
Floral Phyllotaxis and Floral Architecture in Calycanthaceae (Laurales)
23
Floral Phyllotaxis and Floral Architecture in Calycanthaceae (Laurales) Author(s): Yannick M. Staedler, Peter H. Weston, and Peter K. Endress Source: International Journal of Plant Sciences, Vol. 168, No. 3 (March/April 2007), pp. 285- 306 Published by: The University of Chicago Press Stable URL: http://www.jstor.org/stable/10.1086/510417 . Accessed: 15/08/2013 15:47 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access to International Journal of Plant Sciences. http://www.jstor.org This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PM All use subject to JSTOR Terms and Conditions
Transcript of Floral Phyllotaxis and Floral Architecture in Calycanthaceae (Laurales)

Floral Phyllotaxis and Floral Architecture in Calycanthaceae (Laurales)Author(s): Yannick M. Staedler, Peter H. Weston, and Peter K. EndressSource: International Journal of Plant Sciences, Vol. 168, No. 3 (March/April 2007), pp. 285-306Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/10.1086/510417 .
Accessed: 15/08/2013 15:47
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toInternational Journal of Plant Sciences.
http://www.jstor.org
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

FLORAL PHYLLOTAXIS AND FLORAL ARCHITECTURE INCALYCANTHACEAE (LAURALES)
Yannick M. Staedler,1,* Peter H. Weston,y and Peter K. Endress*
*Institute of Systematic Botany, University of Zurich, Zollikerstrasse 107, 8008 Zurich, Switzerland;and yRoyal Botanic Gardens and Domain Trust, Mrs Macquaries Road, Sydney,
New South Wales 2000, Australia
Calycanthaceae are sister to the remainder of Laurales (magnoliid clade), an order seemingly showing highvariability in floral phyllotaxis. Previous studies of floral phyllotaxis in this family focused on one species,Calycanthus floridus. Therefore, a comparative study of floral phyllotaxis, comprising species of all four generaof the family, was undertaken. Floral phyllotaxis is spiral according to the Fibonacci pattern for all speciesstudied. The more or less extensive transition zone from the vegetative decussate to the floral spiral phyllotaxiscomprises three to 19 organs. There is usually a gradual transition between floral organ types, especiallybetween different functional parts of the perianth and between stamens, staminodes, and carpels. Thetransition between staminodes and carpels sometimes shows irregularities in organ sequence along theontogenetic spiral. There is a tendency to form series of organs of the same kind in Fibonacci numbers. Thecaedehisce via laterally hinged valves in Sinocalycanthus but via simple longitudinal slits in all other genera. Foodbodies are present on the stamens and inner tepals of Calycanthus and Sinocalycanthus but differ in extent. Allpreviously described calycanthaceous fossil flowers (Araripia, Virginianthus, Jerseyanthus) appear to showspiral floral phyllotaxis. In the three-dimensionally preserved Virginianthus and Jerseyanthus, arrangement ofandroecium organs in series of Fibonacci numbers appears to be present.
Keywords: Calycanthaceae, floral phyllotaxis, organ series, Fibonacci spiral, anthers, food body.
Introduction
Calycanthaceae, a small and distinctive family of basal an-giosperms comprising four genera and 10 species (Zhou et al.2006) of temperate shrubs or tropical trees, have relativelylarge flowers (fig. 1), which are pollinated by beetles, thrips,or flies (Grant 1950; Badrutt 1992; Vogel 1998; Worboysand Jackes 2005). The fossil record of reproductive structuresof Calycanthaceae is relatively abundant (Mai 1987; Friiset al. 1994; Herendeen et al. 1999; Mohr and Eklund 2003;Crepet et al. 2005) and goes back to the early Albian (Friiset al. 1994). Some of these fossils are extremely well pre-served (Crepet et al. 2005). Early studies recognized the affin-ity of Calycanthaceae to Monimiaceae s.l. and Lauraceae(Baillon 1868). This affinity was acknowledged in the ma-jor classification systems of the past 50 yr (Cronquist 1957,1981, 1988; Takhtajan 1959, 1987, 1997; Thorne 1974,1992, 2000; Dahlgren and Bremer 1985). Molecular phy-logenetic analyses (Renner 1998, 1999, 2004; Qiu et al.1999, 2005; Soltis et al. 2000) settled the family as sister tothe rest of Laurales. These analyses also divided the re-maining Laurales into two clades of three families each: theSiparunaceae-Atherospermataceae-Gomortegaceae clade (85species; Renner 2004) and the Monimiaceae-Hernandiaceae-Lauraceae clade (2750–3250 species; Renner 2004). Within
Calycanthaceae, Idiospermum is sister to the rest of the family,and Chimonanthus is sister to the Calycanthus-Sinocalycanthusclade (Li et al. 2004; Zhou et al. 2006).
From accounts on various groups of Laurales, it appearsthat their floral phyllotaxis is diverse. Floral phyllotaxisis, however, well documented only for some Hernandiaceae,Lauraceae, Hortonia of Monimiaceae, and Calycanthus flor-idus of Calycanthaceae. Taxa studied in Hernandiaceae(Shutts 1959; Kubitzki 1969; Endress and Lorence 2004) andLauraceae (Singh and Singh 1985; Endress 1987; Hyland1989) show whorled phyllotaxis (exceptions with irregularphyllotaxis in few Lauraceae; Endress 1990). Hortonia(Monimiaceae) (Endress 1980a), C. floridus (Calycanthaceae)(Unger 1852; Baillon 1868; Eichler 1878; Prantl 1889; Erbarand Leins 1983), and Chimonanthus praecox (Calycantha-ceae; only perianth studied; Nelson 1954) show spiral phyl-lotaxis. Information about floral phyllotaxis of the otherfamilies of the order is scanty and sometimes conflicting.
Floral phyllotaxis has been mapped on cladograms ofbasal angiosperms, either for the perianth (Zanis et al. 2003)or for the whole flower (Doyle and Endress 2000; Ronse DeCraene et al. 2003; Endress and Doyle 2007). These analysesshow several transitions from spiral to whorled floral phyllo-taxis and vice versa: in the branch leading to Nymphaeales,at the base of eudicots, at the base of the magnoliid clade,within Magnoliales, and within Laurales. Calycanthaceae, atthe base of Laurales, are thus in a position close to such atransition. But because former studies of Calycanthaceae wererestricted to a single species, C. floridus, and because basal
1 Author for correspondence; telephone 41-44-634-8432; fax 41-
44-634-8403; e-mail [email protected].
Manuscript received July 2006; revised manuscript received September 2006.
285
Int. J. Plant Sci. 168(3):285–306. 2007.
� 2007 by The University of Chicago. All rights reserved.
1058-5893/2007/16803-0003$15.00
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

angiosperms often show variability of floral phyllotaxis at vari-ous taxonomic levels (see Endress 1987; Doust 2001), a studyon all genera of Calycanthaceae is needed.
Floral phyllotaxis often differs from vegetative phyllotaxisin a plant. This change, usually discrete in flowers with feworgans, may be more gradual in flowers with numerous or-gans. The notion of floral merism has usually been appliedfor whorled flowers. However, in spiral flowers, floral organsalso tend to occur in preferred numbers and tend to appearin series. A series is a set of organs of the same kind, rela-tively regularly surrounding the floral center. A series of aspiral flower corresponds to some extent to a whorl of awhorled flower, but it appears less regular because the neigh-boring organs are not equidistant (in contrast to a whorl).Organs of an organ series appear in a number belonging tothe mathematical series used to define the type of spiral phyl-lotaxis. In flowers showing Fibonacci spiral phyllotaxis, thesenumbers belong to the Fibonacci mathematical series (1, 2,3, 5, 8, 13, 21, . . .), as has been described from some basal-most angiosperms (Endress 2001). In previous literature, theterm ‘‘Kreis’’ (Hirmer 1931) and the expression ‘‘Fibonaccicycles’’ (Nelson 1954) were used for series as we considerthem. We prefer the term ‘‘series’’ because ‘‘Kreis’’ and ‘‘cy-cle’’ are sometimes also used for a whorled organization. Theterm ‘‘cycle’’ implies either a whorl or one full turn on theontogenetic spiral. However, the series represents more thanone turn (approximately one and a half turns for five organs,approximately two and a half turns for eight organs, etc.).
The average angle between two consecutively initiated or-gans of a series (a, the divergence angle) in a spiral system islinked to the mathematical series used to describe the systemby the relation a ¼ limn�!‘360�ðUn=Unþ1Þ, where Un is amember of the relevant mathematical series. For Fibonaccispiral phyllotaxis, the average divergence angle, also calledthe golden angle, is 137.50. . .�.
Calycanthaceae flowers have a floral cup, with differentorgans inserted at different sites on the cup. The perianth is in-serted on the outer surface of the cup, up to its rim. Stamens areusually inserted on the rim, staminodes on the inside of the cup,and carpels (topographically) below the staminodes and at thebottom of the cup. This organization is present in all describedspecies (Baillon 1868; Diels 1912; Rauh and Reznik 1951;Cheng and Chang 1963; Nicely 1965; Blake 1972; Dengler1972; Wilson 1976). Other floral parameters vary substantiallyin the family in response to pollination ecology, from quite largebeetle-pollinated trap flowers to fly-pollinated, somewhat moreopen flowers (Delpino 1870; Grant 1950; Vogel 1998).
In this study, six species from all four genera of Calycantha-ceae (two species each of Calycanthus and Chimonanthus andthe sole species of Sinocalycanthus and Idiospermum) arestudied and compared in their floral architecture, floral phy-llotaxis (including vegetative to floral phyllotaxis transition),and occurrence of organ series. We also focus on perianthand androecium structure, whereas gynoecium structure willbe treated in a future study. Systematic aspects within the fam-ily and morphological interpretation of fossil calycanthaceous
Fig. 1 Calycanthaceae flowers at anthesis. A, Calycanthus floridus (end of female phase). B, Calycanthus occidentalis (end of female phase). C,
Sinocalycanthus chinensis (full female phase). D, Chimonanthus nitens (end of female phase–beginning of male phase). E, Chimonanthus praecox(full female phase). F, Idiospermum australiense (full female phase). Bar ¼ 5 mm.
286 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

flowers based on the results on extant flowers are discussed.Figure 1 introduces the floral diversity of the family, figures 2–8relate to the single species studied, and figures 9–14 providecomparisons across the family.
Material and Methods
The following collections were used for this study (S ¼Yannick M. Staedler; BGZ ¼ cultivated in the Botanic Gar-den of the University of Zurich; organ number ¼ collectionused for calculation of average floral organ numbers).
Calycanthus floridus L. (one individual, BGZ): S004-71,S004-80, and S005-47 (organ number), S004-34 (figs. 2, 9E),S004-4 (fig. 10A, 10D, 10G), S004-62 (fig. 13B, 13G, 13L),S004-80 (fig. 14E–14H), S004-125, S004-150 (immaturefruits), S005-94 (mature fruits with cup partially severed),S005-112 (intact mature fruit).
Calycanthus occidentalis Hooker and Arnott (one individ-ual, BGZ): S004-111, S004-147, and 005-79 (organ num-ber), S004-67 (figs. 3, 9F), S005-48, S005-33, and S005-33(fig. 10B, 10E, 10H), S004-157, S004-189 (immature fruits),S005-108 (mature fruit).
Fig. 2 Calycanthus floridus, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B, C, Level of
staminode-bearing part of floral cup. D–H, Level of gynoecium. I, Pedicel. S ¼ staminode, C ¼ carpel. Numbers indicate floral organ number on
the ontogenetic spiral. Bar ¼ 1 mm.
287STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

Fig. 3 Calycanthus occidentalis, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B, C, Level of
staminode-bearing part of floral cup. D–G, Level of gynoecium. H, Pedicel. S ¼ staminode, C ¼ carpel. Numbers indicate floral organ number on
the ontogenetic spiral. Bar ¼ 1 mm.
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

Chimonanthus nitens Oliver (two individuals numbered1 and 2, BGZ): J. B. Bachelier 004-136, 004-134, 004-198(individual 1; fig. 11A, 11D, 11G); P. K. Endress 03-111 (in-dividual 2; figs. 5, 9A); S004-231 (individual 1, organ num-ber), S004-232 (individual 2, organ number).
Chimonanthus praecox (L.) Link (one individual, BGZ): P.K. Endress 04-3 (fig. 13C, 13H, 13M; organ number), P. K.Endress 03-112 (figs. 6, 9B); S004-97, S004-131, and S004-163 (fig. 11B, 11E, 11H), S005-81 (immature fruits), S005-96, S006-7 (mature fruits).
Idiospermum australiense (Diels) S. T. Blake: B. P. M. Hy-land 2569 (Noah Creek, Northern Queensland, Australia;figs. 7, 9C, 15); P. H. Weston NSW 607200, 004-10, 004-77,004-79, 004-245, 005-84, 005-88, and 005-89 (Royal Bo-tanic Gardens, Sydney; organ number), 004-8, 004-11 and005-84 (Royal Botanic Gardens, Sydney; fig. 11C, 11F, 11I),
004-238 (fig. 13D, 13I, 13N); David H. Lorence 9471NTBG 840223 (National Tropical Botanical Garden, Kauai,Hawaii; immature fruits).
Sinocalycanthus chinensis (W. C. Cheng and S. Y. Chang)W. C. Cheng and S. Y. Chang (three individuals numbered1–3, BGZ): S004-112 (individual 1; organ number), S004-92(individual 2; organ number), S004-104 (individual 3; organnumber), S004-56 (individual 1; figs. 4, 9D), S004-121 (fig.8A, 8B; fig. 14A–14D), S005-20 (individual 1; fig. 10C, 10F,10I), S005-90 (individual 1; immature fruits), S005-97 (indi-vidual 1; mature fruits).
Plant material was fixed and stored in 70% ethanol. Serialmicrotome sections were made after embedding in Kulzer’sTechnovit 7100 (2-hydroxyethyl methacrylate) (Igersheimand Cichocki 1996). The sections were stained with toluidineblue and ruthenium red. For scanning electron microscopy
Fig. 4 Sinocalycanthus chinensis, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B–D, Level ofstaminode-bearing part of floral cup. E–H, Level of gynoecium. I, J, Pedicel. Bar ¼ 1 mm.
289STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

(SEM) studies, specimens were critical-point dried, sputter-coated with gold, and studied at 20 kV with a HitachiS-4000 scanning electron microscope.
For the study of phyllotaxis, the center of a flower needsto be determined. We did this in two ways. First, if a trans-verse section comprises many densely packed organs, whichis the case in the region of the rim of the floral cup of mostspecies, the center of the flower is estimated to be betweenthe three last initiated organs. Second, if a transverse sectionhas relatively few, loosely packed organs, which is the caselower in the floral cup, straight lines are drawn that more orless divide the floral cup in two halves, and the center of thefloral cup is estimated from the intersections of these lines. In
both cases, the relative position of organs is assessed usingthe center of the median vascular bundle of each organ in atransverse section close to, or just below, the base of the or-gan. These points of reference are then used to measure theangles between subsequent organs.
Divergence angles of the innermost organs conspicuouslyvary because of stochastic variation in the flower shape in itscenter and measurement errors in determination of the flowercenter. We use an artificial boundary between phyllotaxistypes based on statistical considerations. Systems are consid-ered to show a decussate phyllotaxis when the divergence an-gles alternate from 90� to 180� 6 10�. Organs are consideredto show Fibonacci spiral phyllotaxis if the average divergence
Fig. 5 Chimonanthus nitens, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B, Level of base of
staminodes. B–D, Level of gynoecium. E–J, Pedicel. Bar ¼ 1 mm.
290 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

angle equals the golden angle 6 1� and if at least one of twoconsecutive angles falls within the interval of average diver-gence angle 6 standard deviation. We will refer to organs be-longing to the zone between the decussate phyllotaxis zoneand the Fibonacci spiral phyllotaxis zone as organs of thephyllotaxis transition zone.
The transition between leaves and bracts is gradual; wethus treated as bracts all the modified leaves in the vicinity offloral organs. The transition between bracts and tepals is gradual,with no precise delimitation. Endress (1980a, 1980c) proposed(for spiral flowers of Austrobaileya and Hortonia) four crite-ria to—arbitrarily—set such a delimitation: ‘‘(1) beginning ofshort internodes, (2) change of phyllotaxis from decussate tospiral, (3) beginning of an increase in size of the phyllomes,(4) beginning of branched vasculature in the phyllomes’’(Endress 1980c, p. 396). We have used phyllotaxis change to
delimit bracts and tepals. The organs of the phyllotaxis tran-sition zone have a protective function often until late in floralbud development and are often partially colored (white, yel-low, or red). They play a role usually of sepals (in all generaexcept in Calycanthus) and occasionally of petals (in Calycan-thus floridus), and they have thus been treated as tepals. Wedid not use the other criteria proposed by Endress (1980a,1980c) because they could not be applied to the family as awhole.
Floral organ numbers were counted for 10 flowers per spe-cies, and means, modes (printed in bold when present), andstandard deviations were calculated. Divergence angles weremeasured for a late floral bud of all studied species, and themean angle and its standard error were calculated for eachspecies. Divergence angles were cursorily examined in addi-tional flowers of each species.
Fig. 6 Chimonanthus praecox, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B, Level ofstaminode-bearing part of floral cup. C, Level of base of innermost staminodes. C–E, Level of gynoecium. F–I, Pedicel. S ¼ staminode, C ¼ carpel.
Numbers indicate floral organ number on the ontogenetic spiral. Bar ¼ 1 mm.
291STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

Results
Calycanthus floridus. Flowers are solitary in the axilof a leaf of the previous year and are preceded by a pair (ortwo pairs) of foliage leaves. Just before anthesis, stamens, sta-minodes, and carpels are hidden by the tepals, and tepal tipsare appressed to each other. The flowers are then ovate in longi-tudinal section, and stigmas appear receptive. Tepals aredark reddish brown. Progressive flower opening during an-thesis (fig. 1A) is concomitant with style wilting and antherdehiscence.
In the sectioned floral bud studied for phyllotaxis (S004-43; fig. 2), the floral organs show Fibonacci spiral phyllo-taxis, with an average divergence angle of 137.44� (65.51�)for the 80 innermost organs (all organs except decussatebracts and transition organs). Divergence angles between theinnermost perianth organs show values all close to the goldenangle (fig. 12). Divergence angles between the remaining or-gans are also close to the golden angle (fig. 12). Two almostopposite organs are inserted on the pedicel of the floral bud(fig. 2I) (these organs alternate with the preceding pair of fo-liage leaves). The transition zone from decussate to spiralphyllotaxis comprises three organs (fig. 9E; fig. 10A, 10D).In later development, the ratio of floral organ to floral apexdiameter decreases (fig. 10G). Unexpectedly, it was observedthat along the ontogenetic spiral, the first carpel is followedby the two last staminodes and only then by the remainingcarpels (fig. 2D).
The 10 studied flowering shoots (S004-71, S004-80, andS005-47) comprise 2–4 (2:2 6 0:6) leaves, two opposite-decussate bractlike organs (seven out of 10 shoots), (26–)28(–29)(27:7 6 1) tepals, 13(–16) (13:8 6 1) stamens, (16–)20(–22)(19:2 6 1:3) staminodes, and (25–)34(–35) (31:7 6 3:6) car-
pels (the apparent modes of organ numbers are in boldface;all numbers are in plain text when no modes are apparent).Nicely (1965) reported larger ranges of organ numbers: 15–30 tepals, 10–15(–20) stamens, 15–25 staminodes, and 10–35 carpels (the numbers in Nicely 1965 for this and otherspecies were reported as groups of five, which seems artifi-cial). In the 10 flowers here studied, tepals do not behave asexpected from observation of the sections alone: at the endof the female phase of anthesis, a series of eight tepals (7:6 6 1)are spreading and later are reflexed, while the remainder at firststay upright and only later spread. The eight innermost tepalsusually have an incurved apex. The arrangement of tepals inthe sectioned floral bud (fig. 9E) and average tepal number of28 in the 10 buds studied may indicate that the perianth iscomposed of two 13-merous series (but see observation on an-thetic flowers). Stamens usually form a 13-merous series, withoften a less well-developed additional stamen behaving like astaminode (inflexed during the male phase of anthesis) (fig. 9E).Staminodes seem to form several 8-merous series at the rim ofthe cup (fig. 9E) and below (fig. 2B, 2C) (but see observationson fruits).
During fruit development (S004-125, S004-150), all sta-minodes enlarge, whereas all outer organs have abscised. Theinitially smaller innermost staminodes show the strongestgrowth in breadth and length and eventually become evenlarger than the outer staminodes. Growth of the staminodesafter anthesis results in the occlusion of the floral cup andcomplete inclusion of the carpels after wilting of the stigmasand styles. Staminodes form a ca. 21-merous series aroundthe rim of the floral cup. In mature fruits (S005-94, S005-112), they become reflexed upon fruit desiccation, but thisprocess does not open the floral cup enough to enable nutletrelease. A mature fruit (S005-94) was found with the side of
Fig. 8 Sinocalycanthus chinensis, laterally hinged valves of stamens. A, Late female phase of flowering, closed anther showing stomiumfollowing pollen sac outline bifurcating at the end of the pollen sac (white arrowhead). B, Full male phase of flowering, open anther showing
opened laterally hinged valves. Bar ¼ 300 mm.
Fig. 7 Idiospermum australiense, transverse section series of late floral bud. Vasculature in gray. A, Level of rim of floral cup. B, Level of
staminode-bearing part of floral cup. C–E, Level of inner staminodes, and carpel. F, Level of base of carpel. Bar ¼ 1 mm.
293STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

the floral cup mechanically severed, perhaps by a bird, butapparently with no nutlets missing. Van der Pijl (1982) men-tions dispersal of mature nutlets by squirrels.
Calycanthus occidentalis. Inflorescences are terminal withup to 15 flowers in a thyrsoid. After a variously long vegetativegrowth, the shoot apical meristem initiates a terminal flowerpreceded by a pair of at least partially reduced leaves, which
may subtend two- to seven-flowered cymes. Up to four flowersin the inflorescence usually mature; the others usually abort.The flowers are similar in shape to those of C. floridus butlarger, and tepals are deep red (fig. 1B). Flowering proceeds asin C. floridus.
In the sectioned floral bud studied for phyllotaxis (S004-67; fig. 3), the floral organs show Fibonacci spiral phyllo-taxis, with an average divergence angle of 137.48� (65.61�)
Fig. 9 Transverse sections at level of rim of floral cup, showing organ series. A, Chimonanthus nitens, with ontogenetic spiral and divergence
angles shown for bracts and tepals. A9, Close-up of flower center, with ontogenetic spiral and divergence angles shown for stamens, staminodes,
and carpels. B, Chimonanthus praecox. C, Idiospermum australiense. D, Sinocalycanthus chinensis. E, Calycanthus floridus. F, Calycanthusoccidentalis. Bar ¼ 1 mm.
294 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

for the 142 innermost organs (all organs except decussatebracts and transition organs). The divergence angles of theoutermost floral organs and the innermost floral organs showthe strongest deviation from the golden angle (fig. 12). Thetransition zone from decussate to spiral phyllotaxis comprisesnine organs (figs. 3F–3H, 9F; fig. 10B, 10E). In later develop-ment, the ratio of floral organ to floral apex diameter de-creases (fig. 10H). Unexpectedly, along the ontogenetic spiral,the first carpel is followed by the last staminode and onlythen by the remaining carpels (fig. 3D).
The 10 flowers studied for organ numbers (S004-111,S004-147, and S005-79) were the terminal flowers of 10 in-
florescences. They comprise 0–2–4 (1:8 6 0:9) opposite-decussate bractlike organs, (42–)45–47(–50) (46:1 6 2:5) tepals,22–27 (25:1 6 1:6) stamens, 36–50 (41:6 6 4:2) staminodes,and 35–43 (39:3 6 3:7) carpels. Nicely (1965) reported muchsmaller organ numbers with larger ranges: 15–30 tepals, 10–15 stamens, 15–25 staminodes, and 10–20 carpels. In the 10flowers here studied, no differentiation of the perianth into ob-vious series was observed during anthesis. In the sectionedfloral bud, the perianth seems to be arranged in two or three13-merous series (fig. 9F) (but see observations on antheticflowers). Stamens form two series in all flowers studied (alsoin the sectioned floral bud; fig. 9F). The two series show a
Fig. 10 Transition from decussate to spiral organ initiation. A, D, G, Calycanthus floridus. B, E, H, Calycanthus occidentalis. C, F, I,Sinocalycanthus chinensis. A–C, First organs of the transition zone. D–F, Somewhat older, initiation of further organs of the transition zone. G–I,Somewhat older, spiral initiation of organs. Bar ¼ 100 mm.
295STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

different behavior during the male phase of flowering: theouter, 21-merous series reflexes during the male phase of an-thesis, while the inner, 3–5-merous series inflexes (as do sta-minodes). In the 10 flowers studied and in the sectioned floralbud, staminodes also appear to form two 21-merous seriesfrom their average number of 42 (fig. 3B, 3C) (but see obser-vations on fruits).
During fruit development, all staminodes enlarge, whereasall outer organs have abscised. The initially smaller inner-
most staminodes show the strongest growth in breadth andlength and become larger than the outer staminodes (S004-157 and S004-189). Growth of the staminodes after anthesisresults in the occlusion of the floral cup and complete inclu-sion of the carpels after wilting of the stigmas and styles. Infruit, staminodes form two ca. 21-merous series on the rimof the floral cup. The staminodes desiccate during fruit matu-ration (observation on mature fruit S005-108); this processopens the cup wide, enabling nutlet release.
Fig. 11 Transition from decussate to spiral organ initiation. A, D, G, Chimonanthus nitens. B, E, H, Chimonanthus praecox. C, F, I,Idiospermum australiense. A–C, First organs of the transition zone, with scars of preceding decussate organs visible. D–F, Somewhat older,initiation of further organs of the transition zone. G–I, Somewhat older, spiral initiation of organs. Bar ¼ 100 mm.
296 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions
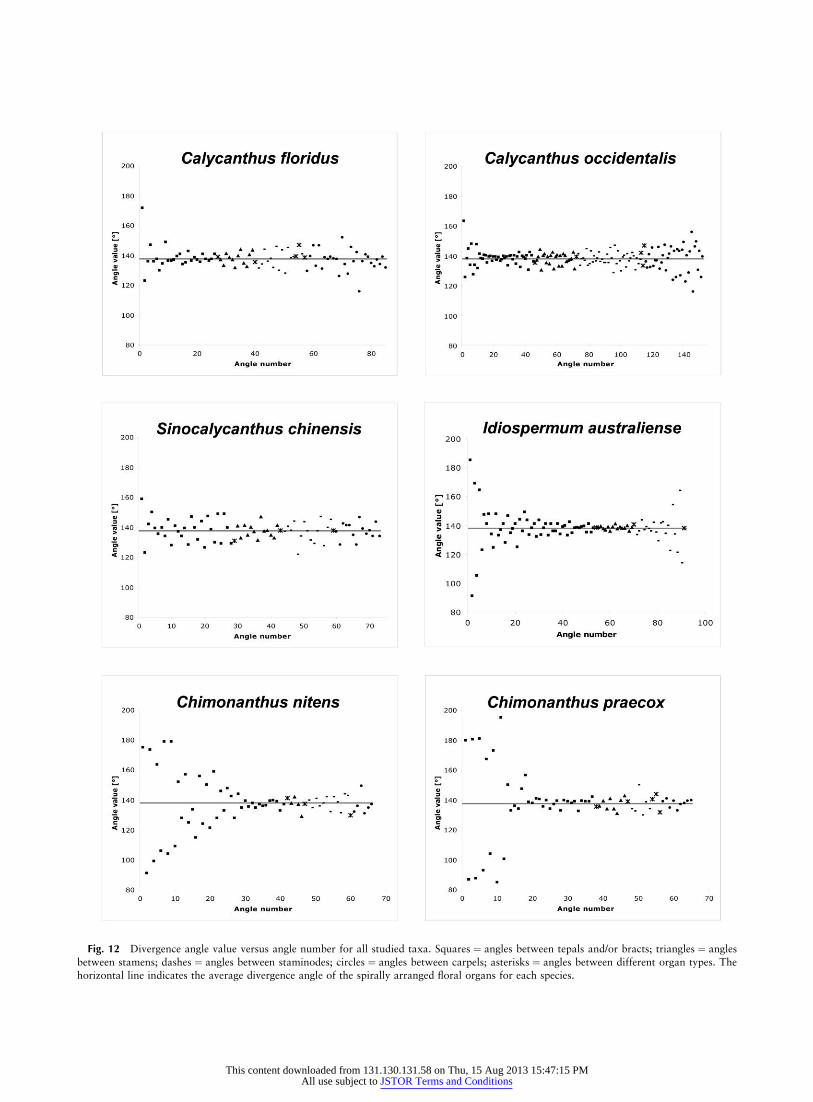
Fig. 12 Divergence angle value versus angle number for all studied taxa. Squares ¼ angles between tepals and/or bracts; triangles ¼ angles
between stamens; dashes ¼ angles between staminodes; circles ¼ angles between carpels; asterisks ¼ angles between different organ types. Thehorizontal line indicates the average divergence angle of the spirally arranged floral organs for each species.
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions
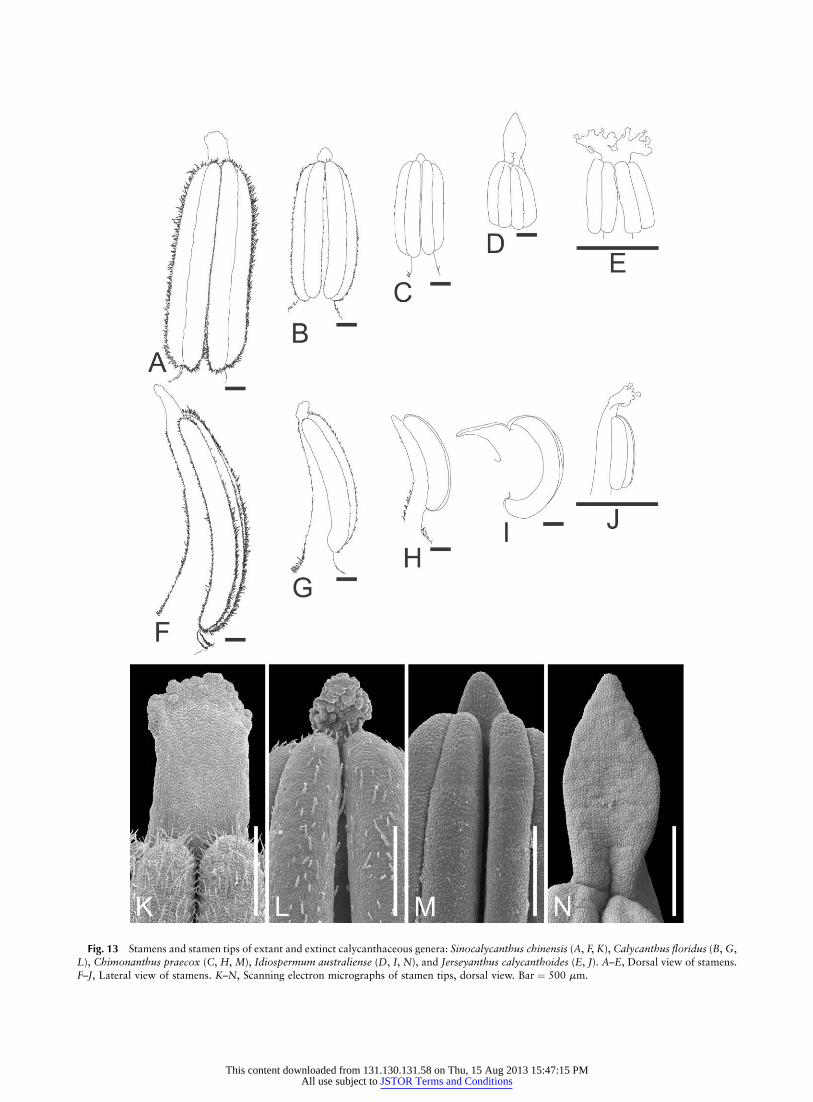
Fig. 13 Stamens and stamen tips of extant and extinct calycanthaceous genera: Sinocalycanthus chinensis (A, F, K), Calycanthus floridus (B, G,
L), Chimonanthus praecox (C, H, M), Idiospermum australiense (D, I, N), and Jerseyanthus calycanthoides (E, J). A–E, Dorsal view of stamens.
F–J, Lateral view of stamens. K–N, Scanning electron micrographs of stamen tips, dorsal view. Bar ¼ 500 mm.
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

Sinocalycanthus chinensis. Flowers are solitary and ter-minate long shoots. After vegetative growth, resulting in twoto four leaf pairs, the shoot apex turns into the flower apex.At the beginning of anthesis, the white outer tepals are al-most spreading and the yellow inner tepals form a roundedcuplike structure (fig. 1C). The reproductive organs are moreor less exposed during anthesis.
In the sectioned floral bud studied for phyllotaxis (S004-56; fig. 4), the floral organs show Fibonacci spiral phyllo-taxis, with an average divergence angle of 137.40� (66.06�)for the 68 innermost organs (all organs except decussate bractsand transition organs). Most divergence angles are close tothe golden angle (fig. 12). The transition zone from decussateto spiral phyllotaxis comprises five organs (fig. 4I, 4J; fig.
9D; fig. 10C, 10F). In later development, the ratio of floralorgan to floral apex diameter decreases (fig. 10I).
The 10 flowers studied for organ number (three to four ofeach of the three individuals S004-92, S004-104, and S004-112) are preceded by two bractlike organs (eight out of 10flowers), or bractlike organs are lacking (two flowers); flow-ers comprise (22–)26(–28) (25:5 6 1:9) tepals, 15(–19) (16:3 6
1:6) stamens, (14–)15(–17) (15:4 6 0:9) staminodes, and(12–)15(–16) (14:4 6 1:5Þ carpels. Cheng and Chang (1964)reported different ranges of organ numbers: 26–33 tepals,18–19 stamens, 11–12 staminodes, and 11–12 carpels. In the10 flowers here studied, the perianth is constituted of threeca. 8-merous series. The outermost series comprises mostlycaducous sepaloid tepals (with one or two white petaloid
Fig. 14 Comparison of inner tepals in Sinocalycanthus chinensis (A–D) and Calycanthus floridus (E–H). A, B, Photographs of the abaxial (A)
and adaxial (B) side of an inner tepal of S. chinensis. C, D, Scanning electron micrographs of the dorsal surface of an inner tepal (feeding tissue). E,F, Scanning electron micrographs of the dorsal (E) and lateral (F) surface of an innermost tepal of C. floridus. G, H, Detail of the tepal tip (feeding
tissue). Bar ¼ 2 mm (A, B, E, F), 300 mm (C, G), or 50 mm (D, H).
299STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

members). The middle series is composed of eight spreading,broad, thin, and smooth white tepals (fig. 1C). The inner se-ries is composed of eight upright, small, stout tepals (fig. 1C;fig. 14A, 14B) plus one to two smaller innermost tepals ofsimilar shape. All these inner tepals are covered with a yel-low papillate epidermis in their distal and lateral parts (fig.14A–14D); the remaining surface is smooth. The smoothpart of the inner tepals is white except for two small redpatches at the base. The inner eight tepals are coherent by se-cretion during the female phase of anthesis (fig. 1C). The ex-tra one or two tepals could thus play a role in stiffening theinner tepal series by bridging neighboring tepals. During themale phase of anthesis, coherence of the inner tepals is lostas the tepals become slightly reflexed. In the sectioned floralbud, the perianth seems to form two ca. 8-merous series (fig.9D) (but see observations on anthetic flowers). Stamens con-sistently form an outer series of 13 large, well-developed sta-mens, plus 2–3–4 smaller stamens that form a 5-merous seriestogether with 3–2–1 staminodes (also in the sectioned floralbud; fig. 9D). The thecae do not open by single longitudinalslits but open by valves, which originate by bifurcations ofthe dehiscence line at both of its ends (upper end shown infig. 8A, 8B). In the sectioned floral bud, the staminodes notassociated with stamens form a ca. 13-merous series (fig. 4C,4D) (but see observations on fruits).
During fruit development, all staminodes enlarge, whereasall outer organs have abscised (S005-90). The staminodes of theouter series do not usually show conspicuous growth, but thestaminodes of the inner series, on the contrary, conspicuouslyenlarge, ultimately leading to the occlusion of the floral cupand complete inclusion of the carpels after wilting of the stig-mas and styles. The enlarged staminodes become reflexed uponfruit desiccation (S005-97). This process is associated with awidening of the orifice of the cup, which enables nutlet release.
Chimonanthus nitens. Flowers are solitary in the axis ofevergreen foliage leaves. The stamens and stigmas are exposedduring anthesis. The flowers are narrowly funnel shaped atearly anthesis. The whitish petaloid tepals progressively spreadto an almost horizontal position during anthesis (fig. 1D).Three outer tepals reflex during anthesis.
In the sectioned floral bud studied for phyllotaxis (Endress03-111; fig. 5), the floral organs show Fibonacci spiral phyl-lotaxis, with an average divergence angle of 137.83� (64.38�)for the 37 innermost organs (all organs except decussatebracts and tepals of the transition zone). Divergence anglesbetween the last floral organs initiated show the strongest de-viation from the golden angle (fig. 12). The sectioned floralbud comprises 10 decussate organs; the transition zone fromdecussate to spiral phyllotaxis comprises 19 organs (figs. 5B–5E, 9A; fig. 11A, 11D). In later development, the ratio of flo-ral organ to floral apex diameter does not conspicuouslydecrease (fig. 11G).
The 10 flowers studied for organ number (five in each ofthe two specimens, S004-231 and S004-232) comprise 8–10(9:5 6 1:2) decussate bracts, 2–4–5 (2:8 6 1:4) sepaloid tepals,2–3–5 (3 6 0:9) recurved petaloid tepals, 13–19 (15:9 6 1:9)not recurved petaloid tepals, 5–6 (5:2 6 0:4) stamens, 10–11–12 (11:3 6 0:3) staminodes, and 7–9 (7:6 6 0:7) carpels. Nicely(1965) reported similar organ numbers with larger ranges:10–20 tepals, 5 stamens, 5–12 staminodes, and 5–15 carpels.In the 10 flowers here studied, the exposed part of bracts andsepaloid tepals is brownish and hard; these organs are proba-bly not involved in pollinator attraction. The petaloid tepalsform several series with varying patterns. The outermost, re-curved petaloid tepals usually form a 3-merous series. Themiddle petaloid tepals are the largest and the longest; theyform either two 5-merous series or one 8-merous series (as inthe sectioned floral bud; fig. 9A). The innermost tepals areshorter and slightly clawed; they form a 5-merous series inall flowers studied (also in the sectioned floral bud; fig. 9A).The stamens form a 5-merous series in most flowers studied(also in the sectioned floral bud; fig. 9A); however, an addi-tional smaller stamen is present in three of the 10 flowersstudied. In the sectioned floral bud, the staminodes seem toform an outer 8-merous and an inner 5-merous series (figs.5B, 9A). The role of the staminodes in fruit construction andfruitlet dispersal has not been studied because fruiting mate-rial was not available.
Chimonanthus praecox. Flowers are solitary in the axilof fallen foliage leaves of the preceding season. The stamens
Fig. 15 Idiospermum australiense, transverse section of late floral bud, rim of the floral cup (from fig. 7A). Only the spirally arranged organsare shown, with different sets of parastichies. A, Set of five parastichies. B, Set of eight parastichies. C, Set of 13 parastichies. Bar ¼ 1 mm.
300 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

and stigmas are exposed during anthesis. The flowers arebroadly funnel shaped at early anthesis. They comprise yellowand brownish red petaloid tepals (fig. 1E); the yellow onesspread to a more or less horizontal position during anthesis.
In the sectioned floral bud studied for phyllotaxis (Endress03-112; fig. 6), the floral organs show Fibonacci spiral phyl-lotaxis, with an average divergence angle of 137.78� (63.79�)for the 46 innermost organs (all organs except decussatebracts and transition organs). Most organs show divergenceangles close to the golden angle (fig. 12). The sectioned floralbud comprises six decussate organs, and the transition zonefrom decussate to spiral phyllotaxis comprises 13 organs (alltepals in fig. 6E; the 12 outer tepals in fig. 6C, 6D; the 10outer tepals in fig. 6B; the nine outer tepals in fig. 9B; seealso fig. 11B, 11E). In later development, the ratio of floralorgan to floral apex diameter does not conspicuously de-crease (fig. 11H). Unexpectedly, along the ontogenetic spiral,the first carpel is followed by the last staminode and onlythen by the remaining carpels (fig. 6C).
The 10 flowers studied for organ number (Endress 04–3)comprise 12–14–16 (14:4 6 1:3) decussate bracts, 5–6–7(6:4 6 0:9) sepaloid tepals, 15–16–18 (16:1 6 0:9) petaloidtepals, 6–8 (7:3 6 0:9) stamens, 8–12 (10:4 6 1:4) stamino-des, and 8–9–10 (9:3 6 0:7) carpels. Nicely (1965) describedsimilar organ numbers with larger ranges: 15–20 tepals, 5–10 stamens, 10–15 staminodes, and 5–15 carpels. In the 10flowering shoots here studied, the 21 first organs (decussatebracts and phyllotaxis transition tepals) are short, broad-based organs with a tip that is hard and brown (the last or-gans before the petaloid tepals are yellow and petaloid attheir base); these organs do not seem to be involved in polli-nator attraction. The petaloid tepals of the 10 studied flowersform on the average two 8-merous series (also present in thesectioned floral bud; fig. 9B). The first, outer series comprisesall the elongate, yellow tepals. The second, inner series com-prises all the short, clawed, partially or entirely brownish-redtepals (see also Nelson 1954). The brownish-red tepals se-crete nectar by small glands (nectarioles; Vogel 1998). Thetwo innermost perianth series, together with the stamens,represent the optical attractant (fig. 1E) and reward systemof the flower. The stamens often form an 8-merous series (sixof the 10 flowers studied) or a series of six or seven stamensand two or one staminodes (in the sectioned floral bud, sevenstamens are present; fig. 9B). The remaining staminodes forma ca. 8-merous series (also in the sectioned floral bud; figs.6C, 9B).
During fruit development (S005-81), all staminodes en-large, whereas all outer organs have abscised. Growth of thestaminodes after anthesis results in the occlusion of the floralcup and complete inclusion of the carpels after wilting of thestigmas and styles. Staminodes show no movement duringfruit desiccation. Fruitlets may be released through decay ofthe tissue of the staminodes (S005-96). Mature fruits werefound with the cup mechanically severed (vertically slashed)and the nutlets missing (S006-7), probably taken by birds.
Idiospermum australiense. The flowers are terminal oraxillary, solitary, in botryoids of three to 11 flowers, or inthyrsoids (Y. Staedler, personal observation on herbariumand living material). At the beginning of anthesis, the anthersand stigma(s) are hidden (Worboys and Jackes 2005); the
flowers are obovate in longitudinal section. During anthesis,the flowers progressively open. At the end of anthesis, mosttepals are spreading or are recurved (fig. 1F). Tepals arecreamy white at first and become progressively flushed withpink, then red, and finally dull reddish purple.
In the sectioned floral bud studied for phyllotaxis (Hyland2569; fig. 7), the floral organs show Fibonacci spiral phyllo-taxis, with an average divergence angle of 137.67� (66.38�)for the 78 innermost organs (all organs except decussatebracts and transition organs). The last initiated organs showthe largest deviation from the golden angle. Divergence an-gles close to the golden angle are observed for the innermostperianth series and for the stamen series (fig. 12). The sec-tioned floral bud comprises four decussate organs and nineorgans in the transition zone from decussate to spiral phyllo-taxis (fig. 9C; fig. 11C, 11F). In later development, the ratioof floral organ to floral apex diameter decreases (fig. 11I).
The 10 studied flower buds (Weston NSW 607200, 004-10, 004-77, 004-79, 004-245, 005-84, 005-88, and 005-89)comprised 6–10–12 (8:6 6 2:3) bractlike organs, 32–35–38–48 (38:2 6 4:4) tepals, 13–14–15–16 (14:2 6 1:4) stamens,13–16–18–21 (16:2 6 3:1) staminodes, and 0–1–2 (1:2 6 0:8)carpels. Blake (1972) reported 30–40 tepals, 13–15 stamens,8–10 staminodes, and 0–2 carpels. Worboys (2003) reported32–48 tepals, 10–17 stamens, 8–23 staminodes, and 1–5 car-pels in newly found southern populations. Wilson (1976) re-ported 48–50 tepals, 12–13 stamens, 8–10 staminodes, and0–2 carpels. In the 10 late floral buds and flowers here stud-ied, the caducous decussate bracts serve to protect the youngfloral bud. The following 20 tepals are either caducous budprotection organs or noncaducous spatulate, reflexed, opti-cal, and olfactory display organs (Worboys and Jackes 2005)or are noncaducous intermediate organs between these twotypes. These tepals may be arranged in three series of eightor five organs (fig. 9C). The innermost tepals form consis-tently two series of 13 tepals (also in the sectioned floralbud; fig. 9C). The outer series of 13 tepals comprises mostlyspreading, large, spatulate, clawed, olfactory and opticallyattractive organs and 2–3 smaller, thicker, acuminate tepals(fig. 1F). The innermost 13 tepals are vertical, short, stout,with parallel and straight margins and an incurved broadlyacuminate apex (fig. 1F). These tepals seem to be involvedin the construction of the sheltered environment of thefloral center characteristic for the observed beetle pollination(Worboys and Jackes 2005). A series of 13 stamens is almostalways present on the rim of the cup (fig. 9C), and an addi-tional series of three seemingly functional, smaller stamens issometimes present inside the cup, topographically just belowthe first stamen series (also present in the sectioned floralbud; fig. 9C). The frequent staminode numbers (13, 16, 18,21) may indicate one or two series (21 staminodes werefound in the sectioned floral bud). Staminodes close the cupat the end of the female phase of anthesis (Worboys andJackes 2005). This could play a role in protection against pre-dation by the numerous larvae found in the flowers (Worboysand Jackes 2005). Staminodes persist until quite late in fruitdevelopment, as observed on immature fruits (Lorence 9471NTBG 840223) (see also Endress 1983). But they play norole in diaspore release; only one fruitlet develops, and theentire fruit is the diaspore. Most flowers of the collection by
301STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

P. H. Weston from a cultivated tree (Weston NSW 607200,004-10, 004-77, 004-79, 004-245, 005-84, 005-88, and 005-89) comprise one carpel (five out of 12) or two carpels (fiveout of 12); few had no carpel (two out of 12). In the collec-tion from a wild tree by B. P. M. Hyland (Hyland 2569),flowers with two carpels were rare (less than 10% of thestudied flowers), and around half of the flowers had only onecarpel; the rest had no carpel. Because we favored large latebuds, this could be a collecting artifact (if flowers with twocarpels are indeed larger than flowers with only one or nocarpel), or it might reflect the less competitive environmentof the cultivated plant than the wild tree from which thespecimens were collected.
Discussion
Floral Architecture
Flowers of Calycanthaceae open at anthesis (Chimonan-thus; fig. 1D, 1E), are closed during the female phase andlater open during the male phase (Calycanthus, Idiospermum;fig. 1A, 1B, 1F), or contain a sheltered but not completelyclosed chamber that opens at the transition from female tomale phase, as the tips of the tepals move apart from eachother (Sinocalycanthus; fig. 1C). Flowers always have a floralcup, on the upper rim of which stamens are borne. Staminodesare always positioned at the entrance of the floral cup. Theovary is always hidden in the cup (figs. 2–7), with stigmasexposed during the female phase of anthesis (Chimonanthus,Sinocalycanthus) or hidden (Calycanthus, Idiospermum). Thepresence of a floral cup is also pronounced in other Laurales(Endress 1980b; Endress and Igersheim 1997).
An outstanding feature found in this study is that the thecaein Sinocalycanthus open via laterally hinged valves, not simplelongitudinal slits as in the other Calycanthaceae studied (fig.8A, 8B). Sinocalycanthus flowers have the largest stamens ofstudied Calycanthaceae (fig. 13A–13J), with an especiallylarge connective, a feature found to be correlated with valvatedehiscence (Hufford and Endress 1989). This is of special in-terest because this opening mode is present in a number ofMagnoliales (Endress and Hufford 1989) but occurs rarely inLaurales (Endress and Hufford 1989). Because it is also com-mon in fossil Cretaceous stamens (Friis et al. 1991) and pres-ent in the Cretaceous genus Jerseyanthus (fig. 13E, 13J) (seebelow), our new results may give additional weight to an in-terpretation of such anthers as plesiomorphic in Laurales.
Floral Phyllotaxis and Transition between Floral Organs
All Calycanthaceae flowers here studied show spiral phyl-lotaxis according to the Fibonacci pattern. Optimizationstudies (Doyle and Endress 2000; Ronse De Craene et al.2003; Zanis et al. 2003; Endress and Doyle 2007) indicatethat the ancestral character state may be whorled for theperianth in the magnoliid clade (a clade comprising two sub-clades of two orders each, the Laurales-Magnoliales subcladeand the Piperales-Canellales subclade; Qiu et al. 1999, 2005;Zanis et al. 2002). Both spiral and whorled floral phyllotaxisare also present in the Magnoliales as well as in more basalangiosperms (Doyle and Endress 2000; Endress and Iger-
sheim 2000; Endress 2001). This suggests two main alterna-tive hypotheses. (1) The ancestral floral phyllotaxis inmagnoliids is whorled, and a transition to spiral phyllotaxishas occurred early in the evolution of Laurales, probably atleast at the early-middle Albian (to which Virginianthus caly-canthoides belongs, the earliest known fossil with Calycan-thaceae-like flowers and probably with spiral phyllotaxis;Friis et al. 1994). Whorled floral phyllotaxis in Laurales isknown since the early Cenomanian from the Lauraceae-likeflowers of Mauldinia (Drinnan et al. 1990). (2) The ancestralfloral phyllotaxis in magnoliids is spiral, and whorled floralphyllotaxis among magnoliids evolved independently severaltimes. To test these hypotheses more rigorously, future stud-ies of floral phyllotaxis in other families of Laurales andother magnoliids are needed.
The transition zone from vegetative to floral phyllotaxis ex-tends over 11–19 organs in Chimonanthus, nine organs inIdiospermum and Calycanthus occidentalis, and three to fiveorgans in Calycanthus floridus and Sinocalycanthus (fig. 12).Flowers of Chimonanthus need 5–7 mo from initiation to an-thesis, and the floral apex in the early spiral phase is ca. 100–150 mm in diameter (fig. 11D, 11E). In contrast, flowers ofCalycanthus and Sinocalycanthus need 3–4 mo from initia-tion to anthesis, and the floral apex in the early spiral phaseis ca. 200–300 mm in diameter (fig. 10D–10F). Thus, tran-sitions with many organs appear to be correlated withrelatively small and slow-growing floral apices, whereas tran-sitions with only few organs appear to be correlated with rel-atively large and fast-growing floral apices. A large organprimordium size relative to the floral apex may have an inhib-itory role in the transition process by allowing less placementfreedom for the new primordium. The growth rate of the flo-ral apex seems to play a crucial role in phyllotaxis transition.
The transition from one organ type to the next is unidirec-tional in flowers of Calycanthaceae. However, in the tran-sition from staminodes to carpels, irregularities in organidentity sequence were found in three taxa (C. floridus, C.occidentalis, and Chimonanthus praecox). The first carpelpreceded the last staminode or the two last staminodes onthe ontogenetic spiral (figs. 2D, 3D, 6C). Despite the irregu-larities in sequence of organ identity, the positions of the or-gans are very close to ideal Fibonacci positions. During theinitiation of the last staminodes and the first carpels, the nowconcave floral apex has reached its maximal size, whichcould explain the observed irregularities. Apparently, undersuch extreme conditions organ identity depends on the local,sectorial neighborhood rather than on the position on theontogenetic spiral if subsequent organs are small and notcontiguous. Shorter plastochrons than in the perianth couldcontribute to emphasizing such a phenomenon. Such extremecases with minute organ primordia and large floral or shootapex would favor a view as proposed by Plantefol (1949),who focused on parastichies rather than the ontogenetic spi-ral as determinants for organ initiation. Similar irregularitiesin sequence were also found and discussed in heads of Aster-aceae, here with regard to bracts, ray flowers, and disk flow-ers (Hirmer 1931; Bachmann 1983; Battjes and Bachmann1996; Battjes and Prusinkiewicz 1998). Because these systemsare entire inflorescences, they are larger than single flowersand therefore more easily accessible for experimental studies.
302 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

The easy switch from staminodes to carpels and vice versamay suggest that staminodes of Calycanthaceae are in theiridentity closer to carpels than to stamens, thus strengtheningthe view of Hiepko (1965), who based his hypothesis that thestaminodes of Calycanthaceae should rather be interpretedas carpellodes on observations he made on young stages: thedistinct transition between stamens and staminodes but theimpossibility of distinguishing between staminodes and car-pels. However, the presence of organs with only one very re-duced pollen sac, especially in taxa with numerous stamens(Calycanthus, Sinocalycanthus, and Idiospermum), and simi-lar behavior of fertile stamens and staminodes during anthesis,do not support a carpel identity, at least of the outermoststaminodes. It seems reasonable to suggest that the staminodesof Calycanthaceae have a distinct morphology and that theymay be intermediate organs between stamens and carpels (asa side effect of spiral organization) that have acquired a newfunction in the closure of the floral cup. Gene expression stud-ies would be needed to resolve the identity of these organs de-cisively.
Organ Series
The perianth organs of Calycanthaceae, although they arealways arranged following a continuous spiral, show differ-entiation into functional categories (bud protection vs. at-traction, reward, or construction of a sheltered environmentfor pollinators). The differentiation of the perianth almost al-ways includes the presence of intermediate organs, in accor-dance with the fading-borders model of floral developmentalgenetics (Buzgo et al. 2004, 2005). Organs can neverthelessbe grouped into subcategories. The organ number of a sub-category is often a Fibonacci number (see Nelson 1954 forthe perianth of Chimonanthus praecox). The innermost peri-anth organs seem to be always involved in pollinator attrac-tion, sometimes including reward (C. praecox; probably alsoSinocalycanthus; see Vogel 1998 for both taxa) and some-times in the construction of a shelter for pollinators. The lat-ter is the case in the beetle flowers of Calycanthus (Grant1950; Nicely 1965; Badrutt 1992) and Idiospermum (Worboysand Jackes 2005) and could also be the case in Sinocalycan-thus (Y. M. Staedler, personal observation of small Coleop-tera and Heteroptera between the reflexed outer stamens andthe inner tepals in female phase flowers at night).
Stamens, more often than other organs, appear in series:series of five or eight in Chimonanthus, series of 13 in C. flor-idus, Sinocalycanthus, and Idiospermum, and series of 21 inC. occidentalis (fig. 9). Few additional, often reduced innerstamens are present in Calycanthus, Sinocalycanthus, andIdiospermum. They form a series with staminodes (fig. 9D).The regularity of stamen number is probably due to the de-velopmental constraints of their position on the rim of thefloral cup. The outer and inner stamen series sometimes donot behave similarly during anthesis. The inner series tendsto behave like staminodes: in Calycanthus, they inflex at theend of anthesis, thus contributing to floral cup closure (whilethe outer stamens reflex).
Staminodes often appear in series of a higher Fibonaccinumber than series of stamens (usually 13 or 21); this isprobably a result of their late initiation when the floral apex
is at maximal size and of their relatively narrow shape. Thestaminodes always play a role in the closure of the floral cupduring fruit development and often during anthesis (Worboysand Jackes 2005; this study). At fruit maturity, they reopenthe cup and allow nutlet release in at least two taxa (C. occi-dentalis and Sinocalycanthus), and they may play this rolealso in C. floridus. Staminode decay may also play a role innutlet release in C. praecox. In Idiospermum, the staminodesand even the stamens seem to persist until late fruit matura-tion. At maturity, the staminodes disintegrate together withthe floral cup.
Floral Structure and Intrafamilial Relationship
Two recent phylogenetic studies of Calycanthaceae, usingnrITS (Li et al. 2004) and combining ITS, trnL–F, and trnC–D data (Zhou et al. 2006), have confirmed affinity of most ofthe genera of the Calycanthaceae (Cheng and Chang 1963,1964; Blake 1972): Sinocalycanthus sister to or nested in Caly-canthus, Chimonanthus sister to Calycanthus plus Sinocaly-canthus, and Idiospermum sister to the remainder of thefamily. Segregation of Calycanthus chinensis into a separategenus, Sinocalycanthus (Cheng and Chang 1964), was basedon differences in the perianth (breadth, color, and surfacedifferentiation of tepals; fig. 1A–1C) and lack of smell ofthe flowers (although this does not correspond to our ob-servations). Other differences between Sinocalycanthus andCalycanthus include valvate dehiscence of the anthers of Sin-ocalycanthus (this study) versus dehiscence by slits in Caly-canthus (Endress and Hufford 1989) and all other extantCalycanthaceae (this study) and postgenital coherence of in-ner tepal series. Instead of discrete conical food bodies (fig.14E–14H), ‘‘feeding tissue’’ (papillate, cytoplasm-rich tissuesimilar to that of the food bodies on inner tepal and stamentips in Calycanthus) is present on the yellow inner tepal pe-riphery and partially on stamen and staminode tips (Vogel1998; this study: fig. 14A–14D). All of these characters canbe interpreted as autapomorphies of Sinocalycanthus, butnone of them can be interpreted unequivocally as autapomor-phies of Calycanthus.
In contrast, successful hybridization between Sinocalycan-thus chinensis and C. floridus (Lasseigne et al. 2001) suggestsa placement of Sinocalycanthus in Calycanthus. The feedingtissue of Sinocalycanthus (tip of the inner tepals, stamens,and staminodes; figs. 13K, 14A–14D) is present on the sameorgans at the same place as the food bodies of Calycanthus(figs. 13L, 14E–14H), and they seem to differ only in sizeand perhaps in amount of secretion produced (much moreabundant in Calycanthus; fig. 14D, 14H). Feeding tissue isnot present on the petal and anther tips in Chimonanthusand Idiospermum (fig. 13M, 13N). The postgenital coherenceof the inner tepals during the female phase of anthesis (pres-ent only in Sinocalycanthus) could be correlated with pro-duction of secretion by the feeding tissue during the femalephase and its cessation during the male phase (concomitantlywith continuous opening of the flower). Calycanthus flowerssmell of fermented fruits and are pollinated by small Coleop-tera (Grant 1950; Badrutt 1992). Although Cheng and Chang(1964) described the flower of Sinocalycanthus as scentless,we detected a faint smell of overripe pears that appeared to
303STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

be stronger at night. At night, flower visitors were observedin a cultivated plant, among which were a few small, yet un-determined Coleoptera. These observations are consistentwith the putative generalized cantharophily of these flowerspredicted by Vogel (1998) and thus with a close functionalsimilarity of the flowers of Calycanthus and Sinocalycan-thus. Another functional similarity is the wide fruit openingin C. occidentalis and S. chinensis at maturity. These struc-tural/functional similarities support the close relationship ofSinocalycanthus and Calycanthus found in molecular studies.
Interpretation of Fossil Calycanthaceous Flowers
The new results on floral structure in Calycanthaceae canbe used to improve the interpretation of the floral structureof fossil calycanthaceous flowers. Araripia florifera is basedon an impression fossil of a flowering twig with 6(–7) flowers(2 probably mature and 4–5 in earlier stages of development)of the early Cretaceous (Aptian or possibly Albian) of theCrato Formation, Brazil (Mohr and Eklund 2003). The larg-est flower or flower bud is ca. 10 mm long and 6 mm broad.It comprises at least 11–13 helically arranged straplike tepalswith appressed tips, and the tepals are inserted on a floralcup comprising at least six to seven globular structures.These flowers or floral buds are reminiscent of early femalephase C. floridus flowers. A side view of a flower of C. flori-dus (in early female phase or just before anthesis) displays anumber of tepals similar to the number seen in the fossil. Thetepals of the fossil seem spirally arranged, but the mode ofpreservation prevents any detailed observations.
Virginianthus calycanthoides is based on a single well-preserved charcoalified flower of the early Cretaceous (early-middle Albian) of Virginia. (Friis et al. 1994). The flower isca. 2.8 mm long and 2.2 mm broad. Phyllotaxis is probablyspiral, but preservation does not allow identification of se-ries, except for staminodes (an 8-merous series) and perhapsfor the perianth, which comprises at least 12 tepals in two se-ries (perhaps a 5-merous and an 8-merous series). The exactnumber of stamens is not known (30–40); the anthers are ex-tremely extrorse and dehisce via laterally hinged valves.
Jerseyanthus calycanthoides is based on extremely well-preserved charcoalified clearly calycanthaceous fossil flowersfrom the late Cretaceous (Turonian) of New Jersey. (Crepetet al. 2005). The flowers are 1.3–2.8 mm long and 1.5–3 mmbroad. The perianth parts are numerous and appear spirallyarranged, although no full set of parastichies could be drawnfrom the illustrations. The androecium was described as com-prising a whorl of 12 ‘‘staminode-stamen’’ pairs with eachstaminode abaxial to and overarching a latrorse to extrorsestamen (Crepet et al. 2005). However, 13 stamens could bepresent in CPC 1487 (figs. 1–3 in Crepet et al. 2005). In ex-tant Calycanthaceae, no trace of whorled arrangement wasfound in any part of the flowers, but series of 13 stamenswere consistently found in C. floridus, Sinocalycanthus, andIdiospermum. In Idiospermum, the rim of the floral cup iscrowded with organs (tepals and stamens); successive sets ofparastichies rapidly increase in steepness (fig. 10A–10C); asthe organs are dorsiventrally flattened, the parastichies of theset of 13 look almost like orthostichies (fig. 15C). Thus, eachouter stamen is almost—but not exactly—on the same radius
as one of the innermost tepals (the theoretical angle betweena tepal and its nearest stamen is 12.4�). If the stamens and‘‘outer staminodes’’ of Jerseyanthus were spirally arranged,the outer staminodes and the stamens would not be perfectlyaligned, which indeed appears to be the case (figs. 11, 13 inCrepet et al. 2005). In conclusion, Jerseyanthus flowers areprobably spiral, as supported by the following evidence: (1)absence of any whorled phyllotaxis in the studied extantmembers of the family, (2) the common occurrence of seriesof 13 stamens in extant Calycanthaceae, and (3) presence inJerseyanthus of a set of 13 steep parastichies almost (but notcompletely) like orthostichies as in extant Calycanthaceae(especially Idiospermum; fig. 15C). The perianth of Calycan-thaceae often shows conspicuous differentiations between itsouter and inner organs (Idiospermum, Chimonanthus, andSinocalycanthus). The inner tepals of Sinocalycanthus areshort and have a papillate epidermis on most of their surface(fig. 14A, 14B). What has been interpreted as outer stami-nodes in Jerseyanthus also shows these features. Given theirmorphological similarity to similarly placed organs of flowersof the extant Calycanthaceae, we suggest that the ‘‘outerstaminodes’’ of Jerseyanthus should rather be considered te-pals. The two thecae of the anther of Jerseyanthus open vialaterally hinged valves (‘‘staminal dehiscence ‘valvate’ dehis-cence slit I-shaped’’ [Crepet et al. 2005, p. 1476]) and have amodern counterpart in Sinocalycanthus, as found in thisstudy, whereas the thecae of the anthers in all other extantCalycanthaceae appear to open with single longitudinal slits.An additional unusual feature of the anthers in Jerseyanthusis a ramified connective appendage, which may representfood bodies (fig. 13E, 13J) (Crepet et al. 2005). All extantCalycanthaceae have connective appendages, but they arenot ramified; only in Sinocalycanthus may there be incipientramification (fig. 13A–13D, 13F–13I, 13K–13N).
Detrusandra mystagoga is based on charcoalified flowersof the late Cretaceous (Turonian) of New Jersey (Crepet andNixon 1998), with additional flowers found in central Geor-gia from the Santonian (Herendeen et al. 1999). The flowersare 1.0–2.5 mm long and 1.1–3.2 mm broad. Despite superfi-cial resemblance to Calycanthaceae, the fossil was placed bya cladistic analysis close to Eupomatiaceae (Crepet and Nixon1998). Character states consistent with exclusion from Caly-canthaceae are scalariform perforation plates of the vesselelements, laminar introrse stamens, absence of staminodes,bilobed, sessile stigmas, and at least five ovules per carpel(Crepet and Nixon 1998). Character states of D. mystagogathat are inconsistent with membership of the crown group ofEupomatiaceae are presence of a perianth, a well-developedfloral cup that totally encloses the carpels, and completelyapocarpous carpels. Nevertheless, this fossil could still be-long to the stem group of the Eupomatiaceae. Spiral floralphyllotaxis of Detrusandra does not help in the systematicplacement because floral phyllotaxis is also spiral in both Ca-lycanthaceae and Eupomatiaceae (Endress 2003).
Acknowledgments
We thank David H. Lorence, National Tropical BotanicalGarden, Lawai, Kauai, Hawaii, for some flowers and the
304 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

fruits of Idiospermum, and Bernie P. M. Hyland, Com-monwealth Scientific and Industrial Research Organisation,Atherton, Queensland, Australia, for some floral buds of Idi-ospermum. We also thank Julien Bachelier for collection of
some of the material of the Botanic Garden of the Universityof Zurich. Andre Chanderbali, Larry Hufford, and BruceKirchoff are acknowledged for critically reading the manu-script.
Literature Cited
Bachmann K 1983 Evolutionary genetics and the genetic control ofmorphogenesis in flowering plants. Evol Biol 16:157–208.
Badrutt S 1992 Struktur und Funktion der Bluten bei Calycantha-
ceae. Diploma thesis, University of Zurich.
Baillon H 1868 Histoire des plantes. Vol 1. Hachette, Paris.Battjes J, K Bachmann 1996 Numerical canalization in asteracean
heads. Pages 185–204 in PDS Caligari, DJN Hind, eds. Compositae:
biology and utilization. Vol 2. Royal Botanic Gardens, Kew.Battjes J, P Prusinkiewicz 1998 Modeling meristic characters of
asteracean flowerheads. Pages 281–312 in D Barabe, RV Jean, eds.
Symmetry in plants. World Scientific, Singapore.
Blake ST 1972 Idiospermum (Idiospermaceae), a new genus andfamily for Calycanthus australiensis. Contrib Queensl Herbar 12:
1–37.
Buzgo M, PS Soltis, S Kim, DE Soltis 2005 The making of the flower.
Biologist 52:149–154.Buzgo M, PS Soltis, DE Soltis 2004 Floral developmental morphol-
ogy of Amborella trichopoda (Amborellaceae). Int J Plant Sci 165:
925–947.
Cheng WS, SY Chang 1963 Calycanthus sect. Sinocalycantus. SciSilvae 8:1.
——— 1964 Genus novum Calycanthacearum chinae orientalis.Acta Phytotaxon Sin 9:137–139.
Crepet WL, KC Nixon 1998 Two new fossil flowers of magnoliid
affinity from the Late Cretaceous of New Jersey. Am J Bot 85:
1273–1288.
Crepet WL, KC Nixon, MA Gandolfo 2005 An extinct calycanthoidtaxon, Jerseyanthus calycanthoides, from the Late Cretaceous of
New Jersey. Am J Bot 92:1475–1485.
Cronquist A 1957 Outline of a new system of families and orders of
the dicotyledons. Bull Jard Bot Brux 27:13–40.——— 1981 An integrated system of classification of flowering
plants. Columbia University Press, New York.
——— 1988 The evolution and classification of flowering plants.2nd ed. New York Botanical Garden, New York.
Dahlgren R, K Bremer 1985 Major clades of the angiosperms.
Cladistics 1:349–368.
Delpino F 1870 Altri apparecchi dicogamici recentemente observati.Nuovo G Bot Ital 2:51–64.
Dengler NG 1972 Ontogeny of the vegetative and floral apex of
Calycanthus occidentalis. Can J Bot 50:1349–1356.
Diels L 1912 Die Auffindung von Calycanthus im australischenRegenwald. Bot Jahrb Syst 48:7–11.
Doust AN 2001 The developmental basis of floral variation in
Drimys winteri (Winteraceae). Int J Plant Sci 162:697–717.
Doyle JA, PK Endress 2000 Morphological phylogenetic analysis ofbasal angiosperms: comparison and combination with molecular
data. Int J Plant Sci 161(suppl):S121–S153.
Drinnan AN, PR Crane, EM Friis, KR Pedersen 1990 Lauraceousflowers from the mid-Cretaceous of eastern North America. Bot
Gaz 151:370–384.
Eichler AW 1878 Bluthendiagramme. Vol 2. Engelmann, Leipzig.
Endress PK 1980a Floral structure and relationships of Hortonia(Monimiaceae). Plant Syst Evol 133:199–221.
——— 1980b Ontogeny, function and evolution of extreme floral
construction in Monimiaceae. Plant Syst Evol 134:79–120.
——— 1980c The reproductive structure and systematic position ofthe Austrobaileyaceae. Bot Jahrb Syst 101:393–433.
——— 1983 Dispersal and distribution in some small archaic relict
angiosperm families (Austrobaileyaceae, Eupomatiaceae, Himan-
tandraceae, Idiospermoideae-Calycanthaceae). Sonderbd NatwissVer Hambg 7:201–217.
——— 1987 Floral phyllotaxis and floral evolution. Bot Jahrb Syst
108:417–438.——— 1990 Patterns of floral construction in ontogeny and phylog-
eny. Biol J Linn Soc 39:153–175.
——— 2001 The flowers in extant basal angiosperms and inferences
on ancestral flowers. Int J Plant Sci 162:1111–1140.——— 2003 Early floral development and the nature of the calyptra
in Eupomatiaceae. Int J Plant Sci 164:489–503.
Endress PK, JA Doyle 2007 Floral phyllotaxis in basal angiosperms:
development and evolution. Curr Opin Plant Biol (forthcoming).Endress PK, L Hufford 1989 The diversity of stamen structures and
dehiscence patterns among Magnoliidae. Bot J Linn Soc 100:45–85.
Endress PK, A Igersheim 1997 Gynoecium diversity and systematics
of the Laurales. Bot J Linn Soc 125:93–168.——— 2000 Reproductive structures of the basal angiosperm
Amborella trichopoda (Amborellaceae). Int J Plant Sci 161(suppl):
S237–S248.Endress PK, DH Lorence 2004 Heterodichogamy of a novel type in
Hernandia (Hernandiaceae) and its structural basis. Int J Plant Sci
165:753–763.
Erbar C, P Leins 1983 Zur Sequenz von Blutenorganen bei einigenMagnoliiden. Bot Jahrb Syst 103:433–449.
Friis EM, PR Crane, KR Pedersen 1991 Stamen diversity and in situpollen of Cretaceous angiosperms. Pages 197–224 in S Blackmore,
SH Barnes, eds. Pollen and spores: patterns of diversification.Clarendon, Oxford.
Friis EM, H Eklund, KR Pedersen, PR Crane 1994 Virginianthuscalycanthoides gen. et sp. nov.: a calycanthaceous flower from thePotomac Group (Early Cretaceous) of eastern North America. Int J
Plant Sci 155:772–785.
Grant V 1950 The pollination of Calycanthus occidentalis. Am J Bot
37:294–297.Herendeen PS, S Magallon-Puebla, R Lupia, PR Crane, J Kobylin-
ska 1999 A preliminary conspectus of the Allon flora from the Late
Cretaceous (Late Santonian) of central Georgia, USA. Ann Mo Bot
Gard 86:407–471.Hiepko P 1965 Vergleichend-morphologische und entwicklungs-
geschichtliche Untersuchungen uber das Perianth bei den Poly-
carpicae. Bot Jahrb Syst 84:359–508.
Hirmer M 1931 Zur Kenntnis der Schraubenstellungen im Pflanzen-reich. Planta 14:132–206.
Hufford LD, PK Endress 1989 The diversity of anther structures and
dehiscence patterns among Hamamelididae. Bot J Linn Soc 99:301–346.
Hyland BPM 1989 A revision of Lauraceae in Australia (excluding
Cassytha). Aust Syst Bot 2:135–367.
Igersheim A, O Cichocki 1996 A simple method for microtomesectioning of prehistoric charcoal specimens, embedded in 2-
hydroxyethyl methacrylate (HEMA). Rev Palaeobot Palynol 92:
389–393.
305STAEDLER ET AL.—FLORAL PHYLLOTAXIS IN CALYCANTHACEAE
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions

Kubitzki K 1969 Monographie der Hernandiaceen. Bot Jahrb Syst89:78–209.
Lasseigne FT, PR Fantz, JC Raulston, GB Straley 2001 x Sinocaly-calycanthus raulstonii (Calycanthaceae): a new intergeneric hybridbetween Sinocalycanthus chinensis and Calycanthus floridus. Am
Soc Hortic Sci 36:765–767.
Li J, J Ledger, T Ward, P del Tredici 2004 Phylogenetics of
Calycanthaceae based on molecular and morphological data with aspecial reference to divergent paralogues of the nrDNA ITS region.
Harv Pap Bot 9:69–82.
Mai HM 1987 Neue Arten nach Fruchten und Samen aus dem Tertiar
von Nordwestsachsen und der Lausitz. Feddes Repert 98:105–126.Mohr BAR, H Eklund 2003 Araripia florifera, a magnoliid angio-
sperm from the lower Cretaceous Crato Formation. Rev Palaeobot
Palynol 126:279–292.Nelson E 1954 Gesetzmassigkeiten der Gestaltwandlung im Bluten-
bereich: ihre Bedeutung fur das Problem der Evolution. Nelson,
Chernex-Montreux.
Nicely KA 1965 A monographic study of the Calycanthaceae.Castanea 30:38–81.
Plantefol L 1949 L’ontogenie de la fleur. Masson, Paris.
Prantl K 1889 Calycanthaceae. Pages 92–94 in A Engler, K Prantl,
eds. Die naturlichen Pflanzenfamilien III. Vol 2. Engelmann, Leipzig.Qiu YL, O Dombrovska, JH Lee, LB Li, BA Whitlock, F
Bernasconi-Quadroni, JS Rest, et al 2005 Phylogenetic analyses
of basal angiosperms based on nine plastid, mitochondrial, andnuclear genes. Int J Plant Sci 166:815–842.
Qiu YL, J Lee, F Bernasconi-Quadroni, DE Soltis, PS Soltis, MJ Zanis,
EA Zimmer, Z Chen, V Savolainen, MW Chase 1999 The earliest
angiosperms: evidence from mitochondrial, plastid and nucleargenomes. Nature 402:404–407.
Rauh W, H Reznik 1951 Histogenetische Untersuchungen an Bluten-
und Infloreszenzachsen. I. Teil. Die Histogenese becherfomiger
Bluten- und Infloreszenzachsen, sowie der Blutenachsen einigerRosoideen. Sitzungsber Heidelb Akad Wiss Math-Natwiss Kl 3:1–71.
Renner SS 1998 Phylogenetic affinities of Monimiaceae based on
cpDNA gene and spacer sequences. Perspect Plant Ecol Syst Evol 1:
61–77.——— 1999 Circumscription and phylogeny of the Laurales: evi-
dence from molecular and morphological data. Am J Bot 86:
1301–1315.——— 2004 Variation in diversity among Laurales, Early Cretaceous
to present. Biol Skr 55:441–458.
Ronse De Craene LP, PS Soltis, DE Soltis 2003 Evolution of floralstructures in basal angiosperms. Int J Plant Sci 164(suppl):
S329–S363.
Shutts CF 1959 The comparative morphology of the Hernandiaceae.PhD diss, Indiana University. University Microfilms, Ann Arbor, MI.
Singh V, A Singh 1985 Floral organogenesis in Cinnamomumcamphora. Phytomorphology 35:61–67.
Soltis DE, PS Soltis, MW Chase, ME Mort, DC Albach, MJ Zanis, VSavolainen, et al 2000 Angiosperm phylogeny inferred from 18S
rDNA, rbcL, and atpB sequences. Bot J Linn Soc 133:381–461.
Takhtajan AL 1959 Die Evolution der Angiospermen. Fischer, Jena.
——— 1987 Systema magnoliophytorum. Nauka, Leningrad.——— 1997 Diversity and classification of flowering plants. Colum-
bia University Press, New York.
Thorne RF 1974 A phylogenetic classification of the Annoniflorae.Aliso 8:147–209.
——— 1992 An updated phylogenetic classification of the flowering
plants. Aliso 13:365–389.
——— 2000 The classification and geography of the floweringplants: dicotyledons of the class Angiospermae (subclasses Magno-
liidae, Ranunculidae, Caryophyllidae, Dilleniidae, Rosidae, Aster-
idae, and Lamiidae). Bot Rev 66:441–647.
Unger F 1852 Botanische Briefe. Gerold, Vienna.van der Pijl L 1982 Principles of dispersal in higher plants. 3rd ed.
Springer, Berlin.
Vogel S 1998 Remarkable nectaries: structure, ecology, organophy-letic perspectives. II. Nectarioles. Flora 193:1–29.
Wilson CL 1976 Floral anatomy of Idiospermum australiense(Idiospermaceae). Am J Bot 63:987–996.
Worboys SJ 2003 Polycarpelly in Idiospermum australiense (Caly-canthaceae). Austrobaileya 6:553–556.
Worboys SJ, BR Jackes 2005 Pollination processes in Idiospermumaustraliense (Calycanthaceae), an arborescent basal angiosperm of
Australia’s tropical rain forest. Plant Syst Evol 251:107–117.Zanis MJ, PS Soltis, YL Qiu, EA Zimmer, DE Soltis 2003 Phyloge-
netic analyses and perianth evolution in basal angiosperms. Ann Mo
Bot Gard 90:129–150.
Zanis MJ, DE Soltis, PS Soltis, S Mathews, MJ Donoghue 2002 Theroot of the angiosperms revisited. Proc Natl Acad Sci USA 99:
6848–6853.
Zhou S, SS Renner, J Wen 2006 Molecular phylogeny and intra- andintercontinental biogeography of Calycanthaceae. Mol Phylogenet
Evol 39:1–15.
306 INTERNATIONAL JOURNAL OF PLANT SCIENCES
This content downloaded from 131.130.131.58 on Thu, 15 Aug 2013 15:47:15 PMAll use subject to JSTOR Terms and Conditions




















