
Implications of iron solubilization on soil phosphorus release in seasonally flooded forests of the...
6
Short communication Implications of iron solubilization on soil phosphorus release in seasonally flooded forests of the lower Orinoco River, Venezuela Noemi Chacon * , Saul Flores, Ana Gonzalez Centro de Ecologı ´a, Instituto Venezolano de Investigaciones Cientı ´ficas, Apdo. 21827, Caracas 1020 A, Venezuela Received 19 April 2005; received in revised form 7 October 2005; accepted 26 October 2005 Available online 4 January 2006 Abstract The microbial reduction of Fe oxides is thought to contribute with the release of P in sedimentary environments. However, secondary reactions of the bioproduced Fe(II) with P in solution, can lead to a decrease in the soluble P concentration. In this study, we examined how the reduction of Fe(III) affects the soluble P concentration, when the soils of a seasonally flooded forest gradient are subjected to anaerobic conditions. Soil samples were collected during the dry season from two zones subjected to different flooding intensity: MAX and MIN zones that were inundated 8 and 2 months per year, respectively. When anaerobic conditions were applied to soils from both zones, a clear stimulatory effect on the Fe(III) reduction was observed. However, bioproduced Fe(II) underwent secondary chemical reactions, masking the extend of Fe(III) reduction of these soils. Iron was reduced mainly during the first 15 days of the anaerobic incubation and it was stimulated by a pulse of labile carbon. Iron dissolution did not lead to an increase of the soluble P content. However, in both zones P was high and positively correlated with Fe(II), implying that soil P mobilization was linked to Fe dissolution. In the MIN zone, soluble P concentration decreased, probably as a consequence of the secondary reactions of solubilized P with other non-redox sensitive soils elements. Fe solubilization also had an effect on the activity of acid phosphatase and consequently in the solubilization of P from the organic pool. In conclusion, the P cycle in these soils is strongly coupled to C and Fe cycles. q 2005 Elsevier Ltd. All rights reserved. Keywords: Anaerobic soils; Iron dissolution; Phosphorus mobilization Extensive areas of river watersheds in the tropics are covered with forests, some of which are seasonally flooded (Kubitzky, 1989). These forests are characterized by annual oscillations between aquatic and terrestrial phases (Junk et al., 1989). The productivity of these forests in the humid tropics is thought to be limited by the phosphorus (P) supply (Vitousek, 1984). Soils in this region are nearly depleted of primary P-containing minerals and most of this element is found pre- dominantely chemisorbed on amorphous minerals and some is also found on crystalline iron (Fe) and aluminum (Al) oxides (Lo ´pez-Herna ´ndez, 1977; Hsu, 1977; Schwertmann and Taylor, 1977; Parfitt, 1978; Parfitt and Smart, 1978). For these ecosystems, soil P mobilization has been associated with the biogeochemistry of Fe (Gambrel and Patrick, 1978; Baldwin and Mitchell, 2000; Chacon et al., 2005a). Water in soil restricts gas diffusion and limits the oxygen availability in soil (Smith and Tiedje, 1979), with a consequent decrease in the soil redox potential (Gambrel and Patrick, 1978). Under such circumstances, microbial dissolution of Fe(III) oxides can take place (Lovley et al., 1991; Baldwin et al., 1997; Baldwin and Mitchell, 2000) and the phosphate anion chemically sorbed to these iron oxides surfaces can be released (Baldwin and Mitchell, 2000). Indeed, several studies have found that P concentration increases when soils become waterlogged (Moore et al., 1992; Szilas et al., 1998; Ferrando et al., 2002). However, Fe(II) produced by the reduction of Fe(III) oxides may undergo chemical transformations, affect- ing the concentration of P in solution. For example, ferric hydrous oxides can be reduced to amorphous ferrous hydroxide gel complexes (Fe(OH) 2 )(Ponnamperuma et al., 1967), with a greater surface area than the original ferric compound (Holford and Patrick, 1979). This results in an increase in P sorption capacity (Patrick and Khalid, 1974; Willet and Higgins, 1978; Holford and Patrick, 1979; Phillips and Greenway, 1998). Phosphorus concentration may also decrease during flooding due to the precipitation of ferrous phosphate (vivianite, Fe 3 (PO 4 ) 2 $8H 2 O) (Zachara et al., 1998). In a previous study, along a flooded gradient, Chaco ´n et al. (2005a) found that both labile and moderately labile soil P Soil Biology & Biochemistry 38 (2006) 1494–1499 www.elsevier.com/locate/soilbio 0038-0717/$ - see front matter q 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.soilbio.2005.10.018 * Corresponding author. Tel.: C58 212 5041415; fax: C58 212 5041088. E-mail address: [email protected] (N. Chacon).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Implications of iron solubilization on soil phosphorus release in seasonally flooded forests of the...

Short communication
Implications of iron solubilization on soil phosphorus release in seasonally
flooded forests of the lower Orinoco River, Venezuela
Noemi Chacon *, Saul Flores, Ana Gonzalez
Centro de Ecologıa, Instituto Venezolano de Investigaciones Cientıficas, Apdo. 21827, Caracas 1020 A, Venezuela
Received 19 April 2005; received in revised form 7 October 2005; accepted 26 October 2005
Available online 4 January 2006
Abstract
The microbial reduction of Fe oxides is thought to contribute with the release of P in sedimentary environments. However, secondary reactions
of the bioproduced Fe(II) with P in solution, can lead to a decrease in the soluble P concentration. In this study, we examined how the reduction of
Fe(III) affects the soluble P concentration, when the soils of a seasonally flooded forest gradient are subjected to anaerobic conditions. Soil
samples were collected during the dry season from two zones subjected to different flooding intensity: MAX andMIN zones that were inundated 8
and 2 months per year, respectively. When anaerobic conditions were applied to soils from both zones, a clear stimulatory effect on the Fe(III)
reduction was observed. However, bioproduced Fe(II) underwent secondary chemical reactions, masking the extend of Fe(III) reduction of these
soils. Iron was reduced mainly during the first 15 days of the anaerobic incubation and it was stimulated by a pulse of labile carbon. Iron
dissolution did not lead to an increase of the soluble P content. However, in both zones P was high and positively correlated with Fe(II), implying
that soil P mobilization was linked to Fe dissolution. In the MIN zone, soluble P concentration decreased, probably as a consequence of the
secondary reactions of solubilized P with other non-redox sensitive soils elements. Fe solubilization also had an effect on the activity of acid
phosphatase and consequently in the solubilization of P from the organic pool. In conclusion, the P cycle in these soils is strongly coupled to C and
Fe cycles.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: Anaerobic soils; Iron dissolution; Phosphorus mobilization
Extensive areas of river watersheds in the tropics are
covered with forests, some of which are seasonally flooded
(Kubitzky, 1989). These forests are characterized by annual
oscillations between aquatic and terrestrial phases (Junk et al.,
1989). The productivity of these forests in the humid tropics is
thought to be limited by the phosphorus (P) supply (Vitousek,
1984). Soils in this region are nearly depleted of primary
P-containing minerals and most of this element is found pre-
dominantely chemisorbed on amorphous minerals and some is
also found on crystalline iron (Fe) and aluminum (Al) oxides
(Lopez-Hernandez, 1977; Hsu, 1977; Schwertmann and
Taylor, 1977; Parfitt, 1978; Parfitt and Smart, 1978). For
these ecosystems, soil P mobilization has been associated with
the biogeochemistry of Fe (Gambrel and Patrick, 1978;
Baldwin and Mitchell, 2000; Chacon et al., 2005a).
Water in soil restricts gas diffusion and limits the oxygen
availability in soil (Smith and Tiedje, 1979), with a consequent
0038-0717/$ - see front matter q 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.soilbio.2005.10.018
* Corresponding author. Tel.: C58 212 5041415; fax: C58 212 5041088.
E-mail address: [email protected] (N. Chacon).
decrease in the soil redox potential (Gambrel and Patrick,
1978). Under such circumstances, microbial dissolution of
Fe(III) oxides can take place (Lovley et al., 1991; Baldwin
et al., 1997; Baldwin and Mitchell, 2000) and the phosphate
anion chemically sorbed to these iron oxides surfaces can be
released (Baldwin and Mitchell, 2000). Indeed, several studies
have found that P concentration increases when soils become
waterlogged (Moore et al., 1992; Szilas et al., 1998; Ferrando
et al., 2002). However, Fe(II) produced by the reduction of
Fe(III) oxides may undergo chemical transformations, affect-
ing the concentration of P in solution. For example, ferric
hydrous oxides can be reduced to amorphous ferrous hydroxide
gel complexes (Fe(OH)2) (Ponnamperuma et al., 1967), with a
greater surface area than the original ferric compound (Holford
and Patrick, 1979). This results in an increase in P sorption
capacity (Patrick and Khalid, 1974; Willet and Higgins, 1978;
Holford and Patrick, 1979; Phillips and Greenway, 1998).
Phosphorus concentration may also decrease during flooding
due to the precipitation of ferrous phosphate (vivianite,
Fe3(PO4)2$8H2O) (Zachara et al., 1998).
In a previous study, along a flooded gradient, Chacon et al.
(2005a) found that both labile and moderately labile soil P
Soil Biology & Biochemistry 38 (2006) 1494–1499
www.elsevier.com/locate/soilbio
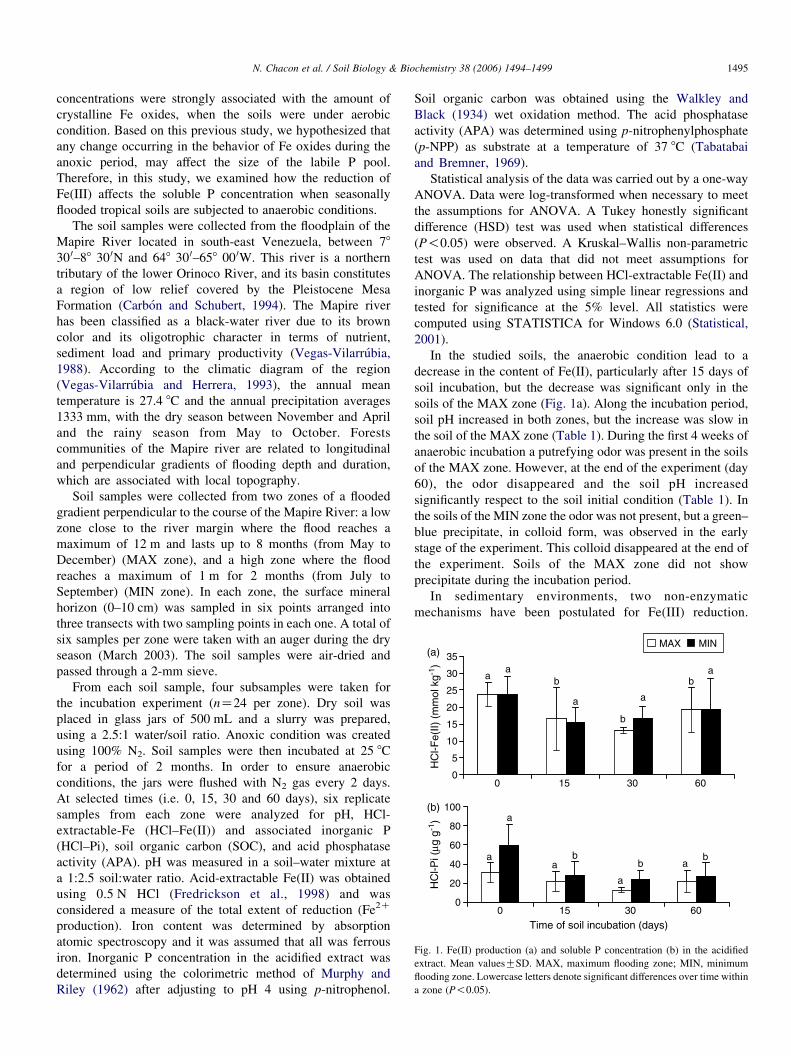
Fig. 1. Fe(II) production (a) and soluble P concentration (b) in the acidified
extract. Mean valuesGSD. MAX, maximum flooding zone; MIN, minimum
flooding zone. Lowercase letters denote significant differences over time within
a zone (P!0.05).
N. Chacon et al. / Soil Biology & Biochemistry 38 (2006) 1494–1499 1495
concentrations were strongly associated with the amount of
crystalline Fe oxides, when the soils were under aerobic
condition. Based on this previous study, we hypothesized that
any change occurring in the behavior of Fe oxides during the
anoxic period, may affect the size of the labile P pool.
Therefore, in this study, we examined how the reduction of
Fe(III) affects the soluble P concentration when seasonally
flooded tropical soils are subjected to anaerobic conditions.
The soil samples were collected from the floodplain of the
Mapire River located in south-east Venezuela, between 78
30 0–88 30 0N and 648 30 0–658 00 0W. This river is a northern
tributary of the lower Orinoco River, and its basin constitutes
a region of low relief covered by the Pleistocene Mesa
Formation (Carbon and Schubert, 1994). The Mapire river
has been classified as a black-water river due to its brown
color and its oligotrophic character in terms of nutrient,
sediment load and primary productivity (Vegas-Vilarrubia,
1988). According to the climatic diagram of the region
(Vegas-Vilarrubia and Herrera, 1993), the annual mean
temperature is 27.4 8C and the annual precipitation averages
1333 mm, with the dry season between November and April
and the rainy season from May to October. Forests
communities of the Mapire river are related to longitudinal
and perpendicular gradients of flooding depth and duration,
which are associated with local topography.
Soil samples were collected from two zones of a flooded
gradient perpendicular to the course of the Mapire River: a low
zone close to the river margin where the flood reaches a
maximum of 12 m and lasts up to 8 months (from May to
December) (MAX zone), and a high zone where the flood
reaches a maximum of 1 m for 2 months (from July to
September) (MIN zone). In each zone, the surface mineral
horizon (0–10 cm) was sampled in six points arranged into
three transects with two sampling points in each one. A total of
six samples per zone were taken with an auger during the dry
season (March 2003). The soil samples were air-dried and
passed through a 2-mm sieve.
From each soil sample, four subsamples were taken for
the incubation experiment (nZ24 per zone). Dry soil was
placed in glass jars of 500 mL and a slurry was prepared,
using a 2.5:1 water/soil ratio. Anoxic condition was created
using 100% N2. Soil samples were then incubated at 25 8C
for a period of 2 months. In order to ensure anaerobic
conditions, the jars were flushed with N2 gas every 2 days.
At selected times (i.e. 0, 15, 30 and 60 days), six replicate
samples from each zone were analyzed for pH, HCl-
extractable-Fe (HCl–Fe(II)) and associated inorganic P
(HCl–Pi), soil organic carbon (SOC), and acid phosphatase
activity (APA). pH was measured in a soil–water mixture at
a 1:2.5 soil:water ratio. Acid-extractable Fe(II) was obtained
using 0.5 N HCl (Fredrickson et al., 1998) and was
considered a measure of the total extent of reduction (Fe2C
production). Iron content was determined by absorption
atomic spectroscopy and it was assumed that all was ferrous
iron. Inorganic P concentration in the acidified extract was
determined using the colorimetric method of Murphy and
Riley (1962) after adjusting to pH 4 using p-nitrophenol.
Soil organic carbon was obtained using the Walkley and
Black (1934) wet oxidation method. The acid phosphatase
activity (APA) was determined using p-nitrophenylphosphate
(p-NPP) as substrate at a temperature of 37 8C (Tabatabai
and Bremner, 1969).
Statistical analysis of the data was carried out by a one-way
ANOVA. Data were log-transformed when necessary to meet
the assumptions for ANOVA. A Tukey honestly significant
difference (HSD) test was used when statistical differences
(P!0.05) were observed. A Kruskal–Wallis non-parametric
test was used on data that did not meet assumptions for
ANOVA. The relationship between HCl-extractable Fe(II) and
inorganic P was analyzed using simple linear regressions and
tested for significance at the 5% level. All statistics were
computed using STATISTICA for Windows 6.0 (Statistical,
2001).
In the studied soils, the anaerobic condition lead to a
decrease in the content of Fe(II), particularly after 15 days of
soil incubation, but the decrease was significant only in the
soils of the MAX zone (Fig. 1a). Along the incubation period,
soil pH increased in both zones, but the increase was slow in
the soil of the MAX zone (Table 1). During the first 4 weeks of
anaerobic incubation a putrefying odor was present in the soils
of the MAX zone. However, at the end of the experiment (day
60), the odor disappeared and the soil pH increased
significantly respect to the soil initial condition (Table 1). In
the soils of the MIN zone the odor was not present, but a green–
blue precipitate, in colloid form, was observed in the early
stage of the experiment. This colloid disappeared at the end of
the experiment. Soils of the MAX zone did not show
precipitate during the incubation period.
In sedimentary environments, two non-enzymatic
mechanisms have been postulated for Fe(III) reduction.
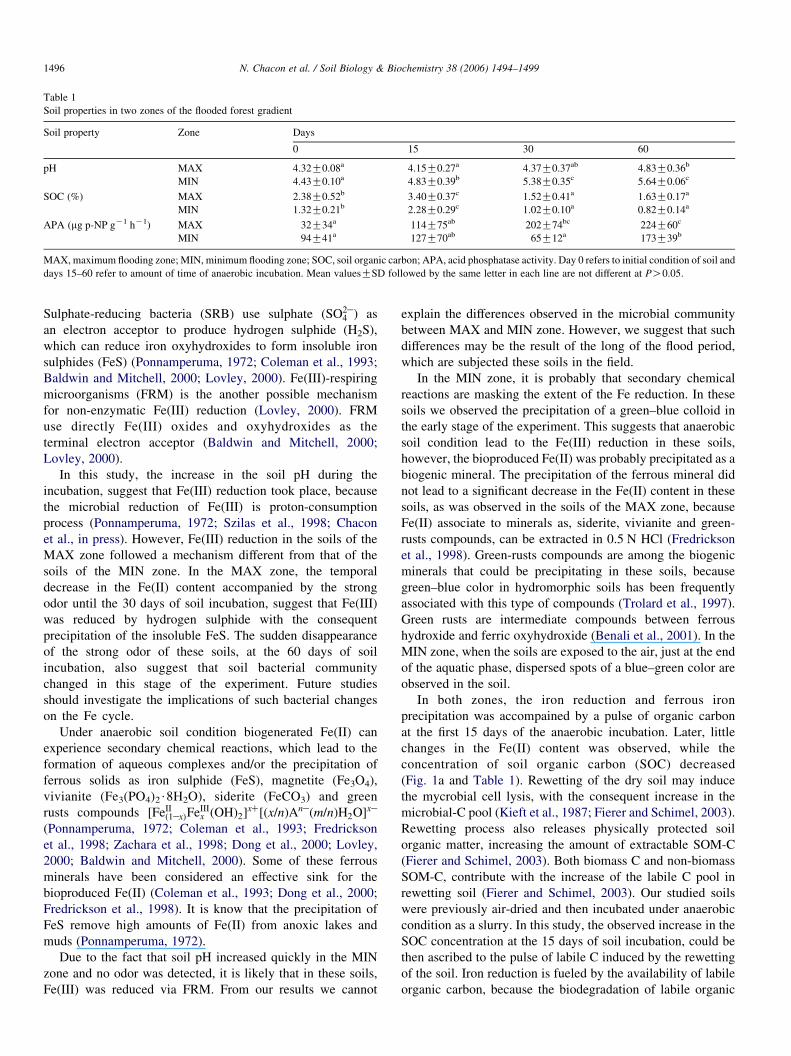
Table 1
Soil properties in two zones of the flooded forest gradient
Soil property Zone Days
0 15 30 60
pH MAX 4.32G0.08a 4.15G0.27a 4.37G0.37ab 4.83G0.36b
MIN 4.43G0.10a 4.83G0.39b 5.38G0.35c 5.64G0.06c
SOC (%) MAX 2.38G0.52b 3.40G0.37c 1.52G0.41a 1.63G0.17a
MIN 1.32G0.21b 2.28G0.29c 1.02G0.10a 0.82G0.14a
APA (mg p-NP gK1 hK1) MAX 32G34a 114G75ab 202G74bc 224G60c
MIN 94G41a 127G70ab 65G12a 173G39b
MAX, maximum flooding zone; MIN, minimum flooding zone; SOC, soil organic carbon; APA, acid phosphatase activity. Day 0 refers to initial condition of soil and
days 15–60 refer to amount of time of anaerobic incubation. Mean valuesGSD followed by the same letter in each line are not different at PO0.05.
N. Chacon et al. / Soil Biology & Biochemistry 38 (2006) 1494–14991496
Sulphate-reducing bacteria (SRB) use sulphate (SO2K4 ) as
an electron acceptor to produce hydrogen sulphide (H2S),
which can reduce iron oxyhydroxides to form insoluble iron
sulphides (FeS) (Ponnamperuma, 1972; Coleman et al., 1993;
Baldwin and Mitchell, 2000; Lovley, 2000). Fe(III)-respiring
microorganisms (FRM) is the another possible mechanism
for non-enzymatic Fe(III) reduction (Lovley, 2000). FRM
use directly Fe(III) oxides and oxyhydroxides as the
terminal electron acceptor (Baldwin and Mitchell, 2000;
Lovley, 2000).
In this study, the increase in the soil pH during the
incubation, suggest that Fe(III) reduction took place, because
the microbial reduction of Fe(III) is proton-consumption
process (Ponnamperuma, 1972; Szilas et al., 1998; Chacon
et al., in press). However, Fe(III) reduction in the soils of the
MAX zone followed a mechanism different from that of the
soils of the MIN zone. In the MAX zone, the temporal
decrease in the Fe(II) content accompanied by the strong
odor until the 30 days of soil incubation, suggest that Fe(III)
was reduced by hydrogen sulphide with the consequent
precipitation of the insoluble FeS. The sudden disappearance
of the strong odor of these soils, at the 60 days of soil
incubation, also suggest that soil bacterial community
changed in this stage of the experiment. Future studies
should investigate the implications of such bacterial changes
on the Fe cycle.
Under anaerobic soil condition biogenerated Fe(II) can
experience secondary chemical reactions, which lead to the
formation of aqueous complexes and/or the precipitation of
ferrous solids as iron sulphide (FeS), magnetite (Fe3O4),
vivianite (Fe3(PO4)2$8H2O), siderite (FeCO3) and green
rusts compounds ½FeIIð1KxÞFeIIIx ðOHÞ2�
xC½ðx=nÞAnKðm=nÞH2O�xK
(Ponnamperuma, 1972; Coleman et al., 1993; Fredrickson
et al., 1998; Zachara et al., 1998; Dong et al., 2000; Lovley,
2000; Baldwin and Mitchell, 2000). Some of these ferrous
minerals have been considered an effective sink for the
bioproduced Fe(II) (Coleman et al., 1993; Dong et al., 2000;
Fredrickson et al., 1998). It is know that the precipitation of
FeS remove high amounts of Fe(II) from anoxic lakes and
muds (Ponnamperuma, 1972).
Due to the fact that soil pH increased quickly in the MIN
zone and no odor was detected, it is likely that in these soils,
Fe(III) was reduced via FRM. From our results we cannot
explain the differences observed in the microbial community
between MAX and MIN zone. However, we suggest that such
differences may be the result of the long of the flood period,
which are subjected these soils in the field.
In the MIN zone, it is probably that secondary chemical
reactions are masking the extent of the Fe reduction. In these
soils we observed the precipitation of a green–blue colloid in
the early stage of the experiment. This suggests that anaerobic
soil condition lead to the Fe(III) reduction in these soils,
however, the bioproduced Fe(II) was probably precipitated as a
biogenic mineral. The precipitation of the ferrous mineral did
not lead to a significant decrease in the Fe(II) content in these
soils, as was observed in the soils of the MAX zone, because
Fe(II) associate to minerals as, siderite, vivianite and green-
rusts compounds, can be extracted in 0.5 N HCl (Fredrickson
et al., 1998). Green-rusts compounds are among the biogenic
minerals that could be precipitating in these soils, because
green–blue color in hydromorphic soils has been frequently
associated with this type of compounds (Trolard et al., 1997).
Green rusts are intermediate compounds between ferrous
hydroxide and ferric oxyhydroxide (Benali et al., 2001). In the
MIN zone, when the soils are exposed to the air, just at the end
of the aquatic phase, dispersed spots of a blue–green color are
observed in the soil.
In both zones, the iron reduction and ferrous iron
precipitation was accompained by a pulse of organic carbon
at the first 15 days of the anaerobic incubation. Later, little
changes in the Fe(II) content was observed, while the
concentration of soil organic carbon (SOC) decreased
(Fig. 1a and Table 1). Rewetting of the dry soil may induce
the mycrobial cell lysis, with the consequent increase in the
microbial-C pool (Kieft et al., 1987; Fierer and Schimel, 2003).
Rewetting process also releases physically protected soil
organic matter, increasing the amount of extractable SOM-C
(Fierer and Schimel, 2003). Both biomass C and non-biomass
SOM-C, contribute with the increase of the labile C pool in
rewetting soil (Fierer and Schimel, 2003). Our studied soils
were previously air-dried and then incubated under anaerobic
condition as a slurry. In this study, the observed increase in the
SOC concentration at the 15 days of soil incubation, could be
then ascribed to the pulse of labile C induced by the rewetting
of the soil. Iron reduction is fueled by the availability of labile
organic carbon, because the biodegradation of labile organic

N. Chacon et al. / Soil Biology & Biochemistry 38 (2006) 1494–1499 1497
matter leads to the production of electrons which can be
microbially transfered to the Fe(III), an important electron
acceptor under anaerobic soil conditions (Lovley and Phillips,
1986; Quantin et al., 2001; 2002; Stemmler and Berthelin,
2003; Chacon et al., in press). The depletion of organic C in the
later stages of the experiment (30–60 days), suggest that Fe(III)
reduction was limited by the supply of labile organic C in this
stage of the experiment.
It has been reported that in waterlogged soils, the
microbial reduction of Fe(III) is greatly stimulated by
organic matter inputs. In natural areas oscillating between
terrestrial and aquatic phases, the microbial reduction of
Fe(III) will primarily be carried out during the initial period
of soil anoxia because pulses of labile C are available during
the aquatic phase as a result of both the leaching of the
accumulated litter during the terrestrial phase and by the
pulse of labile C induced by the rewetting of the soils (Kieft
et al., 1987; Junk, 1997; O‘Connell et al., 2000; Fierer and
Schimel, 2003).
During the first 15 days of anaerobic incubation, the content
of soluble P declined respect to the initial levels in the soils of
the MIN zone, but there was no significant changes in the
content of P in the soils of the MAX zone (Fig. 1b). In both
zones, P content was high and positively correlated with Fe(II)
content (MAX: r2Z0.53, P!0.0001; MIN: r2Z0.41,
P!0.0001). A recent study by Chacon et al. (2005a) suggests
that the microbial reduction of Fe(III) is an important
mechanism for P release in soils subjected to long flooding
periods. Although, we cannot corroborate this hypothesis with
the available data from this study, the positive relationship
between both Fe(II) and P content allow us to suggests that the
P mobilization in the studied soils is linked to Fe(II)
dissolution. However, under anaerobic soil condition Fe and
P undergo secondary chemical reactions that could be
preventing to observe a significant increase in the content of
both elements.
The sharp decrease in the P content in the soils of the MIN
zone could be ascribed to secondary reactions of P with other
non-redox sensitive soil components. In oxidized soil samples
of the MIN zone, Chacon et al. (2005a) reported high contents
of humic–Al-complexes (174.01G19.01 mmol kgK1)
compared to the soils of the MAX zone (99.65G35.36 mmol kgK1). Aluminum is a non-redox sensitive
element and the organo-mineral–Al complexes are considered
active surfaces that control the soil’s sorption capacity (Gerke
and Jungk, 1991; Darke and Walbridge, 2000; Chacon and
Dezzeo, 2004).
Acid phosphatase activity (APA) increased gradually in
the soils of the MAX zone (Table 1). However, in the soils
of the MIN zone, APA showed no significant changes during
the first 30 days of soil incubation, but it increased
significantly at the end of the experiment (Table 1). The
differences in the APA behavior could be explained through
the interaction of this enzyme with various soil components.
According to Chacon et al. (2005b), during the non-flooded
phase, acid phosphatase seems to be adsorbed by minerals of
the fine fraction of the soils (clayCsilt) in the MAX zone
and by organo-mineral–Al–complexes in the MIN zone. As
was previously disscussed, organo-mineral–Al-complexes are
not sensitive to the changes in the soil redox condition.
Therefore, it is logical that APA in the soil of the MIN zone
did not respond to the anaerobic soil condition. The gradual
increases of the enzyme activity as the Fe dissolution took
place in the MAX soils, suggests that probably immobilized
enzyme is being released during the Fe(III) reduction. It has
been reported that the activity of acid phosphatase in solution
is much higher than the activity of the immobilized enzyme
(Rao et al., 1996; Shindo et al., 2002). Even though a net
change in the P content was not observed in the soils of the
MAX zone, it is clear that the increase in the enzimatic
activity, can lead to the transference of P from the organic
pool to the soil solution.
Our findings showed that anaerobic condition coupled with
inputs of labile carbon stimulate iron reduction in the studied
soils. We propose that two different mechanisms are involved
in Fe(III) reduction: in the MAX zone, Fe is indirectly reduced
by the activity of sulphate-reducing bacteria, through the
production of sulphide, while in the MIN zone, Fe is reduced
directly by Fe(III)-respiring microorganisms. Under anoxic
soil condition P mobilization was linked to the Fe dissolution.
However, both biogenerated Fe(II) and the released P,
underwent secondary chemical reactions, which could be
masking the extend of iron reduction and consequently the
transference of P from this pool to the soil solution. Fe
solubilization seems to have a significant effect on the activity
of the immobilized acid phosphatase and consequently in the
solubilization of P from the organic pool. The results in this
study allowed us to suggest a number of hypotheses concerning
the mechanisms that regulate the iron dissolution and
phosphorus release in seasonal flooded forests. However,
future investigations should include long-term studies in the
field in combination with simulations in the laboratory that
allow to understand, the composition and activity of the soil
microbial community, linked to the geochemical
transformations.
Acknowledgements
The authors thank two anonymous reviewers who provided
helpful comments on this manuscript.
References
Baldwin, D.S., Mitchell, A.M., 2000. The effects of drying and re-flooding on
the sediment and soil nutrient dynamics of lowland river-floodplain
systems: a synthesis. Regulated Rivers—Research and Management 16,
457–467.
Baldwin, D.S., Mitchell, A.M., Rees, G., 1997. Chemistry and microbial
ecology: processes at the microscale. In: Klomp, N., Lunt, I. (Eds.),
Frontiers in Ecology–Building the Links. Elsevier, Oxford, pp. 171–179.
Benali, O., Abdelmoula, M., Refait, P., Genin, J.M.R., 2001. Effect of
ortophosphate on the oxidation products of Fe(II)–Fe(III) hydroxycarbo-
nate: the transformation of green rust to ferrihydrite. Geochimica et
Cosmochimica Acta 65, 1715–1726.

N. Chacon et al. / Soil Biology & Biochemistry 38 (2006) 1494–14991498
Carbon, J., Schubert, C., 1994. Late Cenozoic history of the eastern Llanos of
Venezuela: geomorphology and stratigraphy of the Mesa formation.
Quaternary International 21, 91–100.
Chacon, N., Dezzeo, N., 2004. Phosphorus fractions and sorption processes in
soil samples taken in a forest-savanna sequence of the Gran Sabana in
southern Venezuela. Biology and Fertility of Soils 40, 14–19.
Chacon, N., Dezzeo, N., Munoz, B., Rodrıguez, J.M., 2005a. Implications of
soil organic carbon and the biogeochemistry of iron and aluminum on soil
phosphorus distribution in flooded forests of the lower Orinoco River,
Venezuela. Biogeochemistry 73, 555–556.
Chacon, N., Dezzeo, N., Flores, S., 2005b. Effect of particle-size distribution,
soil organic carbon content and organo-mineral aluminum complexes on
acid phosphatases of seasonally flooded forest soils. Biology and Fertility
of Soils 41, 69–72.
Chacon, N., Silver, W.L., Dubinsky, E.A., Cusack, D.F., in press. Iron
reduction and soil phosphorus solubilization in humid tropical forests soils:
the roles of labile carbon pools and an electron shuttle compound.
Biogeochemistry.
Coleman, M.L., Hedrick, D.B., Lovley, D.R., White, D.C., Pye, K., 1993.
Reduction of Fe(III) in sediments by sulphate-reducing bacteria. Nature
361, 436–438.
Darke, A.K., Walbridge, M.R., 2000. Al and Fe biogeochemistry in a floodplain
forest: implications for P retention. Biogeochemistry 51, 1–32.
Dong, H., Fredrickson, J.K., Kennedy, D.W., Zachara, J.M., Kukkadapu, R.K.,
Onstott, T.C., 2000. Mineral transformation associated with the microbial
reduction of magnetite. Chemical Geology 169, 299–318.
Ferrando, M., Mercado, G., Hernandez, J., 2002. Dinamica del hierro y
disponibilidad de fosforo durante perıodos cortos de anaerobiosis en los
suelos. Agrociencia VI, 1–9.
Fierer, N., Schimel, J.P., 2003. A proposed mechanism for the pulse in
carbon dioxide. Production commonly observed following the rapid
rewetting of a dry soil. Soil Science Society of America Journal 67,
798–805.
Fredrickson, J.K., Zachara, J.M., Kennedy, D.W., Dong, H., Onstott, T.C.,
Hinman, N.W., Li, S., 1998. Biogenic iron mineralization accompanying
the dissimilatory reduction of hydrous ferric oxide by a groundwater
bacterium. Geochimica et Cosmochimica Acta 62, 3239–3257.
Gambrel, R.P., Patrick, W.H., 1978. Chemical and microbiological properties
of anaerobic soils and sediments. In: Hook, D.D., Crawford, R.M.M. (Eds.),
Plant Life in Anaerobic Environments. Ann Arbor Science, Ann Arbor, MI,
pp. 233–247.
Gerke, J., Jungk, A., 1991. Separation of phosphorus bound to organic
matrices from inorganic phosphorus in alkaline soil extracts by
ultrafiltration. Communications in Soil Science and Plant Analysis 22,
1621–1630.
Holford, I.C.R., Patrick, W.H., 1979. Effects of reduction and pH changes on
phosphate sorption and mobility in an acid soil. Soil Science Society of
America Journal 43, 292–296.
Hsu, P.H., 1977. Aluminum oxides and oxyhidroxides. In: Dixon, J.B.,
Weed, S.B. (Eds.), Minerals in Soil Environments. Soil Science Society
America, Madison, WI, pp. 99–143.
Junk, W.J., 1997. General aspects of floodplain ecology with special reference
to Amazonian floodplains. In: Junk, W.J. (Ed.), The Central Amazon
Floodplain. Ecology of a Pulsing System Ecological Studies. Springer,
Berlin, pp. 3–20.
Junk, W.J., Bayley, P.B., Sparks, R.E., 1989. The flood pulse concept in
river–floodplain systems. In: Dodge D.P., (Ed.). Proceedings of the
International Large River Symposium. Canadian Special Publication of
Fisheries and Aquatic Sciences 106, pp. 110–127.
Kieft, T., Soroker, E., Firestone, M., 1987. Microbial biomass response to a
rapid increase in water potential when dry soil is wetted. Soil Biology &
Biochemistry 19, 119–126.
Kubitzky, K., 1989. The ecogeographical differentiation of Amazonian
inundation forests. Plant Systematics and Evolution 162, 285–304.
Lopez-Hernandez, D., 1977. La quımica del fosforo en suelos acidos.
Universidad Central de Venezuela, Ediciones de la Biblioteca. Caracas.
Lovley, D.R., 2000. Fe(III) and Mn(IV) reduction. In: Lovley, D.R. (Ed.),
Environmental Microbe–Metal Interactions. ASM Press, Washington, DC,
pp. 3–30.
Lovley, D.R., Phillips, E.J.P., 1986. Organic matter mineralization with
reduction of ferric iron in anaerobic sediments. Applied and Environmental
Microbiology 51, 683–689.
Lovley, D.R., Phillips, E.J.P., Lonergan, D.J., 1991. Enzymatic versus
nonenzymatic mechanims for Fe (III) reduction in aquatic sediments.
Environmental Science & Technology 25, 1062–1067.
Moore, P.A., Reddy, K.R., Graetz, D.A., 1992. Nutrient transformations in
sediments as influenced by oxygen supply. Journal of Environmental
Quality 21, 387–393.
Murphy, J., Riley, J.P., 1962. A modified single solution method for the
determination of phosphate in natural waters. Analitica Chimica Acta 27,
31–36.
O’Connel, M., Daldwin, D.S., Robertson, A.I., Rees, G., 2000. Release
and bioavailability of dissolved organic matter from floodplain litter:
influence of origin and oxygen levels. Freshwater Biology 45,
333–342.
Parfitt, R.L., 1978. Anion adsorption by soils and soil materials. Advances in
Agronomy 30, 1–50.
Parfitt, R.L., Smart, R.St C., 1978. The mechanism of sulfate adsorption on iron
oxides. Soil Science Society of America Journal 42, 48–50.
Patrick, W.K., Khalid, R.A., 1974. Phosphate release and sorption by soils
and sediments: effect of aerobic and anaerobic conditions. Science
186, 53–55.
Phillips, I.R., Greenway, M., 1998. Changes in water-soluble exchangeable
ions, cation exchange capacity, and phosphorusmax in soils under
alternanting waterlogged and drying conditions. Communications in Soil
Science and Plant Analysis 29, 51–65.
Ponnamperuma, F.N., 1972. The chemistry of submerged soils. Advances in
Agronomy 26, 29–96.
Ponnamperuma, F.N., Tianco, E.M., Loy, T., 1967. Redox equilibria in flooded
soils, I. The iron hydroxide systems. Soil Science 103, 374–382.
Quantin, C., Becquer, T., Rouiller, J.H., Berthelin, J., 2001. Oxide weathering
and trace metal release by bacterial reduction in a New Caledonia Ferralsol.
Biogeochemistry 53, 323–340.
Quantin, C., Becquer, T., Rouiller, J.H., Berthelin, J., 2002. Redistribution of
metals in a New Caledonia Ferralsol after microbial weathering. Soil
Science Society of America Journal 66, 1797–1804.
Rao, M.A., Gianfreda, L., Palmiero, F., Violante, A., 1996. Interaction of acid
phosphatase with clays, organic molecules and organo-mineral complexes.
Soil Science 161, 751–760.
Schwertmann, U., Taylor, R.M., 1977. Iron oxides. In: Dixon, J.B., Weed, S.B.
(Eds.), Minerals in Soil Environments. Soil Science Society America,
Madison, WI, pp. 145–180.
Shindo, H., Watanabe, D., Onaga, T., Urakawa, M., Nakahara, O., Huang, Q.,
2002. Adsorption, activity, and kinetics of acid phosphatase as influenced
by selected oxides and clay minerals. Soil Science and Plant Nutrition 48,
763–767.
Smith, M.S., Tiedje, J.M., 1979. Phases of denitrification following oxygen
depletion in soil. Soil Biology & Biochemistry 11, 261–267.
Statistica 2001. Statistica for windows. StatSoft Inc., Tulsa, OK.
Stemmler, S.H., Berthelin, J., 2003. Microbial activity as a major factor in the
mobilization of iron in the humid tropics. European Journal of Soil Science
54, 725–733.
Szilas, C.P., Borgaard, O.K., Hansen, H.C.B., 1998. Potential iron and
phosphate mobilization during flooding of soil material. Water Air and Soil
Pollution 106, 97–109.
Tabatabai, M.A., Bremner, J.M., 1969. Use of p-nitrophenyl phosphate for
assay of soil phosphatase activity. Soil Biology & Biochemistry 1,
301–307.
Trolard, F., Genin, J.M.R., Abdelmoula, M., Bourrie, G., Humbert, B.,
Herbillon, A., 1997. Identification of a green rust mineral in a
reductimorphic soil by Mossbauer and Raman spectroscopies. Geochimica
et Cosmochimica Acta 61, 1107–1111.

N. Chacon et al. / Soil Biology & Biochemistry 38 (2006) 1494–1499 1499
Vegas-Vilarrubia, T. 1988. Aproximacion a una clasificacion de los rıos de
aguas negras venezolanos atendiendo a las caracterısticas de sus sustancias
humicas y de sus variables fısico-quımicas. MSc thesis, Instituto
Venezolano de Investigaciones Cientıficas, Caracas, DC.
Vegas-Vilarrubia, T., Herrera, R., 1993. Effects of periodic flooding on the
water chemistry and primary production of the Mapire systems
(Venezuela). Hidrobiologıa 262, 31–42.
Vitousek, P.M., 1984. Literfall, nutrient cycling, and nutrient limitation in
tropical forest. Ecology 65, 285–298.
Walkley, A., Black, A., 1934. An examination of the Degtjareff method for
determining soil organic matter and a proposed modification of the chromic
acid titration method. Soil Science 37, 29–38.
Willet, I.R., Higgins, M.L., 1978. Phosphate sorption by reduced and
reoxidized rice soils. Australian Journal of Soil Research 16, 319–326.
Zachara, J.M., Fredrickson, J.K., Li, S.W., Kennedy, D.W., Smith, S.C.,
Gassman, P.L., 1998. Bacterial reduction of crystalline Fe3C oxides in
single phase suspensions and subsurface materials. American Mineralogist
83, 1426–1443.




















