
Modelling of ammonia volatilisation in fertilised and flooded ...

Oecologia (2003) 136:193–204DOI 10.1007/s00442-003-1259-7
E C O P H Y S I O L O G Y
Omar R. Lopez · Thomas A. Kursar
Does flood tolerance explain tree species distributionin tropical seasonally flooded habitats?
Received: 19 November 2001 / Accepted: 17 March 2003 / Published online: 13 May 2003� Springer-Verlag 2003
Abstract In the tropics, seasonally flooded forests (SFF)harbor fewer tree species than terra firme (i.e. non-flooded) forests. The low species diversity of tropicalflooded forests has been ascribed to the paucity of specieswith adaptations to tolerate flooding. To test the hypoth-esis that flooding is the only factor restricting mostspecies from SFF, we compared plant morphological andphysiological responses to flooding in 2-month oldseedlings of 6 species common to SFF and 12 speciescommon to terra firme forests. Although floodingimpaired growth, total biomass, maximum root lengthand stomatal conductance in most species, responsesvaried greatly and were species-specific. For example,after 90 days, flooding reduced leaf area growth by 10–50% in all species, except in Tabebuia, a common speciesfrom non-flooded habitats. Similarly, flooding had a 5–45% negative effect on total biomass for all species,except in 1 SFF and 1 terra firme species both of whichhad more biomass under flooding. A principal componentanalysis, using the above responses to flooding, providedno evidence that SFF and terra firme species differed intheir responses to flooding. Flooding also caused reduc-tions in root growth for most species. Rooting depth androot: shoot ratios were significantly less affected byflooding in SFF than in terra firme species. Althoughflood tolerance is critical for survival in flooded habitats,we hypothesize that responses to post-flooding eventssuch as drought might be equally important in seasonal
habitats. Therefore, we suggest that the ability to growroots under anoxia might be critical in predicting successin inundated habitats that also experience a strong dryseason.
Keywords Panama · Root growth · Seasonality ·Stomatal conductance · Swamps
Introduction
In the tropics, low diversity forests are commonlyassociated with seasonally waterlogged conditions (Davisand Richards 1933; Janzen 1974; Peters et al. 1989; Hart1990; Richards 1996). Shortly after flooding, oxygendepletion at the rhizosphere could result in reducedstomatal conductance, lower photosynthesis, plant hor-monal imbalance, poor water and nutrient uptake,ultimately compromising plant growth and survival(Pezeshki 1994). Thus the reduced species richness oftropical swamps has been ascribed to the inability of terrafirme species (species from non-flooded forests) to copewith the stressful conditions resulting from flooding(Black et al. 1950; Campbell et al. 1992; Duivenvoorden1996).
Contrary to this assertion, recent studies suggest thatflood tolerance of tropical trees might be more commonthan previously thought, although the extent to whichtropical trees are flood tolerant has not been thoroughlyinvestigated. For example, Genipa americana L. (Rubi-aceae), a widely distributed Neotropical species found inflood-prone habitats in Brazil, but most commonlyoccurring in terra firme elsewhere, is a flood-tolerantspecies (Andrade et al. 1999). Likewise, Hymeneacourbaril L. var. stilocarpa (Caesalpinioideae), a dryforest species, and Corisia speciosa St. Hill (Bomba-caceae), a terra firme species, were found to be highlyflood tolerant (Joly and Crawford 1982). Thus, floodtolerance might be common among species that experi-ence little or no flooding in their natural habitat.
O. R. Lopez ()) · T. A. KursarDepartment of Biology, University of Utah,257 S., 1400 E., Salt Lake City, UT 84112-0840, USAe-mail: [email protected].: +1-828-4880178
T. A. KursarSmithsonian Tropical Research Institute,Apartado 2072, Balboa, Anc�n, Panama
Present address:O. R. Lopez, Department of Botany,University of Wisconsin-Madison,430 Lincoln Dr., Madison, WI 53706-1381, USA

To test the hypothesis that flood tolerance regulatesspecies diversity in seasonally flooded forests (SFF), weevaluated flood tolerance in tropical tree species. For thispurpose we investigated growth, physiological and mor-phological responses to flooding in 2-month-old seedlingsof 18 Neotropical tree species. We studied 6 species fromSFF, and 12 terra firme species that are common inlowland tropical forests, but that very rarely experienceflooding (see Table 1). If the hypothesis of flood toleranceis true, we would expect terra firme species to besignificantly less tolerant to flooding than SFF species.Furthermore, light demanding and pioneer species appearuncommon in some SFF (Parolin 2000; Lopez, personalobservation). Thus, to see whether light-demandingspecies are more susceptible to flooding than shade-tolerant species, we included 6 light-demanding species, 2from SFF and 4 from terra firme forests.
Materials and methods
Study site and species
Flooding experiments were carried out in a screened house fromDecember 1998 to September 2000 on Barro Colorado Island(BCI), a research facility administered by the Smithsonian TropicalResearch Institute in central Panama (9�90N, 79�510W). A completedescription of the site and its vegetation can be found in Leigh et al.(1996) and Croat (1978), respectively. Mean annual rainfall on BCIis 2,600 mm with a pronounced dry season from mid-December tothe beginning of May (Windsor 1990). A total of 18 species werestudied (Table 1). Among the SFF species Prioria is unique as itforms extensive mono-dominant stands in SFF of Central America
from Nicaragua to Colombia (Mayo-Mel�ndez 1965; Linares andMart�nez 1991). With the exception of Pachira, which is mainlyfound in riverine habitats, the other five SFF species also occur innon-flooded forests. Although terra firme species are most commonin lowland non-flooded tropical forests, Anacardium, Tabebuia,Dipteryx and Virola also occur at very low densities in riparian orSFF (Linares and Mart�nez 1991; Grauel and Kursar 1999). Theterra firme and SFF species include both shade-tolerant and light-demanding species (Table 1).
Experimental design
Seeds of terra firme species were collected on BCI where treesnever experience flooding, while seeds of SFF species werecollected at Rio Balsas, Dari�n (8�070N, 77�520W) and broughtback to BCI within 48 h. Seeds were germinated in 0.5-l plasticpots containing sandy loam soil (Utah State University SoilsLaboratories, Logan, Utah). Seedlings were grown under shadedconditions (0.8 mol photons m�2 day�1) and transplanted 3 weeksprior to the experiment into plastic pots (0.5 m tall � 0.2 mdiameter), one plant per pot. Seedlings were about 10 weeks oldand of similar size within each species (e.g. leaf area and seedlingheight) when assigned to the treatments. Flooded and non-floodedtreatments comprised a total of nine or ten plants each, for allspecies, and were maintained under shaded conditions. Floodingwas maintained in water-filled containers for 90 days providing awater level at least 3 cm above the soil surface. Non-flooded plants(=control) were well watered but never flooded.
Measurements
Soil redox potentials were subsampled for all species at a depth of0.3 m after 1, 2, 3, 5, 6, 13, 21, 30, 45, 80, and 90 days in floodedand non-flooded pots. Soil redox potentials were taken using aPlatinum (Pt) redox electrode, coupled with a silver (Ag/AgCl)electrode (Jensen Instruments, Tacoma, Wash.). Redox potentials
Table 1 Species name, family, growth form, light environment andhabitat. Light environment follows Welden et al. 1999, R. Conditpersonal communication and personal observation (CT canopy tree,
Sub-CT sub-canopy tree,SFFseasonally flooded forests,TF terrafirme forests)
Family Growth form Light environment Habitat Reference
SFF species
Carapa guianensis Meliaceae CT Shade-tolerant SFF and TF Jim�nez 1994Pachira aquatica Bombacaceae River edges Light-demanding SFF Jim�nez 1994Pentaclethra macroloba Fabaceae CT Shade-tolerant SFF and TF Hartshorn 1972Prioria copaifera Fabaceae CT Shade-tolerant SFF and TF Condit et al. 1993,
Grauel and Kursar 1999Pterocarpus officinalis Fabaceae CT Light-demanding SFF Grauel and Kursar 1999Pterocarpus sp. Fabaceae Unknown SFF Personal observation
Terra firme species
Anacardium excelsum Anacardiaceae CT Light-demanding TF and SFF Grauel and Kursar 1999,Condit et al. 1996
Aspidosperma cruenta Apocynaceae CT Shade-tolerant TF Condit et al. 1996Brosimum alicastrum Moraceae CT Shade-tolerant TF Condit et al. 1996Calophyllum longifolium Clusiaceae CT Shade-tolerant TF Condit et al. 1996Croton billbergianus Euphorbiaceae Sub-CT Light-demanding TF Personal observationDipteryx panamensis Fabaceae CT Light-demanding TF and SFF Grauel and Kursar 1999,
Condit et al. 1996Guazuma ulmifolia Tiliaceae CT Light-demanding TF Personal observationGustavia superba Lecythidaceae Sub-CT Shade-tolerant TF Condit et al. 1996, Sork
1985Tabebuia rosea Bignoniaceae CT Shade-tolerant TF and SFF Grauel and Kursar 1999,
Condit et al. 1996Tachigali versicolor Fabaceae CT Shade-tolerant TF Condit et al. 1996Tetragastris panamensis Burseraceae CT Shade-tolerant TF Condit et al. 1996Virola surinamensis Myristicaceae CT Shade-tolerant TF and SFF Berry et al. 1995
194
were corrected (converted to Eh) by adding +242 mV to thepotential given by the platinum electrode. Redox potentials werenot adjusted for pH changes due to negligible pH differencesbetween the two treatments (average pH =6.7).
Leaf number and leaf area were measured every 15 days fromthe beginning of the experiment using a portable leaf area meter(LI-3000, Li-Cor, Lincoln, Neb.). At the end of the experiment(90 days), plants were harvested and seedling height, maximumroot depth and stem diameter at 0.05 m above the soil weremeasured. Additionally, morphological responses to flooding werenoted, specifically the formation of hypertrophied lenticels andadventitious roots. After 90 days, plants were harvested, dividedinto above- and belowground parts and dried at 70�C for 72 h forbiomass.
Stomatal conductance (gst) was measured after 1, 2, 5, 12, 15and 30 days of treatment with a steady-state porometer (LI-1600,Li-Cor, Lincoln, Neb.) on leaves that had matured prior to thetreatments. Measurements were conducted in a growth chamber(Model M-Sun, Environmental Growth Chambers, Chagrin Falls,Ohio) between 0900 and 1300 hours at saturating light (500–600 mmol m—2 s�1, photosynthetic photon flux density, 25–30�Cand 70–75% relative humidity). gst was measured on a minimum ofsix plants per treatment and at least two leaves per plant. Plantswere allowed to acclimate for 8–10 min prior to each measurementand measured in pairs, one control and one flooded plant perspecies.
Data analysis
The effect of flooding on soil redox potential was tested using ananalysis of variance (ANOVA). However, the comparison at90 days was made by Student’s t-test. Flooding effects on leaf areagrowth and gst in each species were analyzed by repeated measuresANOVA. After excluding several highly correlated parameters andnormalizing the data when needed (log x), the effects of flooding oneach species were analyzed by multivariate analysis-of-variance(MANOVA, F-test) using five parameters: height relative growthrate {RGRheight=[ln (final height cm)�ln (initial height)]/3 months},leaf mass per area, total biomass, root: shoot ratios and maximumroot depth. Following this, each parameter was tested independent-ly and the alpha level was adjusted by taking into consideration thenumber of tests performed (k=5). Therefore the effect of floodingon each parameter was tested at the P=0.01 level (Bonferronicorrection; Sokal and Rohlf 1995; Cobin and Mitchel 2000). Weused ANOVA to test for treatment (flooding, non-flooding) andhabitat (SFF, terra firme) main effects and their interaction onseedling performance. Species effects were nested within habitatand treated as random effects. Principal component analysis (PCA)was used to determine species grouping patterns with respect togrowth, morphological and physiological responses to flooding(Fernandez et al. 1993; Golluscio and Sala 1993). Data analysis wasconducted with the statistical software JUMP v 3.2.1. (SASInstitute, Cary, N.C.).
Results
Soil redox potential
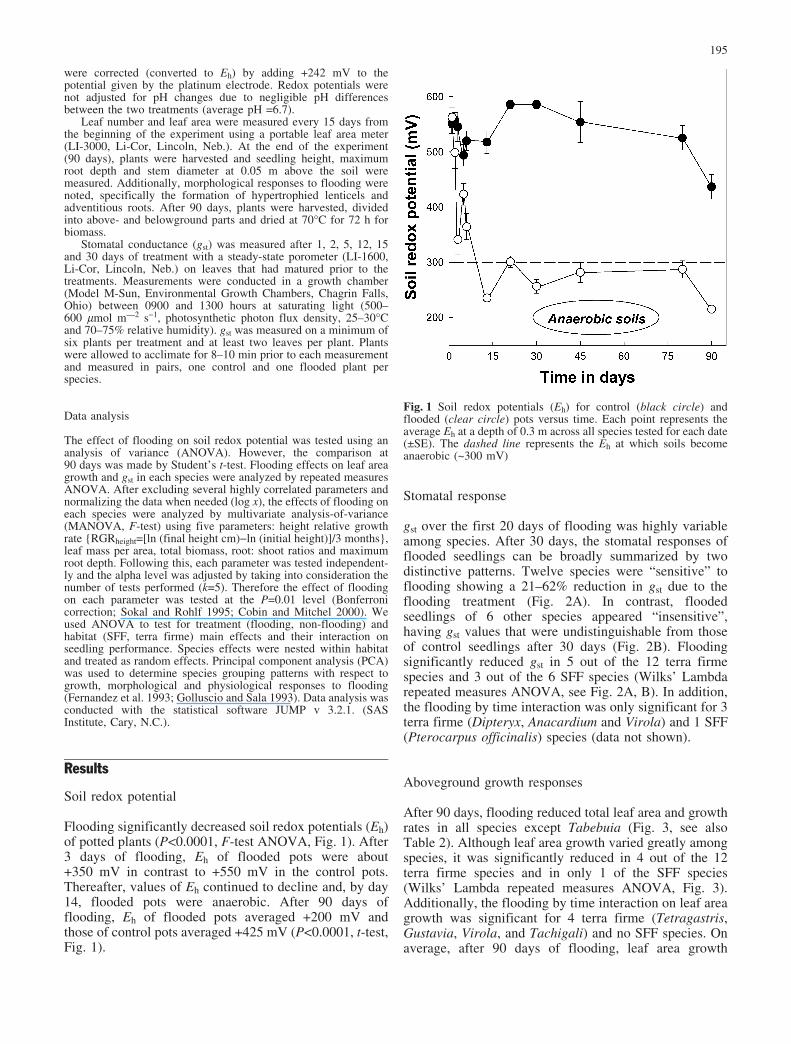
Flooding significantly decreased soil redox potentials (Eh)of potted plants (P<0.0001, F-test ANOVA, Fig. 1). After3 days of flooding, Eh of flooded pots were about+350 mV in contrast to +550 mV in the control pots.Thereafter, values of Eh continued to decline and, by day14, flooded pots were anaerobic. After 90 days offlooding, Eh of flooded pots averaged +200 mV andthose of control pots averaged +425 mV (P<0.0001, t-test,Fig. 1).
Stomatal response
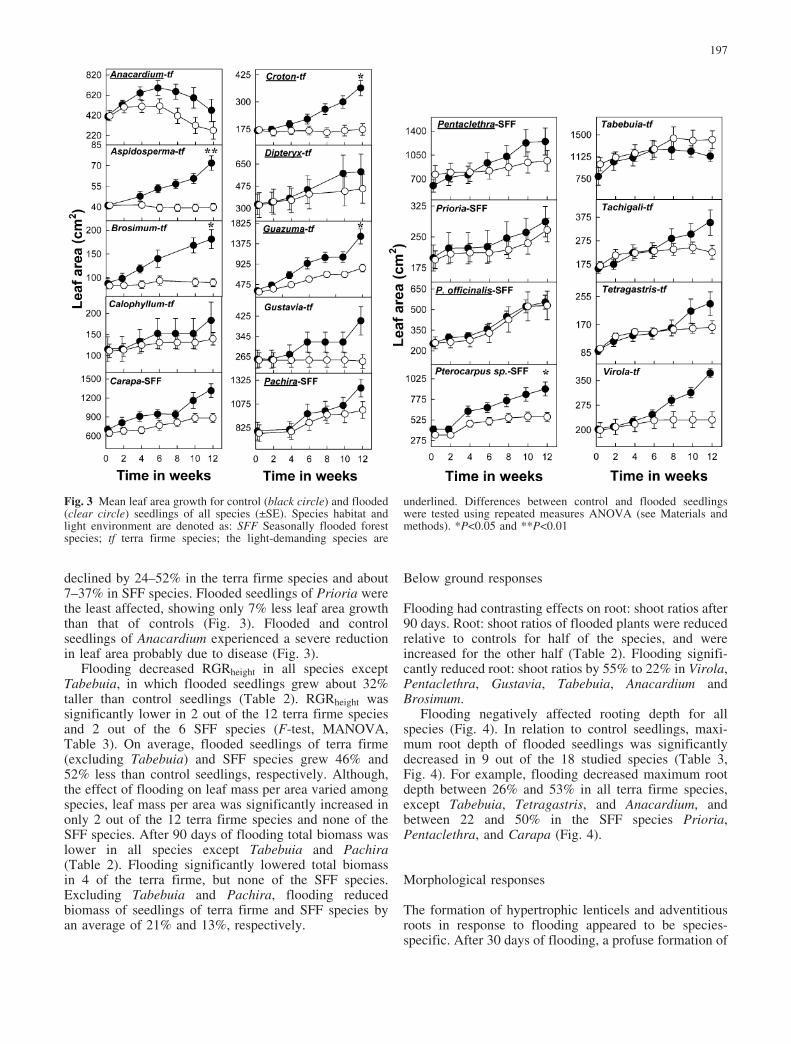
gst over the first 20 days of flooding was highly variableamong species. After 30 days, the stomatal responses offlooded seedlings can be broadly summarized by twodistinctive patterns. Twelve species were “sensitive” toflooding showing a 21–62% reduction in gst due to theflooding treatment (Fig. 2A). In contrast, floodedseedlings of 6 other species appeared “insensitive”,having gst values that were undistinguishable from thoseof control seedlings after 30 days (Fig. 2B). Floodingsignificantly reduced gst in 5 out of the 12 terra firmespecies and 3 out of the 6 SFF species (Wilks’ Lambdarepeated measures ANOVA, see Fig. 2A, B). In addition,the flooding by time interaction was only significant for 3terra firme (Dipteryx, Anacardium and Virola) and 1 SFF(Pterocarpus officinalis) species (data not shown).
Aboveground growth responses
After 90 days, flooding reduced total leaf area and growthrates in all species except Tabebuia (Fig. 3, see alsoTable 2). Although leaf area growth varied greatly amongspecies, it was significantly reduced in 4 out of the 12terra firme species and in only 1 of the SFF species(Wilks’ Lambda repeated measures ANOVA, Fig. 3).Additionally, the flooding by time interaction on leaf areagrowth was significant for 4 terra firme (Tetragastris,Gustavia, Virola, and Tachigali) and no SFF species. Onaverage, after 90 days of flooding, leaf area growth
Fig. 1 Soil redox potentials (Eh) for control (black circle) andflooded (clear circle) pots versus time. Each point represents theaverage Eh at a depth of 0.3 m across all species tested for each date(€SE). The dashed line represents the Eh at which soils becomeanaerobic (~300 mV)
195

Fig. 2A, B Mean stomatalconductance during the first30 days of the experiment forcontrol (black circle) andflooded (clear circle) seedlingsof all species (€SE). A Speciesfor which flooding resulted in asignificant decrease in gst at30 days, i.e. “sensitive” species.B Species for which floodingdid not have an effect on gst at30 days, i.e. “insensitive” spe-cies. Species habitat and lightenvironment are denoted as:SFF Seasonally flooded forestspecies; tf terra firme species;and the light-demanding speciesare underlined. Differences be-tween control and floodedseedlings were analyzed by re-peated measures ANOVA (seeMaterials and methods).*P<0.05, **P<0.01,***P<0.001
196
declined by 24–52% in the terra firme species and about7–37% in SFF species. Flooded seedlings of Prioria werethe least affected, showing only 7% less leaf area growththan that of controls (Fig. 3). Flooded and controlseedlings of Anacardium experienced a severe reductionin leaf area probably due to disease (Fig. 3).
Flooding decreased RGRheight in all species exceptTabebuia, in which flooded seedlings grew about 32%taller than control seedlings (Table 2). RGRheight wassignificantly lower in 2 out of the 12 terra firme speciesand 2 out of the 6 SFF species (F-test, MANOVA,Table 3). On average, flooded seedlings of terra firme(excluding Tabebuia) and SFF species grew 46% and52% less than control seedlings, respectively. Although,the effect of flooding on leaf mass per area varied amongspecies, leaf mass per area was significantly increased inonly 2 out of the 12 terra firme species and none of theSFF species. After 90 days of flooding total biomass waslower in all species except Tabebuia and Pachira(Table 2). Flooding significantly lowered total biomassin 4 of the terra firme, but none of the SFF species.Excluding Tabebuia and Pachira, flooding reducedbiomass of seedlings of terra firme and SFF species byan average of 21% and 13%, respectively.
Below ground responses
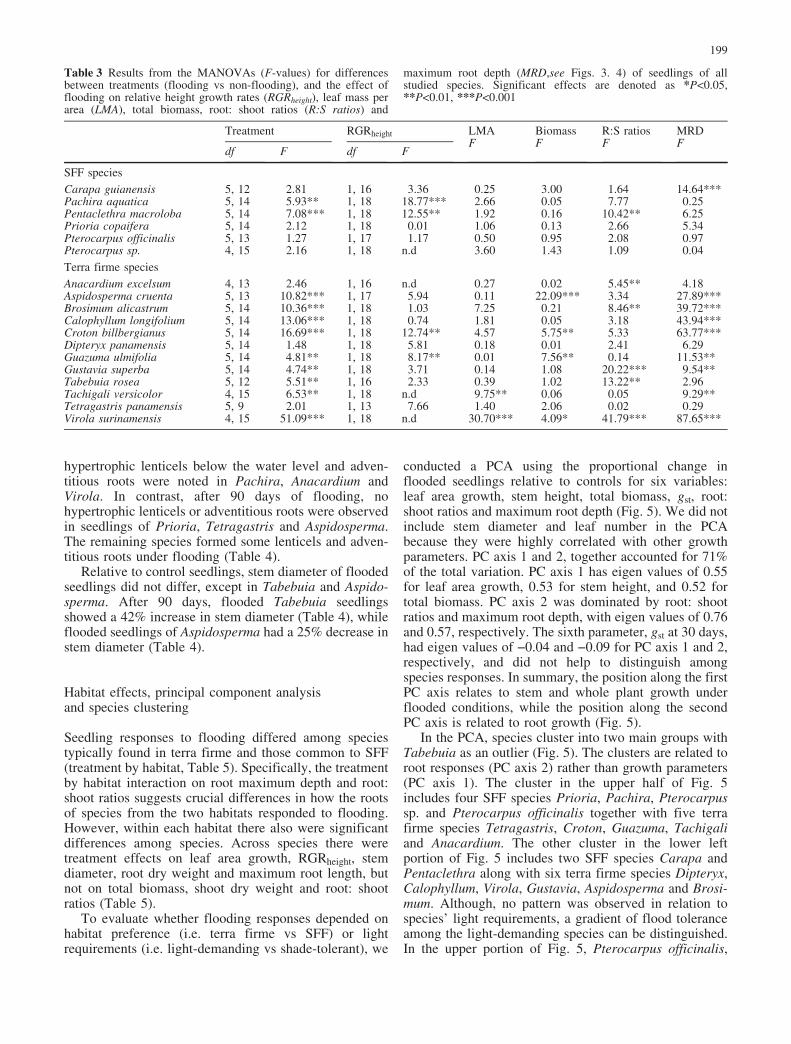
Flooding had contrasting effects on root: shoot ratios after90 days. Root: shoot ratios of flooded plants were reducedrelative to controls for half of the species, and wereincreased for the other half (Table 2). Flooding signifi-cantly reduced root: shoot ratios by 55% to 22% in Virola,Pentaclethra, Gustavia, Tabebuia, Anacardium andBrosimum.
Flooding negatively affected rooting depth for allspecies (Fig. 4). In relation to control seedlings, maxi-mum root depth of flooded seedlings was significantlydecreased in 9 out of the 18 studied species (Table 3,Fig. 4). For example, flooding decreased maximum rootdepth between 26% and 53% in all terra firme species,except Tabebuia, Tetragastris, and Anacardium, andbetween 22 and 50% in the SFF species Prioria,Pentaclethra, and Carapa (Fig. 4).
Morphological responses
The formation of hypertrophic lenticels and adventitiousroots in response to flooding appeared to be species-specific. After 30 days of flooding, a profuse formation of
Fig. 3 Mean leaf area growth for control (black circle) and flooded(clear circle) seedlings of all species (€SE). Species habitat andlight environment are denoted as: SFF Seasonally flooded forestspecies; tf terra firme species; the light-demanding species are
underlined. Differences between control and flooded seedlingswere tested using repeated measures ANOVA (see Materials andmethods). *P<0.05 and **P<0.01
197
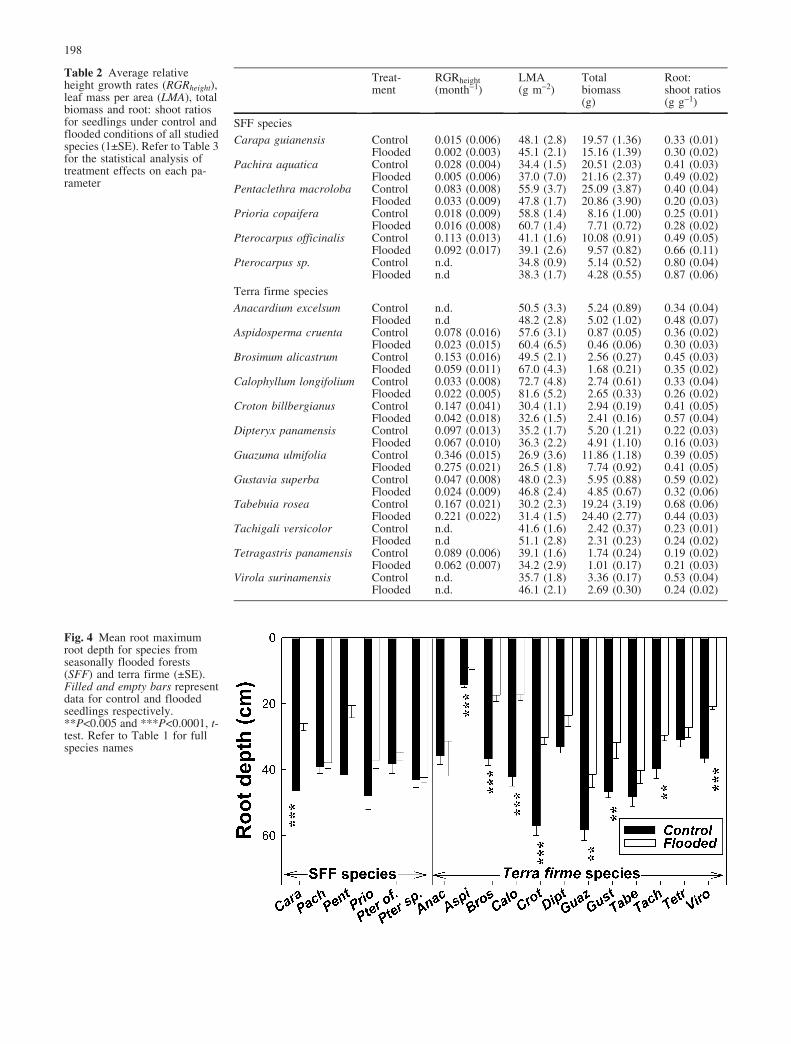
Table 2 Average relativeheight growth rates (RGRheight),leaf mass per area (LMA), totalbiomass and root: shoot ratiosfor seedlings under control andflooded conditions of all studiedspecies (1€SE). Refer to Table 3for the statistical analysis oftreatment effects on each pa-rameter
Treat-ment
RGRheight(month�1)
LMA(g m�2)
Totalbiomass(g)
Root:shoot ratios(g g�1)
SFF species
Carapa guianensis Control 0.015 (0.006) 48.1 (2.8) 19.57 (1.36) 0.33 (0.01)Flooded 0.002 (0.003) 45.1 (2.1) 15.16 (1.39) 0.30 (0.02)
Pachira aquatica Control 0.028 (0.004) 34.4 (1.5) 20.51 (2.03) 0.41 (0.03)Flooded 0.005 (0.006) 37.0 (7.0) 21.16 (2.37) 0.49 (0.02)
Pentaclethra macroloba Control 0.083 (0.008) 55.9 (3.7) 25.09 (3.87) 0.40 (0.04)Flooded 0.033 (0.009) 47.8 (1.7) 20.86 (3.90) 0.20 (0.03)
Prioria copaifera Control 0.018 (0.009) 58.8 (1.4) 8.16 (1.00) 0.25 (0.01)Flooded 0.016 (0.008) 60.7 (1.4) 7.71 (0.72) 0.28 (0.02)
Pterocarpus officinalis Control 0.113 (0.013) 41.1 (1.6) 10.08 (0.91) 0.49 (0.05)Flooded 0.092 (0.017) 39.1 (2.6) 9.57 (0.82) 0.66 (0.11)
Pterocarpus sp. Control n.d. 34.8 (0.9) 5.14 (0.52) 0.80 (0.04)Flooded n.d 38.3 (1.7) 4.28 (0.55) 0.87 (0.06)
Terra firme species
Anacardium excelsum Control n.d. 50.5 (3.3) 5.24 (0.89) 0.34 (0.04)Flooded n.d 48.2 (2.8) 5.02 (1.02) 0.48 (0.07)
Aspidosperma cruenta Control 0.078 (0.016) 57.6 (3.1) 0.87 (0.05) 0.36 (0.02)Flooded 0.023 (0.015) 60.4 (6.5) 0.46 (0.06) 0.30 (0.03)
Brosimum alicastrum Control 0.153 (0.016) 49.5 (2.1) 2.56 (0.27) 0.45 (0.03)Flooded 0.059 (0.011) 67.0 (4.3) 1.68 (0.21) 0.35 (0.02)
Calophyllum longifolium Control 0.033 (0.008) 72.7 (4.8) 2.74 (0.61) 0.33 (0.04)Flooded 0.022 (0.005) 81.6 (5.2) 2.65 (0.33) 0.26 (0.02)
Croton billbergianus Control 0.147 (0.041) 30.4 (1.1) 2.94 (0.19) 0.41 (0.05)Flooded 0.042 (0.018) 32.6 (1.5) 2.41 (0.16) 0.57 (0.04)
Dipteryx panamensis Control 0.097 (0.013) 35.2 (1.7) 5.20 (1.21) 0.22 (0.03)Flooded 0.067 (0.010) 36.3 (2.2) 4.91 (1.10) 0.16 (0.03)
Guazuma ulmifolia Control 0.346 (0.015) 26.9 (3.6) 11.86 (1.18) 0.39 (0.05)Flooded 0.275 (0.021) 26.5 (1.8) 7.74 (0.92) 0.41 (0.05)
Gustavia superba Control 0.047 (0.008) 48.0 (2.3) 5.95 (0.88) 0.59 (0.02)Flooded 0.024 (0.009) 46.8 (2.4) 4.85 (0.67) 0.32 (0.06)
Tabebuia rosea Control 0.167 (0.021) 30.2 (2.3) 19.24 (3.19) 0.68 (0.06)Flooded 0.221 (0.022) 31.4 (1.5) 24.40 (2.77) 0.44 (0.03)
Tachigali versicolor Control n.d. 41.6 (1.6) 2.42 (0.37) 0.23 (0.01)Flooded n.d 51.1 (2.8) 2.31 (0.23) 0.24 (0.02)
Tetragastris panamensis Control 0.089 (0.006) 39.1 (1.6) 1.74 (0.24) 0.19 (0.02)Flooded 0.062 (0.007) 34.2 (2.9) 1.01 (0.17) 0.21 (0.03)
Virola surinamensis Control n.d. 35.7 (1.8) 3.36 (0.17) 0.53 (0.04)Flooded n.d. 46.1 (2.1) 2.69 (0.30) 0.24 (0.02)
Fig. 4 Mean root maximumroot depth for species fromseasonally flooded forests(SFF) and terra firme (€SE).Filled and empty bars representdata for control and floodedseedlings respectively.**P<0.005 and ***P<0.0001, t-test. Refer to Table 1 for fullspecies names
198
hypertrophic lenticels below the water level and adven-titious roots were noted in Pachira, Anacardium andVirola. In contrast, after 90 days of flooding, nohypertrophic lenticels or adventitious roots were observedin seedlings of Prioria, Tetragastris and Aspidosperma.The remaining species formed some lenticels and adven-titious roots under flooding (Table 4).
Relative to control seedlings, stem diameter of floodedseedlings did not differ, except in Tabebuia and Aspido-sperma. After 90 days, flooded Tabebuia seedlingsshowed a 42% increase in stem diameter (Table 4), whileflooded seedlings of Aspidosperma had a 25% decrease instem diameter (Table 4).
Habitat effects, principal component analysisand species clustering
Seedling responses to flooding differed among speciestypically found in terra firme and those common to SFF(treatment by habitat, Table 5). Specifically, the treatmentby habitat interaction on root maximum depth and root:shoot ratios suggests crucial differences in how the rootsof species from the two habitats responded to flooding.However, within each habitat there also were significantdifferences among species. Across species there weretreatment effects on leaf area growth, RGRheight, stemdiameter, root dry weight and maximum root length, butnot on total biomass, shoot dry weight and root: shootratios (Table 5).
To evaluate whether flooding responses depended onhabitat preference (i.e. terra firme vs SFF) or lightrequirements (i.e. light-demanding vs shade-tolerant), we
conducted a PCA using the proportional change inflooded seedlings relative to controls for six variables:leaf area growth, stem height, total biomass, gst, root:shoot ratios and maximum root depth (Fig. 5). We did notinclude stem diameter and leaf number in the PCAbecause they were highly correlated with other growthparameters. PC axis 1 and 2, together accounted for 71%of the total variation. PC axis 1 has eigen values of 0.55for leaf area growth, 0.53 for stem height, and 0.52 fortotal biomass. PC axis 2 was dominated by root: shootratios and maximum root depth, with eigen values of 0.76and 0.57, respectively. The sixth parameter, gst at 30 days,had eigen values of �0.04 and �0.09 for PC axis 1 and 2,respectively, and did not help to distinguish amongspecies responses. In summary, the position along the firstPC axis relates to stem and whole plant growth underflooded conditions, while the position along the secondPC axis is related to root growth (Fig. 5).
In the PCA, species cluster into two main groups withTabebuia as an outlier (Fig. 5). The clusters are related toroot responses (PC axis 2) rather than growth parameters(PC axis 1). The cluster in the upper half of Fig. 5includes four SFF species Prioria, Pachira, Pterocarpussp. and Pterocarpus officinalis together with five terrafirme species Tetragastris, Croton, Guazuma, Tachigaliand Anacardium. The other cluster in the lower leftportion of Fig. 5 includes two SFF species Carapa andPentaclethra along with six terra firme species Dipteryx,Calophyllum, Virola, Gustavia, Aspidosperma and Brosi-mum. Although, no pattern was observed in relation tospecies’ light requirements, a gradient of flood toleranceamong the light-demanding species can be distinguished.In the upper portion of Fig. 5, Pterocarpus officinalis,
Table 3 Results from the MANOVAs (F-values) for differencesbetween treatments (flooding vs non-flooding), and the effect offlooding on relative height growth rates (RGRheight), leaf mass perarea (LMA), total biomass, root: shoot ratios (R:S ratios) and
maximum root depth (MRD,see Figs. 3. 4) of seedlings of allstudied species. Significant effects are denoted as *P<0.05,**P<0.01, ***P<0.001
Treatment RGRheight LMAF
BiomassF
R:S ratiosF
MRDF
df F df F
SFF species
Carapa guianensis 5, 12 2.81 1, 16 3.36 0.25 3.00 1.64 14.64***Pachira aquatica 5, 14 5.93** 1, 18 18.77*** 2.66 0.05 7.77 0.25Pentaclethra macroloba 5, 14 7.08*** 1, 18 12.55** 1.92 0.16 10.42** 6.25Prioria copaifera 5, 14 2.12 1, 18 0.01 1.06 0.13 2.66 5.34Pterocarpus officinalis 5, 13 1.27 1, 17 1.17 0.50 0.95 2.08 0.97Pterocarpus sp. 4, 15 2.16 1, 18 n.d 3.60 1.43 1.09 0.04
Terra firme species
Anacardium excelsum 4, 13 2.46 1, 16 n.d 0.27 0.02 5.45** 4.18Aspidosperma cruenta 5, 13 10.82*** 1, 17 5.94 0.11 22.09*** 3.34 27.89***Brosimum alicastrum 5, 14 10.36*** 1, 18 1.03 7.25 0.21 8.46** 39.72***Calophyllum longifolium 5, 14 13.06*** 1, 18 0.74 1.81 0.05 3.18 43.94***Croton billbergianus 5, 14 16.69*** 1, 18 12.74** 4.57 5.75** 5.33 63.77***Dipteryx panamensis 5, 14 1.48 1, 18 5.81 0.18 0.01 2.41 6.29Guazuma ulmifolia 5, 14 4.81** 1, 18 8.17** 0.01 7.56** 0.14 11.53**Gustavia superba 5, 14 4.74** 1, 18 3.71 0.14 1.08 20.22*** 9.54**Tabebuia rosea 5, 12 5.51** 1, 16 2.33 0.39 1.02 13.22** 2.96Tachigali versicolor 4, 15 6.53** 1, 18 n.d 9.75** 0.06 0.05 9.29**Tetragastris panamensis 5, 9 2.01 1, 13 7.66 1.40 2.06 0.02 0.29Virola surinamensis 4, 15 51.09*** 1, 18 n.d 30.70*** 4.09* 41.79*** 87.65***
199
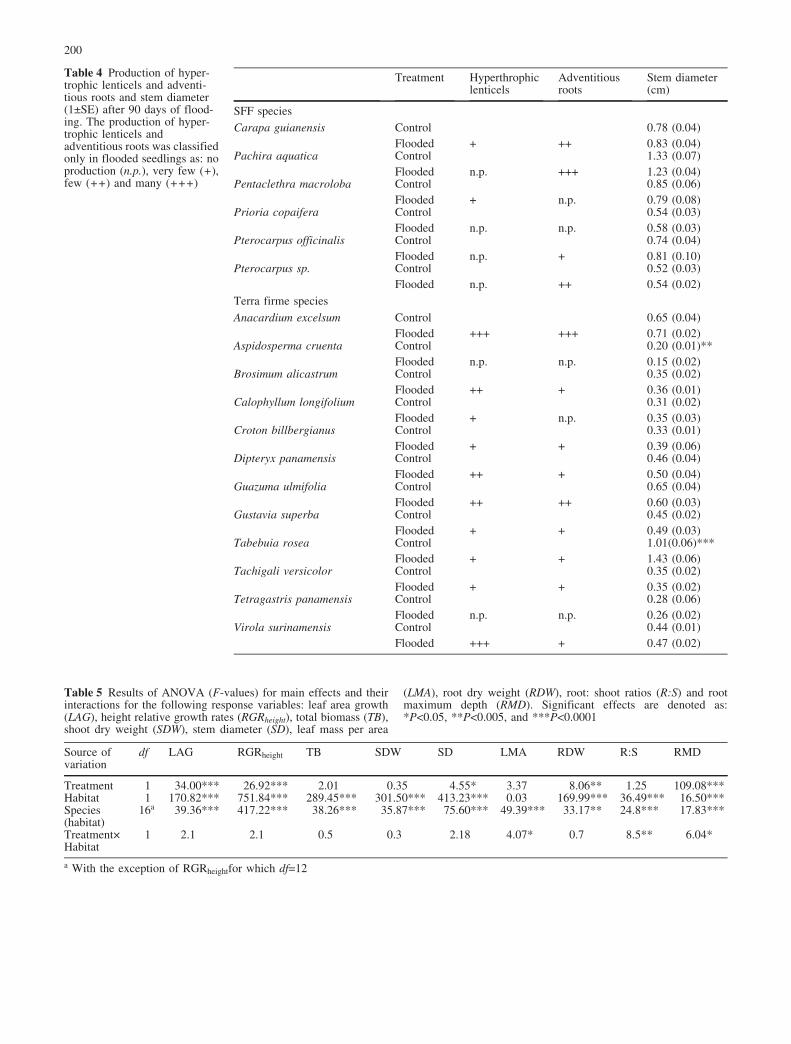
Table 4 Production of hyper-trophic lenticels and adventi-tious roots and stem diameter(1€SE) after 90 days of flood-ing. The production of hyper-trophic lenticels andadventitious roots was classifiedonly in flooded seedlings as: noproduction (n.p.), very few (+),few (++) and many (+++)
Treatment Hyperthrophiclenticels
Adventitiousroots
Stem diameter(cm)
SFF species
Carapa guianensis Control 0.78 (0.04)Flooded + ++ 0.83 (0.04)
Pachira aquatica Control 1.33 (0.07)Flooded n.p. +++ 1.23 (0.04)
Pentaclethra macroloba Control 0.85 (0.06)Flooded + n.p. 0.79 (0.08)
Prioria copaifera Control 0.54 (0.03)Flooded n.p. n.p. 0.58 (0.03)
Pterocarpus officinalis Control 0.74 (0.04)Flooded n.p. + 0.81 (0.10)
Pterocarpus sp. Control 0.52 (0.03)Flooded n.p. ++ 0.54 (0.02)
Terra firme species
Anacardium excelsum Control 0.65 (0.04)Flooded +++ +++ 0.71 (0.02)
Aspidosperma cruenta Control 0.20 (0.01)**Flooded n.p. n.p. 0.15 (0.02)
Brosimum alicastrum Control 0.35 (0.02)Flooded ++ + 0.36 (0.01)
Calophyllum longifolium Control 0.31 (0.02)Flooded + n.p. 0.35 (0.03)
Croton billbergianus Control 0.33 (0.01)Flooded + + 0.39 (0.06)
Dipteryx panamensis Control 0.46 (0.04)Flooded ++ + 0.50 (0.04)
Guazuma ulmifolia Control 0.65 (0.04)Flooded ++ ++ 0.60 (0.03)
Gustavia superba Control 0.45 (0.02)Flooded + + 0.49 (0.03)
Tabebuia rosea Control 1.01(0.06)***Flooded + + 1.43 (0.06)
Tachigali versicolor Control 0.35 (0.02)Flooded + + 0.35 (0.02)
Tetragastris panamensis Control 0.28 (0.06)Flooded n.p. n.p. 0.26 (0.02)
Virola surinamensis Control 0.44 (0.01)Flooded +++ + 0.47 (0.02)
Table 5 Results of ANOVA (F-values) for main effects and theirinteractions for the following response variables: leaf area growth(LAG), height relative growth rates (RGRheight), total biomass (TB),shoot dry weight (SDW), stem diameter (SD), leaf mass per area
(LMA), root dry weight (RDW), root: shoot ratios (R:S) and rootmaximum depth (RMD). Significant effects are denoted as:*P<0.05, **P<0.005, and ***P<0.0001
Source ofvariation
df LAG RGRheight TB SDW SD LMA RDW R:S RMD
Treatment 1 34.00*** 26.92*** 2.01 0.35 4.55* 3.37 8.06** 1.25 109.08***Habitat 1 170.82*** 751.84*** 289.45*** 301.50*** 413.23*** 0.03 169.99*** 36.49*** 16.50***Species(habitat)
16a 39.36*** 417.22*** 38.26*** 35.87*** 75.60*** 49.39*** 33.17** 24.8*** 17.83***
Treatment�Habitat
1 2.1 2.1 0.5 0.3 2.18 4.07* 0.7 8.5** 6.04*
a With the exception of RGRheightfor which df=12
200
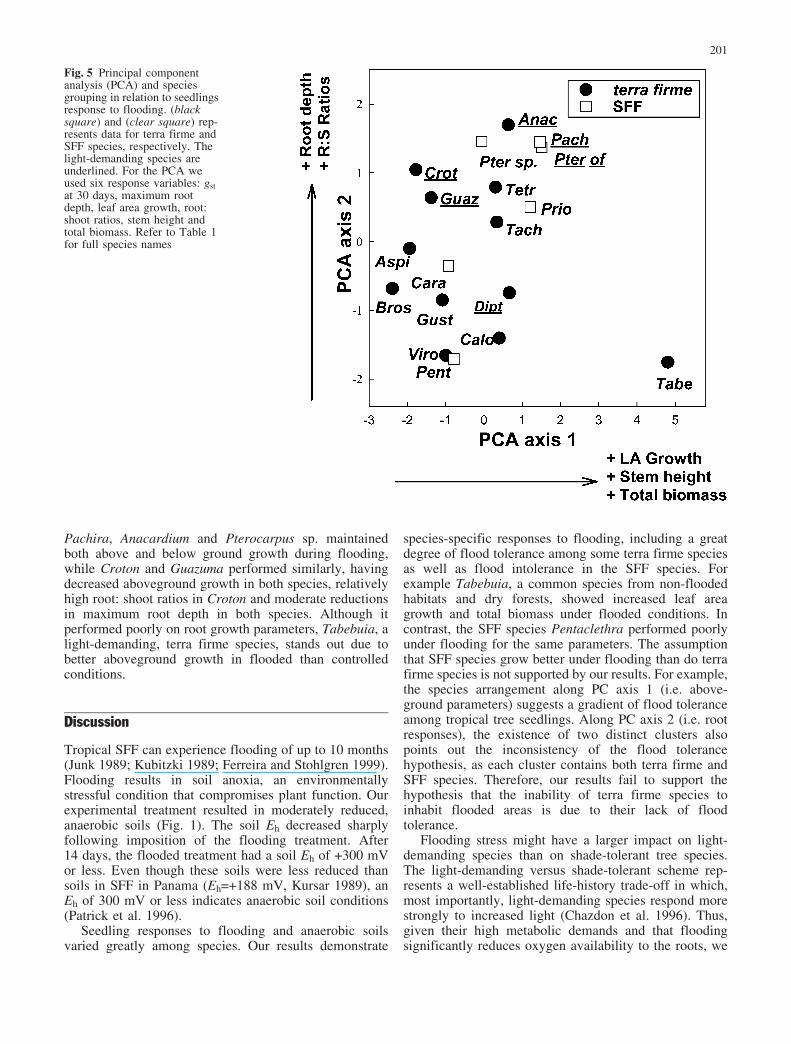
Pachira, Anacardium and Pterocarpus sp. maintainedboth above and below ground growth during flooding,while Croton and Guazuma performed similarly, havingdecreased aboveground growth in both species, relativelyhigh root: shoot ratios in Croton and moderate reductionsin maximum root depth in both species. Although itperformed poorly on root growth parameters, Tabebuia, alight-demanding, terra firme species, stands out due tobetter aboveground growth in flooded than controlledconditions.
Discussion
Tropical SFF can experience flooding of up to 10 months(Junk 1989; Kubitzki 1989; Ferreira and Stohlgren 1999).Flooding results in soil anoxia, an environmentallystressful condition that compromises plant function. Ourexperimental treatment resulted in moderately reduced,anaerobic soils (Fig. 1). The soil Eh decreased sharplyfollowing imposition of the flooding treatment. After14 days, the flooded treatment had a soil Eh of +300 mVor less. Even though these soils were less reduced thansoils in SFF in Panama (Eh=+188 mV, Kursar 1989), anEh of 300 mV or less indicates anaerobic soil conditions(Patrick et al. 1996).
Seedling responses to flooding and anaerobic soilsvaried greatly among species. Our results demonstrate
species-specific responses to flooding, including a greatdegree of flood tolerance among some terra firme speciesas well as flood intolerance in the SFF species. Forexample Tabebuia, a common species from non-floodedhabitats and dry forests, showed increased leaf areagrowth and total biomass under flooded conditions. Incontrast, the SFF species Pentaclethra performed poorlyunder flooding for the same parameters. The assumptionthat SFF species grow better under flooding than do terrafirme species is not supported by our results. For example,the species arrangement along PC axis 1 (i.e. above-ground parameters) suggests a gradient of flood toleranceamong tropical tree seedlings. Along PC axis 2 (i.e. rootresponses), the existence of two distinct clusters alsopoints out the inconsistency of the flood tolerancehypothesis, as each cluster contains both terra firme andSFF species. Therefore, our results fail to support thehypothesis that the inability of terra firme species toinhabit flooded areas is due to their lack of floodtolerance.
Flooding stress might have a larger impact on light-demanding species than on shade-tolerant tree species.The light-demanding versus shade-tolerant scheme rep-resents a well-established life-history trade-off in which,most importantly, light-demanding species respond morestrongly to increased light (Chazdon et al. 1996). Thus,given their high metabolic demands and that floodingsignificantly reduces oxygen availability to the roots, we
Fig. 5 Principal componentanalysis (PCA) and speciesgrouping in relation to seedlingsresponse to flooding. (blacksquare) and (clear square) rep-resents data for terra firme andSFF species, respectively. Thelight-demanding species areunderlined. For the PCA weused six response variables: gstat 30 days, maximum rootdepth, leaf area growth, root:shoot ratios, stem height andtotal biomass. Refer to Table 1for full species names
201

would predict light-demanding species to be moresusceptible to flooding than shade-tolerant species. How-ever, this was not the case in our study. Response of light-demanding species to flooding did not differ from that ofshade-tolerant species, though a gradient of toleranceamong the light-demanding species was apparent (seeFig. 5). Light-demanding SFF species Pterocarpus offi-cinalis and Pachira, along with the terra firme speciesAnacardium, performed relatively well under flooding inabove- and belowground parameters. Although, the twolight-demanding terra firme species, Croton and Guazu-ma, performed similarly, they experienced reducedaboveground growth but maintaining root growth.Dipteryx, another light-demanding terra firme speciesappears to maintain aboveground growth at the expense ofroot growth. Although no distinction between light-demanding and shade-tolerant species in relation toflooding exists, the susceptibility of light-demandingtropical species to flooding under high light regimesremains to be tested (see Collins and Battaglia 2002).
In the following sections, we examine how stomatalbehavior and the production of hypertrophic lenticels andadventitious roots relate to other measures of plantperformance under flooding. Reduction in gst and CO2assimilation is a common response to flooding (Pezeshki1994; Gravatt and Kirby 1998; Lopez and Kursar 1999).Flood sensitive species are characterized by a sharpreduction in gst immediately after exposure to low oxygenavailability (Pezeshki 1993; Gravatt and Kirby 1998). Forexample, seedlings of Quercus lyrata L. (Fagaceae), atemperate flood-tolerant species, showed a 54% reductionin gst after 3 days of flooding (Pezeshki 1993). In ourstudy, although large reductions in gst were observed after30 days, with the exception of Virola, we did not observea drop in gst at 12–15 days when soils became anaerobic.On average, after 15 days of flooding, terra firme specieshad a decrease in gst of 25%, with only Virola decreasingby more than 40%. Interestingly, after 5 days of floodingthe SFF species, Pterocarpus sp. and Pterocarpus offici-nalis showed a 53 and 40% decrease in gst, respectively.This suggests that even though SFF species thrive inflooded habitats, some of them experience a remarkabledegree of flooding stress.
Morphological adaptations expressed as the formationof hypertrophic lenticels, adventitious roots and increasedaerenchyma formation are frequent among flood-tolerantspecies. These adaptations minimize the effects offlooding on physiological processes at the leaf level suchas CO2 assimilation and gst in temperate species (Hook etal. 1970; Kozlowski 1984), as well as in some tropicalspecies (Nu�ez-Elisea et al. 1999). Only three of ourspecies did not produce visible morphological adaptationsand of the nine species with decreased gst under flooding,all but one had one of the indicated morphologicaladaptations. In the flooded plants, maintenance or areduction followed by recovery of gst was not related toproduction of lenticels or adventitious roots. Both Pachi-ra and Aspidosperma maintained similar gst values inflooded and control seedlings. While Pachira produced
adventitious roots, Aspidosperma produced neither ad-ventitious roots nor hypertrophic lenticels. Tabebuia hada third pattern; flooded seedlings produced adventitiousroots but failed to maintain a high gst. Our results suggestthat the relationship between the production of hypertro-phic lenticels or adventitious roots and the maintenance ofgst upon flooding is not straightforward. Rather, otherprocesses such as oxygen diffusion to the roots may bemore critical for maintenance of gst under flooding.Lastly, neither gst, hypertrophic lenticels nor adventitiousroots are closely correlated to flood tolerance.
Flooding generally reduces tree growth (Kozlowski1982). Among the studied species, the terra firme speciesTabebuia appears unique, as flooded seedlings had 25%more leaf area relative to control seedlings (Fig. 3).Flooding reduced leaf area to 30–60% of that of controlseedlings in all the other 11 terra firme species. Amongthe SFF species Prioria and Pterocarpus officinalisshowed no reduction in leaf area, while Carapa, Pachiraand Pentaclethra exhibited non-significant leaf areareduction of about 35% (Fig. 3). Consequently, we foundno treatment by habitat interaction for leaf area growth(Table 5). Flooding effect on RGRheight was slightlygreater in SFF than in terra firme species (Table 2);however no interaction effects were found (Table 5). Lackof treatment by habitat interaction among abovegroundresponses suggests that these parameters are not indica-tive of adaptations to tropical SFF habitats.
The reductions in gst, leaf area growth and RGRheight ofsome of the SFF species and the flood tolerance amongterra firme species suggest that current explanations forlow diversity in SFF should be reevaluated. If floodtolerance alone does not explain species diversity in SFF,what determines a species’ ability to thrive in inundatedhabitats? SFF are remarkably dynamic environmentscharacterized by alternating flooding and dry conditions(Golley et al. 1975; Junk 1989; Parolin 2001). Thus, inaddition to flooding, other environmental variables arelikely to influence species composition in SFF. Theability of seedlings to thrive in SFF may not rely on floodtolerance itself, but on a seedling’s tolerance or resistanceto drought. This is likely the case in SFF, since reductionsin root depth and root: shoot ratios due to flooding aremore likely to expose seedlings to drought stress underhigh evaporative demands during the dry season (Sena-Gomes and Kozlowski 1988; Lopez and Kursar 1999;Parolin 2001). Drought resistance associated with greaterroot depth or root biomass may be the primary physio-logical factor defining the species composition of habitatswith flooding followed by drought. Tyree et al. (1998)found no significant differences in xylem vulnerability tocavitation between non-flooded dipterocarp forests anddrought-prone heath (white sand) forests in Malaysia,suggesting that survival of occasional dry spells in eitherhabitat relies primarily on regulation of stomatal apertureand the development of deep roots.
In Panama, where the dry season can exert severewater stress in the plant community (Tobin et al. 1999),drought resistance is associated with greater root depth
202

(Rundel and Becker 1987; Wright 1992). Tobin et al.(1999) found that, during the dry season, the shallowrooted shrubs Psychotria horizontalis Sw. (Rubiaceae)and Hybanthus prunifolius (Schult.) Schultze (Violaceae)experienced midday water potentials as low as �4.5 and�3.4 MPa., respectively. In contrast, deeply rooted shrubsOuratea lucens (H.B.K.) Engler in Mart. and Swartziasimplex (Sw.) Spreng. (Caesalpinioideae) maintainedmidday water potentials above �1.3 MPa. Thus, after aperiod of flooding in a SFF, shallow-rooted seedlings maybe at a disadvantage in surviving a drought. Consistentwith this hypothesis is the observation that two of thethree parameters that showed treatment by habitat inter-actions were maximum root depth and root: shoot ratios(Table 5). Specifically, under flooding, rooting depthswere lowered by 32% in terra firme species and only 20%in SFF species. Similarly, root: shoot ratios were loweredby 10% in terra firme species but only 3% in SFF species.Hence the SFF species experienced less inhibition of rootgrowth, leading us to propose that inhibition of rootgrowth under flooding may be a good predictor ofabundance in SFF that also experience drought. Thisinterpretation is consistent with recent work suggestingthat post-flooding responses may play a very importantrole in determining a species’ distribution and abundancein flooded habitats (Crawford et al. 1994; Crawford andBraendle 1996). For example, although phenology ofAlchornea sidifolia Mll Arg. (Euphorbiaceae) trees didnot differ between flooded and non-flooded sites, cambialgrowth of waterlogged trees was restricted to the dryseason (Callado et al. 2001). Thus, the occurrence ofsome terra firme species in SFF might be related to theirability to exploit good growing conditions, i.e. mild dryseasons and not necessarily to flooding adaptations(Worbes et al. 1992).
Ecological factors such as seed dispersal also arecritical in determining species establishment and survivalin flooded forests. Variation in the onset of the floodingperiod may constrain the timing of seed dispersal andseedling establishment in SFF (Kubitzki and Ziburski1994; Parolin 2000). Lopez (2001) found that relative toSFF, the seeds of terra firme species are significantly lesscapable of floating and have lower germination followinglong-term flotation. In particular, seeds of Tabebuiarosea, the only terra firme species that performed betterunder flooded conditions, are incapable of floating morethan 10 days (Lopez 2001). Even though the physiologyof Tabebuia seedlings may be compatible with the SFFabiotic environment, it may fail to disperse effectivelyinto SFF.
We conclude that the ability of a species to thrive onSFF might be better explained by a combination ofadaptive strategies. First, flood tolerance is a complextrait involving more than one adaptation. While floodtolerance is crucial for survival in flood-prone habitats,responses to post-flooding events might be equallyimportant. The ability to grow roots under anoxia andwater stress resistance is probably critical for survival inSFF. Species-specific differences in root growth during
flooding (i.e. root: shoot ratios, maximum root depth) alsomay predict drought resistance in habitats that alsoexperience a strong dry season. Lastly, establishment inSFF also is determined by the ability of a species todisperse seeds and germinate after being in water for longperiods of time. Therefore, we propose that examining thespatio-temporal variation in flooding and drought of SFFmight prove critical for understanding the low speciesdiversity in these habitats, as relatively few species mayhave the complex suite of traits necessary to disperse andestablish under flood-drought cycles.
Acknowledgements We would like to thank the SmithsonianTropical Research Institute for its excellent logistical support atBCI, Panama. Lissy Coley, Krista Farris Lopez, Jeff Dukes, RickCondit, David Mason, R.M.M Crawford and an anonymousreviewer provided valuable comments on the manuscript. EdmundoAyarza, Dora Alvarez, Blexein Contreras, Delfin Jaramillo and BillGrauel provided assistance in Dari�n and at BCI. This research wassupported by a NSF Dissertation Improvement Grant (IBN-9902211 to O.L. and T.A.K.), a grant from the InternationalTropical Timber Organization to the Autoridad Nacional delAmbiente of the Republic of Panama and a doctoral fellowshipfrom the Panamanian National Secretariat for Science and Tech-nology (SENACYT).
References
Andrade ACS, Ramos FN, De Souza AF, Loureiro MB, Bastos R(1999) Flooding effects in seedlings of Cytharexylum myri-anthum Cham. and Genipa americana L.: responses of twoneotropical lowland tree species. Rev Bras Bot [Supl] 22:281–285
Berry PE, Holst B, Yatskievych K (1995) Flora of the VenezuelanGuyana, vol 1. Introduction. Missouri Botanical Garden,Timber Press, Portland, Ore.
Black GA, Dobzhansky T, Pavan C (1950) Some attempts toestimate species diversity and population density of trees inAmazonian forests. Bot Gaz 111:413–425
Callado CH, Neto SJD, Scarano FR, Barros CF, Costa CG (2001)Periodicity of growth rings in some flood-prone trees of theAtlantic Rain Forests in Rio de Janeiro, Brazil. Trees 15:492–497
Campbell DG, Stone JL, Rosas A Jr (1992) A comparison of thephytosociology and dynamics of three (vrzea) forests ofknown ages, Rio Juru, Western Brazilian Amazon. Bot J LinnSoc 108:213–237
Chazdon RL, Pearcy RW, Lee DW, Fetcher N (1996) Photosyn-thetic responses of tropical forest plants to contrasting lightenvironments. In: Mulkey SS, Chazdon RL, Smith AP (eds)Tropical forest plant ecophysiology. Chapman and Hall, NewYork, pp 5–55
Cobin RJ, Mitchell RJ (2000) To Bonferroni or not to Bonferroni:when and how are the questions. Bull Ecol Soc Am 81:246–248
Collins BS, Battaglia LL (2002) Microenvironmental heterogeneityand Quercus michauxii regeneration in experimental gaps. ForEcol Manage 155:279–290
Condit R, Hubbell SP, Foster RB (1993) Mortality and growth of acommercial hardwood “el cativo”, Prioria copaifera, inPanama. For Ecol Manage 63:107–112
Condit R, Hubbell SP, Foster RB (1996) Changes in tree speciesabundance in a Neotropical forest: impact of climate change. JTrop Ecol 12:231–256
Crawford RMM, Braendle R (1996) Oxygen deprivation stress in achanging environment. J Exp Bot 47:145–159
Crawford RMM, Walton JC, Wollenweber-Ratzer B (1994)Similarities between post-ischaemic injury to animal tissues
203

and post-anoxic injury in plants. Proc R Soc Edinb 102B:325–332
Croat TB (1978) Flora of Barro Colorado Island. StanfordUniversity Press, Palo Alto, Calif.
D’Arcy WG (1987) Flora of Panama checklist. Missouri BotanicalGarden, St. Louis, Mo.
Davis TAW, Richards PW (1933) The vegetation of MorabilliCreek, British Guiana: an ecological study of a limited area oftropical rain forest. Part II. J Ecol 22:106–155
Duivenvoorden JF (1996) Patterns of tree species richness in rainforest of middle Caquet area, Colombia, NW Amazonia.Biotropica 28:142-158
Fernndez AR, Laffarga JM, Ortega F (1993) Strategies inMediterranean grassland annuals in relation to stress anddisturbance. J Veg Sci 4:313–322
Ferreira LV, Stohlgren TJ (1999) Effects of river level fluctuationon plant richness, diversity and distribution in a floodplainforest in Central Amazonia. Oecologia 120:582–587
Golley FB, McGinnis JT, Clements RG, Child GI, Duever MJ(1975) Mineral cycling in a tropical moist forest ecosystem.University of Georgia Press, Athens, Ga.
Golluscio RA, Sala OE (1993) Plant functional types and ecolog-ical strategies in Patagonian forbs. J Veg Sci 6:99–105
Grauel W, Kursar TA (1999) Species diversity and stand dynamicsof Cativo (Prioria copaifera, Griseb.) forests in Darienprovince, Panama. In: Kleinn C, Kohl M (eds) Proceedings ofthe IUFRO S4.11 International Symposium: Long-term obser-vations and research in forestry. CATIE, Costa Rica, pp 69–77
Gravatt DA, Kirby CJ (1998) Patterns of photosynthesis and starchallocation in seedlings of four bottomland hardwood treespecies subjected to flooding. Tree Physiol 18:411–417
Hart TB (1990) Monospecific dominance in tropical rain forest.Trends Ecol Evol 5:6-10
Hartshorn GS (1972) The ecological life history and populationdynamics of Pentaclethra macroloba a tropical wet forestdominant, and Stryphnodendron excelsum, an ocassional asso-ciate. Ph.D. Dissertation, University of Washington
Hook DD, Langdon OG, Stubbs J, Brown CL (1970) Effect ofwater regimes on the survival, growth and morphology oftupelo seedlings. For Sci 16:304-311
Janzen DH (1974) Tropical blackwater rivers, animals, and mastfruiting by the Dipterocarpaceae. Biotropica 6:69-103
Jim�nez JA (1994) Los manglares del Pac�fico centroamericano.Editorial Fundaci�n UNA, Heredia, Costa Rica
Joly CA, Crawford RMM (1982) Variation in tolerance andmetabolic responses to flooding in some tropical trees. J ExpBot 33:799-809
Junk WJ (1989) Flood tolerance and tree distribution in centralAmazonian floodplains. In: Holm-Nielsen LB, Nielsen IC,Baslev H (eds) Tropical forests. Botanical dynamics, speciationand diversity. Academic Press, San Diego, Calif., pp 45-64
Kozlowski TT (1982) Water supply and tree growth. Part II.Flooding. For Abst 43:145–161
Kozlowski TT (1984). Responses of woody plants to flooding. In:Kozlowski TT (ed) Flooding and plant growth. AcademicPress, Orlando, Fla., pp 129-164
Kubitzki K (1989) The ecogeographical differentiation of Amazo-nian inundation forests. Plant Syst Evol 162:285–304
Kubitzki K, Ziburski A (1994) Seed dispersal in floodplain forest ofAmazonia. Biotropica 26:30–43
Kursar TA (1989) Evaluation of soil respiration and soil CO2concentration in a lowland moist forest in Panama. Plant Soil113:21–29
Leigh EG Jr, Rand AS, Windsor DM (1996) The ecology of atropical forest: seasonal changes and long term changes, 2ndedn. Smithsonian Institution, Washington, D.C.
Linares PR, Mart�nez H (1991) La regeneraci�n natural tempranadel bosque de Cativo en Choc�-Colombia. Corporaci�nNacional de Investigaci�n y Fomento Forestal (CONIF),Bogot, Colombia. Serie T�cnica 30, pp 1–27
Lopez OR (2001) Seed flotation and post-flooding germination intropical terra firme and seasonally flooded forest species. FunctEcol 15:763–771
Lopez OR, Kursar TA (1999) Flood tolerance of four tropical treespecies. Tree Physiol 19:925–932
Mayo-Mel�ndez E (1965) Algunas caracter�sticas ecol�gicas de losbosques inundables del Dari�n, Panam, con miras a su posibleutilizaci�n. Turrialba 14:336–347
Nu�ez-Elisea R, Schaffer B, Fisher JB, Colls AM, Crane JH (1999)Influence of flooding on net CO2 assimilation, growth and stemanatomy of Annona species. Ann Bot 84:771–780
Parolin P (2000) Seed mass in Amazonian floodplain forests withcontrasting nutrient supplies. J Trop Ecol 16:417–428
Parolin P (2001) Morphological and physiological adjustments towaterlogging and drought in seedlings of Amazonian floodplaintrees. Oecologia 128:326–335
Patrick WH, Gambrell RP, Faulkner SP (1996) Redox measure-ments of soils. In: Methods in Soil Analysis Part III, ChemicalMethods. Soil Science Society of America/American Society ofAgronomy, Madison, Wis., pp1255–1273
Peters CM, Balick MJ, Kahn F, Anderson AB (1989) Oligarchicforest of economic plants in Amazonia: utilization and conser-vation of an important tropical resource. Conserv Biol 3:341–349
Pezeshki SR (1993) Differences in patterns of photosyntheticresponses to hypoxia in flood-tolerant and flood-sensitive treespecies. Photosynthetica 28:423–430
Pezeshki SR (1994) Plant responses to flooding. In: Wilkinson RE(ed) Plant environment interactions. Dekker, New York, pp289–321
Richards PW (1996) The tropical rain forest: an ecological study,2nd edn. Cambridge University Press, London
Rundel PW, Becker PF (1987) Cambios estacionales en lasrelaciones h�dricas y en la fenolog�a vegetativa de plantas delestrato bajo del bosque tropical de la Isla de Barro Colorado,Panam. Rev Biol Trop 35:71–84
Sena-Gomes AR, Kozlowski TT (1988) Physiological and growthresponses to flooding of seedlings of Hevea brasiliensis.Biotropica 20:286–293
Sokal RR, Rohlf FJ (1995) Biometry, 2nd edn. Freeman, New YorkSork VL (1985) Germination responses in a large-seeded Neotrop-
ical tree species, Gustavia superba (Lecythidaceae). Biotropica17:130–136
Tobin MF, Lopez OR, Kursar TA (1999) Responses of tropicalunderstory plants to a severe drought: tolerance and avoidanceof water stress. Biotropica 31:570–578
Tyree MT, Pati�o S, Becker P (1998) Vulnerability to drought-induced embolism of Bornean heath and dipterocarp foresttrees. Tree Physiol 18:583–588
Welden CW, Hewett SW, Hubbell SP, Foster RB (1991) Saplingsurvival, growth, and recruitment: relationship to canopy heightin a neotropical forest. Ecology 72:35–50
Windsor DM (1990) Climate and moisture variability in a tropicalforest: long-term records from Barro Colorado Island, Panama.Smithson Contrib Earth Sci 29:1–145
Worbes M, Klinge H, Revilla JD, Martius C (1992) On thedynamics, floristics subdivision and geographical distributionof vrzea forests in Central Amazonia. J Veg Sci 3:553–564
Wright SJ (1992) Seasonal drought, soil fertility and the speciesdensity of tropical forest plant communities. Trends Ecol Evol7:260–263
204
Copyright © 2022 FDOKUMEN




















