
Floral resources and hummingbirds on an island of flooded forest in Central Brazil
9
Flora 206 (2011) 827–835 Contents lists available at ScienceDirect Flora j ourna l h omepage: www.elsevier.de/flora Floral resources and hummingbirds on an island of flooded forest in Central Brazil Francielle Paulina de Araújo, Ana Angélica Almeida Barbosa, Paulo Eugênio Oliveira ∗ Instituto de Biologia, Programa de Pós-Graduac ¸ ão em Ecologia e Conservac ¸ ão de Recursos Naturais, Universidade Federal de Uberlândia, Cx.P. 593, Cep 38400-902, Uberlândia, MG, Brazil a r t i c l e i n f o Article history: Received 13 October 2010 Accepted 17 February 2011 Keywords: Hummingbird-pollinated flowers Cerrado Energy supply Nectar Ornithophilous species Carrying capacity a b s t r a c t Nectar availability for hummingbirds was studied for two and a half years in an island of flooded forest in Uberlândia, MG, in the Cerrado region, the Neotropical savanna areas in Central Brazil. Flowering indi- viduals of ten plant species from eight families that offered resources potentially used by hummingbirds were recorded along transects crisscrossing the area, which were surveyed every other week. The num- ber of flowers was estimated for each plant. The volume and concentration of nectar were measured for a sample of flowers of each species, and the data were used to estimate the caloric content available per flower and the general resource availability in the area throughout the study period. Focal observations of hummingbird visits were carried out on blooming individuals of all plant species. Hummingbird flowers occurred throughout the study period between January 2001 and June 2003. Only three of the studied plants did not present clear ornithophilous features. Six hummingbird species were recorded in the area, but total nectar availability in the area was seldom sufficient for the daily energy requirements of a sin- gle bird. Therefore, it can be inferred from the study that these hummingbirds need to move among the mosaic of different plant physiognomies in the region in order to obtain enough energy for their survival. These movements may also contribute to increasing gene flow and regeneration of plant populations isolated in forest patches as the one studied here. © 2011 Elsevier GmbH. All rights reserved. Introduction The Angiosperms present different kinds of interactions with animals, and in many cases, have mutual adaptations associated with pollination (Bawa, 1990; Engel and Irwin, 2003; Taura and Laroca, 2004). Plants have flowers morphologically adjusted to attract different vectors as well as to facilitate the access to nec- tar or other floral rewards while inducing pollen transport, which is vital for their reproduction (Fenster et al., 2004; Stiles, 1981). Such floral morphological adjustments and adaptations are directed toward specific functional groups of pollinators and may be viewed as adaptive pollination syndromes (Faegri and van der Pijl, 1980; Fenster et al., 2004). Flowers adapted for hummingbird pollination are usually tubular, with bright and contrasting colours, odourless, with diurnal anthesis and abundant nectar (Faegri and van der Pijl, 1980; Rocca and Sazima, 2010; Stiles, 1981; Wilson et al., 2004). Although these birds are not exclusively nectarivo- rous and can search for nectar also in flowers that are pollinated by other animals (Araujo and Sazima, 2003; Buzato et al., 2000; Sick, 1997; Wilson et al., 2004), their mutualism with flowers seems to be vital for their survival (Stiles, 1995). ∗ Corresponding author. E-mail address: [email protected] (P.E. Oliveira). Hummingbirds are exclusive of the American continent, with 83 species in Brazil, and 36 occurring in the Cerrado, the tropical savanna region in central Brazil (Macedo, 2002). They belong to the family Trochilidae, which can be subdivided into two sub-families: Phaethornithinae and Trochilinae (Bleiweiss et al., 1994; Sick, 1997; Stiles, 1981). These small birds visit flowers for nectar and have long and/or curved bills, which fit the size/shape of their flowers. Moreover, they are agile in looking for pendent flowers and have spatial memory as well as other specializations to explore floral resources (Hurly, 1996; Hurly and Healy, 1996; Sick, 1997; Stiles, 1981). Although nectar is not the only food source used by humming- birds, it provides most of the carbohydrates that supply the energy these birds need for continuous flight (Brice and Grau, 1991; Sick, 1997). Foraging behaviour and number of flowers visited are influ- enced by the amount of available nectar in the area (Rojas and Ribon, 1997; Wolf et al., 1976). In order to get enough energy, these mostly nectarivorous birds must forage all day long, visiting hundreds of flowers (Stiles, 1995). Therefore, hummingbirds are important for the maintenance of pollen flow. In some tropical ecosystems where plants depend on cross pollination and pollen transfer, these birds are particularly important pollinating agents (Araujo and Sazima, 2003; Buzato et al., 2000; Freitas and Sazima, 2001; Gobatto-Rodrigues and Stort, 1992; Oliveira, 1998). 0367-2530/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.flora.2011.04.001
Transcript of Floral resources and hummingbirds on an island of flooded forest in Central Brazil

F
FIC
a
ARA
KHCENOC
I
awLati
dbPpovero1b
0d
Flora 206 (2011) 827– 835
Contents lists available at ScienceDirect
Flora
j ourna l h omepage: www.elsev ier .de / f lora
loral resources and hummingbirds on an island of flooded forest in Central Brazil
rancielle Paulina de Araújo, Ana Angélica Almeida Barbosa, Paulo Eugênio Oliveira ∗
nstituto de Biologia, Programa de Pós-Graduac ão em Ecologia e Conservac ão de Recursos Naturais, Universidade Federal de Uberlândia, Cx.P. 593,ep 38400-902, Uberlândia, MG, Brazil
r t i c l e i n f o
rticle history:eceived 13 October 2010ccepted 17 February 2011
eywords:ummingbird-pollinated flowerserradonergy supplyectarrnithophilous species
a b s t r a c t
Nectar availability for hummingbirds was studied for two and a half years in an island of flooded forestin Uberlândia, MG, in the Cerrado region, the Neotropical savanna areas in Central Brazil. Flowering indi-viduals of ten plant species from eight families that offered resources potentially used by hummingbirdswere recorded along transects crisscrossing the area, which were surveyed every other week. The num-ber of flowers was estimated for each plant. The volume and concentration of nectar were measured fora sample of flowers of each species, and the data were used to estimate the caloric content available perflower and the general resource availability in the area throughout the study period. Focal observations ofhummingbird visits were carried out on blooming individuals of all plant species. Hummingbird flowersoccurred throughout the study period between January 2001 and June 2003. Only three of the studied
arrying capacity plants did not present clear ornithophilous features. Six hummingbird species were recorded in the area,but total nectar availability in the area was seldom sufficient for the daily energy requirements of a sin-gle bird. Therefore, it can be inferred from the study that these hummingbirds need to move among themosaic of different plant physiognomies in the region in order to obtain enough energy for their survival.These movements may also contribute to increasing gene flow and regeneration of plant populationsisolated in forest patches as the one studied here.
ntroduction
The Angiosperms present different kinds of interactions withnimals, and in many cases, have mutual adaptations associatedith pollination (Bawa, 1990; Engel and Irwin, 2003; Taura and
aroca, 2004). Plants have flowers morphologically adjusted tottract different vectors as well as to facilitate the access to nec-ar or other floral rewards while inducing pollen transport, whichs vital for their reproduction (Fenster et al., 2004; Stiles, 1981).
Such floral morphological adjustments and adaptations areirected toward specific functional groups of pollinators and maye viewed as adaptive pollination syndromes (Faegri and van derijl, 1980; Fenster et al., 2004). Flowers adapted for hummingbirdollination are usually tubular, with bright and contrasting colours,dourless, with diurnal anthesis and abundant nectar (Faegri andan der Pijl, 1980; Rocca and Sazima, 2010; Stiles, 1981; Wilsont al., 2004). Although these birds are not exclusively nectarivo-ous and can search for nectar also in flowers that are pollinated by
ther animals (Araujo and Sazima, 2003; Buzato et al., 2000; Sick,997; Wilson et al., 2004), their mutualism with flowers seems toe vital for their survival (Stiles, 1995).∗ Corresponding author.E-mail address: [email protected] (P.E. Oliveira).
367-2530/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.flora.2011.04.001
© 2011 Elsevier GmbH. All rights reserved.
Hummingbirds are exclusive of the American continent, with83 species in Brazil, and 36 occurring in the Cerrado, the tropicalsavanna region in central Brazil (Macedo, 2002). They belong to thefamily Trochilidae, which can be subdivided into two sub-families:Phaethornithinae and Trochilinae (Bleiweiss et al., 1994; Sick, 1997;Stiles, 1981). These small birds visit flowers for nectar and havelong and/or curved bills, which fit the size/shape of their flowers.Moreover, they are agile in looking for pendent flowers and havespatial memory as well as other specializations to explore floralresources (Hurly, 1996; Hurly and Healy, 1996; Sick, 1997; Stiles,1981).
Although nectar is not the only food source used by humming-birds, it provides most of the carbohydrates that supply the energythese birds need for continuous flight (Brice and Grau, 1991; Sick,1997). Foraging behaviour and number of flowers visited are influ-enced by the amount of available nectar in the area (Rojas andRibon, 1997; Wolf et al., 1976). In order to get enough energy,these mostly nectarivorous birds must forage all day long, visitinghundreds of flowers (Stiles, 1995).
Therefore, hummingbirds are important for the maintenance ofpollen flow. In some tropical ecosystems where plants depend on
cross pollination and pollen transfer, these birds are particularlyimportant pollinating agents (Araujo and Sazima, 2003; Buzatoet al., 2000; Freitas and Sazima, 2001; Gobatto-Rodrigues and Stort,1992; Oliveira, 1998).
8 lora 20
baCaM1ta1ifma
nMflmap
M
S
(cMU1
awSfc“egtri
awM((tUSpao
dSfttoas
28 F.P. Araújo et al. / F
The interaction between hummingbirds and their flowers haseen widely studied in Brazil and elsewhere in the Americas (Araujond Sazima, 2003; Buzato et al., 2000; Canela and Sazima, 2003;astellanos et al., 2003, 2004; Coelho and Barbosa, 2004; Fariand Araujo, 2010; Freitas and Sazima, 2001; Gonzáles et al., 2005;endonc a and Anjos, 2005; Rocca and Sazima, 2008; Sazima et al.,
996; Temeles et al., 2002). The studies of the particularities ofhese interactions help elucidate ecological patterns and evolution-ry aspects (Buzato et al., 2000; Grant, 1994; Simberlof and Dayan,991), while contributing to clarify the importance of the organ-
sms in the maintenance of plant communities (Bawa, 1990). Butew studies in Brazil and South America as a whole tried to esti-
ate resource offer and carrying capacity of hummingbird-plantssemblages (e.g., Cotton, 2007).
The present study focused on a hummingbird-flower commu-ity that occurs on an island of flooded gallery forest in Uberlândia,G, Brazil. The specific objectives were: (1) to investigate plant
owering phenology and the availability of floral resources to hum-ingbirds; (2) identify the species of hummingbirds that use the
vailable floral resources; (3) and analyze if this forest island canrovide enough energy for year round hummingbird survival.
aterials and methods
tudy site
The study was carried out in an island of flooded gallery forestsensu Ribeiro and Walter, 1998) in a palm swamp in the ecologi-al reserve of “Clube Cac a e Pesca Itororó de Uberlândia” (CCPIU),G, Brazil. The CCPIU comprises a 127 ha natural area located inberlândia, Minas Gerais state, Southeastern Brazil (Lima et al.,989).
The predominant vegetation is cerrado (sensu stricto) but therea is divided by a long stretch of Mauritia flexuosa palm swamp inhich two isolated forest patches are embedded (Appolinario and
chiavini, 2002). The present study was conducted in the largerorest island (18◦59′21′′S and 48◦18′06′′W), an irregularly ellipti-al area of ca. 2 ha, inside the palm swamp that is drained by theCabeceira do Lageado” stream. This forest island presents a cleardge being surrounded by swamp comprised of herbaceous, mostlyrassland, vegetation. The interior of the forest has at least 5 m fromhe border a markedly shaded understory. The distance to open cer-ado areas is less than 50 m and to the nearest other forest patch its less than 1 km.
The climate in the region has two well-defined seasons: a drynd cooler season between April and September, and a rainy andarmer one from October to March (Nimer and Brandão, 1989).ore than 70% of the rains fall between November and March
Cardoso et al., 2009). The specific climatic data for the study periodtotal rainfall and average temperatures; Fig. 1) were obtained fromhe Climatology and Water Resources Laboratory of the Federalniversity of Uberlândia–MG (Meteorological station of Campusanta Mônica). The climate can be classified as Aw in the Köp-en scale with higher temperatures in the summer (above 35 ◦C),nd only occasional frosts in winter (Rosa et al., 1991). But frostsccurred in June 2001 and September 2002 during the study period.
Despite being a preserved area, the occurrence of forest firesuring the dry season has been often reported in CCPIU. On 29eptember 2002, sometime after the above mentioned frost, aorest fire occurred in the reserve that reached a great part ofhe cerrado (sensu stricto), palm swamp area and the border of
he studied forest. Both frosts and fires have transient impactsn the Cerrado biome. They usually destroy inflorescences, leavesnd branches of trees and shrubs, and increase mortality of somepecies (Hoffmann and Solbrig, 2003; Silberbauer-Gottsberger6 (2011) 827– 835
et al., 1977). But even strong forest fires have limited impact on theforest structures (Santiago and Silva-Lima, 2005) and their effectsare not much perceptible already some months after the event.
Sampling method
Plant species were surveyed every other week from January2001 to June 2003 by counting the number of open flowers avail-able on each day along transects. The transects used had differentlength: 147 m, 100 m, 100 m, 83 m, 60 m, 40 m and 35 m, crossingthe forest, 40 m apart from each other. The surveys included bloom-ing individuals, occurring 2 m on each side of the transects, of allputatively ornithophilous species (Faegri and van der Pijl, 1980) andalso some species of different pollination syndromes that presentednectar and floral morphology which would attract hummingbirds.The species recognised as potentially used by hummingbirds arereferred in this text as hummingbird flowers and included the trulyornithophilous or not.
Vouchers of each flowering species observed were collected anddried according to standard protocols (Fidalgo and Bononi, 1984).They were incorporated in the collection of “Herbarium Uberlan-dense” (HUFU) of the Institute of Biology of the Federal Universityof Uberlândia.
Floral morphology
The floral morphology was described from flowers preservedin 70% alcohol. The length and the largest diameter of the corollawere recorded from a sample of four to 15 flowers of each species.As length of the corolla tube was considered the effective length(from the base to the opening; sensu Wolf et al., 1976), and averageand standard deviation were calculated for the measurements ofeach species. Some other floral characteristics, which could be lostafter fixation, such as colour and odour, were recorded in the field.
Hummingbirds and foraging behaviour
The species of hummingbirds existing in the study area wererecorded by focal observations carried out on hummingbird plantsblooming along the transects, mostly in the morning and duringthe whole study period. These observations were made in contin-uous sessions that lasted 1–3 h. Each plant species was observedfor at least 3 h in total, amounting to 119 h of focal observation.The visited plant species, the frequency of visits and the number ofvisited flowers were recorded. Identification of the hummingbirdswas made in the field, based on identification keys and descrip-tions (Grantsau, 1989), and some species were photographed whilevisiting flowers. The hummingbirds were classified based on theirforaging behaviour (territorial, generalists, thieves and trapliners)according to Feinsinger and Colwell (1978).
Floral resources
Nectar production was measured in flowers bagged beforeopening. The volume of nectar for each flower was measured usingglass micropipettes, sugar concentration was obtained using a handrefractometer. Nectar data were taken at the end of the morninghours. The average nectar volume and concentration were used toestimate the amount of reward in calories per flower. Brix refrac-tometric measures, in g sugar/g solution, were converted into mgsugar/�L of nectar (Galetto and Bernadello, 2005). We considered1 mg sugar = 4 calories (Dafni, 1992; Galetto and Bernadello, 2005).
Caloric reward per flower and the number of flowers sampled pro-vided an instantaneous estimate of the resource in the area foreach month of the study period. Although sugars are the majorconstituents of nectar, other solutes affect refractive index and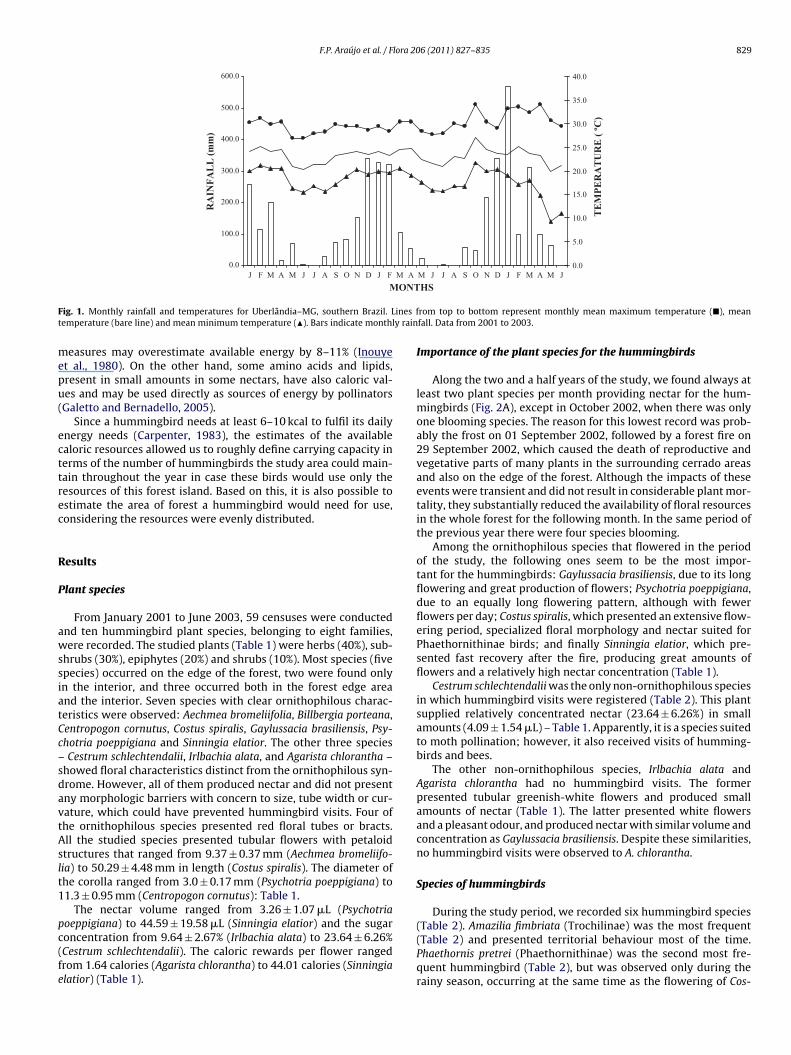
F.P. Araújo et al. / Flora 206 (2011) 827– 835 829
0.0
100.0
200.0
300.0
400.0
500.0
600.0
JMAMFJDNOSAJJMAMFJDNOSAJJMAMFJMONTHS
RA
INFA
LL
(mm
)
0.0
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
TE
MPE
RA
TU
RE
( ºC
)
F ines ft y rain
mepu(
ecttrec
R
P
awssiatCc–sdavtAslt1
pc(fe
ig. 1. Monthly rainfall and temperatures for Uberlândia–MG, southern Brazil. Lemperature (bare line) and mean minimum temperature (�). Bars indicate monthl
easures may overestimate available energy by 8–11% (Inouyet al., 1980). On the other hand, some amino acids and lipids,resent in small amounts in some nectars, have also caloric val-es and may be used directly as sources of energy by pollinatorsGaletto and Bernadello, 2005).
Since a hummingbird needs at least 6–10 kcal to fulfil its dailynergy needs (Carpenter, 1983), the estimates of the availablealoric resources allowed us to roughly define carrying capacity inerms of the number of hummingbirds the study area could main-ain throughout the year in case these birds would use only theesources of this forest island. Based on this, it is also possible tostimate the area of forest a hummingbird would need for use,onsidering the resources were evenly distributed.
esults
lant species
From January 2001 to June 2003, 59 censuses were conductednd ten hummingbird plant species, belonging to eight families,ere recorded. The studied plants (Table 1) were herbs (40%), sub-
hrubs (30%), epiphytes (20%) and shrubs (10%). Most species (fivepecies) occurred on the edge of the forest, two were found onlyn the interior, and three occurred both in the forest edge areand the interior. Seven species with clear ornithophilous charac-eristics were observed: Aechmea bromeliifolia, Billbergia porteana,entropogon cornutus, Costus spiralis, Gaylussacia brasiliensis, Psy-hotria poeppigiana and Sinningia elatior. The other three species
Cestrum schlechtendalii, Irlbachia alata, and Agarista chlorantha –howed floral characteristics distinct from the ornithophilous syn-rome. However, all of them produced nectar and did not presentny morphologic barriers with concern to size, tube width or cur-ature, which could have prevented hummingbird visits. Four ofhe ornithophilous species presented red floral tubes or bracts.ll the studied species presented tubular flowers with petaloidtructures that ranged from 9.37 ± 0.37 mm (Aechmea bromeliifo-ia) to 50.29 ± 4.48 mm in length (Costus spiralis). The diameter ofhe corolla ranged from 3.0 ± 0.17 mm (Psychotria poeppigiana) to1.3 ± 0.95 mm (Centropogon cornutus): Table 1.
The nectar volume ranged from 3.26 ± 1.07 �L (Psychotriaoeppigiana) to 44.59 ± 19.58 �L (Sinningia elatior) and the sugar
oncentration from 9.64 ± 2.67% (Irlbachia alata) to 23.64 ± 6.26%Cestrum schlechtendalii). The caloric rewards per flower rangedrom 1.64 calories (Agarista chlorantha) to 44.01 calories (Sinningialatior) (Table 1).rom top to bottom represent monthly mean maximum temperature (�), meanfall. Data from 2001 to 2003.
Importance of the plant species for the hummingbirds
Along the two and a half years of the study, we found always atleast two plant species per month providing nectar for the hum-mingbirds (Fig. 2A), except in October 2002, when there was onlyone blooming species. The reason for this lowest record was prob-ably the frost on 01 September 2002, followed by a forest fire on29 September 2002, which caused the death of reproductive andvegetative parts of many plants in the surrounding cerrado areasand also on the edge of the forest. Although the impacts of theseevents were transient and did not result in considerable plant mor-tality, they substantially reduced the availability of floral resourcesin the whole forest for the following month. In the same period ofthe previous year there were four species blooming.
Among the ornithophilous species that flowered in the periodof the study, the following ones seem to be the most impor-tant for the hummingbirds: Gaylussacia brasiliensis, due to its longflowering and great production of flowers; Psychotria poeppigiana,due to an equally long flowering pattern, although with fewerflowers per day; Costus spiralis, which presented an extensive flow-ering period, specialized floral morphology and nectar suited forPhaethornithinae birds; and finally Sinningia elatior, which pre-sented fast recovery after the fire, producing great amounts offlowers and a relatively high nectar concentration (Table 1).
Cestrum schlechtendalii was the only non-ornithophilous speciesin which hummingbird visits were registered (Table 2). This plantsupplied relatively concentrated nectar (23.64 ± 6.26%) in smallamounts (4.09 ± 1.54 �L) – Table 1. Apparently, it is a species suitedto moth pollination; however, it also received visits of humming-birds and bees.
The other non-ornithophilous species, Irlbachia alata andAgarista chlorantha had no hummingbird visits. The formerpresented tubular greenish-white flowers and produced smallamounts of nectar (Table 1). The latter presented white flowersand a pleasant odour, and produced nectar with similar volume andconcentration as Gaylussacia brasiliensis. Despite these similarities,no hummingbird visits were observed to A. chlorantha.
Species of hummingbirds
During the study period, we recorded six hummingbird species(Table 2). Amazilia fimbriata (Trochilinae) was the most frequent
(Table 2) and presented territorial behaviour most of the time.Phaethornis pretrei (Phaethornithinae) was the second most fre-quent hummingbird (Table 2), but was observed only during therainy season, occurring at the same time as the flowering of Cos-
830F.P.
Araújo
et al.
/ Flora
206 (2011) 827– 835
Table 1Putative hummingbird flowers and some of their attributes in a forest island in the CCPIU, Uberlândia/MG Brazil. HUFU (Herbario Universidade Federal de Uberlândia).
Plant species Growthform
Occurrence Flowershape
Flowercolour
Corolla length(mean mm ± SD)
Corolla diameter(mean mm ± SD)
Nectar volume (mean�L ± SD)
Sugar concentration(x% ± SD)
Caloric reward perflower (calories)
Herbarium registrationnumber at HUFU
CostaceaeCostus spiralis(Jacq.) Roscoe
Herb Facultative Tubular Red 50.29 ± 4.48(n = 15)
10.64 ± 1.64 (n = 15) 9.13 ± 4.03 (n = 12) 19.96 ± 8.3 (n = 12) 7.86 33301
SolanaceaeCestrumschlechtendalii G.Don
Subshrub Edge Tubular White 31.4 ± 4.26 (n = 15) 3.29 ± 0.26 (n = 15) 4.09 ± 1.54 (n = 14) 23.64 ± 6.26 (n = 14) 4.22 33305
GentianaceaeIrlbachia alata(Aubl.) Maas.
Herb Edge Tubular Greenish-white
29.9 ± 2.51 (n = 15) 10.1 ± 1.1 (n = 15) 9.04 ± 3.04 (n = 14) 9.64 ± 2.67 (n = 14) 3.61 33303
GesneriaceaeSinningia elatior(Kunth) Chantems
Herb Edge Tubular Orange 37.75 ± 2.72(n = 15)
9.1 ± 0.92 (n = 15) 44.59 ± 79.61b (n = 13) 22.59 ± 3.8b (n = 13) 44.01 33306
BromeliaceaeAechmeabromeliifolia(Rudger) Baker
Epiphyte Facultative Tubular Yellow 9.37 ± 0.37 (n = 15) 4 ± 0 (n = 4) 6.59 ± 3.80 (n = 15) 22.03 ± 1.30 (n = 15) 6.344 33309
BromeliaceaeBillbergia porteanaBeer
Epiphyte Interior Tubular Green/lilacand pinka
– – – – – 33304
EricaceaeGaylussaciabrasiliensis
Shrub Edge Tubular Red 15.37 ± 0.47(n = 15)
5.57 ± 0.16 (n = 15) 3.62 ± 1.23 (n = 21) 13.5 ± 5.23 (n = 21) 1.98 33300
(Spreng.) Meisn.EricaceaeAgarista chlorantha(Cham.) G. Don.
Subshrub Edge Tubular White 11.39 ± 0.29(n = 15)
5.3 ± 0.16 (n = 15) 1.82 ± 0.64 (n = 17) 20.8 ± 8.75 (n = 17) 1.64 33721
RubiaceaePsychotriapoeppigiana Müll.Arg.
Subshrub Interior Tubular Yellow/Reda 15.38 ± 1.26(n = 12)
3.0 ± 0.17 (n = 12) 3.26 ± 1.07 (n = 11) 23.14 ± 2.46 (n = 11) 3.31 33302
CampanulaceaeCentropogoncornutus (L.) Druce
Herb Interior Tubular Red 42.65 ± 1.07 (n = 5) 11.3 ± 0.95 (n = 5) – – – 33307
– Data not available.a Bract or calyx colour.b Nectar accumulated more than two days.

F.P. Araújo et al. / Flora 206 (2011) 827– 835 831
Fig. 2. (A) Flowering period and intensity (percentage of flowering individuals given by line thickness) of hummingbird-flowers. Plant species names: Siel, Sinningia elatior;Cosp, Costus spiralis; Iral, Irlbachia alata; Bipo, Billbergia porteana; Gabr, Gaylussacia brasiliensis; Pspo, Psychotria poeppigiana; Aebr, Aechmea bromeliifolia; Agch, Agaristachlorantha; Cesc, Cestrum schlechtendalii; Ceco, Centropogon cornutus. (B) Phenology of hummingbirds (records per month – varied from zero to seven and given by relativel ilia fimc D) Esti il on J
tabrdifi
ta(wC
ine thickness). Hummingbird species names: Phpr, Phaethornis pretrei; Amfi, Amazhrysura. (C) Number of available flowers for hummingbirds, recorded fortnightly. (sland of flooded gallery forest in CCPIU reserve, Uberlândia–MG, southeastern Braz
us spiralis. This species returned to the flowers at regular intervalsnd direction, indicating foraging routes and possibly trapliningehaviour. Along the study period the number of hummingbirdegistered varied widely, although they seem to be more frequenturing the rainy seasons (Fig. 2B). The number of hummingbird
ndividuals observed concomitantly in the area varied from one tove.
Chlorostilbon lucidus (Trochilinae) was observed in the areahroughout the year, but its visits to the flowers were not regular
nd territorial behaviour was less frequent. Eupetomena macrouraTrochilinae) was observed only in the months when resourcesere higher, and it always presented territorial behaviour (Table 2).olibri serrirostris (Trochilinae) was observed all year long, but
briata; Chlu, Chlorostilbon lucidus; Euma, Eupetomena macroura; Hych, Hylocharisimation of available energy per hectare (in calories) on hummingbird plants, in ananuary 2001 to June 2003. Months in gray refer to rainy season.
it rarely visited the plants in the interior or on the forest edge.Hylocharis chrysura (Trochilinae) was spotted only once, visitingtwo different plant species (Gaylussacia brasiliensis and Sinningiaelatior).
Available resources throughout the year
As mentioned above, in the absence of factors such as fire andfrost affecting the vegetation, there was an apparently continuous
flowering of the species used by hummingbirds in the community.Peaks of flower production occurred during both the dry and rainyseason, and they did not occur in the same periods in the differentyears studied (Fig. 2A and B). In 2001, the greatest flower produc-
832 F.P. Araújo et al. / Flora 206 (2011) 827– 835
Table 2Number of hummingbird visits per hour of observation to plant species in an island of flooded gallery forest at CCPIU Uberlândia–MG, southeastern Brazil.
Plant species Hummingbird species
Amfi Chlu Euma Phpr Hych Cose Hours of observation
Psychotria poeppigiana 19.42 1.05 – 0.18 – – 22 h 45 minGaylussacia brasiliensis 6.95 14.15 – 0.22 0.72 X 27 h 46 minCostus spiralis 0.1a 0.03a 0.41 5.71 – X 36 h 16 minSinningia elatior 8.67 – 18.58 0.25 0.5 X 12 hAgarista chlorantha – – – – – – 5 h 20 minAechmea bromeliifolia – – – – – – 3hCestrum schlechtendalii – 13.25 – – – – 4 hBillbergia porteana – – – – – – –Centropogon cornutus – – – – – – 3 h 30 minIrlbachia alata – – – – – – 5 h
A EumaC
tda
yottflC22c
aw2itt(rftooanwfoiwamirmibic
D
P
b
bbreviations of bird names: Amfi, Amazilia fimbriata; Chlu, Chlorostilbon lucidus;
olibri serrirostris. X – non-quantified visits. – no visits recorded.a Nectar robbery only.
ion occurred in the second fortnight of September; in 2002, fourifferent peaks occurred in January, March, August and December;nd in 2003, two peaks occurred in January and February.
These peaks of flowering occurred at different periods of eachear as a result of the contribution of different species. The first peakf flower production, registered September 2001, was caused byhe flowering of Gaylussacia brasiliensis. In January 2002, it was dueo Costus spiralis, though a great amount of Gaylussacia brasiliensisowers were still available. March 2002 the flowering species wereostus spiralis, Irlbachia alata, and Gaylussacia brasiliensis, August002 it was Agarista chlorantha. The flowering peak of December002 was a result of the intense Sinningia elatior flowering, whichontinued up to February 2003 (Fig. 2A).
The estimates of the nectar caloric content indicated that thevailable nectar for the hummingbirds per hectare in the study areaas almost always below the daily requirements of a single bird. In
001, the maximum available energy value, 7.2 kcal/ha, occurredn the second fortnight of September (Fig. 2C). It was due mostlyo the great number of Gaylussacia brasiliensis flowers available athat period. In the other months, the amount of available energyvia nectar) per hectare was always inferior to the minimum energyequirements of a hummingbird. The lowest offer occurred in firstortnight of January and July and in second fortnight of June, whenhere were no flowering plant individuals. There was a frost in sec-nd fortnight of July (28 July) which affected blooming individualsf G. brasiliensis and Costus spiralis and, although it was not as strongs the one that occurred in the following year, reduced to zero theectar offer. In 2002, the minimum required energy for a single birdas never reached (Fig. 2C). The lowest amount of 0.07 kcal/ha was
ound in first fortnight of July, and a maximum of 5.77 kcal/ha wasbtained in second fortnight of March. In 2003, energy amount var-ed from a minimum of 0.149 kcal/ha to a maximum of 8.6 kcal/ha,
ith peaks during the months of January, February, March and Aprils a result of Sinningia elatior flowering. These values, even if esti-ated for 2 ha, which corresponded to the total area of the forest
sland, would be barely enough to keep up with the minimal energyequirements of couples of resident hummingbirds, even during theonths with the greatest nectar availability. Considering the min-
mum and maximum energy offer, a single bird would need to useetween 0.7 and 81 ha of forest to get the nectar for its daily needs,
f the resources were evenly distributed, which is certainly not thease for the patchy distributed forest areas of the Cerrado region.
iscussion
lant species
The number of plant species that could be used by humming-irds in the studied forest (10 species) was the same as that found in
, Eupetomena macroura; Phpr, Phaethornis pretrei; Hych, Hylocharis chrysura; Cose,
high grassland outcrops, the “campos rupestres”, at “Serra do Cur-ral” (Vasconcelos and Lombardi, 1999) and in the gallery forest ofthe Panga Ecological Station (Oliveira, 1998), both in the BrazilianCerrado region. The species richness in the studied area was com-parable to some other open plant communities in Brazil, as coastalRestinga plains (Ormond et al., 1993). However, when compared tocommunities of the Atlantic Forest (see Buzato et al., 2000; Lopes,2002; Rocca and Sazima, 2008; Sazima et al., 1996), the number ofhummingbird plants in our study was two or three times smaller,even including non-ornithophilous species. The additional obser-vation of non-ornithophilous flowers in studies concerning theavailability of floral resources is important because hummingbirds,especially those belonging to the subfamily Trochilinae, are com-mon visitors also of flowers of non-ornithophilous plants (Araujoand Sazima, 2003; Oliveira and Gibbs, 2000). In the studied com-munity, however, we observed only one non-ornithophilous plantspecies receiving hummingbird visits. This contrasts with the for-est type “capões” of the Pantanal, for example, where only sixout of 18 plant species visited by hummingbirds possessed clearornithophilous characteristics (Araujo and Sazima, 2003). Amongwood species in a Cerrado area near Brasília, only one out of 11plant species visited by hummingbirds, was truly ornithophilous(Oliveira and Gibbs, 2000).
Hummingbirds are opportunistic flower visitors, which canbehave as pollinators but also as thieves, interfering nega-tively in the reproduction of various non-ornithophilous plants(Navarro, 1999). In any case, the nectar resources provided bothby ornithophilous and non-ornithophilous plants are important forhummingbird survival.
The relative importance of plant species for hummingbirds
In the community studied, Gaylussacia brasiliensis has to beconsidered an important species when it comes to the availabil-ity of floral resources, because not only does it occur practicallyall along the forest edge, but it also flowers almost continuously.Its flowering was interrupted only by the frost/fire episodes. Thiscontinuous flowering (sensu Newstrom et al., 1994), practically allyear long, helps to keep the pollinators in the area. This speciesattracted hummingbirds of the subfamily Trochilinae (Amazilia fim-briata and Chlorostilbon lucidus) which presented either territorialor opportunistic behaviour. The interaction between the densityof the plants and the amount of flowers that they produce proba-bly defines the foraging behaviour of the birds (Franceschinelli andBawa, 2000; Justino et al., 2011).
Psychotria poeppigiana is an ornithophilous species that presentsa pattern of annual flowering of the extended type (sensu Newstromet al., 1994), which is not linked to rainfall (Coelho and Barbosa,2004). In this sense, Psychotria poeppigiana was the only species

lora 20
tt
oPnroThL
ttEdo
iAoyfo
H
wma(Rfaohsftd2
Phrts
odBppsbw
A
i(fia
F.P. Araújo et al. / F
hat offered steady resources for hummingbirds in the interior ofhe forest.
Costus spiralis, the flowering pattern of which is also annual andf the extended type (sensu Newstrom et al., 1994), can attracthaethornis pretrei throughout the rainy season. This species offersectar in flowers with long and curved floral tubes, adapted toeceive visits from Phaethornithinae and to exclude visits fromther hummingbirds with shorter bills (Araújo and Oliveira, 2007).his association between Costus spiralis and Phaethornithinae birdsas been observed also in Atlantic Forest areas (Buzato et al., 2000;opes, 2002).
The occurrence of hummingbird species seems to depend onhe availability of floral resources, which characterizes an oppor-unistic foraging, as observed elsewhere (Araujo and Sazima, 2003).upetomena macroura, e.g., which was rarely observed in the areauring most of the study period, became one of the most frequentlybserved species during the intense flowering of Sinningia elatior.
Different from other tropical communities, there was no dom-nant ornithophilous plant family in the studied area. In thetlantic Forest, for example, the Bromeliaceae are the dominantrnithophilous family, with many species flowering throughout theear (Buzato et al., 2000). In the censuses carried out in the studiedorest, however, we observed only two sparsely distributed speciesf bromeliads.
ummingbird species
The species richness of hummingbirds recorded in this studyas relatively low when compared with other Neotropical com-unities such as the Atlantic forest (see Buzato et al., 2000; Rocca
nd Sazima, 2008; Sazima et al., 1996), the Colombian AmazoniaLasprilla and Sazima, 2004), and the Subtropical Forest of Costaica (Stiles, 1985). However, it was similar to the number of species
ound in the dry forests of Mexico (Arizmendi and Ornelas, 1990),nd higher than the number of species found in the forest “capões”f the Brazilian Pantanal (Araujo and Sazima, 2003) and in a tropicaligh-altitude grassland area in Brazil (Freitas and Sazima, 2006). Iteems that such forest areas primarily function as refuges for theauna in the Cerrado region and movements between plant forma-ions may be responsible for the maintenance of species richness ofifferent groups of birds (Gottsberger and Silberbauer-Gottsberger,006; Macedo, 2002).
Amazilia fimbriata, Chlorostilbon lucidus (territorial species) andhaethornis pretrei (trapliner) were the most important species ofummingbirds in the studied community. The presence of territo-ial species and trapliners is an important aspect of the diversity ofropical nectarivorous birds and results in a stable coexistence ofpecies that use similar resources (Arizmendi and Ornelas, 1990).
The other species recorded in the area can be considered rarer accidental (sensu Stiles, 1985). Although it was not possible toivide the community into sub-communities, as in richer areas (seeuzato et al., 2000; Rocca and Sazima, 2008; Stiles, 1985), it wasossible to identify a most frequent hummingbird species for eachlant. In this sense, Phaethornis pretrei was associated with Costuspiralis, Amazilia fimbriata with Psychotria poeppigiana, Chlorostil-on lucidus with Gaylussacia brasiliensis, and Eupetomena macrouraith Sinningia elatior.
vailable resources throughout the year
The flowering phenology of the ornithophilous species stud-
ed here can be classified as belonging to the extended typesensu Newstrom et al., 1994), as most of them flowered for overve months with a small daily flower production. This extendednd somewhat sequential flowering (Newstrom et al., 1994) may6 (2011) 827– 835 833
increase pollination assurance and maintain hummingbird pollina-tors in the area.
It is difficult to isolate the determining factors of the pheno-logical patterns of tropical plant communities (Bawa, 1990), butabiotic factors such as precipitation and photoperiod (Morellatoet al., 1990), and biotic factors such as interactions between plantsand animals likely influence the reproductive strategies of plantsof the Cerrado communities (Oliveira and Gibbs, 2002). Compar-ing the flowering periods of the species used by hummingbirdsin this study with those found in cerrado (sensu stricto) surround-ing the palm swamp (A.F. Melazzo, unpublished data), there wasa peak of availability of floral resources in January (rainy season)in these open savanna formations, while in the studied forest, thispeak occurred in September (end of the dry season). This contrast ofresources availability may provide the opportunity for humming-birds to migrate and use floral resources in this mosaic of plantformations (Barbosa and Sazima, 2008; Oliveira and Gibbs, 2000,2002).
The relatively low caloric value available for the hummingbirdsin the study area is another factor that may force the hummingbirdsto use a wider spectrum of plant formations. It seems that bothsavanna and forest areas rarely provide the minimum amount ofnectar necessary to maintain resident hummingbirds over the year(Gottsberger and Silberbauer-Gottsberger, 2006; Oliveira, 1998).Even if we consider that our estimates may reflect only the “imme-diate” volume of nectar, since the values showed the accumulatedvolume during the morning only, floral resource limitation mayforce the use of resources outside of the forest island.
This study reinforces the idea that in the Cerrado biome, floralresources occur in a mosaic, where physiognomical different plantformations like forests, palm swamps and cerrado (sensu stricto),mostly different in floristic composition and phenology (Oliveira,1998; Ribeiro and Walter, 1998), complement each other to main-tain the pollinators in the area throughout the year (Oliveira andGibbs, 2002). Limited in size and resource offer, forest patches as theforest island studied here, provide shelter and will function as com-plementary areas for hummingbirds in the matrix of open savannaformations, which dominates the Cerrado region. Markedly sea-sonal climate and less predictable but common occurrence of frostsand forest fires can completely change resource availability overtime.
Changes in resource availability probably drive hummingbirdmovements among plant physiognomies. Resource tracking, bothspatial and seasonal, has been shown for hummingbirds in othertropical habitats with much less seasonality than the Cerrado, aslowland rainforest (Cotton, 2007). As far as we know, there is nopopulation study for Brazilian hummingbirds, but observations andcolour tagging of P. petrei in another 40 ha forest area in the regionsuggest that this population should comprise less than ten breedingindividuals (Araújo, 2010). But the resource offer in the forest patchstudied here would be low even for populations of such limitedsize. Drought and disturbance have been shown to affect popula-tion size and mobility of tropical hummingbirds. Severe droughtin tropical forest areas may affect overall resource availability andhummingbird population sizes (Stiles, 1992). But the kind of chang-ing availability of resources observed in our study area seems tocreate a mosaic of habitats for the birds which would be more com-parable to transient forest gaps, where patchy distribution of thevegetation leads to mobility and complex changes in hummingbirdassemblages (Feinsinger et al., 1988).
Resource distribution in a mosaic vegetation, separated in timeand space, makes the animals to be mobile links between plant
populations and environmental sub-units (Sekercioglu, 2006). Ithelps maintaining and regenerating plant populations isolated inthe studied forest island (Araújo and Oliveira, 2007). Hence, theresults presented here imply that the conservation of the char-
8 lora 20
apm
A
oeCR
R
A
A
A
A
A
B
B
B
B
B
C
C
C
C
C
C
C
D
E
F
F
F
F
F
F
F
34 F.P. Araújo et al. / F
cteristic mosaic of plant formations, and not only of the legallyrotected gallery forests, is vital for maintenance of active hum-ingbird populations and their plants in the Cerrado Biome.
cknowledgments
We thank Jane Ogilvie and Jessica Forrest for revising the Englishf the manuscript, We also thank Marlies Sazima and the review-rs for valuable suggestions “Cac a e Pesca Itororó de Uberlândia”lub (CCPIU) for permitting our access to the reserve and Brazilianesearch Council – CNPq for supporting this study.
eferences
ppolinario, V., Schiavini, I., 2002. Levantamento Fitossociológico de Espéciesarbóreas de Cerrado (stricto sensu) em Uberlândia – Minas Gerais. Bol. Herb.Ezechias Paulo Heringer 10, 57–75.
raújo, F.P., 2010. A comunidade de plantas ornitófilas da Estac ão Ecológicado Panga: o fluxo de pólen intermediado pelo beija-flor Phaethornis pre-trei (Phaethornithinae). Doctoral Thesis, Universidade Estadual de Campinas,Campinas-Brazil.
raújo, F.P., Oliveira, P.E., 2007. Biologia floral de Costus spiralis (Jacquin) Roscoe(Costaceae) e mecanismos para evitar a autopolinizac ão. Rev. Brasil. Bot. 30,61–70.
raujo, A.C., Sazima, M., 2003. The assemblage of flowers visited by hummingbirdsin the “capões” of southern Pantanal, Mato Grosso do Sul. Brazil. Flora 189, 1–9.
rizmendi, M.C., Ornelas, J.F., 1990. Hummingbirds and their floral resources in atropical dry forest in Mexico. Biotropica 22, 172–180.
arbosa, A.A.A., Sazima, M., 2008. Biologia reprodutiva de plantas herbáceo-arbustivas de uma área de campo sujo de cerrado. In: Sano, S.M., Almeida, S.P.,Ribeiro, J.F. (Eds.), Cerrado Ecologia e Flora. Embrapa Cerrados, Brasília, DF, pp.291–318.
awa, K.S., 1990. Plant-pollinator interactions in a tropical rain forest. Annu. Rev.Ecol. Syst. 21, 399–422.
leiweiss, R., Kirsch, J.A.W., Matheus, J.C., 1994. DNA–DNA Hybridization evidencefor subfamily structure among hummingbirds. Auk 111, 8–19.
rice, A.T., Grau, R.C., 1991. Protein requirements of Costa’s Hummingbirds Calyptecostae. Physiol. Zool. 64, 611–626.
uzato, S., Sazima, M., Sazima, I., 2000. Hummingbird-pollinated floras at threeAtlantic forest sites. Biotropica 32, 824–841.
anela, M.B.F., Sazima, M., 2003. Aechmea pectinata: a hummingbird-dependentBromeliad with inconspicuous flowers from the Rainforest in South-easternBrazil. Ann. Bot. 92, 731–737.
ardoso, E., Moreno, M.I.C., Bruna, E.M., Vasconcelos, H.L., 2009. Mudanc as fitofi-sionômicas no cerrado: 18 anos de sucessão ecológica na Estac ão Ecológica doPanga, Uberlândia–MG. Caminhos de Geografia 10 (32), 254–268.
arpenter, F.L., 1983. Pollination energetics in avian communities: simple conceptsand complex realities. In: Jones, C.E., Little, R.J. (Eds.), Handbook of ExperimentalPollination Biology. Scientific and Academic Editions, New York, pp. 215–234.
astellanos, M.C., Wilson, P., Thomson, J.D., 2003. Pollen transfer by hummingbirdsand bumblebees, and the divergence of pollination modes in Penstemon. Evolu-tion 57, 2742–2752.
astellanos, M.C., Wilson, P., Thomson, J.D., 2004. ‘Anti-bee’ and ‘pro-bird’ changesduring evolution of hummingbird pollination in Penstemon flowers. J. Evol. Biol.17, 876–885.
oelho, C.P., Barbosa, A.A.A., 2004. Biologia reprodutiva de Psychotria poeppigianaMull Arg. (Rubiaceae) em mata de galeria. Acta Bot. Bras. 18, 481–489.
otton, P.A., 2007. Seasonal resource tracking by Amazonian hummingbirds. Ibis149, 135–142.
afni, A., 1992. Pollination Ecology. In: A Practical Approach. Oxford University Press,Oxford.
ngel, E.C., Irwin, R.E., 2003. Linking pollinator visitation rate and pollen receipt. Am.J. Bot. 90, 1612–1618.
aegri, K., van der Pijl, L., 1980. The Principles of Pollination Ecology. Pergamon Press,Oxford.
aria, R.R., Araujo, A.C., 2010. Pollination ecology of Ruellia angustifolia (Ness) Lindauex Rambo (Acanthaceae) in the Serra da Bodoquena, Mato Grosso do Sul. Brazil.Flora 205, 106–111.
enster, C.B., Armbruster, W.S., Wilson, P., Dudash, M.R., Thomson, J.D., 2004. Pol-lination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 35,375–403.
einsinger, P., Colwell, R.D., 1978. Community organization among Neotropicalnectar-feeding birds. Am. Zool. 18, 779–795.
einsinger, P., Busby, W.H., Murray, K.G., Beach, J.H., Pounds, W.Z., Linhart, Y.B., 1988.Mixed support for spatial heterogeneity in species interactions: hummingbirdsin a tropical disturbance mosaic. Am. Nat. 131, 33–57.
idalgo, O., Bononi, V.L.R., 1984. Técnicas de coleta, preservac ão e herbarizac ão dematerial botânico. Instituto de Botânica/Secretaria do Meio Ambiente, São Paulo.
ranceschinelli, E.V., Bawa, K.S., 2000. The effect of ecological factors on the matingsystem of a South American shrub species (Helicteres brevispira). Heredity 84,116–123.
6 (2011) 827– 835
Freitas, L., Sazima, M., 2001. Nectar features in Esterhazya macrodonta, ahummingbird-pollinated Scrophulariaceae in Southeastern Brazil. J. Plant Res.114, 187–191.
Freitas, L., Sazima, M., 2006. Pollination biology in a tropical high-altitude grasslandin Brazil: interactions at the community level. Ann. Miss. Bot. Gard. 93, 465–516.
Galetto, L., Bernadello, G., 2005. Rewards in flowers, Nectar. In: Dafni, A., Kevan,P.G., Husband, B.C. (Eds.), Practical Pollination Biology. Enviroquest, Canada, pp.261–313.
Gonzáles, C., Ornelas, J.F., Jiménez, L., 2005. Between-year changes in functionalgender expression of Palicourea padifolia (Rubiaceae), a distylous, hummingbird-pollinated shrub. Ann. Bot. 95, 371–378.
Grant, V., 1994. Historical development of ornithophily in the western North Amer-ican flora. Proc. Natl. Acad. Sci. U.S.A. 91, 10407–10411.
Grantsau, R., 1989. Os beija-flores do brasil. Expressão e Cultura, Rio de Janeiro.Gobatto-Rodrigues, A.A., Stort, M.N.S., 1992. Biologia floral e reproduc ão de Pyroste-
gia venusta (Ker-Gaw) Miers (Bignoniaceae). Rev. Brasil. Bot. 15, 37–41.Gottsberger, G., Silberbauer-Gottsberger, I., 2006. Life in the Cerrado: A South Amer-
ican Tropical Seasonal Ecosystem. Reta Verlag, Ulm.Hoffmann, W.A., Solbrig, O.T., 2003. The role of topkill in the differential response
of savanna woody species to fire. For. Ecol. Manage. 180, 273–286.Hurly, T.A., 1996. Spatial memory in rufous hummingbirds: memory for reward and
non-rewarded sites. Anim. Behav. 51, 177–183.Hurly, T.A., Healy, S.D., 1996. Spatial memory in rufous hummingbirds: location or
local visual cues? Anim. Behav. 51, 1149–1157.Inouye, D.W., et al., 1980. The effects of nonsugar nectar constituents on estimates
of nectar energy content. Ecology 61, 992–996 (8 authors).Justino, D.G., Maruyama, P.K., Oliveira, P.E., 2011. Floral resource availability and
hummingbird territorial behaviour on a Neotropical savanna shrub. J. Ornithol.,in press.
Lasprilla, L.R., Sazima, M., 2004. Interacciones planta-colibrí en tres comunidadesvegetales de la parte suroriental de Parque Nacional Natural de Chiribiquete,Colombia. Ornit. Neotrop. 15 (Suppl.), 183–190.
Lima, S.C., Rosa, R., Feltran-Filho, A., 1989. Mapeamento do uso do solo no municípiode Uberlândia–MG, através de imagens TM/LANDSAT. Soc. Nat. 1, 127–145.
Lopes, A.V.F. 2002. Polinizac ão por beija-flores em remanescentes da Mata AtlânticaPernambucana, Nordeste do Brasil. Doctoral Thesis, Universidade Estadual deCampinas, Campinas.
Macedo, R.H.F., 2002. Avifauna: ecology, biogeography, and behavior. In: Oliveira,P.S., Marquis, R.J. (Eds.), The Cerrados of Brazil. Columbia University Press, NewYork, pp. 242–265.
Mendonc a, L.B., Anjos, L., 2005. Beija-flores (Aves Trochilidae) e seus recursos floraisem uma área urbana do Sul do Brasil. Rev. Bras. Zool. 22, 51–59.
Morellato, L.P.C., Leitão-Filho, H.F., Rodrigues, R.R., Joly, C.A., 1990. Estratégiasfenológicas de espécies arbóreas em floresta de altitude na Serra do Japi, Jundiaí,São Paulo. Rev. Brasil. Biol. 50, 149–162.
Navarro, L., 1999. Pollination ecology and effect of nectar removal in Macleaniabullata (Ericaceae). Biotropica 31, 618–625.
Newstrom, L.E., Frankie, G.W., Baker, H.G., 1994. A new classification for plant phe-nology based on flowering patterns in lowland tropical rain forest trees at LaSelva, Costa Rica. Biotropica 26, 141–159.
Nimer, E., Brandão, A.M.P.M., 1989. Balanc o hídrico e clima da região dos cerrados.IBGE, Rio de Janeiro.
Oliveira, G.M., 1998. Disponibilidade de recursos florais para beija-flores no cerradode Uberlândia, MG. MSc Dissertation, Universidade de Brasília, Brasília-DF.
Oliveira, P.E., Gibbs, P.E., 2000. Reproductive biology of woody plants in a cerradocommunity of Central Brazil. Flora 195, 311–329.
Oliveira, P.E., Gibbs, P.E., 2002. Pollination and reproductive biology in Cerradoplant communities. In: Oliveira, P.S., Marquis, R.J. (Eds.), The Cerrados of Brazil.Columbia University Press, New York, pp. 329–347.
Ormond, W.T., Pinheiro, M.C.B., Lima, H.A., Correia, M.C.R., Pimenta, M.L., 1993.Estudo das recompensas florais das plantas da Restinga de Maricá – Itaipuac u,RJ. I – Nectaríferas. Bradea 6, 179–195.
Ribeiro, J.F., Walter, B.M.T., 1998. Fitofisionomias do Bioma Cerrado. In: Sano, S.M.,Almeida, S.P. (Eds.), Cerrado: ambiente e flora. EMBRAPA-CPAC, Planaltina, DF,pp. 89–166.
Rocca, M.A., Sazima, M., 2008. Ornithophilous canopy species in the Atlantic rainforest of southeastern Brazil. J. Field Ornithol. 79, 130–137.
Rocca, M.A., Sazima, M., 2010. Beyond hummingbirds-flowers: the other side ofornithophily in the Neotropics. Oecol. Aust. 14, 67–99.
Rojas, R., Ribon, R., 1997. Guilda de aves em Bowdichia virgilioides (Fabaceae:Faboideae) em área de cerrado de Furnas. M. G. Ararajuba 5, 189–194.
Rosa, R., Lima, S.C., Assunc ão, W.L., 1991. Abordagem preliminar das condic õesclimáticas de Uberlândia (MG). Soc. Nat. 3, 91–108.
Santiago, J., Silva Jr., M.C., Lima, L.C., 2005. Fitossociologia da regeneracão arbóreana Mata de Galeria do Pitoco (IBGE-DF), seis anos após fogo acidental. Sci. For.(Piracicaba) 67, 64–77.
Sazima, I., Buzato, S., Sazima, M., 1996. An assemblage of hummingbird-pollinatedflowers in a montane forest in southeastern Brazil. Bot. Acta 109, 149–160.
Sekercioglu, C.H., 2006. Increasing awareness of avian ecological function. TrendsEcol. Evol. 21, 464–471.
Sick, H., 1997. Ornitologia Brasileira. Ed. Nova Fronteira, Rio de Janeiro.
Silberbauer-Gottsberger, I., Morawetz, W., Gottsberger, G., 1977. Frost damage ofCerrado plants in Botucatu Brazil, as related to the geographical distribution ofthe species. Biotropica 9, 253–261.
Simberlof, D., Dayan, T., 1991. The guild concept and the structure of ecologicalcommunities. Ann. Rev. Ecol. Syst. 22, 115–143.

lora 20
S
S
S
S
T
Wilson, P., Castellanos, M.C., Hogue, J.N., Thomson, J.D., Armbruster, W.S., 2004. A
F.P. Araújo et al. / F
tiles, F.G., 1981. Geographical aspects of bird-flower coevolution, with particularreference to Central América. Ann. Miss. Bot. Gard. 68, 323–351.
tiles, F.G., 1985. Seasonal patterns and coevolution in the hummingbird-flower community of a Costa Rican subtropical forest. Ornith. Monogr. 36,757–787.
tiles, F.G., 1992. Effects of a severe drought on the population biology of a tropicalhummingbird. Ecology 73, 1375–1390.
tiles, F.G., 1995. Behavioral, ecological and morphological correlates of foraging forarthropods by the hummingbirds of a tropical wet forest. Condor 97, 853–878.
aura, H.M., Laroca, S., 2004. Biologia da polinizac ão: interac ões entre as abel-has (Hym., Apoidea) e as flores de Vassobia breviflora (Solanaceae). Acta Biol.Paranaense 33, 143–162.
6 (2011) 827– 835 835
Temeles, E.J., Linhart, Y.B., Masonjones, M., Masonjones, H., 2002. The role of flowerwidth in hummingbird bill length-flower length relationships. Biotropica 34,68–80.
Vasconcelos, M.F., Lombardi, J.A., 1999. Padrão sazonal na ocorrência de seis espéciesde beija-flores (Apodiformes: Trochilidae) em uma localidade de campo rupestrena Serra do Curral. Minas Gerais. Ararajuba 7, 71–79.
multivariate search for pollination syndromes among penstemons. Oikos 104,345–361.
Wolf, L.L., Stiles, F.G., Hainsworth, F.R., 1976. Ecological organization of a tropicalhighland hummingbird community. J. Anim. Ecol. 32, 349–379.




















