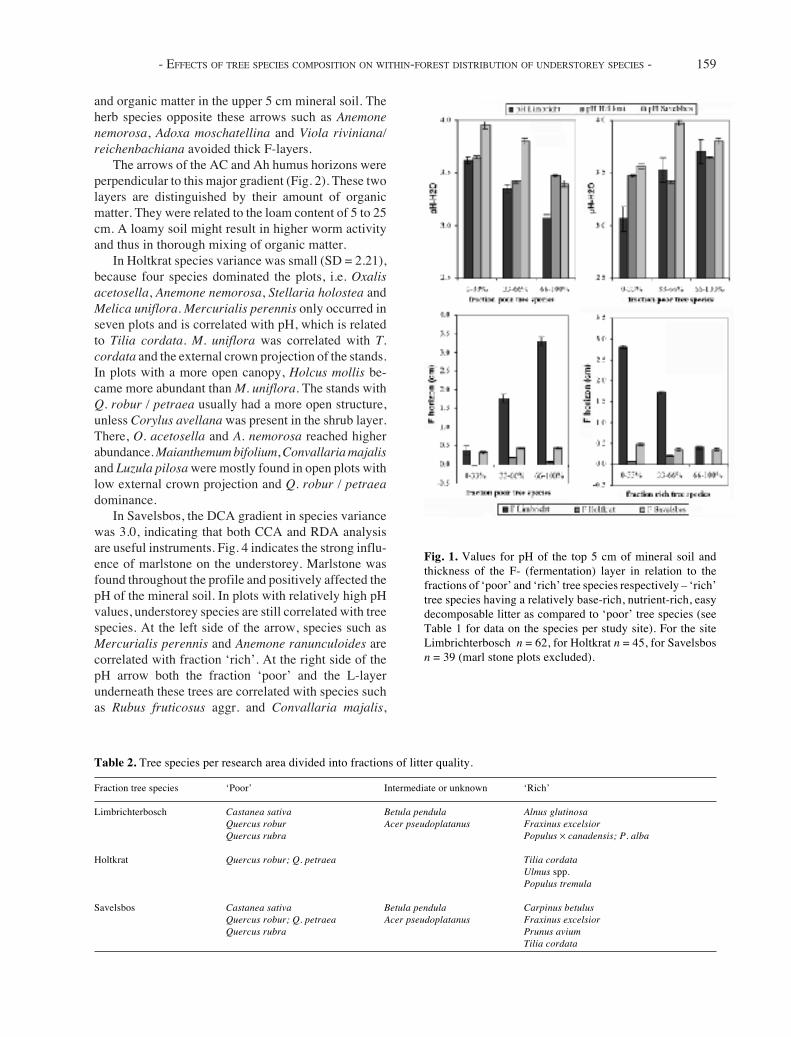
In all three areas significant relationships were foundbetween the composition of the tree canopy and charac-teristics of the humus profile. The relationship of treefractions with the pH of the first 5 cm mineral soil andF-layer are shown in Fig. 1. These relationships weremost pronounced in Limbrichterbosch, where the RDAmodel explained 30.2% (p = 0.005) of the variation inhumus characteristics. The pH of 0 to 25 cm mineralsoil was positively related to fraction ‘rich’ (Populus ×canadensis and Fraxinus excelsior) and negatively toQuercus robur / petraea (fraction ‘poor’). Vice versafor indicators of ectorganic humus profiles (F- and L-layer, organic matter content in the upper 5 cm ofmineral soil).
In Holtkrat the L- and F-layer were small or lackingand the relationships of tree species and humus charac-teristics were less pronounced (explained variance RDAmodel = 13.1%; p = 0.005). The pH was still positivelyrelated to the ‘rich’ fraction and negatively to the ‘poor’fraction (Fig. 1). The organic matter content in theupper 25 cm of mineral soil was positively related to Q.robur / petraea.
In Savelsbos the occurrence of marlstone through-out the hill sequence strongly affected pH values. Inmultivariate analysis only the fraction of ‘poor’ treespecies significantly explained humus characteristics.The thickness of the F- and L-layers showed a positiverelationship to this fraction and the pH to 25 cm anegative. The marlstone plots (mean pH 6.5) are ex-cluded from Fig. 1.
Forest understorey - environment relationship
For Limbrichterbosch, Canonical CorrespondenceAnalysis was conducted including all understoreyspecies (DCA gradient of 5.8). The pH of the first 5 cmof mineral soil explained the major species gradient(Fig. 2). The nitrophilic and forest edge species (e.g.Urtica dioica, Galium aparine) and ancient forest speciesknown to occur in base-rich systems (e.g. Circaealutetiana, ) are positioned at the leftside of the diagram in the positive direction of the 0-5pH arrow. Plagiomnium undulatum, a bryophyte, alsoindicates a base-rich environment. Populus × canadensisand fraction ‘rich’ were positively related to the pH,whereas fraction ‘poor’ was on the opposite side of thepH gradient. The tree species Quercus robur is corre-lated with species such as Lonicera periclymenum andConvallaria majalis in the diagram. Q. robur was alsorelated to two ectorganic humus characteristics: F-layer
‹
- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 159
and organic matter in the upper 5 cm mineral soil. Theherb species opposite these arrows such as Anemonenemorosa, Adoxa moschatellina and Viola riviniana/reichenbachiana avoided thick F-layers.
The arrows of the AC and Ah humus horizons wereperpendicular to this major gradient (Fig. 2). These twolayers are distinguished by their amount of organicmatter. They were related to the loam content of 5 to 25cm. A loamy soil might result in higher worm activityand thus in thorough mixing of organic matter.
In Holtkrat species variance was small (SD = 2.21),because four species dominated the plots, i.e. Oxalisacetosella, Anemone nemorosa, Stellaria holostea andMelica uniflora. Mercurialis perennis only occurred inseven plots and is correlated with pH, which is relatedto Tilia cordata. M. uniflora was correlated with T.cordata and the external crown projection of the stands.In plots with a more open canopy, Holcus mollis be-came more abundant than M. uniflora. The stands withQ. robur / petraea usually had a more open structure,unless Corylus avellana was present in the shrub layer.There, O. acetosella and A. nemorosa reached higherabundance. Maianthemum bifolium, Convallaria majalisand Luzula pilosa were mostly found in open plots withlow external crown projection and Q. robur / petraeadominance.
In Savelsbos, the DCA gradient in species variancewas 3.0, indicating that both CCA and RDA analysisare useful instruments. Fig. 4 indicates the strong influ-ence of marlstone on the understorey. Marlstone wasfound throughout the profile and positively affected thepH of the mineral soil. In plots with relatively high pHvalues, understorey species are still correlated with treespecies. At the left side of the arrow, species such asMercurialis perennis and Anemone ranunculoides arecorrelated with fraction ‘rich’. At the right side of thepH arrow both the fraction ‘poor’ and the L-layerunderneath these trees are correlated with species suchas Rubus fruticosus aggr. and Convallaria majalis,
Fig. 1. Values for pH of the top 5 cm of mineral soil andthickness of the F- (fermentation) layer in relation to thefractions of ‘poor’ and ‘rich’ tree species respectively – ‘rich’tree species having a relatively base-rich, nutrient-rich, easydecomposable litter as compared to ‘poor’ tree species (seeTable 1 for data on the species per study site). For the siteLimbrichterbosch n = 62, for Holtkrat n = 45, for Savelsbosn = 39 (marl stone plots excluded).
Table 2. Tree species per research area divided into fractions of litter quality.
Fraction tree species ‘Poor’ Intermediate or unknown ‘Rich’
Limbrichterbosch Castanea sativa Betula pendula Alnus glutinosaQuercus robur Acer pseudoplatanus Fraxinus excelsiorQuercus rubra Populus × canadensis; P. alba
Holtkrat Quercus robur; Q. petraea Tilia cordataUlmus spp.Populus tremula
Savelsbos Castanea sativa Betula pendula Carpinus betulusQuercus robur; Q. petraea Acer pseudoplatanus Fraxinus excelsiorQuercus rubra Prunus avium
Tilia cordata

160 VAN OIJEN, D. ET AL.
which are indicative of relatively poor soils andspecies poor vegetation (Hommel et al. 1999; Stortelderet al. 1999).
The Spearman correlation results of ancient forestspecies (occurring in at least 5% of all plots) andhumus characteristics are shown in Table 3. Differentcorrelations are found in each area. For example,Anemone nemorosa was correlated with the pH to 25cm and thickness of the ectorganic layers in Limbricht,whereas in the two other forests no correlation was
found. In Limbrichterbosch the critical pH value forthe occurrence of A. nemorosa was 3.4 and the thick-ness of the F-layer should not exceed 1 cm. In Holtkratand Savelsbos these critical values were not reached.This might explain that A. nemorosa was abundantthroughout these forests, because it was not limited(and therefore not correlated) by low pH values andthick F-layers. The same reasoning holds for Adoxamoschatellina, which only occurred in plots withoutan F-layer. In multivariate analyses, as well as in
Fig. 2. CCA biplot of forest undergrowthvegetation and explanatory environmental factorsin Limbrichterbosch. Species with a low fit arenot shown (< 2) to avoid a crowded diagram.Explained variance: 37%; F-value (model): 2.406;P-value: 0.005; P. canadensis = Populus ×canadensis; Q. robur = Quercus robur; Fr. Rich= fraction of trees with more base-rich, nutrient-rich litter; Fr. Poor = Fraction of tree species withacid, nutrient-poor litter; 0-5 pH = pH of first 5cm mineral soil; 0-5 org. and 5-25 org. = organicmatter content of the first 5 and 5 to 25 cmmineral soil, respectively; 5-25 loam = loamcontent 5 to 25 cm mineral soil; F = Fermentationlayer; Ah and AC: humus horizon; Tot. cov.:external crown projection
Table 3. Spearman correlation values of ancient forest species and humus characteristics in three studied ancient forests. * =significant at 0.05-level; ** = significant at 0.01-level. Lim = Limbrichterbosch; Sav = Savelsbos; Hol = Holtkrat; pH0.5 &pH5.25 = pH of the first 5 and 5-25 cm mineral soil; OM0.5 and OM5.25 = Organic matter content (%) of the first 5 and 5-25 cmmineral soil; L = thickness (cm) of the litter horizon; F = thickness (cm) of the fermentation layer; Ahtot = thickness (cm) of theAh horizon; AC = thickness (cm) of the AC horizon; ECP = external crown projection (%).
Species Area pH0.5 pH5.25 OM0.5 OM5.25 L F Ahtot AC ECP
Adoxa moschatellina Lim 0.50** 0.50** –0.30* –0.33* –0.42** –0.61**Sav
Anemone nemorosa Lim 0.64** 0.62** –0.32* –0.69** –0.59** 0.31* 0.14**SavHol
Anemone ranunculoides Sav 0.40** 0.39** 0.33* –0.40** –0.39** –0.40*Circaea lutetiana Lim 0.43** –0.34** –0.35** 0.14**Lamium galeobdolon SavMaianthemum bifolium Hol –0.40*Melica uniflora Hol 0.36* 0.41*Mercurialis perennis Sav 0.38* 0.37* –0.68** –0.39*
Hol 0.50** –0.45** –0.31* 0.16**Milium effusum Lim 0.40** 0.28* –0.46** –0.44** –0.29* 0.10**Oxalis acetosella HolStellaria holostea Hol –0.13**

- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 161
correlation analyses, Mercurialis perennis was clearlyrelated to high pH values of the mineral soil. At a pH of3.8 only one or two specimen were found, whereas atpH levels higher than 4.0, M. perennis was abundant.
In correlation analyses the strong interaction be-tween pH and humus layers was shown. All species thatwere positively related to pH values were negatively
related to the thickness of F- and L-layers.The correlations found in Holtkrat for light regime,
Melica uniflora and Maianthemum bifolium underlinethe Holtkrat RDA results.
Fig. 3. Redundancy Analysis biplot of the for-est undergrowth and explanatory environmen-tal factors in Holtkrat; Total explained speciesvariance = 37.9%; F-value model = 3.768; P-value = 0.005; 0-5 pH = pH first 5 cm mineralsoil; 5-25 org.: organic matter content 5 to 25cm; Tot. cov.: external crown projection.
Fig. 4. CCA biplot of forest undergrowth andexplanatory environmental factors in Savelsbos; Totalexplained variance = 22%; F-value model = 2.558;P-value model = 0.0050; 0-5 pH = pH of the first fivecm mineral soil; L = litter layer; Fr. Poor = fraction oftree species with acid nutrient-poor litter; Fr. Rich =fraction of tree species with base-rich, nutrient-richlitter

162 VAN OIJEN, D. ET AL.
Tree species - forest understorey relationship
In Table 4 the direct relationship of the understorey(occurring in more than 5% of the plots) and treespecies fractions is shown. In Limbrichterbosch, Adoxamoschatellina, Circaea lutetiana, Viola riviniana/reichenbachiana and Polygonatum multiflorum do notoccur beneath a canopy layer of the fraction ‘poor’ treespecies. However, Rubus fruticosus aggr. and Dryopteriscarthusiana thrive in these stands. Convallaria majalisand Lonicera periclymenum have their optimum in aneven mixture of tree fractions. Anemone nemorosaoccurs in low densities in plots with a large fraction‘poor’, but thrives when tree species with base-richlitter are abundant. In Holtkrat A. nemorosa and Oxalisacetosella occurred abundantly throughout the area.Melica uniflora is found in highest abundance and
frequency in plots where Tilia cordata is abundant.Mercurialis perennis also thrives in those plots. As thesection Holtkrat in Table 4 shows, Maianthemumbifolium and Holcus mollis are indicative of the vegeta-tion found underneath a higher abundance of Quercusrobur / petraea. In Savelsbos the marlstone plots andthe species that are restricted to these plots such asAnemone ranunculoides and Fissidens spp. are ex-cluded. In the remaining plots a relationship betweenthe occurrence of tree species and understorey can beseen but the last class only represents four plots. M.perennis reaches highest abundance with a low abun-dance of fraction ‘poor’. A. nemorosa and Hedera helixoccur slightly less in those plots whereas Rubusfruticosus aggr. reaches its optimum. C. majalis is mostabundant in a mixture of tree species with both litterqualities.
Table 4. Fraction of tree species with nutrient-poor, acid litter (fraction ‘poor’) averaged in three classes in relation to forestunderstorey species in frequencies and characteristic abundance between brackets; classes: without superscript = < 5%, + = 5-10%,I = 10-20%; II = 20-40%; III = 40-60%; IV = 60-80%; V = 80-100%. The median abundance of the main tree species are in Braun-Blanquet values (see Table 1) and the tree species fraction in percentages. Marlstone plots in Savelsbos are excluded.
Limbricht 0-33% 33-66% 66-100% Savelsbos 0-33% 33-66% 66-100%Fraction’ poor’ n = 29 n = 11 n = 22 Fraction ‘poor’ n = 19 n = 13 n = 4
Fraxinus excelsior 4 5 1 Carpinus betulus 5 3 0Populus x canadensis 4 1 1 Quercus robur/petraea 5 7 7Quercus robur/petraea 0 7 8 Tilia cordata 6 6 4
Fraction ‘rich’ 85 % 42 % 4 % Fraction ‘rich’ 60% 46% 20%
Hedera helix 14 - - Mercurialis perennis 42 (III) 54 (I) -Adoxa moschatellina 52 (+) - 5 Adoxa moschatellina 26 (I) - 50Circaea lutetiana 34 9 - Milium effusum 16 - -Eurhynchium praelongum 28 9 (+) - Polygonatum multiflorum 11 - -Ranunuculus ficaria 24 (I) 9 (+) - Galium aparine 11 8 -Geum urbanum 14 (+) 9 - Eurhynchium pumilum 11 8 -Poa trivialis 21 9 - Anemone nemorosa 95 (III) 100 (II) 75 (II)Stellaria holostea 10 9 - Hedera helix 79 (II) 85 (+) 50Deschampsia cespitosa 21 (+) 18 - Convallaria majalis 11 15 -Milium effusum 31 27 - Eurhynchium praelongum 5 15 -Glechoma hederacea 24 (+) 9 5 (+) Lamiastrum galeobdolon 16 (II) 31 (I) -Ribes rubrum 14 (+) 9 5 Rubus fruticosus aggr. 42 (+) 31 (+) 75 (II)Atrichum undulatum 24 - 5Viola riviniana/reichenbachiana 10 - 5 Holtkrat 0-33% 33-66% 66-100%Polygonatum multiflorum 17 - 5Mnium hornum 17 - 5 Fraction ‘poor’ n = 11 n = 5 n = 28
Anemone nemorosa 79 (II) 55 (II) 18 (II) Quercus robur/petraea 5 7 9Eurhynchium pumilum 24 18 (+) 5 Tilia cordata 9 7 2
Fraction ‘rich’ 82% 53% 7%
Brachythecium rutabulum 38 36 5 Mercurialis perennis 36 (II) - 11Convallaria majalis 3 (IV) 27 (III) 5 (+) Melica uniflora 100 (II) 100 (I) 89 (+)Lonicera periclymenum 14 27 5 Anemone nemorosa 100 (II) 100 (II) 100 (III)Athyrium filix-femina 7 - 9 Oxalis acetosella 100 (I) 100 (I) 100 (I)Dryopteris carthusiana 7 - 18 (+) Stellaria holostea 55 80 57Rubus fruticosus aggr. 38 (+) 45 (+) 55 (+) Polygonatum multiflorum 9 - 7
Convallaria majalis 9 - 11 (I)Galeopsis tetrahit 9 - 18Ajuga reptans - - 14Maianthemum bifolium - - 21Holcus mollis - - 43 (I)

- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 163
Discussion
Tree species - humus relationship
Tree species affect physical and chemical proper-ties of the humus profile. These effects vary for thesame tree species on the various soil types. For in-stance, on base-rich soil types in Savelsbos the impactof Quercus robur / petraea on the ectorganic part of thehumus profile was not as strong as in Limbrichterbosch,where luvisols (acidification-sensitive) are found. Twotheories support these findings. Firstly, Nordén (1994a-c) concluded that different soil types influence thequality of tree litter. On base-rich or nutrient-rich soils,leaf litter of a certain tree species contains more nutri-ents and base cations than on poorer soils. However,irrespective of the soil quality, certain tree species, likeTilia cordata, anyhow recycle these nutrients better,than other tree species like Quercus robur/petraea).Secondly, base-rich and loamy soil types enhance soilfauna and therefore the decomposition of litter (Neiryncket al. 2000). The buffering capacity of these soils pre-vents the development of F-layers and acidification. Inacidification-sensitive soils with less buffering capac-ity (old river clay, sandy loess and Pleistocene sandswith more than 15% loam) ectorganic humus profilesdevelop in the presence of tree species that are less ableto recycle nutrients and base cations (Hommel & deWaal 2003a).
In our analysis we used tree species and a divisionby litter quality as explanatory variables. The impact oftrees on soil is, however, dependent on a multitude offactors. Tree species differ in qualities such as stemflow,litter quantity, throughfall of nutrients, root systeminteractions and light regime. Experimental studies areneeded to disentangle all these interacting effects be-tween trees and soil. In our observational study thoseunderlying effects were not studied and therefore onlycauses and effects regarding tree species proper andsoil can be addressed.
Humus - forest understorey relationship
Both chemical and physical properties of the soilinfluence the occurrence and establishment of forestunderstorey species. A thick F-layer hampers certainspecies, whereas others are able to penetrate it (Sydes& Grime 1981; Beatty & Sholes 1988). In Limbrich-terbosch observations were also made on the impor-tance of the surface of leaves in the L-layer. Anemonenemorosa was forced to grow tall white stems to cir-cumvent the compact structure of large leaves fromAcer pseudoplatanus. This illustrates that the distribu-tion of A. nemorosa was negatively influenced by this
tree species, despite its base-rich litter. Den Ouden &Vogels (1997) have shown the negative effects of me-chanical obstruction by the L-layer on the developmentof Pinus sylvestris seedlings. The abundance of A.pseudoplatanus in Limbrichterbosch is too low to de-tect this effect in our statistical analysis.
The three research areas are on a sequence of parentmaterial in terms of base-richness, nutrient richnessand loam content. The interactions between tree species,humus characteristics and understorey differ in inten-sity along this sequence. They are strongest on acidifi-cation-sensitive soils in Limbrichterbosch. In Holtkrat,on soils with higher base saturation, relationships arestill strong but the range in vegetation types and humustypes is smaller and the light climate is of greaterimportance in determining species distribution. InSavelsbos parent material and topography greatly in-fluence soil development.
Tree species - forest understorey
The external projection was a significantly explain-ing factor for species distribution in Holtkrat. Corylusavellana, the most abundant shrub in all three forests,was found to have a positive effect on the abundance ofancient forest species. More light on the forest floor isfavourable to more light demanding species, at the costof ancient forest species (De Keersmaeker et al. 2004;Dzwonko & Gawronski 1994). Therefore, the shrublayer, especially underneath tree species with low inter-nal projection such as Fraxinus excelsior and Quercusrobur / petraea, is important for the distribution ofancient forest species (Verheyen 2002).
Dispersal limitations are often assumed to be themost important factor in the distribution of ancientforest species. Newly established forests, even nearancient forests, do not contain the great variety ofspecies found in ancient forests (Bossuyt et al. 1999;Honnay et al. 1999; Ehrlén & Eriksson 2000; Jacquemynet al. 2001). Our research does not reject these conclu-sions, but offers another view. Within ancient forests,dispersal problems are not to be expected. We tried tominimize variation in independent factors such as par-ent material and hydrology. Therefore, we could focuson soil development and interactions with the forestunderstorey and tree species. Large variances in soilproperties are found within small distances. Nonethe-less, trends were found in pH and F-layer that corre-lated with the occurrence of understorey species. Inturn, these soil characteristics were affected by treespecies. Species abundance and distribution in theunderstorey varied between adjacent stands with differ-ent tree species, where independent factors were con-stant. To illustrate: Anemone nemorosa was abundant

164 VAN OIJEN, D. ET AL.
underneath a Fraxinus stand in Limbrichterbosch, butvirtually absent 2 m away beneath Quercus robur /petraea. This cannot be explained by a dispersal prob-lem, but was related to humus characteristics. In newlyestablished forests, without developed forest soils andwith more soil disturbance, suboptimal growing condi-tions for ancient forest species might be improved byplanting tree species with base- and nutrient-rich litter.If those tree species are planted on former heathlandson poor sandy soils they might improve the base satura-tion and keep acidification at bay. On former agricul-tural lands with high P-contents a closed canopy (e.g.Tilia spp.) might favour ancient forest species by limit-ing the competitiveness of ruderal species.
The benefits of tree species with base- and nutrient-rich litter are measured in chemical and physical prop-erties of the soil, including higher base saturation,CEC, pH, soil fauna activity, decomposition rate, totalporosity and lower C:N ratios (Binkley & Valentine1991; Finzi et al. 1998 a,b; Neirynck et al. 2000). Evenwithin the life span of one tree the effect of trees onthese soil properties is found (Neirynck et al. 2000).The right choice in tree species could be a practicalmanagement tool to enhance biodiversity in understoreyand canopy layer and in this way contribute to theconservation of endangered or rare species.
Conclusions
We conclude that canopy species affect character-istics of the humus profile through the nature of theirlitter. Base-rich litter from trees such as Tilia cordata,Fraxinus excelsior and Populus × canadensis enhancesmore endorganic humus profiles. Underneath Quercusrobur / petraea, Q. rubra and Castanea sativa, whichare tree species with more acid litter, ectorganic humusprofiles develop. Characteristics of the humus profile,especially the pH of the upper 25 cm mineral soil andectorganic layers affect the abundance and the distribu-tion of forest species. However, the impact of treespecies on the distribution of forest species throughhumus characteristics varies with different soil types.
Most research on the distribution of ancient forestspecies does not take soil properties, humus profile andtree species into account. To discern between dispersallimitations and recruitment limitation, we recommendthat the independent and dependent environmental fac-tors (e.g. visualized in the humus profile) are investi-gated alongside dispersal factors (see also Verheyen2002; Verheyen et al. 2003).
Acknowledgements. We acknowledge Staatsbosbeheer andNatuurmonumenten for their permission to conduct our re-search in their forests. Kristian Dalsgaard and Niels PeterDalsgaard Jensen kindly allowed us to conduct research inHoltkrat. We thank Obbe Boersma for analysing soil samplesand Wim Braakhekke for constructive comments on ourresearch. The paper benefited from the comments of anony-mous referees.
References
Anon. 1975. Soil taxonomy. A basic system of soil classifi-cation for making and interpreting soil surveys. Agricul-ture Handbook 436. Soil Survey Staff, USDA, Washing-ton, DC, US.
Anon. 2001. SPSS Base 11.0: user’s guide. SPSS, Chicago,IL, US.
Beatty, S.W. & Sholes, O.D.V. 1988. Leaf litter effects onplant species composition of deciduous forest treefallpits. Can. J. For. Res. 18: 553-559.
Bijlsma, R.J., van Blitterswijk, H., Clerkx, A.P.P.M., deJong, J.J., van Wijk, M.N. & van Os, L.J. 2001. Bospadenvoor bosplanten. Bospaden en -wegen als transportroute,vestigingsmilieu, refugium en uitvalsbasis voor bosplanten.Alterra rapport 193, Wageningen, NL.
Binkley, D. & Valentine, D. 1991. Fifty year biogeochemicaleffects of green ash, white pine, and Norway spruce in areplicated experiment. For. Ecol. Manage. 40: 13-25.
Birks, H.J.B. 1986. Late-Quaternary biotic changesinterrestrial and lacustrine environments, with particularreference to north-west Europe. In: Berglund, B.E.,Ralska-Jasiewiczowa, M. (eds.) Handbook of holocenepalaeoecology and palaeohydrology. Wiley, Chicester, UK.
Bossuyt, B., Hermy, M. & Deckers, J. 1999. Migration ofherbaceous plant species across ancient-recent forestecotones in central Belgium. J. Ecol. 87: 628-638.
Brunet, J., von Oheimb, G. & Diekmann, M. 2000. Factorsinfluencing vegetation gradients across ancient-recentwoodland borderlines in southern Sweden. J. Veg. Sci.11: 515-524.
De Keersmaeker, L., Martens, L., Verheyen K., Hermy, M.,De Schrijver, A. & Lust, N. 2004. Impact of soil fertilityand insolation on diversity of herbaceous woodlandspecies colonizing afforestations in Muizen forest (Bel-gium). For. Ecol. Manage. 188: 291-304.
den Ouden, J. & Vogels, D. 1997. Mechanical resistance byan ectorganic soil layer on root development of seedlingPinus sylvestris. Plant Soil 197: 209-217.
Dirkse, G.M. & Martakis, G.F.P. 1998. Species density ofphanerogams and bryophytes in Dutch forests. Biodiv.Conserv. 7: 147-157.
Dzwonko, Z. & Gawronski S. 1994. The role of woodlandfragments, soil type, and dominant species in secondarysuccession on the western Carpathian foothills. Vegetatio111: 149-160.
Ehrlén, J. & Eriksson, O. 2000. Dispersal limitation andpatch occupancy in forest herbs. Ecology 81: 1667-1674.
Faliński, J.B. & Falińska, K. 1986. Vegetation dynamics in

- EFFECTS OF TREE SPECIES COMPOSITION ON WITHIN-FOREST DISTRIBUTION OF UNDERSTOREY SPECIES - 165
temperate lowland primeval forests. Ecological studiesin Bia∏owieża forest. Geobotany 8. Junk, Dordrecht, NL.
Finzi, A.C., van Breemen, N. & Canham, C.D. 1998a. Canopytree-soil interactions within temperate forests: specieseffects on soil carbon and nitrogen. Ecol. Appl. 8: 440-446.
Finzi, A.C., van Breemen, N. & Canham, C.D. 1998b. Canopytree-soil interactions within temperate forests: specieseffects on pH and cations. Ecol. Appl. 8: 447-454.
Graae, B.J., Sunde, P.B. & Fritzbøger, B. 2003. Vegetationand soil differences in ancient opposed to new forests.For. Ecol. Manage. 177: 179-190.
Grashof-Bokdam, C. 1997. Forest species in an agriculturallandscape in The Netherlands: Effects of habitat frag-mentation. J. Veg. Sci. 8: 21-28.
Harmer, R., Peterken, G., Kerr, G. & Poulton, P. 2001.Vegetation changes during 100 years of development oftwo secondary woodlands on abandoned arable land.Biol. Conserv. 101: 291-304.
Heath, G.W., Arnold, M.K. & Edward, C.A. 1966. Studies inleaf litter breakdown. I. Breakdown rates of leaves ofdifferent species. Pedobiologica 6: 1-12.
Hermy, M., Honnay, O., Firbank, L., Grashof-Bokdam, C. &Lawesson, J.E. 1999. An ecological comparison betweenancient and other forest plant species of Europe, and theimplications for forest conservation. Biol. Conserv. 91:9-22.
Hommel, P.W.F.M., van Dort, K.W. & Schaminée, J.H.J.1999. Quercetea robori-petraeae (klasse der eiken- enbeukenbossen op voedselarme grond). In: Stortelder,A.F.H., Schaminée, J.H.J., Hommel, P.W.F.M. (eds). Devegetatie van Nederland Deel 5. Plantengemeenschappenvan ruigten, struwelen en bossen, pp 255-286. OpulusPress, Uppsala, SE. (In Dutch.)
Hommel, P.W.F.M., Spek, Th. & de Waal, R.W. 2002.Boomsoort, strooiselkwaliteit en ondergroei op ver-zuringsgevoelige bodem. Een verkennend literatuur- enveldonderzoek. Alterra rapport 509, Wageningen, NL.
Hommel, P.W.F.M. & de Waal, R.W. 2003a. Boomsoortbepaalt bostype op verzuringsgevoelige bodem. Stratiotes26: 3-19. (In Dutch.)
Hommel, P.W.F.M. & de Waal, R.W. 2003b. Rijke bossenop arme bodems. Alternatieve boomsoortenkeuzeverhoogt soortenrijkdom ondergroei op verzurings-gevoelige bodems. Landschap 4: 192-204. (In Dutch.)
Honnay, O., Hermy, M. & Coppin, P. 1999. Effects of area,age and diversity of forest patches in Belgium on plantspecies richness, and implications for conservation andreforestation. Biol. Conserv. 87: 73-84.
Jacquemyn, H., Butaye, J. & Hermy, M. 2001. Forest plantspecies richness in small, fragmented mixed deciduousforest patches: the role of area, time and dispersal limita-tion. J. Biogeogr. 28: 801-812.
Jenny, H. 1941. Factors of soil formation. A system of quan-titative pedology. McGraw-Hill, New York, NY, US.
Jongman, R.H.G., ter Braak, C.J.F. & van Tongeren, O.F.R.1995. Data analysis in community and landscape ecol-ogy. Cambridge University Press, Cambridge, UK.
Kemmers, R.H. & de Waal, R.W. 1999. Ecologische typering
van bodems. Raamwerk en humusvorm typologie. AlterraRapport 667-1, Wageningen, NL. (In Dutch.)
Klinka, K., Green, R.N & Trowbridge, R.L. 1981. Taxo-nomic classification of humus forms in ecosystems ofBritish Colombia. Land Management Report 8.Vancou-ver Forest Region, Vancouver, CA.
Koerner, W., Dupouey, J.L., Dambrine, E. & Benoît, M.1997. Influence of past land use on the vegetation andsoils of present day forest in the Vosges mountains,France. J. Ecol. 85: 351-358.
Kristiansen, S.M. & Dalsgaard, K. 2000. Soil evolution in theremnants of natural forest vegetation: An example froman old oak-lime coppice wood in Denmark. Dan. J.Geogr. 100: 27-36.
Kuiters, A.T. 1987. Phenolic acids and plant growth in forestecosystems. Thesis VU. Free University Press, Amster-dam, NL.
Maddelein, D., Muys, B., Neirynck, J. & Sioen, G. 1994. Siteclassification in a mixed hardwood forest, Hallerbos,Belgium. with a homogeneous ground vegetation domi-nated by Hyacinthoides nonscripta L. Chouard ex. Rothm.Silva Gandavensis 59: 15-27.
Neirynck, J., Mirtcheva, S., Sioen, G. & Lust, N. 2000.Impact of Tilia platyphyllos Scop., Fraxinus excelsior L.,Acer pseudoplatanus L., Quercus robur L. and Fagussylvatica L. on earthworm biomass and physicochemicalproperties of a loamy topsoil. For. Ecol. Manage. 133:275-286.
Nordén, U. 1994a. Leaf litterfall concentrations and fluxes ofelements in deciduous tree species. Scan. J. For. Res. 9:9-16.
Nordén, U. 1994b. Influence of tree species on acidificationand mineral pools in deciduous forest soils of SouthSweden. Water Air Soil Pollut. 76: 363-381.
Nordén, U. 1994c. Influence of broad-leaved tree species onpH and organic matter content of forest topsoils in Scania,South Sweden. Scan. J. For. Res. 9: 1-8.
Palmer, M.W. 1993. Putting things in even better order: theadvantages of canonical correspondence analysis. Ecol-ogy 74: 2215-2230.
Persson, S., Malmer, N. & Wallén, B. 1987. Leaf litter falland soil acidity during half a century of secondary suc-cession in a temperate deciduous forest. Vegetatio 73:31-45.
Peterken, G.F. 1974. A method of assessing woodland florafor conservation using indicator species. Biol. Conserv.6: 239-245.
Peterken, G.F. 1977. Habitat conservation priorities in Brit-ish and European woodlands. Biol. Conserv. 11: 223-236.
Peterken, G.F. 1993. Woodland conservation and manage-ment. Chapman & Hall, Cambridge, UK.
Petersen, P.M. 1994. Flora, vegetation, and soil in broad-leaved ancient and planted woodland, and scrub onRosnas, Denmark. Nord. J. Bot. 14: 693-709.
Pigott, C.D. 1989. The growth of lime Tilia cordata in anexperimental plantation and its influence on soil devel-opment and vegetation. Q. J. For. 83: 14-24.
Rackham, O. 1980. Ancient woodland; its history, vegetation

166 VAN OIJEN, D. ET AL.
and uses in England. Edward Arnold, London, UK.Scheffer, F., Schachtschabel, P., Blume, H.P. & Hartge, K.H.
1982. Lehrbuch der Bodenkunde. Enke, Stuttgart, DE.Schols, R. 1986. Limbrichterbosch beheersplan. Natuur-
monumenten, s’Graveland, NL.Schoonderwoerd, M. & Nyssen, B. 1999. Moerslag 18.
Historisch beheer en huidige vegetatie van het Savelsbos.Student report WUR, Wageningen, NL. (In Dutch.)
Stortelder, A.H.F., Schaminée, J.H.J. & Hermy, M. 1999.Querco-Fagetea (klasse der eiken- en beukenbossen opvoedselrijke grond). In: Stortelder, A.F.H., Schaminee,J.H.J. & Hommel, P.W.F.M. (eds) De vegetatie vanNederland Deel 5. Plantengemeenschappen van ruigten,struwelen en bossen, pp. 287-331. Opulus, Leiden, NL,Uppsala, SE. (In Dutch.)
Sydes, C. & Grime, J.P. 1981. Effects of tree leaf litter onherbaceous vegetation in deciduous woodland I. Fieldinvestigations. J. Ecol. 69: 237-248.
ter Braak, C.J.F. 1996. Unimodal models to relate species toenvironment. Agricultural Mathematics Group, Wage-ningen, NL.
ter Braak, C.J.F. & Smilauer, P. 1998. Canoco ReferenceManual and User’s Guide to Canoco for Windows. Soft-ware for Canonical Community Ordination. Centre forBiometry, Wageningen, NL.
van der Maarel, E. 1979. Transformation of cover-abundancevalues in phytosociology and its effects on communitysimilarity. Vegetatio 39: 97-114.
Verheyen, K. 2002. The relative importance of seed andrecruitment limitation of vascular plants in secondaryforest succession. Dissertation 511. Katholieke Univer-siteit Leuven, BE.
Verheyen, K., Bossuyt, B., Hermy, M. & Tack, G. 1999. Theland use history, 1278-1990 of a mixed hardwood forestin western Belgium and its relationship with chemicalsoil characteristics. J. Biogeogr. 26: 1115-1128.
Verheyen, K., Guntenspergen, R., Biesbrouck, B. & Hermy,M. 2003. An integrated analysis of the effects of past landuse on forest herb colonization at the landscape scale. J.Ecol. 91: 731-742.
Vleeshouwer, J.J. & Damoiseaux, J.H. 1990. Bodemkaartvan Nederland. Toelichting bij kaartblad 61-62 West enOost Maastricht - Heerlen. SC, Wageningen, NL. (InDutch.)
Whitney, G.G. & Foster, D.R. 1988. Overstorey compositionand age as determinants of the understorey flora of woodsof central New England. J. Ecol. 76: 867-876.
Wulf, M. 1997. Plant species as indicators of ancient wood-land in north-western Germany. J. Veg. Sci. 8: 635-642.
Received 17 March 2004;Accepted 13 July 2005.
Co-ordinating Editor: M. Hermy.
Copyright © 2022 FDOKUMEN




















