
Plant zonation in irregularly flooded salt marshes - Repositório ...
15
Journal of Ecology 2003 91, 951– 965 © 2003 British Ecological Society Blackwell Publishing Ltd. Plant zonation in irregularly flooded salt marshes: relative importance of stress tolerance and biological interactions CÉSAR S. B. COSTA, JULIANO C. MARANGONI and ADRIANA M. G. AZEVEDO Departamento de Oceanografia, Fundação Universidade Federal do Rio Grande (FURG), CP 474, Rio Grande, RS, 96200 – 901, Brazil Summary 1 Most studies of salt marsh plant zonation have been at middle to high latitudes of the northern hemisphere, in euhaline or periodically hypersaline marshes with regular tides. In this study, we examined plant zonation in an irregularly flooded marsh in southern Brazil. Pore water characteristics were compared in four vegetation zones across a marsh elevation gradient. Reciprocal transplants between vegetation zones and removal experiments were performed to examine species interactions in low and mid marshes. 2 There was no distinctive gradient of physical stress across the elevation of irregularly flooded low and mid marshes. Moreover, the three dominant plants, Spartina alterni- flora, Spartina densiflora and Scirpus maritimus, were able to grow across the entire ele- vation gradient, i.e. within zones normally occupied by the other species. The only exception was Spartina alterniflora, which was strongly limited by selective herbivory by the crab Chasmagnathus granulata in the Scirpus maritimus zone. 3 Although intra- and interspecific competition reduced growth of all three species, no competitive hierarchy was found in any vegetation zone. 4 These results suggest that, as in tidal marshes of the north hemisphere, competition is important in structuring salt marsh plant communities. In contrast, however, plant zonation in irregularly flooded marshes cannot be explained by displacement of com- petitive subordinates to physically stressful habitats. The roles of founder effects and selective herbivory in such marshes therefore merits further investigation. Key-words: competition, Grapsidae crab, herbivory, non-tidal marsh Journal of Ecology (2003) 91, 951– 965 Introduction Well-developed coastal salt marshes display a zonation of plant species parallel to the shore. Several studies of salt marsh zonation and community structure have concentrated on the role of the physico-chemical effects of inundation with seawater (Ranwell 1974; Pielou & Routledge 1976; Jefferies 1977; Jefferies et al . 1979; Vince & Snow 1984; Rozema et al . 1988; Zedler et al . 1999; Rogel et al . 2000). There can be no doubt that elevation and salinity are major determinants of distribution per se and also that physico-chemical factors affect the out- come of any biotic interactions (Pennings & Callaway 1992; Callaway & Pennings 2000; Davy et al . 2000; Emery et al . 2001). Understanding zonation requires consideration of a realistic range of environmental variability and evaluation of biotic interactions and is therefore likely to involve field experiments. Most experimental field studies of zonation in salt marsh plants have been conducted at middle and high latitudes of the northern hemisphere and in regularly tidal marshes (Jefferies 1977; Snow & Vince 1984; Scholten et al . 1987; Rozema et al . 1988; Pennings & Bertness 1999; van Wijnen & Bakker 1999; Castillo et al . 2000; Davy et al . 2000; Pennings & Bertness 2001; Pennings & Moore 2001). These studies point to trade-offs between stress tolerance and competitive ability and a competitive displacement of subordinate species to physically stressful habitats (Snow & Vince 1984; Rozema et al . 1988; Bertness 1991a, 1991b; Pennings & Callaway 1992; Bertness & Callaway 1994; Bertness 1999; Pennings & Moore 2001). Nutrient availability (Jefferies 1977; Rozema et al . 1988; Levine et al . 1998; van Wijnen & Bakker 1999; Emery et al . 2001), perturbation by wrack (Bertness & Ellison 1987; Bertness 1999), ice damage (Pennings & Bertness Correspondence: C. S. B. Costa (fax +55 532336602; e-mail [email protected]).
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Plant zonation in irregularly flooded salt marshes - Repositório ...

Journal of Ecology
2003
91
951ndash965
copy 2003 British Ecological Society
Blackwell Publishing Ltd
Plant zonation in irregularly flooded salt marshes relative importance of stress tolerance and biological interactions
CEacuteSAR S B COSTA JULIANO C MARANGONI and ADRIANA M G AZEVEDO
Departamento de Oceanografia Fundaccedilatildeo Universidade Federal do Rio Grande (FURG) CP 474 Rio Grande RS 96200ndash901 Brazil
Summary
1
Most studies of salt marsh plant zonation have been at middle to high latitudes ofthe northern hemisphere in euhaline or periodically hypersaline marshes with regulartides In this study we examined plant zonation in an irregularly flooded marsh insouthern Brazil Pore water characteristics were compared in four vegetation zonesacross a marsh elevation gradient Reciprocal transplants between vegetation zones andremoval experiments were performed to examine species interactions in low and midmarshes
2
There was no distinctive gradient of physical stress across the elevation of irregularlyflooded low and mid marshes Moreover the three dominant plants
Spartina alterni-flora
Spartina densiflora
and
Scirpus maritimus
were able to grow across the entire ele-vation gradient ie within zones normally occupied by the other species The onlyexception was
Spartina alterniflora
which was strongly limited by selective herbivory bythe crab
Chasmagnathus granulata
in the
Scirpus maritimus
zone
3
Although intra- and interspecific competition reduced growth of all three species nocompetitive hierarchy was found in any vegetation zone
4
These results suggest that as in tidal marshes of the north hemisphere competitionis important in structuring salt marsh plant communities In contrast however plantzonation in irregularly flooded marshes cannot be explained by displacement of com-petitive subordinates to physically stressful habitats The roles of founder effects andselective herbivory in such marshes therefore merits further investigation
Key-words
competition Grapsidae crab herbivory non-tidal marsh
Journal of Ecology
(2003)
91
951ndash965
Introduction
Well-developed coastal salt marshes display a zonationof plant species parallel to the shore Several studies ofsalt marsh zonation and community structure haveconcentrated on the role of the physico-chemical effectsof inundation with seawater (Ranwell 1974 Pielou ampRoutledge 1976 Jefferies 1977 Jefferies
et al
1979 Vinceamp Snow 1984 Rozema
et al
1988 Zedler
et al
1999Rogel
et al
2000) There can be no doubt that elevationand salinity are major determinants of distribution
perse
and also that physico-chemical factors affect the out-come of any biotic interactions (Pennings amp Callaway1992 Callaway amp Pennings 2000 Davy
et al
2000Emery
et al
2001) Understanding zonation requiresconsideration of a realistic range of environmental
variability and evaluation of biotic interactions and istherefore likely to involve field experiments
Most experimental field studies of zonation in saltmarsh plants have been conducted at middle and highlatitudes of the northern hemisphere and in regularlytidal marshes (Jefferies 1977 Snow amp Vince 1984 Scholten
et al
1987 Rozema
et al
1988 Pennings amp Bertness 1999van Wijnen amp Bakker 1999 Castillo
et al
2000 Davy
et al
2000 Pennings amp Bertness 2001 Pennings amp Moore2001) These studies point to trade-offs between stresstolerance and competitive ability and a competitivedisplacement of subordinate species to physically stressfulhabitats (Snow amp Vince 1984 Rozema
et al
1988 Bertness1991a 1991b Pennings amp Callaway 1992 Bertness ampCallaway 1994 Bertness 1999 Pennings amp Moore 2001)Nutrient availability (Jefferies 1977 Rozema
et al
1988Levine
et al
1998 van Wijnen amp Bakker 1999 Emery
et al
2001) perturbation by wrack (Bertness amp Ellison1987 Bertness 1999) ice damage (Pennings amp Bertness
Correspondence C S B Costa (fax +55 532336602 e-maildocostafurgbr)
952
C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society
Journal of Ecology
91
951ndash965
2001) and herbivory (Cargill amp Jefferies 1984 Iribarne
et al
1997 Taylor
et al
1997 Bortolus amp Iribarne 1999)as well as positive interactions (eg plant cover allevi-ating saline stress) (Bertness amp Callaway 1994 Bertnessamp Leonard 1997 Hacker amp Bertness 1999 Callawayamp Pennings 2000) may also be locally important inshaping salt marsh plant communities
It is unclear how far this can be extrapolated tomicrotidal and irregularly flooded salt marshes In theseecosystems flooding patterns are primarily driven byvariation in rainfall and wind direction (Eleuterius ampEleuterius 1979 Stout 1984 Costa
et al
1988 Christian
et al
1990 Wang 1997 Holm
et al
2000 Moumlller
et al
2001) and both salinity and the timing and duration offlooding are thus less predictable In contrast to tidalsalt marshes where dominant species tend to be restrictedto specific zones with little habitat overlap (Pielou ampRoutledge 1976 Russell
et al
1985 Bertness 1999)irregularly flooded salt marshes have perennial domin-ants growing across wide ranges of salinity and waterlevel with considerable habitat overlap (Ranwell 1974Eleuterius amp Eleuterius 1979 Odum 1988 Baldwin ampMendelssohn 1998)
Salt marshes occupy 70 km
2
of the intertidal zone ofthe estuarine region of Patos Lagoon in southern Bra-zil (Costa
et al
1997) Diurnal tidal range is very small(astronomical tides less than 05 m) and fluctuations ofwater level and salinity are irregular (Costa
et al
1988Moumlller
et al
2001) The native grasses
Spartina alterni-flora
Lois and
Spartina densiflora
Brong tend to dom-inate mesohaline low- (between
minus
50 cm and +10 cmMWL mean water level of the estuary) and mid marshes(+10 to +30 cm MWL) respectively (Silva
et al
1993Costa 1997a 1997b) while highly productive stands of
Scirpus maritimus
L may opportunistically dominatetransitional areas between low and mid marshes (Costa1998) These dominant species occur over a wide ver-tical range with a high degree of overlap (Costa 1997a)and monospecific stands of all three can often be foundat the same tidal level The local occurrence of such dis-crete patches suggests that interspecific competitionplays an important role in the structure and composi-tion of these communities The herbivorous Grapsidaecrab
Chasmagnathus granulata
Dana 1851 which is oneof the most abundant macroinvertebrates in the saltmarshes along the coasts of southern Brazil and north-east Argentina is found at high density in the intertidalzone (DrsquoIncao
et al
1990 Copertino
et al
1997) feed-ing on
Spartina
spp (DrsquoIncao
et al
1990 Iribarne
et al
1997 Bortolus amp Iribarne 1999 Bortolus
et al
2002)The relative importance of stress tolerance competi-tion and herbivory in the distribution patterns of thesedominant plants however has yet to be examined Suchecological information is particularly important because
S densiflora
has been introduced unintentionally to thecoasts of California (Kittelson amp Boyd 1997) and south-western Spain (Figueroa amp Castellanos 1988 Castillo
et al
2000) where it has been displacing indigenouslow marsh dominants
We use methods identical to those applied in thenorth hemisphere (reciprocal transplantation betweenvegetated zones and removal of neighbours) to test thehypotheses that (i) distributions of the dominantspecies (
Spartina densiflora
Spartina alterniflora
and
Scirpus maritimus
) on irregularly flooded low to mid marshzones in southern Brazil are related to their competitiveperformance and (ii) stress tolerance has a minor rolein zonation of marshes with poorly predictable hydro-logy We therefore tested the applicability of communitystructuring rules based on work in northern-hemispheresalt marshes to marshes with no distinct tidal regimeWe also examined whether selective grazing by
Chas-magnathus granulata
could affect the performance ofdominant plants and thus community structure
Materials and methods
A
c
10 000 km
2
Patos Lagoon located in southernBrazil (30ndash32
deg
S) is the wordrsquos largest choked lagoon(ie characterized by a single narrow but permanentlyopen entrance channel) It receives runoff from the5500 km
2
Mirim Lagoon near the BrazilndashUruguay border(Moumlller
et al
2001 Seeliger amp Costa 2002) forming alarge lagoon complex that only exchanges water withthe Atlantic Ocean through the Patos Lagoon estuaryan inlet 20 km long and 05ndash30 km wide
The study site was located on the north-west side ofPoacutelvora Island (32
deg
01
prime
S 52
deg
06
prime
W) Rio Grande (RSBrazil) Poacutelvora Island is completely covered by 45 haof undisturbed estuarine salt marsh (Costa 1997a) Lowmarshes are mostly colonized by
Spartina alterniflora
while mid marshes are dominated by either
Spartinadensiflora
or
Scirpus maritimus
and a shrub zone domin-ated by
Myrsine parvifolia
ADC and the halophyticfern
Acrosticum danaefolium
Langsd amp Fish occur onthe high marsh Monospecific stands of
S maritimus
and
S densiflora
are also found colonizing mud flats ofthe low marsh The climate is hot-temperate with clearseasonal fluctuations lowest (13
deg
C) and highest (25
deg
C)average air temperatures are observed in July andJanuary respectively Marsh dominants show specificseasonal growth cycles
S maritimus
behaves as a crypto-phyte with aerial parts sprouting from its rhizomes atthe end of winter peak growth at the end of spring andtillers completely dying back in mid-autumn (Seeliger
et al
1997 Costa 1998) Both
Spartina
species are C4phanerophytes with slow winterndashspring growth andgrowth peaks in the summer and autumn (Silva
et al
1993 Copertino
et al
1997 Seeliger
et al
1997)
The variability of water level in the Patos Lagoon estu-ary was assessed by comparing flooding frequencies inthe intertidal zone between years (each annual periodlasts from July until the following June) Water level
953
Irregularly flooded marsh
copy 2003 British Ecological Society
Journal of Ecology
91
951ndash965
and salinity have been monitored daily since July 1992at the southern part of Marinheiro Island (1 km northof the study area) Water level was determined to thenearest cm between 1000 and 1100 h at a fixed stationand related to the mean water level (MWL) of the estuarybetween July 1992 and June 2001
To document variation in physical stress within veg-etation zones at Poacutelvora Island pore water salinity pHand redox potential of the surface sediment (10 cm) weremeasured at 10 sampling points along a 410-m transectThe pore water samples were obtained between Apriland August 1995 (
n
= 10 dates) with a hand vacuumpump (model 2005G2 Soilmoisture Equip CorpSanta Barbara CA USA) by inserting the bottomlesscollection cylinder 10 cm into the sediment Thesampling points corresponded to zones dominanted by
Spartina alterniflora
(points 1 and 2)
S densiflora
(34 5 and 9)
Scirpus maritimus
(6 7 and 8) and high marshshrubs (
Myrsine parvifolia
point 10) respectively InJanuary 2000 three cores (top 10 cm) were randomlycollected in the low and mid marsh vegetation zonesThe sediment was air-dried and passed through a 2-mmsieve before laboratory analysis The total organiccontent was estimated by combustion at 500
deg
C for6 h (Brower amp Zar 1977) Total Kjedahl nitrogenconcentration in the sediment was determined and theconcentration of exchangeable phosphate estimated bycolourimetry (Allen
et al
1976)
Transplants were established in June 1995 Three adja-cent monospecific stands of
Spartina alterniflora
Sdensiflora
and
Scirpus maritimus
(hereafter called lsquozonesrsquo)each covering
c
2500 m
2
were chosen for the experi-ment The elevation of each zone was estimated with alevel and express relative to the water-level monitoringsite to allow us to estimate their flooding frequencies(percentage of days) Thirty blocks of sediment of 20
times
20
times
20 cm including tillers rhizomes and roots ofeach species (hereafter called lsquoturfsrsquo) were excavatedfrom each of the monospecific zones Ten were trans-planted to similar-sized holes in each of the true zones(ie manipulation controls) Ten blocks were placed inthe other species zones In each zone half of the trans-plants (
n
= 5) of each species were placed in direct con-tact with the surrounding vegetation and the other half(
n
= 5) had all neighbouring plants in a 25-cm widestrip around the perimeter removed by clipping theirshoots at ground level All biological measurementswere made within the 0031-m
2
central areas of the turfsdelimited by a PVC ring with 20 cm diameter fixed tothe sediment with metal pins To avoid invasion by rootsof surrounding plants all neighbour-removal areas werecut to a depth of 25 cm with a spade 25 cm outside theirperimeter The clipping and severing were maintainedfor several weeks until neighbours ceased to grow backinto the neighbour-removal area This methodologyallowed us to account for the overall competitive effect
of neighbouring vegetation and the impact of localphysical stresses on target transplanted plants (Bertness1991a 1991b Pennings amp Callaway 1992 Emery
et al
2001 Pennings amp Moore 2001 Pennings
et al
2003)Five times during the period of study (31 August 1995
26 October 1995 24 November 1995 30 January 1996and 27 February 1996 hereafter referred to as lsquolate winter1995rsquo lsquoearly spring 1995rsquo lsquolate spring 1995rsquo lsquoearly summer1996rsquo and lsquomid-summer 1996rsquo respectively) the numberof live tillers reproductive tillers (with flowersseeds)and tillers with crab wounds from
Chasmagnathusgranulata
were determined in each turf A tiller wasconsidered grazed when it had small scratches or part orall of the tiller had been removed The small Grapsid crab
Metasesarma rubripes
Rathbun (1897) (Capitoli
et al
1977) was also observed in the local marsh but it feedsmainly on plant detritus Insect herbivory was negligibleand appeared uniform for all three plant species
The final harvest of all tillers inside PVC rings wasdone by clipping at ground level at the growth peak ofeach species Because of the early growing season of
Smaritimus
(Costa 1998) it was harvested in early summer1996 whereas the two perennial grasses (
S alterniflora
and
S densiflora
) were collected in mid-summer 1996In the laboratory the heights of all tillers were meas-ured and after removing dead plant material dried at80
deg
C for 48 h and weighed Between-treatment differ-ences in height were analysed using only the 10 tallesttillers per turf of
S densiflora
and the five tallest tillersper turf of
S alterniflora
and
S maritimus
(where standsare less dense due to their architecture)
Maximumtiller height was used as indicator of the growth poten-tial of each species under the experimental conditionsas it excludes bias in the average height due to between-turf variation of tiller density and recently sprouted till-ers The percentage of flowering tillers was calculatedincluding only tillers of height equal to or taller thanthe minimum height from which flowering wasobserved (
S densiflora
= 10 cm
S alterniflora
= 45 cm
S maritimus
= 45 cm) Additionally six plants of eachspecies were collected from their natural zones and theKjeldahl N concentration of the leaves was determined(Allen
et al
1976)
Because of the great temporal variability in pore watersalinity pH and redox the assumptions of homogene-ity of variances and normality were not met with the usualtransformations The KruskalndashWallis test was used totest for differences between sampling points and betweenvegetation zones (ie sampling points grouped by localplant cover) at the 5 significance level (Siegel 1975)When significant differences were found for the KruskalndashWallis test pairwise comparisons by MannndashWhitneyU-test were made at the 1 significance level (Siegel1975)
Although transplanted plants were measured fivetimes statistical analyses are presented only for the
954C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
final harvest To test whether grazing intensity by crabsaffected transplanted plant performance the percent-age of tillers per turf with wound marks induced byChasmagnathus was correlated with density and aver-age maximum height of tillers percentage of floweringtillers and live aerial biomass of the three species usingthe non-parametric Spearman correlation (Siegel 1975)
One-way s (Zar 1984) were performed to testfor differences in the concentrations of sediment organicmatter total nitrogen and exchangeable phosphatebetween vegetation zones and of leaf nitrogen contentbetween species Transplant performance was analysedusing two-way within each transplant specieswith vegetation zone (lsquoZrsquo S densiflora S alternifloraand S maritimus) and the presence of competitors (lsquoCrsquowith and without competition) as fixed effects andincluding their interaction term (Z X C) For S alterni-flora biomass and density analyses turfs that werecompletely eaten during the experiment were includedbut given values equal to 0 Analyses of the averages ofmaximum tiller height and percentage of flowering tillerincluded only the surviving turfs and their s weretherefore not balanced Sediment organic matter totalnitrogen and exchangeable phosphate data were square-root transformed leaf nitrogen content data were log10(x)transformed tiller density data were square-root (x + 1)transformed biomass data were log10(x + 1) trans-formed and percentage of flowering tiller data werearcsin square-root transformed to increase homogeneityof variance and normality and all s were per-formed at the 5 significance level When a main effector interaction factor was significant mean comparisonswere performed by Tukey multiple comparison tests atthe 5 significance level (Zar 1984)
The relative competition intensity (RCI) was esti-mated by the relative reduction in the average turfperformance due to neighbours ie the difference in aplant attribute without and with neighbours dividedby that attribute without neighbours Although RCI ismost widely used for biomass (Rozema et al 1988Keddy 1989 Grace 1995 Williams amp McCarthy 2001)we also applied for tiller density and maximum heightto evaluate how competition affects these different aerialcomponents If RCI = 0 there is no effect of a competitorwhereas if it is positive the target species has poorerperformance in the presence of neighbours and com-petition is therefore indicated If the RCI is negativethe target species is benefiting from facilitation
Results
Hydrological data are shown for low-water (autumn)and high-water (winter) periods as well as overall (Fig 1Table 1) The Poacutelvora Island salt marsh extended overonly 36 cm in elevation between a S alterniflora lowmarsh (points 1 and 2 minus1 to +8 cm MWL) and dense
shrub cover on the top of the sand bank that boundedthe island (Fig 2)
The highest values of salinity and pH were observedduring the autumn (Table 1) There were no statisticaldifferences in salinity pH or redox potential of the porewater between zones with different dominant speciesfor either the overall period of monitoring or the low-water and high-water periods (Table 1) Redox poten-tial (overall and autumn analyses) and pH (autumn)were however significantly different between samplingpoints A S densiflora stand just above the drift-line zone(point 3 +14 cm MWL) had more reducing sedimentthan a mid marsh stand of the same species at higherelevation (point 9 +25 cm MWL) (Fig 2 Table 1) Forthe autumn period significantly lower pH values werefound in organic-rich sediments of S densiflora or shrubson higher ground (points 9 and 10) and higher values inmarginal S alterniflorandashS densiflora marshes frequentlywashed by alkaline waters (points 1ndash3)
The analysis of flooding frequency pattern across theintertidal zone showed great variability between years(Fig 3) Maximum interannual amplitudes of variationoccurred in the low marsh and lower mid marsh zones(minus15 to +15 cm MWL) where they ranged from 38 to56 (Fig 3b) Thus at MWL a height colonized by allthree species plants can be flooded for as little as 24or as much as 80 of the time depending on the yearconsidered (Fig 3a)
Daily changes in water level can frequently be aslarge as 10ndash20 cm (Figs 1 and 4a) sufficient to expose
Fig 1 Daily variation in water level and salinity of the PatosLagoon estuary between April and August 1995 Linear andpolynomial curves were fitted to highlight seasonal trends
955Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
or flood most of the marsh During the experimentwater level of the Patos Lagoon estuary varied (Fig 4a)but on average was +11 plusmn 17 cm MWL (plusmn SD) Watersalinity averaged 7 plusmn 8 PSU (slightly lower than the 10-year average of 9 plusmn 8 PSU) Characteristic seasonalpatterns of salinity and water level were also observed(Fig 1) The monospecific zones of the reciprocal trans-plantation experiment dominated by S densiflora Salterniflora and S maritimus were flooded 28 37 and43 of the time during the study period respectively(Fig 4a) The sediment of the S densiflora zone had higherconcentrations of organic matter (84 plusmn 05) (plusmn SE)and total nitrogen (16 plusmn 02 g kgminus1) than the S maritimuszone (Fig 4b) but nitrogen concentrations were notsignificantly different between zones (F = 44 P = 007)
Higher concentrations of exchangeable phosphate werefound in S alterniflora (528 plusmn 07 mg kgminus1) and S mariti-mus (495 plusmn 20 mg kgminus1) zones than in the S densiflorazone (377 plusmn 39 mg kgminus1) (Tukey P lt 005) (Fig 4b)
Chasmagnathus ate tillers of all three species during thestudy although fewer tillers showed damage in spring1995 (data not presented) At the end of the experimentsignificantly higher grazing intensities by Chasmag-nathus were recorded for S alterniflora (Fz = 602 P lt0001) and S densiflora tillers (Fz = 289 P lt 0001)when they were transplanted to the S maritimus zonerather than to other zones (Table 2 Fig 5) Crab and crab
Table 1 Mean values (plusmn SE) of pore water salinity pH and redox potential of 10 sampling points located across the Poacutelvora Islandsalt marsh (see Fig 2) KruskalndashWallis test was applied to compare sampling points and vegetation zones (sampling pointsgrouped according to their plant cover)
Analysis Variables n Average
KruskalndashWallis test
Sampling point Vegetation zone
Overall Salinity (PSU) 100 129 plusmn 08(0ndash26)
47 NS 06 NS
pH 100 67 plusmn 01(54ndash85)
159 NS 70 NS
Redox (mV) 100 +175 plusmn 11(minus115 minus +371)
177 22 NS
Autumn Salinity (PSU) 50 173 plusmn 07 72 NS 19 NS1995 (7ndash26)
pH 50 62 plusmn 01(54ndash74)
208 53 NS
Redox (mV) 50 +215 plusmn 10(+46 minus +371)
195 46 NS
Winter Salinity (PSU) 50 81 plusmn 12 15 NS 05 NS1995 (0ndash21)
pH 50 71 plusmn 01 (61ndash85)
165 NS 66 NS
Redox (mV) 50 +114 plusmn 16(minus115 minus +300)
123 NS 24 NS
NS = not significant P lt 005
Table 2 Results of two-way for crab grazing intensity and attributes of transplanted plants (data aggregated per species)for the factorial effects of zone location (z) and competition (c presenceabsence of neighbours) F-values of not includingthe heavily grazed S maritimus zone are shown (inside boxes) only when this procedure produced changes in the significanceof the tested factors or bifactorial interaction Analysed species Sd = S densiflora Sa = S alterniflora Sm = S maritimus
Sd Sa Sm
Fz Fc Fz times c Fz Fc Fz times c Fz Fc Fz times c
Grazed tillers () 2892 028 003 6019 112 232 147 010 010Tiller density 052 831 009 5195 1016 633 066 293 088
1276 2551 008
Maximum tiller height 5403 9215 418 24614 073 2688 101 1356 258
3746 3418 018
Live biomass 767 1101 376 2233 171 322 275 335 186
001 1972 001 089 1048 011
Flowering tillers () 934 479 083 123 108 105 556 202 022
P lt 005 P lt 001 P lt 0001
956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

952
C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society
Journal of Ecology
91
951ndash965
2001) and herbivory (Cargill amp Jefferies 1984 Iribarne
et al
1997 Taylor
et al
1997 Bortolus amp Iribarne 1999)as well as positive interactions (eg plant cover allevi-ating saline stress) (Bertness amp Callaway 1994 Bertnessamp Leonard 1997 Hacker amp Bertness 1999 Callawayamp Pennings 2000) may also be locally important inshaping salt marsh plant communities
It is unclear how far this can be extrapolated tomicrotidal and irregularly flooded salt marshes In theseecosystems flooding patterns are primarily driven byvariation in rainfall and wind direction (Eleuterius ampEleuterius 1979 Stout 1984 Costa
et al
1988 Christian
et al
1990 Wang 1997 Holm
et al
2000 Moumlller
et al
2001) and both salinity and the timing and duration offlooding are thus less predictable In contrast to tidalsalt marshes where dominant species tend to be restrictedto specific zones with little habitat overlap (Pielou ampRoutledge 1976 Russell
et al
1985 Bertness 1999)irregularly flooded salt marshes have perennial domin-ants growing across wide ranges of salinity and waterlevel with considerable habitat overlap (Ranwell 1974Eleuterius amp Eleuterius 1979 Odum 1988 Baldwin ampMendelssohn 1998)
Salt marshes occupy 70 km
2
of the intertidal zone ofthe estuarine region of Patos Lagoon in southern Bra-zil (Costa
et al
1997) Diurnal tidal range is very small(astronomical tides less than 05 m) and fluctuations ofwater level and salinity are irregular (Costa
et al
1988Moumlller
et al
2001) The native grasses
Spartina alterni-flora
Lois and
Spartina densiflora
Brong tend to dom-inate mesohaline low- (between
minus
50 cm and +10 cmMWL mean water level of the estuary) and mid marshes(+10 to +30 cm MWL) respectively (Silva
et al
1993Costa 1997a 1997b) while highly productive stands of
Scirpus maritimus
L may opportunistically dominatetransitional areas between low and mid marshes (Costa1998) These dominant species occur over a wide ver-tical range with a high degree of overlap (Costa 1997a)and monospecific stands of all three can often be foundat the same tidal level The local occurrence of such dis-crete patches suggests that interspecific competitionplays an important role in the structure and composi-tion of these communities The herbivorous Grapsidaecrab
Chasmagnathus granulata
Dana 1851 which is oneof the most abundant macroinvertebrates in the saltmarshes along the coasts of southern Brazil and north-east Argentina is found at high density in the intertidalzone (DrsquoIncao
et al
1990 Copertino
et al
1997) feed-ing on
Spartina
spp (DrsquoIncao
et al
1990 Iribarne
et al
1997 Bortolus amp Iribarne 1999 Bortolus
et al
2002)The relative importance of stress tolerance competi-tion and herbivory in the distribution patterns of thesedominant plants however has yet to be examined Suchecological information is particularly important because
S densiflora
has been introduced unintentionally to thecoasts of California (Kittelson amp Boyd 1997) and south-western Spain (Figueroa amp Castellanos 1988 Castillo
et al
2000) where it has been displacing indigenouslow marsh dominants
We use methods identical to those applied in thenorth hemisphere (reciprocal transplantation betweenvegetated zones and removal of neighbours) to test thehypotheses that (i) distributions of the dominantspecies (
Spartina densiflora
Spartina alterniflora
and
Scirpus maritimus
) on irregularly flooded low to mid marshzones in southern Brazil are related to their competitiveperformance and (ii) stress tolerance has a minor rolein zonation of marshes with poorly predictable hydro-logy We therefore tested the applicability of communitystructuring rules based on work in northern-hemispheresalt marshes to marshes with no distinct tidal regimeWe also examined whether selective grazing by
Chas-magnathus granulata
could affect the performance ofdominant plants and thus community structure
Materials and methods
A
c
10 000 km
2
Patos Lagoon located in southernBrazil (30ndash32
deg
S) is the wordrsquos largest choked lagoon(ie characterized by a single narrow but permanentlyopen entrance channel) It receives runoff from the5500 km
2
Mirim Lagoon near the BrazilndashUruguay border(Moumlller
et al
2001 Seeliger amp Costa 2002) forming alarge lagoon complex that only exchanges water withthe Atlantic Ocean through the Patos Lagoon estuaryan inlet 20 km long and 05ndash30 km wide
The study site was located on the north-west side ofPoacutelvora Island (32
deg
01
prime
S 52
deg
06
prime
W) Rio Grande (RSBrazil) Poacutelvora Island is completely covered by 45 haof undisturbed estuarine salt marsh (Costa 1997a) Lowmarshes are mostly colonized by
Spartina alterniflora
while mid marshes are dominated by either
Spartinadensiflora
or
Scirpus maritimus
and a shrub zone domin-ated by
Myrsine parvifolia
ADC and the halophyticfern
Acrosticum danaefolium
Langsd amp Fish occur onthe high marsh Monospecific stands of
S maritimus
and
S densiflora
are also found colonizing mud flats ofthe low marsh The climate is hot-temperate with clearseasonal fluctuations lowest (13
deg
C) and highest (25
deg
C)average air temperatures are observed in July andJanuary respectively Marsh dominants show specificseasonal growth cycles
S maritimus
behaves as a crypto-phyte with aerial parts sprouting from its rhizomes atthe end of winter peak growth at the end of spring andtillers completely dying back in mid-autumn (Seeliger
et al
1997 Costa 1998) Both
Spartina
species are C4phanerophytes with slow winterndashspring growth andgrowth peaks in the summer and autumn (Silva
et al
1993 Copertino
et al
1997 Seeliger
et al
1997)
The variability of water level in the Patos Lagoon estu-ary was assessed by comparing flooding frequencies inthe intertidal zone between years (each annual periodlasts from July until the following June) Water level
953
Irregularly flooded marsh
copy 2003 British Ecological Society
Journal of Ecology
91
951ndash965
and salinity have been monitored daily since July 1992at the southern part of Marinheiro Island (1 km northof the study area) Water level was determined to thenearest cm between 1000 and 1100 h at a fixed stationand related to the mean water level (MWL) of the estuarybetween July 1992 and June 2001
To document variation in physical stress within veg-etation zones at Poacutelvora Island pore water salinity pHand redox potential of the surface sediment (10 cm) weremeasured at 10 sampling points along a 410-m transectThe pore water samples were obtained between Apriland August 1995 (
n
= 10 dates) with a hand vacuumpump (model 2005G2 Soilmoisture Equip CorpSanta Barbara CA USA) by inserting the bottomlesscollection cylinder 10 cm into the sediment Thesampling points corresponded to zones dominanted by
Spartina alterniflora
(points 1 and 2)
S densiflora
(34 5 and 9)
Scirpus maritimus
(6 7 and 8) and high marshshrubs (
Myrsine parvifolia
point 10) respectively InJanuary 2000 three cores (top 10 cm) were randomlycollected in the low and mid marsh vegetation zonesThe sediment was air-dried and passed through a 2-mmsieve before laboratory analysis The total organiccontent was estimated by combustion at 500
deg
C for6 h (Brower amp Zar 1977) Total Kjedahl nitrogenconcentration in the sediment was determined and theconcentration of exchangeable phosphate estimated bycolourimetry (Allen
et al
1976)
Transplants were established in June 1995 Three adja-cent monospecific stands of
Spartina alterniflora
Sdensiflora
and
Scirpus maritimus
(hereafter called lsquozonesrsquo)each covering
c
2500 m
2
were chosen for the experi-ment The elevation of each zone was estimated with alevel and express relative to the water-level monitoringsite to allow us to estimate their flooding frequencies(percentage of days) Thirty blocks of sediment of 20
times
20
times
20 cm including tillers rhizomes and roots ofeach species (hereafter called lsquoturfsrsquo) were excavatedfrom each of the monospecific zones Ten were trans-planted to similar-sized holes in each of the true zones(ie manipulation controls) Ten blocks were placed inthe other species zones In each zone half of the trans-plants (
n
= 5) of each species were placed in direct con-tact with the surrounding vegetation and the other half(
n
= 5) had all neighbouring plants in a 25-cm widestrip around the perimeter removed by clipping theirshoots at ground level All biological measurementswere made within the 0031-m
2
central areas of the turfsdelimited by a PVC ring with 20 cm diameter fixed tothe sediment with metal pins To avoid invasion by rootsof surrounding plants all neighbour-removal areas werecut to a depth of 25 cm with a spade 25 cm outside theirperimeter The clipping and severing were maintainedfor several weeks until neighbours ceased to grow backinto the neighbour-removal area This methodologyallowed us to account for the overall competitive effect
of neighbouring vegetation and the impact of localphysical stresses on target transplanted plants (Bertness1991a 1991b Pennings amp Callaway 1992 Emery
et al
2001 Pennings amp Moore 2001 Pennings
et al
2003)Five times during the period of study (31 August 1995
26 October 1995 24 November 1995 30 January 1996and 27 February 1996 hereafter referred to as lsquolate winter1995rsquo lsquoearly spring 1995rsquo lsquolate spring 1995rsquo lsquoearly summer1996rsquo and lsquomid-summer 1996rsquo respectively) the numberof live tillers reproductive tillers (with flowersseeds)and tillers with crab wounds from
Chasmagnathusgranulata
were determined in each turf A tiller wasconsidered grazed when it had small scratches or part orall of the tiller had been removed The small Grapsid crab
Metasesarma rubripes
Rathbun (1897) (Capitoli
et al
1977) was also observed in the local marsh but it feedsmainly on plant detritus Insect herbivory was negligibleand appeared uniform for all three plant species
The final harvest of all tillers inside PVC rings wasdone by clipping at ground level at the growth peak ofeach species Because of the early growing season of
Smaritimus
(Costa 1998) it was harvested in early summer1996 whereas the two perennial grasses (
S alterniflora
and
S densiflora
) were collected in mid-summer 1996In the laboratory the heights of all tillers were meas-ured and after removing dead plant material dried at80
deg
C for 48 h and weighed Between-treatment differ-ences in height were analysed using only the 10 tallesttillers per turf of
S densiflora
and the five tallest tillersper turf of
S alterniflora
and
S maritimus
(where standsare less dense due to their architecture)
Maximumtiller height was used as indicator of the growth poten-tial of each species under the experimental conditionsas it excludes bias in the average height due to between-turf variation of tiller density and recently sprouted till-ers The percentage of flowering tillers was calculatedincluding only tillers of height equal to or taller thanthe minimum height from which flowering wasobserved (
S densiflora
= 10 cm
S alterniflora
= 45 cm
S maritimus
= 45 cm) Additionally six plants of eachspecies were collected from their natural zones and theKjeldahl N concentration of the leaves was determined(Allen
et al
1976)
Because of the great temporal variability in pore watersalinity pH and redox the assumptions of homogene-ity of variances and normality were not met with the usualtransformations The KruskalndashWallis test was used totest for differences between sampling points and betweenvegetation zones (ie sampling points grouped by localplant cover) at the 5 significance level (Siegel 1975)When significant differences were found for the KruskalndashWallis test pairwise comparisons by MannndashWhitneyU-test were made at the 1 significance level (Siegel1975)
Although transplanted plants were measured fivetimes statistical analyses are presented only for the
954C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
final harvest To test whether grazing intensity by crabsaffected transplanted plant performance the percent-age of tillers per turf with wound marks induced byChasmagnathus was correlated with density and aver-age maximum height of tillers percentage of floweringtillers and live aerial biomass of the three species usingthe non-parametric Spearman correlation (Siegel 1975)
One-way s (Zar 1984) were performed to testfor differences in the concentrations of sediment organicmatter total nitrogen and exchangeable phosphatebetween vegetation zones and of leaf nitrogen contentbetween species Transplant performance was analysedusing two-way within each transplant specieswith vegetation zone (lsquoZrsquo S densiflora S alternifloraand S maritimus) and the presence of competitors (lsquoCrsquowith and without competition) as fixed effects andincluding their interaction term (Z X C) For S alterni-flora biomass and density analyses turfs that werecompletely eaten during the experiment were includedbut given values equal to 0 Analyses of the averages ofmaximum tiller height and percentage of flowering tillerincluded only the surviving turfs and their s weretherefore not balanced Sediment organic matter totalnitrogen and exchangeable phosphate data were square-root transformed leaf nitrogen content data were log10(x)transformed tiller density data were square-root (x + 1)transformed biomass data were log10(x + 1) trans-formed and percentage of flowering tiller data werearcsin square-root transformed to increase homogeneityof variance and normality and all s were per-formed at the 5 significance level When a main effector interaction factor was significant mean comparisonswere performed by Tukey multiple comparison tests atthe 5 significance level (Zar 1984)
The relative competition intensity (RCI) was esti-mated by the relative reduction in the average turfperformance due to neighbours ie the difference in aplant attribute without and with neighbours dividedby that attribute without neighbours Although RCI ismost widely used for biomass (Rozema et al 1988Keddy 1989 Grace 1995 Williams amp McCarthy 2001)we also applied for tiller density and maximum heightto evaluate how competition affects these different aerialcomponents If RCI = 0 there is no effect of a competitorwhereas if it is positive the target species has poorerperformance in the presence of neighbours and com-petition is therefore indicated If the RCI is negativethe target species is benefiting from facilitation
Results
Hydrological data are shown for low-water (autumn)and high-water (winter) periods as well as overall (Fig 1Table 1) The Poacutelvora Island salt marsh extended overonly 36 cm in elevation between a S alterniflora lowmarsh (points 1 and 2 minus1 to +8 cm MWL) and dense
shrub cover on the top of the sand bank that boundedthe island (Fig 2)
The highest values of salinity and pH were observedduring the autumn (Table 1) There were no statisticaldifferences in salinity pH or redox potential of the porewater between zones with different dominant speciesfor either the overall period of monitoring or the low-water and high-water periods (Table 1) Redox poten-tial (overall and autumn analyses) and pH (autumn)were however significantly different between samplingpoints A S densiflora stand just above the drift-line zone(point 3 +14 cm MWL) had more reducing sedimentthan a mid marsh stand of the same species at higherelevation (point 9 +25 cm MWL) (Fig 2 Table 1) Forthe autumn period significantly lower pH values werefound in organic-rich sediments of S densiflora or shrubson higher ground (points 9 and 10) and higher values inmarginal S alterniflorandashS densiflora marshes frequentlywashed by alkaline waters (points 1ndash3)
The analysis of flooding frequency pattern across theintertidal zone showed great variability between years(Fig 3) Maximum interannual amplitudes of variationoccurred in the low marsh and lower mid marsh zones(minus15 to +15 cm MWL) where they ranged from 38 to56 (Fig 3b) Thus at MWL a height colonized by allthree species plants can be flooded for as little as 24or as much as 80 of the time depending on the yearconsidered (Fig 3a)
Daily changes in water level can frequently be aslarge as 10ndash20 cm (Figs 1 and 4a) sufficient to expose
Fig 1 Daily variation in water level and salinity of the PatosLagoon estuary between April and August 1995 Linear andpolynomial curves were fitted to highlight seasonal trends
955Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
or flood most of the marsh During the experimentwater level of the Patos Lagoon estuary varied (Fig 4a)but on average was +11 plusmn 17 cm MWL (plusmn SD) Watersalinity averaged 7 plusmn 8 PSU (slightly lower than the 10-year average of 9 plusmn 8 PSU) Characteristic seasonalpatterns of salinity and water level were also observed(Fig 1) The monospecific zones of the reciprocal trans-plantation experiment dominated by S densiflora Salterniflora and S maritimus were flooded 28 37 and43 of the time during the study period respectively(Fig 4a) The sediment of the S densiflora zone had higherconcentrations of organic matter (84 plusmn 05) (plusmn SE)and total nitrogen (16 plusmn 02 g kgminus1) than the S maritimuszone (Fig 4b) but nitrogen concentrations were notsignificantly different between zones (F = 44 P = 007)
Higher concentrations of exchangeable phosphate werefound in S alterniflora (528 plusmn 07 mg kgminus1) and S mariti-mus (495 plusmn 20 mg kgminus1) zones than in the S densiflorazone (377 plusmn 39 mg kgminus1) (Tukey P lt 005) (Fig 4b)
Chasmagnathus ate tillers of all three species during thestudy although fewer tillers showed damage in spring1995 (data not presented) At the end of the experimentsignificantly higher grazing intensities by Chasmag-nathus were recorded for S alterniflora (Fz = 602 P lt0001) and S densiflora tillers (Fz = 289 P lt 0001)when they were transplanted to the S maritimus zonerather than to other zones (Table 2 Fig 5) Crab and crab
Table 1 Mean values (plusmn SE) of pore water salinity pH and redox potential of 10 sampling points located across the Poacutelvora Islandsalt marsh (see Fig 2) KruskalndashWallis test was applied to compare sampling points and vegetation zones (sampling pointsgrouped according to their plant cover)
Analysis Variables n Average
KruskalndashWallis test
Sampling point Vegetation zone
Overall Salinity (PSU) 100 129 plusmn 08(0ndash26)
47 NS 06 NS
pH 100 67 plusmn 01(54ndash85)
159 NS 70 NS
Redox (mV) 100 +175 plusmn 11(minus115 minus +371)
177 22 NS
Autumn Salinity (PSU) 50 173 plusmn 07 72 NS 19 NS1995 (7ndash26)
pH 50 62 plusmn 01(54ndash74)
208 53 NS
Redox (mV) 50 +215 plusmn 10(+46 minus +371)
195 46 NS
Winter Salinity (PSU) 50 81 plusmn 12 15 NS 05 NS1995 (0ndash21)
pH 50 71 plusmn 01 (61ndash85)
165 NS 66 NS
Redox (mV) 50 +114 plusmn 16(minus115 minus +300)
123 NS 24 NS
NS = not significant P lt 005
Table 2 Results of two-way for crab grazing intensity and attributes of transplanted plants (data aggregated per species)for the factorial effects of zone location (z) and competition (c presenceabsence of neighbours) F-values of not includingthe heavily grazed S maritimus zone are shown (inside boxes) only when this procedure produced changes in the significanceof the tested factors or bifactorial interaction Analysed species Sd = S densiflora Sa = S alterniflora Sm = S maritimus
Sd Sa Sm
Fz Fc Fz times c Fz Fc Fz times c Fz Fc Fz times c
Grazed tillers () 2892 028 003 6019 112 232 147 010 010Tiller density 052 831 009 5195 1016 633 066 293 088
1276 2551 008
Maximum tiller height 5403 9215 418 24614 073 2688 101 1356 258
3746 3418 018
Live biomass 767 1101 376 2233 171 322 275 335 186
001 1972 001 089 1048 011
Flowering tillers () 934 479 083 123 108 105 556 202 022
P lt 005 P lt 001 P lt 0001
956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

953
Irregularly flooded marsh
copy 2003 British Ecological Society
Journal of Ecology
91
951ndash965
and salinity have been monitored daily since July 1992at the southern part of Marinheiro Island (1 km northof the study area) Water level was determined to thenearest cm between 1000 and 1100 h at a fixed stationand related to the mean water level (MWL) of the estuarybetween July 1992 and June 2001
To document variation in physical stress within veg-etation zones at Poacutelvora Island pore water salinity pHand redox potential of the surface sediment (10 cm) weremeasured at 10 sampling points along a 410-m transectThe pore water samples were obtained between Apriland August 1995 (
n
= 10 dates) with a hand vacuumpump (model 2005G2 Soilmoisture Equip CorpSanta Barbara CA USA) by inserting the bottomlesscollection cylinder 10 cm into the sediment Thesampling points corresponded to zones dominanted by
Spartina alterniflora
(points 1 and 2)
S densiflora
(34 5 and 9)
Scirpus maritimus
(6 7 and 8) and high marshshrubs (
Myrsine parvifolia
point 10) respectively InJanuary 2000 three cores (top 10 cm) were randomlycollected in the low and mid marsh vegetation zonesThe sediment was air-dried and passed through a 2-mmsieve before laboratory analysis The total organiccontent was estimated by combustion at 500
deg
C for6 h (Brower amp Zar 1977) Total Kjedahl nitrogenconcentration in the sediment was determined and theconcentration of exchangeable phosphate estimated bycolourimetry (Allen
et al
1976)
Transplants were established in June 1995 Three adja-cent monospecific stands of
Spartina alterniflora
Sdensiflora
and
Scirpus maritimus
(hereafter called lsquozonesrsquo)each covering
c
2500 m
2
were chosen for the experi-ment The elevation of each zone was estimated with alevel and express relative to the water-level monitoringsite to allow us to estimate their flooding frequencies(percentage of days) Thirty blocks of sediment of 20
times
20
times
20 cm including tillers rhizomes and roots ofeach species (hereafter called lsquoturfsrsquo) were excavatedfrom each of the monospecific zones Ten were trans-planted to similar-sized holes in each of the true zones(ie manipulation controls) Ten blocks were placed inthe other species zones In each zone half of the trans-plants (
n
= 5) of each species were placed in direct con-tact with the surrounding vegetation and the other half(
n
= 5) had all neighbouring plants in a 25-cm widestrip around the perimeter removed by clipping theirshoots at ground level All biological measurementswere made within the 0031-m
2
central areas of the turfsdelimited by a PVC ring with 20 cm diameter fixed tothe sediment with metal pins To avoid invasion by rootsof surrounding plants all neighbour-removal areas werecut to a depth of 25 cm with a spade 25 cm outside theirperimeter The clipping and severing were maintainedfor several weeks until neighbours ceased to grow backinto the neighbour-removal area This methodologyallowed us to account for the overall competitive effect
of neighbouring vegetation and the impact of localphysical stresses on target transplanted plants (Bertness1991a 1991b Pennings amp Callaway 1992 Emery
et al
2001 Pennings amp Moore 2001 Pennings
et al
2003)Five times during the period of study (31 August 1995
26 October 1995 24 November 1995 30 January 1996and 27 February 1996 hereafter referred to as lsquolate winter1995rsquo lsquoearly spring 1995rsquo lsquolate spring 1995rsquo lsquoearly summer1996rsquo and lsquomid-summer 1996rsquo respectively) the numberof live tillers reproductive tillers (with flowersseeds)and tillers with crab wounds from
Chasmagnathusgranulata
were determined in each turf A tiller wasconsidered grazed when it had small scratches or part orall of the tiller had been removed The small Grapsid crab
Metasesarma rubripes
Rathbun (1897) (Capitoli
et al
1977) was also observed in the local marsh but it feedsmainly on plant detritus Insect herbivory was negligibleand appeared uniform for all three plant species
The final harvest of all tillers inside PVC rings wasdone by clipping at ground level at the growth peak ofeach species Because of the early growing season of
Smaritimus
(Costa 1998) it was harvested in early summer1996 whereas the two perennial grasses (
S alterniflora
and
S densiflora
) were collected in mid-summer 1996In the laboratory the heights of all tillers were meas-ured and after removing dead plant material dried at80
deg
C for 48 h and weighed Between-treatment differ-ences in height were analysed using only the 10 tallesttillers per turf of
S densiflora
and the five tallest tillersper turf of
S alterniflora
and
S maritimus
(where standsare less dense due to their architecture)
Maximumtiller height was used as indicator of the growth poten-tial of each species under the experimental conditionsas it excludes bias in the average height due to between-turf variation of tiller density and recently sprouted till-ers The percentage of flowering tillers was calculatedincluding only tillers of height equal to or taller thanthe minimum height from which flowering wasobserved (
S densiflora
= 10 cm
S alterniflora
= 45 cm
S maritimus
= 45 cm) Additionally six plants of eachspecies were collected from their natural zones and theKjeldahl N concentration of the leaves was determined(Allen
et al
1976)
Because of the great temporal variability in pore watersalinity pH and redox the assumptions of homogene-ity of variances and normality were not met with the usualtransformations The KruskalndashWallis test was used totest for differences between sampling points and betweenvegetation zones (ie sampling points grouped by localplant cover) at the 5 significance level (Siegel 1975)When significant differences were found for the KruskalndashWallis test pairwise comparisons by MannndashWhitneyU-test were made at the 1 significance level (Siegel1975)
Although transplanted plants were measured fivetimes statistical analyses are presented only for the
954C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
final harvest To test whether grazing intensity by crabsaffected transplanted plant performance the percent-age of tillers per turf with wound marks induced byChasmagnathus was correlated with density and aver-age maximum height of tillers percentage of floweringtillers and live aerial biomass of the three species usingthe non-parametric Spearman correlation (Siegel 1975)
One-way s (Zar 1984) were performed to testfor differences in the concentrations of sediment organicmatter total nitrogen and exchangeable phosphatebetween vegetation zones and of leaf nitrogen contentbetween species Transplant performance was analysedusing two-way within each transplant specieswith vegetation zone (lsquoZrsquo S densiflora S alternifloraand S maritimus) and the presence of competitors (lsquoCrsquowith and without competition) as fixed effects andincluding their interaction term (Z X C) For S alterni-flora biomass and density analyses turfs that werecompletely eaten during the experiment were includedbut given values equal to 0 Analyses of the averages ofmaximum tiller height and percentage of flowering tillerincluded only the surviving turfs and their s weretherefore not balanced Sediment organic matter totalnitrogen and exchangeable phosphate data were square-root transformed leaf nitrogen content data were log10(x)transformed tiller density data were square-root (x + 1)transformed biomass data were log10(x + 1) trans-formed and percentage of flowering tiller data werearcsin square-root transformed to increase homogeneityof variance and normality and all s were per-formed at the 5 significance level When a main effector interaction factor was significant mean comparisonswere performed by Tukey multiple comparison tests atthe 5 significance level (Zar 1984)
The relative competition intensity (RCI) was esti-mated by the relative reduction in the average turfperformance due to neighbours ie the difference in aplant attribute without and with neighbours dividedby that attribute without neighbours Although RCI ismost widely used for biomass (Rozema et al 1988Keddy 1989 Grace 1995 Williams amp McCarthy 2001)we also applied for tiller density and maximum heightto evaluate how competition affects these different aerialcomponents If RCI = 0 there is no effect of a competitorwhereas if it is positive the target species has poorerperformance in the presence of neighbours and com-petition is therefore indicated If the RCI is negativethe target species is benefiting from facilitation
Results
Hydrological data are shown for low-water (autumn)and high-water (winter) periods as well as overall (Fig 1Table 1) The Poacutelvora Island salt marsh extended overonly 36 cm in elevation between a S alterniflora lowmarsh (points 1 and 2 minus1 to +8 cm MWL) and dense
shrub cover on the top of the sand bank that boundedthe island (Fig 2)
The highest values of salinity and pH were observedduring the autumn (Table 1) There were no statisticaldifferences in salinity pH or redox potential of the porewater between zones with different dominant speciesfor either the overall period of monitoring or the low-water and high-water periods (Table 1) Redox poten-tial (overall and autumn analyses) and pH (autumn)were however significantly different between samplingpoints A S densiflora stand just above the drift-line zone(point 3 +14 cm MWL) had more reducing sedimentthan a mid marsh stand of the same species at higherelevation (point 9 +25 cm MWL) (Fig 2 Table 1) Forthe autumn period significantly lower pH values werefound in organic-rich sediments of S densiflora or shrubson higher ground (points 9 and 10) and higher values inmarginal S alterniflorandashS densiflora marshes frequentlywashed by alkaline waters (points 1ndash3)
The analysis of flooding frequency pattern across theintertidal zone showed great variability between years(Fig 3) Maximum interannual amplitudes of variationoccurred in the low marsh and lower mid marsh zones(minus15 to +15 cm MWL) where they ranged from 38 to56 (Fig 3b) Thus at MWL a height colonized by allthree species plants can be flooded for as little as 24or as much as 80 of the time depending on the yearconsidered (Fig 3a)
Daily changes in water level can frequently be aslarge as 10ndash20 cm (Figs 1 and 4a) sufficient to expose
Fig 1 Daily variation in water level and salinity of the PatosLagoon estuary between April and August 1995 Linear andpolynomial curves were fitted to highlight seasonal trends
955Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
or flood most of the marsh During the experimentwater level of the Patos Lagoon estuary varied (Fig 4a)but on average was +11 plusmn 17 cm MWL (plusmn SD) Watersalinity averaged 7 plusmn 8 PSU (slightly lower than the 10-year average of 9 plusmn 8 PSU) Characteristic seasonalpatterns of salinity and water level were also observed(Fig 1) The monospecific zones of the reciprocal trans-plantation experiment dominated by S densiflora Salterniflora and S maritimus were flooded 28 37 and43 of the time during the study period respectively(Fig 4a) The sediment of the S densiflora zone had higherconcentrations of organic matter (84 plusmn 05) (plusmn SE)and total nitrogen (16 plusmn 02 g kgminus1) than the S maritimuszone (Fig 4b) but nitrogen concentrations were notsignificantly different between zones (F = 44 P = 007)
Higher concentrations of exchangeable phosphate werefound in S alterniflora (528 plusmn 07 mg kgminus1) and S mariti-mus (495 plusmn 20 mg kgminus1) zones than in the S densiflorazone (377 plusmn 39 mg kgminus1) (Tukey P lt 005) (Fig 4b)
Chasmagnathus ate tillers of all three species during thestudy although fewer tillers showed damage in spring1995 (data not presented) At the end of the experimentsignificantly higher grazing intensities by Chasmag-nathus were recorded for S alterniflora (Fz = 602 P lt0001) and S densiflora tillers (Fz = 289 P lt 0001)when they were transplanted to the S maritimus zonerather than to other zones (Table 2 Fig 5) Crab and crab
Table 1 Mean values (plusmn SE) of pore water salinity pH and redox potential of 10 sampling points located across the Poacutelvora Islandsalt marsh (see Fig 2) KruskalndashWallis test was applied to compare sampling points and vegetation zones (sampling pointsgrouped according to their plant cover)
Analysis Variables n Average
KruskalndashWallis test
Sampling point Vegetation zone
Overall Salinity (PSU) 100 129 plusmn 08(0ndash26)
47 NS 06 NS
pH 100 67 plusmn 01(54ndash85)
159 NS 70 NS
Redox (mV) 100 +175 plusmn 11(minus115 minus +371)
177 22 NS
Autumn Salinity (PSU) 50 173 plusmn 07 72 NS 19 NS1995 (7ndash26)
pH 50 62 plusmn 01(54ndash74)
208 53 NS
Redox (mV) 50 +215 plusmn 10(+46 minus +371)
195 46 NS
Winter Salinity (PSU) 50 81 plusmn 12 15 NS 05 NS1995 (0ndash21)
pH 50 71 plusmn 01 (61ndash85)
165 NS 66 NS
Redox (mV) 50 +114 plusmn 16(minus115 minus +300)
123 NS 24 NS
NS = not significant P lt 005
Table 2 Results of two-way for crab grazing intensity and attributes of transplanted plants (data aggregated per species)for the factorial effects of zone location (z) and competition (c presenceabsence of neighbours) F-values of not includingthe heavily grazed S maritimus zone are shown (inside boxes) only when this procedure produced changes in the significanceof the tested factors or bifactorial interaction Analysed species Sd = S densiflora Sa = S alterniflora Sm = S maritimus
Sd Sa Sm
Fz Fc Fz times c Fz Fc Fz times c Fz Fc Fz times c
Grazed tillers () 2892 028 003 6019 112 232 147 010 010Tiller density 052 831 009 5195 1016 633 066 293 088
1276 2551 008
Maximum tiller height 5403 9215 418 24614 073 2688 101 1356 258
3746 3418 018
Live biomass 767 1101 376 2233 171 322 275 335 186
001 1972 001 089 1048 011
Flowering tillers () 934 479 083 123 108 105 556 202 022
P lt 005 P lt 001 P lt 0001
956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

954C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
final harvest To test whether grazing intensity by crabsaffected transplanted plant performance the percent-age of tillers per turf with wound marks induced byChasmagnathus was correlated with density and aver-age maximum height of tillers percentage of floweringtillers and live aerial biomass of the three species usingthe non-parametric Spearman correlation (Siegel 1975)
One-way s (Zar 1984) were performed to testfor differences in the concentrations of sediment organicmatter total nitrogen and exchangeable phosphatebetween vegetation zones and of leaf nitrogen contentbetween species Transplant performance was analysedusing two-way within each transplant specieswith vegetation zone (lsquoZrsquo S densiflora S alternifloraand S maritimus) and the presence of competitors (lsquoCrsquowith and without competition) as fixed effects andincluding their interaction term (Z X C) For S alterni-flora biomass and density analyses turfs that werecompletely eaten during the experiment were includedbut given values equal to 0 Analyses of the averages ofmaximum tiller height and percentage of flowering tillerincluded only the surviving turfs and their s weretherefore not balanced Sediment organic matter totalnitrogen and exchangeable phosphate data were square-root transformed leaf nitrogen content data were log10(x)transformed tiller density data were square-root (x + 1)transformed biomass data were log10(x + 1) trans-formed and percentage of flowering tiller data werearcsin square-root transformed to increase homogeneityof variance and normality and all s were per-formed at the 5 significance level When a main effector interaction factor was significant mean comparisonswere performed by Tukey multiple comparison tests atthe 5 significance level (Zar 1984)
The relative competition intensity (RCI) was esti-mated by the relative reduction in the average turfperformance due to neighbours ie the difference in aplant attribute without and with neighbours dividedby that attribute without neighbours Although RCI ismost widely used for biomass (Rozema et al 1988Keddy 1989 Grace 1995 Williams amp McCarthy 2001)we also applied for tiller density and maximum heightto evaluate how competition affects these different aerialcomponents If RCI = 0 there is no effect of a competitorwhereas if it is positive the target species has poorerperformance in the presence of neighbours and com-petition is therefore indicated If the RCI is negativethe target species is benefiting from facilitation
Results
Hydrological data are shown for low-water (autumn)and high-water (winter) periods as well as overall (Fig 1Table 1) The Poacutelvora Island salt marsh extended overonly 36 cm in elevation between a S alterniflora lowmarsh (points 1 and 2 minus1 to +8 cm MWL) and dense
shrub cover on the top of the sand bank that boundedthe island (Fig 2)
The highest values of salinity and pH were observedduring the autumn (Table 1) There were no statisticaldifferences in salinity pH or redox potential of the porewater between zones with different dominant speciesfor either the overall period of monitoring or the low-water and high-water periods (Table 1) Redox poten-tial (overall and autumn analyses) and pH (autumn)were however significantly different between samplingpoints A S densiflora stand just above the drift-line zone(point 3 +14 cm MWL) had more reducing sedimentthan a mid marsh stand of the same species at higherelevation (point 9 +25 cm MWL) (Fig 2 Table 1) Forthe autumn period significantly lower pH values werefound in organic-rich sediments of S densiflora or shrubson higher ground (points 9 and 10) and higher values inmarginal S alterniflorandashS densiflora marshes frequentlywashed by alkaline waters (points 1ndash3)
The analysis of flooding frequency pattern across theintertidal zone showed great variability between years(Fig 3) Maximum interannual amplitudes of variationoccurred in the low marsh and lower mid marsh zones(minus15 to +15 cm MWL) where they ranged from 38 to56 (Fig 3b) Thus at MWL a height colonized by allthree species plants can be flooded for as little as 24or as much as 80 of the time depending on the yearconsidered (Fig 3a)
Daily changes in water level can frequently be aslarge as 10ndash20 cm (Figs 1 and 4a) sufficient to expose
Fig 1 Daily variation in water level and salinity of the PatosLagoon estuary between April and August 1995 Linear andpolynomial curves were fitted to highlight seasonal trends
955Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
or flood most of the marsh During the experimentwater level of the Patos Lagoon estuary varied (Fig 4a)but on average was +11 plusmn 17 cm MWL (plusmn SD) Watersalinity averaged 7 plusmn 8 PSU (slightly lower than the 10-year average of 9 plusmn 8 PSU) Characteristic seasonalpatterns of salinity and water level were also observed(Fig 1) The monospecific zones of the reciprocal trans-plantation experiment dominated by S densiflora Salterniflora and S maritimus were flooded 28 37 and43 of the time during the study period respectively(Fig 4a) The sediment of the S densiflora zone had higherconcentrations of organic matter (84 plusmn 05) (plusmn SE)and total nitrogen (16 plusmn 02 g kgminus1) than the S maritimuszone (Fig 4b) but nitrogen concentrations were notsignificantly different between zones (F = 44 P = 007)
Higher concentrations of exchangeable phosphate werefound in S alterniflora (528 plusmn 07 mg kgminus1) and S mariti-mus (495 plusmn 20 mg kgminus1) zones than in the S densiflorazone (377 plusmn 39 mg kgminus1) (Tukey P lt 005) (Fig 4b)
Chasmagnathus ate tillers of all three species during thestudy although fewer tillers showed damage in spring1995 (data not presented) At the end of the experimentsignificantly higher grazing intensities by Chasmag-nathus were recorded for S alterniflora (Fz = 602 P lt0001) and S densiflora tillers (Fz = 289 P lt 0001)when they were transplanted to the S maritimus zonerather than to other zones (Table 2 Fig 5) Crab and crab
Table 1 Mean values (plusmn SE) of pore water salinity pH and redox potential of 10 sampling points located across the Poacutelvora Islandsalt marsh (see Fig 2) KruskalndashWallis test was applied to compare sampling points and vegetation zones (sampling pointsgrouped according to their plant cover)
Analysis Variables n Average
KruskalndashWallis test
Sampling point Vegetation zone
Overall Salinity (PSU) 100 129 plusmn 08(0ndash26)
47 NS 06 NS
pH 100 67 plusmn 01(54ndash85)
159 NS 70 NS
Redox (mV) 100 +175 plusmn 11(minus115 minus +371)
177 22 NS
Autumn Salinity (PSU) 50 173 plusmn 07 72 NS 19 NS1995 (7ndash26)
pH 50 62 plusmn 01(54ndash74)
208 53 NS
Redox (mV) 50 +215 plusmn 10(+46 minus +371)
195 46 NS
Winter Salinity (PSU) 50 81 plusmn 12 15 NS 05 NS1995 (0ndash21)
pH 50 71 plusmn 01 (61ndash85)
165 NS 66 NS
Redox (mV) 50 +114 plusmn 16(minus115 minus +300)
123 NS 24 NS
NS = not significant P lt 005
Table 2 Results of two-way for crab grazing intensity and attributes of transplanted plants (data aggregated per species)for the factorial effects of zone location (z) and competition (c presenceabsence of neighbours) F-values of not includingthe heavily grazed S maritimus zone are shown (inside boxes) only when this procedure produced changes in the significanceof the tested factors or bifactorial interaction Analysed species Sd = S densiflora Sa = S alterniflora Sm = S maritimus
Sd Sa Sm
Fz Fc Fz times c Fz Fc Fz times c Fz Fc Fz times c
Grazed tillers () 2892 028 003 6019 112 232 147 010 010Tiller density 052 831 009 5195 1016 633 066 293 088
1276 2551 008
Maximum tiller height 5403 9215 418 24614 073 2688 101 1356 258
3746 3418 018
Live biomass 767 1101 376 2233 171 322 275 335 186
001 1972 001 089 1048 011
Flowering tillers () 934 479 083 123 108 105 556 202 022
P lt 005 P lt 001 P lt 0001
956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003
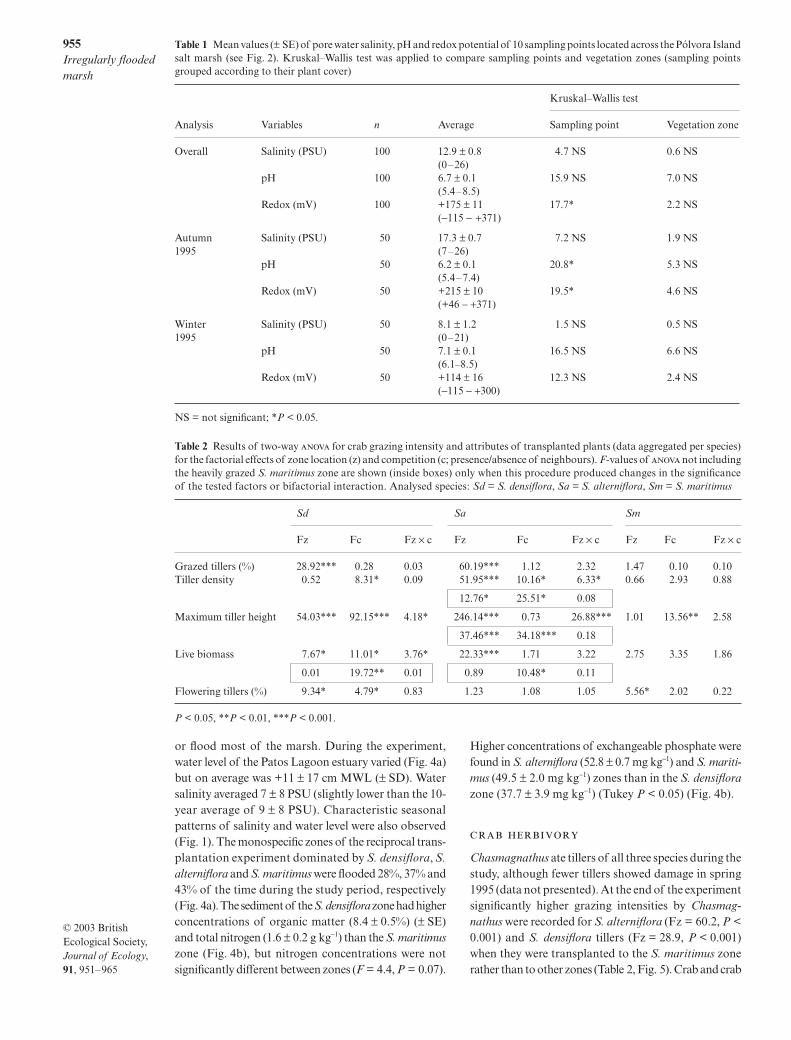
955Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
or flood most of the marsh During the experimentwater level of the Patos Lagoon estuary varied (Fig 4a)but on average was +11 plusmn 17 cm MWL (plusmn SD) Watersalinity averaged 7 plusmn 8 PSU (slightly lower than the 10-year average of 9 plusmn 8 PSU) Characteristic seasonalpatterns of salinity and water level were also observed(Fig 1) The monospecific zones of the reciprocal trans-plantation experiment dominated by S densiflora Salterniflora and S maritimus were flooded 28 37 and43 of the time during the study period respectively(Fig 4a) The sediment of the S densiflora zone had higherconcentrations of organic matter (84 plusmn 05) (plusmn SE)and total nitrogen (16 plusmn 02 g kgminus1) than the S maritimuszone (Fig 4b) but nitrogen concentrations were notsignificantly different between zones (F = 44 P = 007)
Higher concentrations of exchangeable phosphate werefound in S alterniflora (528 plusmn 07 mg kgminus1) and S mariti-mus (495 plusmn 20 mg kgminus1) zones than in the S densiflorazone (377 plusmn 39 mg kgminus1) (Tukey P lt 005) (Fig 4b)
Chasmagnathus ate tillers of all three species during thestudy although fewer tillers showed damage in spring1995 (data not presented) At the end of the experimentsignificantly higher grazing intensities by Chasmag-nathus were recorded for S alterniflora (Fz = 602 P lt0001) and S densiflora tillers (Fz = 289 P lt 0001)when they were transplanted to the S maritimus zonerather than to other zones (Table 2 Fig 5) Crab and crab
Table 1 Mean values (plusmn SE) of pore water salinity pH and redox potential of 10 sampling points located across the Poacutelvora Islandsalt marsh (see Fig 2) KruskalndashWallis test was applied to compare sampling points and vegetation zones (sampling pointsgrouped according to their plant cover)
Analysis Variables n Average
KruskalndashWallis test
Sampling point Vegetation zone
Overall Salinity (PSU) 100 129 plusmn 08(0ndash26)
47 NS 06 NS
pH 100 67 plusmn 01(54ndash85)
159 NS 70 NS
Redox (mV) 100 +175 plusmn 11(minus115 minus +371)
177 22 NS
Autumn Salinity (PSU) 50 173 plusmn 07 72 NS 19 NS1995 (7ndash26)
pH 50 62 plusmn 01(54ndash74)
208 53 NS
Redox (mV) 50 +215 plusmn 10(+46 minus +371)
195 46 NS
Winter Salinity (PSU) 50 81 plusmn 12 15 NS 05 NS1995 (0ndash21)
pH 50 71 plusmn 01 (61ndash85)
165 NS 66 NS
Redox (mV) 50 +114 plusmn 16(minus115 minus +300)
123 NS 24 NS
NS = not significant P lt 005
Table 2 Results of two-way for crab grazing intensity and attributes of transplanted plants (data aggregated per species)for the factorial effects of zone location (z) and competition (c presenceabsence of neighbours) F-values of not includingthe heavily grazed S maritimus zone are shown (inside boxes) only when this procedure produced changes in the significanceof the tested factors or bifactorial interaction Analysed species Sd = S densiflora Sa = S alterniflora Sm = S maritimus
Sd Sa Sm
Fz Fc Fz times c Fz Fc Fz times c Fz Fc Fz times c
Grazed tillers () 2892 028 003 6019 112 232 147 010 010Tiller density 052 831 009 5195 1016 633 066 293 088
1276 2551 008
Maximum tiller height 5403 9215 418 24614 073 2688 101 1356 258
3746 3418 018
Live biomass 767 1101 376 2233 171 322 275 335 186
001 1972 001 089 1048 011
Flowering tillers () 934 479 083 123 108 105 556 202 022
P lt 005 P lt 001 P lt 0001
956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

956C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Fig 2 Pore water salinity pH and redox potential values between April and August 1995 at 10 sampling points located acrossPoacutelvora Island salt marsh (n = 10 sampling dates) Average minimum and maximum observed values as well as first data quarterabove and below the average are shown Vegetation zones are represented by different pattern Lower case letters indicatesignificant differences (MannndashWhitney U-test P lt 001) between sampling points for each environmental characteristic for theoverall monitoring period (see Table 1)
Fig 3 (a) Annual flooding frequency curves for 1995ndash2001 (b) Ten-year (for 1992ndash2001) average flooding frequency andmaximum amplitude of interannual variation in frequency observed at each tidal elevation Graphs were calculated in 5-cmintervals from the water level measurements Topographic marsh zones were defined according to Costa (1997a)
957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

957Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
burrow densities appeared to be uniform for all zonesIn the S maritimus zone regardless of whether or notneighbours were present S alterniflora tillers were morefrequently grazed (average of turfs without competition= 852 of tillers with competition = 944 of tillers)than S densiflora tillers (without competition = 455and with competition = 489) (Fig 5) The lowestpercentage of S alterniflora-grazed tillers occurred inits own zone (P lt 005 Turkey test) S maritimus wasrarely grazed and grazing on S maritimus did not differbetween zones or neighbour removal treatment levels(Table 2 Fig 5)
Chemical analyses showed that the leaf nitrogencontent of S densiflora (54 plusmn 03 mg gminus1) was signi-ficantly lower (F = 222 P lt 0001) than for leaves of Salterniflora (97 plusmn 05 mg gminus1) and S maritimus (121 plusmn27 mg gminus1)
There was no significant correlation between grazingintensity and performance of S maritimus Increasingcrab herbivory however significantly decreased thegrowth of both Spartina species (Fig 6) Although Sdensiflora maximum tiller height live aerial biomassand percentage of flowering tillers were negatively cor-related with grazing intensity by crabs tiller density
Fig 4 Daily variation in water level of the Patos Lagoon estuary between July 1995 and February 1996 (a) and averageconcentrations (plusmn SE) of organic matter total nitrogen and exchangeable phosphate in the surface sediment (b) of the zonesdominated by S densiflora (Sd stripes) S alterniflora (Sa white) and S maritimus (Sm black) Height of each vegetation zone(dashed horizontal lines) Some water level data are missing for October and November 1995 Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones within each sediment parameter
Fig 5 Mean percentage (plusmn SE) of tillers grazed by Chasmagnathus granulata crabs when transplanted into zones dominated byS densiflora (stripes) S alterniflora (white) and S maritimus (black) Lower case letters indicate significant differences (posterioriTukey test P lt 005) between zones and competition treatments (NC = no competition WC = with competition) within each species
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003
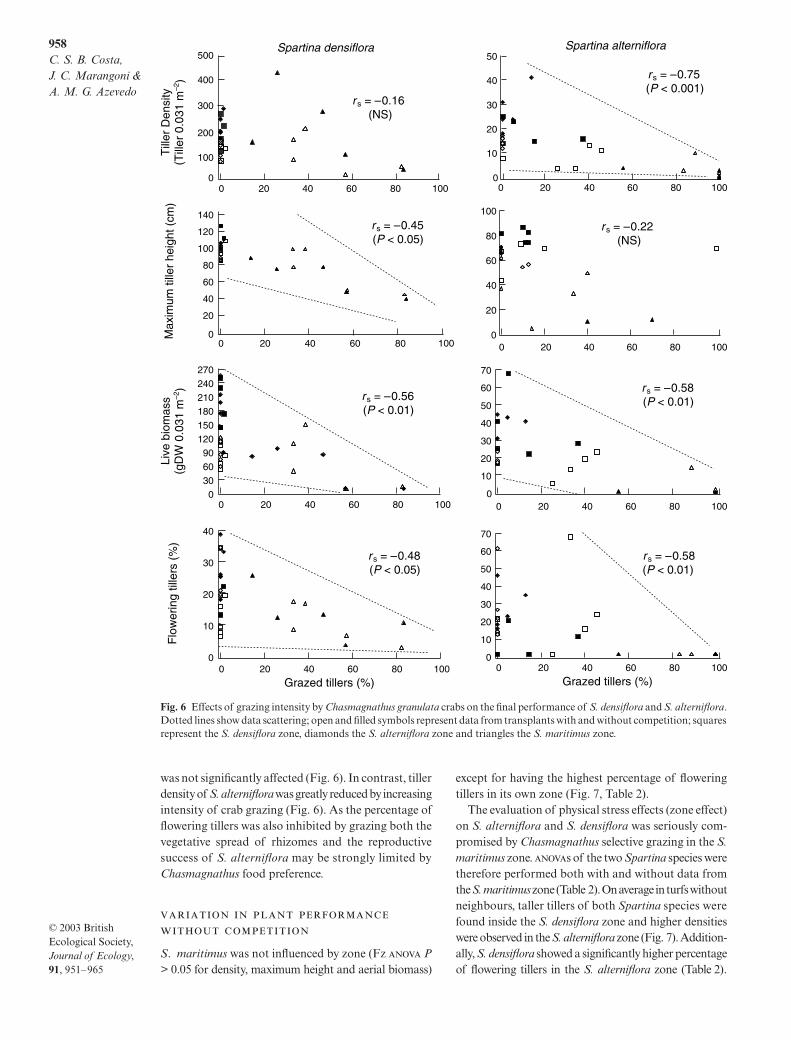
958C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
was not significantly affected (Fig 6) In contrast tillerdensity of S alterniflora was greatly reduced by increasingintensity of crab grazing (Fig 6) As the percentage offlowering tillers was also inhibited by grazing both thevegetative spread of rhizomes and the reproductivesuccess of S alterniflora may be strongly limited byChasmagnathus food preference
S maritimus was not influenced by zone (Fz Pgt 005 for density maximum height and aerial biomass)
except for having the highest percentage of floweringtillers in its own zone (Fig 7 Table 2)
The evaluation of physical stress effects (zone effect)on S alterniflora and S densiflora was seriously com-promised by Chasmagnathus selective grazing in the Smaritimus zone s of the two Spartina species weretherefore performed both with and without data fromthe S maritimus zone (Table 2) On average in turfs withoutneighbours taller tillers of both Spartina species werefound inside the S densiflora zone and higher densitieswere observed in the S alterniflora zone (Fig 7) Addition-ally S densiflora showed a significantly higher percentageof flowering tillers in the S alterniflora zone (Table 2)
Fig 6 Effects of grazing intensity by Chasmagnathus granulata crabs on the final performance of S densiflora and S alternifloraDotted lines show data scattering open and filled symbols represent data from transplants with and without competition squaresrepresent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone
959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

959Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Significant differences in live biomass between zoneswere detected only when data from S maritimus zonewere included (Table 2) The lack of interzone differ-ences in average live biomass of S alterniflora and Sdensiflora (Fig 7) is consistent with their allocations totillering or individual tiller height in each othersrsquo zones
S densiflora and S maritimus showed linear increasesin average tiller densities in all treatments during thestudy but particularly for turfs without competition(Fig 8) At the end of the experiment regardless ofvegetation zone significantly lower tiller densitiesmaximum tiller heights live aerial biomass and tillerflowering of S densiflora were found in turfs growingwith competition (Fc s P lt 005 Table 2) Sig-nificant interactions for S densifora maximum tillerheight and live aerial biomass (Table 2) also suggestvariation in the intensity of neighbour effects betweenzones However the significance of these interactionterms and of the differences between zones (significant
Fz) disappears when data from the S maritimus zoneare excluded from the analysis indicating that dif-ferences in plant performance between zones wererelated to crab grazing effects S maritimus attributesmeasured in summer 1996 were not significantlyaffected by neighbour presence (Table 2 Fig 7) exceptthat the tallest tillers were found in turfs surroundedby neighbours (Fc = 135 P lt 0001) In this specific casethe only significant pairwise comparison betweentreatments (Tukey test P lt 005) was observed betweenturfs with (767 plusmn 11 cm) and without neighbours(685 plusmn 10 cm) inside S maritimus own zone (Fig 7)
S alterniflora showed an increase in tiller densities inSpartina species zones but starting in late spring 1995(Fig 8) whereas its average density in the S maritimuszone fell throughout the experimental period By latesummer 1996 S alterniflora had high tiller densities inturfs without competition (Fc = 102 P lt 005) pro-vided they were in less grazed areas both inside itsconspecific zone (28 plusmn 22 tillers 0031 mminus2) and in theS densiflora zone (19 plusmn 39 tillers 0031 mminus2) (Fig 7)No other significant main effects of the competition
Fig 7 Effects of competition (NC = no competition WC = with competition) and transplant location on mean (plusmn SE) tillerdensity maximum height and flowering and live aerial biomass of the three dominant species Lower case letters indicatesignificant differences (posteriori Tukey test P lt 005) between zones (S densiflora = stripes S alterniflora = white S maritimus= black) and competition treatment within each species
960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

960C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
treatment were observed for S alterniflora in these ana-lyses but significant interactions (tiller density andmaximum height) were detected (Table 2) Two-wayfactorial s performed without the data from theS maritimus zone suggest that S alterniflora densitymaximum tiller height and live biomass were signifi-cantly depressed by conspecifics and by S densifloraneighbours (Table 2 Fig 7) The lack of significantinteractions in these later analyses also suggestedsymmetry between intraspecific and interspecificcompetitive effects
All three species demonstrated a great sensitivity oftiller density and live biomass to the presence of neigh-bours RCI values indicated competition reduced aver-age values by between 36 and 57 in the two Spartinaspecies (Table 3) However the overall performance ofS alterniflora was greatly improved (eg RCI = minus850for live biomass) and live biomass of S densifloraslightly increased (RCI = minus18) by the presence ofneighbours in crab-grazed stands of S maritimusMaximum tiller height of the two Spartina species waslittle affected by congeneric presence (RCI lt 15)
and S maritimus plants were consistently slightlytaller (4ndash12) when surrounded by other plants(negative RCI values) The presence of S alternifloraneighbours inhibited (42) S maritimus tillering andlive biomass but the same parameters were little affectedby conspecifics or S densiflora plants (RCI lt 19reduction)
Discussion
S alterniflora S densiflora and S maritimus were ableto grow across the entire elevation gradient of the irr-egularly flooded low and mid marshes of Poacutelvora IslandAlthough selective herbivory by crabs affected the per-formance of both Spartina species and restricted theinterpretation of the study results strong intra- andinterspecific competition was observed in all monospe-cific zones of the marsh As no competitive hierarchywas detected zonation in irregularly flooded low and midmarshes of southern Brazil seems to not be explainedby the displacement of competitive subordinates tophysically stressful habitats
Fig 8 Fluctuation of mean tiller density of the transplanted species in different zones (S densiflora = stripes S alterniflora =white S maritimus = black) and competition treatments (open and filled symbols represent data from transplants with andwithout neighbours squares represent the S densiflora zone diamonds the S alterniflora zone and triangles the S maritimus zone)
Table 3 Relative competition intensity index (RCI) for different biological attributes of S densiflora (Sd ) S alterniflora (Sa) andS maritimus (Sm) plants transplanted to three vegetation zones Negative values indicate positive interactions
Transplanted zone
Tiller density Maximum tiller height Live biomass
Sd Sa Sm Sd Sa Sm Sd Sa Sm
Sd 36 57 19 14 15 minus4 51 51 4Sa 43 46 42 10 11 minus3 52 41 42Sm 48 minus128 9 8 minus397 minus12 minus18 minus850 10
961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

961Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
The irregularity and the relatively high amplitudeof variation in hydrological conditions appear to ex-plain the lack of a persistent physical stress gradientacross the low and mid marshes of Patos Lagoon estuaryPersistent differences in physical factors betweenvegetation zones have been demonstrated for tidal andnon-tidal salt marshes but are always related to regularpatterns of flooding tides or risingfalling groundwater levels Marked differences of pore water redox(Bertness amp Ellison 1987 Odum 1988 Bertness 1991aCastellanos et al 1994 Portnoy amp Giblin 1997 Paludanamp Morris 1999 Castillo et al 2000 Pennings amp Moore 2001Figueroa et al 2003) and salinity (Pennings amp Callaway1992 Pennings amp Bertness 1999 Rogel et al 2000Pennings amp Moore 2001) have been found across ele-vation gradients at a wide range of latitudes Differ-ences between vegetation zones in surface sediment pHare less common but indirect factors such as organiccontent accumulation in older marshes and salt depo-sition leaching in depressions can produce differencesof 1ndash4 pH units (Vince amp Snow 1984 Bertness amp Ellison1987 Paludan amp Morris 1999 Rogel et al 2000) Sim-ilar ranges of redox salinity and pH values were foundalong Poacutelvora Island marsh but because of the flatlandscape and the large amplitude of daily (Costa et al1988) and seasonal (Garcia et al 2001 Moumlller et al2001) fluctuations in the estuary water level differencesbetween vegetation zones were not persistent in timeand it is unlikely that they impose distribution limitsfor the dominant perennials Furthermore the sedimentsat Poacutelvora Island are typically well oxidized with lowsalinity and nearly neutral pH These are ideal growthconditions for facultative halophytes (Lessmann et al1997 Pennings amp Moore 2001 Pennings et al 2003)suggesting that most dominant plants are not physi-cally limited in this system
Episodes of warming (El Nintildeo) and cooling (La Nintildea)of surface waters in the eastern tropical Pacific arehighly correlated with excessive rainfall and droughtrespectively over the south-western Atlantic (Zhou ampLau 2001 Coelho et al 2002) These events possess astrong temporal stochastic component (Burgers 1999)and are responsible for a great part of the interannualvariability in both the salinity pattern (Garcia et al2001 Seeliger amp Costa 2002) and flooding frequency ofintertidal zones of the Patos Lagoon estuary (Fig 3)Large and small fresh water discharges into the PatosLagoon estuary are associated with periods of strongEl Nino (19971998 data Garcia et al 2001) and LaNina (19992000 data NOAA 2000 2001) events in thePacific Ocean Interannual differences in estuarine waterlevel can cause the physical stress gradient to move up anddown the marsh elevation particularly in low and midmarsh zones Such conditions are not considered by themost common models explaining salt marsh structuring(Bertness 1991a 1991b Pennings amp Callaway 1992 Emery
et al 2001 Pennings amp Bertness 2001) These are basedon the existence of a persistent physical stress gradientand therefore have limited use in the Patos Lagoon
By the end of the growing season on average Chasmagna-thus granulata grazing had reduced S alterniflora bio-mass in the S maritimus zone (averaged over competitiontreatments) by 366 in relation to its own zone Sdensiflora biomass was however reduced by only 10although a strong preference for recently sprouted leaveswhen grazing on S densiflora (Bortolus amp Iribarne 1999)means that grazing intensity of this species was probablyunderestimated Selective herbivory by Chasmagnathusis similar to that of other Grapsidae crabs from mangroves(Robertson 1991 Lee 1998) and is probably related toanatomical and chemical differences between plant species(Lightfoot amp Whitford 1987 Robertson 1991 Lee 1998Pennings et al 1998) Lower sensitivity of S densiflorato grazing seems to be explained by its clumped growthform and a capacity for rapid tillering in response toperturbation of its aerial parts (eg clipping Silva et al1993 or burning Bortolus amp Iribarne 1999) it also haslower palatability (it has more silica bodies in the epidermisand sclerenchyma fibres in its leaves than S alternifloraSantos amp Almeida 2000) S alterniflora also has nitrogenleaf contents double those of S densiflora and S marit-imus has similar levels to S alterniflora but none is presentas easily assimilated compounds such as glycinebethaineand proline (up to 11 in Spartina sp Stewart et al1979 Parsons amp de la Cruz 1980 Cavalieri 1983)
Several herbivores (grazers leaf suckers seed eatersetc) utilize Spartina species in salt marshes around theworld (Adam 1993 Furbish amp Albano 1994 Daehleramp Strong 1995 Bertness 1999 Davy et al 2000)Herbivory by crabs (Robertson 1991 Wolcott amp OrsquoConnor1992 Lee 1998) and other organisms (Louda et al 1990Taylor et al 1997 Silliman amp Zieman 2001 Silliman ampBertness 2002) frequently limits the growth of grazedplants and reduces biomass andor tiller heights Dif-ferential consumption of mangrove propagules by Grap-sidae crabs can affect forest composition and maydetermine the local dominance of some trees (Robertson1991 Lee 1998) Recent findings of strong top-downcontrol of S alterniflora by the gastropod Littorariairrorata on the south-east coast of America (Silliman ampZieman 2001 Silliman amp Bertness 2002) indicate thatherbivores may also play a larger role in marsh plantcommunity structure than is generally recognized(Pennings amp Bertness 2001) Our results suggest thatChasmagnathus herbivory can be very plant-selectivebut evaluating crab effects on South American saltmarshes will require further field experimentation
Few differences in the growth of transplanted plantscould be exclusively related to physiological limitations
962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

962C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
imposed by variations in flooding frequency betweenzones Scirpus maritimus grew similarly in all zones andthe aerial biomass of S alterniflora and S densifloraplants without neighbours was practically identical inthe two Spartina zones For all three species floweringwas stimulated in low zones possibly due to higherconcentrations of exchangeable phosphate (Salisburyamp Ross 1992) or low N P ratios (van Wijnen amp Bakker1999) in these areas Both S alterniflora and S densifloramay benefit from lower flooding stress (Bertness 1991aCastillo et al 2000 Emery et al 2001) or higher nitrogenavailability (Mendelssohn 1979 Levine et al 1998Emery et al 2001) of mid marshes However the resultssuggest that none of the three species experienced physi-ological growth restriction in any vegetation zone ofthe low and mid marshes
The few instances where positive interactions (negativevalues of RCI) were recorded either occurred in theS maritimus zone or were related to growth responses ofS maritimus It is highly improbable that these effectswere related to amelioration of salinity stress by neigh-bouring vegetation (Bertness amp Callaway 1994 Bertness1999 Hacker amp Bertness 1999 Callaway amp Pennings2000 Bortolus et al 2002) because of the prevailing meso-oligohaline conditions (Costa et al 1988 Costa 1997b1998 Seeliger amp Costa 2002) As S alterniflora plantsgrowing beside S maritimus are much more vulnerableto crab grazing than plants in Spartina stands negativevalues of RCI of this species in the S maritimus zonecannot be related to the association offering defenceagainst herbivory (Bertness amp Ellison 1987 Davy et al2000 Pennings amp Bertness 2001) S maritimus tillershave well-developed aerenchyma (Kantrud 1996) andoxidized iron is common near their roots (C S B Costapersonal observation) Small patches of S alternifloracannot efficiently oxidize sediments of low marshes(Bertness 1991a) and amelioration of low oxygen levelsby other species can generate positive interaction betweenmarsh plants at low marsh elevations (Castellanos et al1994 Pennings amp Bertness 2001 Figueroa et al 2003)
Most of the interactions were competitive with Salterniflora and S densiflora demonstrating reductionin tiller density and maximum height live aerial bio-mass and proportion of flowering tillers (in the case ofS densiflora) when surrounded by other plants The lackof significant negative effects on S maritimus could inpart be an experimental artefact related to the shortduration of the experiment (6 months) and the distinc-tive phenological cycles of S maritimus and Spartinaspecies in southern Brazil On average about 41ndash52of the reduction in live aerial biomass of Spartina speciescan be accounted for by intra or interspecific competi-tion The negative interspecific responses (competition)are consistent with observations of growth inhibitionby the presence of other plants (S densiflora Kittelson
amp Boyd 1997 S alterniflora Bertness amp Ellison 1987Bertness 1991a Emery et al 2001 Pennings et al 2003)
Consequently at least for Spartina species intra-and interspecific effects although strong and consistentappear to be symmetrical with no marked variationin competitive intensity between vegetation zones Theevaluation of statistical differences in the magnitudeof intra- or interspecific competitive effects whichwould be reflected in significant interactions betweenthe effects of competition and zone was biased by thezone-dependent crab grazing (within the S maritimuszone) Plant size (maximum height and aerial biomass)which is severely reduced in both species of Spartina byChasmagnathus grazing is a very important componentof competitive ability (Keddy 1989 Bertness 1999) Acompetitive hierarchy does not however appear to bea major process structuring plant community of theseirregularly flooded salt marshes where the dominantspecies show considerable physiological tolerance It isparsimonious to suggest that selective processes shouldfavour stress tolerators as the dominant species of the midmarsh (eg S densiflora) and that competition shouldoccur at all elevations Indeed the mid marsh domin-ant S densiflora can grow vigorously on low marshes(Costa 1997a Kittelson amp Boyd 1997) within its phys-iological constraints of submergence time and mini-mum redox potential which are not very different fromthose of low marsh dominants such as Spartina foliosa(Kittelson amp Boyd 1997) and Spartina maritima(Castillo et al 2000)
S maritimus (in less saline estuaries Ranwell 1974Podlejski 1982 Kantrud 1996 Costa 1998) and S den-siflora (Costa 1997a Castillo et al 2000 Bortolus et al2002) are species capable of acting as salt marsh pio-neers and persisting in more mature communitiesThese two species are certainly generalists and aggres-sive invaders (Figueroa amp Castellanos 1988 Kantrud1996 Kittelson amp Boyd 1997 Castillo et al 2000) thattogether with S alterniflora can monopolize habitatswhether physically stressful or more benign Under thelocal conditions of the low and mid marshes whichfavour meso-oligohaline species and weaken the deter-ministic role of intertidal elevation on flooding stressnone of the three species can be defined as competitivelysubordinate or dominant We therefore concur withEmery et al (2001) that salt marsh stress tolerators canalso be good competitors
The predictably of an environmental stress is adominant factor for specialization (Jefferies et al 1979Davy et al 2000) and for establishmentmaintenanceof a competitive hierarchy (with dominant and sub-ordinate species) (Keddy 1989) The occurrence of a par-ticular situation every day and distinct scenarios eachyear means that local dominance at Patos Lagoonmay be determined more by a lottery mechanism or by
963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

963Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
establishment of propagules and subsequent pre-emption of resources than by stress tolerance and com-petition as in North Atlantic salt marshes (Bertness1991a 1991b 1999) Although events occurring duringthe earliest establishment phases of the life history canbe of primary importance in regulating plant speciesdistribution along environmental gradients (Keddy 1989Davy et al 2000) seedlings and vegetative propagulesof all three studied species are able to colonize low marshmud flats (Kantrud 1996 Copertino et al 1997 Kittelsonamp Boyd 1997 Costa 1998 Castillo et al 2000 Bortoluset al 2002) and similar proportions of survivors ofSpartina species (35ndash50) were observed 2 years aftertransplantation to southern Brazil mud flats (Costa ampMarangoni 1997) Alternative models considering theimportance of founder effects on community structurein salt marshes have already been suggested by Bertnessamp Ellison (1987) Scholten et al (1987) and Levine et al(1998) and their application to irregularly flooded saltmarshes needs to be investigated Additionally as con-sumers can modify the competitive outcome by grazinglocal dominants or newly arrived migrants (Robertson1991 Furbish amp Albano 1994 Taylor et al 1997 Lee 1998)Chasmagnathus granulata could be an important deter-minant of the local dominance of S maritimus or Sdensiflora during simultaneous establishment with Salterniflora or secondary succession after disturbancein the low- and mid-salt marsh of the Patos Lagoonestuary Further studies are necessary to test thesehypotheses
Acknowledgements
We would like to thank Anthony Davy Paul AdamLindsay Haddon and the anonymous referees whosecomments have greatly improved the manuscriptMark Bertness and Bryan Silliman provided helpfulcomments on earlier drafts of the manuscript The workwas supported by CNPq (Brazilian Government) grants20047088ndash4 PDE (CSBC) 52212494ndash1 (AMGA)and a post-graduation fellowship (JCM)
References
Adam P (1993) Saltmarsh Ecology Cambridge UniversityPress New York
Allen SE Grimshaw HM Parkinson JA Quarmby Camp Roberts JD (1976) Chemical analysis Methods in PlantEcology (ed SB Chapman) pp 411ndash466 BlackwellScientific Oxford
Baldwin AH amp Mendelssohn IA (1998) Effects of salinityand water level on coastal marshes an experimental test ofdisturbance as a catalyst for vegetation change AquaticBotany 61 255ndash268
Bertness MD (1991a) Zonation of Spartina patens andSpartina alterniflora in a New England salt marsh Ecology72 138ndash148
Bertness MD (1991b) Interspecific interactions among highmarsh perennials in a New England salt marsh Ecology72 125ndash137
Bertness MD (1999) The Ecology of Atlantic ShorelinesSinauer Associates Sunderland Massachusetts
Bertness MD amp Callaway R (1994) Positive interactions incommunities Trends in Ecology and Evolution 9 191ndash193
Bertness MD amp Ellison AM (1987) Determinants of pat-tern in a New England salt marsh plant communityEcological Monographs 57 (2) 129ndash147
Bertness MD amp Leonard GH (1997) The role of positiveinteractions in communities lessons from intertidalhabitats Ecology 78 1976ndash1989
Bortolus A amp Iribarne O (1999) Effects of the SW Atlanticburrowing crab Chasmagnathus granulata on a Spartina saltmarsh Marine Ecology Progress Series 178 79ndash88
Bortolus A Schwindt E amp Iribarne O (2002) Positive plantndashanimal interactions in the high marsh of an Argentineancoastal lagoon Ecology 83 733ndash742
Brower JE amp Zar JH (1977) Field and Laboratory Methodsfor General Ecology Wm C Brown Dubuque
Burgers G (1999) The El Nintildeo stochastic oscillator ClimateDynamics 15 521ndash531
Callaway RM amp Pennings SC (2000) Facilitation maybuffer competitive effects indirect and diffuse interactionsamong salt marsh plants American Naturalist 156 (4)416ndash424
Capitoli RR Bemvenuti CE amp Gianuca NM (1977)Occurrence and bio-ecologic observations on Metasesarmarubripes crab in the estuarine region of Lagoa dos PatosAtlacircntica (Rio Grande) 2 (1) 50ndash62
Cargill SM amp Jefferies RL (1984) The effects of grazing bylesser snow geese on the vegetation of a sub-arctic saltmarsh Journal of Applied Ecology 21 669ndash686
Castellanos EM Figueroa ME amp Davy AJ (1994)Nucleation and facilitation in saltmarsh successioninteractions between Spartina maritima and Arthrocnemumperenne Journal of Ecology 82 239ndash248
Castillo JM Fernandez-Baco L Castellanos EM Luque CJFigueroa ME amp Davy AJ (2000) Lower limits of Spar-tina densiflora and S maritima in a Mediterranean saltmarsh determined by different ecophysiological tolerancesJournal of Ecology 88 801ndash812
Cavalieri AJ (1983) Proline and glycinebethaine accumula-tion by Spartina alterniflora Loisel in response to NaCl andnitrogen in a controlled environment Oecologia 57 20ndash24
Christian RR Bryant WL Jr amp Brinson MM (1990)Juncus roemerianus production and decomposition alonggradients of salinity and hydroperiod Marine EcologyProgress Series 68 137ndash145
Coelho CAS Uvo CB amp Ambrizzi T (2002) Exploringthe impacts of the tropical Pacific SST on the precipitationpatterns over South America during ENSO periodsTheoretical and Applied Climatology 71 185ndash197
Copertino M Costa CSB amp Seeliger U (1997) Dinacircmicapopulacional de Spartina alterniflora em pacircntanos salgadosdo estuaacuterio da Lagoa dos Patos Rio Grande RS Annals ofVIII Seminaacuterio Regional de Ecologia (ed JE Santos)pp 295ndash312 UFSCarPrograma de Poacutes-Graduaccedilatildeo emEcologia e Recursos Renovaacuteveis Satildeo Carlos Brazil
Costa CSB (1997a) Tidal marshes and wetlands Sub-tropical Convergence Environments the Coast and Sea in theWarm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 24ndash26 Springer-VerlagBerlin
Costa CSB (1997b) Irregularly flooded marginal marshesSubtropical Convergence Environments the Coast and Sea inthe Warm-Temperate Southwestern Atlantic (eds U SeeligerC Odebrecht amp JP Castello) pp 73ndash77 Springer-VerlagBerlin
Costa CSB (1998) Production ecology of Scirpus maritimusin southern Brazil Ciecircncia E Cultura (SBPC) 50 (4) 273ndash280
Costa CSB amp Marangoni JC (1997) Criaccedilatildeo de marismascom espeacutecies nativas experimento piloto no sul do Brasil
964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

964C S B Costa J C Marangoni amp A M G Azevedo
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Annals of VII Congresso Latino Americano de Ciecircncias DoMar Volume I (eds AM Paiva Filho amp RR Weber)pp 200ndash201 Instituto Oceanograacutefico da Universidade deSatildeo PauloAssociaccedilatildeo Latino-americana de Investigadoresem Ciecircncias do Mar Santos Brazil
Costa CSB Seeliger U amp Kinas PG (1988) The effect ofwind velocity and direction on the salinity regime in thelower Patos Lagoon estuary Ciecircncia E Cultura (SBPC) 40(9) 909ndash912
Costa CSB Seeliger U Oliveira CPL amp Mazo AMM(1997) Distribuiccedilatildeo funccedilotildees e valores das marismas epradarias submersas no estuaacuterio da Lagoa dos Patos (RSBrasil) Atlacircntica (Rio Grande) 19 67ndash85
DrsquoIncao F Silva KG Ruffino ML amp Braga AC (1990)Haacutebito alimentar do caranguejo Chasmagnathus granulataDana 1851 na Barra do Rio Grande RS (DecapodaGrapsidae) Atlacircntica (Rio Grande) 12 (2) 85ndash93
Daehler CC amp Strong DR (1995) Impact of high herbivoredensities on introduced smooth cordgrass Spartina alterniflorainvading San Francisco Bay California Estuaries 18 409ndash417
Davy AJ Costa CSB Yallop AR Proudfoot AM ampMohamed MF (2000) Biotic interactions in plant communi-ties of saltmarshes British Saltmarshes (eds BR SherwoodBG Gardiner amp T Harris) pp 109ndash127 Linnean Societyof London London
Eleuterius LN amp Eleuterius CK (1979) Tide levels and saltmarsh zonation Bulletin of Marine Science 29 (3) 394ndash400
Emery NC Ewanchuk PJ amp Bertness MD (2001) Com-petition and salt-marsh plant zonation stress toleratorsmay be dominant competitors Ecology 82 2471ndash2485
Figueroa ME amp Castellanos EM (1988) Vertical structureof Spartina maritima and Spartina densiflora in mediter-ranean marshes Plant Form and Vegetation Structure(eds MJA Werger PJM van der Aart HJ During ampJTA Verhoeven) pp 105ndash108 SPB Academic The Hague
Figueroa ME Castillo JM Redondo S Luque TCastellanos EM Nieva FJ et al (2003) Facilitated inva-sion by hybridization of Sarcocornia species in a salt-marshsuccession Journal of Ecology 91 616ndash626
Furbish CE amp Albano M (1994) Selective herbivory andplant community structure in a Mid-Atlantic salt marshEcology 75 1015ndash1022
Garcia AM Vieira JP amp Winemiller KO (2001) Dynamicsof shallow-water fish assemblage of the Patos Lagoonestuary (Brazil) during cold and warm ENSO episodesJournal of Fish Biology 59 1218ndash1238
Grace JB (1995) On the measurement of plant competitionintensity Ecology 76 305ndash308
Hacker SD amp Bertness MD (1999) Experimental evidencefor the factors maintaining plant species diversity in a NewEngland salt marsh Ecology 80 2064ndash2073
Holm GO Jr Sasser CE Peterson GW amp Swenson EM(2000) Vertical movement and substrate characteristics ofoligohaline marshes near a high-sediment riverine systemJournal of Coastal Research 16 164ndash171
Iribarne O Bortolus A amp Botto F (1997) Between-habitatdifferences in burrow characteristics and trophic modes inthe southeastern Atlantic burrowing crab Chasmagnathusgranulata Marine Ecology Progress Series 155 137ndash145
Jefferies RL (1977) Growth responses of coastal halophytesto inorganic nitrogen Journal of Ecology 65 847ndash865
Jefferies RL Davy AJ amp Rudmik T (1979) The growthstrategies of coastal halophytes Ecological Processes inCoastal Environments (eds RL Jefferies amp AJ Davy)pp 243ndash268 Blackwell Scientific Oxford
Kantrud HA (1996) The Alkali (Scirpus maritimus L) andSaltmarsh (S Robustus Pursh) Bulrushes a LiteratureReview Information and Technology Report 6 NationalBiological Service Jamestown North Dakota httpwwwnpwrcusgsgovresource literatrbulrushbulrushhtm(last update 16 July 1997)
Keddy PA (1989) Competition Chapman amp Hall LondonKittelson PM amp Boyd MJ (1997) Mechanisms of expansion
for an introduced species of cordgrass Spartina densiflorain Humboldt Bay California Estuaries 20 770ndash778
Lee SY (1998) Ecological role of grapsid crabs in mangroveecosystem a review Marine and Freshwater Research 49335ndash343
Lessmann JM Mendelssohn IA Hester MW ampMcKee KL (1997) Population variation in growth responseto flooding of three marsh grasses Ecological Engineering8 31ndash47
Levine JM Brewer JS amp Bertness MD (1998) Nutrientscompetition and plant zonation in a New England saltmarsh Journal of Ecology 86 285ndash292
Lightfoot DC amp Whitford WG (1987) Variation in insectdensities on desert creosotebush is nitrogen a factorEcology 68 547ndash557
Louda SM Keeler KH amp Holt RD (1990) Herbivoreinfluences on plant performance and competitive inter-actions Perspectives on Plant Competition (eds JB Graceamp D Tilman) pp 414ndash444 Academic Press San Diego
Mendelssohn IA (1979) The influence of nitrogen level formand application method on the growth response of Spartinaalterniflora North Carolina Estuaries 2 (2) 106ndash112
Moumlller OO Castaing P Dalomon J amp Lazure P (2001)The influence of local and non-local forcing effects on thesubtidal circulation of Patos Lagoon Estuaries 24 (2)297ndash311
NOAA (2000) Climate of 1999 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch1999annann99html (last update 12 January 2000)
NOAA (2001) Climate of 2000 ndash Annual Review NationalClimatic Data CenterNOAA Homepage Asheville NorthCarolina http lwfncdcnoaagovoaclimateresearch2000annannhtml (last update 12 January 2001)
Odum WE (1988) Comparative ecology of tidal freshwaterand salt marshes Annual Review of Ecology and Systematics19 147ndash176
Paludan C amp Morris JT (1999) Distribution and speciationof phosphorus along a salinity gradient in intertidal marshsediment Biogeochemistry 45 197ndash221
Parsons KA amp de La Cruz A (1980) Energy flow andgrazing behavior of concephaline grasshoppers in a Juncusroemerianus marsh Ecology 61 1045ndash1050
Pennings SC amp Bertness MD (1999) Using latitudinal vari-ation to examine effects of climate on coastal salt marshpattern and process Current Topics in Wetland Biogeochemistry3 100ndash111
Pennings SC amp Bertness MD (2001) Salt marsh commu-nities Marine Community Ecology (eds MD BertnessSD Gaines amp ME Hay) pp 289ndash316 Sinauer AssociatesSunderland Massachusetts
Pennings SC amp Callaway RM (1992) Salt marsh plantzonation the relative importance of competition andphysical factors Ecology 73 681ndash690
Pennings SC Carefoot TH Siska EL Chase ME ampPage TA (1998) Feeding preferences of a generalist salt-marsh crab relative importance of multiple plant traitsEcology 79 1968ndash1979
Pennings SC amp Moore DJ (2001) Zonation of shrubs inwestern Atlantic salt marshes Oecologia 126 587ndash594
Pennings SC Selig ER Houser LT amp Bertness MD (2003)Geographic variation in positive and negative interactionsamong salt marsh plants Ecology 84 1527ndash1538
Pielou EC amp Routledge RD (1976) Salt marsh vegetationlatitudinal gradients in the zonation patterns Oecologia24 311ndash321
Podlejski V (1982) Phenology and seasonal above-groundbiomass in two Scirpus maritimus marshes in the CamargueFolia Geobotanica and Phytotaxonomica (Praha) 17 225ndash236
965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003

965Irregularly flooded marsh
copy 2003 British Ecological Society Journal of Ecology 91 951ndash965
Portnoy JW amp Giblin AE (1997) Effects of historic tidalrestrictions on salt marsh sediment chemistry Biogeochemistry36 275ndash303
Ranwell DS (1974) The salt marsh to tidal woodland tran-sition Hydrobiological Bulletin (Amsterdam) 8 139ndash151
Robertson AI (1991) Plantndashanimal interactions and thestructure and function of mangrove forest ecosystemsAustralian Journal of Ecology 16 433ndash443
Rogel JA Ariza FA amp Silla RO (2000) Soil salinity andmoisture gradients and plant zonation in Mediterraneansalt marshes of southeast Spain Wetlands 20 357ndash372
Rozema J Scholten MCT Blaauw PA amp Diggelen J(1988) Distribution limits and physiological toleranceswith particular reference to the salt marsh environmentPlant Population Ecology (eds AJ Davy MJ Hutchings ampAR Watkinson) pp 137ndash164 Blackwell Scientific Oxford
Russell PJ Flows TJP amp Hutchings MJ (1985) Compar-ison of niche breadths and overlaps of halophytes on saltmarshes of differing diversity Vegetatio 61 171ndash178
Salisbury FB amp Ross CW (1992) Plant PhysiologyWadsworth San Diego
Santos M amp Almeida SL (2000) Anatomia foliar de SpartinaSchreber (Poaceae) da Ilha de Santa Catarina (FlorianopolisSC) Insula (Florianopolis) 29 1ndash24
Scholten M Blaauw PA Stroetenga M amp Rozema J(1987) The impact of competitive interactions on the growthand distribution of plant species in salt marshes VegetationBetween Land and Sea (eds AHL Huiskes CWPM Blomamp J Rozema) pp 270ndash281 Dr Junk Publishers Dordrecht
Seeliger U amp Costa CSB (2002) The Patos-Mirim basinslagoons and estuary south Brazil natural and human forc-ing factors South American Basins Loicz Global ChangeAssessment and Synthesis of River CatchmentndashCoastal SeaInteraction and Human Dimensions LOICZ Reports ampStudies 21 (eds LD Lacerda HH Kremer B KjerveW Salomons JI Marshall amp CJ Crossland) pp 105ndash112LOICZIPO Texel the Netherlands
Seeliger U Costa CSB amp Abreu PC (1997) Energy flowand habitats in the Patos Lagoon estuary SubtropicalConvergence Environments the Coast and Sea in the Warm-Temperate Southwestern Atlantic (eds U Seeliger C Odebrechtamp JP Castello) pp 65ndash70 Springer-Verlag Berlin
Siegel S (1975) Estatiacutestica Natildeo-Parameacutetrica Para as CiecircnciasDo Comportamento McGraw-Hill Satildeo Paulo
Silliman BR amp Bertness MD (2002) A trophic cascade re-gulates salt marsh primary production Proceedings of theNational Academy of Sciences (USA) 99 10500ndash10505
Silliman BR amp Zieman JC (2001) Top-down control ofSpartina alterniflora production by periwinkle grazing in aVirginia salt marsh Ecology 82 2830ndash2845
Silva CP Pereira CMP amp Dorneles LPP (1993) Espeacuteciesde gramiacuteneas e crescimento de Spartina densiflora Brongem uma marisma da Laguna dos Patos RS Brasil Cadernode Pesquisa Seacuterie Botanica (Santa Cruz Do Sul) 5 95ndash108
Snow AA amp Vince SW (1984) Plant zonation in anAlaskan salt marsh II an experimental study of the role ofedaphic conditions Journal of Ecology 72 669ndash684
Stewart GR Larher F Ahmad I amp Lee JA (1979) Nitro-gen metabolism and salt-tolerance in higher plant halo-phytes Ecological Processes in Coastal Environments(eds RL Jefferies amp AJ Davy) pp 211ndash227 BlackwellScientific Oxford
Stout JP (1984) The Ecology of Irregularly Flooded SaltMarshes of Northeastern Gulf of Mexico a Community Pro-file Biological Report 85(71) Fish and Wildlife ServiceMinerals Management ServiceUS Department of InteriorWashington
Taylor KL Grace JB amp Marx BD (1997) The effects ofherbivory on neighbour interactions along a coastal marshgradient American Journal of Botany 84 709ndash715
Vince SW amp Snow AA (1984) Plant zonation in an Alaskansalt marsh I Distribution abundance and environmentalfactors Journal of Ecology 72 651ndash667
Wang FC (1997) Dynamics of intertidal marshes nearshallow estuaries in Lousiana Wetlands Ecology andManagement 5 131ndash143
van Wijnen HJ amp Bakker JP (1999) Nitrogen and phos-phorus limitation in a coastal barrier salt marsh theimplications for vegetation succession Journal of Ecology87 265ndash272
Williams AC amp McCarthy BC (2001) A new index of inter-specific competition for replacement and additive designsEcological Research 16 29ndash40
Wolcott DL amp OrsquoConnor NJ (1992) Herbivory in crabsadaptations and ecological considerations AmericanZoologist 32 370ndash381
Zar JH (1984) Biostatistical Analysis Prentice HallEnglewood Cliffs New York
Zedler JB Callaway JC Desmond JS Vivian-Smith GWilliams GD Sullivan G et al (1999) Californian salt-marsh vegetation an improved model of spatial patternEcosystems 2 19ndash35
Zhou J amp Lau KM (2001) Principal modes of interannualand decadal variability of summer rainfall over SouthAmerica International Journal of Climatology 21 1623ndash1644
Received 13 August 2002 revision accepted 21 May 2003




















